
Abstract
Amidst several global environmental challenges, climate change severely threatens biodiversity, leading to shifts in species distributions, and, in extreme cases, local or global extinctions. Here, we modeled the current and future distributions of biodiversity hotspots for terrestrial venomous snake species across India and evaluated shifts under two climate change scenarios that represent future greenhouse gas concentrations for the years 2050 and 2070. Additionally, to assess potential changes in human-snake conflict zones, we emphasized the four major species of medical importance (hereafter “big four”): Bungarus caeruleus, Naja naja, Daboia russelii, and Echis carinatus. We compiled 4966 occurrence records of 30 species obtained from citizen science platforms, open-access repositories, social media groups, and scientific literature, which were further thinned to 2931 unique locations. We developed species distribution models using MaxEnt by integrating species-specific sets of least-correlated bioclimatic variables. Species-specific distribution maps were overlaid to identify regional hotspots and their predicted spatial shifts. Our projections indicated that around ~ 3% of India’s land area could undergo hotspot turnover by 2070 (in worst-case scenario), including substantial contractions in the Western Ghats and northeast India, and expansions in central India. The consensus habitat suitability for the big four showed a significant positive effect on state-wide snakebite records (β = 1.15 ± 0.4, p < 0.01). Future scenarios suggest increasing snakebite risk in parts of northern India, including Himalaya and northeast India, and southern elevated ranges, such as the Western Ghats. Our study provides the first nationwide assessment of climate-driven distributional shifts in venomous snakes in India, highlighting the need to integrate climate-driven conservation planning with adaptive public health strategies to minimize biodiversity loss and human-snake conflict under future climate change scenarios.
Similar content being viewed by others
Introduction
Climate change is rapidly reshaping global biodiversity patterns1,2,3. Ectothermic animals are among the most vulnerable groups as they rely on environmental temperatures to regulate physiological functions4,5,6,7,8. Among ectotherms, reptiles typically have narrow tolerances and limited dispersal capacities, which make them especially sensitive to rapid climate changes and habitat loss9,10. Despite growing evidence of climate-induced range shifts and extinction risks across vertebrates, reptiles remain underrepresented in global and regional vulnerability assessments1,11,12,13,14.
To understand and forecast species responses to climate change, the World Climate Research Programme launched the Coupled Model Intercomparison Project (CMIP), offering simulations of historical and future climate dynamics under different greenhouse gas emission trajectories15. CMIP findings support global assessments by the Intergovernmental Panel on Climate Change (IPCC) and are widely used in ecological forecasting studies, including reptilian responses to climate scenarios16,17. According to the IPCC Sixth Assessment Report (2022), ecosystems in tropical and polar regions are projected to face conditions beyond historical baselines by mid-century18. Such anomalies are likely to drive elevational and latitudinal range shifts and intensify extinction risk due to habitat reduction or reconfiguration1,11,14,17,19.
Recent global assessments predict significant contractions in climatically suitable habitats for reptiles, particularly in tropical systems where many species already exist near their physiological thresholds1,2,14,17. These changes are expected to cause local extinctions, colonization in novel areas, and altered community assemblages, ultimately impacting ecosystem functioning and stability1,2,13,14,20,21. Forest-dependent and range-restricted reptiles are likely to face a high magnitude of climate change-induced range loss, whereas widespread snakes may expand their ranges by exploiting agricultural lands and peri-urban habitats, thereby increasing the risk of human-snake conflict2,22,23,24,25,26. For instance, projected expansions of venomous snakes in border areas of India, Nepal and Bangladesh could elevate envenomation risk into areas with limited clinical preparedness and antivenom availability2,21.
India harbors over 700 reptile species, nearly half of which are endemic27,28. Of these, 285 are snakes, including more than 40 terrestrial venomous snake species primarily from the families Elapidae and Viperidae28,29,30,31. India also reports the highest snakebite-caused mortalities worldwide, with an estimated 50,000 fatalities annually, disproportionately affecting rural and economically marginalized communities32,33,34,35, However, the actual numbers are likely higher due to underreporting from marginally literate areas35,36. This crisis is largely caused by four widespread and medically important species, collectively known as the “big four”: Common Krait (Bungarus caeruleus), Spectacled Cobra (Naja naja), Russell’s Viper (Daboia russelii), and Saw-scaled Viper (Echis carinatus)30,37,38,39. These species are ecologically dominant and highly tolerant of human-modified environments, which perpetually make them a persistent public health concern40,41. Snakebite envenoming is officially recognized as a Neglected Tropical Disease by the World Health Organization, with India contributing over one-third of the estimated 81,000 to 138,000 global deaths annually34,35,42,43.
Climate-driven changes in the distribution of these species may reshape the geography of snakebite risk in India. Regions currently unaffected could become vulnerable, while some high-risk areas may see declining exposure due to range contraction2,44. Climate projections suggest a potential rise of up to 4 °C in India by the end of the century, emphasizing the urgency of biodiversity response assessments and public health preparedness45,46. Despite these concerns, very few studies have attempted to forecast how venomous snake distributions might respond to future climate conditions and how this may alter human-snake conflict44,47,48,49. Here, we model the current and future distributions of terrestrial venomous snakes across India under two greenhouse gas emission scenarios (climate change scenarios), one moderate (Representative Concentration Pathways RCP 4.5) and one extreme (RCP 8.5), for the years 2050 and 2070. Using species distribution models and citizen science occurrence data, we aim to: (1) identify current and future biodiversity hotspots of terrestrial venomous snakes in India (i.e., areas with predicted high species richness), (2) analyze range shift dynamics of the big four species, and evaluate their spatial congruence with present snakebite incidence patterns.
Materials and methods
Study area
India has a wide range of habitat and climatic diversity across its 28 states and eight union territories, which harbor one of the world’s richest species and ecosystem assemblages, making it a globally important region for biodiversity research (Fig. 1). India’s tropical monsoon climate is characterized by distinct seasonal variability in rainfall and temperature, primarily driven by the southwest and northeast monsoon systems50,51. The country is divided into nine biogeographic zones based on physiography, climate, and vegetation: Trans-Himalaya, Himalaya, Semi-arid, Gangetic plain, Deccan peninsula, Desert, Northeast, Western Ghats, and Coasts52. Additionally, India hosts four globally recognized biodiversity hotspots—the Himalaya, Indo-Burma, Western Ghats, and Sundaland (i.e., areas with high endemic species richness)53,54,55.

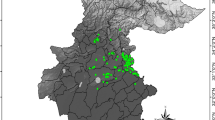
Distribution of spatially thinned occurrence records (n = 2931) for 30 terrestrial venomous snakes species across India, shown with state boundaries and major geographic features. The inset indicates the study area within South Asia. The map was generated using ArcMap version 10.8 (www.support.esri.com).
The Himalaya is geologically the youngest mountain range with ongoing tectonic activity56. Their elevations reach up to 6100 m, and temperatures often reach sub-zero. The region supports cold-adapted and range-restricted reptiles, which makes it particularly vulnerable to climatic shifts57,58. The Gangetic Plain spreads over ~ 16% of the country’s landmass and is heavily influenced by monsoon rainfall. Summer temperatures can exceed 45 °C. Intensive agriculture dominates this fertile alluvial region59. The Western Ghats extend over 1600 km along the western coast and receive annual precipitation exceeding 3000–4000 mm. The region holds mosaics of vegetation types, including grasslands, moist deciduous, and evergreen forests, and is recognized for high reptile endemism and climate sensitivity58,60. The Deccan Peninsula includes varied hill ranges, such as the Vindhyas, Satpuras, Eastern Ghats, and the Chhota Nagpur Plateau. The region spans semi-arid to moist semi-evergreen areas and harbors both generalist and habitat-specialist reptile communities58. The Thar Desert is located west of the Aravalli Mountain range and is characterized by low rainfall, extreme diurnal temperature variation, and sparse xeric vegetation58. Recent irrigation activities have been notably intensified due to the expansion of the Indira Gandhi Canal, which has transformed extensive grasslands into agricultural fields61, which could potentially allow the expansion of more generalist but highly venomous snake species. India’s coastal zone stretches ~7000 km along the Arabian Sea, the Bay of Bengal, and the Indian Ocean, supports mangroves and moist deciduous forests, and provide habitat for several medically important snakes58,62. The northeastern region, including the eastern Himalaya, is a topographically complex and rainfall-rich area, containing dense evergreen and montane forests that support some of India’s rare herpetofaunal diversity58.
