
Abstract
The autophagy receptor p62 is degraded via autophagy under hyperosmotic stress, but whether this involves the formation of biomolecular condensates (p62 bodies) remains unclear. Using human cells, we found that p62 bodies formed within 1 minute of hyperosmotic stress, and increased with stress severity. They formed faster and under milder stress than stress granules, a classic condensate, and exhibited liquid-like properties. Unlike stress granules, p62 bodies frequently colocalized with LC3 and WIPI-2, and were degraded via autophagy. Correlative light and electron microscopy revealed that these p62 bodies were more compact than stress granules and were often associated with the autophagic isolation membrane. Autophagy receptors NBR1 and TAX1BP1, but not OPTN1 or NDP52, behaved similarly to p62, and p62 bodies preferentially contained K63-linked ubiquitin chains. p62 body formation was also observed in human epithelial organoids in association with WIPI-2. Collectively, these results indicate that p62 bodies function as a platform of degradation under hyperosmotic stress.
Similar content being viewed by others
Introduction
Human tissues are frequently exposed to drastic changes in the extracellular environment, with epithelial tissues (e.g., renal tubule and digestive mucosa) subject to severe osmotic fluctuations1. Defects in osmotic homeostasis contribute to various human diseases, making intracellular osmotic balance critical for maintaining cell function and survival2. When cells are exposed to high osmotic pressure, water diffuses out across the plasma membrane, leading to intracellular molecular crowding. This environmental condition eventually results in the accumulation of misfolded proteins, which are toxic to cells3,4,5. To maintain homeostasis, cells eliminate misfolded proteins through two major degradation systems: the ubiquitin–proteasome system and autophagy6. There are two distinct types of autophagy: nonselective bulk autophagy and selective autophagy7,8. In selective autophagy, autophagy receptors that recognize both polyubiquitinated substrates and autophagic isolation membrane mediate autophagosome formation. The established autophagy receptor p62 (also known as SQSTM1) has an LC3-interacting region (LIR) domain and a ubiquitin-associated (UBA) domain at its C-terminus. In vitro reconstitution experiments have shown that p62 is capable of forming liquid droplets in cooperation with polyubiquitin chains9,10. In mammalian cells, endogenous p62 forms cytoplasmic condensates called p62 bodies in response to oxidative or translational stress10,11. Previous studies have reported autophagy-mediated p62 degradation under hyperosmotic stress12,13,14; however, it remains unclear whether p62 body formation mediates this process.
Here, using human cell lines, we found that hyperosmotic stress rapidly induced the formation of p62 bodies containing K63-linked ubiquitinated proteins, which were subsequently engulfed by the isolation membrane for lysosomal degradation. The behavior, composition, and morphology of the p62 bodies differed from those of stress granules or proteasome foci, two condensates formed under hyperosmotic stress15,16,17,18,19. Importantly, p62 body-mediated autophagy was also observed in epithelial organoids, suggesting that this mechanism plays a role in maintaining cellular homeostasis in tissues under physiologically fluctuating extracellular osmolality.
Results
p62 bodies form more rapidly and in response to lower hyperosmotic stress than stress granules
First, we examined p62 behavior under hyperosmotic stress, comparing it with G3BP stress granule assembly factor 1 (G3BP1), a stress granule marker that undergoes phase separation20, in T24 human bladder carcinoma cell line and U2OS human osteosarcoma cells. Under normosmotic culture conditions, p62 was detected as a diffuse signal with fine and occasional large puncta in the cytoplasm, while G3BP1 mainly exhibited a diffuse signal, regardless of culture medium exchange. After exposure to hypertonic medium with increasing concentrations of sucrose (0.1–0.6 M), intense punctate signals for p62 (hereafter referred to as p62 bodies) appeared in varying numbers and sizes, depending on the sucrose concentration (Fig. 1A and Supplementary Fig. 1A). Quantitative analysis revealed that, as hypertonicity increased, the number of p62 bodies increased significantly at 0.1–0.4 M sucrose, while their size decreased at 0.4 and 0.6 M sucrose (Fig. 1B, C). G3BP1-positive puncta were not observed at 0.1 M sucrose, but did become visible at 0.2−0.6 M sucrose (Fig. 1A, B, C and Supplementary Fig. 1A). Importantly, the two structures showed little-to-no colocalization (Fig. 1A and Supplementary Fig. 1A). The same results were observed in experiments using mannitol or sodium chloride as the solute in T24 cells (Supplementary Fig. 1B). These results suggest that p62 bodies and stress granules form as distinct condensates under hyperosmotic stress, with p62 bodies forming at lower levels of hyperosmotic stress than stress granules.

A Immunostaining of T24 cells exposed to hyperosmotic stress using antibodies against p62 (PM045) and G3BP1. Cells were cultured under normosmotic conditions without medium exchange (No Me), or shifted to DMEM containing indicated concentrations of sucrose for 20 min. Boxed regions are enlarged and shown as insets. Scale bars = 20 µm and 2 µm (insets). B Quantification of the number of p62 bodies and stress granules in T24 cells and U2OS cells (images are shown in Supplementary Fig. 1A). The number of cytoplasmic p62 (green) or G3BP1 (red) puncta per area was quantified in 45 cells from three independent experiments. Data are presented as mean ± SD. Each value was statistically compared to “0.1 M” group using one-way ANOVA followed by Tukey’s test. ** P < 0.01, *** P < 0.001, NS: not significant. C Quantification of the size of p62 puncta in T24 cells and U2OS cells. A minimum of 150 puncta was measured from three independent experiments. Data are presented as mean ± SD. Each value was statistically compared to “0.1 M” group using one-way ANOVA followed by Tukey’s test. *** P < 0.001, NS not significant.
