
Abstract
This study addresses the challenge of managing complex epidemic dynamics exhibited by a novel Susceptible-Infected-Recovered-Deceased (SIRD) model. The introduced model incorporates a biologically and behaviorally mediated nonlinear incidence rate alongside a saturated treatment function, realistically capturing both physical transmission risks and the human behavioral responses during an outbreak. Our comprehensive analysis establishes the mathematical rigor of the system through positivity, boundedness, and a detailed investigation of equilibrium points and bifurcations (transcritical, saddle-node, and Hopf), which reveal the potential for complex, oscillatory, and chaotic epidemic patterns. To mitigate the disease burden, we apply Pontryagin’s Maximum Principle to derive optimal time-dependent prevention and treatment control strategies. Furthermore, we introduce a highly efficient machine learning framework based on a Logistic-Map Reservoir Computer (LMRC) for accurate, real-time forecasting of these complex dynamics. Numerical simulations validate the theoretical findings and reveal the model’s rich dynamical behaviors, including stable limit cycles and chaotic regimes driven by parameter variations. The integrated analytical, control, and machine learning approach provides a robust toolkit for public health policymakers, offering data-driven insights crucial for designing adaptive, cost-effective interventions that directly support the achievement of Sustainable Development Goal 3 (Good Health and Well-being).
Similar content being viewed by others

Introduction
The global burden of infectious diseases remains profound and dynamic, posing continuous threats to public health, economic stability, and societal well-being in the community where the 21 st century has witnessed a series of devastating outbreaks, from Ebola and Zika to the unprecedented COVID-19 pandemic, underscoring the world’s vulnerability to both emerging and re-emerging pathogens1,2. This kind of landscape is not defined by sudden pandemics alone; a significant and growing concern is the persistent challenge of endemic diseases like tuberculosis and various respiratory infectious diseases, which continue to cause substantial morbidity and mortality, particularly in vulnerable populations3,4. Various drivers such as climate change, urbanization, and antimicrobial resistance are exacerbating these risks, making the accurate prediction and effective management of disease dynamics more critical than ever5.
Mathematical modeling has long served as a cornerstone for understanding and controlling various kinds of infectious diseases, where classical compartmental frameworks such as the Susceptible–Infected–Recovered (SIR) model and its extension to include deceased individuals (SIRD) provide fundamental insights into outbreak trajectories, reproduction numbers, and the impact of different intervention mechanisms6,7,8,9. However, traditional integer-order ordinary differential equation (ODE) models typically assume homogeneous mixing, memoryless transitions, and constant parameters where these assumptions oversimplify the long incubation periods, waning immunity, and behavioral feedback loops observed in real epidemics10,11. To overcome such limitations, fractional-order calculus has been integrated into epidemiological modeling, endowing the system with memory and hereditary properties that capture the dependence of current dynamics on the entire history of the disease5,12,13. Nowadays various studies have demonstrated that fractional-order models offer a more refined representation of diseases ranging from COVID-19 and monkeypox to chickenpox and maize streak epidemics14,15,16,17,18.
Despite these advances, significant gaps remain, since most of the existing mathematical models employ linear or bilinear incidence rates, which fail to account for saturation effects due to behavioral changes or intervention fatigue during high-prevalence outbreaks3,6. Similarly, while the saturated treatment functions have been explored7,11, their integration with psychologically mediated nonlinear incidence and detailed bifurcation analysis, including backward, Hopf, and saddle-node bifurcations, remains underexplored. Furthermore, many optimal control studies focus on single-disease contexts or simpler structures, lacking a unified SIRD framework that combines these realistic nonlinearities with a comprehensive dynamical systems analysis19,20,21. Recent research studies have begun to address co-dynamics (e.g., pneumonia–malnutrition, malnutrition TB, hepatitis B pair-formation) and game-theoretic behavioral feedback22,23,24,25,26,27, yet they often rely on linear incidence and ignore healthcare capacity constraints, and other studies have explored public health education as a standalone control for diseases such as conjunctivitis and norovirus28,29, or culling and vaccination for rabies30, while still using bilinear incidence and missing the interplay between behavioral saturation and treatment constraints. The integration of optimal control with COVID-19 transmission has also been examined31, but without the dual nonlinearities considered here. On the methodological frontier, deep neural networks and reservoir computing have shown promise for forecasting epidemic trajectories, but their integration with mechanistic models and optimal control strategies is still nascent32,33,34. A thorough understanding of the bifurcation structures underlying such complex models requires rigorous mathematical tools, as detailed in classical texts on dynamical systems35.
Our study bridges these gaps by formulating and analyzing a novel SIRD model that incorporates: a nonlinear saturated incidence rate capturing both super-linear transmission and prevalence-dependent behavioral inhibition; and a saturated treatment function reflecting realistic diminishing returns under strained healthcare resources. Also, We provide a rigorous qualitative analysis of equilibria, stability, and bifurcations (transcritical, saddle-node, Hopf), revealing previously undocumented routes to chaos. Using Pontryagin’s maximum principle, we derive time-dependent optimal prevention and treatment strategies that jointly minimize disease burden and intervention costs, and finally, we develop a Logistic-Map Reservoir Computer (LMRC) for efficient, accurate real-time forecasting of the resulting complex dynamics, outperforming conventional recurrent neural networks where this integrated analytical, control, and machine-learning toolkit offers public health policymakers a robust, data-driven framework for designing adaptive, cost-effective interventions, directly supporting Sustainable Development Goal 3 (Good Health and Well-being).
The remainder of this paper is organized as follows. "Mathematical representation of the model" Section details the mathematical formulation of the general SIRD model. "Qualitative analysis of the SIRD model" Section is devoted to the rigorous qualitative analysis of the model. "Optimal control scheme" Section extends the model into an optimal control framework. "Forecasting model dynamics using logistic-map reservoir computer" Section presents a machine learning approach for forecasting epidemic dynamics. "Numerical simulations and discussions of results" Section provides comprehensive numerical simulations. Finally, "Conclusions and future directions" Section concludes the paper by summarizing the key findings and suggesting directions for future research.
Mathematical representation of the model
This study proposes a deterministic, compartmental SIRD model to investigate the transmission dynamics of infectious disease within a closed population. The model stratifies the overall living population N(t) into three distinct, time-dependent epidemiological classes: susceptible individuals S(t), who are at risk of infection; infected individuals I(t), who are contagious and actively contribute to disease transmission; and recovered individuals R(t), who have gained temporary immunity after clearing the infection, expressed as \(N(t) = S(t) + I(t) + R(t)\) for all \(t \ge 0\). A fourth compartment, D(t), tracks the density of deceased individuals who have succumbed to either the disease or natural causes and are subsequently removed from the population. Crucially, while all deaths (from both natural causes at rate \(\mu\) and disease-induced mortality at rate \(\xi\)) feed into the D compartment, the living population dynamics are driven exclusively by interactions among S, I, and R. The core dynamics of this system are governed by a nonlinear, saturated incidence rate that captures complex contagion behaviors and a saturated treatment function reflecting realistic healthcare constraints, as detailed in the following foundational assumptions. Indeed, the quadratic incidence rate is not merely a mathematical choice but serves to describe “superspreading” events and synergistic transmission dynamics often observed in densely populated urban centers. The model’s behaviors in this case can mimic the curved parabolic trajectory of increasing (or decreasing) incidence over time. Furthermore, the saturated treatment function is explicitly grounded in the “healthcare system strain” observed globally during the COVID-19 pandemic, where therapeutic delivery is constrained by finite hospital infrastructure.
Model assumptions
-
I.
The total population \(N(t) = S(t) + I(t) + R(t)\) is dynamically sized and divided into three mutually exclusive epidemiological compartments: Susceptible (S), Infected (I), and Recovered (R). A fourth compartment tracks the cumulative density of Deceased individuals (D) removed from the living population.
-
II.
The model is designed for a general infectious disease characterized by direct person-to-person transmission via respiratory droplets or close contact, aligning with the model’s focus on contact-driven spread.
-
III.
The force of infection is modeled by a nonlinear saturated incidence rate \(\dfrac{(1-w)\beta S I^2}{1+aI^2}\), where:
-
The quadratic term \(I^2\) in the numerator hypothesizes a contagion mechanism where transmission risk increases super-linearly with the infected density, representing aggregation effects or increased symptomatic shedding at higher prevalence.
-
The parameter \(\beta> 0\) is the baseline transmission coefficient, encapsulating the intrinsic contagiousness of the pathogen and baseline contact rate.
-
The term \((1-w)\), where \(0 \le w < 1\), models the efficacy of prevention controls (e.g., mask-wearing, improved ventilation, isolation), scaling down the effective contact rate.
-
The denominator \((1+aI^2)\), with saturation constant \(a> 0\), incorporates a prevalence-dependent inhibitory effect, capturing behavioral changes or psychological factors that reduce mixing as the epidemic becomes more severe.
-
-
IV.
The susceptible population experiences logistic growth, \(p S (1 - S/K)\), rather than constant recruitment. This represents births and new admissions into the population (intrinsic rate \(p> 0\)), constrained by a maximum carrying capacity K reflective of physical space or resource limits in settings like schools, hospitals, or communities.
-
V.
Individuals in the infected compartment I face three possible exits: disease-induced mortality at a constant rate \(\xi> 0\), natural (non-disease) mortality at a constant rate \(\mu> 0\), or recovery through a combination of natural immune response and medical treatment.
-
VI.
The recovery process is split into two pathways: a constant fraction of infected individuals recovers naturally at rate \(\sigma \ge 0\), and an additional, controlled recovery flux is provided via medical treatment.
-
VII.
The treatment rate is modeled by a saturated function \(\dfrac{v I}{1+bI}\), where:
-
The parameter \(v \ge 0\) represents the treatment control effort, proportional to the availability and administration rate of therapeutics.
-
The saturation constant \(b> 0\) models the diminishing per-capita effectiveness of treatment due to resource limitations (e.g., finite hospital beds, staff, or drug supplies).
-
As I grows large, the treatment term approaches the constant capacity v/b.
-
-
VIII.
Recovered individuals do not acquire permanent immunity. They revert to the susceptible compartment at a constant waning rate \(r> 0\), which is epidemiologically significant for many infectious diseases as neither natural infection nor vaccination may confer lifelong protection.
-
IX.
Individuals who die from disease (\(\xi I\)) or natural causes (\(\mu I\)) enter the deceased compartment D. This compartment is cleared at a constant rate \(\alpha> 0\), representing the coordinated removal, burial, or cremation of deceased individuals.
-
X.
The population is assumed to be homogeneously mixed. All susceptible individuals have an equal probability of contact with any infected individual, and the contact rate is implicitly incorporated into the transmission coefficient \(\beta\).
-
XI.
The model does not explicitly include an Exposed (E) compartment, assuming the latent period between infection and becoming infectious is negligible compared to the infectious period or is implicitly contained within the infected stage.
-
XII.
New entrants into the susceptible compartment come only from logistic growth/births (assumed disease-free) and waning immunity. The model does not currently include a vaccinated class or maternal antibody protection.
-
XIII.
All model parameters (\(p, K, \beta , a, \mu , \xi , \sigma , v, b, r, \alpha\)) are non-negative constants over the simulation period, assuming environmental conditions and pathogen virulence are time-invariant.
-
XIV.
The model is initiated at time \(t=0\) with non-negative populations in all compartments: \(S(0)> 0, I(0) \ge 0, R(0) \ge 0, D(0) \ge 0\). The analysis focuses on the domain where solutions remain biologically meaningful.
-
XV.
The model is formulated as a system of ordinary differential equations, neglecting stochastic effects. This is appropriate for large population sizes where random events have minimal impact on the macroscopic dynamics.
-
XVI.
Apart from the specified logistic growth and mortality, the population is considered closed to external migration. The model is designed for an isolated community or specific institutional settings (e.g., schools, workplaces, or communities).
-
XVII.
The model assumes that treatment immediately moves individuals from the infected to the recovered compartment, ignoring any possible hospitalization stage or treatment delay.
Furthermore, the strong biological and empirical rationale behind the nonlinear incidence term, \(\frac{(1-w)\beta S I^2}{1 + a I^2}\), is supported by the following key observations:
-
1.
The \(I^2\) dependency in the numerator accounts for “super-linear” transmission. This models scenarios where infection risk scales nonlinearly with prevalence, such as in high-contact social environments or when high environmental viral loads increase the probability of a susceptible individual being exposed to multiple infectious particles simultaneously. This aligns with empirical observations of “power-law” dynamics in respiratory disease transmission.
-
2.
The denominator \((1 + aI^2)\) introduces a saturation effect representing behavioral and psychological feedback. As infection levels rise, individuals naturally adopt self-protective measures (e.g., social distancing or reduced mobility) even in the absence of mandates. The parameter a quantifies this inhibitory effect, preventing the force of infection from growing boundlessly and reflecting the finite limits of human interaction.
-
3.
The inclusion of \((1-w)\) provides a measurable link to prevention efficacy (e.g., mask-wearing), allowing the model to simulate the impact of non-pharmaceutical interventions. Together, these terms allow the model to capture complex wave behaviors that standard mass-action models cannot represent.
Indeed, the model’s complexity is essential for capturing non-trivial epidemiological phenomena that simpler models fail to describe. In what follows, we highlight three key insights that arise solely from this structure:
-
1.
Resource Limits: The saturated treatment term \(\frac{vI}{1+bI}\) provides a realistic assessment of healthcare capacity. It allows policymakers to determine the “threshold of system collapse,” identifying the precise point where medical resources are overwhelmed by infection volume. In other words, the models takes into account that the there is a maximum effective treatment rate - hospitals, staff, ICU beds, etc. cannot increase indefinitely. As infections rise, each additional patient receives less effective care on average.
-
2.
Adaptive Contact Rates: By incorporating behavioral inhibition (a), the model better simulates the real scenario that when infections rise, people reduce contacts wear masks, and avoid gatherings. This presents a more accurate prediction of “peak flattening” caused by social self-organization.
-
3.
The obtained results reveal that \(\mathcal {R}_0 = 0\). In terms of epidemiological interpretation, this signifies that a single infected individual cannot trigger a massive outbreak in a fully susceptible population under this framework. Instead, the system requires a “critical mass” of infected individuals to initiate an epidemic wave.
Thus, the complexity introduces useful practical consideration into the model. We have refined "Qualitative analysis of the SIRD model" Section to more clearly connect these mathematical features to public health strategy.
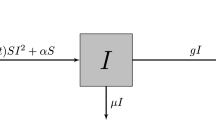
Model flow diagram
According to the model assumptions, state variable descriptions and parameters described in Table 1 above, the dynamics of infectious disease transmission are depicted in the schematic diagram in Fig. 1.

