
Abstract
Alcoholic liver disease (ALD) is a disease with high incidence, limited therapies, and poor prognosis. The present study aims to investigate the effect of riboflavin on ALD and explore its potential therapeutic mechanisms. C57BL/6 mice were divided into the control, alcohol, and alcohol+ riboflavin groups. 16S rRNA-seq and RNA-seq analysis were utilized to analyze the polymorphism of intestinal microbiota and the transcriptome heterogeneity respectively. KEGG and GO enrichment analysis were performed. CIBERSORTx was applied to evaluate the immune cell infiltration level. Publicly available transcriptome data of ALD was enrolled and combined with the RNA-seq data to identify the immune subtypes of ALD. Pathological and histology analysis demonstrated that riboflavin reversed the progression of ALD. 16S rRNA-seq results showed that riboflavin could regulate alcohol-induced intestinal microbiota alteration. Intestinal microbiota polymorphism analysis indicated that VLIDP may contribute to the progression of ALD. Based on the VLIDP pathway, two subtypes were identified. Immune microenvironment analysis indicated that the upregulated inflammatory factors may be important regulators of ALD. In conclusion, intestinal microbiota homeostasis was associated with the protective effect of riboflavin against ALD, which was likely mediated by modulating inflammatory cell infiltration. Riboflavin emerges as a promising therapeutic candidate for the management of ALD.

Similar content being viewed by others

Introduction
Alcoholic liver disease (ALD) is a kind of serious liver diseases induced by alcohol, which was associated with increased death risk of acute liver failure in clinic1,2,3,4. There were therapeutic approaches for ALD during clinical practice including alcohol withdrawal, surveillance for infections and antibiotic therapy and liver transplantation5. However, the therapy strategies could not satisfy the clinical demands. Therefore, it is necessary to explore effective therapeutic methods for ALD.
Vitamin plays pivotal roles in ALD. Vitamin E has hepatoprotective effects against liver injury induced by chronic alcohol exposure6. Vitamin C dramatically attenuates neutrophil infiltration thus relieving alcohol-induced liver injury7. Riboflavin, also known as vitamin B2, is a member of the vitamin B family8. Riboflavin depletion ariboflavinosis can induce liver and intestinal inflammation9. Sanches et al. found that riboflavin can protect against ischemia and reperfusion-induced liver injury in mice10. A study showed that riboflavin affects the development of parenteral nutrition-associated liver disease through modulating the mitochondrial electron transport chain11. Riboflavin can ameliorate liver fibrosis via regulating the AMPK/PGC1α/HO‑1 signaling pathway in rats12. Furthermore, our previous in vitro experiments found that riboflavin has protective effect on alcohol-treated liver cells13. However, the anti-ALD effect of riboflavin in vivo and the underlying mechanisms remain unelucidated.
Intestinal microbiota plays important roles in many liver diseases, including ALD14, nonalcoholic fatty liver disease (NAFLD)15,16,17, and hepatocellular carcinoma (HCC)18. Some agents were shown to protect liver injury by regulating intestinal microbiota. LV et al. found that ganoderic acid A can improve lipid metabolism and modulate the intestinal microbiota composition thus attenuating ALD19. Flavonoids extract of Hippophae rhamnoides L. can ameliorate ALD through the regulation of intestinal microbiota imbalance20.
The aforementioned findings indicate that the intestinal microbiota could serve as a promising therapeutic target for ALD. Our present study aims to explore the effects of riboflavin on ALD and related alteration in intestinal microbiota homeostasis.
Materials and methods
Animal model establishment and treatment
C57BL/6 mice (male, 20–22 g, 8 weeks) were purchased from Liaoning Changsheng Biotechnology Co., Ltd. The mice were randomly divided into three groups, including control, alcohol, and alcohol + riboflavin (AR) groups. The control group was fed with control liquid diet (bio-serv, USA), alcohol and AR group were fed with alcoholic liquid diet (bio-serv, USA) for 6 weeks to induce ALD model21. At the same time, AR group was administered with riboflavin 12.5 mg/kg by gavage every day for 6 weeks. For alcohol-treated mice, we gradually increased the volume ratio of alcohol in liquid feed to make the mouse adapt to the alcohol liquid diet according to the instructions of manufacture’s introductions (bio-serv, USA). For the first week (7 days), the volume ratio of alcohol liquid feed and control liquid feed was 1:2 for the first 3 days, and the volume ratio of alcohol liquid feed and control liquid feed was 2:1 for the last 4 days. The experimental procedure was pre-approved by the Experimental Animal Ethic Committee of College of Pharmacy, Harbin Medical University, China (No. IRB3016722).
Histological analysis
Liver tissues were fixed with 4% paraformaldehyde overnight, dehydrated and paraffin-embedded. Then the liver specimens were cut into 5-μm thick slices. Following that, the tissue samples of paraffin slices were stained with Hematoxylin-Eosin/HE Staining Kit (WANLEIBIO, China), according to the manufacturer’s instructions. All images of slices were captured by a light microscope.