For time-course experiments in T24 and U2OS cells, we employed a hyperosmotic condition of 0.3 M sucrose, which was considered to be of sufficiently low stress to allow the detection of both p62 bodies and stress granules. Under this condition, p62 bodies appeared within 1 min, whereas stress granules were first detectable after 5 min and became distinct by 10 min, indicating that p62 bodies form more rapidly than stress granules (Fig. 2A, B and Supplementary Fig. 2A). We also observed that the number of p62 bodies decreased from 10 to 20 min (Fig. 2B). Live-cell imaging demonstrated that p62 bodies simultaneously emerged across the cytoplasm upon addition of hypertonic solution (final concentration of ~0.38 M sucrose) and their size increased over time (Supplementary Fig. 2B and Supplementary Movie 1). Moreover, careful observations revealed that a subset of p62 bodies occasionally undergo fusion (Fig. 2C and supplementary Fig. 2C), which may account for the observed decrease in the number of p62 bodies (Fig. 2B). Additionally, the fluorescence intensity of the cytoplasmic area surrounding the p62 bodies decreased (Fig. 2D and Supplementary Fig. 2D), suggesting that they grow by recruiting diffusely distributed p62. After shifting the cells to isotonic conditions, the p62 bodies disappeared within 5 min and the diffuse distribution of p62 was restored (Fig. 2E, F), indicating that they are reversible structures. These characteristics strongly suggest that p62 undergoes liquid–liquid phase separation in response to hyperosmotic stress. Given that multivalent interactions between p62 and ubiquitinated proteins promote condensate formation21, we examined p62 mutants lacking such interactions. In p62-deficient Huh-1 cells expressing green fluorescent protein (GFP)-tagged p62 proteins with mutations in the PB1 domain (K7A/D69A) or UBA domain (F406A), hyperosmotic stress-induced p62 body formation was significantly reduced (Fig. 2G). Taken together, these results indicate that p62 bodies form via phase separation under hyperosmotic stress, independently of stress granules, and form more rapidly and at lower stress intensities than stress granules.

A Immunostaining of T24 cells exposed to hyperosmotic stress using antibodies against p62 (PM045) and G3BP1. Cells were cultured under normosmotic conditions without medium exchange (No Me), or shifted to DMEM with 0.3 M sucrose for 1, 5, 10, or 20 min. Boxed regions are enlarged and shown as insets. Scale bars = 20 µm and 2 µm (insets). B Quantification of the number of p62 bodies and stress granules in T24 cells and U2OS cells (images are shown in Supplementary Fig. 2A). The number of cytoplasmic p62 (green) or G3BP1 (red) puncta per area was quantified in 45 cells from three independent experiments. Data are presented as mean ± SD. Each value was statistically compared to “No Me” group using one-way ANOVA followed by Tukey’s test. ** P < 0.01, *** P < 0.001, NS: not significant. C Time-lapse images of mCherry-p62 in U2OS cells after medium exchange with hypertonic media as described in the “Materials and methods” (a final concentration of about 0.38 M sucrose). A representative sequence of still images from the region (i), indicated in Supplementary Fig. 2B, is shown. Two additional regions, (ii) and (iii), are presented in Supplementary Fig. 2C. Arrows indicate fusion events between p62 bodies. See also Supplementary Movie 1. Scale bar = 2 μm. D Quantification of fluorescence intensity of p62 body-free cytoplasm. Cytoplasmic areas devoid of p62 bodies were randomly selected (n = 10, representative regions shown in Supplementary Fig. 2D) and analyzed using Fiji ImageJ. Data are presented as mean ± SD. Each value was statistically compared to “2 min” group using one-way ANOVA with Tukey’s test. *P < 0.05, ***P < 0.001. E Immunostaining of T24 cells following osmotic stress release. Cells were cultured in DMEM with 0.4 M sucrose for 20 min (no wash), followed by a shift to isotonic DMEM for 1 or 5 min, and fixed for immunostaining with anti-p62 antibodies (PM045). Boxed regions are enlarged and shown as insets. Scale bars = 20 µm and 2 µm (insets). F Quantification of the number of p62 bodies in T24 and U2OS cells following osmotic stress release. The number of cytoplasmic p62 puncta per area was quantified in 30 cells from three independent experiments. Data are presented as mean ± SD. Each value was statistically compared to “no wash” group using one-way ANOVA followed by Tukey’s test. *** P < 0.001. G p62 body formation in p62 mutant-expressing Huh-1 cells. p62-knockout (p62KO) Huh-1 cells were transfected with GFP-tagged wild-type (WT) or mutant (K7A/D69A or F406V) p62, and incubated in DMEM without medium exchange (No Me) or with 0.3 M sucrose for 20 min. Boxed regions are enlarged and shown as insets. Scale bars = 20 µm and 2 µm (insets).
p62 bodies are degraded by autophagy under hyperosmotic stress
Next, we examined the relationship between autophagy and the two condensates, p62 bodies and stress granules, under hyperosmotic conditions in T24 and U2OS cells. Consistent with previous studies12,13,14,22, punctate structures of the autophagosome marker LC38 appeared under hyperosmotic stress, increasing in number with increasing osmolarity (Fig. 3A, C and Supplementary Fig. 3A). Double immunofluorescence microscopy for LC3 and p62 showed that 70–80% of p62 bodies overlapped with LC3 at 0.1 M sucrose, and this fraction decreased to ~30% at 0.4 M sucrose (Fig. 3A and Supplementary Fig. 3A, D). This decrease was statistically significant in T24 and U2OS when the extent of colocalization was quantified by Manders’ coefficients (Fig. 3D). We then examined to what extent LC3-positive puncta colocalize with p62 bodies or G3BP1. Double IF staining for LC3 and p62 or G3BP1 at 0.4 M sucrose, a condition that induces obvious stress granule formation, showed that a large proportion of LC3-positive puncta colocalized with p62 bodies, whereas they were rarely associated with G3BP1-positive puncta (Fig. 3A, B and Supplementary Fig. 3A, B). The difference between p62 and G3BP1 was statistically significant in T24 and U2OS, as quantified by Manders’ coefficients (Fig. 3E). This colocalization of LC3 with p62 was also observed in experiments using mannitol or sodium chloride in T24 cells (Supplementary Fig. 3C). In time-course experiments examining the appearance of isolation membrane/autophagosomes using the marker WIPI-2 in T24 cells, WIPI-2 was detected on p62 bodies at 10 min, but not at 5 min, post osmotic challenge (Supplementary Fig. 3E). Given that p62 body formation began within 1 min (Fig. 2A and Supplementary Movie 1), this result suggests that p62 body formation is followed by the association of the isolation membrane. Western blot (WB) analysis showed that, while p62 protein levels significantly decreased in medium containing 0.1–0.8 M sucrose, with the lowest levels observed at 0.2 M sucrose. In contrast, the levels of stress granule markers G3BP1 and YB1 remained unchanged in T24 and U2OS cells (Fig. 3F, G, Supplementary Information). These results strongly suggest that p62 bodies, but not stress granules, are degraded by autophagy under hyperosmotic stress.