Schematic diagram for model (1).
Mathematical formulation
Based on the schematic diagram, the transmission dynamics can be described by the following system of differential equations:
then by taking \((1-w) \beta =u,\) then the model can be written as follows:
where \(S(0)>0, I(0)\ge 0, R(0)\ge 0, D(0)\ge 0\) for all \(t\ge 0\).
Qualitative analysis of the SIRD model
Fundamental properties of the system’s solutions
The initial step in analyzing the proposed system, described by the set of differential equations in (2), is to confirm that its solutions remain physically meaningful. Specifically, we must demonstrate that all state variables are non-negative and bounded for any non-negative initial conditions.
Theorem 3.1
For any non-negative initial conditions, the solution \((S(t), I(t), R(t), D(t))\) of system (2) is guaranteed to remain non-negative for all subsequent times \(t> 0\).
Proof
By evaluating the rate of change of each state variable at the boundary where that variable is zero, we can confirm the non-negativity of the solution components. From system (2), we have:
The expressions on the right-hand side clearly indicate that the flow is directed inward or along the boundary of the non-negative orthant. Consequently, any solution \((S(t), I(t),\) \(R(t), D(t))\) originating from non-negative initial conditions will remain non-negative for all \(t> 0\).
Theorem 3.2
Assume that the initial conditions of the model are non-negative, then every solution \((S(t), I(t), R(t), D(t))\) of (2) is restricted to the compact and positively invariant region
Proof
First, we consider the total living population \(N = S+I+R\). Summing the first three equations of the system (4) yields the rate of change for N:
which leads to the inequality:
By completing the square on the right-hand side, we obtain:
Applying the theory of differential inequalities, the solution for N(t) is bounded by
This implies that N approaches the upper bound \(\dfrac{p K}{4\mu }\) as \(t \rightarrow \infty\). Similarly, for the deceased class D, the rate of change is given by:
Since \(I \le N \le \dfrac{pK}{4\mu }\), we can establish the following inequality:
The solution for D(t) is bounded by \(D(t) \le \dfrac{pK(\mu +\xi ) }{4\mu \alpha }+\left( D(0)-\dfrac{pK(\mu +\xi ) }{4\mu \alpha }\right) e^{-\alpha t}\), and D approaches \(\dfrac{pK(\mu +\xi ) }{4\mu \alpha }\) as \(t \rightarrow \infty\). Thus, all solutions of system (2) are confined within the region \(\Omega\).
Given the nonlinear nature of system (2), a complete analytical solution is generally unattainable. Therefore, we proceed by investigating the qualitative behavior of the system, focusing on the existence and stability of its equilibrium points. Since the dynamics of S, I, and R are independent of D, we simplify the analysis by considering the reduced three-dimensional subsystem:
Equilibrium points analysis
The model system (4) possesses several equilibrium points, which represent steady-state solutions.
-
1.
The Trivial Equilibrium E(0, 0, 0) is always present.
-
2.
The disease-free equilibrium \(E_0 \left( S^{0} = \dfrac{K ( p - \mu )}{p}, 0, 0 \right)\) exists provided that the intrinsic growth rate p is strictly greater than the natural mortality rate \(\mu\) (\(p> \mu\)).
-
3.
An endemic equilibrium \(E^{*} (S^{*}, I^{*}, R^{*})\) occurs when the disease persists in the population, requiring \(I^{*}> 0\). At this steady state, setting the system derivatives to zero yields the following relationships for \(S^{*}\) and \(R^{*}\) as explicit functions of \(I^{*}\):
To determine the values of \(I^{*}\), we substitute these expressions back into the stationary state equation of the susceptible population. Utilizing the equilibrium condition \(\dfrac{u S^{*} {I^{*}}^2}{1+a {I^{*}}^2} = (\sigma +\mu +\xi )I^{*} + \dfrac{vI^{*}}{1+bI^{*}}\), the first equation becomes:
To clear the rational fractions and consolidate Eq. (6) into a single polynomial, we first define several auxiliary constants based on the model’s parameters:
Using these constants, the expressions for \(S^{*}\) and \(R^{*}\) simplify to:
Substituting these simplified forms into Eq. (6) and multiplying the entire expression by the common denominator \(u^2 K (\mu +r) {I^{*}}^2 (1+bI^{*})^2\) eliminates all rational terms, yielding the following expansion:
Expanding this expression and grouping the terms by powers of \(I^{*}\) results in a 6th-order polynomial equation:
The coefficients \(\ell _i\) are explicitly defined as:
Since all fundamental parameters are non-negative, the leading coefficient \(\ell _6 < 0\) and the constant term \(\ell _0 < 0\) are both strictly negative. Applying Descartes’ Rule of Signs to the sequence \(\{\ell _6, \ell _5, \ell _4, \ell _3, \ell _2, \ell _1, \ell _0\}\), the polynomial must exhibit an even number of sign changes because it begins and ends with the same sign. Consequently, the system (4) will support zero, two, or four positive endemic equilibria depending on the intermediate control and transmission parameters. This analytical result rigorously establishes the mathematical foundation for the backward bifurcations and bistable regimes observed in the model. The complexity of the coefficients \(\ell _i\) prevents their explicit listing. However, an application of Descartes’ rule of signs to the polynomial in \(I^{*}\) allows us to conclude that the system (4) can support either two positive endemic equilibrium points or none, depending on the signs of the coefficients \(\ell _i\), see Table 2.
Local stability analysis of equilibria
The local stability of the equilibrium points is determined by the eigenvalues of the Jacobian matrix evaluated at each point.
Theorem 3.3
The trivial equilibrium \(E(0,0,0)\) is locally asymptotically stable if \(p < \mu\), and it becomes unstable when \(p> \mu\).
Proof
The Jacobian matrix J of the reduced system (4) evaluated at the trivial point \(E\) is:
The eigenvalues of \(J_E\) are \(\lambda _1 = p - \mu\), \(\lambda _2 = -(\mu + \xi + \sigma + v)\), and \(\lambda _3 = -(\mu + r)\). Since \(\lambda _2\) and \(\lambda _3\) are always negative, the stability is governed by \(\lambda _1\). If \(p < \mu\), then \(\lambda _1 < 0\), and all eigenvalues are negative, implying that the trivial equilibrium is locally asymptotically stable. Conversely, if \(p> \mu\), then \(\lambda _1> 0\), rendering the equilibrium unstable.
Theorem 3.4
The disease-free equilibrium \(E_0 \left( S^{0} = \dfrac{K\left( p - \mu \right) }{p}, 0, 0 \right)\) is locally asymptotically stable iff \(p> \mu\), and unstable otherwise.
Proof
The Jacobian matrix evaluated at the disease-free equilibrium \(E_0\) is:
The eigenvalues of \(J_{E_0}\) are \(\lambda _1 = -(\mu + r)\), \(\lambda _2 = \mu - p\), and \(\lambda _3 = -(\mu + \xi + \sigma + v)\). For local asymptotic stability, all eigenvalues must be negative. This condition is satisfied if and only if \(p> \mu\), which ensures \(\lambda _2 < 0\). Therefore, \(E_0\) is locally asymptotically stable under this condition and unstable otherwise.
Theorem 3.5
The endemic equilibrium \(E^{*}(S^{*}, I^{*}, R^{*})\) is locally asymptotically stable provided that the Routh-Hurwitz conditions \(d_1> 0\), \(d_2> 0\), \(d_3> 0\), and \(d_1 d_2> d_3\) are satisfied, where \(d_1\), \(d_2\), and \(d_3\) are the coefficients of the characteristic polynomial.
Proof
The Jacobian matrix at the endemic equilibrium \(E^{*}\) is given by:
where the entries are: \(c_{{11}} = - \frac{{uI^{*2} }}{{1 + aI^{*2} }} - \frac{{2pS^{*} }}{K} - \mu + p,c_{{12}} = - \frac{{2uS^{*} I^{*} }}{{\left( {1 + aI^{*2} } \right)^{2} }},c_{{13}} = r,\)\(c_{{21}} = \frac{{uI^{*2} }}{{1 + aI^{*2} }},\)\(c_{{22}} = \frac{{2uS^{*} I^{*} }}{{1 + aI^{{*2}} }} - \frac{{2auS^{*} I^{{*3}} }}{{\left( {1 + aI^{{*2}} } \right)^{2} }} + \frac{{bvI^{*} }}{{(1 + bI^{*} )^{2} }} - \frac{v}{{1 + bI^{*} }} - \mu - \xi I^{*} - \sigma ,\)\(c_{{32}} = \frac{{\sigma (1 + bI^{*} )^{2} + v}}{{(1 + bI^{*} )^{2} }},\) and \(c_{33}=-\mu -r.\)
The characteristic equation of \(J_{E^{*}}\) is:
where the coefficients are defined as: \(d_1=-(c_{11}+c_{22}+c_{33}),\)\(d_2=c_{11} c_{22}+c_{11} c_{33}+c_{22} c_{33}-c_{12} c_{21},\)\(d_3=c_{12} c_{21} c_{33}-c_{13} c_{21} c_{32}-c_{11} c_{22} c_{33}.\) The Routh–Hurwitz stability criterion ensures that all roots of Eq. (9) have negative real parts, which is the condition for local asymptotic stability of \(E^{*}\). This criterion requires that \(d_1> 0\), \(d_2> 0\), \(d_3> 0\), and \(d_1 d_2> d_3\).
To visually support the theoretical stability results for the trivial point E and the disease-free equilibrium point \(E_0\), we illustrate their stability and instability regions in Fig. 2, using the parameter values provided in Table 3. The regions of instability are highlighted in yellow, while the stable regions are shown in various colors.