Determination of liver biochemical indicators
Liver alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were detected according to the manufacture’s instructions using Alanine aminotransferase Assay Kit (Jiancheng Company, China) and Aspartate aminotransferase Assay Kit (Jiancheng Company, China), respectively22,23. Liver total cholesterol (TC) and triglyceride (TG) contents were detected according to the manufacture’s instructions using TG kit (Solarbio Inc, China) and TC kit (Solarbio Inc, China), respectively24,25.
16S rRNA sequencing
The 16S rRNA sequencing (16S rRNA-seq) was detected by Gene Denovo company, China. Microbial DNA was extracted from stool samples using the HiPure Stool DNA Kits (Magen, China) according to the manufacturer’s instructions. The V3 and V4 regions of the 16S rRNA gene were amplified using PCR (Eastwin scientific equipmentsinc, China). The 341F Primer sequence: CCTACGGGNGGCWGCAG. The 806R Primer sequence: GGACTACHVGGGTATCTAAT. AMPure XP Beads (Beckman, USA) was used to purify DNA according to the manufacturer’s instruction. Illumina DNA Prep Kit (Illumina, USA) was used to construct the cDNA library. The Qubit 3.0 (Thermo Fisher Scientific, USA) was used for the quantification of cDNA library. ABI StepOnePlus Real-Time PCR System (Life Technologies, USA) was used to measure the quality of cDNA library. A novaseq 6000 (Illumina, USA) was used for sequencing under PE250 model and got the 16S rRNA-seq date that was used for the further analysis The data was shown in Supplementary Table 1.
RNA sequencing
The RNA sequencing (RNA-seq) was detected by Shanghai Outdo Biotech company, China. Mouse liver tissue RNA was extract by Trizol Reagent (Invitrogen Life Technologies, USA). NanoDrop 2000 (Thermo Fisher Scientific, USA) and the agarose gel electrophoresis measure the quality of the total RNA. mRNA was purified from total RNA using poly-T oligo-attached magnetic beads. Fragmentation was carried out using divalent cations under elevated temperature in an Illumina proprietary fragmentation buffer. First strand cDNA was synthesized using random oligonucleotides and Super Script II. Second strand cDNA synthesis was subsequently performed using DNA Polymerase I and RNase H. Remaining overhangs were converted into blunt ends via exonuclease/polymerase activities and the enzymes were removed. After adenylation of the 3′ ends of the DNA fragments, Illumina PE adapter oligonucleotides were ligated to prepare for hybridization. To select cDNA fragments of the preferred 400-500 bp in length, the library fragments were purified using the AMPure XP system (Beckman Coulter, USA). Qubit® 2.0 Fluorometer (Thermo Fisher Scientific, USA) was used to detect the concentration of the cDNA library. Bioanalyzer 2100 system (Agilent, USA) was used to measure the size and quantification of the cDNA library. NovaSeq 6000 (Illumina, USA) was used for sequencing of the cDNA library under PE150 model and got the RNA-seq data The data was shown in Supplementary Table 2.
Analysis of functional differences in intestinal microbiota
To analyze the different relative abundances of bacterial taxa within different groups, Krona was used to visualize the abundance statistics for each taxa and the R package “ggplot2” was used to visualize community composition in stacked bar plots. The abundance of species was represented in circular layouts using Circus. Welch’s t-test was employed to compare species calculated by “Vegan” package between groups. The pathway activity of OTUs (Operational Taxonomic Units) were analyzed via PICRUSt. Welch’s t-test was used to determine whether there are functional differences between groups.
Data acquisition
Transcriptome data for GSE28619 and clinic information was acquired from the GEO database. The data was transformed in to the Log2 (TPM + 1)13 (Supplementary Fig. S1). Integration with the ALD cohort. Meta-cohort was constructed based on the integration of GSE28619 and our in-housed RNA-seq data by R package “sva”.
The biological alteration was mediated by colon microbial polymorphism. 16S rRNA-seq is the extraction of the sample total DNA, followed by amplification of the target fragment region, sequencing, and then data analysis for the diversity of microbial communities in the sample. After obtaining reads, multiple data processing processes were carried out. Tag splicing and OTUs clustering are used for strict quality control. According to the species annotation and abundance information of the OTUs, the KEGG pathway functional annotation in the PICRUSt2 software bacteria/archaea (16S) was conducted. The abundance information of each pathway was counted. GSVA algorithm was employed to evaluate the pathway activity based on the transcriptome data.
ALD subtypes identification
Consensus clustering analysis was applied to identify the VLIDPA (Valine, leucine and isoleucine degradation pathway active) subtype and VLIDPI (Valine, leucine and isoleucine degradation pathway inactive) subtype based on Valine, leucine and isoleucine degradation pathway (VLIDP). VLIDP signatures were obtained from the KEGG database (https://www.kegg.jp/). Relatively stable clusters were determined via the Cumulative distribution function (CDF) curve. The process was executed using the Consensus Cluster Plus R package26. In order to assess the activity of VLIDP to validated the robustness of identified subtypes, we used the GSVA R package27. This package allows for gene set enrichment analysis based on a non-parametric, unsupervised method, providing a continuous score representing the pathway activity for each sample.
