
Abstract
Within- and between-city contexts and interactions shape our experiences of city life. However, a gap in understanding is how the wider landscape context of cities and the interactions with hinterlands influence urban ecology and health hazards. Using a meta-ecosystem framework, we fill this gap for the tick-borne Lyme disease ecological system by comparing the tick and Lyme disease hazards of urban and hinterland sites for 16 UK cities. We discover that the environmental hazards of ticks and Lyme disease of urban greenspaces are two- and threefold lower, respectively, than those of hinterland woodlands. Crucially, urban tick and Lyme disease hazards are shaped by tick abundance and the landcover (woodland and built-up) of hinterlands, but not of cities themselves. This highlights how rural–urban interactions form meta-ecosystems, and urban eco-epidemiology can depend on the characteristics of the surrounding rural landscape. Therefore, to better understand urban ecological processes and to mitigate disease risk in cities, it may be necessary to consider environmental factors in the hinterland such as landcover and disease hazard outside cities.
Similar content being viewed by others
Main
Our experiences of city life are shaped by our interactions with the social, cultural, economic, political and structural context of the areas within the city that we frequent, and this context can be influenced by economic, political and cultural interactions between separate cities. In addition to the within- and between-city interactions, cities may be influenced by the surrounding landscape context within which the city lies. For example, adjacent rural landcover can mitigate the urban heat island effect inside cities1; rural intensive livestock farming can impact the downstream water quality of cities2; various hazards originating in the hinterland, such as flood, fire and smoke, can disrupt city infrastructure3; and the movement of people can result in the spread of diseases between rural and urban areas4,5.
Vertebrate wildlife species that carry parasites and pathogens can move between rural and urban environments and colonize cities from the hinterland6,7, facilitated by ‘green corridors’8,9. We suggest that urban areas and their hinterlands can be viewed as two distinct but coupled socio-ecological systems that are linked by movements of resources (such as economic wealth, food or water) or vectors (such as vehicles, people or wildlife), thus forming a ‘meta-ecosystem’10. The meta-ecosystem concept has been used successfully to better understand interactions across many systems and scales. Classic examples include freshwater or marine nutrient flow on terrestrial ecosystems, particularly affecting soil nutrients, plant biomass and diversity11,12. However, the meta-ecosystem concept has not been applied to rural–urban socio-ecosystems, even though they are likely to interact and affect each other. In particular, how the environmental characteristics (such as landcover) of rural areas affect terrestrial ecological processes in urban areas, such as ecological interactions underlying disease emergence, is rarely addressed. Yet, understanding how hinterlands affect terrestrial urban disease risk is of fundamental importance, especially where disease mitigation strategies rely on the assumption that within-city factors are the prime risk factors.
One of the most important emerging environmental disease threats of Northern Hemisphere urban greenspaces in terms of prevalence and public interest is that of ticks and tick-borne diseases. Ixodes ricinus ticks are found extensively throughout Europe, western Asia and North Africa and have been reported in urban or peri-urban greenspaces in 24 European countries13. Ixodes ricinus ticks are generalist parasites that feed on the blood of most terrestrial vertebrate species and are able to transmit many pathogens of medical and veterinary importance. These include the tick-borne encephalitis virus complex, Anaplasma phagocytophilum, spotted fever rickettsiae and the Borrelia burgdorferi sensu lato (s.l.) complex of bacteria that causes Lyme disease14. Lyme disease is the most prevalent and widespread vector-borne disease of humans in the Northern Hemisphere15 and is increasing in many areas, especially in higher latitude regions, including the United Kingdom16.
Despite ticks and Lyme disease being reported in many urban greenspaces13, rural woodlands are the habitats most associated with higher tick densities and Lyme disease hazard17,18,19. There are multiple reasons for this, including the fact that roe deer (Capreolus capreolus), the most abundant and widespread deer species in the United Kingdom and Europe, prefer woodland and woodland edge habitats20. Deer are often the most important hosts for I. ricinus ticks and are the main drivers of tick abundance21,22. Compared to other habitats, woodlands also often have higher diversity and abundance of other I. ricinus hosts, such as rodents and birds23, that transmit B. burgdorferi s.l. bacteria to feeding ticks24. Furthermore, the mild and humid micro-climate created by woodland canopies and understory vegetation aid tick activity and survival17.
The most important ecological parameter to consider when assessing the public health threat from tick-borne diseases in the environment is the density of infected nymphs, which is termed the ‘environmental disease hazard’. In this Article we test the overarching hypothesis that urban and rural socio-ecosystems interact, acting as a meta-ecosystem, to shape the environmental tick and Lyme disease hazards of cities. One probable mechanism for tick population establishment in cities is the movement of terrestrial vertebrates into urban greenspaces from surrounding rural woodlands6,7 and the transport of ticks with them (a source–sink dynamic). Another plausible mechanism is the expansion of cities (urbanization) into rural habitats that already have existing tick populations, such that previously rural areas become new urban greenspaces that still retain these ticks and hosts. These mechanisms assume that there are more ticks25,26 and vertebrate hosts23,27 in rural than urban environments. We therefore predict (1) greater environmental hazards of ticks and Lyme disease in rural woodland than in urban greenspaces, and if either of these mechanisms of urban–rural interactions occurs, we also predict greater environmental hazards of ticks and Lyme disease in (2) cities whose hinterlands have higher tick densities, more optimal habitat (woodland) and less unsuitable habitat (built-up cover), and (3) cities that themselves have more woodland and less built-up cover. Habitat connectivity between the city and its rural woodlands for tick hosts, such as deer, could also be important in determining movements of ticks between rural hinterland woodland and cities. Thus, if there is not sufficient habitat connectivity between the city and its rural hinterland woodlands, there may not be more ticks in cities, even when they and their rural hinterlands have more ticks and woodland. Therefore, we also predict (4) greater hazards of ticks and Lyme disease in cities that have higher connectivity with rural woodlands (Fig. 1).