(a–c) 3D stability and instability regions of the trivial point E, (d–i) 3D stability and instability regions of the disease-free equilibrium point \(E_0\).
Threshold analysis and derivation of \(\mathcal {R}_{0}\)
To determine the reproduction number, we linearize the system around the DFE, \(E_{0} = (S^{*}, 0, 0, 0)\), where \(S^{*} = K(1 - \mu /p)\). Defining \(\mathcal {F}\) as the rate of new infections and \(\mathcal {V}\) as the rate of transitions out of the infected class, it follows that:
Calculating the Jacobian matrix F for the new infection rate at \(E_0\):
Since \(F=0\), the spectral radius \(\rho (FV^{-1})\) results in \(\mathcal {R}_0 = 0\). This result indicates that the model is governed by “density-dependent” transmission thresholds rather than single-case propagation.The interpretation of \(\mathcal {R}_0\) can be updated to reflect that while \(\mathcal {R}_0=0\) at the origin, the system exhibits stable endemic states for higher initial values of I, a characteristic of nonlinear synergistic contagion.
In terms of epidemiological interpretation, this signifies that a single infected individual cannot trigger a massive outbreak in a fully susceptible population under this framework. Instead, the system requires a “critical mass” of infected individuals to initiate an epidemic wave.
Transcritical bifurcation analysis
The system’s dynamics exhibit a critical change in stability at a specific parameter value, which is investigated using Sotomayor’s theorem. This analysis reveals that a transcritical bifurcation occurs when the population growth rate p equals the natural mortality rate \(\mu\).
Theorem 3.6
System (4) undergoes a transcritical bifurcation at the trivial equilibrium \(E\) as the parameter \(p\) crosses the critical value \(p^{*} = \mu\).
Proof
When the bifurcation parameter is set to its critical value, \(p^{*} = \mu\), the Jacobian matrix \(J_E\) evaluated at the trivial equilibrium \(E\) of model (4) is:
The characteristic equation for \(J_E\) at \(p^{*} = \mu\) is:
This yields the eigenvalues:
The presence of a zero eigenvalue indicates that a bifurcation may occur. We now determine the eigenvectors \(\mathcal {{\textbf {V}}}\) and \(\mathcal {{\textbf {W}}}\) corresponding to the zero eigenvalue of \(J_E\) and its transpose \(J_E^T\), respectively:
Let \(\mathcal {{\textbf {F}}}\) be the vector field of the system:
To satisfy the conditions of Sotomayor’s theorem for a transcritical bifurcation, we must verify three conditions. First, the partial derivative of the vector field with respect to the parameter p, evaluated at the equilibrium, must be orthogonal to \(\mathcal {{\textbf {W}}}\):
The second condition involves the mixed partial derivative:
The third condition involves the second-order partial derivative of the vector field:
Since the second and third conditions are non-zero, Sotomayor’s theorem confirms that a transcritical bifurcation occurs at \(p^{*} = \mu\). This bifurcation signifies the exchange of stability between the trivial equilibrium E and the disease-free equilibrium \(E_0\).

