
Abstract
Diabetes-associated cognitive dysfunction represents a global health challenge, yet the mechanisms by which anesthetics modulate cognitive function in diabetic states remain poorly understood. We systematically compared the effects of 2-hour brief exposure to sevoflurane (SEV) and propofol (PRO) on cognitive function and neuropathology in streptozotocin (STZ) -induced diabetic mice. Morris water maze and Y-maze tests revealed that SEV significantly exacerbated spatial memory and learning deficits in mice, while PRO showed no significant effects. Additionally, diabetic mice exhibited reduced NeuN+ neurons, increased β-amyloid deposition, and decreased SYN expression in the hippocampal CA1 region as examined by Immuno-fluorescence staining. Neither short-term SEV nor PRO exposure aggravated neuronal structural damage. Further transcriptomics revealed both anesthetics affected hippocampal neuron differentiation, but SEV uniquely perturbed fatty acid metabolism pathways. Metabolomics identified SEV-induced disruptions in lipid metabolism, marked by elevated hippocampal free fatty acids, phospholipids, as well as reduced lysophospholipids and acylcarnitine. Integrated multi-omics analysis demonstrated that SEV impaired cognition by suppressing fatty acid oxidation and dysregulating glycerophospholipid metabolism. These findings highlight the critical impact of anesthetic selection in diabetic populations.
Similar content being viewed by others
Introduction
Diabetes mellitus (DM), a global metabolic disorder, has seen its complications extend beyond traditional cardiovascular and renal pathologies to encompass central nervous system impairments. Emerging evidence indicates that 45% of patients with type 2 diabetes (T2DM) exhibit mild cognitive impairment (MCI)1, with dementia risk elevated by about two-fold compared to non-diabetic populations2. Similarly, individuals with type 1 diabetes (T1DM) face accelerated cognitive decline, particularly under conditions of poor glycemic control or prolonged disease duration. The pathogenesis of diabetes-associated cognitive dysfunction involves multifaceted interactions, including cerebral insulin resistance, impaired synaptic plasticity, neuroinflammation and oxidative stress evoked by advanced glycation end products (AGEs), and blood-brain barrier hyperpermeability induced by microvascular lesion3,4,5. Notably, diabetes not only accelerates Alzheimer’s disease (AD)-like pathologies, such as β-amyloid (Aβ) deposition, but also drives neurodegeneration through distinct metabolic pathways independent of AD hallmarks6.
Perioperative anesthesia is a common clinical scenario for patients, which presents underappreciated risks for cognitive outcomes7,8. Commonly used inhalational anesthetics, such as sevoflurane (SEV), and intravenous anesthetics, like propofol (PRO), exhibit varying effects on cognitive function. SEV, despite their hemodynamic stability9, raise concerns due to preclinical evidence that SEV induced hypotension, decreased cerebral blood flow, and hippocampal inflammation. SEV evoked postoperative cognitive decline through multiple mechanism, including promoting neuronal ferroptosis10, activating NLRP3 inflammasome11 and tau phosphorylation12. Compared to SEV, PRO seems to be safer regarding the incidence of postoperative neurocognitive impairment, especially for the elders13,14,15. However, some studies have reported that PRO anesthesia is associated with a higher incidence of postoperative cognitive dysfunction compared to SEV anesthesia16,17. These conflicting findings may be related to variations in patient characteristics, surgical procedures, and anesthesia duration across studies. In preclinical studies, PRO serves as an intravenous anesthetic with antioxidative properties, which has been reported to mitigate neural injury and attenuate cognitive function18,19,20,21.
Current preclinical research predominantly focuses on neurotoxicity in healthy or AD models, largely overlooking DM-specific contexts12,22. Recent clinical studies have reported increased susceptibility to postoperative delirium and long-term cognitive deficits in DM populations, potentially linked to glycemic fluctuations, oxidative stress, and anesthetic neurotoxicity23,24,25. Animal studies suggest that while brief SEV exposure in healthy subjects induces negligible cognitive effects, it exacerbates hippocampal neuroinflammation and synaptic dysfunction in DM models via glial cell activation and insulin-like growth factor 1 (IGF1) signaling disruption26,27. Though PRO seems to be beneficial for DM complications, conflicting evidence suggests an exacerbated insulin resistance after exposure with PRO28. These discrepancies highlight context-dependent anesthetic effects modulated by metabolic status, exposure duration, and individual susceptibility, underscoring the need for mechanistic clarity to guide clinical practice.
In this study, we hypothesized that SEV and PRO, with different mechanisms, exert distinct effects on cognitive function in streptozotocin (STZ)-induced T1DM mice. Integrated behavioral, histological, hippocampal transcriptomic, and untargeted metabolomic analyses further revealed that the stronger pro-cognitive-impairment effect of SEV is associated with its disruption of fatty acid and phospholipid metabolism.
Results
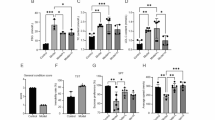
SEV, but not PRO, exacerbates cognitive deficits in STZ-Induced diabetic mice
As shown in Fig. 1B, administration of STZ led to a significant increase in fast blood glucose levels, confirming a T1DM phenotype. Both PRO and SEV exposure did not lead to a significant change in blood glucose. The body weight of STZ-treated mice was significantly reduced (before anesthesia: P = 0.040, endpoint: P < 0.001), while exposure to both anesthetics showed no obvious effect on their weight (Figure S1A). In Morris water maze test (Fig. 1C), all groups demonstrated progressive reductions in escape latency during trial period. However, SEV-exposed DM mice displayed prolonged latency compared to the mice in control (CON) group by the second trial day (2nd P = 0.036, 3rd −5th P < 0.001), with all DM groups showing consistently longer latencies than corresponding CON from the third day onward (P < 0.001). On the last two days during trial period, SEV-treated DM mice exhibited further latency increases relative to untreated DM mice (4th P = 0.024, 5th P = 0.041). During the test period, the number of entries (Fig. 1D) (P = 0.0307) and residence time in the quadrant where the platform was located in the MWM (Fig. 1E) (P = 0.0108) were significantly lower in all three groups of DM model mice compared with the CON mice. And the reduction was more pronounced in the SEV-anesthetized DM mouse model (vs. CON, P < 0.001, P < 0.001), whose target zone duration was shorter than that of the unanesthetized DM mice (mean rank: 4.17 vs. 13.00). In Y maze test, STZ treatment and anesthetic exposure appeared to have no significant effect on the total number of arm entries in mice (Fig. S1B). DM mice showed significant reduced spontaneous alternation rates (Fig. 1F) (P < 0.001), which were further diminished by SEV anesthesia (mean: 28.32 vs. 38.76) without a statistical significance. As for PRO, its exposure did not produce any significant effect on the performance of DM mice in the MWM and Y maze test. Meanwhile, PRO-treated DM mice showed no additional behavioral change in MWM and Y maze test compared to the untreated DM mice. Notably, neither SEV nor PRO altered glucose levels or cognitive performance in non-DM mice. These findings demonstrate that STZ-induced T1DM mice develop cognitive deficits, which are selectively exacerbated by SEV but not PRO.

