
Abstract
Molecular interactions between therapeutic antibodies and neonatal Fc receptor (FcRn) are major contributors to the long serum half-life of antibodies. Therefore, the in vitro affinity of IgG for FcRn is an important indicator of in vivo pharmacokinetic properties. However, the molecular basis of IgG-FcRn interactions is not fully understood from experimental and structural perspectives. Affinity evaluation using surface plasmon resonance is difficult. The molecular mechanism by which domains other than Fc or surface charges affect FcRn binding remains unclear. In this study, we developed an engineered FcRn-immobilized column for affinity chromatography. We analyzed various types of antibodies to link physicochemical characteristics and FcRn affinity. Furthermore, analysis using charge-engineered mutants indicated that the lateral surface of L chain also affects the FcRn affinity. Finally, we presented a structural model of the IgG-FcRn complex to discuss the molecular mechanism of how the charges at L chain affect the FcRn affinity.
Similar content being viewed by others


Introduction
Therapeutic antibody drugs have proven to be one of the most important therapeutic modalities for the treatment of a wide range of diseases. Over the past three decades, the number of therapeutic antibodies in clinical use has increased dramatically1. Moreover, evolving biotechnologies have enabled scientists worldwide to enhance the therapeutic efficacy of these drugs. Next-generation, engineered therapeutic antibody drugs, such as antibody drug conjugates (ADCs), bispecific antibodies, and Fc-engineered antibodies, have delivered remarkable breakthroughs for the treatment of intractable diseases2,3,4.
IgG1 is the predominant molecular format of therapeutic antibody drugs. IgG1 comprises two Fab domains and an Fc domain. The Fc domain, in particular, has been intensively engineered. This is because the Fc domain governs the multifaceted characteristics of therapeutic antibodies, such as Fc-mediated effector functions (antibody-dependent cell-mediated cytotoxicity (ADCC), antibody-dependent cell-mediated phagocytosis (ADCP), complement-dependent cytotoxicity (CDC)), and a long serum half-life in patients.
The long serum half-life of therapeutic antibodies in patients, that is a pharmacokinetic (PK) property, has a great impact on therapeutic efficacy. Estimating PK properties of therapeutic antibody in humans remains challenging, since multiple parameters—such as FcRn affinity of the antibody, administration dose, administration route, patient body weight, properties of target molecules, and underlying conditions—affect PK properties. Furthermore, multiple mechanisms are involved in the regulation of the PK properties of administered therapeutic antibodies, including phagocytosis by the reticuloendothelial system (phagocytic immune cells such as macrophages and monocytes), target (antigen)-mediated internalization, and target-independent pinocytosis5,6,7. Importantly, the neonatal Fc receptor (FcRn) plays a key role in the pinocytosis-mediated antibody clearance pathway because of its function of protection and recycling of IgG8,9. Human FcRn is located on diverse cell types, including epithelial cells, endothelial cells, and immune cells. FcRn comprises an MHC-class-I-like molecule and a β2-microglobulin (β2m). FcRn plays pivotal roles in IgG homeostasis, albumin homeostasis, maternal IgG transport, and antigen presentation. Notably, the interaction between IgG and FcRn is pH-dependent10. At acidic pH, IgG binds to FcRn with moderate affinity, whereas at neutral pH, the affinity is absent. In endosomes, only FcRn-captured IgG is protected from lysosomal degradation at acidic pH and can be recycled back into the blood circulation at neutral pH. To predict PK properties of therapeutic antibodies in humans, in vivo studies using human FcRn transgenic mice has been widely used. However, such studies could pose a few issues, including non-specific cellular uptake and anti-drug antibody (ADA) in some cases11,12,13,14. Therefore, it is crucially important to first establish a thorough molecular-level understanding of in vitro FcRn affinity as a critical element in predicting in vivo PK properties.
Notably, the molecular basis of the IgG-FcRn interaction is not fully understood from experimental and structural perspectives. From an experimental perspective, the affinity evaluation of IgG-FcRn interaction is known to be difficult for the following reasons: (1) the binding stoichiometry between IgG and FcRn is not 1:1, thus the surface plasmon resonance (SPR) results are inconsistent depending on the FcRn-immobilization level15; (2) the experimental methods used for affinity evaluation (such as SPR, enzyme-linked immunosorbent assay (ELISA), and bio-layer interferometry (BLI)) are based on the hypothesis that the analyte samples contain homogeneous molecular species. However, IgG characteristics are intrinsically heterogeneous. Therefore, these methods may yield inconsistent results if the samples are highly heterogeneous. (3) IgG-FcRn interactions are pH-dependent. Therefore, an affinity measurement at a single pH is not sufficient as an indicator of PK properties. From a structural perspective, the detailed structure of the complex of full-length IgG and FcRn has not yet been revealed. Based on the crystal structure, several studies have reported that the Fab arms are located close to the membrane surface when IgG binds to FcRn, and the effect of Fab on IgG-FcRn binding has been studied16,17,18,19,20,21,22,23,24,25. Brinkhaus et al. discussed that Fab impairs FcRn binding due to this structural hindrance26. Moreover, several studies have shown that the amino acid sequence or surface charges in the variable region also affect IgG-FcRn affinity11,12,27,28,29,30,31. However, the molecular mechanisms by which domains other than Fc, or surface charges, affect the FcRn-binding remain unclear25.