a,b, Under the assumption that there are more ticks and greater Lyme disease hazard in rural than urban environments (prediction 1), we predict (a) higher likelihoods of ticks and Lyme disease hazard in urban greenspaces where the hinterland has more woodland, less built-up cover and higher tick densities (prediction 2), and where the city has more woodland, less built-up cover (prediction 3) and higher connectivity (prediction 4) to hinterland woodlands. Conversely, we predict (b) lower likelihood of ticks and Lyme disease hazard in urban greenspaces where the hinterland has less woodland, more built-up cover and lower tick densities (prediction 2), and where the city has less woodland, more built-up cover (prediction 3) and lower connectivity (prediction 4) to hinterland woodlands.
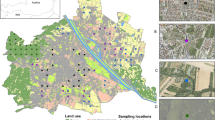
To test these predictions and fill the gaps in knowledge of how urban–rural interactions shape urban disease risk, we characterized the environmental hazards of ticks and Lyme disease in 16 small cities (population size 50,000–100,00028) and their hinterlands throughout the United Kingdom. Cities were chosen to encompass a broad range of hinterland landcovers and to cover a wide geographic area of the United Kingdom from northern Scotland to south-central England and Wales (Fig. 2, Supplementary Fig. 3 and Supplementary Table 1). By doing so, we formulated generalizable principles of how urban–rural interactions shape the tick and Lyme disease hazards of cities, as distinct from studying ticks and Lyme disease hazard in a city29. For a more complete characterization of these health hazards of urban United Kingdom, we mapped our new hazard values alongside the only other previously published urban UK values in five cities in southern England (Fig. 2).

a,b, Values for the 16 cities surveyed here (large symbols) and five cities surveyed in previous studies in southern England (small symbols) for tick hazard (the density of questing I. ricinus nymphs (DON); a) and Lyme disease hazard (density of nymphs infected with B. burgdorferi s.l. (DIN); b). For the 16 cities, the inner circle represents each city and the outer buffer represents each hinterland. Colors represent mean values over all urban sites (inner circle) and rural hinterland sites (outer circle) surveyed. Published data were compiled for Bath39, Bristol39, London38,41,50,51,52, Salisbury25 and Southampton39; these were not included in statistical analyses. Credit: the map contains public sector information from Open Geography Portal (ONS) licensed under the Open Government Licence v3.0 (UK outline).
Results and discussion
Tick and Lyme disease hazards of cities and hinterlands
We measured the density of questing (host-awaiting) nymphal I. ricinus ticks by counting nymphs on ‘blanket drag’ transects30, and the density of nymphs infected with B. burgdorferi s.l. using molecular testing, at five urban greenspaces within each city (0–5 km from the city center) and five hinterland woodlands (5–10 km from the city center) for each of the 16 cities in 2022 and 2023.
Ixodes ricinus nymphs were detected in six (37.5 %) of the 16 cities, but in twice as many, that is, 12 (75.0%), of the hinterlands (Fig. 2). Borrelia burgdorferi s.l. infected nymphs were detected in five (31.3%) of the cities and in eight (50.0%) of the hinterlands (Fig. 2 and Supplementary Table 1).
After taking into account confounding factors between cities (climate, month of survey and ground vegetation), statistical models showed that both the tick hazard index (the likelihood of finding a nymph on a transect; P < 0.001, Table 1, Fig. 3 and Supplementary Table 2) and Lyme disease hazard (the likelihood of finding an infected nymph at a site; P < 0.001, Table 1, Fig. 3, Supplementary Tables 2 and 3) were significantly higher in hinterland woodlands than urban greenspaces. This supported our first prediction of higher hazards in rural than urban environments and is not restricted to the cities that we surveyed in the United Kingdom. Similar patterns of I. ricinus and Lyme disease hazard increasing from the urban center outwards to the peri-urban environments have been reported from the south of England25 and continental Europe26,31,32. This suggests a general principle that recreation in urban parks is substantially less hazardous than recreation in nearby rural woodlands. This is probably because I. ricinus ticks and bacteria in the B. burgdorferi s.l. complex are carried by a wide range of terrestrial vertebrate species33 and, although some individual urban greenspaces can maintain a high abundance of certain vertebrate species34, the diversity and abundance of vertebrate species is generally higher in rural habitats23,27. Although we captured similar wooded habitats between rural and urban sites through our sampling design, urban woodlands can have fundamentally different properties compared to rural woodlands27. For example, rural woodlands are usually surrounded by other ‘green’ habitats such as pastures, and are often less fragmented than urban woodlands35. This can contribute to a higher abundance and diversity of vertebrates36,37, including deer, which are important I. ricinus hosts21,22. However, it is important to keep in mind that, although Lyme disease environmental hazard may be lower in cities than in hinterlands, the high numbers of urban greenspace users could have significant implications for public health at the population level.

The hazard of ticks is shown in orange (left) and Lyme disease hazard in blue (right). a,b, Rural hinterland woodlands had higher indices of I. ricinus nymph density (likelihood of nymphs on 10-m transects; 1,624 transects in hinterland sites, 1,630 transects in urban greenspaces; a) and Lyme disease hazard (likelihood of a site having I. ricinus nymphs infected with B. burgdorferi s.l.; 112 site visits in hinterland sites, 109 site visits in urban greenspaces; b). Statistical analyses were performed using GLMMs with binomial error structure and included site, city and survey month as random intercepts. P values are based on two-sided Wald Z-tests from the GLMM summary output. c,d, Higher I. ricinus nymph density in hinterlands was associated with higher hazard indices of ticks (c) and Lyme disease (d) in urban greenspaces. Statistical analyses were performed using GLMMs with binomial error structure and included site, city and survey month as random intercepts. P values are based on two-sided Wald Z-tests from the GLMM summary output. e,f, A higher percentage of woodland cover in hinterlands was associated with a higher urban tick hazard index (e), but urban Lyme disease hazard was not affected (f). Statistical analyses were performed using GLMMs with binomial error structure and included site, city and survey month as random intercepts. P values are based on two-sided Wald Z-tests from the GLMM summary output. g,h, A higher percentage of built-up cover in hinterlands was associated with a lower tick hazard index (g), but urban Lyme disease hazard index was not affected (h). Statistical analyses were performed using GLMMs with binomial error structure and included site, city and survey month as random intercepts. P values are based on two-sided Wald Z-tests from the GLMM summary output. Data are presented as model-predicted means ± 95% confidence intervals (CIs), with error bars (a,b) and shaded ribbons (c–h) representing CIs. Two-sided likelihood ratio tests comparing the full model to a reduced model using analysis of variance were used to assess the effect of each predictor, without adjustment for multiple comparisons. Test statistics and P values for each panel: χ² = 19.54, degrees of freedom (d.f.) = 1, P = 9.8 × 10−6 (a); χ² = 25.53, d.f. = 1, P = 4.6 × 10−7 (b); χ² = 23.8, d.f. = 1, P = 1.1 × 10−6 (c); χ² = 11.3, d.f. = 1, P = 7.6 × 10−4 (d); χ² = 2.69, d.f. = 1, P = 0.10 (e); χ² = 0.001, d.f. = 1, P = 1 (f); χ² = 7.85, d.f. = 1, P = 0.005 (g); χ² = 0.001, d.f. = 1, P = 1 (h).