Effects of anesthetics on learning and memory in T1DM mice induced by STZ. (A) Schematic diagram of the experimental timeline. (B) Blood glucose levels at baseline, post-STZ induction, and post-anesthesia (n = 10). (C-E) Mice performance in Morris water maze test (n = 6). (C) Escape latency during training days, (D) number of platform crossings, and (E) time spent in the target quadrant during the probe test. (F) Spontaneous alternation rates in the Y-maze (n = 6). *P < 0.05, **P < 0.01, ***P < 0.001 vs CON group; #P < 0.05 vs DM group; @P < 0.05, @@P < 0.01, @@@P < 0.001 vs. DM + PRO group; &&&P < 0.001 vs. DM + SEV group. Data are mean ± SEM. P values were determined by two-way ANOVA followed by Bonferroni’ s multiple comparisons (C, D, F) or Kruskal-Wallis test (B, E)
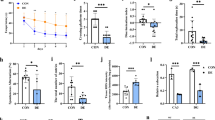
Neither short-term SEV nor PRO exposure significantly affect diabetes-associated hippocampal lesion
To investigate the effects of anesthetics on hippocampal structure and neuron damage, IF staining was performed to assess mature neurons (NeuN), synaptic integrity (SYN), and Aβ1−42 deposition. As shown in Fig. 2A, DM mice exhibited a significant reduction in NeuN+ neurons within the hippocampal CA1 region compared to CON mice (P = 0.0127), with disorganized neuronal arrangement. DM mice exposed to anesthetics did not show a further marked reduction in NeuN+ neurons compared to untreated DM mice. While SEV-treated DM mice showed a trend toward greater neuronal loss (mean: 0.409 vs. 0.622). Aβ1−42 deposition in the hippocampal dentate gyrus was notably elevated in DM mice (P < 0.001), though anesthetic exposure did not amplify this effect (Fig. 2B). SYN displayed reduced expression (P < 0.001) and disrupted spatial organization in hippocampus tissue of DM mice compared to CON mice, transitioning from dense clusters to sparse, fragmented patterns (Fig. 2C). Neither SEV nor PRO administration significantly altered SYN expression levels or distribution. These findings corroborate the association between DM-induced cognitive decline and neuronal/synaptic degeneration, while indicating that short-term, single-dose anesthetic exposure does not exacerbate structural damage in the hippocampus under these experimental conditions.

Effects of anesthetics on hippocampal neuronal pathology in T1DM mice.(A-C) Representative immunofluorescence images and semi-quantitative analysis of (A) NeuN-, (B) Aβ1−42-, and (C) SYN-positive area in the hippocampal region (n = 6). Scale bars, 50 µm (NeuN, SYN) or 100 µm (Aβ1−42). *P < 0.05, **P < 0.01, ***P < 0.001 vs. CON group; @@P < 0.01, @@@P < 0.001 vs. DM + PRO group; &&&P < 0.001 vs. DM + SEV group. Data are mean ± SEM. P values were determined by two-way ANOVA followed by Bonferroni’ s multiple comparisons
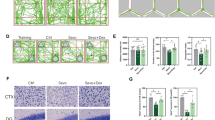
SEV and PRO anesthesia elicit divergent hippocampal gene expression profiles in diabetic mice
To compare the impacts of PRO and SEV on hippocampal gene expression in DM mice, RNA sequencing was performed on hippocampal tissues from CON, DM, and DM mice anesthetized by PRO or SEV. Principal component analysis (PCA) revealed distinct transcriptional profiles between CON and DM groups, with further divergence observed in PRO- or SEV-treated DM mice compared to untreated DM counterparts (Fig. 3A). Differentially expressed genes (DEGs) were identified using thresholds of |fold change| >1.5 and adjusted P < 0.05. Comparative analysis demonstrated that PRO-treated DM mice exhibited the highest number of hippocampal DEGs relative to untreated DM mice (Fig. 3B), whereas SEV treatment induced more uniform transcriptional shifts with pronounced polarization between upregulated and downregulated genes. Heatmap visualization of 822 DEGs between DM and DM + PRO groups highlighted distinct expression patterns across samples (Fig. 3C). Gene Ontology (GO) enrichment analysis revealed that PRO anesthesia predominantly modulated biological processes such as “neuron differentiation” and “regulation of ion transmembrane transport” in DM hippocampi (Fig. 3D). Similarly, analysis of 503 DEGs between DM and DM + SEV groups (Fig. 3E) identified enrichment in “neuron differentiation,” “hindbrain development,” and “cellular response to retinoic acid,” with SEV exerting stronger regulatory effects on “neuron differentiation” compared to PRO. Notably, Notably, in addition to the aforementioned biological processes, SEV induced significant alterations in hippocampal tissue lipid metabolism-related “fatty acid beta-oxidation”, “fatty acid catabolic process,” and “regulation of fatty acid oxidation” in DM mice(Fig. 3F). This suggests that SEV-induced cognitive impairment may be associated with disrupted central lipid metabolism, which might represent a regulatory mechanism distinct from PRO. These findings suggest that both anesthetics influence hippocampal neuronal differentiation, but the more pronounced transcriptional impact of SEV may underlie its exacerbation of DM-associated cognitive impairment.