Schlothauer et al. have reported an analytical FcRn-immobilized affinity chromatography32. This analytical technology is highly useful and widely used for in vitro FcRn affinity evaluation. In the current study, we developed an engineered FcRn-immobilized column for affinity chromatography. The thermal stability of the immobilized FcRn was increased to resist the harsh environment of liquid chromatography. Using the FcRn column, we analyzed methionine-oxidized antibody and FcRn affinity-engineered mutants. Moreover, we analyzed 13 therapeutic monoclonal antibodies to elucidate the mechanism of how the amino acid sequence in the variable region affects the FcRn affinity. Based on the results, we further investigated the mechanism of positive charges in the L chain affect FcRn affinity by mutation analysis. Through these studies, we elucidated the structural insights into IgG-FcRn interactions. In addition, although the correlation between the elution profile of the FcRn column and in vivo data was not addressed, it was demonstrated that the method is useful for quality assessment of therapeutic antibodies.
Results
Development of the FcRn column
The proteinaceous ligand for an affinity column must be sufficiently stable to resist the harsh environment (e.g., high salt concentration, high pH, low pH, and high column pressure). To obtain a suitable ligand for the affinity chromatography column, FcRn was mutated by error-prone polymerase chain reaction (PCR). The obtained mutant contained seven mutations: C48R, N55D, R169L, N173D, Q209L, C251S, and K272E. The mutation sites in FcRn are shown in Fig. 1a. The seven mutation sites in FcRn were distant from the Fc-binding site. Thus, the binding interaction between IgG and FcRn is not altered upon these mutations. The thermal stability of the wild type and the mutant FcRn was evaluated using differential scanning calorimetry (DSC) (Fig. 1b). The melting temperature (Tm) of the wild type was 54.1 °C, whereas that of the mutant was 61.7 °C. The purified mutant FcRn protein was immobilized on the surface of the resin (Fig. 1c).

a The mutation sites of FcRn are shown. The six mutation sites in FcRn are colored in yellow. K272E is not displayed in the figure because the structure of the FcRn C-terminus was not determined in the crystal structure. b Thermal stability of FcRn wild type and mutant. DSC thermogram of wild type (black) and mutant (blue) FcRn are shown. c A schematic view of IgG, FcRn, and resin is represented. The model was assembled using the crystal structures of human FcRn (steel blue), β2m (medium aquamarine), Fc (light gray), Fab H chain (light salmon), and Fab L chain (lemon chiffon). PDB IDs of each structure are as follows: adalimumab Fab, 4NYL; Fc, 4w4N; and FcRn, 4N0U. The image of the resin is shown as a sphere colored in teal.
Analysis of methionine oxidized antibody using the FcRn column
First, we evaluated the effect of methionine oxidation of antibodies on the FcRn interaction. It is well known that oxidation of Met252 and Met428 in the Fc region impairs FcRn-binding and decreases its long serum half-life. Stracke et al. showed that exposure to H2O2 resulted in the oxidation of Met252 and Met42833. We prepared oxidized adalimumab and analyzed using the developed FcRn column. The overall chromatograms of the oxidized adalimumab at each H2O2 concentration are shown in Fig. 2. The non-oxidized adalimumab showed a sharp peak at approximately pH 7.7. As the H2O2 concentration increased, several peaks appeared before the peak of non-oxidized adalimumab, and overall peaks became lower and broader.

An overall view of the FcRn affinity chromatography is shown on the left. A zoomed-in view of the FcRn affinity chromatography is shown on the right. The solid lines represent the chromatogram of absorbance at 280 nm. The concentration of H2O2 used for the oxidation is shown on the right side. The fitted curves for each peak are colored in green, orange, blue, gray, and purple. The % gradient and pH values are shown on top.
Analysis of FcRn affinity-engineered mutants
To assess the effects of FcRn affinity-engineering, we prepared seven mutants (six with increased affinity, and one with impaired affinity) and analyzed them using the FcRn column. The chromatograms are shown in Fig. 3a. The six mutants with increased affinity (IH, LS, N434H, QA, QL, and YTE) showed delayed peaks compared to adalimumab. In contrast, the null mutant (LOW) showed a markedly earlier elution (flow-through), indicating that the mutant was not bound to the FcRn column. The peaks of the eluted antibodies were almost single, except for the IH mutant. Interestingly, the IH elution peak was split into several broad peaks.

a FcRn affinity chromatography of FcRn affinity engineered mutants. b Overall view of the ion-exchange chromatography. c A zoomed-in view of the ion-exchange chromatography. Solid lines represent the chromatogram of absorbance at 280 nm. The pH of elution is shown on top.
Ion-exchange chromatography was performed to assess the charge heterogeneity, especially for the IH mutant (Fig. 3b, c). The elution peaks reflected the overall charge of the antibody molecules. Thus, the order of antibody elution differed between the FcRn column and ion-exchange chromatography. Consistent with the results of FcRn affinity chromatography, only the IH mutant exhibited split and broad peaks.
Analysis of pH-dependent FcRn affinity
FcRn affinity chromatography is a method to assess pH-dependent FcRn affinity of IgG. Since antibodies that elute excessively late in FcRn affinity chromatography are likely to bind FcRn strongly at neutral pH, and therefore cannot exhibit extended serum half-life, we considered it important to reveal the correlation between the pH dependence of the FcRn affinity and the elution profiles of FcRn affinity chromatography. As an orthogonal assessment, we measured the FcRn affinity of the FcRn affinity-engineered mutants using SPR at pH 6.00, 6.25, 6.50, and 7.40 (Fig. 4 and Supplemental Table 1). To decrease the influence of bivalent binding of FcRn on the sensor chip, the amount of immobilized FcRn was reduced34. Overall, the affinities of these mutants were decreased with increasing pH (Fig. 4b). At pH 7.40, while the IH and LS mutants showed weak binding to FcRn, the other antibodies (adalimumab, N434H, QA, QL, and YTE) did not bind to FcRn. In particular, for the mutants, the plot of affinity at pH 6.0 versus the elution volume showed a modest correlation (Fig. 4c, d).