Many of these biogeographic zones contain ecotones that are especially vulnerable to climate change, functioning either as corridors or barriers for species range shifts63. The elevational and climatic heterogeneity across India makes it particularly suitable for examining spatial responses of reptiles to climate change.
Checklist of study species
A preliminary checklist of 44 terrestrial venomous snake species was compiled, consisting of 19 species from the family Elapidae and 25 from the family Viperidae (Supp. Table 1). This list was constructed using peer-reviewed literature, biodiversity portals, and verified citizen science datasets. The use of large-scale biodiversity databases has become increasingly systematic and robust, along with the increasing applications of data-driven ecology64. Geneviève et al.65 and Chowdhury et al.66,67 highlighted the applications of open-access repositories, citizen science, and social media platforms in understanding the spatial and temporal resolutions of biodiversity assessments in addition to mobilizing species occurrence records to bridge geographic and taxonomic knowledge gaps in underrepresented taxa.
For this study, we selected data sources based on accessibility, authenticity, and the presence of metadata to ensure reliability and consistency. These included established citizen science repositories, such as Global Biodiversity Information Facility (GBIF; www.gbif.org; n = 67 records)68, Indian Biodiversity Portal (www.indiabiodiversity.org; n = 594)69, iNaturalist (www.inaturalist.org; n = 2270)70, VertNet (www.vertnet.org; n = 1)71 and Reptile Database (www.reptile-database.reptarium.cz; n = 2)72. We filtered occurrence records from 1980 onwards to ensure recent taxonomic and technological advancements. Additionally, we compiled species records from Facebook groups (n = 1181), such as Indian Snakes (facebook.com/groups/indiansnakes; n = 564) and Snakes of India (facebook.com/groups/SnakesofIndia; n = 617), from start of January 2011 to end of December 2022. These records were included only when the location and identification were provided and validated through photographs and expert confirmation. Furthermore, we conducted a comprehensive literature search using Google Scholar (https://scholar.google.com/) and ResearchGate (https://www.researchgate.net/), to target studies through combinations of keywords such as “venomous snakes,” “Elapids,” “Vipers,” “Ophiofauna,” “India,” “Protected areas,” and “Occurrence records.” This search yielded 853 additional records. Wherever the precise coordinate data were absent, the centroid of the nearest identifiable administrative boundary (village, taluka, or district; within ~ 1 km) was assigned. Species occurrence points were broadly validated using field guides and expert opinions.
We curated and extracted occurrence information, including species name, geographic location (latitude and longitude), year of the observation reporting, and the respective sources. To reduce sampling bias and spatial autocorrelation, we spatially filtered occurrence data for each species at 1 km2 distance using the SDM Toolbox version 2.4 in ArcMap20. Following minimum data thresholds highlighted in species distribution modeling literature73,74, species with fewer than 10 thinned records were excluded from further spatial analyses. As a result, 14 species were removed, and the final dataset resulted in 2931 records belonging to 30 species (16 Elapidae and 14 Viperidae). We assumed that the current prediction models also represent the climatic tendencies of excluded species75.
We created species-specific 99% Minimum Convex Polygons (MCPs) using the filtered occurrence data (n = 2931) to outline the species-specific extent of climatic suitability. This framework reduces the prediction risk to climatically similar but biologically inaccessible areas76. We applied a 100 km buffer around each species’ MCP and combined with the local state boundaries, considering the reporting uncertainty of occurrence records and possible near-future reports. The buffered MCP was defined as the modeling extent for each species.
Predictor variables selection
To estimate spatial shifts in species distributions under climate change, we used bioclimatic variables obtained from WorldClim version 2 (www.worldclim.org; Supp. Table 2)77. The dataset includes 19 bioclimatic variables at a spatial resolution of 30 arc-seconds (~ 1 km), available for current and future climate conditions extracted from the Coupled Model Intercomparison Project Phase 5 (CMIP5)15. Future climate projections were based on Representative Concentration Pathways (RCPs), which were standardized greenhouse gas concentration trajectories highlighted by the IPCC to describe alternative climate futures18,78. We used two scenarios: RCP 4.5, representing a stabilization pathway with emission peaking around 2040 and then declining, and RCP 8.5, indicating a high-emission worst-case scenario with continued increase through 21st century. Projections obtained for mid-century (2050; average of 2041–2060), and late-century (2070; average of 2061–2080). The same set of bioclimatic predictors was used consistently for projections across the four temporal scenarios (RCP 4.5 2050, RCP 4.5. 2070, RCP 8.5 2050, and RCP 8.5 2070).
Raster layers were converted to ASCII format, and raster values were extracted at the occurrence points using current climatic data. Records with missing or invalid raster values due to imprecise coordinates were excluded from the analysis. We computed pairwise Pearson’s correlation coefficients (r) among predictor variables for each species to reduce multicollinearity and to improve model stability. We retained a subset of ecologically meaningful and statistically least-correlated predictors (r < |0.7|) for each species independently79,80. The final set of variables thus represented species-specific climatic drivers and formed the basis for both current and future predictive models.
Spatial modeling
Spatial shift in biological hotspots
We modeled future species distributions using the presence-only framework implemented in MaxEnt software version 3.4.481,82 for each of the 30 venomous snake species. Each model was trained using 70% of the filtered occurrence records and tested with the remaining 30% to evaluate predictive performance (including pseudoabsences)83. Pseudoabsences were generated at a ratio of 1:100 relative to the number of occurrence points per species (Supp. Table 1). To reduce overprediction in areas lacking records, we generated species-specific sampling bias surfaces by assigning a weight of 1.0 to the grid cells within a 1 km radius of occurrence points, 0.5 to cells within a 10 km buffer around 1 km buffer, and 0.01 to all other cells (at 1 km resolution). These layers were further rasterized and converted to ASCII format for model use. To minimize variability and improve robustness, we conducted 10 replicate model runs for species with > 100 presence records and 50 replicates with < 100 presence records84. Models were generated in logistic format, which provides continuous relative values of habitat suitability ranging from 0 (unsuitable) to 1 (highly suitable)73,85. We kept MaxEnt settings at default (regularization multiplier 1, convergence threshold 10e − 6, prevalence value 0.5, and 500 maximum iterations)81. We enabled feature classes including linear, quadratic, product, threshold, hinge and their combinations73,84. To classify suitable vs. unsuitable areas, we applied the ten-percentile logistic training presence threshold (10p-LPT)73,86,87. For each species, cells above this threshold were defined as suitable habitat, while those below were unsuitable. This binarization was applied across five climatic scenarios: current, 2050 RCP 4.5, 2070 RCP 4.5, 2050 RCP 8.5, and 2070 RCP 8.578. With slight adjustments from Bertelsmeier et al.88, we also used species-specific linear regression to evaluate the temporal trends in suitable habitat area between current (2020), mid-century (2050), and late-century (2070) projections under both RCP scenarios.
Further, we overlaid all binary outputs within each scenario to produce cumulative suitability maps, where pixel values represented the number of species with suitable habitat at that area (i.e., predicted species richness)39,40,44,89. To delineate venomous snake hotspots, we classified pixels with richness > 50% of the maximum observed values as hotspots (coded as 1) while the remainder as non-hotspots (0)90,91,92.