Differential expressed genes analysis
Differential expression analysis was performed by limma R package. The differentially expressed genes (DEGs) were obtained in the threshold of |log2FC| > 1 and adjusted P < 0.05. Volcano plot of the DEGs were presented using ggplot2 R packages.
Functional enrichment analysis
Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis was conducted by R package “cluster Profiler”28. KEGG pathway enrichment get functional pathways that affect ALD29. Gene Ontology (GO) analysis includes biological processes, cellular components and molecular functions, enabling us to determine the functional differences between the two subtypes30. Disease ontology (DO) analysis was conducted to obtain the related disease enrichment results. The biological pathways and processes were identified by Reactome analysis.
Characterization of inflammation immune microenvironment
CIBERSORTx algorithm was applied to evaluate the infiltration level of 22 immune cells in the meta data set. Analysis was conducted based on the transcriptome data of immune cell acquired from CIBERSORTx (https://cibersortx.stanford.edu/). Immune infiltration data was visualized via R package “Vioplot”.
Statistics and reproducibility
Data were presented as mean ± standard error of mean (SEM). Statistical comparison among three or more groups was analyzed by one-way ANOVA and Kruskal-Wallis Test. Species comparisons and functional differences between two groups were calculated by Welch’s t-test and Wilcoxon Rank Sum Test. A two-tailed P < 0.05 was considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Results
Riboflavin could protect against ALD
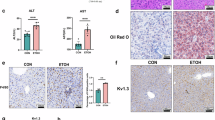
A flowchart was shown in Fig. 1a, which showed the process of mouse treatment. The results showed that alcohol induced liver steatosis, and riboflavin alleviated the effect of alcohol. The morphological observation of the liver showed that riboflavin inhibited alcohol-induced liver injury (Fig. 1b). The rate of mouse liver weight to the body weight result shown that alcohol increased the rate, which was decreased by riboflavin (Fig. 1c). HE staining showed that alcohol induced steatosis and inflammatory cell infiltration in liver tissues, which was alleviated by riboflavin (Fig. 1d). In addition, ALT, AST, TG, and TC levels were higher in the alcohol group than in the control group, which was attenuated by riboflavin (Fig. 1e–h). These results suggest that riboflavin alleviates ALD in mice.

a A flowchart showed the whole process of mouse treatment. b The representative diagram of liver tissues shown that alcohol induced steatosis changes, and riboflavin alleviate the effect of alcohol diet. The black arrow in the figure was to show the steatosis changes part of the liver. c The rate of liver weight to the body weight result shown that alcohol increased the rate of mouse liver weight to the body weight, which was decreased by riboflavin. d HE staining representative images (400×) shown the hepatic steatosis and inflammatory cell infiltration of the alcohol groups mouse livers, which was alleviated by riboflavin. e ALT level was upregulated in the alcohol induced mouse liver tissues, which was alleviated by riboflavin. f AST level was upregulated in the alcohol induced mouse liver tissues, which was alleviated by riboflavin. g TG level was upregulated in the alcohol induced mouse liver tissues, which was alleviated by riboflavin. h TC level was upregulated in the alcohol induced mouse liver tissues, which was alleviated by riboflavin. *P < 0.05, **P < 0.01, ***P < 0.001vs. Control; #P < 0.05, ###P < 0.001 vs. Alcohol. n = 5. Data were presented as mean ± standard error of mean (SEM). AR alcohol + riboflavin, ALD alcoholic liver disease.
The effect of alcohol and riboflavin on mouse intestinal microbiota compositions
The relative abundance of OTUs and taxonomic groups can distinguish the differences in bacterial profiles among groups. The species abundance of intestinal microbiota in mice treated with alcohol or riboflavin was different (Fig. 2a). The Venn diagram showed that there were 68 species of bacteria at the intersection of the three groups (Fig. 2b). Biomarkers as taxonomic units in different groups were explored by LEfSe analysis (Fig. 2c, d). According to the results, there were 12, 11, and 19 species of bacteria that were the most abundant in the control group, alcohol group and AR group, respectively (Fig. 2c). And the results indicated that there were differences in intestinal microbiota composition among all groups (Fig. 2d). Then we compared the differences in bacteria abundance among the three groups. These results showed that alcohol downregulated the abundance of 5 bacteria species (Akkermansia, Leuconostoc, Clostridium sensu stricto 13, Rikenella and Harryflintia), and upregulated the abundance of the 10 bacteria species (Corynebacterium 1, Acinetobacter, Escherichia-Shigella, Sporosarcina, Clostridium sensu stricto 1, Lactobacillus, Bifidobacterium, Candidatus Stoquefichus, Christensenellaceae R-7 group and Romboutsia), which effect was alleviated by riboflavin (Fig. 3). These results showed that alcohol could regulate the abundance of intestinal microbiota, which could be alleviated by riboflavin.

a The stacked bar plots of the relative proportion of the top 10 most abundant bacterial genera. b The Venn diagram of shared and unique bacterial genera among the three groups. c Biomarker taxon generated from LEfSe analysis (LDA > 4). d Taxonomic differences of phylum, class, order, family, and genus. AR Alcohol + riboflavin, ALD Alcoholic liver disease.