a SPR sensorgrams of FcRn binding on the sensor chip. The measurements were performed at pH 7.40 (top), pH 6.50, pH 6.25, and pH 6.00 (bottom). The black solid lines represent sensorgrams. The blue solid lines represent fitted sensorgrams. The highest analyte (antibody) concentration is shown above each sensorgram. The sensorgrams at pH 7.4 could not be fitted using the kinetic model. b The determined affinity (dissociation constant, KD) is plotted. The affinity values at pH 6.50 (blue), pH 6.25 (magenta), and pH 6.00 (green) are shown. c The correlation between FcRn affinity at pH 6.0 versus elution volume of FcRn affinity chromatography is shown. d The correlation for FcRn affinity-engineered mutants and its R-squared value for linearity is shown.
Analysis of 13 therapeutic monoclonal antibodies
Multiple studies have shown that the charges of antibodies affect the FcRn affinity12,27,29,31,35. However, which part of IgG has an impact on FcRn affinity, and to what extent the impact differs among several antibodies required exploration. We analyzed 13 different therapeutic monoclonal antibodies using FcRn affinity chromatography (Fig. 5a, b). Although these 13 antibodies are all IgG1 (three mutations in anifrolumab (L234F, L235E, and P331S) are distant from the binding interface of Fc-FcRn), the elution profiles differed remarkably. Among these 13 antibodies, omalizumab eluted first, and adalimumab eluted last.

a The FcRn affinity chromatography of therapeutic monoclonal antibodies is shown on the left. b A zoomed-in view of the chromatography. Solid lines represent the chromatogram of absorbance at 280 nm. The pH of elution is shown on top.
Furthermore, we assessed the correlation between the elution volume and the charges of each domain or chain of the antibodies. The amino acid sequence-based isoelectric point (pI) of each domain/chain was calculated and plotted (Fig. 6). The pI was calculated for the following domains/chains: whole, H chain, Fab, Fab H chain, L chain, VH and VL, VH, VL, CDR H and CDR L, CDR H, and CDR L. The highest R-squared value for linearity was 0.53, which was derived from the “L chain” and “CDR H and CDR L”. Overall, the R-squared values related to the L chain (L chain, 0.53; VL, 0.41; CDR L, 0.22) were higher than those for the H chain (H chain, 0.35 × 10−4; Fab H chain, 0.02; VH, 0.01; CDR H, 0.16). Although a clear correlation between the elution volume and pI was not observed, it is reasonable to conclude that the pI of the L chain, rather than that of the H chain, has an impact on FcRn affinity.

The x-axis shows the elution volume in FcRn affinity chromatography. The y-axis represents the pI for each domain/chain. Orange dotted lines indicate the 95% confidence band. The turquoise dotted lines indicate the 95% prediction band. The plot was generated using GraphPad Prism software.
Analysis of the charges in the L chain of antibodies
To focus on how the pI of the L chain affects FcRn affinity, we compared the amino acid sequences of the variable region of the L chain of the 13 antibodies (Fig. 7a). We observed that positively or negatively charged residues were located in variable regions other than the complementarity-determining region (CDR). In addition, the pattern of the charged residues varied depending on the antibody. Specifically, the Ser60 in adalimumab (numbered according to adalimumab) is an aspartic acid (negatively charged) in anifrolumab, durvalumab, and ipilimumab. The Ser77 in adalimumab is an arginine (positively charged) in anifrolumab, durvalumab, ipilimumab, and rituximab. The Gln79 in adalimumab is a glutamic acid (negatively charged) in anifrolumab, daratumumab, durvalumab, ipilimumab, necitumumab, ofatumumab, and rituximab. Thus, we hypothesized that the charged residues at these three positions might also affect FcRn affinity. These three residues are located on the lateral surface of the L chain (Fig. 7b).

a Comparison of amino acid sequences of the L chain for 13 antibodies. S60, S77, and Q79 (according to the numbering of adalimumab) are highlighted using arrows. The acidic amino acids are colored in red. The basic amino acids are colored in indigo blue. b The structure of adalimumab Fab is shown (PDB ID, 4NYL). c Ion exchange chromatography of adalimumab and charge-engineered mutants. d The FcRn affinity chromatography of adalimumab and charge-engineered mutants. e Elution volume of FcRn affinity chromatography of adalimumab and charge-engineered mutants. Error bars represent data from 3 independent experiments (n = 3). f SPR sensorgrams of FcRn binding on the sensor chip. The measurements were performed at pH 7.4 and pH 6.0. The black solid lines represent sensorgrams. The blue solid lines represent fitted sensorgrams. The sensorgrams at pH 7.4 could not be fitted using the kinetic model.
To test this hypothesis, we prepared charge-engineered adalimumab mutants and performed FcRn affinity chromatography. S60, S77, and Q79 of adalimumab L chain were mutated to charged residues: S60D, S77R, and Q79E, respectively. Ion-exchange chromatography analysis showed that the negative charge-introduced mutants (S60D and Q79E) eluted earlier than the parent adalimumab (Fig. 7c). Additionally, the S60D/Q79E mutant eluted earlier than the S60D and Q79E. In contrast, the elution peak of the positive charge-introduced mutant (S77R) was delayed. These results confirmed that the charge of the adalimumab was modified by the mutagenesis. Subsequently, we performed the FcRn affinity chromatography using these adalimumab L chain mutants (Fig. 7d, e). Consistent with the results of ion-exchange chromatography, the elution of S60D was earlier than the parent adalimumab, that of Q79E was slightly earlier, that of S60D/Q79E was further earlier, and that of S77R was delayed. The SPR results also supported the increase or decrease in FcRn affinity upon these mutations (Fig. 7f, Supplemental Table 1). These results strongly indicated that the charge on the lateral surface of the L chain also affects FcRn affinity.