To assess spatial shifts in hotspots under climate change, we used the “Distribution changes between binary SDMs” tool under the SDM toolbox in ArcMap version 10.520. We did pairwise comparisons between the current scenario and each of the four future projections, these comparisons will help in identifying regions of predicted habitat loss (range contraction/population extinction) and gain (range extension/population colonization)93,94. We further calculated area turnover in habitat suitability using the formula (formula 1)95,96:
…….
Where the units of loss, gain, and constant areas of habitat suitability for all venomous snake species are in km2.
Potential human–big four conflict
To assess how climate change may affect the distribution of the big four and consequently alter the snakebite risk scenario, we used species distribution models to predict habitat suitability for each of the big four species under five climatic scenarios: current (2020), RCP 4.5 2050, RCP 8.5 2050, RCP 8.5 2050 and RCP 8.5 207026,97. We generated a consensus habitat suitability surface by averaging the predicted suitability values of the big four under each scenario, thereby producing an integrated climatic suitability layer.
We extracted state- and union territory-level snakebite records from Ministry of Health and Family Welfare 2020 for the years of 2016 to 201843. For standardization, we calculated snakebite cases as incidence density (number of cases km− 2 year− 1). Literacy ratio of all states and union territories from the 2001 census was used to represent education levels98, under the assumption that higher literacy promotes improved reporting of snakebite incidents. Median of literacy ratio was considered threshold to categorize states and union territories into high and low literacy classes. These were also grouped into five broad geographic zones: northern (Himachal Pradesh, and Uttarakhand), central (Bihar, Chhattisgarh, Jharkhand, Madhya Pradesh, Maharashtra, Odisha, Uttar Pradesh, and West Bengal), western (Chandigarh, Dadar & Nagar Haveli, Daman & Diu, Delhi, Gujarat, Haryana, Punjab, and Rajasthan), southern (Andhra Pradesh, Telangana, Goa, Karnataka, Kerala, Puducherry, and Tamil Nadu) and northeastern zones (Arunachal Pradesh, Assam, Manipur, Meghalaya, Mizoram, Nagaland, Sikkim, and Tripura). Jammu & Kashmir, Ladakh, and Islands were excluded from the analysis, as large fraction of these bodies were outside 99% MCPs of big fours.
We fitted generalized linear mixed effect models to assess the relationship between snakebite density and the mean climatic suitability for the big four, with elevation included as an additive fixed effect. Literacy class and geographic zone were included as categorical random intercepts. Model selection was based on secondary AIC (AICc), accounting for small sample size (n = 31)99. Models were compared with their candidate models, taking into consideration the combination of fixed effect variables (pbigfour and elevation) and the null model (~1).
Finally, to identify future risk hotspots, we subtracted future habitat suitability surfaces from the current surface, which resulted in relative difference maps that indicated areas of relative potential gain (colonization) and relative loss (extinction). Colonization zones were interpreted as emerging risk regions where snakebite incidences may increase due to range expansion of the big four, while extinction zones were identified where suitability difference showed decline in suitable habitats93. The inferences were made through calculated turnover of colonization and extinction areas using the following formula (formula 2)95,96:
….
Where extinction, colonization and stable areas were measured in km2.
Results
Species presence records and predictor variables
A total of 30 terrestrial venomous snake species were included in the climatic suitability analysis (Fig. 1), 16 from the family Elapidae and 14 from the family Viperidae. After thinning at 1 km², the dataset retained 2931 unique occurrence records (Fig. 1). Species < 10 records were excluded from prediction analysis.
Among Elapidae, Naja naja and Bungarus caeruleus had the highest number of thinned records, whereas Naja sagittifera had the fewest. Among Viperidae, Echis carinatus had the highest number of records and Craspedocephalus strigatus the lowest (Fig. 1; Supp. Table 1).

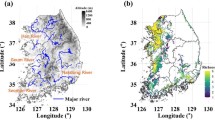
Projected spatial changes in venomous snake hotspots in India relative to the current distribution under four future climate scenarios: (A) 2050 RCP 4.5, (B) 2050 RCP 8.5, (C) 2070 RCP 4.5, and (D) 2070 RCP 8.5. Green shading indicates colonization (expansion of hotspots), blue shading indicates extinction (contraction of hotspots), and grey shading shows current hotspots’ persistence in the future (≥ 10 species). The map was generated using ArcMap version 10.8 (www.support.esri.com).
Spatial shift in biological hotspots
Overall, hotspot modeling revealed substantial changes in venomous snake distributions under future scenarios. Considering all 30 species, prediction analysis under the 2070 RCP 8.5 scenario revealed a net gain of ~ 47,000 km2 (colonization) and a net loss of ~ 48,500 km2 (extinction), corresponding to a turnover of 23.88% and affecting ~ 2.66% of India’s landmass (Table 1). Of all hotspots identified under the current scenario, only 72.93% persisted to 2070 RCP 8.5. Existing hotspot distributions were mainly concentrated in the Western Ghats, central India, and northeastern states (Fig. 2A–D). Overall, the largest contraction and fragmentation predicted to occur in the Western Ghats, particularly in northern Karnataka, southern and eastern Maharashtra, while a few pockets in central India and northern Western Ghats showed hotspot emergence under future climates, especially in severe climate scenarios (Fig. 2B, D). Northeastern India, especially the north Bengal region exhibited consistent declines in suitable habitat across all scenarios, but more intense in the worst-case, suggesting concentrated losses of snake species richness.
Species-specific regression analysis (Supp. Tables 4, 5; Supp. Fig. 4) looked at the relationship between suitable habitat area and time (current, 2050, 2070) under varied climate scenarios. In the moderate scenario (RCP 4.5), Bungarus fasciatus, B. sindanus, and Calliophis melanurus exhibited significant declines in suitable habitat area (p < 0.1) (Supp. Table 4), which are very widely distributed species as of the current understanding. Several species, including B. andamanensis, B. caeruleus, B. fasciatus, B. sindanus, B. walli, C. melanurus, Craspedocephalus malabaricus, Daboia russelii, Hypnale hypnale, Naja naja, N. sagittifera, and Ovophis monticola, demonstrated a strong model fit (R² > 0.6), indicating largely a consistent decline in habitat suitability over time (Supp. Fig. 4; Supp. Table 4). These species also include three of the big four as well. In contrast, species distributed in limited biogeographic zones, such as, B. lividus, C. bibroni, C. gramineus, C. macrolepis, and N. kaouthia exhibited increase in suitable habitat areas over time with robust model fits (Supp. Table 4; Supp. Fig. 4).
Under the worst-case scenario (RCP 8.5), significant contraction was observed in B. andamanensis, C. melanurus, N. sagittifera, O. monticola, and Trimeresurus septentrionalis, whereas N. kaouthia significantly expanded. B. andamanensis, N. sagittifera, and O. monticola have small range of distribution, and higher emission scenario may push these species towards extinction (Supp. Table 4; Supp. Fig. 4). Additional expansions with strong model fit were observed in C. bibroni, C. castoe, C. gramineus, C. macrolepis, C. peltopelor, N. kaouthia, Ophiophagus hannah, and T. salazar, including some of the important species, such as King cobra and pit vipers. Proportional fraction of suitable habitat for the species within each biogeographic zones is given in Supp. Fig. 3.
Prediction inferences were based on models exhibiting high mean AUC of 0.72 to 0.99 (Supp. Table 3). Species-specific distribution maps with respect to climatic scenarios are shown in Supp. Fig. 1 (species 1–30). These results demonstrate that climate change may simultaneously drive both local extinctions and expansions, which will further reshape the geography of snake biodiversity hotspots across India.
Potential human–big four conflict
The best fit generalized linear mixed-effects model indicated that climatic suitability of the big four shows a strong positive relation with snakebite incidence density (β = 1.15 ± 0.4, p < 0.01) across mainland Indian states and union territories (Table 3; Supp. Fig. 5). The significant p-value supports our expectation that higher climatic suitability corresponds to greater snakebite risk. Elevation had a weak and non-significant effect (β = − 0.11 ± 0.31, p > 0.05) (Table 3; Supp. Fig. 5). Random intercepts for geographic zone and literacy class explained additional variance of 61% and 24%, respectively (R2 = 0.65), interpreting the influence of spatial and socio-demographic heterogeneity (Table 3; Supp. Fig. 6).