2D Stability and instability regions of trivial point E and disease-free equilibrium point \(E_0\).

A transcritical bifurcation diagram concerning \(p\), while maintaining the parameters listed in Table 3.

(a) Solution trajectories converge to the trivial equilibrium point E, (b) Solution trajectories converge to the disease-free equilibrium point \(E_0\).
The results presented in Figs. 3, 4, and 5 collectively illustrate the critical threshold dynamics that govern whether the disease persists or is eliminated from the population. Figure 3 delineates the parameter space (in terms of \(p\) and \(\mu\)) where either the trivial equilibrium \(E\) or the disease-free equilibrium \(E_0\) is stable, clearly defining the conditions for disease extinction or absence. This theoretical stability is corroborated by the time-domain simulations in Fig. 5, which show solution trajectories converging to the stable trivial point when \(p < \mu\) (panel a) or to the stable disease-free point when \(p> \mu\) (panel b). The transition between these two stable states is precisely characterized by the transcritical bifurcation shown in Fig. 4, occurring at the critical parameter value \(p = \mu\). From an epidemiological perspective, this bifurcation represents a crucial reproduction threshold: when \(p> \mu\), the population’s intrinsic growth rate is sufficient to sustain the susceptible class and allow the disease to settle at the endemic level \(E_0\); conversely, when \(p < \mu\), the population declines, leading to the eventual extinction of both the host population and the disease at E.
Saddle-node bifurcation analysis
The possibility of a saddle-node bifurcation, which corresponds to the sudden appearance or disappearance of a pair of equilibrium points, is investigated at the endemic equilibrium \(E^{*}\).
Theorem 3.7
The system (4) exhibits a saddle-node bifurcation at the endemic equilibrium \(E^{*}(S^{*}, I^{*}, R^{*})\) when \(p = p^{**}\), provided that a specific non-degeneracy condition on the second-order derivatives of the vector field is satisfied.
Proof
A saddle-node bifurcation occurs when the characteristic Eq. (9) has a single zero eigenvalue, implying that p is at a critical value \(p^{**}\). We define the vector field \({\textbf {F}}(S, I, R, p)\) as:
The partial derivative of \({\textbf {F}}\) with respect to the parameter p is:
The Jacobian matrix \(D {\textbf {F}} (E^{*};p)\) is given by (8). The eigenvectors \({\textbf {V}}\) and \({\textbf {W}}\) associated with the zero eigenvalue of \(D{\textbf {F}}(E^{*})\) and \(D^T{\textbf {F}}(E^{*})\), respectively, are:
For the saddle-node bifurcation to occur, two non-degeneracy conditions must be met. The first condition is:
The second condition involves the second-order derivative of the vector field:
If both of these conditions are satisfied, Sotomayor’s theorem confirms that the system (4) undergoes a saddle-node bifurcation at \(E^{*}(S^{*}, I^{*}, R^{*})\) when \(p = p{**}\).
Hopf bifurcation analysis
We now investigate the conditions under which the endemic equilibrium \(E^{*}\) loses stability, leading to the emergence of isolated periodic solutions (limit cycles) through Hopf bifurcation. As the deceased class \(D(t)\) does not affect the dynamics of the other three compartments, the analysis focuses on the reduced three-dimensional model:
The Jacobian matrix \(J(E^{*})\) at the endemic equilibrium \(E^{*} = (S^{*}, I^{*}, R^{*})\) is:
where the entries are:
The characteristic equation of \(J(E^{*})\) is a cubic polynomial in \(\lambda\):
where the coefficients are:
A Hopf bifurcation occurs when the characteristic Eq. (14) admits a pair of purely imaginary eigenvalues, \(\lambda = \pm i\omega\) (\(\omega> 0\)), and one negative real eigenvalue. Substituting \(\lambda = i\omega\) into (14) and separating the real and imaginary parts yields:
Since \(\omega> 0\), the second equation gives \(\omega ^2 = C_2\). Substituting this into the first equation provides the necessary condition for the Hopf bifurcation:
For the bifurcation to be non-degenerate, the real part of the complex conjugate eigenvalues must cross the imaginary axis with a non-zero speed as the bifurcation parameter \(\eta\) is varied:
where \(\eta\) is a chosen parameter (e.g., u, v, a, b, p).
To determine the direction and stability of the resulting periodic orbit, we utilize the center manifold reduction and normal form theory. We first translate the equilibrium to the origin using the transformation:
The system then takes the form:
where \(F_i\) represent the nonlinear terms (quadratic and higher). At the Hopf bifurcation point, \(J(E^{*})\) has eigenvalues \(\lambda _{1,2} = \pm i\omega\) and \(\lambda _3 = -C_1 < 0\). By applying a linear transformation using the eigenvector matrix P:
such that \(P^{-1} J P = \text {diag}(i\omega , -i\omega , -C_1)\), and setting:
the system is transformed into:
The dynamics on the center manifold are approximated by setting \(z = h(x,y)\), leading to the reduced two-dimensional system:
where \(g^i\) contain the quadratic and cubic nonlinearities. Finally, the first Lyapunov coefficient \(\Gamma\) is calculated to determine the nature of the bifurcation:
where the subscripts denote partial derivatives evaluated at the origin. The sign of \(\Gamma\) dictates the stability of the bifurcating limit cycle in the way that two cases arise:
-
If \(\Gamma < 0\), the bifurcation is supercritical, resulting in a stable limit cycle.
-
If \(\Gamma> 0\), the bifurcation is subcritical, resulting in an unstable limit cycle.
For the current model, the occurrence of a Hopf bifurcation suggests that sustained, periodic oscillations in the infection levels can emerge even without external periodic forcing.
Summarizing previously obtained results, Fig. 2 presents three-dimensional stability and instability regions for the trivial equilibrium \(E\) and the disease-free equilibrium \(E_0\) of the SIRD model. These regions provide critical insight into how parameter variations govern the long-term behavior of the epidemic. For the trivial equilibrium \(E = (0,0,0)\), the stability condition \(p < \mu\) implies that when the intrinsic growth rate of the susceptible population is lower than the natural mortality rate, the population cannot sustain itself and inevitably collapses to extinction. Biologically, this corresponds to a scenario where deaths outpace new entries into the population, leading to depopulation regardless of disease dynamics. Conversely, when \(p> \mu\), the trivial equilibrium becomes unstable, allowing the population to persist. The disease-free equilibrium \(E_0\) exhibits local asymptotic stability precisely when \(p> \mu\), as established in Theorem 3.4, meaning that the population can maintain itself and the disease fails to invade. The colorful regions in Fig. 2 indicate stable parameter domains, while yellow regions denote instability. The bifurcation analysis is carried out and reveals that the recruitment rate \(p\) and natural mortality \(\mu\) act as critical switches: their relative magnitude determines whether the system settles into a disease-free state or permits infection establishment. From a public health perspective, this underscores that demographic factors-birth rates and baseline mortality-are not mere background conditions but active determinants of epidemic vulnerability.
The analytical results of bifurcation analysis are validated using the numerical continuation MATCONT package, see Fig. 6 below, and show agreement with theoretical results. The branch point (BP) refers to the transcritical bifurcation that exists at the threshold value \(p=\mu\), as previously obtained from analytical bifurcation analysis. Moreover, the bifurcation diagrams and Lyapunov exponents plots are also presented in revised manuscript to complement bifurcation analysis and numerical simulations of the model; see Fig. 7. It is verified that the maximum Lyapunov exponent has a positive value for some values of the model’s parameters, which confirms the existence of chaotic behaviors.

Results of numerical continuation of trivial and disease-free equilibrium points of the model. The branch point (BP) refers to transcritical bifurcation that exists at the threshold value \(p=\mu\).