Mapping the tick densities and Lyme disease hazards of the 16 cities we surveyed alongside published values from urban greenspaces in five cities in southern England showed that the United Kingdom is characterized by very low tick and Lyme disease hazard in the central regions (data from this study), flanked by higher hazards in the south of England (data from published studies25,38,39,40,41) and in Scotland, especially further north (data from this study; Fig. 2 and Supplementary Tables 1 and 3). A recent study of tick bites on humans, dogs and cats from 2013 to 2020 similarly showed that these central regions of England had the lowest number of records for I. ricinus bites in the United Kingdom42, suggesting this broad geographic pattern is generalizable and long term rather than being particular to our cities or study period.
Variation in tick and Lyme disease hazards among cities
To gain insights into why some cities have urban greenspaces with more ticks and higher Lyme disease hazard than other cities, we developed generalized linear mixed effect models (GLMMs) that statistically took into account the variance due to confounding factors between cities (which was important because tick and/or Lyme disease hazards could vary with latitude, population size, year, annual cumulative rainfall and ground vegetation density16,18).
The first two models tested part of our second prediction, that the tick and Lyme disease hazards of cities are influenced by tick densities in the hinterlands. In support of this prediction, cities had substantially higher indices of tick hazard (P < 0.001) and Lyme disease hazard (P = 0.002) in their greenspaces if hinterland woodlands had higher nymph densities (Table 1, Fig. 3 and Supplementary Table 2), and there were no instances where ticks or Lyme disease pathogens were found in a city but not in its hinterland (Fig. 2).
We then tested the final part of our second prediction, that urban tick and Lyme disease hazards are influenced by hinterland landcover. We used landcover metrics in a ring around the city between 5 and 10 km from the city centroid to represent ‘hinterland’. This area was chosen because all hinterland woodlands that we surveyed for ticks were between 5 and 10 km from the center, and the built-up areas of all 16 cities areas were contained within 5 km of the city centroid. We used percentage landcovers to represent habitats for ticks and hosts that are deemed most suitable (cover of broadleaved and coniferous woodlands) and less suitable (built-up cover)18,19,43,44. As predicted, hinterland woodland cover was retained in the best tick hazard model, indicating its influence on urban tick hazard (P = 0.08; Table 1, Fig. 3 and Supplementary Table 2). Previous studies have reported small-scale (within 100 m), within-ecosystem effects of tree or shrub cover on ticks and tick-borne pathogens in adjacent pastures44,45, but the particularly important and novel finding here is that rural woodlands are associated with the tick hazard of cities, that is, in a different socio-ecosystem several kilometers away. Given that hinterlands with more woodland cover are likely to also have higher densities of deer and other tick hosts such as small mammals and birds, two potential mechanisms might be at work: (1) there is higher likelihood of some of these hosts dispersing and establishing in adjacent urban greenspaces and (2) when there is urban encroachment, parts of these wooded hinterlands (with high densities of ticks and vertebrate hosts) may become new urban greenspaces that retain ticks and hosts.
We also found that less suitable hinterland habitat (built-up cover) was negatively associated with urban tick hazard index (P = 0.02; Table 1, Fig. 3 and Supplementary Table 2). We checked for a correlation between explanatory variables before running models, and hinterland woodland and built-up covers did not correlate, so this result is not due to more built-up cover equating to less woodland cover. Furthermore, both woodland and built-up cover were entered in the same statistical model, so the effect of built-up hinterland on urban ticks is after having taken into account the effect of hinterland woodland. This result thus suggests an inhibitory effect of hinterland built-up cover on tick hazard in the adjacent city’s greenspaces.
Despite the effect of hinterland landcover on urban ticks, we found little evidence that hinterland woodland or built-up cover measurably influenced the Lyme disease hazard of urban greenspaces (Table 1, Fig. 3 and Supplementary Table 2). This differential influence is interesting and could reflect different movement or dispersal distances between different host types. No nationwide data are available on densities or movements of wildlife hosts in and around cities in the United Kingdom, so we could not quantitatively examine which wildlife hosts contribute most to tick and Lyme disease hazards in cities. Research in rural environments repeatedly demonstrates the importance of deer in maintaining populations of generalist tick species including I. ricinus21,22,33,46, and deer are abundant in many rural, suburban and even some urban areas47,48. This evidence, along with our findings that urban ticks, but not Lyme disease hazard, are influenced by hinterland landcovers suggest that rural–urban tick transport may be driven primarily by wider-ranging tick hosts that do not carry the pathogens, such as deer24.
Transport of infected ticks from pathogen-carrying hosts such as small mammals, squirrels and birds24 must be frequent enough to seed pathogen into some urban greenspaces. However, the dispersal distances of these pathogen-carrying hosts are often shorter than for deer, so more local peri-urban and urban landcover, rather than hinterland landcover, may drive B. burgdorferi s.l. in urban greenspaces9,26,39,49. We hypothesize that, once transported, the ticks drop off into the ground vegetation and, when they emerge as the next instar the following spring, they could feed on local urban wildlife, starting new B. burgdorferi s.l. transmission cycles, influenced by local, within-greenspace characteristics9,26,39,49.