Impacts of SEV and PRO on hippocampal gene expression in T1DM mice. (A-F) Transcriptomic analysis of hippocampus from mice (n = 3). (A) Principal component analysis (PCA) of hippocampal gene expression profiles. (B) Number of DEGs across comparisons. (C-D) (C) Heatmap and (D) Gene Ontology (GO) enrichment analysis of 602 DEGs between DM and DM + PRO groups. (E-F) (E) Heatmap and (F) GO enrichment analysis of 503 DEGs between DM and DM + SEV groups
SEV disturbed hippocampal metabolic profiles in diabetic mice associated to compromised lipid disposal
To validate the impact of SEV on hippocampal fatty acid metabolism identified in the transcriptomic analysis, we examined the effects of SEV and PRO treatment on the expression of fatty acid metabolism-related genes in DM mice. As shown in Fig. 4A, SEV treatment downregulated the transcription of genes involved in fatty acid oxidation, including Cpt1a (P = 0.0608), and Hadh (P = 0.0509), as well as their positive regulators Prkag1 (P = 0.0973), and Prkag2 (P = 0.0941). Conversely, expression of genes that inhibit fatty acid oxidation, including Mlycd (P = 0.0509), Acaca (P = 0.0014), and Acacb (P = 0.0144), was markedly upregulated. In contrast, PRO treatment exerted no significant effects on the expression of these fatty acid metabolism-related genes. To further verify SEV-induced perturbations in fatty acid metabolic homeostasis, we conducted metabolomic profiling of hippocampal tissues from DM mice with or without SEV treatment, identifying 172 differentially abundant metabolites. The volcano plot (Fig. 4B) revealed that SEV treatment significantly elevated metabolites including myristic acid, palmitic acid, and multiple phospholipids, while reducing levels of acetylcarnitine, dodecanoylcarnitine, arachidonic acid, and several lysophospholipids. KEGG enrichment analysis (Fig. 4C) of differential metabolites demonstrated that SEV treatment robustly disrupted hippocampal metabolic pathways, including “glycerophospholipid metabolism”, “glycerolipid metabolism”, “fatty acid metabolism”, and “biosynthesis of unsaturated fatty acids”. Collectively, these resulting s corroborate the metabolomic findings and confirm that SEV treatment induces alterations in hippocampal fatty acid metabolism and compromises phospholipid metabolism homeostasis (Fig. 4D).

Metabolic effects of SEV on hippocampal profiles in T1DM mice. (A) mRNA levels of genes related to fatty acid metabolism in mouse hippocampus (n = 3). (B and C) Metabolomics analysis of hippocampus from mice (n = 6). (B) Volcano plot illustrating the differential abundant metabolites. (C) KEGG pathway enrichment analysis of differential metabolites between DM and DM + SEV groups. (D) Alteration of lipid metabolism pathway induced by SEV. *P < 0.05, **P < 0.01. Data are mean ± SEM. P values were determined by one-way ANOVA followed by Bonferroni’ s multiple comparisons
Discussion
Anesthetics have been reported to be a double-edged sword exerting beneficial or harmful effects on cognitive function. In specific state with existing cognitive impairment, such as DM-associated cognitive decline and AD, the effects of anesthetics require extra attention29,30. This study reveals that SEV exacerbates cognitive impairment in STZ-induced T1DM mice, while PRO does not exhibit similar effects. Behavioral tests demonstrated that SEV anesthesia aggravated spatial memory deficits in T1DM mice, consistent with prior findings in T2DM models where SEV exposure worsened cognitive decline via hippocampal neuroinflammation26. However, SEV exposure in the Y-maze test showed a trend toward reduced working memory in DM mice, though it was not statistically significant. Previous literature has reported that SEV exposure can impair working memory in type 2 diabetic rats31. Therefore, we speculate that the lack of a significant effect observed in this study might be related to the limited sample size.
The absence of cognitive deterioration in PRO-treated T1DM mice observed in this study may be attributed to its unique pharmacological properties. A clinical trial comparing PRO and SEV in elderly DM patients reported fewer perioperative cardiovascular events and hemodynamic perturbations with PRO, potentially leading milder central nervous system impact32. Furthermore, preclinical evidence highlights PRO’ s antioxidant and anti-inflammatory capacities. PRO treatment can mitigate oxidative stress and neuroinflammation in animal models of neurodegeneration, thereby preserving neuronal integrity and cognitive function19,33. These intrinsic neuroprotective properties, position PRO as a safer anesthetic choice for populations with preexisting metabolic and cognitive vulnerabilities. Furthermore, this study found that SEV exposure exhibited a tendency to elevate blood glucose levels in DM mice, though not significantly. However, previous literature has reported the phenomenon of elevated blood glucose in both surgical patients and animals under SEV anesthesia34,35. These findings suggest the potential adverse effects of SEV on glycemic control, indicating that its use in diabetic patients should be approached with caution.
This present study observed neuronal damage in the hippocampi of T1DM model mice, characterized by reduced NeuN+ neuron counts, diminished SYN expression, and Aβ deposition. However, short-term, single-dose anesthetic exposure did not induce significant structural alterations in hippocampal cellular architecture. This finding contrasts with prior studies reporting that SEV exacerbates hippocampal neuroinflammation in DM states26, leading to neuronal apoptosis and synaptic dysfunction. The discrepancy may stem from differences in exposure duration and dosage, since we employed brief anesthetic exposure below the threshold required to trigger overt neuronal injury. Continuous or high-concentration SEV exposure has been shown to elevate hippocampal ROS levels, impair mitochondrial function, suppress BDNF signaling, and reduce dendritic spine density36,37,38. The absence of such structural changes in our study suggests that short-term SEV exposure primarily induces functional synaptic disruption rather than irreversible neurodegeneration. This implies that cognitive deficits mediated by SEV may arise from early-stage functional or metabolic perturbations, preceding detectable histopathological damage. Therefore, it is necessary to evaluate anesthetic neurotoxicity through functional assays in addition to conventional morphological analyses.