(a) Solution time series, (b) 3D phase portrait, (c−e) bifurcation diagrams with respect to parameter \(a \in [0.4, 0.42]\), and (d) Lyapunov exponents (LEs) plots of the model.
Optimal control scheme
This section focuses on extending the SIRD model (1) by integrating optimal control theory. The primary objective is to formulate a dynamic strategy that effectively minimizes the prevalence of infection and the cumulative impact of disease-related mortality, while simultaneously accounting for the economic and logistical costs associated with the implementation of public health interventions. To achieve this, we introduce two time-dependent control variables:
-
Prophylactic Control, w(t): This control variable, constrained by \(0 \le w(t) \le w_{max} < 1\), represents the intensity of preventive measures, such as public awareness campaigns, social distancing mandates, or hygiene promotion efforts. Its function is to reduce the effective disease transmission rate, \(\beta\).
-
Therapeutic Control, v(t): This variable, with bounds \(0 \le v(t) \le v_{max}\), quantifies the resources dedicated to treating infected individuals, including the provision of medical care and facilities. Its application is designed to increase the rate at which infected individuals transition to the recovered class.
By substituting the constant parameters w and v in the original model (1) with these dynamic control functions, we establish the following controlled system of differential equations:
with the initial conditions \(S(0)> 0, I(0) \ge 0, R(0) \ge 0, D(0) \ge 0\).
Definition of the objective functional
The goal of the optimal control problem is to find the control functions that minimize a functional J over a fixed time interval \([0, T_f]\). This functional is constructed to balance the minimization of the disease burden with the minimization of the intervention costs. The burden is quantified by the number of infected individuals I(t) and the cumulative deceased population D(t). The costs of the controls are modeled quadratically. The objective functional is formally defined as:
where \(A_1, A_2, A_3, A_4\) are positive weighting constants. The term \(A_1 I(t)\) represents the societal cost associated with the infected population, while \(A_4 D(t)\) accounts for the long-term societal and logistical costs related to the deceased population. The terms \(\frac{A_2}{2} w(t)^2\) and \(\frac{A_3}{2} v(t)^2\) represent the costs incurred for implementing the prophylactic and therapeutic controls, respectively.
We aim to identify the optimal control pair \((w^{*}(t), v^{*}(t))\) that satisfies:
where the set of admissible controls \(\mathcal {U}\) is defined as:
Derivation of optimal control conditions
To derive the necessary conditions that the optimal controls must satisfy, we employ Pontryagin’s Maximum Principle (PMP)36,37. The first step is to define the Hamiltonian function, \(\mathcal {H}\), for the system:
where \(\lambda _S, \lambda _I, \lambda _R,\) and \(\lambda _D\) are the adjoint variables (or co-state variables) corresponding to the state variables S, I, R, and D, respectively.
Theorem 4.1
Given the optimal controls \(w^{*}(t)\) and \(v^{*}(t)\) and the corresponding optimal state trajectories \(S^{*}, I^{*}, R^{*}, D^{*}\) of the system (17), there exists a set of adjoint variables \(\lambda _S(t), \lambda _I(t), \lambda _R(t), \lambda _D(t)\) that satisfy the following conditions:
-
1.
Adjoint equations The co-state variables satisfy the following system of differential equations, derived from \(\frac{d\lambda _i}{dt} = -\frac{\partial \mathcal {H}}{\partial x_i}\):
$$\begin{aligned} {\left\{ \begin{array}{ll} \dfrac{d\lambda _S}{dt} = - \dfrac{\partial \mathcal {H}}{\partial S} = -\lambda _S \left[ p\left( 1-\dfrac{2S}{K}\right) - \dfrac{(1-w)\beta I^2}{1+aI^2} - \mu \right] - \lambda _I \left[ \dfrac{(1-w)\beta I^2}{1+aI^2} \right] , \\ \\ \dfrac{d\lambda _I}{dt} = - \dfrac{\partial \mathcal {H}}{\partial I} = -A_1 - (\lambda _S - \lambda _I) \left[ \dfrac{2(1-w)\beta S I}{(1+aI^2)^2} \right] + \lambda _I \left[ \sigma +\mu +\xi + \dfrac{v}{(1+bI)^2} \right] \\ \qquad \qquad \qquad - \lambda _R \left[ \dfrac{v}{(1+bI)^2} + \sigma \right] - \lambda _D[\mu +\xi ], \\ \\ \dfrac{d\lambda _R}{dt} = - \dfrac{\partial \mathcal {H}}{\partial R} = -\lambda _S r + \lambda _R(\mu +r), \\ \\ \dfrac{d\lambda _D}{dt} = - \dfrac{\partial \mathcal {H}}{\partial D} = \lambda _D \alpha - A_4. \end{array}\right. } \end{aligned}$$(22) -
2.
Transversality conditions Since the final time \(T_f\) is fixed and the final states are free, the adjoint variables must satisfy the following terminal conditions:
$$\begin{aligned} \lambda _S(T_f) = 0, \quad \lambda _I(T_f) = 0, \quad \lambda _R(T_f) = 0, \quad \lambda _D(T_f) = 0. \end{aligned}$$(23) -
3.
Optimality conditions The optimal controls \(w^{*}(t)\) and \(v^{*}(t)\) are determined by minimizing the Hamiltonian \(\mathcal {H}\) over the admissible control set \(\mathcal {U}\). This yields the following characterizations:
$$\begin{aligned} w^{*}(t) = \min \left\{ w_{max}, \max \left\{ 0, \frac{(\lambda _S - \lambda _I) \beta S I^2}{A_2(1+aI^2)} \right\} \right\} , \end{aligned}$$(24)$$\begin{aligned} v^{*}(t) = \min \left\{ v_{max}, \max \left\{ 0, \frac{(\lambda _I - \lambda _R) I}{A_3(1+bI)} \right\} \right\} . \end{aligned}$$(25)
Proof
The adjoint system (22) is a direct consequence of the PMP condition \(\frac{d\lambda _i}{dt} = -\frac{\partial \mathcal {H}}{\partial x_i}\). Notably, the inclusion of the term \(A_4 D\) in the objective functional results in the constant term \(-A_4\) in the \(\frac{\partial \mathcal {H}}{\partial D}\) expression, which is reflected in the \(\lambda _D\) adjoint equation. The transversality conditions (23) are standard for a fixed-time, free-endpoint optimal control problem. The characterization of the optimal controls is obtained by setting the partial derivatives of the Hamiltonian with respect to the controls to zero:
By incorporating the constraints imposed by the admissible control set \(\mathcal {U}\), the optimal controls are expressed in the form of a bounded function, leading to the final characterizations in (24) and (25).
The combination of the state system (17) and the adjoint system (22), along with the initial conditions for the state variables, the terminal conditions for the adjoint variables, and the algebraic optimality conditions (24–25), forms a Two-Point Boundary Value Problem (TPBVP). Given the complexity and nonlinearity of this coupled system, an analytical solution is typically intractable, necessitating the use of numerical techniques, such as the iterative forward-backward sweep method, for its resolution.
Forecasting model dynamics using logistic-map reservoir computer
A machine learning-based prediction scheme relying on a logistic-map reservoir computer32, incorporating virtual nodes and finite memory, is utilized in this section as an efficient and accurate approach for learning and forecasting the nonlinear dynamical behaviors of the present model. The approach achieves competitive performance with minimal architectural complexity, making it particularly suitable for low-cost hardware implementations and fast learning of nonlinear dynamical systems.
Formulation of the prediction problem
Assume that the discrete-time sequence of training data is given by
where \(s_k\) denotes the input signal sampled from a dynamical variable (e.g. x(t)) and \(r_k\) denotes the target signal (e.g. y(t)) at discrete time index k. The aim is to learn a mapping that predicts future outputs \(r_{k+1}, r_{k+2}, \dots\) from the present and past values of \(s_k\).
Logistic-map reservoir construction
Each input \(s_k\) is mapped to a control parameter \(\alpha _k\) via a linear rescaling,
where \((\alpha _{\min },\alpha _{\max })\) defines the admissible parameter range. The reservoir dynamics are generated by iterating a logistic-type nonlinear map
from a fixed initial condition \(\xi _0\). For each \(\alpha _k\), the map is iterated Q times, producing a virtual-node vector
These virtual nodes represent the instantaneous reservoir response to the input \(s_k\).
Temporal embedding and reservoir state
To incorporate memory, a temporal embedding of depth M is employed. The full reservoir state corresponding to input \(s_k\) is defined as
where \(\{\lambda _j\}_{j=1}^{M}\) are fixed memory weights satisfying \(0 \le \lambda _j \le 1\) and \(\lambda _M = 1\). The state dimension is therefore MQ. Collecting all states yields the reservoir matrix
Given sufficiently large memory depth M and virtual-node count Q, the mapping
embeds the input time series into a high-dimensional feature space that is linearly separable for generic nonlinear dynamical responses.
Training and prediction algorithm
Algorithm 1: Logistic-map temporal reservoir computing
-
1.
Input: Training data \(\{(s_k,r_k)\}_{k=1}^{N}\), parameters \((\alpha _{\min },\alpha _{\max })\), Q, M.
-
2.
Rescale each \(s_k \rightarrow \alpha _k\).
-
3.
Generate virtual nodes \(\varvec{\phi }_k\) via Q iterations of the map.
-
4.
Construct embedded states \(\varvec{\psi }_k\) using the previous M inputs.
-
5.
Form the reservoir matrix \(\varvec{\Psi }\).
-
6.
Compute linear readout weights
$$\textbf{w} = \textbf{r}\,\varvec{\Psi }^{\dagger },$$where \(\textbf{r}=[r_1,\ldots ,r_N]\). The optimal readout weights are obtained using the Moore–Penrose pseudoinverse.
-
7.
Prediction: For new inputs, repeat steps 2–4 and compute
$$\hat{r}_{k+1} = \textbf{w}\,\varvec{\psi }_k.$$

Schematic of the logistic-map-based temporal reservoir computing framework. Scalar inputs are transformed into high-dimensional spatiotemporal states via virtual nodes and finite memory, followed by a linear readout.
Now, we assess the capability of the proposed Logistic-Map Reservoir Computer (LMRC) to learn and forecast the temporal evolution of different state variables arising from the SIRD model. The objective is to demonstrate that the same reservoir architecture can accurately reproduce qualitatively distinct dynamical behaviors generated by the epidemiological system.