Across scenarios, the most distinct expansion of potential snakebite risk was observed under the 2070 RCP 4.5 scenario, with an increase projected in the Deccan Plateau, Gangetic Plains, and Himalayan foothills (Fig. 3). The 2050 RCP 8.5 scenario showed the most extensive decline in suitable habitat, indicating potential range contractions in the consensus surface (Fig. 3). Notably, the 2070 RCP 8.5 scenario exhibited both extensive expansion (~ 255,600 km2) and contraction (~ 146,200 km2), suggesting strong population displacements rather than general range shifts (Table 2).

Projected changes in the potential distribution of big four venomous snake species (Bungarus caeruleus, Naja naja, Echis carinatus, Daboia russelii) across India under four climate change scenarios: (A) RCP 4.5 2050, (B) RCP 8.5 2050, (C) RCP 4.5 2070, and (D) RCP 8.5 2070. RCP 4.5 represents a moderate stabilization scenario and RCP 8.5 represents a high-emission worst-case scenario. Green indicates expansion (colonization), and blue shading indicates contraction (extinction). The map was generated using ArcMap version 10.8 (www.support.esri.com).
The proportional change in the big four’s distribution relative to state and union territory areas is presented in Figs. 3 and 4. Colonization trends were most evident in northern and northeastern states, including Uttarakhand, Uttar Pradesh, Bihar, West Bengal (north Bengal region), western Meghalaya, and the Western Ghats in Kerala. Expansions were also projected in northern and northeastern states, including Himachal Pradesh, Sikkim, Manipur, Mizoram, Tripura, Assam, and Kerala, and some of the central states, such as West Bengal, Jharkhand, Chhattisgarh, Uttar Pradesh, and Madhya Pradesh (Figs. 3 and 4). These regions may face elevated snakebite risk under all modeled climatic scenarios. Conversely, states such as Karnataka, Tamil Nadu, and parts of Gujarat consistently showed a decline in suitability, suggesting a reduction in big four distributions and potentially reduced snakebite risk. Similar declines were projected in southern states, such as Goa, Puducherry, Andhra Pradesh, Telangana, and Maharashtra (Figs. 3 and 4). Overall, under the 2070 RCP 4.5 scenario, ~ 262,000 km² of new habitat became suitable, while ~ 105,000 km² was lost, highlighting a positive balance of expansion (Table 2).

Proportional change in the potential distribution of the big four venomous snakes relative to the total area of each state and union territory under four climate scenarios. Blue bars represent the moderate emissions pathway (RCP 4.5; light—2050 and dark—2070), while green bars indicate the worst-case scenario (RCP 8.5; light—2050 and dark—2070). Positive values indicate expansion of colonization of new areas whereas negative values indicate habitat loss. The labels on the x-axis indicate the geographic zones, such as brown (southern), blue (western), black (central), pink (northeast), and red (northern states).
Discussion
Current hotspots and future shifts
Our nationwide analysis shows that climate change is likely to substantially reshape the distribution of venomous snake biodiversity hotspots across India. The magnitude of turnover scales with carbon-emission scenarios, with the worst-case pathway (RCP 8.5, 2070) causing ~ 95,600 km2 area change in the hotspot distribution, and even moderate scenario (RCP 4.5, 2070), resulting in ~66,800 km2 of turnover (Table 1; Fig. 2). Such outcomes emphasize on the urgency of dynamic conservation planning rather than static gap-analyses.
Precipitation-related variables were found to be dominant predictors of hotspot distribution, especially seasonality and dry-season moisture (Supp. Figure 2) (similar findings to1,2,49. These findings align with global evidence that rainfall strongly limits reptile distributions by influencing refuge humidity, prey access, and activity windows1,2,100. Model projections indicate persistent suitability in the Western Ghats, central India, and parts of the northeast, though with substantial spatial change in habitats (Fig. 2). In the Western Ghats, steep gradients and high endemism form a mountain-top squeeze strategy where elevational shift rapidly exhausts available habitat, which further leads to habitat fragmentation, population isolation, and extinction risks (Fig. 2)14,101,102,103. Central India, in contrast, shows potential expansion of suitable climatic space potentially as hydrothermal windows broaden, which are more likely to allow disturbance-tolerant generalist species (e.g., Echis carinatus, Naja naja) to spread across agro-mosaics and peri-urban landscapes (Fig. 2)22,23,24,25,26. Conversely, parts of north Bengal may lose suitability despite high annual rainfall, due to increasing intra-seasonal dry spells104,105. In these cases, the landscape may host an influx of exotic species106, that may get benefits from growing human infrastructures107. These regional contrasts highlight how precipitation redistribution, rather than alone temperature, influences ectothermic responses in monsoon-driven tropical systems1,2,49.
Species-level responses
Species’ trajectories towards habitat selection and survival diverge according to traits under climate change108. Disturbance-tolerant and thermally flexible taxa (e.g., N. kaouthia, C. gramineus) are projected to expand in central and northeastern regions, due to their ability to exploit human-modified areas (Supp. Figures 1, 4)22,23,107. In contrast, small-ranged montane specialists (e.g., Ovophis monticola) face prominent contractions under high-emission scenarios following mountain-top squeeze theory, which has been widely projected in tropical montane reptiles (Supp. Figure 2)102,103. Several widespread taxa (such as Bungarus fasciatus, B. sindanus, C. melanurus, C. malabaricus) also exhibit projected range declines (Supp. Figure 1), likely linked to intensifying dry-season stress and shift in monsoon timing. These trait-based differences emphasize that climate change will intensify generalist–specialist contrasts, expanding generalists and promoting biotic homogenization while contracting habitat-restricted lineages22,23. Additionally, the impact of climate change includes inhibition of dispersal in case of habitat unavailability around, which trigger to the local or regional extinction of the species1,14,37. A few prior studies have made attempts to model the impacts of climate change on venomous snakes, that include central, south and southeast Asian Naja cobras. The studies have evidently informed that climate change will trigger the habitat contraction of these species, as similar to our findings [26, 21, 49].
Big four and snakebite risk
Understanding the possible range shifts of the big four species is critical for mitigation planning during future human–snake conflicts in India1,2,40. Our mixed-effect models showed a strong positive relationship between climatic suitability and snakebite case density across states (β = 1.15 ± 0.4, p < 0.01) (Table 3; Supp. Figure 5). Projected increases in risk concentrate along the Himalayan foothills, central highlands, and parts of the northeast, while peninsular states such as Karnataka, Tamil Nadu, and Gujarat show declines (Fig. 3) (also observed by1. These patterns are biologically plausible, as wetter and warmer conditions extend snake activity windows, agricultural timing synchronize with their peak activity, and disturbance-tolerant species thrive in agro-ecosystems and irrigation areas, offering high likelihood of spatial and temporal overlap with people109. Socio-demographic factors and geographic factors played a key role in reporting the cases (Supp. Figure 6).
By 2070, a coherent northward and upslope shift is projected for the big four, aligning to global studies stressing on latitudinal or elevational reptile shifts under warming110,111,112. This underpins the need for anticipatory public health interventions, including pre-positioning antivenom, rural health centers, and prevention campaigns during crop cycles and seasonal activity. In projected decline zones, long-term surveillance is still necessary, as micro-refugia, such as crop lands and urban heat islands, may tolerate localized risk21,113. Importantly, reductions in snake populations (species richness) could diminish ecosystem services, such as rodent control, which inevitably cause novel uncertainties for agro-ecosystem and public health114. From human concern, snakebite often leaves lasting physical (e.g., tissue loss, amputations, or musculoskeletal impairment) and psychological sequelae (e.g., anxiety, depression, post-traumatic stress), which influence quality of life and work capacity109,115. In rural and low-income areas, these effects propagate into household economic insecurity and social marginalization115. In essence, snakebite, being a neglected tropical disease, causes high mortality and long-term disability, yet remains underexplored and underprioritized despite its heavy socioeconomic toll109,115,116. Integrating snakebite into broader neglected tropical disease strategies, through prevention, timely antivenom access, and post-bite rehabilitation, can substantially reduce human suffering in vulnerable rural communities116,117, as collective range of big four is predicted to experience more expansion than reduction in both climatic scenarios by the end of the century (Table 2).