Integrating transcriptomic and metabolomic analysis, our results further reveal a novel pathway involving dysregulated neuronal differentiation and lipid metabolism. This finding contrasts with earlier studies which focused on microglial activation and pro-inflammatory cytokine release. RNA-Seq results indicated that exposure to both anesthetics significantly affected the biological process of “neuron differentiation,” which is consistent with previous findings. SEV has been shown to suppress CREB transcription by upregulating HDAC9, thereby reducing neurotrophin-3 levels and impairing adult hippocampal neurogenesis39. Moreover, non‑coding RNAs such as Rik‑203 have been reported to mediate SEV’s effects on neural progenitor cells by modulating downstream GSK‑3β levels40. In contrast, the impact of PRO on neuronal differentiation is closely associated with exposure dosage. Studies have reported that a low dose (10 µM) promotes neuronal differentiation of neural progenitor cells, whereas a high concentration (200 µM) inhibits neuronal differentiation and drives cells toward a glial fate41. This shift is related to high‑dose‑induced autophagy and intracellular calcium overload. Correspondingly, it has also been indicated that clinical‑dose PRO increases neuronal fate choice via a non‑γ‑aminobutyric acid type A mechanism42. These findings suggest that, in terms of influencing neuronal differentiation, PRO appears to be relatively safer for clinical use. Among the biological processes significantly affected by exposure to these anesthetics, SEV exhibits a particularly notable regulatory effect on fatty acid metabolism at the transcriptional level. This finding prompts us to further investigate its impact on hippocampal lipid homeostasis through a multi-omics approach.
Our present study identified a significant alternation of fatty acid and phospholipid metabolism in T1DM mice treated with SEV. Numerous studies have suggested a closely association between cognitive dysfunction and abnormal central lipid metabolism. In aged-relative cognitive deficit rats, amount of polyunsaturated fatty acids (PUFAs) in phospholipids decreases while the amount of oleic acid and stearic acid elevates43. Similarly, our study revealed that the majority of elevated phospholipids in the hippocampus of SEV-exposed mice predominantly contained oleoyl or stearoyl acyl chains. Corresponding to the observed accumulation of fatty acids, RNA-Seq results also indicated that SEV suppresses fatty acid beta‑oxidation. This effect was likewise observed in non‑DM mice treated with SEV, revealing a neurotoxic mechanism whereby the inhibition of fatty acid oxidation FAO in neural stem/progenitor cells impairs neurogenesis44. Conversely, the abundance of PUFAs and lysophospholipids containing PUFAs, such as docosahexaenoic acid and arachidonic acid, was reduced. Furthermore, previous literature reports an association between decreased cerebrospinal fluid levels of lysophosphatidic acid (LPA) 22:6 and major depressive disorder, indicating significant role in central nervous system45. LPA activates receptors (LPARs) that support cognitive function46,47. The observed decrease in LPA 22:6 may therefore be a factor by which SEV worsens cognitive impairment in T1DM mice. As storage reservoirs for fatty acids, relative proportions between phospholipids and lysophospholipids reflect the metabolic status of fatty acids. Our findings of increased phospholipid content alongside decreased lysophospholipid content in SEV-anesthetized DM mice potentially indicate reduced fatty acid utilization in hippocampus. This interpretation is further supported by elevated levels of saturated fatty acids, including myristic acid and palmitic acid, and a concomitant decrease in the abundance of acylcarnitines, which are intermediate metabolites of beta-oxidation. These data collectively demonstrate abnormal fatty acid catabolism induced by SEV exposure. Notably, prior studies have documented pre-existing metabolic alterations in the brain tissue of DM animal models characterized by elevated fatty acid levels and reduced acylcarnitines48. This suggests that SEV treatment may exacerbate underlying lipid metabolic dysregulation, thereby aggravating DM-associated cognitive dysfunction. Medicines that promote fatty acid oxidation, such as fenofibrate, a PPARα agonist49,50, and metformin, an AMPK activator51,52, have demonstrated efficacy in ameliorating cognitive impairment in animal models. Considering the established clinical use of these metabolic modulators in DM patients, their therapeutic potential in preventing or treating anesthesia-associated cognitive dysfunction in this population warrants further investigation.
Our study integrates phenotypic and molecular evidence to clarify how SEV promotes cognitive impairment under DM conditions. However, several limitations of this study should be noted. The modest sample size, potential gender bias in animal selection, and the use of a single exposure duration constrain the generalizability of the findings. Future studies with more comprehensive designs are needed to validate these results and explore the underlying mechanisms in greater depth. Furthermore, our lipid metabolism analysis focused only on the DM state. It therefore remains unclear whether the effects of SEV are independent of DM, especially since SEV has been reported to disrupt central fatty acid metabolism even in non-diabetic models44,53. For investigating mechanism, a determination on lipid metabolism-related protein should been encompassed in the future research, which were not addressed in the present study. Additionally, we did not perform direct neurofunctional assessments, which limits the ability to establish clear links between the observed phenotypic and molecular changes. Further research should investigate how SEV-induced disturbances in lipid metabolism affect learning and memory, for example by examining pathways involving neuroinflammation, synaptic plasticity, or neuronal electrophysiology.
In summary, we elucidate the differential effects and mechanisms of SEV and PRO on cognitive function in T1DM mice. Multi-omics profiling uncovers disruptions in lipid metabolism and neuronal differentiation. These findings provide a new perspective on anesthetic exposure in diabetic cognitive impairment and enrich the mechanistic framework of neurotoxicity. Beyond theory, they highlight potential targets for future validation and inform strategies to optimize perioperative care. Translating these insights into clinical practice, however, requires further experimental and clinical investigation.
Methods
Animals
Six-week-old male C57BL/6 mice were obtained from Charles River Experimental Animal Co., Ltd (Beijing, China) and housed in a specific pathogen-free (SPF) facility with a temperature-controlled environment at 22–24 °C and a 12-hour light/dark cycle. All animal experiments were approved by the Animal Care and Use Committee of the School of Basic Medical Sciences, Jilin University and were conducted in accordance with the Guide for the Care and Use of Laboratory Animals (8th edition)54.