Against our third prediction, we found no effect of within-city woodland or built-up cover (within 5 km of the city centroid) on urban greenspace tick and Lyme disease hazard indices. This finding indicates that the hazards of ticks and Lyme disease of cities may be primarily shaped by hinterland factors (ticks and landcover), rather than by city-level landcover within the cities themselves. We had expected that cities containing more woodland cover would be more attractive to immigrating vertebrate hosts carrying ticks. Although we found no effect of within-city landcover on ticks or infected ticks per unit area of greenspace, it could still be the case that cities with more woodland cover may have more area over the whole city that contains ticks or tick-borne pathogens.
Contrary to our fourth prediction, we found no association between connectivity at the urban–hinterland interface and urban tick or Lyme disease hazards26,49. The fact that hinterland tick density and landcover does shape the urban hazards of ticks and Lyme disease, but urban–hinterland connectivity does not, implies that the cities in our study may have enough connectivity with their hinterlands despite the wide range of connectivity values (Supplementary Fig. 3). An example is Falkirk, which had ticks and B. burgdorferi s.l. in some urban greenspaces, despite low overall urban–hinterland connectivity. However, Falkirk had good connectivity in just one direction, which may have been enough for hosts carrying ticks to disperse into the city. This would suggest that even a small amount of connectivity could allow the rural environment to influence some of the ecological processes of cities.
Our key finding, that hinterland characteristics (tick density, woodland cover, built-up cover), rather than within-city landcover, influence the hazards of ticks and Lyme disease of cities, supports our hypothesis that rural and urban socio-ecosystems are linked to form a meta-ecosystem. Because ticks cannot move more than a few meters on their own, their dispersal is determined by the movement of their terrestrial vertebrate hosts. Accordingly, one likely mechanism for this ecosystem coupling is through animal movements, transporting ticks through the rural–urban interface as a means of establishing tick populations in cities. However, an additional mechanism explaining ticks and tick-borne pathogens in cities could be operating alongside that of host movements. This is the dynamic expansion of cities (urbanization) into the rural hinterland where wildlife and ticks are long established. No immigration of hosts is necessary if some urban greenspaces were, until they were enveloped by urban developments, rural woodlands with resident hosts and ticks. Further targeted research of greenspace history and animal tracking would be needed to tease these two mechanisms apart.
In this Article we have been asking why some cities have more ticks or tick-borne pathogen hazards in their parks than other cities, and we found that landcover in the hinterland is more important than within-city landcovers. However, within any single city, there will be differences in tick and tick-borne pathogen abundance between individual parks. At this finer spatial scale within a city, studies in North America and Europe have revealed that the proximity, connectivity or amount of nearby greenspace, tree cover or deer populations influence how individual greenspaces differ in their ticks and tick-borne pathogens9,26,39,49.
Conclusion
We included a wide range of cities and hinterlands across a country to examine how and why cities differ in their tick and Lyme disease hazards, an emerging environmental health issue for urban populations. We have described how the central regions of England have very low tick and Lyme disease hazards and that cities have lower hazards than their hinterlands, implying that recreation in urban parks is considerably safer from these hazards than in rural woodlands.
Our study provides generalizable principles about how urban socio-ecosystems interact with their rural hinterland contexts as part of an urban–rural meta-ecosystem. We show that one of the manifestations of this meta-ecosystem is that landcover and/or tick abundance in the rural ecosystem are key determinants of the hazards of ticks and Lyme disease of cities, probably reflecting differences in movement patterns between the main tick hosts and pathogen hosts.
Although we now know that differences in hinterland (but not within-city) characteristics determine the variation in tick and Lyme disease hazards between cities, previous research has shown that individual greenspace characteristics shape variation in these hazards between greenspaces within a city26,39. Therefore, disease mitigation strategies might be most effective if they target both spatial scales—the individual greenspace characteristics within a city, as well as targeting key hinterland areas. However, different districts often have separate administrative governance bodies. In these cases, collaboration between urban and rural governance bodies could enable more effective strategies to mitigate hazards to urban dwellers3.
Methods
Survey sites
We measured the relative abundance of questing I. ricinus nymphs and their infections with B. burgdorferi s.l. in 162 sites across 16 UK cities. The cities were spread throughout most of the United Kingdom, and our intention was to encompass a broad geographic area and to cover a wide range of woodland cover among the hinterlands (Fig. 2). To limit the confounding effects of city size, and to allow inclusion of Scottish cities (most of which are small), which helped broaden the range of hinterland landcovers, we used urban areas defined as ‘small cities’ with a population size of 50,000–100,000 (ref. 28). We did not survey cities in the southern regions of England, as I. ricinus tick densities and Lyme disease hazard have been studied previously in several southern England urban centers (Bristol, Bath, Salisbury, Southampton and London25,39,40,41,50,51,52; Fig. 2 and Supplementary Table 3). The densities of I. ricinus and Lyme disease hazard in urban environments in the regions we chose have not been published previously.
The cities included were (from north to south) Inverness, Perth, Falkirk, Dunfermline, Kilmarnock, Carlisle, Durham, Scarborough, Harrogate, Keighley, Halifax, Burnley, Macclesfield, Wrexham, Cannock and King’s Lynn (Fig. 2 and Supplementary Fig. 1). Ten sites in and around each city were surveyed: five were urban greenspaces within each city within 5 km of the city center, and five were in the hinterland woodlands between 5 and 10 km of the city center. The exception was Burnley, where 12 sites were surveyed (six urban and six rural). All rural hinterland sites were areas with woodland accessible to the public, and all urban greenspaces were public amenity areas such as local-authority-managed public parks with some tree cover or community woodlands. Each of the 162 sites was surveyed at least once, and cities for which questing nymphs were found in urban greenspaces were surveyed twice, resulting in 221 site visits.