A, B Immunostaining of T24 cells exposed to hyperosmotic stress. Cells were cultured under normosmotic conditions without medium exchange (No Me) or shifted to DMEM containing indicated concentrations of sucrose for 20 min. After fixation, the cells were stained with antibodies against LC3 (green) and p62 (H00008878-MO1, red, A), or G3BP1(red, B). Boxed regions are enlarged and shown as insets. Scale bars = 20 µm and 2 µm (insets). C Quantification of the number of LC3-positive puncta (green) in T24 and U2OS cells. The number of cytoplasmic LC3-positive puncta per area was quantified in 30 cells from three independent experiments. Data are presented as mean ± SD. Each value was statistically compared to “0 M” group using one-way ANOVA followed by Tukey’s test. * P < 0.05, *** P < 0.001. D Quantification of the colocalization of p62 with LC3 in T24 and U2OS cells using Manders’ coefficients. The fraction of p62-positive area overlapping with LC3 was measured in 30 cells from three independent experiments. Data are presented as mean ± SD. Each value was statistically compared to “0.1 M” group using one-way ANOVA followed by Tukey’s test. *** P < 0.001, NS: not significant. E Quantification of the colocalization of LC3 with p62 or G3BP1 in T24 and U2OS cells using Manders’ coefficients. The fraction of LC3-positive area overlapping with p62 or G3BP1 was measured in 30 cells from three independent experiments. T24 and U2OS cells were exposed to DMEM containing 0.4 M sucrose for 20 min. Data are presented as mean ± SD. Statistical comparisons were conducted using Welch’s t-test. *** P < 0.001. Image data for U2OS cells in panels C, D and E are shown in Supplementary Fig. 3A, B. F WB analysis of T24 and U2OS cells exposed to hyperosmotic stress using indicated antibodies. Cells were cultured under normosmotic conditions without medium exchange (No Me) or shifted to DMEM containing indicated concentrations of sucrose for 1 h followed by cell lysis. G Quantification of WB data in panel F. Band intensities were normalized to GAPDH and expressed relative to “0 M” condition. Data are presented as mean ± SD from three independent experiments. Each value was statistically compared to “0 M” condition using one-way ANOVA followed by Tukey’s test. * P < 0.05, ** P < 0.01, *** P < 0.001.
p62 bodies and stress granules exhibit distinct morphological characteristics
Because 70–80% of p62 bodies overlapped with LC3 (Supplementary Fig. 3D), and no prior study has directly compared their fine structures with stress granules, we performed correlative light and electron microscopy (CLEM) analysis. First, we analyzed U2OS cells stably expressing mCherry-tagged p62 and GFP-tagged G3BP1. The p62 bodies contained condensed materials comprising fibrillar and particulate structures, and were often fully or partially surrounded by isolation membranes (Fig. 4A and Supplementary Fig. 4A). In contrast, stress granules exhibited irregular shapes and comprised fibrillar and particulate structures that appeared coarser than those in p62 bodies. Similar findings were observed with specific antibody-labeled endogenous p62 and G3BP1 proteins in the human retinal pigment epithelial cell line ARPE-19 (Fig. 4B and Supplementary Fig. 4B), in which the formation of p62 bodies and their colocalization with LC3 were comparable to those in T24 and U2OS cells (Supplementary Fig. 4C). Although this procedure may have caused some leakage of cytoplasmic components, it excluded the effects of protein overexpression and fluorescent tagging. These observations suggest that p62 bodies and stress granules are morphologically distinct, and that p62 bodies are more likely to be enclosed by isolation membranes under hyperosmotic stress (Fig. 4B and Supplementary Fig. 4B).

A CLEM images of U2OS cells stably expressing GFP-G3BP1 and mCherry-p62, cultured in DMEM with 0.3 M sucrose for 20 min. CLEM images (fluorescence, EM, and merged images) are shown on the left, with the regions (ii) and (iii) enlarged below, as indicated. The region (sup) is shown in Supplementary Fig. 4A. The black-flamed EM image is enlarged on the right (iv); the boxed regions are further magnified below (v). Red arrows and green arrowheads indicate p62- and G3BP1-positive structures, respectively. B CLEM images of ARPE-19 cells cultured in DMEM with 0.3 M sucrose for 20 min and immunolabeled using anti-p62 (PM045) and anti-G3BP1 antibodies. The layout is the same as (A). The region (sup) is shown in Supplementary Fig. 4B. Red arrows and green arrowheads indicate p62- and G3BP1-positive structures, respectively. Scale bars = 20 µm (i), 5 µm (ii), 1 µm (iii and iv), 200 nm (v).
Hyperosmotic stress-induced p62 bodies primarily contain NBR1, TAX1BP1, and K63-linked ubiquitin chains
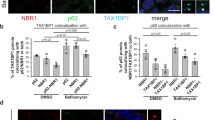
NBR1, TAX1BP1, NDP52, and optineurin (OPTN) contain UBA and LIR domains, functioning as p62-like autophagy receptors7. WB analysis of hypertonicity-induced receptor degradation showed that NBR1 and TAX1BP1, but not NDP52 or OPTN, were reduced under hyperosmotic stress, similar to p62, in both T24 and U2OS cells (Fig. 5A, B, Supplementary Information). Furthermore, upon 0.2 M sucrose treatment for 1 h, the degradation of p62, NBR1, and TAX1BP1 was blocked by treatment with a lysosomal inhibitor, Bafilomycin A1 (BafA1), but not by a proteasome inhibitor, epoxomicin (Fig. 5C, D, Supplementary Information). Similar to p62 bodies, the number of NBR1 and TAX1BP1 puncta increased after 20 min of osmotic stress induced by 0.2 M sucrose, and these puncta overlapped with p62 bodies (Supplementary Fig. 5A). Although prolonged stress (1 h) reduced the number of p62 bodies, BafA1-treatment increased the fraction of p62 bodies in WIPI-2-positive puncta (Supplementary Fig. 5B, C), indicating the incorporation of p62 into autophagosomes. Under the same hyperosmotic conditions, BafA1 treatment also significantly enhanced the colocalization of NBR1 or TAXBP1 with p62 (Fig. 5E, F). When GFP-tagged receptor proteins were compared, a significant fraction of NBR1 and TAX1BP1, but only a small fraction of NDP52 and OPTN, were recruited to p62 bodies (Supplementary Fig. 5D). These results indicate that NBR1 and TAX1BP1 are components of p62 bodies and are specifically degraded by autophagy under hyperosmotic stress.