Experimental protocols
After a one-week acclimation period, the fifty mice were randomly divided into two groups based on body weight. DM group mice received intraperitoneal injections of 50 mg/kg STZ (dissolved in 0.1 M citrate buffer, pH 4.5) for five consecutive days to induce T1DM55, while CON group mice were injected with citrate buffer alone. On day 6, fasting blood glucose levels were measured in DM mice, and animals with levels > 11.0 mmol/L were considered successful T1DM models for subsequent experiments. Two weeks post-induction, CON mice were randomly allocated into three subgroups: CON (no treatment), CON + PRO, and CON + SEV. Similarly, DM mice were divided into DM (no treatment), DM + PRO, and DM + SEV groups. The CON + PRO and DM + PRO groups received continuous intraperitoneal PRO infusion (50 mg/kg/h) for anesthesia induction, whereas CON + SEV and DM + SEV groups underwent isoflurane inhalation (3% induction, 1.5% maintenance in 30% O₂/70% N₂). The doses of SEV56 and PRO57,58 were based on those reported in previous literature to induce cognitive impairment in animals. All anesthetic exposures lasted 2 h. After discontinuing SEV exposure, mice were placed in a chamber receiving a gas mixture of 70% air and 30% oxygen. Recovery of the animals was determined by observing their ability to resume normal, spontaneous locomotor activity. Following recovery, the mice were immediately transferred to the behavioral testing room and allowed to acclimate to the environment for 2 h before behavioral testing. Mice were euthanized with carbon dioxide 1 h after completing the behavioral tests on day 6. After collecting blood through the orbital venous plexus, systemic blood vessels of the mice were perfused with saline via the heart. After opening the skull, the whole brain was removed and bisected along the midline. It was then either directly fixed in 4% paraformaldehyde (PFA) solution, or the hippocampus was further dissected and preserved in RNA-later, or snap-frozen in liquid nitrogen and stored at −80 °C for subsequent experiments. The experimental scheme is summarized in Fig. 1A. Researchers involved in animal grouping do not engage in data collection and analysis.
Blood glucose test
Glucose levels in serum were tested using Blood Glucose Content Assay Kit (BC2495, Solarbio) following the manufacturer’s protocol.
Morris water maze (MWM) test
The experimental procedure was conducted as described previously59, including training trials and a probe test. All tests were conducted between 10:00 and 16:00. The MWM test was performed in a circular opaque pool measuring 120 cm in diameter and 40 cm in depth. The water temperature was maintained constant at 23 ± 1 °C. The pool was virtually divided into four equal quadrants. A circular platform (7.5 cm in diameter) was placed in one of the quadrants, designated as the target quadrant, submerged approximately 1 cm below the water surface. A video camera was mounted about 80 cm above the center of the pool to track the trajectory of the mice. Briefly, from the 1 st day to the 5th day, a platform was positioned in a fixed location within the target quadrant, and all mice were trained once daily. Both escape latency and swimming path were recorded. On the 6th day, the platform was removed, and the time spent in the target quadrant and the number of platform crossings were monitored for 60 s. The same experimenters performed all procedures throughout the MWM test.
Y maze test
As described previously60, the Y-maze consisted of three identical arms (each 35 cm long with 15 cm high walls) arranged at 120° angles, and was constructed from opaque white plastic. A removable partition was inserted at the entrance of each arm to restrict the mouse’s movement before the trial began. Mice were placed in a fixed arm, and after removing the partitions, the sequence of mouse entry into different arms was recorded over 5 min via a camera mounted 50 cm above the apparatus. The spontaneous alternation is defined as consecutive entries into three different arms, and spontaneous alternation rate was calculated according to previous studies60. To eliminate odor interference between trials, the interior of the maze was wiped with 75% ethanol after each mouse test. All tests were conducted between 10:00 and 16:00.
Immunofluorescence
After 2-day incubation in PFA, brain tissue was dehydrated through a graded ethanol series, cleared in xylene, embedded in paraffin, and sectioned into 5-µm slices. Next, the sections were deparaffinized in xylene and rehydrated through a graded ethanol series. Antigen retrieval was performed by incubating sections in citrate buffer (pH 6.0) at 95 °C for 20 min. After cooling to room temperature, sections were permeabilized with 0.3% Triton X-100 in PBS for 15 min and blocked with 5% normal goat serum for 1 h at RT. Primary antibodies against NeuN (1:300, Abcam, ab177487), Aβ1–42 (1:500, CST, # 14974), and synaptophysin (SYN, 1:300, Abcam, ab 32127) were applied separately and incubated overnight at 4 °C. After washing with PBST (0.1% Tween-20), sections were incubated with Alexa Fluor 594-conjugated goat anti-rabbit IgG secondary antibody (1:1000, Invitrogen, R37117) for 2 h at RT in the dark. Nuclei were counterstained with DAPI (Sigma-Aldrich, St. Louis, MO, USA) for 5 min. Sections were visualized under a light microscope (Ni-U, Nikon, Shanghai, China). Fluorescence intensity was quantified with ImageJ software (v 1.53c).
RNA-seq
Hippocampal tissues from mice were harvested and immediately preserved in RNAlater (Vazyme, Nanjing, China). 2–3 biological replicates were pooled to create one sample for tests, resulting in 3 pooled samples per group for these assays. Total RNA extraction and subsequent RNA sequencing were conducted by Wekomo (Shenzhen, China). Sequencing libraries were prepared using the NEBNext® Ultra™ RNA Library Prep Kit for Illumina® (NEB, MA, USA) according to the manufacturer’s protocol. Paired-end sequencing (150-bp reads) was performed on an Illumina NovaSeq™ 6000 platform (Illumina, CA, USA). Raw sequencing data were processed using R (v 4.2.0). Sequence quality was assessed, and low-quality reads were filtered using FastQC (v 0.1.3). Reads were then aligned to the reference genome using HISAT2 (v 1.24.0). Transcript assembly and quantification of gene-level read counts were performed using Stringtie (v 2.2.3). Differential gene expression analysis, including data normalization, was conducted using DESeq2 (v 1.46.0). Functional enrichment analysis was performed using ClusterProfiler (v 4.12.6). Data visualization was implemented using ggplot2 (v 3.5.1).
Untargeted metabolomic analysis
To characterize the metabolites in hippocampus, UPLC-MS/MS analysis was performed on a Vanquish UHPLC System (Thermo Fisher Scientific, USA) equipped with an ACQUITY UPLC HSS T3 column (2.1 × 100 mm, 1.8 μm) (Waters, Milford, MA, USA) and a coupled Q Exactive Focus Orbitrap mass spectrometer (Thermo Fisher Scientific, USA). The column was maintained at 40 ℃ and injection volume was set at 2 µL. For the positive ion mode, the mobile phases consisted of (A) 0.1% (v/v) formic acid aqueous solution and (B) 0.1% formic acid in acetonitrile. For the negative ion mode, the mobile phases consisted of (A) 10 mM ammonium formate aqueous solution and (B) acetonitrile. The gradient was set as follows: 0–1 min, 5% B; 1–9 min, 5%−98% B; 9–10 min, 98% B; 10–10.5 min, 98%−5% B; 10.5–15 min, 5% B. The flow rate of mobile phases was set at 0.3 mL/min. Mass spectrometry data were acquired using full scan accompanied by a data-dependent MS2 mode for precursor and fragment ion detection. Analyses were performed in both positive and negative ionization modes with spray voltages of + 3.50 kV and − 2.50 kV. Scan ranges were set to m/z 100–1000 for MS1 and m/z 50–1000 for MS2.