India’s infrastructure development is marked by pronounced regional disparities, with southern and northern states generally having more developed infrastructure118. Notably, the northern states are projected to experience a relatively higher likelihood of snakebite cases, as our models indicate a greater probability of range shift of the big four species in these regions, particularly towards the late 21st century. At the same time, if we look at species-specific predictions, substantial portions of climatically suitable habitat for the big four snakes are projected to decline in other areas, especially in southern India. Such contraction zones may provide ecological space for other venomous species to become locally dominant26, necessitating additional medical preparedness and region-specific antivenom strategies. Our study is among the first to explicitly describe species distribution projections of the medically important big four, relating it with snakebite incidence data across India, thereby joining ecological modeling with public health outcomes and providing a framework for anticipating region-specific risks under climate change.
Limitations and potential biases
Our models rely on presence-only data, which remain spatially biased despite efforts to mitigate this through thinning and sampling bias layers119,120. In India, large scattered patches in central and northern states are under-sampled in terms of efforts representing scientific studies and citizen science datasets on venomous snakes, which may bias hotspot prediction113, but this assessment represents the most comprehensive description of terrestrial venomous snakes’ distributions in India to date. This can definitely be enhanced in the later stage by incorporating land use change and other stressors. Also, snakebite incidence data were available only as state-level aggregates for 2016–201843, which were found relatively most reliable from government sources. However, the data lacked the fine-scale spatial resolution to capture fine-scale heterogeneity in snakebite-prone areas. However, to partly address this, we standardized snakebite incidence as cases km− 2 year− 1 and such consideration smoothened the local variations and generalized the risk at underreported regions.
This modeling framework assumes quasi-equilibrium with species distribution and current climate121,122. However, dispersal limits and rapid environmental change may increase the variations from projected niches122,123. Moreover, our models are solely based on climatic predictors, and do not explicitly reveal effects of land use change, prey availability, and human pressures2, which are known to shape species distributions and their habitat selection on a finer scale. Also, our dataset excluded > 10 species due to their insufficient occurrence records (as n < 10), which may restrict the actual representation of hotspot distribution. Nevertheless, we assumed that the included species adequately capture the broader biogeographic and ecological patterns of venomous snake richness in India.
Conclusions
Our study provides the first nationwide assessment of climate-driven distributional shifts in venomous snake species in India, linking biodiversity dynamics with snakebite risk. The projected reconfiguration of snake species hotspots indicates large-scale habitat shifts that could alter ecosystem services while intensifying snakebite risk in vulnerable regions. In particular, the projected change in the range of the big four medically important species suggests a potential increase in human-snake interactions and snakebite risk, particularly in northern and central India. These findings can inform biodiversity conservation priorities and guide targeted snakebite management strategies, including antivenom allocation and strengthening rural health preparedness. Conservation efforts should prioritize vulnerable habitats, especially in the Western Ghats and northeast India. Integrating biodiversity conservation with public health planning will be critical for promoting coexistence under future climate scenarios.
Data availability
The data generated and used in the paper will be made available to the reader. The reader can write to the corresponding author with a reasonable request.
Change history
14 December 2025
The original online version of this Article was revised: In the original version of this Article the Supplementary Information files contained tracked changes. The original Article has been corrected.
References
Biber, M. F., Voskamp, A. & Hof, C. Potential effects of future climate change on global reptile distributions and diversity. Glob. Ecol. Biogeogr. 32 (4), 519–534. https://doi.org/10.1111/geb.13646 (2023).
Martinez, P. A. et al. Climate change-related distributional range shifts of venomous snakes: a predictive modelling study of effects on public health and biodiversity. Lancet Planet. Health. https://doi.org/10.1016/S2542-5196(24)00005-6 (2024).
Urban, M. C. et al. Improving the forecast for biodiversity under climate change. Science 353, 6304. https://doi.org/10.1126/science.aad8466 (2016).
Dayananda, B., Bezeng, S. B., Karunarathna, S. & Jeffree, R. A. Climate change impacts on tropical reptiles: likely effects and future research needs based on Sri Lankan perspectives. Front. Ecol. Evol. 9, 688723. https://doi.org/10.3389/fevo.2021.688723 (2021).
Diele-Viegas, L. M. & Rocha, C. F. D. Thermal ecology and the potential effects of climate change on neotropical lizards. In Effects of Climate Change on Amphibians and Reptiles. https://doi.org/10.5772/intechopen.74343 (IntechOpen, 2018).
Evans, G. W. Projected behavioral impacts of global climate change. Annu. Rev. Psychol. 70, 449–474. https://doi.org/10.1146/annurev-psych-010418-103023 (2019).
Gardner, T. A., Barlow, J. & Peres, C. A. Paradox, presumption and pitfalls in conservation biology: the importance of habitat change for amphibians and reptiles. Biol. Conserv. 138 (1–2), 166–179. https://doi.org/10.1016/j.biocon.2007.04.017 (2007).
Kellogg, W. W. & Schware, R. Climate Change and Society: Consequences of Increasing Atmospheric Carbon (Routledge, 2019).
Lopez-Alcaide, S. & Macip-Ríos, R. Effect of temperature on activity patterns in reptiles. In Wildlife Management: Concepts and Applications, vol. 9 (2011).
Velásquez-Tibatá, J., Salaman, P. & Graham, C. H. Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Envriron. Chang. 13 (2), 235–248 (2013).
Freeman, B. G., Scholer, M. N., Ruiz-Gutierrez, V. & Fitzpatrick, J. W. Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. Proc. Natl. Acad. Sci. 115 (47), 11982–11987. https://doi.org/10.1073/pnas.1804224115 (2018).
Lourenco-de-Moraes, R. et al. Climate change will decrease the range size of snake species under negligible protection in the Brazilian Atlantic forest hotspot. Sci. Rep. 9, 8523. https://doi.org/10.1038/s41598-019-44732-z (2019).
Massot, M., Clobert, J. & Ferrière, R. Climate warming, dispersal Inhibition and extinction risk. Glob. Change Biol. 14 (3), 461–469. https://doi.org/10.1111/j.1365-2486.2007.01514.x (2007).
Farooq, H., Harfoot, M., Rahbek, C. & Geldmann, J. Threats to reptiles at global and regional scales. Curr. Biol. 34 (10), 2231–2237. https://doi.org/10.1016/j.cub.2024.04.007 (2024).
Noce, S., Caporaso, L. & Santini, M. A new global dataset of bioclimatic indicators. Sci. Data 7, 398 https://doi.org/10.1038/s41597-020-00726-5 (2020).
Archis, J. N., Akcali, C., Stuart, B. L., Kikuchi, D. & Chunco, A. J. Is the future already here? The impact of climate change on the distribution of the Eastern coral snake (Micrurus fulvius). PeerJ 6, e4647. https://doi.org/10.7717/peerj.4647 (2018).
Cerasoli, F. et al. Determinants of habitat suitability models transferability across geographically disjunct populations: insights from Vipera ursinii ursinii. Ecol. Evol. 11 (9), 3991–4011 (2021).
Pörtner, H. O. et al. Technical summary. In Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II To the Sixth Assessment Report of the Intergovernmental Panel on Climate Change (ed Pörtner, H. O.) 37–118. https://doi.org/10.1017/9781009325844.002 (Cambridge University Press, 2022).