Logistic-Map Reservoir Computer predictions for two different state variables of the SIRD model when the disease-free steady state is stable. Solid curves denote true trajectories, while dashed curves represent reservoir predictions over the training and forecasting intervals.

Logistic-Map Reservoir Computer predictions for two different state variables of the SIRD model when the disease-free steady state is unstable. Solid curves denote true trajectories, while dashed curves represent reservoir predictions over the training and forecasting intervals.
Figure 9 displays the combined training and testing results for two state variables of the SIRD model when the disease-free steady state is asymptotically stable. Figure 10 displays the combined training and testing results for two state variables of the SIRD model when the model exhibits sustained oscillatory dynamics (stable limit cycle). For both scenarios, the training phase is performed over the interval \(t\in [10,30)\), after which the trained reservoir is used to forecast the system evolution on the unseen interval \(t\in [30,80)\). The values of Q and M are set at 3 and 100 respectively. In addition, \(\alpha _{min}=3.6\) and \(\alpha _{max}=4\) in numerical simulations. The obtained results in the figures show that the LMRC prediction is virtually indistinguishable from the true SIRD trajectory throughout both phases. Moreover, the reservoir accurately captures the nonlinear oscillations, preserving both phase and amplitude without visible distortion or drift. This excellent agreement is further supported by small error values, with training and testing RMSEs which are found to be S(t) Training RMSE = 0.00125003, S(t) Test RMSE = 0.0012479, D(t) Training RMSE = 0.0000063, and D(t) Test RMSE = 0.0000079, in Fig. 9. Also, it is determined that for Fig. 10: S(t) Training RMSE = 0.000278637, S(t) Test RMSE = 0.000246156, I(t) Training RMSE = 0.00013731, and I(t) Test RMSE = 0.000134219.
The results in the figures clearly demonstrate that the proposed LMRC framework can accurately learn and forecast multiple state variables of the SIRD model, even when these variables exhibit different dynamical behaviors. The very low RMSE values confirm that the reservoir acts as a high-efficient predictor of the underlying nonlinear system. From an applied perspective, LMRC approach is found to be attractive for data-driven analysis of epidemiological models, enabling efficient long-horizon prediction and real-time emulation of complex compartmental dynamics. Table 4 reports examples of the execution time (computational cost) and prediction accuracy of well-known conventional forecasting methods for the susceptible population S(t) of the proposed SIRD model in the more complicated case of oscillatory behavior. Several important observations can be drawn from these results. First, the Logistic-Map Reservoir Computer (LMRC) achieves the smallest testing errors among all methods, see Fig. 9, with RMSE values on the order of \(10^{-4}\). This indicates an excellent capability to learn and generalize the underlying deterministic dynamics of the SIRD system. Although the LMRC requires slightly more execution time than the standard Echo State Network (ESN), its substantially improved test accuracy highlights a favorable accuracy–efficiency trade-off. Second, the ESN exhibits a very small training error, comparable to that of the LMRC, while requiring the lowest computational time among all methods. However, its test RMSE increases by more than an order of magnitude relative to its training error, revealing a noticeable degradation in generalization performance. Third, the recurrent neural network baselines based on gated architectures of Recurrent Neural Networks (RNNs), namely GRU (Gated Recurrent Unit) and LSTM (Long Short-Term Memory), show significantly larger training and testing errors. Despite their higher representational capacity and longer training times, both models fail to accurately reproduce the SIRD dynamics over the prediction horizon.
Consequently, these results demonstrate that reservoir-based approaches, and in particular the LMRC, are better suited for short- and medium-term forecasting of deterministic epidemic models than deep recurrent neural networks, offering superior accuracy with substantially lower computational complexity.

Training and testing predictions of the susceptible population S(t) using different data-driven methods. In all panels, the time axis starts at the beginning of the training interval. Solid lines denote the true SIRD solution, while dashed lines represent model predictions.
Numerical simulations and discussions of results
The numerical simulations performed in MATLAB, utilizing the classical fourth-order Runge-Kutta method, were crucial for validating the complex theoretical dynamics of the proposed SIRD model. While the analytical work established conditions for stability and bifurcation, the nonlinearities in the incidence and treatment functions make closed-form solutions intractable. Numerical integration bridges this gap, providing concrete visual evidence of the system’s behavior under various parameter sets. These simulations confirm the existence and properties of equilibria, illustrate the routes to bifurcation, and ultimately demonstrate the model’s practical utility for forecasting and intervention planning. Without this computational step, the rich dynamics, including multi-stability and chaos, would remain as theoretical possibilities without verification.

(a, b) Solution trajectories converge to the trivial equilibrium point E. (c, d) Solution trajectories converge to the disease-free equilibrium point \(E_0\).

Dynamic transitions as parameter p varies: (a–c) Stable limit cycle for \(p=0.5215\); (d–f) Period-doubling bifurcation leading to a 2-cycle for \(p=0.5242\); (g–i) Further period-doubling for \(p=0.5135\), indicating route to chaos.

Further progression towards chaotic dynamics: (a–c) Complex multi-periodic behavior for \(p=0.5131\); (d–f) Emergence of chaotic attractor for \(p=0.513\), confirmed by strange attractor geometry.

Bifurcation summary: (a, b) Fully developed chaotic regime for \(p=0.5127\); (c, d) Return to stable endemic equilibrium for \(p=0.37\), demonstrating the parameter-sensitive nature of system dynamics.

Chaotic attractor projections in different phase planes (\(p=0.5134, a=0.4\)): (a) Susceptible time series showing aperiodic oscillations; (b) S-I plane; (c) S-R plane; (d) I-R plane. The irregular, non-repeating patterns confirm chaotic dynamics.
Figures (12, 13, 14, 15, and 16) collectively demonstrate the remarkable richness of dynamical behaviors exhibited by the proposed SIRD model ranging from stable endemic equilibria to periodic oscillations and fully developed chaos. As the recruitment rate \(p\) is varied, Figs. 13 and 14 reveal a classical route to chaos via period-doubling bifurcations, beginning with a stable limit cycle at \(p = 0.5215\), transitioning to a 2-cycle at \(p = 0.5242\), and eventually yielding chaotic attractors at \(p = 0.513\) and \(p = 0.5127\) as shown in Figs. (14, 15, and 16). It is revealed that even a deterministic epidemiological model can generate intrinsically unpredictable outbreak patterns.
Biologically, this implies that when transmission rates are high, treatment capacity is saturated (modeled by the term \(vI/(1+bI)\)), and population turnover is rapid, disease dynamics may become fundamentally unpredictable over long time horizons.

Optimal control application in limit cycle regime (\(p=0.5142\)): Time series of (a) Susceptible, (b) Infected, (c) Recovered, (d) Deceased populations, and corresponding optimal controls (e) Prevention w(t) and (f) Treatment v(t). Controls adaptively increase during outbreak peaks to suppress oscillations.

Optimal control in multi-periodic regime (\(p=0.5135\)): Control efforts become more frequent and intense to manage the complex oscillatory dynamics, demonstrating the need for adaptive strategies.

Optimal control in chaotic regime (\(p=0.5131\)): Despite chaotic underlying dynamics, the optimal control framework successfully identifies intervention patterns that reduce disease burden, though with more irregular control profiles.

Comparison of chaotic regime with and without control (\(p=0.5134, a=0.4\)): Optimal controls significantly dampen the chaotic oscillations, reducing the amplitude of infection peaks and cumulative deaths.

Prevention-only control strategy (\(p=0.5142\)): With treatment control \(v(t)=0\), prevention efforts alone cannot fully suppress oscillations, highlighting the need for combined interventions.

Prevention-only strategy in multi-periodic regime (\(p=0.5135\)): Limited efficacy of single-intervention approach becomes more pronounced as dynamics become more complex.

Prevention-only strategy in chaotic regime (\(p=0.5131\)): Single control fails to stabilize chaotic dynamics, emphasizing the necessity of integrated control measures.

Treatment-only control strategy (\(p=0.5142\)): With prevention control \(w(t)=0\), treatment alone reduces infection but cannot prevent recurrent outbreaks due to ongoing transmission.

Treatment-only strategy in multi-periodic regime (\(p=0.5135\)): Treatment reduces severity but cannot eliminate oscillations, supporting the superiority of combined interventions.