Discussion
Multiple mechanisms, including phagocytosis, target-mediated internalization, and target-independent pinocytosis, are involved in the regulation of therapeutic antibody pharmacokinetics. In particular, the molecular interactions between therapeutic antibodies and FcRn affect the long serum half-life of antibodies. Thus, the in vitro FcRn affinity of IgG is an important indicator of its in vivo PK properties. However, the molecular basis of the IgG-FcRn interaction is not fully understood from experimental and structural perspectives. In this study, we developed an engineered FcRn-immobilized column. Using this FcRn column, we performed affinity chromatography and analyzed various types of antibodies to reveal the correlations between antibody characteristics and FcRn affinity. Furthermore, we focused on the effect of charges in the L chain on FcRn affinity from a structural perspective.
The FcRn affinity chromatography enabled an evaluation of FcRn affinity from the elution profile. The elution profiles of Met-oxidized antibodies and FcRn-affinity engineered antibodies were consistent with the previously reported results11,32,33,36,37,38. As the elution peaks of the Met-oxidized antibody were clearly separated, it is evident that this method possesses sufficient specificity as a performance characteristic to identify Met-oxidation of antibodies. The elution profiles of antibodies reflected their affinity for FcRn. Meanwhile, the antibody with abrogated binding to FcRn flows through the column without being captured. Hence, in contrast to SPR, FcRn affinity chromatography can be used to determine the molecular populations in samples based on FcRn affinity. Borrok et al. reported that the high affinity for FcRn at both acidic and neutral pH leads to rapid serum clearance10. To achieve FcRn-mediated half-life extension, antibody molecules must be released from FcRn when the antibodies are recycled back into the blood circulation. Given this, FcRn affinity evaluation at a single pH (e.g., pH 6.0) is not sufficient to indicate PK properties. Since the most important parameter to be evaluated is pH-dependent FcRn affinity, the pH gradient-based FcRn affinity chromatography is a highly effective methodology.
Using the FcRn column, we showed that the elution profiles varied depending on the antibodies, indicating that the sequence of the Fab region affected FcRn affinity. Given that the Fab arms are accommodated on the membrane surface when IgG binds to FcRn, as reported by Brinkhaus et al., it is reasonable that Fab also interacts with FcRn26. Furthermore, we showed that the charges in the L chain, rather than those in the H chain, affected FcRn affinity. These results were consistent with previous studies11,12,27,28,29,30,31. Since the involvement of the Fab domain in the IgG-FcRn interaction is not a highly specific interaction, the relationship between the coordinates of positive charges (i.e., their quantity and positions) and the FcRn affinity remains unclear. However, our study showed that the effect of positive charges on FcRn affinity is not solely dependent on the positive charges of a particular domain, such as Fab, VL, or CDR. The mutation analysis suggested that the charges on the lateral surface of the L chain also affect FcRn affinity.
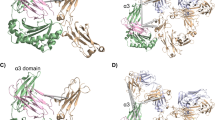
Based on these findings, we assembled a structural model of the complex of full-length IgG, FcRn, and human serum albumin (HSA) using the crystal structures (Fig. 8). Multiple studies have discussed the IgG-FcRn binding model on the membrane surface, including “standing-up”, “lying down”, “Y-shape”, “T-shape”, and “reclined model”22,23,24,26. We speculated the “reclined model” is reasonable considering that (1) the binding stoichiometry between IgG and FcRn is possibly 1:2, (2) the structural clash between the Fab arm and membrane surface should be avoided, and (3) FcRn plays an important role in salvaging and maintaining HSA in blood circulation, thus the simultaneous binding of HSA and IgG to FcRn should not be compromised22,39. In our model, the lateral surface of the L chain touches the bottom of the α3 domain of FcRn (or C-terminus near the membrane surface) and β2m. The model indicated that multiple negatively charged residues (shown in red) are present at the bottom region of FcRn and β2m. These negatively charged residues may interact with the Fab of IgG. Thus, this model may explain the mechanism by which charges in the L chain affect the FcRn affinity. In contrast, an elevated pI of IgG (positively charged) results in a shortened serum half-life7,35,40,41. This is presumably because the positively charged IgG can be captured by the negatively charged cell surface, resulting in increased nonspecific cellular uptake (clearance). Considering the contradiction between FcRn affinity-derived serum half-life extension and the non-specific cellular uptake-derived rapid clearance, the coordinates of positive charges (quantity and positions) are remarkably important.

a The assembled crystal structures of the complex of adalimumab Fab, Fc, FcRn, β2m, and HSA. PDB IDs of each structure are as follows: adalimumab Fab, 4NYL; Fc, 4w4N; and FcRn-β2m, 4N0U; and HSA, 4N0U. Each molecule is colored as follows: Fab H chain, light salmon; Fab L chain, lemon chiffon; Fc, light gray; FcRn, steel blue; β2m, medium aquamarine; and HSA, pale violet red. b Top and side view of the model of the complex on membrane surface. c Top view (left) and rotated view (right) of the Fc-FcRn complex. d Positively charged, negatively charged, and neutral residues in the FcRn and β2m are colored blue, red, and white, respectively. The models were generated by aligning the crystal structures using UCSF Chimera and ChimeraX.