Metabolites in hippocampus were extracted with a pre-cooled mixed reagent (acetonitrile: methanol: water = 9:4:2). Briefly, frozen tissues were homogenized with the reagent at a ratio of 100 µL per 10 mg of tissue at −20 °C. After standing for 30 min, the mixture was centrifuged at 4 °C and 12, 000 rpm for 15 min, and the supernatant was collected for analysis.
Raw data files were converted to mzXML format using MSConvert (https://proteowizard.sourceforge.io/). Subsequent processing for feature detection, retention time correction, and peak alignment was performed using the XCMS package (v3.12.0) in R. Metabolite identification was conducted by matching accurate mass and MS/MS fragmentation patterns against the following databases: HMDB, METLIN, LipidMAPS, KEGG61, and ChEBI.
Real time qPCR
Total RNA was isolated from hippocampus tissue using the RNA-easy Isolation Reagent (Beyotime, Shanghai, China). Subsequently, cDNA was synthesized from 0.5 to 1 µg of total RNA using the HiScript® III RT SuperMix Kit (Vazyme, Nanjing, China). Quantitative real-time PCR (qRT-PCR) was performed using ChamQ SYBR Color qPCR Master Mix (Vazyme) on a StepOnePlus™ Real-Time PCR System (Applied Biosystems, MA, USA). Gene expression levels were normalized to Actin and quantified using the 2-ΔΔCt method. The specific primer sequences used are listed in Table 1.
Statistical analysis
Statistical analysis was performed with GraphPad Prism 8.0.1 (GraphPad Software, Boston, MA, USA, www.graphpad.com). The Shapiro-Wilk test was used to assess the normality of the data. For data with normal distributions, one-way and two-way ANOVA with Bonferroni’ s post-hoc test was utilized to evaluate differences among more than two groups and adjust for multiple comparisons; otherwise, the Kruskal-Wallis test was applied. The data are presented as Mean ± standard error of mean (SEM), and a P value < 0.05 was considered statistically significant.
Data availability
Raw data have been deposited to National Center for Biotechnology Information (NCBI) under the BioProject number PRJNA1305529. And the associated SRA metadata are available at https://dataview.ncbi.nlm.nih.gov/object/PRJNA1306985?reviewer=c8fe6somn8pv49f7o957n17pu1.
References
You, Y. et al. The prevalence of mild cognitive impairment in type 2 diabetes mellitus patients: a systematic review and meta-analysis. Acta Diabetol. 58, 671–685. https://doi.org/10.1007/s00592-020-01648-9 (2021).
Dove, A. et al. The impact of diabetes on cognitive impairment and its progression to dementia. Alzheimer’s Dement. 17, 1769–1778 (2021).
Biessels, G. J. & Despa, F. Cognitive decline and dementia in diabetes mellitus: mechanisms and clinical implications. Nat. Reviews Endocrinol. 14, 591–604. https://doi.org/10.1038/s41574-018-0048-7 (2018).
van Sloten, T. T., Sedaghat, S., Carnethon, M. R., Launer, L. J. & Stehouwer, C. D. A. Cerebral microvascular complications of type 2 diabetes: stroke, cognitive dysfunction, and depression. Lancet Diabetes Endocrinol. 8, 325–336. https://doi.org/10.1016/S2213-8587(19)30405-X2025/06/15 (2020).
Little, K. et al. Common pathways in dementia and diabetic retinopathy: Understanding the mechanisms of diabetes-related cognitive decline. Trends Endocrinol. Metabolism. 33, 50–71 (2022).
Kullmann, S. et al. Brain insulin resistance at the crossroads of metabolic and cognitive disorders in humans. Physiol. Rev. 96, 1169–1209. https://doi.org/10.1152/physrev.00032.20152025/06/15 (2016).
Bilotta, F. & Qeva, E. And Matot, I. Anesthesia and cognitive disorders: a systematic review of the clinical evidence. Expert Rev. Neurother. 16, 1311–1320. https://doi.org/10.1080/14737175.2016.1203256 (2016).
Liu, H. et al. The association between diabetes mellitus and postoperative cognitive dysfunction: a systematic review and meta-analysis. International J. Surgery 111, 2633–2650 (2025).
Dhande, K. et al. Patient Acceptance and Cost of Intravenous Propofol and Inhalational Sevoflurane for Induction of Anaesthesia : A Prospective Randomized Comparative Study.. Cureus 12, e7687 (2020).
Lv, T. et al. Sevoflurane causes neurotoxicity and cognitive impairment by regulating Hippo signaling pathway-mediated ferroptosis via upregulating PRKCD. Exp. Neurol. 377, 114804 (2024).
Zhou, Y. et al. Microglial pyroptosis in hippocampus mediates sevolfurane-induced cognitive impairment in aged mice via ROS-NLRP3 inflammasome pathway. Int. Immunopharmacol. 116, 109725 (2023).
Liang, F. et al. Sevoflurane anaesthesia induces cognitive impairment in young mice through sequential Tau phosphorylation. Br. J. Anaesth. 131, 726–738.https://doi.org/10.1016/j.bja.2023.06.0592025/06/15 (2023).
Geng, Y., Wu, Q. & Zhang, R. Effect of propofol, sevoflurane, and isoflurane on postoperative cognitive dysfunction following laparoscopic cholecystectomy in elderly patients: A randomized controlled trial. J. Clin. Anesth. 38, 165–171 (2017).
Zhang, Y. et al. Propofol compared with Sevoflurane general anaesthesia is associated with decreased delayed neurocognitive recovery in older adults. Br. J. Anaesth. 121, 595–604.https://doi.org/10.1016/j.bja.2018.05.0592025/06/15 (2018).
Cao, S. J. et al. Delirium in older patients given Propofol or Sevoflurane anaesthesia for major cancer surgery: a multicentre randomised trial. Br. J. Anaesth. 131, 253–265.https://doi.org/10.1016/j.bja.2023.04.0242025/06/15 (2023).