A WB analysis of T24 and U2OS cells exposed to hyperosmotic stress using the indicated antibodies. Cells were cultured under normosmotic conditions without medium exchange (No Me), or shifted to DMEM containing indicated concentrations of sucrose for 1 h followed by cell lysis. B Quantification of WB data in panel A. Band intensities were normalized to GAPDH, and expressed relative to “0 M” condition. Data are presented as mean ± SD from three independent experiments. Each value was statistically compared to “0 M” condition using one-way ANOVA followed by Tukey’s test. * P < 0.05, ** P < 0.01, *** P < 0.001. C WB analysis of T24 and U2OS cells exposed to hyperosmotic stress with lysosomal or proteasomal inhibitor. Cells were cultured under normosmotic conditions without medium exchange (No Me), or shifted to DMEM containing 0.2 M sucrose and DMSO (vehicle), BafA1 (100 nM), or Epoxomicin (3.6 µM) for 1 h, before cell lysis. Indicated antibodies were used. D Quantification of WB data in panel C. Band intensities were normalized to GAPDH, and expressed relative to “No Me” condition. Data are presented as mean ± SD from three independent experiments. Each value was statistically compared to “No Me” condition using one-way ANOVA followed by Tukey’s test. * P < 0.05, ** P < 0.01, *** P < 0.001, NS not significant. E Immunostaining of T24 cells exposed to hyperosmotic stress with lysosomal inhibitor. Cells were cultured in DMEM containing 0.2 M sucrose and DMSO (vehicle) or BafA1 (100 nM) for 1 h. They were fixed for immunostaining with antibodies against p62 (H00008878-MO1) and NBR1 or TAX1BP1. Boxed regions are enlarged and shown as insets. Scale bars = 20 µm and 2 µm (insets). F Quantification of colocalization in panel E using Manders’ coefficients. The fraction of p62-positive area overlapping with NBR1, or TAX1BP1 was quantified in 20 cells from a representative experiment. Data are presented as mean ± SD. Statistical comparisons were carried out using Welch’s t-test.
Because p62 bodies contain polyubiquitin chains9,10,11 and are associated with proteasome foci, which form in the nucleus under hyperosmotic stress and mainly contain K48-linked chains19,23,24,25, we sought to investigate the types of ubiquitin chains in hyperosmotic stress-induced p62 bodies. Double IF revealed the appearance of ubiquitin-positive puncta in both the cytoplasm and nucleus of T24 cells treated with 0.2 M sucrose. Notably, p62 colocalized with ubiquitin-positive puncta only in the cytoplasm (Fig. 6A). As expected, stress granules marked by YB1, which was confirmed to colocalizes with G3BP1 but not with WIPI-2 (Supplementary Fig. 6A, B), did not contain ubiquitin (Fig. 6B). Triple immunofluorescence microscopy showed that, while proteasome foci labeled with antibodies against the proteasome subunit PSMD4 primarily contained K48-linked ubiquitin in the nucleus, p62 bodies in the cytosol were predominantly associated with K63-linked ubiquitin (Fig. 6C). Quantitative analysis revealed that PSMD4 exhibited a higher degree of colocalization with K48-linked ubiquitin puncta than p62, whereas p62 showed greater colocalization with K63-linked ubiquitin puncta than PSMD4 (Fig. 6D). Even after treatment with BafA1, the colocalization of p62 puncta with K63-linked ubiquitin, but not with K48-linked ubiquitin, was significantly increased (Fig. 6 E, F). These results suggest that hyperosmotic stress-induced autophagy contributes to the degradation of p62/NBR1/TAX1BP1 as well as K63-polyubiquitinated proteins in the cytosol.

Immunostaining of T24 cells exposed to hyperosmotic stress, using antibodies against p62 (PM045) and ubiquitin (A) or YB1 and ubiquitin (B). Cells were cultured under normosmotic conditions without medium exchange (No Me), or shifted to DMEM with 0.2 M (A) or 0.4 M (B) for 20 min. C Immunostaining of T24 cells exposed to hyperosmotic stress, using antibodies against p62 (GP62-C), PSMD4, and K48-linked or K63-linked polyubiquitin chains. Boxed regions are enlarged and shown as insets. D Quantification of colocalization in panel C using Manders’ coefficients. The fraction of PSMD4- or p62-positive area overlapping with K48-linked (top) or K63-linked (bottom) polyubiquitin was quantified in 20 cells from a representative experiment. Data are presented as mean ± SD. Statistical comparisons were conducted using Welch’s t-test. E Immunostaining of T24 cells exposed to hyperosmotic stress with lysosomal inhibitor. Cells were cultured in DMEM containing 0.2 M sucrose and DMSO (vehicle) or BafA1 (100 nM) for 1 h, before cell fixation. Antibodies against p62 and K48-linked or K63-linked polyubiquitin chains are used. Boxed regions are enlarged and shown as insets. White dashed lines show nuclear membrane. Scale bars = 20 µm and 2 µm (insets). F Quantification of colocalization in E using Manders’ coefficients. The fraction of p62-positive area overlapping with K48-linked (left) or K63-linked (right) polyubiquitin was quantified in 20 cells from a representative experiment. Data are presented as mean ± SD. Statistical comparisons were carried out using Welch’s t-test. NS not significant.
Hyperosmotic stress-induced p62 body formation in an epithelial organoid mimicking human corneal tissue
To determine whether similar p62 bodies can be induced in multicellular tissues, we utilized an epithelial organoid that mimics human corneal epithelium (Supplementary Fig. 7). Exposure to hypertonic medium containing 0.2 M sucrose induced the formation of p62 bodies across all layers of the stratified epithelium, many of which were positive for WIPI-2 (Fig. 7A, C). YB1-positive stress granules also formed under 0.4 M sucrose, but were distinct from p62 bodies (Fig. 7B, D). Triple immunofluorescence microscopy of p62, WIPI-2, and YB1 in the organoid exposed to 0.4 M sucrose revealed that WIPI-2 localized to p62 bodies, but not YB1-positive stress granules (Fig. 7E). Electron microscopy further showed the presence of typical autophagosome-like structures in the cytoplasm following hypertonic stress (Fig. 7F). These findings suggest that p62 bodies are formed for autophagic degradation in cultured stratified epithelium under hyperosmotic stress.