a–e Alcohol decreased abundance of intestinal microbiota compared with control group, which was alleviated by riboflavin. f–o Alcohol increased the abundance of intestinal microbiota compared with control group, which was alleviated by riboflavin. AR alcohol + riboflavin, ALD alcoholic liver disease.
Intestinal microbiota abundance heterogeneity mediated biology function
We then analyzed the characteristics of intestinal microbiota. The differential bacteria species between control and alcohol groups, and between alcohol and AR groups were shown in Fig. 4a, b. These bacteria species maybe potentially serve as biomarkers for alcohol consumption. KEGG analysis was used to predict the enriched metabolic pathways, and to evaluate the potential function of intestinal microbiota. The results of KEGG enrichment analysis showed that there were 46 pathways identified between the control and alcohol groups, and 59 pathways identified between alcohol and AR groups (Fig. 4c, d).

a Random forest contribution scatter plot of different intestinal microbiota between control and alcohol groups. b Random forest contribution scatter plot of different intestinal microbiota between alcohol and AR groups. c The PICRUSt-generated KEGG pathway functional profiles of intestinal microbiota between control and alcohol groups. d The PICRUSt-generated KEGG pathway functional profiles of intestinal microbiota between alcohol and AR groups. The bar presents the 95% confidence intervals. AR alcohol + riboflavin, ALD alcoholic liver disease.
Identification of VLIDPA and VLIDPI subtype in ALD
We further analyzed the important pathways that involved in ALD and the treatment of riboflavin. The results of 16S rRNA-seq showed that 46 pathways were associated with ALD, and 59 pathways were associated with the treatment of riboflavin. There were 14 pathways identified among three groups (Fig. 5a). Based on the RNA-seq data, our results showed that there were 82 pathways identified between the control and alcohol groups, and 47 pathways identified between alcohol and AR groups. There were 21 pathways identified among three groups (Fig. 5b). Then we found that there was an overlapped pathway between 16S rRNA-seq and RNA-seq data, which was VLIDP (Fig. 5c). The KEGG analysis results were shown in Supplementary Table 3 and Supplementary Table 4. The GSVA scores showed the activity of the pathway among three groups (Fig. 5d). Then we clustered the samples using the CDF curve. According to the CDF curve and the area under the curve, the optimal number of clusters was 2 (Fig. 5e, f). Two subtypes were identified based on the K-Means clustering, which were well identified (Fig. 5g). The identified two ALD subtypes showed different VLIDP activity. The ALD subtype with high activity was named as the VLIDPA and the another one with low activity was named as the VLIDPI (Fig. 5h).

a The upset diagram of shared and specific bacterial genera among the three groups. b The upset diagram of shared and specific pathway among the three groups. c The Venn diagram of the enriched pathway number between 16S rRNA-seq and RNA-seq data. d Comparison of GSVA scores among the three groups. e CDF curve for the consensus clustering analysis. f Relative change in area under CDF curve for the consensus clustering analysis. g The heatmap showed that the two subtypes were successfully structured. h Boxplot of GSVA scores between two subtypes. *P < 0.05,**P < 0.01,***P < 0.001. AR alcohol + riboflavin, ALD alcoholic liver disease, CDF cumulative distribution function.
The enrichment analysis of the DEGs between VLIDPA and VLIDPI
We then screened the DEGs between VLIDPA and VLIDPI subtypes. A total of 867 DEGs were identified, with 292 upregulated and 575 downregulated (Fig. 6a). We then conducted the enrichment analysis on DEGs based on KEGG, GO, DO and Reactome databases (Fig. 6b and Fig. S2). According to KEGG enrichment analysis, we found that these DEGs were involved in signaling pathways such as PI3K-Akt signaling pathway, focal adhesion, and ECM-receptor interaction. According to GO enrichment analysis, these DEGs are mainly enriched in extracellular matrix structural constituent, endoplasmic reticulum lumen, fatty acid metabolic process, basement membrane and other signaling pathways. Reactome enrichment results showed that DEGs were mainly enriched in the extracellular matrix organization, metabolism, interleukin-4 and interleukin-13 and other signaling pathways. DO enrichment analysis found that DEGs were mainly enriched in liver cancer, which implied that the DEGs may be involved in the initiation of liver cancer. It was in accordance with a study that alcohol was a risk factor for liver diseases, which could increase the death rate of cirrhosis and liver cancer31.

a Volcano plot of DEGs between VLIDPA and VLIDPI subtypes. b KEGG enrichment analysis of DEGs. c Heat map of immune characteristics between VLIDPA and VLIDPI subtypes. *P <0.05,**P < 0.01,***P <0.001. AR Alcohol + riboflavin, ALD Alcoholic liver disease.