Rowe, K. C. et al. Spatially heterogeneous impact of climate change on small mammals of montane California. Proc. Royal Soc. B: Biol. Sci. 282 (1799), 20141857. https://doi.org/10.1098/rspb.2014.1857 (2015).
Brown, J. L., Bennett, J. R. & French, C. M. SDMtoolbox 2.0: the next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 5, e4095. https://doi.org/10.7717/peerj.4095 (2017).
Chowdhury, M. A. W. et al. Favourable climate niche in low elevations outside the flood zone characterizes the distribution pattern of venomous snakes in Bangladesh. J. Trop. Ecol. 38 (6), 437–450. https://doi.org/10.1017/S0266467422000359 (2022).
McKinney, M. L. & Lockwood, J. L. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 14 (11), 450–453. https://doi.org/10.1016/S0169-5347(99)01679-1 (1999).
Naaf, T. & Wulf, M. Habitat specialists and generalists drive homogenization and differentiation of temperate forest plant communities at the regional scale. Biol. Conserv. 143 (4), 848–855. https://doi.org/10.1016/j.biocon.2009.12.027 (2010).
Frishkoff, L. O. et al. Countryside biogeography: the controls of species distributions in human-dominated landscapes. Curr. Landsc. Ecol. Rep. 4, 15–30. https://doi.org/10.1007/s40823-019-00037-5 (2019).
Williams, J. J., Bates, A. E. & Newbold, T. Human-dominated land uses favour species affiliated with more extreme climates, especially in the tropics. Ecography 43 (3), 391–405. https://doi.org/10.1111/ecog.04806 (2019).
Nori, J., Carrasco, P. A. & Leynaud, G. C. Venomous snakes and climate change: ophidism as a dynamic problem. Clim. Change. 122, 67–80. https://doi.org/10.1007/s10584-013-1019-6 (2014).
Jangid, A. K. & Kamdar, A. Indian reptiles in the iucn red list. In Reptiles of India, v. 1.26 (eds Kamdar, A. et al.) (Indian Foundation for Butterflies, 2022).
Kamdar, A. et al. Reptiles of India, v. 1.26 (Indian Foundation for Butterflies, 2022).
Uetz, P., Freed, P., Aguilar, R. & Hošek, J. (eds) The Reptile Database. http://www.reptile-database.org (Accessed 23 November 2022) (2022).
Whitaker, R. & Martin, G. Diversity and distribution of medically important snakes of India. In Clinical Toxinology in Asia Pacific and Africa 115–136. https://doi.org/10.1007/978-94-007-6386-9_16 (2015).
Whitaker, R. & Captain, A. Snakes of India: the Field Guide (Draco Books, 2008).
Kasturiratne, A. et al. The global burden of snakebite: a literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 5 (11), e218. https://doi.org/10.1371/journal.pmed.0050218 (2008).
Longbottom, J. et al. Vulnerability to snakebite envenoming: a global mapping of hotspots. Lancet 392 (10148), 673–684. https://doi.org/10.1016/S0140-6736(18)31224-8 (2018).
Menon, J. C. et al. ICMR task force project-survey of the incidence, mortality, morbidity and socio-economic burden of snakebite in india: A study protocol. PLoS ONE. 17 (8), e0270735. https://doi.org/10.1371/journal.pone.0270735 (2022).
Suraweera, W. et al. Trends in snakebite deaths in India from 2000 to 2019 in a nationally representative mortality study. Elife 9, e54076. https://doi.org/10.7554/eLife.54076 (2020).
Government of India. Ministry of Health and Family Welfare. Management of Snakebite, Standard Treatment Guideline. https://nhm.gov.in/images/pdf/guidelines/nrhm-guidelines/stg/Snakebite_Full.pdf (Accessed 10 March 2024) (2016).
Chinnasamy, R. & Prabakaran, N. Living with snakes in india: the intensifying health crisis over snakebites—Challenges and way ahead. Asian J. Conserv. Biology. 8 (2), 220–223 (2019).
Gopal, G. et al. The concept of big four: road map from snakebite epidemiology to antivenom efficacy. Int. J. Biol. Macromol. 242 (1), 124771. https://doi.org/10.1016/j.ijbiomac.2023.124771 (2023).
Yousefi, M., Kafash, A., Khani, A. & Nabati, N. Applying species distribution models in public health research by predicting snakebite risk using venomous snakes’ habitat suitability as an indicating factor. Sci. Rep. 10 (1), 18073. https://doi.org/10.1038/s41598-020-74682-w (2020).
Srinivasulu, A., Srinivasulu, B. & Srinivasulu, C. Ecological niche modelling for the conservation of endemic threatened squamates (lizards and snakes) in the Western Ghats. Global Ecol. Conserv. 28, e01700. https://doi.org/10.1016/j.gecco.2021.e01700 (2021).
Whitaker, R. & Martin, G. Diversity and distribution of the medically important snakes of India. In Clinical Toxinology (eds Gopalakrishnakone, P. et al.) https://doi.org/10.1007/978-94-007-6288-6_16-1 (Springer, 2014).
WHO (World Health Organization). Snakebite Envenoming. https://www.who.int/news-room/fact-sheets/detail/snakebite-envenoming (Accessed 10 January 2024) (2023).
Government of India & Ministry of Health and Family Welfare. State/UT-Wise Cases and Deaths Due to Snake Bite During the Years 2016–18 (Government of India, 2020).
Zacarias, D. & Loyola, R. Climate change impacts on the distribution of venomous snakes and snakebite risk in Mozambique. Clim. Change. 152 (1), 195–207. https://doi.org/10.1007/s10584-018-2338-4 (2018).
Crowell, H. L. et al. Thermal ecology and baseline energetic requirements of a large-bodied ectotherm suggest resilience to climate change. Ecol. Evol. 11 (12), 8170–8182. https://doi.org/10.1002/ece3.7649 (2021).
Thuiller, W. Climate change and the ecologist. Nature 448 (7153), 550–552 (2007).
Martín, G. et al. Implications of global environmental change for the burden of snakebite. Toxicon 10 (9), 100069. https://doi.org/10.1016/j.toxcx.2021.100069 (2021).
Martín, G. et al. Integrating snake distribution, abundance and expert-derived behavioural traits predicts snakebite risk. J. Appl. Ecol. 59 (2), 611–623. https://doi.org/10.1111/1365-2664.14081 (2021).
Chowdhury, M. A. W., Müller, J. & Varela, S. Climate change and the increase of human population will threaten conservation of Asian cobras. Sci. Rep. 11, 18113. https://doi.org/10.1038/s41598-021-97553-4 (2021).
Mani, M. S. Physical features. In Ecology and Biogeography in India (ed Mani, M. S.) 11–59 (Dr. W. Junk, 1974).
Varikoden, H., George, C., Ancy, P. & Babu, C. A. A comprehensive review on the recent changes in Indian summer monsoon rainfall and its relations with global climate drivers based on the CMIP6 simulations. Atmos. Res. 326, 108302. https://doi.org/10.1016/j.atmosres.2025.108302 (2025).
Rodgers, W. A. & Panwar, S. H. Biogeographical Classification of India (1988).
Reid, W. V. Biodiversity hotspots. Trends Ecol. Evol. 13 (7), 275–280 (1998).
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403, 853–858. https://doi.org/10.1038/35002501 (2000).
Chitale, V. S., Behera, M. D. & Roy, P. S. Global biodiversity hotspots in india: significant yet under studies. Curr. Sci. 108 (2), 149 (2015).
Zilio, L. D., Hetényi, G., Hubbard, J. & Bollinger, L. Building the himalaya from tectonic to earthquake scales. Nat. Reviews Earth Environ. 2, 251–268. https://doi.org/10.1038/s43017-021-00143-1 (2021).
Chettri, B. & Tamang, R. Status and trends of herpetofaunal diversity in the himalaya biodiversity hotspot. In Biodiversity Hotspot of the Himalaya 215–244 (Apple Academic Press, 2024).