Treatment-only strategy in chaotic regime (\(p=0.5131\)): Single-intervention approach fails to control chaotic outbreaks, demonstrating the critical importance of prevention in reducing transmission.
The detailed simulation results from Figs. 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, and 24) vividly illustrate the complex, parameter-dependent behavior of the epidemic system. Figure 10 demonstrates that for a specific critical value of the recruitment rate p, the system exhibits a stable limit cycle, confirming the theoretical prediction of a Hopf bifurcation where an endemic equilibrium loses stability. As p is finely tuned further (Figs. 11, 12, 13, and 14), the model transitions into chaotic regimes, as evidenced by the strange attractors in phase space and sensitive dependence on initial conditions. Epidemiologically, this is highly significant: it implies that in a certain parameter regime potentially reflecting high transmission and saturated healthcare long-term epidemic forecasting becomes inherently unpredictable. Small changes in population influx or contact rates can shift outcomes from predictable endemic cycles to erratic, chaotic outbreaks, complicating public health planning.
Subsequent Figs. (15, 16, 17, 18, 19, 20, 21, 22, 23, and 24) explore the impact of the optimal control strategies derived via Pontryagin’s Maximum Principle. A detailed comparison of the subfigures reveals key insights: the simultaneous application of both prevention (w(t)) and treatment (v(t)) controls (Figs. 15, 16, and 17) is most effective in suppressing infection peaks and reducing cumulative deaths. When controls are applied singularly (e.g., prevention-only in Figs. (18, 19, and20) or treatment-only in Figs. 21, 22, 23, and 24), the system’s response is notably less robust, often failing to suppress oscillations or chaos. The epidemiological significance is clear: an integrated strategy that reduces transmission while simultaneously enhancing case management is paramount for controlling diseases characterized by nonlinear incidence and treatment saturation. The time-varying nature of the optimal controls, which intensify during outbreak peaks, provides a template for efficient, adaptive resource allocation that minimizes both disease burden and intervention costs.
Summarizing the results of simulated optimal control strategies in the next points as follows:
-
1.
Numerical implementation across different dynamic regimes: We have introduced comprehensive numerical simulations demonstrating the application of optimal control across varying, complex epidemiological states: limit cycle (\(p=0.5142\)), multi-periodic (\(p=0.5135\)), and chaotic regimes (\(p=0.5131\)). The numerical results (Figs. 15, 16, 17, and 18) confirm that the optimal control framework successfully identifies intervention patterns that dampen chaotic oscillations, significantly reducing the amplitude of infection peaks and the cumulative number of deaths.
-
2.
Policy interpretation:
a- The optimal control scheme is applied for different cases illustrating the necessity of combined interventions. To provide direct policy insights, we numerically simulated the effects of implementing only a single intervention. Figures (19, 20, and 21) model a prevention-only strategy where treatment control is zero (\(v(t)=0\)). The results reveal that prevention efforts alone cannot fully suppress oscillations, and this limited efficacy becomes increasingly pronounced as the underlying dynamics become more complex. Conversely, Figs. (22, 23, and 24) depict a treatment-only strategy where prevention control is disabled (\(w(t)=0\)). While treatment mitigates infection severity, it fails to prevent recurrent outbreaks because of the ongoing, uncontrolled transmission. From a policy perspective, these specific numerical outcomes definitively support the superiority and absolute necessity of combined, integrated control measures to stabilize chaotic epidemic outbreaks.
b- The optimal adaptive public health responses are examined. In particular, the time-series profiles of the controls (w(t) and v(t)) in Figs. 15 and 16 offer a crucial policy directive regarding timing. The optimal controls adaptively increase specifically during outbreak peaks to suppress oscillations. Furthermore, as the system enters a multi-periodic regime, control efforts must become more frequent and intense to manage the complex oscillatory dynamics, directly demonstrating the need for highly responsive, adaptive public health strategies rather than static, constant policies.
Validation of the proposed model with real epidemiological data
In this subsection, we validate the proposed mathematical model using historical influenza data from Canada, spanning from the 47th week of 2017 to the 22nd week of 201838. The numerical fitting was performed using an optimization routine in MATLAB. To isolate the optimal parameter set, we minimized the Sum of Squared Errors (SSE) objective function, defined mathematically as:
Here, n is the total number of temporal observation points, \(Y_j\) represents the empirical cumulative number of infected cases recorded at week j, and \(I(t_j)\) denotes the corresponding cumulative number of infections generated by the model over the identical timeframe. The fitting procedure requires setting explicit initial conditions for the state variables. Relying on demographic records38, the population compartments at \(t=0\) were initialized as follows: a susceptible population of \(S(0) = 36,708,083\), an initial infected pool of \(I(0) = 9\), zero initially recovered individuals (\(R(0) = 0\)), and (\(D(0) = 0\)). Through this minimization process, we successfully estimated nine of the twelve fundamental model parameters. The finalized parameter set is documented in Table 5.
To visually assess the accuracy of the optimization, Fig 27 illustrates the alignment between the empirical cumulative data and the model predictions.

Cumulative infected population density.
Finally, it is important to note that the LMRC can be successfully integrated with the epidemiological model in two scenarios.
First, for the uncontrolled epidemiological model, the LMRC is employed as accurate predictive model for the epidemiological dynamics. More specifically, the values of state variables are observed for adequate time interval. Then, the LMRC is trained using this data set and utilized to forecast possible future behaviors. It is depicted that the LMRC exhibit excellent prediction capability for different types of dynamical behaviors that can be induced by the model.
Second, for the controlled epidemiological model, LMRC can be linked to the optimal control framework presented in Section 4 by developing a surrogate modeling approach that leverages LMRC to approximate the mapping from the system’s state to the optimal control actions.
The LMRC surrogate construction proceeds as follows. First, we generate a comprehensive training dataset for a representative ensemble of initial conditions and parameter values. For each solution trajectory, we record the triplet
where \(w^{*}(t_k)\) and \(v^{*}(t_k)\) are the optimal controls at discrete time points \(t_k\). The LMRC is then trained on \(\mathcal {D}\) to learn the functional relationship
where \(\mathcal {F}_{\text {LMRC}}\) denotes the trained reservoir mapping. Once trained, the LMRC surrogate provides several significant advantages. First, inference is extremely fast: given a current state \((S, I, R, D)\) and time \(t\), the LMRC produces the corresponding optimal controls \((w^{*}, v^{*})\) in milliseconds, independent of the original ODE system’s complexity. Second, the surrogate eliminates the need to solve the adjoint system and the TPBVP repeatedly, making it particularly attractive for real-time control applications.
To validate the surrogate approach, we compare the LMRC-predicted controls against the true optimal controls. Figure 28 demonstrates the excellent agreement between the surrogate predictions and the true optimal controls for different scenarios including (a-b) two control inputs \(u_1(t)=w(t); u_2(t)=v(t)\), (c)\(u_1(t)\)-only control input, and (d) \(u_2(t)\)-only control input. This reservoir computing modeling framework thus bridges the gap between rigorous optimal control theory and practical, real-time epidemic intervention, providing public health policymakers with an efficient tool suitable for instantaneous decision support (Fig 28).