Characterizing tick hazard
The environmental hazard of ticks (the density of questing I. ricinus nymphs) was measured using standard blanket drag transects, where each transect consisted of dragging a 1-m × 1-m square blanket material for 10 m (ref. 30). All nymphal ticks on the blanket were counted, collected and stored in 70% ethanol in 1.5-ml Eppendorf tubes for later pathogen analysis. At each of the 221 site visits, we conducted 15 transects per visit, except at 19 site visits, where 12 or 13 transects were conducted due to constraints of time and weather. Surveys were carried out between May and August in 2022 and 2023 (Supplementary Table 4). Although urban woodlands differ in many ways to rural woodlands, we strove to minimize the effect of habitat differences on ticks and Lyme disease hazard by surveying habitats that were as similar as possible across all sites. We achieved this by surveying primarily in areas of tree cover, with only a small proportion of transects in open grassland habitat, and surveying a range of ground vegetation types for both urban greenspaces and hinterland sites.
Because temperature and relative humidity affect the proportion of ticks that are active53, surveys were conducted only when the temperature was more than 10 °C, and transects were not conducted in the rain.
For ground vegetation, a density index was measured using a sward stick at the start (0 m), middle (5 m) and end (10 m) of each transect30. The sward stick had colored bands every 5 cm, and the vegetation density index was the number of these colored bands that were hidden by the vegetation when the sward stick was placed vertically in the vegetation at arm’s length.
Molecular analyses for Lyme disease hazard
To estimate the environmental hazard of Lyme disease (the density of infected I. ricinus nymphs) we tested each nymph for the presence of the B. burgdorferi s.l. complex of pathogens by using DNA extraction on each individual nymph and subjecting it to polymerase chain reaction (PCR). DNA was extracted using ammonia hydroxide54, by boiling each nymph at 100 °C in 150 µl of 0.7 M NH4OH for 15 min. The tubes were briefly centrifuged and heated at 100 °C for another 15 min with their lids open, until 70–100 µl of solution remained. The DNA was used as a template in a real-time PCR targeting the 23S rDNA gene55 for B. burgdorferi s.l. The qPCR used in this study55 was implemented using Meridian Bioscience MyTaq Mix (Scientific Laboratory Supplies) in a CFX Connect real-time PCR detection system (Bio-Rad). Each reaction of 25 μl contained 12.5 μl of master mix, 1 μl of each primer at 10 pmol μl−1 (forward, 5′-CGAGTCTTAAAAGGGCGATTTAGT; reverse, 5′-GCTTCAGCCTGGCCATAAATAG), 1 μl of probe at 3.2 pmol μl−1 (5′-FAM-AGATGTGGTAGACCCGAAGCCGAGTG-TAMRA), 7.5 μl of nuclease free water and 2 μl of DNA extract using a program consisting of 5 min at 95 °C, followed by 40 cycles of 15 s at 95 °C and 1 min at 57 °C. A threshold cycle or c(t) value of 36 or lower was considered a positive result. Due to unforeseen issues, the original assay was switched for another protocol targeting ospA21 part way through the study. In this protocol, each reaction of 20 μl contained 10 μl of master mix, 1 μl of primers (forward, 5′-AATATTTATTGGGAATAGGTCTAA; reverse, 5′-CTTTGTCTTTTTCTTTRCTTACAA G)/probe (5′-FAM-AAGCAAAATGTTAGCAGCCTTGA-BHQ-1) mix, 6 μl of nuclease free water and 3 μl of DNA extract using a program consisting of 5 min at 95 °C, followed by 60 cycles of 5 s at 94 °C and 35 s at 60 °C, with a final incubation at 37 °C for 20 s. All other parameters and equipment remained unchanged. For every five DNA samples tested we included a negative (PCR grade water) control. A c(t) value of 40 or lower was considered a positive result. For both assays, each positive sample was viewed separately to ensure the quality and smoothness of the curve. A subset of previously tested samples (n = 282) was retested using the ospA assay to ensure result consistency, and the two methods produced identical results (in other words, prevalence estimates were not affected by the protocol).
Landcover and connectivity metrics
For each of the 16 cities, for both urban (5-km radius from the city center) and rural (between 5 km and 10 km from the city center) areas, we extracted landcover metrics from the 2021 UKCEH landcover map43 using the packages sf56 and raster57 in R version 4.2.2. The landcover metrics we used were landcover map’s aggregate classes as follows: (1) percentage of woodland (broadleaved and coniferous) cover to represent the habitat in the United Kingdom usually associated with higher I. ricinus tick and Lyme disease hazard18,44 and (2) percentage of ‘built-up area and gardens’ (urban and suburban) cover to represent a landcover that we assume is one of the least suitable habitats for many of the key tick hosts such as deer (Supplementary Fig. 3). ‘Urban’ is characterized by more dense buildings and a lower vegetation signature than ‘suburban’43. Although suburban areas can include suitable habitat for deer and ticks, the landcover data we used categorizes areas of trees separately as ‘woodland’. The UKCEH landcover map 2021 information warns that ‘Suburban and Urban lie on a continuum and confusion is expected’43, hence our use of their aggregate built-up category.
For each city, we also created a connectivity metric to estimate the connectivity between the city and its rural hinterland. This was needed because, if there is little connectivity between the two socio-ecosystems, there may be no establishment of ticks in a city, even if its hinterland has a lot of woodland and ticks. For each city, we estimated connectivity for the city–hinterland interface, which was best represented by a 2-km-wide ring around each city, between 4 and 6 km from the city center. We applied the concept of ‘least-cost paths’, an estimate of the resistance of moving through the landscape, used to estimate connectivity between habitat patches26. Using the UKCEH 2021 landcover map, we attributed a cost to each landcover class, which represents the assumed cost to a deer moving through the landscape, based on published literature9,58,59,60,61. For example, moving through a built-up area has a cost of 1,000 compared to moving through woodland, which has a cost of only 1 (Supplementary Table 5). We computed the least-cost paths between woodland patches for each city using the planar network with two-dimensional nodes on a non-Euclidian resistance surface function from the grainscape package62, and calculated the average path cost for each urban–hinterland interface ring. This method does not distinguish patch sizes of woodland, assigning one cost for each pathway from one woodland patch to another. No path costs are calculated for deer movement within woodlands, even if they are large. Therefore, to allow for woodland patch size, with the assumption that moving within a woodland is lower cost than moving through other landcovers, we divided the average path cost by the average woodland patch size in the urban–hinterland interface and used this as our connectivity metric (Supplementary Fig. 2). A high metric value represents a high cost to deer movement (that is, low connectivity).