Deuti, K., Ganesh, S. R. & Chandra, K. Diversity, distribution and endemicity of herpetofauna in different biogeographic zones and biodiversity hotspots of India. In Biodiversity in India: Status, Issues and Challenges (eds Kaur, S. et al.) https://doi.org/10.1007/978-981-16-9777-7_7 (Springer, 2022).
Misra, A. K. Climate change impact, mitigation and adaptation strategies for agricultural and water resources. In Ganga Plain (India). Mitigation and Adaptation Strategies for Global Change, vol. 18, 673–689. https://doi.org/10.1007/s11027-012-9381-7 (2013).
Ganesh, S. R., Bhupathy, S., David, P., Sathishkumar, N. & Srinivas, G. Snake fauna of high wavy Mountains, Western Ghats, india: species richness, status, and distribution pattern. Russian J. Herpetology. 21 (1), 53–64 (2014).
Jhajharia, D. et al. Reference evapotranspiration under changing climate over the Thar desert in India. Meteorol. Appl. 22 (3), 425–435. https://doi.org/10.1002/met.1471 (2014).
Patra, A. et al. Conflict between sea snakes and humans on the Coast of West Bengal and Odisha, india: perception, attitudes and incidents. Trans. Royal Soc. Trop. Med. Hygiene. 119 (6), 671–678. https://doi.org/10.1093/trstmh/trae086 (2025).
Chatterjee, S. et al. Biodiversity Significance of North East India 71 (WWF-India, 2006).
Johnston, M. P. Secondary data analysis: A method of which the time has come. Qualitative Quant. Methods Libr. 3 (3), 619–626 (2014).
Geneviève, L. D. Participatory approaches and open data on venomous snakes: A neglected opportunity in the global snakebite crisis? PLoS Negl. Trop. Dis. 12 (3), e0006162. https://doi.org/10.1371/journal.pntd.0006162 (2018).
Chowdhury, S. et al. Increasing biodiversity knowledge through social media: A case study from tropical Bangladesh. BioScience 73 (6), 453–459. https://doi.org/10.1093/biosci/biad042 (2023).
Chowdhury, S. et al. Protected areas and the future of insect conservation. Trends Ecol. Evol. 38 (1), 85–95. https://doi.org/10.1016/j.tree.2023.04.006 (2023).
GBIF.org. GBIF Occurrence Download. https://doi.org/10.15468/dl.vd6h6h (2023).
Indiabiodiversity.org (11 January 2023). India Biodiversity Portal Occurrence Download. http://www.indiabiodiversity.org (2023).
iNatualist.org (11 January 2023). iNaturalist Occurrence Download. http://www.inaturalist.org (2023).
Vertnet.org (11 January 2023). Vertnet Portal Occurrence Download. http://www.vertnet.org (2023).
Reptile-database.reptarium.cz. Reptile-Database Portal. https://raptile-database.reptarium.cz (2023).
Pearson, R. G., Raxworthy, C. J. & Nakamura, M. Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J. Biogeogr. 34 (1), 102–117. https://doi.org/10.1111/j.1365-2699.2006.01594.x (2007).
van Proosdij, A. S., Sosef, M. S., Wieringa, J. J. & Raes, N. Minimum required number of specimen records to develop accurate species distribution models. Ecography 39 (6), 542–552. https://doi.org/10.5061/dryad.8sb8v (2016).
Feeley, K. J. & Silman, M. R. The data void in modeling current and future distributions of tropical species. Glob. Change Biol. 17 (1), 626–630. https://doi.org/10.1111/j.1365-2486.2010.02239.x (2010).
Barve, N. et al. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Model. 222 (11), 1810–1819. https://doi.org/10.1016/j.ecolmodel.2011.02.011 (2011).
Fick, S. E. & Hijmans, R. J. WorldClim 2: new 1km Spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37 (12), 4302–4315. https://doi.org/10.1002/joc.5086 (2017).
Moss, R. et al. The next generation of scenarios for climate change research and assessment. Nature 463, 747–756. https://doi.org/10.1038/nature08823 (2010).
Dormann, C. F. Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36 (1), 27–46. https://doi.org/10.1111/j.1600-0587.2012.07348.x (2013).
Hirzel, A. H., Le Lay, G., Helfer, V., Randin, C. & Guisan, A. Evaluating the ability of habitat suitability models to predict species presences. Ecol. Model. 199 (2), 142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017 (2006).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modelling of species geographic distributions. Ecol. Model. 190, 231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026 (2006).
Warren, D. L. & Seifert, S. N. Ecological niche modelling in maxent: the importance of model complexity and the performance of model selection criteria. Ecol. Appl. 21 (2), 335–342. https://doi.org/10.2307/29779663 (2011).
Javidan, N. et al. Evaluation of multi-hazard map produced using maxent machine learning technique. Sci. Rep. 11 (1), 6496. https://doi.org/10.1038/s41598-021-85862-7 (2021).
Baldwin, R. A. Use of maximum entropy modelling in wildlife research. Entropy 11 (4), 854–866. https://doi.org/10.3390/e11040854 (2009).
Phillips, S. J. & Dudík, M. Modeling of species distributions with maxent: new extensions and a comprehensive evaluation. Ecography 31, 161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x (2008).
Meza-Joya, F. L., Rojas-Morales, J. A. & Ramos, E. Predicting distributions of rare species: the case of the false coral snake Rhinobothryum bovallii (Serpentes: Colubridae). Phyllomedusa J. Herpetol. 19 (2), 141–164. https://doi.org/10.11606/issn.2316-9079.v19i2p141-164 (2020).
Muliya, S. K. et al. Addressing Wallacean shortfall using small sampling approach: a case study with endemic Lycodon flavicollis (Squamata: Colubridae) Mukherjee & Bhupathy, 2007. J. Asia-Pacific Biodivers. 14 (2), 159–168. https://doi.org/10.1016/j.japb.2020.12.005 (2021).
Bertelsmeier, C., Luque, G. M. & Courchamp, F. The impact of climate change changes over time. Biol. Conserv. 167, 107–115. https://doi.org/10.1016/j.biocon.2013.07.038 (2013).
Yousefi, M. et al. Identifying high snakebite risk area under climate change for community education and antivenom distribution. Sci. Rep. 13 (1), 8191. https://doi.org/10.1038/s41598-023-35314-1 (2023).
Li, W., Wang, Z., Ma, Z. & Tang, H. A regression model for the Spatial distribution of red-crown crane in Yancheng biosphere Reserve, China. Ecol. Model. 103, 115–121 (1997).
Manel, S., Dias, J. M., Buckton, S. T. & Ormerod, S. J. Alternative methods for predicting species distribution: an illustration with Himalayan river birds. J. Appl. Ecol. 36, 734–747 (1999).
Barbosa, A. M., Real, R., Olivero, J. & Vargas, J. M. Otter (Lutra lutra) distribution modeling at two resolution scales suited to conservation planning in the Iberian Peninsula. Biol. Conserv. 114, 337–387. https://doi.org/10.1016/S0006-3207(03)00066-1 (2003).
Peterson, A. T. et al. B. Future projections for Mexican faunas under global climate change scenarios. Nature 416 (6881), 626–629. https://doi.org/10.1038/416626a (2002).
Venne, S. & Currie, D. J. Can habitat suitability estimated from maxent predict colonisations and extinctions? Divers. Distrib. 27 (5), 873–886. https://doi.org/10.1111/ddi.13238 (2021).
Hu, J. & Jiang, Z. Climate change hastens the conservation urgency of an endangered ungulate. PLoS ONE 6 (8), e22873. https://doi.org/10.1371/journal.pone.0022873 (2011).
Yu, H. et al. Evaluating species richness, turnover, and range shifts under climate change for fluvial fishes in Northeastern and Midwestern USA. Ecol. Process. 14, 43. https://doi.org/10.1186/s13717-025-00612-1 (2025).