A, B Double immunostaining of sections of epithelial organoid exposed to hyperosmotic stress. The organoids were cultured in DMEM containing sucrose as indicated for 20 min. After fixation, sections were prepared and stained with antibodies against p62 (GP62-C, red) and WIPI-2 (green, A) or YB1 (green, B). Boxed regions are enlarged and shown on the right. Scale bars = 20 µm and 5 µm (insets). Quantification of the data in A (C) and B (D). The number of p62 bodies and WIPI-2- or YB1-positive puncta (green) was measured. Data are presented as mean ± SD from four culture wells. Statistical comparisons were carried out using Welch’s t-test. E Triple immunostaining of sections of corneal organoid exposed to hyperosmotic stress, using antibodies against p62 (GP62-C), WIPI-2, and YB1. The organoids were incubated in DMEM with 0 M or 0.4 M sucrose for 20 min. Boxed regions are enlarged and shown on the right. White arrows indicate colocalization of WIPI-2 with p62, but not with YB1. Scale bars = 20 µm and 5 µm (insets). F EM analysis of epithelial organoids exposed to hyperosmotic stress. The organoids were cultured in DMEM with 0 M (i–iv) or 0.4 M sucrose (v-viii) for 20 min. Boxed region in (i and ii) or (v and vi) is enlarged and shown as indicated. Scale bars = 10 µm (i and v), 2 µm (ii and vi), 500 nm (iii, iv, vii, and viii).
Discussion
Degradation of p62 via autophagy following hyperosmotic stress has been shown in several cell lines12,13,14,26. However, it remains unclear whether this process is mediated by the formation of p62 bodies. In the present study, we demonstrated that hyperosmotic stress induces the formation of p62 bodies that are frequently associated with the isolation membrane, facilitating their autophagic degradation. The p62 body formation occurs very rapidly within 1 min throughout the cytoplasm, followed by gradual fusion events and/or the recruitment of surrounding p62 molecules, leading to their sequestration by autophagosomes. Their formation requires multivalent interactions among p62 molecules or with ubiquitin chains, the properties of which are consistent with the characteristics of liquid–liquid phase separation. These p62 bodies predominantly contain K63-linked polyubiquitin chains and two other autophagy receptors, NBR1 and TAX1BP1, in line with the known features of canonical p62 bodies10,11,27,28. Consistent with evidence that p62 bodies serve as platforms for autophagosome formation and degradation under oxidative stress11, we observed similar behavior under hyperosmotic stress. A key distinction is that, while hyperosmotic stress was found to induce the rapid formation of p62 bodies (within 1 min) followed by autophagy, oxidative stress-induced p62 body formation occurs more gradually (over 1 h), with autophagy typically assessed post-stress removal11,29. It should be noted that LC3-positive p62 puncta are also detectable during starvation-induced autophagy by light microscopy30,31. However, our previous study using structured illumination microscopy and immuno-electron microscopy revealed that p62 localizes to specific subdomains of the autophagosome membranes, occasionally forming aggregate-like structures31. These findings support a bulk degradation mechanism rather than a canonical selective autophagic degradation of p62 bodies, as previously observed11 and in the present study. Nevertheless, whether p62 body formation also plays a functional role in the starvation-induced autophagy remains to be elucidated. Taken together, our findings demonstrate for the first time that the formation of p62 bodies mediates the rapid autophagic clearance of cytoplasmic proteins under hyperosmotic stress.
To further characterize p62 bodies, we compared them with stress granules, which are well-studied cytosolic condensates formed under hyperosmotic stress20. We found that p62 bodies, but not stress granules, were degraded by hyperosmotic stress-induced autophagy. This observation aligns well with the proposed functions of the two condensates: stress granules transiently sequester untranslated mRNAs to protect them from degradation under stress conditions32,33, whereas p62 bodies facilitate the degradation of polyubiquitinated proteins, which must be removed to ensure cell survival5. This distinction is further supported by our electron microscopy analysis, which revealed that p62 bodies exhibit a more compact morphology compared with stress granules, and are frequently surrounded by isolation membrane (Fig. 4A and Supplementary Fig. 4A). In contrast to the liquid-like behavior typically attributed to stress granules, the observed compaction of p62 bodies suggests that they possess gel-like properties, consistent with a previous report on oxidative stress-induced p62 bodies11. Furthermore, this morphological compaction may enhance the wetting effect that facilitates autophagosome formation on the surface of p62 bodies34, a process that is likely impeded by the irregular surface of stress granules.
Compared with G3BP1-labeled stress granules, p62 puncta formed more rapidly and under lower levels of hyperosmotic stress, leading us to conclude that p62 bodies are more sensitive to this type of stress. However, it should be noted that IGF2BP, an RNA-binding protein and a component of stress granules, rapidly (<1 min) forms clusters in response to hyperosmotic stress, preceding the recruitment of G3BP1 and T cell-restricted intracellular antigen-1 (TIA1)35. This suggests that stress granule formation is also sensitive to hyperosmotic stress, but requires more time to mature to a level detectable by conventional fluorescence microscopy.
Apart from classical stress granule components, various other molecules have recently been shown to undergo phase separation in response to hyperosmotic stress, including DCP1A36, RAD23B19, ASK337, WNK138, and BAG239. Among them, RAD23B forms nuclear condensates with ubiquitinated proteins, proteasomes, and p97, functioning in nuclear protein degradation. In contrast, cytoplasmic BAG2 condensates do not associate with p62, mediating ubiquitin-independent proteasomal degradation. Here, we have identified p62 bodies as an additional type of cytoplasmic degradation platform that is activated under hyperosmotic stress. Given that p62 bodies preferentially sequester K63-linked ubiquitinated proteins, as suggested by previous studies10,27, cells may utilize this mechanism to clear K63-linked polyubiquitinated proteins, thereby maintaining homeostasis under osmotic stress. Furthermore, this mechanism may contribute to the rapid osmotic compensation achieved by liberating water molecules from proteins, as recently proposed by Watson et al.40. It is still unknown which specific proteins are preferentially ubiquitinated and incorporated into p62 bodies under hyperosmotic stress. Addressing this will require further investigation.
What is the physiological significance of these findings in higher organisms? We demonstrated that treatment with 0.1–0.2 M sucrose (400–500 mOsmol/L) promoted efficient autophagic degradation of p62 in T24 and U2OS cells (Fig. 3D). In addition, 0.2 M sucrose (500 mOsmol/L) induced the formation of both p62 bodies and autophagosomes in a human epithelial organoid mimicking corneal epithelium. In higher organisms, the osmolarity of interstitial fluid in the inner renal medullar ranges from 600 to 1700 mOsm/kg1. Additionally, the oral cavity and upper digestive tracts are frequently exposed to foods or beverages, with osmolarities ranging from approximately 300 to over 2000 mOsm/kg41,42, indicating that such levels of hypertonicity employed in the present study can exist under physiological conditions. However, it is also known that cells in higher organisms are equipped with various mechanisms to respond to hyperosmotic stress, which differ among cell types1,2. Therefore, p62 body formation may not necessarily occur in all tissues under such conditions, and this should be further clarified in future studies. The osmolarity of human serum is normally maintained within a narrow range of 285 – 295 mOsm/kg. In patient with hyperglycemic hyperosmolar state, a serious diabetic emergency, the serum osmolarity can exceed 320 mOsmol/kg43. In addition, in patients exhibiting the most severe pathological grade of dry eye, tear osmolarity increased to 344 mOsm/L on average2. However, these high tonicities are still lower than the hyperosmotic conditions used in this study or several previous studies that employed 400 mOsm/kg or more1. Therefore, the development of more sensitive in vivo models would be valuable for elucidating the pathological relevance of p62 body-mediated autophagy.