The statistical models needed to take into account other variables that we were not specifically interested in, but that might affect tick and Lyme disease hazards. For each city, we also extracted latitude, population size63 and annual cumulative rainfall, averaged for the period 1991–2020 from the UK Met Office online dataset64 (Supplementary Table 4).
Statistical analysis
All statistical analyses were performed in R version 4.2.2. For each of the models, we checked for potential collinearity between explanatory variables using variance inflation factors (VIFs), discarding variables scoring more than four, and Pearson’s correlation tests. We assessed whether continuous variables should be in a quadratic form using an F-test, and continuous variables were tested in both forms (linear and polynomial) against the response variable before model fit was assessed. Model selection was done using the dredge function from the MuMIN package65, and models with the lowest AICc were selected. Model diagnostics were assessed using the DHARMa package66. Statistical analyses were conducted on data collected from the 16 cities and hinterlands as part of this study. Previously published data from five cities in the south of England (published data) presented on the map (Fig. 2) were not included in the analyses.
The nymph count data distribution was strongly zero-inflated; that is, a large proportion of tick survey transects had zero nymphs and a large proportion of sites had zero infected nymphs. This caused both Poisson and negative binomial zero-inflated GLMMs to have poor model diagnostics if our response variables were the number of nymphs and infected nymphs per 10-m transect to represent questing nymph density and Lyme disease hazard, respectively. Therefore, for all analyses, we used binomial GLMMs with a logit link, with presence or absence of nymphs on each 10-m transect analyzed at the individual transect level (to represent questing nymph density), and the presence or absence of B. burgdorferi s.l. infected nymphs analyzed at the site visit level (to represent Lyme disease hazard), as the response variables. The values output from the models were therefore the predicted likelihood of a transect having at least one nymph (tick hazard index) or infected nymph (Lyme disease hazard index).
Differences between cities and their hinterland
To test our first prediction that hinterlands have higher tick and Lyme disease hazards than urban greenspaces, the initial full models included site type (urban or hinterland) and the following additional fixed effects that needed to be accounted for: year, ground vegetation density, latitude, city population size and annual cumulative rainfall. Site, city and month of survey were entered as random effects.
Effect of rural nymph densities
To test whether tick and Lyme disease hazards in urban greenspaces are influenced by nymph densities in the surrounding hinterland, we used nymph presence (transect level) and Lyme disease hazard (site visit level) in urban greenspaces as the response variables. The full models included the density of nymphs in the surrounding hinterland, year, ground vegetation density, latitude, population size and annual cumulative rainfall. Site, city and month of survey were entered as random effects.
Effects of hinterland landcover
To test the effect of hinterland landcover surrounding each city on urban tick and Lyme disease hazards, the full model included woodland and built-up cover in the surrounding hinterland of each city (5–10 km from city center), year, ground vegetation density, population size and annual cumulative rainfall. City latitude was not included as it was correlated with proportion of built-up cover (VIF = 16). Site, city and month were entered as random effects.
Effects of city landcover
To test the effect of landcover within each city on urban ticks and Lyme disease hazards, the full model included woodland and built-up cover within each city (0–5 km from city center), year, latitude, ground vegetation density, population size and annual cumulative rainfall. Site, city and month were entered as random effects.
Effects of urban–hinterland connectivity
To test the effect of the urban–hinterland interface connectivity for each city on urban tick and Lyme disease hazards, the full model included connectivity index, latitude, year, ground vegetation density and annual cumulative rainfall. City population size was not included as it was correlated with the connectivity metric (VIF = 7). Site, city and month were entered as random effects.
A table detailing each model is available in Supplemenary Table 6.
Reporting Summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this Article.
Data availability
Datasets used for data analysis are available at https://doi.org/10.24433/CO.6388252.v1. Any inquiry or request for materials should be addressed to S.L.G.
Code availability
The codes produced to conduct statistical analysis can be accessed at https://doi.org/10.24433/CO.6388252.v1.
References
Yang, M. et al. Mitigating urban heat island through neighboring rural land cover. Nat. Cities 1, 522–532 (2024).
Wen, Y., Schoups, G. & van de Giesen, N. Organic pollution of rivers: combined threats of urbanization, livestock farming and global climate change. Sci. Rep. 7, 43289 (2017).
Joines, M. et al. Cross-boundary risks of hinterland hazards to city infrastructure. Environ. Res. Infrastruct. Sustain. 4, 035004 (2024).
Andrianaivoarimanana, V. et al. Multiple introductions of Yersinia pestis during urban pneumonic plague epidemic, Madagascar, 2017. Emerg. Infect. Dis. 30, 289–298 (2024).
Johansen, I. C., Rodrigues, P. T. & Ferreira, M. U. Human mobility and urban malaria risk in the main transmission hotspot of Amazonian Brazil. PLoS ONE 15, e0242357 (2020).
Dickman, C. R. & Doncaster, C. P. The ecology of small mammals in urban habitats. II. Demography and dispersal. J. Anim. Ecol. 58, 119–127 (1989).
Harris, S. Distribution, habitat utilization and age structure of a suburban fox (Vulpes vulpes) population. Mammal Rev. 7, 25–38 (1977).
Castillo-Contreras, R. et al. Urban wild boars prefer fragmented areas with food resources near natural corridors. Sci. Tot. Environ. 615, 282–288 (2018).
VanAcker, M. C., Little, E. A. H., Molaei, G., Bajwa, W. I. & Diuk-Wasser, M. A. Enhancement of risk for Lyme disease by landscape connectivity, New York, New York, USA. Emerg. Infect. Dis. 25, 1136–1143 (2019).
Loreau, M., Mouquet, N. & Holt, R. D. Meta-ecosystems: a theoretical framework for a spatial ecosystem ecology. Ecol. Lett. 6, 673–679 (2003).