Araújo, S. C. M., Ceron, K. & Guedes, T. B. Use of Geospatial analyses to address snakebite hotspots in mid-northern Brazil—A direction to health planning in shortfall biodiversity knowledge areas. Toxicon 213, 43–51. https://doi.org/10.1016/j.toxicon.2022.03.012 (2022).
Government of India. Census of India 2001 (Office of Registrar General India, 2001).
Burnham, K. P. & Anderson, D. R. (eds) Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach (Springer, 2002).
Shine, R. & Brown, G. P. Adapting to the unpredictable: reproductive biology of vertebrates in the Australian wet-dry tropics. Philosophical Trans. Royal Soc. B. 363, 363–373. https://doi.org/10.1098/rstb.2007.2144 (2008).
Varikoden, H., Revadekar, J. V., Kuttippurath, J. & Babu, C. A. Contrasting trends in Southwest monsoon rainfall over the Western Ghats region of India. Clim. Dyn. 52 (7), 4557–4566. https://doi.org/10.1007/s00382-018-4397-7 (2019).
Freeman, B. G., Lee-Yaw, J. A., Sunday, J. M. & Hargreaves, A. L. Expanding, shifting and shrinking: the impact of global warming on species’ elevation distributions. Glob. Ecol. Biogeogr. 27 (11), 1268–1276. https://doi.org/10.1111/geb.12774 (2018).
Freeman, B. G., Scholer, M. N., Ruiz-Gutierrez, V. & Fitzpatrick, J. W. Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. PNAS 115, 11982–11987. https://doi.org/10.1073/pnas.1804224115 (2018).
Singh, D., Tsiang, M., Rajaratnam, B. & Differnbaugh, N. S. Observed changes in extreme wet and dry spells during the South Asian summer monsoon season. Nat. Clim. Change. 4, 456–461. https://doi.org/10.1038/nclimate2208 (2014).
Mishra, V., Thirumalai, K., Singh, D. & Aadhar, S. Future exacerbation of hot and dry summer monsoon extremes in India. NPJ Clim. Atmospheric Sci. 3, 10. https://doi.org/10.1038/s41612-020-0113-5 (2020).
Acharya, B. K. & Chettri, B. Effect of climate change on birds, herpetofauna and butterflies in Sikkim himalaya: a preliminary investigation. In Climate Change in Sikkim: Patterns, Impacts and Initiatives (eds Arawatia, M. L. & Tambe, S.) 141–160 (Information and Public Relations Department, 2012).
Hulbert, A. C., Hall, J. M., Mitchell, T. S. & Warner, D. A. Use of human-made structures facilitates persistence of a non-native ectotherm. Biol. Invasions. 22, 2017–2031. https://doi.org/10.1007/s10530-020-02236-2 (2020).
Pellerin, F., Cote, J., Bestion, E. & Aguilée, R. Matching habitat choice promotes species persistence under climate change. Oikos 128 (2), 221–234. https://doi.org/10.1111/oik.05309 (2019).
Goldstein, E. et al. Climate change maladaptation for health: agricultural practice against shifting seasonal rainfall affects snakebite risk for farmers in the tropics. iScience 26 (2), 105946. https://doi.org/10.1016/j.isci.2023.105946 (2023).
Lenoir, J. & Svenning, J. C. Latitudinal and elevational range shifts under contemporary climate change. In Encyclopedia of Biodiversity (ed. Levin, S.) 690–709. https://doi.org/10.1016/B978-0-12-384719-5.00375-0 (Academic Press, 2024).
Moreno-Rueda, G. R. E. G., O. R. I. O., Pleguezuelos, J. M., Pizarro, M. & Montori, A. Northward shifts of the distributions of Spanish reptiles in association with climate change. Conserv. Biol. 26 (2), 278–283. https://doi.org/10.1111/j.1523-1739.2011.01793.x (2011).
Walther, G. R. et al. Ecological responses to recent climate change. Nature 416, 389–395. https://doi.org/10.1038/416389a (2002).
Alatawi, A. S., Gilbert, F. & Reader, T. Modelling terrestrial reptile species richness, distributions and habitat suitability in Saudi Arabia. J. Arid Environ. 178, 104153. https://doi.org/10.1016/j.jaridenv.2020.104153 (2020).
Ventura-Rojas, P. D., González-Romero, A., Moreno, C. E. & Sosa, V. J. Effect of rainfall, temperature and climate change on the ecology of the rodents of arid zones: a review. Mammal Rev. 55 (2), e12372. https://doi.org/10.1111/mam.12372 (2025).
Arias-Rodríguez, J. & Gutiérrez, J. M. Circumstances and consequences of snakebite envenomings: a qualitative study in south-eastern Costa Rica. Toxins 12 (1), 45. https://doi.org/10.3390/toxins12010045 (2020).
Gutiérrez, J. M. et al. The need for full integration of snakebite envenoming within a global strategy to combat the neglected tropical diseases: the way forward. PLoS Negl. Trop. Dis. 7 (6), e2162. https://doi.org/10.1371/journal.pntd.0002162 (2013).
Williams, D. J. et al. Strategy for a globally coordinated response to a priority neglected tropical disease: snakebite envenoming. PLoS Neglected Trop. Disease. 13 (2), e0007059. https://doi.org/10.1371/journal.pntd.0007059 (2019).
Garai, N., Roy, A. & Pramanick, K. Unravelling sustainable development at the sub-national scale in India. Environ. Dev. 57, 101336. https://doi.org/10.1016/j.envdev.2025.101336 (2026).
Nolan, V., Gilbert, F. & Reader, T. Solving sampling bias problems in presence-absence or presence-only species data using zero-inflated models. J. Biogeogr. 49 (1), 215–232. https://doi.org/10.1111/jbi.14268 (2021).
Phillips, S. J. et al. Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol. Appl. 19 (1), 181–197. https://doi.org/10.1890/07-2153.1 (2009).
Pearson, R. G. & Dawson, T. P. Predicting the impacts of climate change on the distribution of species: are bioclimatic envelope models useful? Glob. Ecol. Biogeogr. 12 (5), 361–371. https://doi.org/10.1046/j.1466-822X.2003.00042.x (2003).
Araújo, M. B., Pearson, R. G. & Rahbek, C. Equilibrium of species’ distributions with climate. Ecography 28 (5), 693–695 (2005).
Inman, R., Esque, T. C. & Nussear, K. E. Dispersal limitations increase vulnerability under climate change for reptiles and amphibians in the Southwestern united States. J. Wildl. Manage. 87 (1), e22317. https://doi.org/10.1002/jwmg.22317 (2022).
IUCN Red List. The IUCN Red List of Threatened Species. http://www.iucnredlist.org (Accessed 13 February 2024) (2024).
WPA 1972 Amendment. The Wildlife (Protection) Amendment Act, 2022. No. 18 of 2022: The Gazette of India 165 (2022).
Acknowledgements
Authors thank citizen science efforts, such as GBIF, VertNet, iNaturalist, Indian Biodiversity Portal and Reptile Database. Also, authors thank Jason D. Gerard for help in identifying the distribution polygons for the species. Authors acknowledge guidance from Subject experts. This manuscript has the technical contribution number 7438 of the Clemson University Experiment Station. Authors thank all post contributors and reviewers from the Snakes of India and Indian Snakes facebook groups, and express their gratitude to the group administrators, Jose Louies and Gnaneswar Ch, for their permission to utilize the information.
Author information
Authors and Affiliations
Contributions
Conceptualization: SK, AKJ, CPS, ASData collection and curation: SK, AKJ, SSPC, DKJData analysis: SK, AKJManuscript writing: AKJ, SSPC, DKJ, ASManuscript review and editing: SK, AKJ, SSPC, DKJ, AS, JTA, CPS.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Khandve, S., Jangid, A.K., Sreelekha Suresh, P.C. et al. Climate change triggering shifts in venomous snakes hotspots and snakebite risk in India. Sci Rep 15, 40983 (2025). https://doi.org/10.1038/s41598-025-24731-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-24731-z