Characterize the immune microenvironment of ALD subtypes
To further explore the role of VLIDP in ALD, we characterize the inflammatory immune microenvironment differences in the two subtypes according to the cytokines and immune cell infiltration features (Fig. 6c). Compared with the VLIDPA subtype, the immune cell infiltration types including macrophages M0, macrophages M1, dendritic cells resting, mast cells resting were higher, and plasma cells, T cells CD4 memory resting, T cells follicular helper, macrophages M2, eosinophils were lower in VLIDPI subtype. Among them, macrophages M0, macrophages M1, dendritic cells resting, mast cells resting may play a crucial role in ALD (Fig. 7a). Macrophages may be an important regulator of ALD by immune microenvironment analysis. Detailed differential expression status of cytokines related genes between the two groups were shown in Fig. 7b. Compared with VLIDPA subtype, the inflammatory factors, such as IL15, IL18, CXCL1, CXCL10, and CXCL9, were higher in VLIDPI subtype, while IL10, IL2, CXCL14, CCL4, and CCL5 were lower in VLIDPI subtype. Finally, Sankey diagram showed the relation between the cytokines and the immune cell (Fig. 7c).

a Violin diagram of the immune infiltration difference between the VLIDPA and VLIDPI subtypes. b Box diagram of differentially expressed cytokines between the VLIDPA and VLIDPI subtypes. c Sankey diagram of the distribution relationship between immune cytokines and immune cells. *P < 0.05,**P < 0.01,***P < 0.001. AR Alcohol + riboflavin, ALD Alcoholic liver disease.
Discussion
Alcohol is a risk factor of various serious liver disease, such as liver cancer32, alcohol-induced liver injury33, cirrhosis34 and so on. Vitamins are essential element for human and are involved in the development of liver diseases35,36. Riboflavin can protect against liver injury through reducing the oxidative stress injury and regulating the balance of intestinal inflammation9,10. In addition, our previous study found that riboflavin inhibited the level of ALT and AST to exert protective effect on alcohol-treated AML-12 hepatocytes13. In the present study, we further explored the protect effect of riboflavin on ALD in vivo and explored its potential mechanisms.
We established alcohol-induced mice liver injury model. Alcohol increased liver ALT, AST, TG, TC levels compared with that in controls. This was in accordance with previous studies that alcohol induced liver injury37,38. Our results also found that the coadministration of riboflavin exerted protective effects against alcohol induced liver injury. However, the mechanism of riboflavin on ALD was still not fully understood.
With the development of intestinal microbiota research, the relationship between ALD and intestinal microbiota has been revealed. Intestinal microbiota played regulatory roles in the development of ALD. A study showed that alcohol feeding induced the dysregulation of intestinal microbiota and alcoholic steatohepatitis in mouse39. Intestinal microbiota composition was different in alcohol dependence syndrome patients with or without cirrhosis40. A study suggested that riboflavin could enhance host resistance to influenza virus by regulating the compositions of intestinal microbiota41.
To investigate the involvement of intestinal microbiota in the protective effect of riboflavin on ALD, 16S rRNA-seq was used to obtain microbiota profiles. We found that there were differences in the species compositions, abundance, species of bacteria among the three groups. To investigate the effect of bacteria, we further analyzed the abundance of bacteria compared among three groups. These results showed that alcohol diet downregulated the abundance of five bacteria species and upregulated the abundance of the ten bacteria species. While, coadministration of riboflavin alleviated the effect of alcohol on bacteria abundance.
Akkermansia has been reported to reduce acetaminophen mediated hepatotoxicity by reducing the related ALT and AST levels, maintaining intestinal barrier function, and promoting short-chain fatty acid secretion42. In addition, a study showed that Akkermansia could alleviate liver fibrosis, inflammation and liver histopathological damages induced by high-fat diet/CCl443. Our results showed that alcohol diet could downregulate the abundance of the Akkermansia, which could be alleviated by riboflavin. The above results showed that Akkermansia may be involved in alcohol-induced liver injury, and the liver protective effect of riboflavin. Corynebacterium parvum could induced liver injury44,45. Our results showed that alcohol can increase the abundance of Corynebacterium 1, which may increase the liver injury risk, and riboflavin downregulated the abundance of Corynebacterium 1.
Bifidobacterium was a kind of beneficial bacteria, which was associated with the pathogenesis and progression of liver diseases, including NAFLD46, HCC18, and ALD47. Yoon et al. found that Bifidobacterium could improve liver function in NAFLD48. Bifidobacterium could reduce the liver inflammation and oxidative stress, and could relieve the compositions of intestinal microbiota and liver injury induced by alcohol49. Therefore, these studies implied that Bifidobacterium may have protective effect on ALD. Our results showed that alcohol could increase the abundance of Bifidobacterium, which may be associated with liver injury. In addition, the abundance of Bifidobacterium was downregulated in AR group compared with alcohol group. That may due to the protective effect of riboflavin against ALD. These results showed that intestinal microbiota may play an important role in the protective effect of riboflavin against ALD.
To explore the potential mechanism of intestinal microbiota in ALD, functional enrichment analysis was performed. The results showed that there were 14 enriched pathways identified using 16S rRNA-seq data and 21 enriched pathways identified using RNA-seq data. And there was one overlapped pathway between the two kinds of sequencing data, which was VLIDP. The KEGG analysis results were shown in Supplementary Table 3 and Supplementary Table 4. Then ALD samples were divided into two subtypes based on the CDF curve, which were identified as VLIDPA and VLIDPI.