Schoen, J. et al. Cognitive function after Sevoflurane vs propofol-based anaesthesia for on-pump cardiac surgery: a randomized controlled trial. Br. J. Anaesth. 106, 840–850.https://doi.org/10.1093/bja/aer0912025/06/15 (2011).
Duan, G. Y. et al. Cognitive function and delirium following Sevoflurane or Propofol anesthesia for valve replacement surgery: A multicenter randomized controlled trial. Kaohsiung J. Med. Sci. 39, 166–174 (2023).
Liang, C., Du, F., Cang, J. & Xue, Z. Pink1 attenuates propofol-induced apoptosis and oxidative stress in developing neurons. J. Anesth. 32, 62–69 (2018).
Zhang, Y. et al. Propofol prevents oxidative stress and apoptosis by regulating iron homeostasis and targeting JAK/STAT3 signaling in SH-SY5Y cells. Brain Res. Bull. 153, 191–201 (2019).
Nie, Y. et al. Propofol Attenuates Isoflurane-Induced Neurotoxicity and Cognitive Impairment in Fetal and Offspring Mice. Anesthesia & Analgesia 131, 1616–1625 (2020).
Liu, H. et al. Propofol improves sleep deprivation-induced sleep structural and cognitive deficits via upregulating the BMAL1 expression and suppressing microglial M1 polarization. CNS Neurosci. Ther. 30, e14798 (2024).
Villalobos, D. et al. Perioperative changes in neurocognitive and alzheimer’s disease-related cerebrospinal fluid biomarkers in older patients randomised to isoflurane or Propofol for anaesthetic maintenance. Br. J. Anaesth. 131, 328–337.https://doi.org/10.1016/j.bja.2023.04.0192025/06/15(2023).
Peivandi Yazdi, A. et al. The effect of spinal and general anesthesia on serum lipid peroxides and total antioxidant capacity in diabetic patients with lower limb amputation surgery. Archives Bone Joint Surg. 6, 312–317 (2018).
Park, S. J., Oh, A. R., Lee, J. H., Yang, K. & Park, J. Association of preoperative blood glucose level with delirium after non-cardiac surgery in diabetic patients. Korean J. Anesthesiol. 77, 226–235. https://doi.org/10.4097/kja.23301 (2024).
Maniaci, A. et al. Neurological and Olfactory Disturbances After General Anesthesia. Life 15, 344 (2025).
Li, D. et al. Sevoflurane Induces Exaggerated and Persistent Cognitive Decline in a Type II Diabetic Rat Model by Aggregating Hippocampal Inflammation. Front. Pharmacol. 29, 8–2017 (2017).
Liao, W. T., Xiao, X. Y., Zhu, Y. & Zhou, S. P. The effect of Celastrol on learning and memory in diabetic rats after Sevoflurane inhalation. Archives Med. Sci. 14, 370–380 (2018).
Lou, P. H. et al. Propofol (Diprivan®) and Intralipid® Exacerbate Insulin Resistance in Type-2 Diabetic Hearts by Impairing GLUT4 Trafficking. Anesthesia & Analgesia 120, 329–340 (2015).
Fodale, V., Quattrone, D., Trecroci, C., Caminiti, V. & Santamaria, L. B. Alzheimer’s disease and anaesthesia: implications for the central cholinergic system. Br. J. Anaesth. 97, 445–452.https://doi.org/10.1093/bja/ael2332025/06/15 (2006).
Pennings, C. H., Van Boxtel, M., De Korte-De Boer, D., Buhre, W. & Vossen, C. J. Anaesthesia as a risk factor for long-term cognitive decline: Results of the prospective MAAS cohort study. European J. Anaesthesiol. | EJA 42, 468–477 (2025).
E, A., EM, T., AM, T. & MS, A. Effect of Sevoflurane and isoflurane on Post-Anaesthesia cognitive dysfunction in normal and type II diabetic rats. Arch. Razi Inst. 78, 8. https://doi.org/10.22092/ARI.2022.358340.2202 (2023).
Jia, L., Li, H. & Li, T. Efficacy of Sevoflurane and Propofol anesthesia on perioperative adverse cardiovascular events and hemodynamics in elderly patients with diabetes. Clin. Ther. 46, 246–251. https://doi.org/10.1016/j.clinthera.2024.01.006 (2024).
Wang, X., Yang, X., Han, F., Gao, L. & Zhou, Y. Propofol improves brain injury induced by chronic cerebral hypoperfusion in rats. Food Sci. Nutr. 9, 2801–2809. https://doi.org/10.1002/fsn3.1915 (2021).
Kitamura, T., Ogawa, M., Kawamura, G., Sato, K. & Yamada, Y. The Effects of Sevoflurane and Propofol on Glucose Metabolism Under Aerobic Conditions in Fed Rats. Anesthesia & Analgesia 109, 1479–1485 (2009).
Kaushal, A., Bindra, A. & Dube, S. K. Effect of sevoflurane versus desflurane on blood glucose level in patients undergoing intracranial neurosurgery: A randomised controlled study. Indian J. Anaesthesia 66, 769–775 (2022).
Guo, S. et al. Repeated exposure to Sevoflurane impairs the learning and memory of older male rats. Life Sci. 192, 75–83. https://doi.org/10.1016/j.lfs.2017.11.025 (2018).
Zhu, X. et al. Sevoflurane increases intracellular calcium to induce mitochondrial injury and neuroapoptosis. Toxicol. Lett. 336, 11–20. https://doi.org/10.1016/j.toxlet.2020.11.002 (2021).
Wang, X. et al. Beta-Site Amyloid Precursor Protein-Cleaving Enzyme Inhibition Partly Restores Sevoflurane-Induced Deficits on Synaptic Plasticity and Spine Loss. International J. Mol. Sciences 23, 6637 (2022).
Li, X. et al. Histone deacetylase 9 plays a role in sevoflurane-induced neuronal differentiation Inhibition by inactivating cAMP-response element binding protein transcription and inhibiting the expression of neurotrophin-3. FASEB J. 37, e23164. https://doi.org/10.1096/fj.202300168R (2023).