Estimating the PK properties of therapeutic antibodies in humans remains challenging because multiple parameters, such as FcRn affinity of the antibody, administration dose, administration route, patient body weight, properties of target molecules, and underlying conditions, affect PK properties. Therefore, extensive efforts have been made to understand the correlation between FcRn affinity and PK properties. Schlothauer et al., Grevys et al., and Gjølberg et al. conducted comprehensive analyses, including affinity measurements, FcRn affinity column chromatography, in vitro cell-based assays, and experiments using human FcRn transgenic mice (in vivo)11,21,32,37. The results obtained using human FcRn transgenic mice did not always correlate with those of other experiments. As discussed in the literature, in vivo studies using human FcRn transgenic mice pose a few issues, including non-specific cellular uptake and ADA11,13. Grevys et al. revealed that a highly positively charged anti-IL 12/23 p40 antibody (briakinumab) had a shorter serum half-life than that of anti-IL 12/23 p40 antibody (ustekinumab), even though the elution of briakinumab by FcRn affinity chromatography was markedly later than that of ustekinumab11. As discussed above, this was presumably due to the rapid clearance of positively charged IgG via nonspecific cellular uptake. Furthermore, FcRn-mediated antibody recycling is a dynamic and complex event at the membrane surface that involves a heterogeneous IgG:FcRn binding ratio (1:1 to 1:2) and a transition from planar to tubular membrane structures42,43. Therefore, our FcRn affinity chromatography-based method is useful mainly for FcRn affinity analysis (quality assessment), and it remains challenging to extrapolate the results of FcRn affinity chromatography to in vivo/in human PK properties in some cases. Nevertheless, the progressive accumulation of knowledge regarding the structure and affinity of IgG-FcRn interactions could pave the way for more accurate prediction of PK properties.
Meanwhile, we were unable to determine the structure of the IgG-FcRn complex using transmission electron microscopy (TEM), even at low-to-medium resolution. Sun et al. visualized the IgG-FcRn complex using electron microscopy and discussed the Fab-FcRn interactions based on the conformation of IgG44. An increase in the resolution of the structure of the IgG-FcRn complex will unquestionably advance our understanding of IgG-FcRn interactions.
In conclusion, we elucidated structural insights into IgG-FcRn interactions using engineered FcRn-immobilized affinity chromatography. We hope that this study will lead to better and effective characterization methods for therapeutic antibodies. Moreover, we hope that our findings will encourage the development of novel molecular designs of therapeutic antibodies.
Materials and methods
Design of the FcRn column
Thermally stabilized human FcRn α-chain mutants were obtained as described previously45. Briefly, using the human FcRn α-chain gene as a template, error-prone PCR was performed to introduce one or two amino acid substitutions, thereby generating an FcRn mutant library. Subsequently, mutant FcRn proteins were expressed in Escherichia coli strain BL21 DE3 (NIPPON GENE, Cat# 312-06534) using the FcRn mutant library. FcRn mutants with improved thermal stability were identified based on their residual activity after heat treatment. By combining multiple mutations, an engineered human FcRn mutant with enhanced thermal stability was obtained.
The protein immobilized onto the resin included a human FcRn α-chain and β2m. The sequence of human FcRn α-chain was linked at the C-terminus of human β2m via a 5 × (GGGGS) linker. In other words, the protein consisting of β2m-GGGGS linker-FcRn (hereinafter referred to as FcRn) was immobilized on a TSKgel resin (particle size, 10 μm). The FcRn protein was expressed using E. coli strain BL21 (DE3) transformed with an expression vector. The cells were harvested and lysed using buffer (50 mM Bis-Tris Propane, 2.4 mM MgSO4, 150 mM NaCl, 5 units/mL benzonase, 0.006% (w/v) lysozyme, 0.6% (v/v) Triton X-100, pH 10.0). The extracted proteins were purified using Ni-NTA agarose (Wako, Cat# 143-09763) and IgG-Sepharose (Cytiva, Cat# 17061801). The C-terminus of FcRn was immobilized using an optimized, highly oriented coupling method. The amino acid sequence is shown in Supplemental Fig. 1. The FcRn-immobilized resins were packed in a solid support (4.6 mm I.D. × 50 mm). The analytical performance of the column was retained beyond 200 uses.
Differential scanning calorimetry (DSC)
The thermal stability of FcRn was determined using a PEAQ-DSC instrument (Malvern, UK). Wild-type FcRn was purchased from Gamma Proteins, Ltd. (Cat# HUFCRN-U). Mutant FcRn was the protein expressed and purified in-house. Protein samples were concentrated and buffer-exchanged with 50 mM phosphate, 150 mM NaCl (pH 6.0). The protein samples at a concentration of 10 μM were subjected to DSC measurements from 25 to 100 °C at a scan rate of 90 °C/h. The thermograms were normalized by subtracting the signal from the reference cell containing only buffer. The melting temperatures (Tm) were calculated by a standard fitting procedure using an evaluation software using a non-two-state model.
FcRn affinity chromatography
The FcRn affinity chromatography was performed using an ÄKTA Avant 25 (Cytiva, Sweden). Mobile phase A was 50 mM 2-morpholinoethanesulfonic acid (MES), 150 mM NaCl (pH 6.5), and mobile phase B was 50 mM Tris-HCl (pH 8.5). A total of 50 µg of antibody sample was diluted at least 10-fold with the mobile phase A to adjust the pH. A linear gradient of buffer B (0–100%) at a flow rate of 0.4 mL/min was applied to the FcRn column for 20 min to elute the antibody. The elution positions of the peaks in the chromatograms were determined based on the elution volume of the largest peak. The fitting of the chromatogram was performed using the Microsoft Excel Solver tool to minimize the differences between the data and the model.