Methods
Antibodies
A list of all primary and secondary antibodies, dilutions used for each technique, and supplier information are provided in Supplementary Table 1.
Plasmid construction
Plasmids and primers used in this study are listed in Supplementary Tables 2 and 3, respectively. Using human cDNA as template, G3BP1 was amplified with forward (G3BP1 Fw) and reverse (G3BP1 Rv) primers for insertion into the mammalian expression vector GFP-pcDNA at the BamHI and XhoI sites, resulting in GFP-G3BP1-pcDNA. The GFP-G3BP1 fusion gene was amplified with primers GFP Fw and pLVSIN Rv and cloned into pLVSIN-puro (Clontech) at the EcoRI and BamHI sites, resulting in GFP-G3BP1-pLVSIN-puro. An mCherry-p62 fusion gene from mCherry-p62-C2 (provided by Masaaki Komatsu in Juntendo University) was amplified with primers mCherry Fw and p62 Rv and inserted into the lentiviral expression vector pLVSIN-hyg (Clontech) at the XhoI and BamHI sites, resulting in mCherry-p62-pLVSIN-hyg. The GFP-tagged NBR1, TAX1BP1, NDP52, and OPTN plasmids were provided by Michael Lazarou (Walter and Eliza Hall Institute of Medical Research)44. The GFP-p62WT, GFP-p62K7A/D69A, and GFP-p62F406A plasmids were kindly provided by Masaaki Komatsu (Juntendo University)45.
Cell culture and transfection, and generation of a stable cell line
T24 cells (JCRB0711) were purchased from the JCRB Cell Bank (National Institutes of Biomedical Innovation, Japan), and ARPE-19 cells from ATCC. U2OS cells were kindly provided by Yuki Osaki (Sapporo Medical University). The Huh-1 cell line lacking p62 was provided by Masaaki Komatsu (Juntendo University)46. The cells were cultured in high-glucose DMEM (08458-16, Nacalai) supplemented with 9.09% FBS (173012, Lot: BCCC3916, Sigma-Aldrich). They were regularly tested for mycoplasma contamination. To induce hyperosmotic stress, cells were shifted to DMEM supplemented with the indicated concentrations of sucrose, mannitol, or salts. For the experiments shown in Fig. 2G and Supplementary Fig. 5C, Huh-1 or U2OS cells were transfected with plasmids for 48 h using FuGENE HD Transfection Reagent (Promega). To generate U2OS cells stably expressing both mCherry-p62-pLVSIN-hyg and GFP-G3BP1-pLVSIN-puro, 10 μg/mL puromycin (InvivoGen) and 200 µg/mL hygromycin (InvivoGen) were used for selection.
Immunofluorescence microscopy
Cells grown on coverslips (Matsunami Glass Ind., C012001) were fixed with 4% paraformaldehyde (16005, Sigma-Aldrich) in phosphate-buffered saline (PBS) for 20 min and permeabilized with 0.1% Triton X-100 in PBS for 20 min. Cells were blocked in PBS containing 0.1% Tween 20 (PBST) with 0.4% bovine serum albumin (BSA) for 30 min, and then incubated with primary antibodies in PBST for 1–3 h. The coverslips were incubated with secondary antibodies in PBST for 1 h. Images were acquired with an FV1000 confocal microscope (Olympus) equipped with a PlanApo N 60× lens (NA 1.42 oil).
Quantification of puncta
Two images containing approximately 20 cells were acquired for each experimental condition. Using Fiji ImageJ (1.54 h47), puncta were detected after background subtraction with a rolling ball radius of 3.0, followed by adjustment of the threshold value, which was set individually for each experiment. After applying the watershed function, puncta of appropriate sizes and circularities were counted, and the results were normalized to cell area and expressed as the number of puncta per 100 μm2. The colocalization rate was quantified using Manders’ coefficients in Fiji ImageJ software. Puncta were detected after background subtraction with a rolling ball radius of 3.0, followed by adjustment of the threshold value, which was set individually for each experiment. LC3-positive puncta were further processed using “Despeckle” function to remove background noise. Manders’ coefficients were calculated using the “coloc2” plugin. In some experiments, colocalization was also assessed manually by counting puncta and calculating the ratio of double-positive puncta to either of single-positive puncta using Photoshop (Adobe). Diameter of p62 puncta was measured in 50 or more puncta using the ruler tool in Photoshop.
Live-cell imaging
U2OS cells stably expressing GFP-G3BP1 and mCherry-p62 were seeded on a glass-bottom dish (3971-035, Iwaki). Images were acquired by spinning disk SD-OSR confocal microscope (Olympus) equipped with a UPlanSApo 100× lens (NA 1.35 oil). To apply hyperosmotic stress, DMEM containing 0.5 M sucrose was applied and exchanged twice at half volume each, resulting in a final concentration of about 0.38 M sucrose. Time-lapse images were then captured every 30 s for 40 min. The movie file was generated using Basic MetaMorph software (Molecular Devices). Fluorescence intensity of the p62 body-free cytoplasmic area was quantified using Fiji ImageJ (1.54 h47).
Western blot analysis
T24 and U2OS cells were washed once in cold PBS prior to lysis in PBS containing 1% sodium dodecyl sulfate with boiling for 7 min. After sonication with a UR-20P (Tomy), 10 µg of each total cell lysate was loaded onto 5–20% gradient gels (194–15021, Wako) for electrophoresis, followed by transfer to polyvinylidene fluoride membranes (IPVH00010; Merck Millipore). The blots were blocked with 5% skim milk in PBST and probed with the indicated antibodies. The band intensity was analyzed by ImageQuant TL (GE Healthcare). Band intensities were normalized to GAPDH. At least three independent experiments were performed.