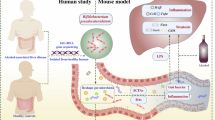
Subsequently, we performed enrichment analysis to analyze the potential biology function of intestinal microbiota. Our findings revealed that the inflammatory factors and immune cell infiltration were associated with the protective effect of riboflavin on ALD. This suggested that immune cell infiltration could be a potential underlying mechanism and play an important role in ALD. A study has demonstrated that pro-inflammatory factor IL-17 was upregulated in alcohol-induced hepatic steatosis, inflammation, fibrosis, and HCC. Inhibition of IL-17 could suppress the progression of alcohol-induced HCC50. In our results, when comparing with VLIDPA subtype, the VLIDPI subtype exhibits higher expression levels of inflammatory factors such as IL-18, CXCL1, CXCL10, and CXCL9. This was in accordance with the finding that inhibiting NLRP3 inflammasome activity and the subsequent IL-18 release could protect against ALD51. In addition, there is evidence that alcohol enhanced the expression of CXCL1 in both LO2 cells and mouse liver tissues52. These findings revealed that the protective effects of riboflavin against ALD were associated with the regulatory effect on intestinal microbiota and inflammatory cell infiltration (Fig. 8 and Supplementary Data 1). However, the exact bacteria mediating the protective effects of riboflavin remain to be elucidated.

Irregular pink circle is shown as the intestine. The red arrow is shown the effect of alcohol. Green thick line is shown the effect of riboflavin.
Conclusions
Intestinal microbiota homeostasis was associated with the protective effect of riboflavin against ALD, which effect was related to the regulatory effect on inflammatory cell infiltration. Riboflavin may be a potential effective agent for ALD.
Data availability
The 16S rRNA-seq data was shown in Supplementary Table 1. The RNA-seq data was shown in Supplementary Table 2. The transcriptome data for GSE28619 and clinic information was acquired from the GEO database. The data that support the findings of this study are available from the corresponding author upon request. All other data are available from the corresponding author (or other sources, as applicable) on reasonable request. Numerical source data for all graphs in the manuscript can be found in supplementary data 1 file. And the 16S rRNA-seq data (GSE273370) and RNA-seq data (GSE273208) are uploaded into the GEO.
Abbreviations
- ALD:
-
Alcoholic liver disease
- AR:
-
Alcohol + riboflavin
- ALT:
-
Alanine aminotransferase
- AST:
-
Aspartate aminotransferase
- CDF:
-
Cumulative distribution function
- DO:
-
Disease ontology
- DEGs:
-
Differentially expressed genes
- GO:
-
Gene Ontology
- HCC:
-
Hepatocellular carcinoma
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- NAFLD:
-
Nonalcoholic fatty liver disease
- OTU:
-
Operational Taxonomic Units
- RNA-seq:
-
RNA sequencing
- SEM:
-
Standard error of mean
- TG:
-
Triglyceride
- TC:
-
Total cholesterol
- VLIDPA:
-
Valine, leucine and isoleucine degradation pathway active
- VLIDPI:
-
Valine, leucine and isoleucine degradation pathway inactive
- VLIDP:
-
Valine, leucine and isoleucine degradation pathway
- 16S rRNA-seq:
-
16S rRNA sequencing
References
Puri, P. et al. Alcohol consumption is associated with the severity and outcome of acute liver injury/failure. Liver Int. 40, 360–367 (2020).
Fan, M. et al. Bile acid-mediated activation of brown fat protects from alcohol-induced steatosis and liver injury in mice. Cell Mol. Gastroenterol. Hepatol. 13, 809–826 (2022).
Li, F. et al. Deficiency of cathelicidin attenuates high-fat diet plus alcohol-induced liver injury through FGF21/adiponectin regulation. Cells 10, 3333 (2021).
Suneja, U. et al. Fulminant hepatic failure requiring liver transplantation due to accidental alcohol ingestion in a 7-month-old baby. Am. J. Emerg. Med. 34, 2466.e1–2466.e2 (2016).
Singal, A. K., Bataller, R., Ahn, J., Kamath, P. S. & Shah, V. H. ACG clinical guideline: alcoholic liver disease. Am. J. Gastroenterol. 113, 175–194 (2018).
Shi, Y. et al. Endoplasmic reticulum-targeted inhibition of CYP2E1 with vitamin E nanoemulsions alleviates hepatocyte oxidative stress and reverses alcoholic liver disease. Biomaterials 288, 121720 (2022).
Baek, S. M. et al. Vitamin C alleviates alcoholic liver injury by suppressing neutrophil infiltration in senescence marker protein 30-knockout mice irrespective of its antioxidant effects. Life Sci. 297, 120228 (2022).
Northrop-Clewes, C. A. & Thurnham, D. I. The discovery and characterization of riboflavin. Ann. Nutr. Metab. 61, 224–230 (2012).
Zhu, Y. Y. et al. Riboflavin bioenriched soymilk alleviates oxidative stress mediated liver injury, intestinal inflammation, and gut microbiota modification in B(2) depletion-repletion mice. J. Agric. Food Chem. 70, 3818–3831 (2022).