Hocking, M. D. & Reynolds, J. D. Impacts of salmon on riparian plant diversity. Science 331, 1609–1612 (2011).
Stapp, P., Polis, G. A. & Sánchez Piñero, F. Stable isotopes reveal strong marine and El Niño effects on island food webs. Nature 401, 467–469 (1999).
Hansford, K. M., Wheeler, B. W., Tschirren, B. & Medlock, J. M. Questing Ixodes ricinus ticks and Borrelia spp. in urban green space across Europe: a review. Zoonoses Public Health 69, 153–166 (2022).
Sprong, H. et al. Control of Lyme borreliosis and other Ixodes ricinus-borne diseases. Parasit. Vectors 11, 145 (2018).
Steere, A. C., Coburn, J. & Glickstein, L. The emergence of Lyme disease. J. Clin. Invest. 113, 1093–1101 (2004).
Gilbert, L. The impacts of climate change on ticks and tick-borne disease risk. Annu. Rev. Entomol. 66, 373–388 (2021).
Estrada-Peña, A. Distribution, abundance and habitat preferences of Ixodes ricinus (Acari: Ixodidae) in northern Spain. J. Med. Entomol. 38, 361–370 (2001).
Gilbert, L. in Ecology and Prevention of Lyme Borreliosis 161–171 (Wageningen Academic, 2016); https://doi.org/10.3920/978-90-8686-838-4_11.
Rosso, F. et al. Temporal changes in tick-borne pathogen prevalence in questing Ixodes ricinus across different habitats in the North-Eastern Italian Alps. MicrobiologyOpen 13, e010 (2024).
Hewison, A. J. et al. The effects of woodland fragmentation and human activity on roe deer distribution in agricultural landscapes. Can. J. Zool. 79, 679–689 (2001).
Gandy, S., Kilbride, E., Biek, R., Millins, C. & Gilbert, L. No net effect of host density on tick-borne disease hazard due to opposing roles of vector amplification and pathogen dilution. Ecol. Evol. 12, e9253 (2022).
Gilbert, L., Maffey, G. L., Ramsay, S. L. & Hester, A. J. The effect of deer management on the abundance of Ixodes ricinus in Scotland. Ecol. Appl. 22, 658–667 (2012).
McKinney, M. L. Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst. 11, 161–176 (2008).
Kurtenbach, K., Sewell, H.-S., Ogden, N. H., Randolph, S. E. & Nuttall, P. A. Serum complement sensitivity as a key factor in Lyme disease ecology. Infect. Immun. 66, 1248–1251 (1998).
Hansford, K. M. et al. Ticks and Borrelia in urban and peri-urban green space habitats in a city in southern England. Ticks Tick Borne Dis. 8, 353–361 (2017).
Heylen, D. et al. Ticks and tick-borne diseases in the city: role of landscape connectivity and green space characteristics in a metropolitan area. Sci. Tot. Environ. 670, 941–949 (2019).
Gilbert, O. L. The Ecology of Urban Habitats (Springer, 1991).
Dijkstra, L. & Poelman, H. Cities in Europe. The New OECD-EC Definition RF 01/2012 (European Commission, 2012).
McDonnell, M. J. in Urban Ecology: Patterns, Processes and Applications (eds Niemelä, J et al.) 5–13 (Oxford Univ. Press, 2011).
Gilbert, L. Altitudinal patterns of tick and host abundance: a potential role for climate change in regulating tick-borne diseases? Oecologia 162, 217–225 (2010).
Mathews-Martin, L. et al. Questing tick abundance in urban and peri-urban parks in the French city of Lyon. Parasit. Vectors 13, 576 (2020).
Pangrácová, L. et al. Ixodes ricinus abundance and its infection with the tick-borne pathogens in urban and suburban areas of eastern Slovakia. Parasit. Vectors 6, 238 (2013).
Gray, J., Kahl, O. & Zintl, A. What do we still need to know about Ixodes ricinus? Ticks Tick Borne Dis. 12, 101682 (2021).
Dondina, O. et al. Spatial and habitat determinants of small-mammal biodiversity in urban green areas: lessons for nature-based solutions. Urban For. Urban Green. 104, 128641 (2025).
Vizzari, M. & Sigura, M. Landscape sequences along the urban-rural-natural gradient: a novel geospatial approach for identification and analysis. Landsc. Urban Plan. 140, 42–55 (2015).
Bailey, S. Increasing connectivity in fragmented landscapes: an investigation of evidence for biodiversity gain in woodlands. For. Ecol. Manag. 238, 7–23 (2007).
Humphrey, J. W. et al. What can studies of woodland fragmentation and creation tell us about ecological networks? A literature review and synthesis. Landsc. Ecol. 30, 21–50 (2015).
Greenfield, B. P. J. Environmental parameters affecting tick (Ixodes ricinus) distribution during the summer season in Richmond Park, London. Biosci. Horiz. Int. J. Student Res. 4, 140–148 (2011).
Hansford, K. M. et al. Impact of green space connectivity on urban tick presence, density and Borrelia infected ticks in different habitats and seasons in three cities in southern England. Ticks Tick Borne Dis. 14, 102103 (2023).
Hansford, K. M. et al. Ixodes ricinus and Borrelia burgdorferi sensu lato in the Royal Parks of London, UK. Exp. Appl. Acarol. 84, 593–606 (2021).
Nelson, C., Banks, S., Jeffries, C. L., Walker, T. & Logan, J. G. Tick abundances in South London parks and the potential risk for Lyme borreliosis to the general public. Med. Vet. Entomol. 29, 448–452 (2015).
Gandy, S. L., Hansford, K. M. & Medlock, J. M. Possible expansion of Ixodes ricinus in the United Kingdom identified through the Tick Surveillance Scheme between 2013 and 2020. Med. Vet. Entomol. 37, 96–104 (2023).
Marston, C., Rowland, C. S., O’Neil, A. W. & Morton, R. D. Land Cover Map 2021 (Land Parcels, GB). NERC EDS Environmental Information Data Centre https://doi.org/10.5285/398dd41e-3c08-47f5-811f-da990007643f (2022).