CLEM analysis
U2OS cells stably expressing GFP-G3BP1 and mCherry-p62 were grown on coverslips with grid lines (Matsunami Glass Ind., GC1310), then shifted to hyperosmotic media and fixed with 4% paraformaldehyde in PBS for 20 min. Fluorescence images were acquired at specific position numbers using a confocal microscope FV1000, as described above. The cells were fixed again with 0.1 M phosphate buffer (pH 7.4) with 2% paraformaldehyde and 2% glutaraldehyde (3042, Nissin EM) overnight, followed by treatment with 1% osmium tetroxide (K95709505, Merck Millipore) and 1.5% potassium ferrocyanide (152560, Wako) in 0.1 M phosphate buffer (pH 7.4) for 1 h. Cells were then dehydrated in ethanol, incubated in propylene oxide, and embedded in epoxy resin (Nissin EM). Areas containing cells of interest were trimmed from the resin based on the fluorescence images, and serial ultrathin (60-nm thickness) sections were prepared and observed with a JEM1400 electron microscope (JEOL). Fluorescence and EM images were positionally aligned based on multiple p62 bodies and stress granules, using Photoshop CS6 software (Adobe, RRID:SCR_014199). For CLEM analysis with antibody labeling, ARPE-19 cells grown on coverslips with grid lines were exposed to hyperosmotic stress and then fixed with 4% paraformaldehyde in PBS for 20 min. The cells were permeabilized in liquid nitrogen for 30 s, blocked in PBS containing 0.4% BSA for 30 min, and then incubated with primary antibodies in PBS for 1 h followed by secondary antibodies in PBS for 1 h. The procedures for fluorescence and EM image acquisition are the same as above.
Analyses of epithelial organoid
A three-dimensional culture system of human corneal epithelium was purchased from J-TEC (LabCyte CORNEA model). After exposing the apical surface of the epithelium to hypertonic or isotonic medium for 20 min, the samples were fixed in 4% paraformaldehyde in PBS overnight at 4 °C, embedded in Tissue-Tek O.C.T. compound (4583, Sakura Finetek Japan), and sliced with a cryo-microtome (CM3050S, Leica) at 50-μm thickness. Sections were permeabilized in PBS containing 0.1% Triton X-100 for 20 min, blocked in PBST containing 0.4% BSA for 30 min, and then incubated with primary antibodies in PBST for 1 h followed by secondary antibodies in PBST for 30 min. Fluorescence images were obtained using an FV1000 fluorescence microscope. For each of four culture wells, puncta were counted in a 200 × 20 μm region of interest using Fiji ImageJ as described above.
For electron microscopy analysis, the samples were fixed in 0.1 M phosphate buffer (pH 7.4) with 2% paraformaldehyde and 2% glutaraldehyde overnight, followed by treatment with 1% osmium tetroxide and 1.5% potassium ferrocyanide in 0.1 M phosphate buffer (pH 7.4) for 1 h. The samples were dehydrated, embedded in epoxy resin, and observed as described above.
Statistics and reproducibility
Statistical significance was assessed using one-way analysis of variance (ANOVA) with Tukey’s test with SPSS software (IBM), or Welch’s t-test using Microsoft Office 16 Excel, as specified in the figure legends. The number of analyzed cells, culture wells, and independent experiments is also indicated in the figure legends 1.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data supporting the results of this study in the article and in the Supplementary Information are available from the corresponding authors on reasonable request. Newly generated plasmids in this study (Supplementary Table 2) are also available from the corresponding authors upon reasonable request. Uncropped blot images are included in Supplementary Information. The dataset used for quantification analyses is in Supplementary Data 1.
Change history
13 April 2026
Since the version of the article initially published, the second sentence in the Data availability section has been amended to “Newly generated plasmids in this study (Supplementary Table 2) are also available from the corresponding authors upon reasonable request” in the HTML and PDF versions of the article.
References
Burg, M. B., Ferraris, J. D. & Dmitrieva, N. I. Cellular response to hyperosmotic stresses. Physiol. Rev. 87, 1441–1474 (2007).
Brocker, C., Thompson, D. C. & Vasiliou, V. The role of hyperosmotic stress in inflammation and disease. Biomol. Concepts 3, 345–364 (2012).
Kitamura, A. et al. Increased intracellular crowding during hyperosmotic stress. Sci. Rep. 13, 11834 (2023).
Choe, K. P. & Strange, K. Genome-wide RNAi screen and in vivo protein aggregation reporters identify degradation of damaged proteins as an essential hypertonic stress response. Am. J. Physiol. Cell Physiol. 295, C1488–C1498 (2008).
Burkewitz, K., Choe, K. & Strange, K. Hypertonic stress induces rapid and widespread protein damage in C. elegans. Am. J. Physiol. Cell Physiol. 301, C566–C576 (2011).
Dikic, I. Proteasomal and autophagic degradation systems. Annu. Rev. Biochem. 86, 193–224 (2017).
Lamark, T. & Johansen, T. Mechanisms of selective autophagy. Annu. Rev. Cell Dev. Biol. 37, 143–169 (2021).
Yamamoto, H., Zhang, S. & Mizushima, N. Autophagy genes in biology and disease. Nat. Rev. Genet. 24, 382–400 (2023).
Sun, D., Wu, R., Zheng, J., Li, P. & Yu, L. Polyubiquitin chain-induced p62 phase separation drives autophagic cargo segregation. Cell Res. 28, 405–415 (2018).
Zaffagnini, G. et al. p62 filaments capture and present ubiquitinated cargos for autophagy. EMBO J. 37, https://doi.org/10.15252/embj.201798308 (2018).
Kageyama, S. et al. p62/SQSTM1-droplet serves as a platform for autophagosome formation and anti-oxidative stress response. Nat. Commun. 12, 16 (2021).
Nunes, P. et al. Hypertonic stress promotes autophagy and microtubule-dependent autophagosomal clusters. Autophagy 9, 550–567 (2013).
Tamura, N., Kageyama, S., Komatsu, M. & Waguri, S. Hyperosmotic stress induces unconventional autophagy independent of the Ulk1 Complex. Mol. Cell. Biol. 39, https://doi.org/10.1128/mcb.00024-19 (2019).
Zhu, H. et al. Hyperosmotic stress stimulates autophagy via the NFAT5/mTOR pathway in cardiomyocytes. Int. J. Mol. Med. 42, 3459–3466 (2018).
Gomes, E. & Shorter, J. The molecular language of membraneless organelles. J. Biol. Chem. 294, 7115–7127 (2019).
van Leeuwen, W. & Rabouille, C. Cellular stress leads to the formation of membraneless stress assemblies in eukaryotic cells. Traffic 20, 623–638 (2019).