Zhang, L., Yan, J., Liu, Q., Xie, Z. & Jiang, H. LncRNA Rik-203 contributes to anesthesia neurotoxicity via microRNA-101a-3p and GSK-3β-mediated neural differentiation. Sci. Rep. 9, 6822. https://doi.org/10.1038/s41598-019-42991-4 (2019).
Qiao, H. et al. Propofol Affects Neurodegeneration and Neurogenesis by Regulation of Autophagy via Effects on Intracellular Calcium Homeostasis. Anesthesiology 127, 490 (2017).
Liang, C., Sun, M., Zhong, J., Miao, C. & Han, X. The role of Pink1-Mediated mitochondrial pathway in Propofol-Induced developmental neurotoxicity. Neurochem. Res. 46, 2226–2237. https://doi.org/10.1007/s11064-021-03359-1 (2021).
Ulmann, L., Mimouni, V., Roux, S., Porsolt, R. & Poisson, J. P. Brain and hippocampus fatty acid composition in phospholipid classes of aged-relative cognitive deficit rats. Prostaglandins Leukot. Essent. Fat. Acids. 64, 189–195. https://doi.org/10.1054/plef.2001.0260 (2001).
Jia, P. et al. Sevoflurane impairs neurogenesis and cognitive function by inhibiting fatty acid β-oxidation in neural stem/progenitor cells of neonatal rats. Chinese Med. Journal 139, 282–295 (9900).
Omori, W. et al. Reduced cerebrospinal fluid levels of lysophosphatidic acid docosahexaenoic acid in patients with major depressive disorder and schizophrenia. Int. J. Neuropsychopharmacol. 24, 948–955. https://doi.org/10.1093/ijnp/pyab044 (2021).
Kim, S. et al. Hippocampus-dependent cognitive enhancement induced by systemic Gintonin administration. J. Ginseng Res. 40, 55–61. https://doi.org/10.1016/j.jgr.2015.05.001 (2016).
Ha, Y. et al. Korean Red Ginseng Marc-Derived Gintonin Improves Alzheimer’s Cognitive Dysfunction by Upregulating LPAR1. Am. J. Chin. Med. 53, 17–261 (2025).
Norman, J. E., Nuthikattu, S., Milenkovic, D. & Villablanca, A. C. Sex Modifies the Impact of Type 2 Diabetes Mellitus on the Murine Whole Brain Metabolome. Metabolites 13, 1012 (2023).
Rizk, F. H. et al. Fenofibrate improves cognitive impairment induced by High-Fat High-Fructose diet: A possible role of Irisin and heat shock proteins. ACS Chem. Neurosci. 13, 1782–1789. https://doi.org/10.1021/acschemneuro.2c00186 (2022).
Liu, T. et al. PPARα agonist Fenofibrate prevents postoperative cognitive dysfunction by enhancing fatty acid oxidation in mice. Translational Neurosci. 14, 20220317 (2023).
Oliveira, W. H. et al. Effects of Metformin on inflammation and short-term memory in streptozotocin-induced diabetic mice. Brain Res. 1644, 149–160. https://doi.org/10.1016/j.brainres.2016.05.013 (2016).
Nibber, A., Singh, H., Burnet, P., Lennox, B. & Minichino, A. Investigating the pro-cognitive and anti-depressant efficacy of metformin: A systematic review and meta-analysis of randomised controlled trials. J. Affect. Disord. 310, 52–59. https://doi.org/10.1016/j.jad.2022.04.156 (2022).
Jiang, S. et al. Neonatal Sevoflurane exposure disrupted fatty acids metabolism, leading to hypomyelination and neurological impairments. Biomed. Pharmacother. 191, 118495. https://doi.org/10.1016/j.biopha.2025.118495 (2025).
Guide for the Care and Use of Laboratory Animals (8th edition). (National Academies Press (US), (2011).
Furman, B. L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protocols Pharmacol. https://doi.org/10.1002/0471141755.ph0547s70 (2015).
Liu, C., Li, J., Liu, R. & Zhao, G. The role of intestinal homeostasis in sevoflurane-induced Myelin development and cognitive impairment in neonatal mice. Front. Cell. Infect. Microbiol. 15, 2025 (2025).
Alaçam, B., Başarır, Ş. N., Tuna, A. T., Palabıyık, O. & Çakıroğlu, H. The effect of amantadine on recovery, postoperative cognitive dysfunction and pain after Propofol anesthesia in mice. Behav. Brain. Res. 477, 115290. https://doi.org/10.1016/j.bbr.2024.115290 (2025).
Wang, Y. L. et al. Paraventricular thalamus controls consciousness transitions during Propofol anaesthesia in mice. Br. J. Anaesth. 130, 698–708. https://doi.org/10.1016/j.bja.2023.01.016 (2023).
Bromley-Brits, K., Deng, Y. & Song, W. Morris Water Maze Test for Learning and Memory Deficits in Alzheimer’s Disease Model Mice. JoVEhttps://doi.org/10.3791/2920(2011).
Prieur, E. A. K. & Jadavji, N. M. Assessing Spatial Working Memory Using the Spontaneous Alternation Y-maze Test in Aged Male Mice. Bio-protocol. 9, e3162 (2019).
Kanehisa, M., Sato, Y., Kawashima, M., Furumichi, M. & Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44, D457–D462. https://doi.org/10.1093/nar/gkv (2015). 1070%J Nucleic Acids Research.
Acknowledgements
We would like to express our gratitude to the Scientific Research Center of China-Japan Union Hospital of Jilin for their support, as well as to the School of Public Health at Jilin University for their excellent support in laboratory animal core facility and behavior tests.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 82003874).
Author information
Authors and Affiliations
Contributions
Guoqing Zhao Fang Wang and Xinyue Liu conceived and designed the study. Xinyue Liu, Kai Yu and Chang Liu performed the animal experiments. Kai Yu and Wang Yuqin performed the omic and statistical analysis. Xinyue Liu and Yuqin Wang prepared the manuscript. Guoqing Zhao supervised the project. All the authors offered valuable criticism throughout the study and conducted a thorough review of the manuscript. All the authors declare that the study is reported in accordance with ARRIVE guidelines.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Liu, X., Wang, F., Liu, C. et al. Multi-omics analysis reveals sevoflurane exacerbates cognitive impairment in diabetic mice by disrupting lipid metabolism. Sci Rep 16, 9543 (2026). https://doi.org/10.1038/s41598-026-37737-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-37737-y