Sanches, S. C. et al. Riboflavin (vitamin B-2) reduces hepatocellular injury following liver ischaemia and reperfusion in mice. Food Chem. Toxicol. 67, 65–71 (2014).
Wan, S. et al. Differential metabolomic analysis of liver tissues from rat models of parenteral nutrition-associated liver disease. Biomed. Res. Int. 2020, 9156359 (2020).
Tang, N. et al. Riboflavin ameliorates mitochondrial dysfunction via the AMPK/PGC1alpha/HO‑1 signaling pathway and attenuates carbon tetrachloride‑induced liver fibrosis in rats. Exp. Ther. Med. 24, 608 (2022).
Qi, W. et al. The implementation of drug reposition for alcoholic hepatitis based on a sub-pathway integration strategy. Ann. Transl. Med. 8, 208 (2020).
Kong, L. et al. Alcoholic fatty liver disease inhibited the co-expression of Fmo5 and PPARalpha to activate the NF-kappaB signaling pathway, thereby reducing liver injury via inducing gut microbiota disturbance. J. Exp. Clin. Cancer Res. 40, 18 (2021).
Li, H. et al. Sequentially fermented dealcoholized apple juice intervenes fatty liver induced by high-fat diets via modulation of intestinal flora and gene pathways. Food Res. Int. 156, 111180 (2022).
Feng, J. et al. Marine chitooligosaccharide alters intestinal flora structure and regulates hepatic inflammatory response to influence nonalcoholic fatty liver disease. Mar. Drugs 20, 383 (2022).
Li, Y. et al. Astaxanthin alleviates nonalcoholic fatty liver disease by regulating the intestinal flora and targeting the AMPK/Nrf2 signal axis. J. Agric. Food Chem. 70, 10620–10634 (2022).
Zhang, X. et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 70, 761–774 (2021).
Lv, X. C. et al. Ganoderic acid A from Ganoderma lucidum protects against alcoholic liver injury through ameliorating the lipid metabolism and modulating the intestinal microbial composition. Food Funct. 13, 5820–5837 (2022).
Zhao, H. et al. Protective effect of flavonoids extract of Hippophae rhamnoides L. on alcoholic fatty liver disease through regulating intestinal flora and inhibiting TAK1/p38MAPK/p65NF-kappaB pathway. J. Ethnopharmacol. 292, 115225 (2022).
Mandrekar, P., Ambade, A., Lim, A., Szabo, G. & Catalano, D. An essential role for monocyte chemoattractant protein-1 in alcoholic liver injury: regulation of proinflammatory cytokines and hepatic steatosis in mice. Hepatology 54, 2185–2197 (2011).
Du, H. et al. Effects of Danhong injection on dyslipidemia and cholesterol metabolism in high-fat diets fed rats. J. Ethnopharmacol. 274, 114058 (2021).
Chen, Y. et al. Gingerenone A alleviates ferroptosis in secondary liver injury in colitis mice via activating Nrf2-Gpx4 signaling pathway. J. Agric Food Chem. 70, 12525–12534 (2022).
Huang, J., Chen, S., Cai, D., Bian, D. & Wang, F. Long noncoding RNA lncARSR promotes hepatic cholesterol biosynthesis via modulating Akt/SREBP-2/HMGCR pathway. Life Sci. 203, 48–53 (2018).
Wang, Y. et al. The hepatitis B virus promotes the progression of non-alcoholic fatty liver disease through incomplete autophagy. Free Radic. Biol. Med. 204, 326–336 (2023).
Wilkerson, M. D. & Hayes, D. N. ConsensusClusterPlus: a class discovery tool with confidence assessments and item tracking. Bioinformatics 26, 1572–1573 (2010).
Hanzelmann, S., Castelo, R. & Guinney, J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 14, 7 (2013).
Yu, G., Wang, L. G., Han, Y. & He, Q. Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS 16, 284–287 (2012).
Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30 (2000).
Gene Ontology C. Gene Ontology Consortium: going forward. Nucleic Acids Res. 43, D1049–D1056 (2015).
Liu, Y. et al. Contribution of alcohol use to the global burden of cirrhosis and liver cancer from 1990 to 2019 and projections to 2044. Hepatol. Int. 17, 1028–1044 (2023).
Xia, P., Liu, D. H., Wang, D., Wen, G. M. & Zhao, Z. Y. SLC3A2, as an indirect target gene of ALDH2, exacerbates alcohol-associated liver cancer via the sphingolipid biosynthesis pathway. Free Radic. Biol. Med. 206, 125–133 (2023).
Tien, S. et al. PTTG1 alleviates acute alcoholic liver injury by inhibiting endoplasmic reticulum stress-induced hepatocyte pyroptosis. Liver Int. 43, 840–854 (2023).
Tran, A. et al. Can alcohol control policies reduce cirrhosis mortality? An interrupted time-series analysis in Lithuania. Liver Int. 42, 765–774 (2022).
Lu, J. F. et al. Neohesperidin attenuates obesity by altering the composition of the gut microbiota in high-fat diet-fed mice. FASEB J. 34, 12053–12071 (2020).