Methionine oxidation
Adalimumab samples were subjected to H2O2 according to the protocols to induce methionine oxidation in the Fc region33,46,47,48. Adalimumab (Humira®) was purchased from Japanese pharmaceutical distributors. Adalimumab samples were buffer-exchanged and diluted to 1 mg/mL in a buffer containing 50 mM MES, 150 mM NaCl (pH 6.5). H2O2 was added to the antibody samples at 0.03, 0.05, 0.1, 0.25, and 0.5%. The reaction mixture was incubated for 17 h at 4 °C. The oxidation reactions were stopped by adding a buffer containing 5 mM L-methionine. L-methionine in the quenched sample was subsequently removed by buffer exchange using Amicon Ultra (Merck, Cat# UFC901024).
Preparations of adalimumab mutants
We prepared seven adalimumab mutants whose FcRn affinities were engineered in previous studies. The mutants were as follows: IH (P257I/N434H), LS (M428L/N434S), N434H, QA (T307Q/N434A), QL (T250Q/M428L), YTE (M252Y/S254T/T256E), and LOW (I253A/ H310A/H435A)19,49,50,51,52,53,54,55. The adalimumab wild type used in this study was derived from a therapeutic drug (Humira®), not prepared in-house. Expression vectors encoding the adalimumab variants were constructed in the previous study38. Antibodies were produced by using the Expi293 expression system (ThermoFisher Scientific, Cat# A14635). Expi293 cells (50 mL culture) were transfected using 50 µg pFuse vector (25 µg of H chain and 25 µg of L chain) and ExpiFectamin 293 Transfection Kit (ThermoFisher Scientific, Cat# A14525). After 6 days of cultivation, the supernatant was collected by centrifugation (40,000 × g, 30 min). The obtained supernatant was filtered through a 0.22 μm bottle top filter (Corning, Cat# 430626) and was subjected to protein A purification. The mutant LOW was subjected to protein L purification. After washing the column with 20 mM Tris-HCl, 150 mM NaCl (pH 7.2), the antibody samples were eluted with 50 mM glycine (pH 3.0) and neutralized with 1 M Tris-HCl (pH 8.0). The eluted antibody samples were further purified by size exclusion chromatography using a HiLoad 26/60 Superdex 200-pg column (Cytiva) equilibrated with 20 mM histidine, 80 mM NaCl (pH 6.0). Expression vectors of the charge-engineered mutants were prepared by Sino Biologicals (Beijing, China). The charge-engineered mutants were expressed as described above for the FcRn affinity-engineered mutants.
Ion exchange chromatography
Ion exchange chromatography was performed using a BioPro IEX SF 4.6 mm I.D. × 100 mm column (YMC, Cat# SF00S03-1046WP). Mobile phase A was CX-1 pH Gradient Buffer A (pH 5.6) (ThermoFisher Scientific, Cat# 085346), and mobile phase B was CX-1 pH Gradient Buffer B (pH 10.2) (ThermoFisher Scientific, Cat# 085348). A total of 50 µg of antibody sample was diluted at least 10-fold with mobile phase A to adjust the pH, and then the sample was applied to a Waters ACQUITY UPLC H-class system. A linear gradient of mobile phase B (0–100%) at a flow rate of 0.3 mL/min was applied to the column for 60 min to elute the antibody.
Surface plasmon resonance (SPR)
A Biacore 8 K instrument (Cytiva) was used to evaluate the binding of antibodies to FcRn. An Avi-tagged recombinant human FcRn-β2m was purchased (ACROBiosystems, Cat# FCM-H82W4). FcRn was immobilized on the CAP chip using the Biotin CAPture Kit (Cytiva, Cat# 28920234). FcRn (0.3 µg/mL) was injected for 60 sec, and the aimed immobilized level was 20 RU. The running buffer was 50 mM phosphate, 150 mM NaCl (pH 6.0). The regeneration solution was 20 mM Tris-HCl, 150 mM NaCl (pH 7.4). The association and dissociation time was 180 and 300 s, respectively. The concentrations of antibodies were as follows. The adalimumab was diluted using the running buffer by 2-fold serial dilutions from 400 to 12.5 nM. The adalimumab mutants were diluted using the running buffer in 2-fold serial dilutions from 100 to 3.1 nM.
Therapeutic monoclonal antibody sample
Adalimumab (Humira®), anifrolumab (Saphnelo®), bevacizumab (Avastin®), burosumab (Crysvita®), daratumumab (Darzalex®), durvalumab (Imfinzi®), ipilimumab (Yervoy®), necitumumab (Portrazza®), ofatumumab (Kesimpta®), omalizumab (Xolair®), rituximab (Rituxan®), trastuzumab (Herceptin®), and ustekinumab (Stelara®) were purchased from Japanese pharmaceutical distributors.
Amino acid sequence-based isoelectric point (pI) calculation
The amino acid sequence-based pI of the antibodies was calculated using the ProtParam tool (ExPASy)56.
Statistics and reproducibility
Error bars represent standard deviation. At least three experiments were conducted for SPR and chromatography using independently prepared samples.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The sequence of FcRn was obtained from UniProt entry; P55899 FCGRN_HUMAN. All data are available in the article itself and its Supplemental data. All other data are available from the corresponding author on reasonable request.
References
Crescioli, S. et al. Antibodies to watch in 2024. mAbs 16, 2297450 (2024).
Carter, P. J. & Lazar, G. A. Next generation antibody drugs: pursuit of the ‘high-hanging fruit. Nat. Rev. Drug Discov. 17, 197–223 (2018).
Dumontet, C., Reichert, J. M., Senter, P. D., Lambert, J. M. & Beck, A. Antibody-drug conjugates come of age in oncology. Nat. Rev. Drug Discov. 22, 641–661 (2023).
Labrijn, A. F., Janmaat, M. L., Reichert, J. M. & Parren, P. Bispecific antibodies: a mechanistic review of the pipeline. Nat. Rev. Drug Discov. 18, 585–608 (2019).