Boeynaems, S. et al. Protein phase separation: a new phase in cell biology. Trends Cell Biol. 28, 420–435 (2018).
Protter, D. S. W. & Parker, R. Principles and properties of stress granules. Trends Cell Biol. 26, 668–679 (2016).
Yasuda, S. et al. Stress- and ubiquitylation-dependent phase separation of the proteasome. Nature 578, 296–300 (2020).
Bounedjah, O. et al. Macromolecular crowding regulates assembly of mRNA stress granules after osmotic stress: new role for compatible osmolytes. J. Biol. Chem. 287, 2446–2458 (2012).
Komatsu, M. p62 bodies: phase separation, NRF2 activation, and selective autophagic degradation. IUBMB Life 74, 1200–1208 (2022).
Peña-Oyarzun, D. et al. Hyperosmotic stress stimulates autophagy via polycystin-2. Oncotarget 8, 55984–55997 (2017).
Iriki, T. et al. Senescent cells form nuclear foci that contain the 26S proteasome. Cell Rep. 42, 112880 (2023).
Lee, J., Le, L., Kim, E. & Lee, M. J. Formation of non-nucleoplasmic proteasome foci during the late stage of hyperosmotic stress. Cells 10, https://doi.org/10.3390/cells10092493 (2021).
Uriarte, M. et al. Starvation-induced proteasome assemblies in the nucleus link amino acid supply to apoptosis. Nat. Commun. 12, 6984 (2021).
Dafre, A. L., Schmitz, A. E. & Maher, P. Hyperosmotic stress initiates AMPK-independent autophagy and AMPK- and autophagy-independent depletion of thioredoxin 1 and glyoxalase 2 in HT22 nerve cells. Oxid. Med. Cell Longev. 2019, 2715810 (2019).
Seibenhener, M. L. et al. Sequestosome 1/p62 is a polyubiquitin chain binding protein involved in ubiquitin proteasome degradation. Mol. Cell. Biol. 24, 8055–8068 (2004).
Turco, E. et al. Reconstitution defines the roles of p62, NBR1 and TAX1BP1 in ubiquitin condensate formation and autophagy initiation. Nat. Commun. 12, 5212 (2021).
Ichimura, Y. et al. Phosphorylation of p62 activates the Keap1-Nrf2 pathway during selective autophagy. Mol. Cell 51, 618–631 (2013).
Bjorkoy, G. et al. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J. Cell Biol. 171, 603–614 (2005).
Eino, A. et al. Sqstm1-GFP knock-in mice reveal dynamic actions of Sqstm1 during autophagy and under stress conditions in living cells. J. Cell Sci. 128, 4453–4461 (2015).
Youn, J. Y. et al. Properties of stress granule and P-body proteomes. Mol. Cell 76, 286–294 (2019).
Hofmann, S., Kedersha, N., Anderson, P. & Ivanov, P. Molecular mechanisms of stress granule assembly and disassembly. Biochim. Biophys. Acta Mol. Cell Res. 1868, 118876 (2021).
Agudo-Canalejo, J. et al. Wetting regulates autophagy of phase-separated compartments and the cytosol. Nature 591, 142–146 (2021).
Zeng, W. J. et al. Initiation of stress granule assembly by rapid clustering of IGF2BP proteins upon osmotic shock. Biochim. Biophys. Acta Mol. Cell Res. 1867, 118795 (2020).
Jalihal, A. P. et al. Multivalent proteins rapidly and reversibly phase-separate upon osmotic cell volume change. Mol. Cell 79, 978–990.e975 (2020).
Watanabe, K. et al. Cells recognize osmotic stress through liquid-liquid phase separation lubricated with poly(ADP-ribose). Nat. Commun. 12, 1353 (2021).
Boyd-Shiwarski, C. R. et al. WNK kinases sense molecular crowding and rescue cell volume via phase separation. Cell 185, 4488–4506.e4420 (2022).
Carrettiero, D. C. et al. Stress routes clients to the proteasome via a BAG2 ubiquitin-independent degradation condensate. Nat. Commun. 13, 3074 (2022).
Watson, J. L. et al. Macromolecular condensation buffers intracellular water potential. Nature 623, 842–852 (2023).
Fordtran, J. S. & Locklear, T. W. Ionic constituents and osmolality of gastric and small-intestinal fluids after eating. Am. J. Dig. Dis. 11, 503–521 (1966).
Wendland, B. E. & Arbus, G. S. Oral fluid therapy: sodium and potassium content and osmolality of some commercial “clear” soups, juices and beverages. Can. Med. Assoc. J. 121, 564–566,568,571 (1979).
Umpierrez, G. E. et al. Hyperglycemic crises in adults with diabetes: a consensus report. Diab. Care 47, 1257–1275 (2024).
Lazarou, M. et al. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 524, 309–314 (2015).
Faruk, M. O. et al. Phase-separated protein droplets of amyotrophic lateral sclerosis-associated p62/SQSTM1 mutants show reduced inner fluidity. J. Biol. Chem. 297, 101405 (2021).
Inami, Y. et al. Persistent activation of Nrf2 through p62 in hepatocellular carcinoma cells. J. Cell Biol. 193, 275–284 (2011).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Acknowledgements
We thank Masaaki Komatsu (Juntendo University), Michael Lazarou (Walter and Eliza Hall Institute of Medical Research), and Yuki Ohsaki (Sapporo Medical University) for providing research materials as described in the Materials and Methods section. We also thank Katsuyuki Kanno for technical support in electron microscopy, Masaaki Komatsu (Juntendo University) for critical reading our manuscript prior to submission, Michelle Kahmeyer-Gabbe, PhD, from Edanz (https://jp.edanz.com/ac) for editing a draft of this manuscript, and all members of our department for helpful discussions. This work was supported by a Grant-in-Aid for Scientific Research (B) (23H02660, 20H03415 to S.W. and N.T.) and a Grant-in-Aid for Scientific Research (C) (22K06794, 19K16478 to N.T.).
Author information
Authors and Affiliations
Contributions
N.T. designed and conceptualized the project, and performed all experiments. N.T and S.W. analyzed the data and wrote the manuscript. S.W. further conceptualized and supervised the project, and reviewed and edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editors: Giulia Bertolin and Kaliya Georgieva. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tamura, N., Waguri, S. Rapid activation of p62 body-mediated autophagy in human cells under hyperosmotic stress. Commun Biol 9, 1 (2026). https://doi.org/10.1038/s42003-025-09190-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-025-09190-6