The true and predicted optimal control inputs obtained by LMRC for (a, b) two control inputs case, (c)\(u_1(t)\)-only control input case, and (d) \(u_2(t)\)-only control input case.
Conclusions and future directions
The present work successfully established a comprehensive nonlinear SIRD model, incorporating dual nonlinearities in both the incidence and treatment functions to accurately reflect real-world epidemiological complexities such as behavioral saturation and resource constraints. The rigorous analytical investigation confirmed the model’s mathematical integrity and uncovered a rich spectrum of dynamical behaviors, including multiple stable states, and complex transitions driven by transcritical, saddle-node, and Hopf bifurcations. These findings underscore the potential for complex, non-trivial epidemic patterns, such as sustained oscillations and chaotic regimes, which are highly sensitive to parameter variations. A core contribution of this work is the development of an optimal control framework, derived using Pontryagin’s Maximum Principle. This framework demonstrated conclusively that integrated, adaptive strategies combining time-dependent prevention and treatment efforts are significantly more effective than static or single-measure interventions in mitigating infection peaks and minimizing the overall disease burden. This provides a clear, actionable roadmap for public health policy.
Furthermore, the study introduced a novel, highly efficient machine learning component: the Logistic-Map Reservoir Computer (LMRC). This LMRC was shown to accurately forecast the complex, nonlinear temporal evolution of the epidemic, outperforming conventional methods in both accuracy and computational efficiency. This makes the LMRC a valuable tool for real-time scenario analysis and adaptive policy adjustments. The integrated analytical, optimal control, and machine learning approach presented here offers a comprehensive toolkit for managing infectious disease outbreaks. By providing a method to predict complex dynamics and design effective, resource-aware interventions, this work directly contributes to the global effort to achieve the Sustainable Development Goal 3 (Good Health and Well-being), particularly target 3.3 on combating communicable diseases. The insights derived from this model are crucial for policymakers aiming to develop resilient public health systems and ensure preparedness for future pandemics.
Future research directions can include extending the model to fractional-order derivatives to incorporate memory effects, incorporating spatial heterogeneity through reaction-diffusion frameworks, integrating real-world epidemiological data for specific diseases to enhance predictive power, and developing hybrid models that combine mechanistic modeling with advanced machine learning techniques for improved forecasting accuracy.
Data availability
The datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
References
Teklu, S. W. Mathematical analysis of the transmission dynamics of COVID-19 infection in the presence of intervention strategies. J. Biol. Dyn. 16(1), 640–664 (2022).
Liu, B. et al. Mathematical assessment of monkeypox disease with the impact of vaccination using a fractional epidemiological modeling approach. Sci. Rep. 13(1), 13550 (2023).
Khajanchi, S., Bera, S. & Roy, T. K. Optimal control and stability analysis of a fractional-order epidemic model with nonlinear incidence and treatment. Chaos Solitons Fractals 167, 113046 (2023).
Khan, A. A., Amin, R., Ullah, S., Sumelka, W. & Altanji, M. Numerical simulation of a Caputo fractional epidemic model for the novel coronavirus with the impact of environmental transmission. Alex. Eng. J. 61(7), 5083–5095 (2022).
Elsonbaty, A., Adel, W., Sabbar, Y. & El-Mesady, A. Nonlinear dynamics and optimal control of a fractional order cotton leaf curl virus model incorporating climate change influences. Part. Differ. Equ. Appl. Math. 10, 100727 (2024).
Saha, P. & Ghosh, U. Global dynamics and control strategies of an epidemic model having logistic growth, non-monotone incidence with the impact of limited hospital beds. Nonlinear Dyn. 105(1), 971–996 (2021).
Lashari, A. A. Optimal control of an SIR epidemic model with a saturated treatment. Appl. Math. Inf. Sci. 10(1), 185 (2016).
El-Mesady, A., Ahmed, N., Elsonbaty, A. & Adel, W. Transmission dynamics and control measures of reaction–diffusion pine wilt disease model. Eur. Phys. J. Plus 138(12), 1078 (2023).
Zhang, T., Kang, R., Wang, K. & Liu, J. Global dynamics of an SEIR epidemic model with discontinuous treatment. Adv. Differ. Equ. 2015(1), 361 (2015).
Cao, Q., Liu, Y. & Yang, W. Global dynamics of a diffusive SIR epidemic model with saturated incidence rate and discontinuous treatments. Int. J. Dyn. Control 10(6), 1770–1777 (2022).
Hu, Y., Wang, H. & Jiang, S. Analysis and optimal control of a two-strain SEIR epidemic model with saturated treatment rate. Mathematics 12(19), 3026 (2024).
Higazy, M. et al. Theoretical analysis and computational modeling of nonlinear fractional-order victim-two predators model.. Results Phys. 32, 105139 (2022).
El-Mesady, A., Aldakhil, A. & Elsonbaty, A. On nonlinear dynamical analysis of a fractional-order two-strains Nipah virus model.. Part. Differ. Equ. Appl. Math. 11, 100900 (2024).
Jose, S. A., Yaagoub, Z., Joseph, D., Ramachandran, R. & Jirawattanapanit, A. Computational dynamics of a fractional order model of chickenpox spread in Phuket province. Biomed. Signal Process. Control 91, 105994 (2024).
Shah, K., Alrabaiah, H., Zeb, A. & Ullah, A. Dynamics and optimal control of a fractional-order monkeypox epidemic model with quarantine and vaccination. Alex. Eng. J. 86, 202–217 (2024).
Bonyah, E., Badu, K. & Asamoah, J. K. K. Fractional optimal control dynamics of coronavirus model with symptomatic and asymptomatic infections. Sci. Rep. 13, 22458 (2023).
Elsadany, A. A., Sabbar, Y., Adel, W. & El-Mesady, A. Dynamics of a novel discrete fractional model for maize streak epidemics with linear control. Int. J. Dyn. Control 13(1), 5 (2025).
Teklu, S. W. Impacts of optimal control strategies on the HBV and COVID-19 co-epidemic spreading dynamics. Sci. Rep. 14(1), 5328 (2024).
Teklu, S. W. et al. Bifurcation and optimal control analysis for a fractional-order model of drug-resistant HBV infection. Comput. Biol. Med. 198, 111209 (2025).
Gümüş, M., Teklu, S. W. & Sezgin, A. Optimal intervention design for tonsillitis transmission via compartmental modeling with stability analysis and control strategies. Sci. Rep. 15(1), 27737 (2025).
Madani, Y. Synergistic malaria control through memory effects, seasonal awareness, and multi-strategy optimization in a fractional-order modeling framework. Boundary Value Probl. https://doi.org/10.1186/s13661-026-02223-x (2026).
Ndendya, J. Z. & Liana, Y. A. Mathematical modeling and analysis of the co-dynamics of pneumonia and malnutrition in children under five years. Microbe 8, 100489 (2025).
Liana, Y. A., Ndendya, J. Z. & Shaban, N. The nutritional nexus: Modeling the impact of malnutrition on TB transmission. Sci. Afr. 27, e02516 (2025).
Msigwa, A. I. & Ndendya, J. Z. A pair-formation model for hepatitis B virus transmission dynamics. Microbe 11, 100692 (2026).
Ullah, M. S., Islam, M. S. & Wang, J. Modeling and simulating cholera transmission dynamics with evolutionary game theory. Numer. Algebra Control Optim. 18, 208 (2025).
Akter, M. et al. An innovative fractional-order evolutionary game theoretical study of personal protection, quarantine, and isolation policies for combating epidemic diseases. Sci. Rep. 14(1), 14464 (2024).
Kabir, K. A., Islam, M. S. & Sharif Ullah, M. Understanding the impact of vaccination and self-defense measures on epidemic dynamics using an embedded optimization and evolutionary game theory methodology. Vaccines 11(9), 1421 (2023).
Ndendya, J. Z. & Liana, Y. A. A deterministic mathematical model for conjunctivitis incorporating public health education as a control measure. Model. Earth Syst. Environ. 11(3), 216 (2025).
Ndendya, J. Z., Mwasunda, J. A. & Mbare, N. S. Modeling the effect of vaccination, treatment and public health education on the dynamics of norovirus disease. Model. Earth Syst. Environ. 11(2), 150 (2025).
Ndendya, J. Mathematical modeling of culling and vaccination for dog rabies disease transmission with optimal control and sensitivity analysis approach. Clin. Mol. Epidemiol. 2, 2 (2025).
Ndendya, J. Z., Mlay, G. & Rwezaura, H. Mathematical modelling of COVID-19 transmission with optimal control and cost-effectiveness analysis. Comput. Methods Programs Biomed. Update 5, 100155 (2024).
Arun, R., Sathish Aravindh, M., Venkatesan, A. & Lakshmanan, M. Reservoir computing with logistic map. Phys. Rev. E 110(3), 034204 (2024).
Shah, K., Khan, A., Abdeljawad & Thabet, E. Using deep neural network and fractional derivative to investigate evolution of virulence (2026).
Mamun-Ur-Rashid Khan, M. & Tanimoto, J. A new concept of optimal control for epidemic spreading by vaccination: Technique for assessing social optimum employing Pontryagin’s maximum principle. AIP Adv. 15(7), 075018 (2025).
Perko, L. Differential Equations and Dynamical Systems (Springer, 2013).
Pontryagin, L. S., Boltyanskii, V. G., Gamkrelidze, R. V. & Mishchenko, E. F. The Mathematical Theory of Optimal Processes (Wiley, 1962).
Lenhart, S. & Workman, J. T. Optimal Control Applied to Biological Models (Chapman and Hall/CRC, 2007).
Nikbakht, R., Baneshi, M. R., Bahrampour, A. & Hosseinnataj, A. Comparison of methods to estimate basic reproduction number\((R_0)\) of influenza, using Canada 2009 and 2017–18 A (h1n1) data. J. Res. Med. Sci. 24, 67 (2019).
Saha, P., Mondal, B. & Ghosh, U. Dynamical behaviors of an epidemic model with partial immunity having nonlinear incidence and saturated treatment in deterministic and stochastic environments. Chaos Solitons Fractals 174, 113775 (2023).
Acknowledgements
The authors extend their appreciation to Prince Sattam bin Abdulaziz University for funding this research work through the project number (PSAU/2025/01/39026).
Funding
The authors extend their appreciation to Prince Sattam bin Abdulaziz University for funding this research work through the project number (PSAU/2025/01/39026).
Author information
Authors and Affiliations
Contributions
All authors contributed equally. All authors have read and agreed to the submitted version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Human and animal participants
This article does not contain any studies with human participants performed by any of the authors.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Elsonbaty, A., Ramaswamy, R., Padmaja, S. et al. Analysis, control, and forecasting the dynamics of SIRD models with saturated treatment and nonlinear incidence. Sci Rep 16, 15459 (2026). https://doi.org/10.1038/s41598-026-52772-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-52772-5