Bell, H. et al. Retinol and retinyl esters in patients with alcoholic liver disease. J. Hepatol. 8, 26–31 (1989).
Dai, W. et al. Protection of Ficus pandurata Hance against acute alcohol-induced liver damage in mice via suppressing oxidative stress, inflammation, and apoptosis. J. Ethnopharmacol. 275, 114140 (2021).
Zhou, Z. S. et al. Fisetin ameliorates alcohol-induced liver injury through regulating SIRT1 and SphK1 pathway. Am. J. Chin. Med. 50, 2171–2184 (2022).
Yan, A. W. et al. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology 53, 96–105 (2011).
Dubinkina, V. B. et al. Links of gut microbiota composition with alcohol dependence syndrome and alcoholic liver disease. Microbiome 5, 141 (2017).
Li, Y. et al. Riboflavin attenuates influenza virus through cytokine-mediated effects on the diversity of the gut microbiota in MAIT cell deficiency mice. Front. Microbiol. 13, 916580 (2022).
Xia, J. et al. Akkermansia muciniphila ameliorates acetaminophen-induced liver injury by regulating gut microbial composition and metabolism. Microbiol. Spectr. 10, e0159621 (2022).
Keshavarz Azizi Raftar, S. et al. The protective effects of live and pasteurized Akkermansia muciniphila and its extracellular vesicles against HFD/CCl4-induced liver injury. Microbiol. Spectr. 9, e0048421 (2021).
Yokochi, S., Ishiwata, Y., Hashimoto, H., Ninomiya, F. & Suzuki, T. Hepatoprotective effect of propagermanium on Corynebacterium parvum and lipopolysaccharide-induced liver injury in mice. Scand. J. Immunol. 48, 183–191 (1998).
Arthur, M. J., Kowalski-Saunders, P. & Wright, R. Effect of endotoxin on release of reactive oxygen intermediates by rat hepatic macrophages. Gastroenterology 95, 1588–1594 (1988).
Mayneris-Perxachs, J. et al. Iron status influences non-alcoholic fatty liver disease in obesity through the gut microbiome. Microbiome 9, 104 (2021).
Lei, C. et al. Enteric VIP-producing neurons maintain gut microbiota homeostasis through regulating epithelium fucosylation. Cell Host Microbe 30, 1417–1434.e8 (2022).
Yoon, S. J. et al. Bifidobacterium-derived short-chain fatty acids and indole compounds attenuate nonalcoholic fatty liver disease by modulating gut-liver axis. Front. Microbiol. 14, 1129904 (2023).
He, Q. et al. Intake of Bifidobacterium lactis Probio-M8 fermented milk protects against alcoholic liver disease. J. Dairy Sci. 105, 2908–2921 (2022).
Ma, H. Y. et al. IL-17 signaling in steatotic hepatocytes and macrophages promotes hepatocellular carcinoma in alcohol-related liver disease. J. Hepatol. 72, 946–959 (2020).
Choudhury, A. et al. Inhibition of HSP90 and activation of HSF1 diminish macrophage NLRP3 inflammasome activity in alcohol-associated liver injury. Alcohol Clin. Exp. Res. 44, 1300–1311 (2020).
Jiang, Y. et al. Ethanol promoting the upregulation of C-X-C motif chemokine ligand 1 (CXCL1) and C-X-C Motif Chemokine Ligand 6 (CXCL6) in models of early alcoholic liver disease. Bioengineered 13, 4688–4701 (2022).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant No. 82370269), the Natural Science Foundation of Heilongjiang Province (Grant No. LH2021H018), the Heilongjiang Postdoctoral Science Foundation (Grant No. LBH-Q21134). National Conference on Undergraduate Innovation and Entrepreneurship (No. S202110226059).
Author information
Authors and Affiliations
Contributions
Jiang Y.N. and Sun H. contribute to conception and design of the study. Jiang Y.N. was the first correspondence author. Shen X.Y., Jiang Y.N., Sun H., Di Y.Z. and Li W.H. wrote or contributed to the writing of the manuscript. Shen X.Y., Shi C.P., Zhi F.N., and Ma W.J. performed the experiment. Xu J.C. and Luo K.P. did the bioinformatic analysis. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
Institutional Review Board Statement: The experiment methods were fully implemented in accordance with the Guide for the Care and Use of Laboratory Animals (NIH Publication No. 85-23, revised 1996). The experimental procedure was pre-approved by the Experimental Animal Ethic Committee of College of Pharmacy, Harbin Medical University, China (No. IRB3016722).
Peer review
Peer review information
Communications Biology thanks Mohamed Bou Saleh and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Sabina Leanti La Rosa and Joao Valente.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Shen, X., Shi, C., Xu, J. et al. Intestinal microbiota homeostasis analysis in riboflavin-treated alcoholic liver disease. Commun Biol 7, 1030 (2024). https://doi.org/10.1038/s42003-024-06722-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-024-06722-4
This article is cited by
-
Postbiotic Potential of Newly Isolated Riboflavin-Overproducing Lactiplantibacillus plantarum Strains
Probiotics and Antimicrobial Proteins (2025)