Kontermann, R. E. Strategies for extended serum half-life of protein therapeutics. Curr. Opin. Biotechnol. 22, 868–876 (2011).
Keizer, R. J., Huitema, A. D., Schellens, J. H. & Beijnen, J. H. Clinical pharmacokinetics of therapeutic monoclonal antibodies. Clin. pharmacokinet. 49, 493–507 (2010).
Dostalek, M., Gardner, I., Gurbaxani, B. M., Rose, R. H. & Chetty, M. Pharmacokinetics, pharmacodynamics and physiologically-based pharmacokinetic modelling of monoclonal antibodies. Clin. pharmacokinet. 52, 83–124 (2013).
Roopenian, D. C. & Akilesh, S. FcRn: the neonatal Fc receptor comes of age. Nat. Rev. Immunol. 7, 715–725 (2007).
Bussel, J. B., Cines, D. B. & Blumberg, R. S. Neonatal Fc receptor—biology and therapeutics. N. Engl. J. Med. 392, 1621–1635 (2025).
Borrok, M. J. et al. pH-dependent binding engineering reveals an FcRn affinity threshold that governs IgG recycling. J. Biol. Chem. 290, 4282–4290 (2015).
Grevys, A. et al. Antibody variable sequences have a pronounced effect on cellular transport and plasma half-life. iScience 25, 103746 (2022).
Datta-Mannan, A. et al. Balancing charge in the complementarity-determining regions of humanized mAbs without affecting pI reduces non-specific binding and improves the pharmacokinetics. mAbs 7, 483–493 (2015).
Chirmule, N., Jawa, V. & Meibohm, B. Immunogenicity to therapeutic proteins: impact on PK/PD and efficacy. AAPS J. 14, 296–302 (2012).
Opolka-Hoffmann, E. et al. The impact of immunogenicity on therapeutic antibody pharmacokinetics: a preclinical evaluation of the effect of immune complex formation and antibody effector function on clearance. mAbs 13, 1995929 (2021).
Reusch, J., Andersen, J. T., Rant, U. & Schlothauer, T. Insight into the avidity-affinity relationship of the bivalent, pH-dependent interaction between IgG and FcRn. mAbs 16, 2361585 (2024).
Burmeister, W. P., Gastinel, L. N., Simister, N. E., Blum, M. L. & Bjorkman, P. J. Crystal structure at 2.2 A resolution of the MHC-related neonatal Fc receptor. Nature 372, 336–343 (1994).
Burmeister, W. P., Huber, A. H. & Bjorkman, P. J. Crystal structure of the complex of rat neonatal Fc receptor with Fc. Nature 372, 379–383 (1994).
West, A. P. Jr & Bjorkman, P. J. Crystal structure and immunoglobulin G binding properties of the human major histocompatibility complex-related Fc receptor(,). Biochemistry 39, 9698–9708 (2000).
Martin, W. L., West, A. P. Jr, Gan, L. & Bjorkman, P. J. Crystal structure at 2.8 A of an FcRn/heterodimeric Fc complex: mechanism of pH-dependent binding. Mol. Cell 7, 867–877 (2001).
Jensen, P. F. et al. A Two-pronged binding mechanism of IgG to the neonatal Fc receptor controls complex stability and IgG serum half-life. Mol. Cell. Proteom. MCP 16, 451–456 (2017).
Gjolberg, T. T. et al. Biophysical differences in IgG1 Fc-based therapeutics relate to their cellular handling, interaction with FcRn and plasma half-life. Commun. Biol. 5, 832 (2022).
Trabjerg, E. et al. Identification of a direct interaction between the Fab domains of IgG antibodies and human FcRn upon IgG-FcRn complex formation. Commun. Biol. 8, 922 (2025).
Booth, B. J. et al. Extending human IgG half-life using structure-guided design. mAbs 10, 1098–1110 (2018).
Pyzik, M. et al. The neonatal Fc receptor (FcRn): a misnomer? Front. Immunol. 10, 1540 (2019).
Suzuki, T. et al. Importance of neonatal FcR in regulating the serum half-life of therapeutic proteins containing the Fc domain of human IgG1: a comparative study of the affinity of monoclonal antibodies and Fc-fusion proteins to human neonatal FcR. J. Immunol. 184, 1968–1976 (2010).
Brinkhaus, M. et al. The Fab region of IgG impairs the internalization pathway of FcRn upon Fc engagement. Nat. Commun. 13, 6073 (2022).
Liu, S., Verma, A., Kettenberger, H., Richter, W. F. & Shah, D. K. Effect of variable domain charge on in vitro and in vivo disposition of monoclonal antibodies. mAbs 13, 1993769 (2021).
Rossini, S. et al. V region of IgG controls the molecular properties of the binding site for neonatal FC receptor. J. Immunol. 205, 2850–2860 (2020).
Ollier, R. et al. Improved antibody pharmacokinetics by disruption of contiguous positive surface potential and charge reduction using alternate human framework. mAbs 15, 2232087 (2023).
Piche-Nicholas, N. M. et al. Changes in complementarity-determining regions significantly alter IgG binding to the neonatal Fc receptor (FcRn) and pharmacokinetics. mAbs 10, 81–94 (2018).
Schoch, A. et al. Charge-mediated influence of the antibody variable domain on FcRn-dependent pharmacokinetics. Proc. Natl. Acad. Sci. USA 112, 5997–6002 (2015).
Schlothauer, T. et al. Analytical FcRn affinity chromatography for functional characterization of monoclonal antibodies. mAbs 5, 576–586 (2013).
Stracke, J. et al. A novel approach to investigate the effect of methionine oxidation on pharmacokinetic properties of therapeutic antibodies. mAbs 6, 1229–1242 (2014).