Halos, L. et al. Ecological factors characterizing the prevalence of bacterial tick-borne pathogens in Ixodes ricinus ticks in pastures and woodlands. Appl. Environ. Microbiol. 76, 4413–4420 (2010).
Gilbert, L., Brunker, K., Lande, U., Klingen, I. & Grøva, L. Environmental risk factors for Ixodes ricinus ticks and their infestation on lambs in a changing ecosystem: implications for tick control and the impact of woodland encroachment on tick-borne disease in livestock. Agric. Ecosyst. Environ. 237, 265–273 (2017).
Mysterud, A. et al. Contrasting emergence of Lyme disease across ecosystems. Nat. Commun. 7, 11882 (2016).
Honda, T., Iijima, H., Tsuboi, J. & Uchida, K. A review of urban wildlife management from the animal personality perspective: the case of urban deer. Sci. Total Environ. 644, 576–582 (2018).
Medlock, J. M. et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 6, 1 (2013).
Estrada-Peña, A. Effects of habitat suitability and landscape patterns on tick (Acarina) metapopulation processes. Landsc. Ecol. 20, 529–541 (2005).
Dobson, A. D. M., Taylor, J. L. & Randolph, S. E. Tick (Ixodes ricinus) abundance and seasonality at recreational sites in the UK: hazards in relation to fine-scale habitat types revealed by complementary sampling methods. Ticks Tick Borne Dis. 2, 67–74 (2011).
Sorouri, R., Ramazani, A., Karami, A., Ranjbar, R. & Guy, E. C. Isolation and characterization of Borrelia burgdorferi strains from Ixodes ricinus ticks in southern England. Bioimpacts 5, 71–78 (2017).
Vollmer, S. A. et al. Host migration impacts on the phylogeography of Lyme borreliosis spirochaete species in Europe. Environ. Microbiol. 13, 184–192 (2011).
Gilbert, L., Aungier, J. & Tomkins, J. L. Climate of origin affects tick (Ixodes ricinus) host-seeking behavior in response to temperature: implications for resilience to climate change? Ecol. Evol. 4, 1186–1198 (2014).
Ammazzalorso, A. D., Zolnik, C. P., Daniels, T. J. & Kolokotronis, S.-O. To beat or not to beat a tick: comparison of DNA extraction methods for ticks (Ixodes scapularis). PeerJ 3, e1147 (2015).
Courtney, J. W., Kostelnik, L. M., Zeidner, N. S. & Massung, R. F. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 42, 3164–3168 (2004).
Edzer, P. & Bivand, R. Simple features for R: standardized support for spatial vector data. R J. 10, 439–446 (2018).
Hijmans, R. J. & van Etten, J. Raster: geographic analysis and modeling with raster data. R version 2.0-12 https://CRAN.R-project.org/package=raster (2022).
Balčiauskas, L., Wierzchowski, J., Kučas, A. & Balčiauskienė, L. Habitat suitability based models for ungulate roadkill prognosis. Animals 10, 1345 (2020).
Girardet, X., Conruyt-Rogeon, G. & Foltête, J.-C. Does regional landscape connectivity influence the location of roe deer roadkill hotspots? Eur. J. Wildl. Res. 61, 731–742 (2015).
Gurrutxaga, M., Rubio, L. & Saura, S. Key connectors in protected forest area networks and the impact of highways: a transnational case study from the Cantabrian Range to the Western Alps (SW Europe). Landsc. Urban Plan. 101, 310–320 (2011).
Loro, M., Ortega, E., Arce, R. M. & Geneletti, D. Assessing landscape resistance to roe deer dispersal using fuzzy set theory and multicriteria analysis: a case study in Central Spain. Landsc. Ecol. Eng. 12, 41–60 (2016).
Chubaty, A. M., Galpern, P. & Doctolero, S. C. The r toolbox grainscape for modelling and visualizing landscape connectivity using spatially explicit networks. Methods Ecol. Evol. 11, 591–595 (2020).
Estimates of the Population for the UK, England, Wales, Scotland and Northern Ireland (Office for National Statistics, 2024).
UK Climate Averages 1991–2020 (Met Office, 2020); https://www.metoffice.gov.uk/research/climate/maps-and-data/uk-climate-averages
Barton, K. Mu-MIn: Multi-model inference. R version 0.12.2/r18 https://R-Forge.R-project.org/projects/mumin/ (2009).
Hartig, F. DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R version 0.4.6; https://cran.r-project.org/web/packages/DHARMa/index.html (2022).
Acknowledgements
We thank I. Jones, H. Ravenswater, N. Broer, Y. Sun, S. Ackers-Johnson, M. Collins, A. Reynolds, K. Fife, A. Creton and A. Coole for help with fieldwork. We thank town councils, landowners and land managers across the United Kingdom for granting us access to sites used in our survey. We are grateful for comments on the paper from M. Dennis, G. Clay and J. Huck. This work is an output from the project ‘Maximising ecosystem services in urban environments’ (MEaSURE), funded by The Natural Environment Research Council UK (reference NE/W003120/1).
Author information
Authors and Affiliations
Contributions
L.G. and D.J. initiated the project and secured funding. L.G. and J.L.H. designed the methodology. L.G., G.P., R.J.B. and J.L.H. collected the data. G.P. conducted molecular analyses. S.L.G. analyzed the data and generated figures. K.W. sourced some landcover metrics. S.L.G. and L.G. led the writing of the paper. All authors contributed to reviewing and editing the paper. All authors read and approved the final paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Cities thanks Gerardo Acosta, Meredith VanAcker and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information (download PDF )
Supplementary Tables 1–6, Figs. 1–3 and references.
Source data
Source Data Fig. 2 (download CSV )
Spreadsheet to produce map in Fig. 2.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gandy, S.L., Hall, J.L., Plahe, G. et al. The dependence of urban tick and Lyme disease hazards on the hinterlands. Nat Cities 2, 948–957 (2025). https://doi.org/10.1038/s44284-025-00320-z
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s44284-025-00320-z