Abdiche, Y. N. et al. The neonatal Fc receptor (FcRn) binds independently to both sites of the IgG homodimer with identical affinity. mAbs 7, 331–343 (2015).
Igawa, T. et al. Reduced elimination of IgG antibodies by engineering the variable region. Protein Eng. Des. Sel. PEDS 23, 385–392 (2010).
Cymer, F., Schlothauer, T., Knaupp, A. & Beck, H. Evaluation of an FcRn affinity chromatographic method for IgG1-type antibodies and evaluation of IgG variants. Bioanalysis 9, 1305–1317 (2017).
Grevys, A. et al. A human endothelial cell-based recycling assay for screening of FcRn targeted molecules. Nat. Commun. 9, 621 (2018).
Suzuki, T., Hashii, N., Tada, M. & Ishii-Watabe, A. The influence of antibody engineering on Fc conformation and Fc receptor binding properties: analysis of FcRn-binding engineered antibodies and an Fc fusion protein. mAbs 13, 1923366 (2021).
Schmidt, M. M. et al. Crystal structure of an HSA/FcRn complex reveals recycling by competitive mimicry of HSA ligands at a pH-dependent hydrophobic interface. Structure 21, 1966–1978 (2013).
Boswell, C. A. et al. Effects of charge on antibody tissue distribution and pharmacokinetics. Bioconjugate Chem. 21, 2153–2163 (2010).
Khawli, L. A. et al. Charge variants in IgG1: Isolation, characterization, in vitro binding properties and pharmacokinetics in rats. mAbs 2, 613–624 (2010).
Gan, Z., Ram, S., Vaccaro, C., Ober, R. J. & Ward, E. S. Analyses of the recycling receptor, FcRn, in live cells reveal novel pathways for lysosomal delivery. Traffic 10, 600–614 (2009).
He, W. et al. FcRn-mediated antibody transport across epithelial cells revealed by electron tomography. Nature 455, 542–546 (2008).
Sun, Y., Estevez, A., Schlothauer, T. & Wecksler, A. T. Antigen physiochemical properties allosterically effect the IgG Fc-region and Fc neonatal receptor affinity. mAbs 12, 1802135 (2020).
Asaoka, Y., Hatayama, K., Tsumoto, K., Tomita, M. & Ide, T. Engineering of recombinant human Fcgamma receptor I by directed evolution. Protein Eng. Des. Sel. PEDS 25, 835–842 (2012).
Bertolotti-Ciarlet, A. et al. Impact of methionine oxidation on the binding of human IgG1 to Fc Rn and Fc gamma receptors. Mol. Immunol. 46, 1878–1882 (2009).
Liu, H., Gaza-Bulseco, G., Xiang, T. & Chumsae, C. Structural effect of deglycosylation and methionine oxidation on a recombinant monoclonal antibody. Mol. Immunol. 45, 701–708 (2008).
Liu, D. et al. Structure and stability changes of human IgG1 Fc as a consequence of methionine oxidation. Biochemistry 47, 5088–5100 (2008).
Datta-Mannan, A. et al. Humanized IgG1 variants with differential binding properties to the neonatal Fc receptor: relationship to pharmacokinetics in mice and primates. Drug Metab. Dispos. Biol. Fate Chem. 35, 86–94 (2007).
Zalevsky, J. et al. Enhanced antibody half-life improves in vivo activity. Nat. Biotechnol. 28, 157–159 (2010).
Deng, R. et al. Pharmacokinetics of humanized monoclonal anti-tumor necrosis factor-alpha antibody and its neonatal Fc receptor variants in mice and cynomolgus monkeys. Drug Metab. dispos. Biol. Fate Chem. 38, 600–605 (2010).
Yeung, Y. A. et al. A therapeutic anti-VEGF antibody with increased potency independent of pharmacokinetic half-life. Cancer Res. 70, 3269–3277 (2010).
Hinton, P. R. et al. Engineered human IgG antibodies with longer serum half-lives in primates. J. Biol. Chem. 279, 6213–6216 (2004).
Dall’Acqua, W. F. et al. Increasing the affinity of a human IgG1 for the neonatal Fc receptor: biological consequences. J. Immunol. 169, 5171–5180 (2002).
Kim, J. K. et al. Mapping the site on human IgG for binding of the MHC class I-related receptor, FcRn. Eur. J. Immunol. 29, 2819–2825 (1999).
Gasteiger, E. et al. Protein identification and analysis tools on the ExPASy server. The Proteomics Protocols Handbook. https://doi.org/10.1385/1-59259-890-0:571 (2005).
Acknowledgements
This research was supported in part by AMED under Grant Number JP22ak0101188, and JP25ak0101280.
Author information
Authors and Affiliations
Contributions
M.K., T.S., N.I., and Y.T. designed the experiments. M.K., T.S., and N.I. conducted the research and analyzed the data. M.K. wrote the manuscript. M.K., T.S., N.I., R.O., T.Y., L.H., Y.T., H.S., T.I., S.N., K.T., and A.I. contributed to the preparation and discussion of this manuscript.
Corresponding author
Ethics declarations
Competing interests
N.I., R.O., T.Y., L.H., Y.T., and T.I. are employees of Tosoh Corporation. The other authors declare no competing financial interests. All the authors declare no non-financial competing interests.
Peer review
Peer review information
Communications Biology thanks Jordan Dimitrov and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Theam Soon Lim and Laura Rodríguez Pérez.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kiyoshi, M., Suzuki, T., Inoue, N. et al. Integrative structural insights into the IgG-FcRn interactions revealed by engineered FcRn-immobilized affinity chromatography. Commun Biol 9, 513 (2026). https://doi.org/10.1038/s42003-026-09789-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-026-09789-3
