
Abstract
Calvarial nerves, along with vasculature, influence skull formation during development and following injury, but it remains unclear how calvarial nerves are spatially distributed during postnatal growth and aging. Studying the spatial distribution of nerves in the skull remains a challenge due to a lack of methods to quantify 3D structures in intact bone. To visualize calvarial 3D neurovascular architecture, we imaged nerves and endothelial cells with lightsheet microscopy. We employed machine-learning-based segmentation to facilitate high-resolution characterization from post-natal day 0 (P0) to 80 weeks. We found that TUBB3+ nerve density decreased with aging with the frontal bone demonstrating earlier onset age-related nerve loss than the parietal bone. In addition, nerves in the periosteum and dura mater exhibited similar yet distinct temporal patterns of nerve growth and loss. While no difference was observed in TUBB3+ nerves during skeletal maturation (P0 → 12 weeks), we did observe an increase in the volume of unmyelinated nerves in the dura mater. Regarding calvarial vasculature, larger CD31hiEmcn- vessel fraction increased with aging, while CD31hiEmcnhi vessel fraction was reduced. Throughout all ages, calvarial nerves maintained a preferential spatial association with CD31hiEmcnhi vessels, however, this association decreased with aging. Additionally, we used a model of Apert syndrome to explore the impact of suture-related disease on neurovascular architecture. Collectively, this 3D, spatiotemporal characterization of calvarial nerves throughout the lifespan and provides new insights into age-induced neurovascular architecture.
Similar content being viewed by others
Introduction
A dense network of peripheral nerves reside throughout bone and have known pro-osteogenic roles during development and after injury.1 Following initial ossification in utero, murine bones continue to develop, then remodel throughout adulthood. Ultimately, their capacity to effectively remodel and regenerate diminishes, and this age-dependent loss is thought to be primarily mediated by cellular senescence.2 Osteoprogenitor cells reduce their numbers and proliferative capacity with aging, resulting in a loss of bone formation and increased bone fragility.3,4 Other cell types in the bone microenvironment also mediate bone loss with aging. In aged fracture healing studies, macrophages maintain a pro-inflammatory phenotype and exhibit reduced bone induction capacity compared to macrophages in young mice.5 Likewise, in aged long bones, Type H (CD31hiEmcnhi) blood vessel density declines, resulting in decreased perfusion, increased hypoxia and reactive oxygen species production, and significantly lower levels of hematopoietic factors.6 While these cell types have known roles in modulating bone structure throughout the lifespan, the peripheral nerves have not been implicated in mediating changes in bone structure during aging.
Bone contains sensory nerves, which generate the pain associated with headaches and bone fracture, as well as sympathetic nerves.7,8,9,10 Apart from skeletal pain, sensory nerves play roles in successful healing of bone fracture injuries. Additionally, both nerve subtypes are closely linked with blood vessels and play a role in their growth and differentiation.11,12,13 In bone, nerves are found in close vicinity of blood vessels, both in homeostasis and in bone fracture.8,9,10,14,15,16,17,18,19,20,21 CD31hiEndomucin (Emcn)hi blood vessels are a vascular subtype that is crucial for bone homeostasis, development, and response to injury, however, they have yet to be implicated in nerve signaling pathways.22,23 In addition to aging models, we explored the neurovascular architecture in an FGFR-related craniosynostosis syndrome, characterized by premature suture closure, as previous studies have found that inhibiting nerves through TrkA inhibition can lead to enhanced sutural closure and dysregulation of skeletal stem cell proliferation.24 Craniosynostosis syndromes are characterized by suture fusion early in development. A majority of craniosynostosis syndromes are associated with mutations in the FGFR2 gene and demonstrate fusion of one or multiple calvarial sutures.25 While skull development is affected in these syndromes, changes in cartilage, brain, upper face, airway, and mandible morphology have also been noted.25,26,27,28,29,30,31 Thus, the impact of these mutations is widespread and could involve changes to other factors within the sutural niche, such as nerves and vasculature.
Imaging the spatial distribution of nerves provides insights into how their interactions with other cell types change with age or disease. Recently, confocal imaging in combination with thick cryosection immunostaining was used to generate a neuroskeletal atlas of the murine long bone periosteum, which identified three patterns of periosteal innervation.32 In contrast, there remains a knowledge gap in our understanding of the complete 3D distribution of nerves in the skull, how they change with time, and their association with the vasculature. Similar to long bone, early studies in the murine calvarium identified nerves in small regions of the periosteum using traditional histology.16,21 Similarly, histologic cross-sections have been used to show that nerves traverse throughout the interparietal, parietal, and frontal bones, with nerves in the periosteum and dura mater that branched into the diploë regions. However, due to limitations in 3D imaging of large sections of tissue at high spatial resolution,33 a complete quantitative characterization of nerve distribution in the calvaria has not been feasible until now. A recently developed quantitative lightsheet microscopy (QLSM) workflow provided high-resolution 3D images of the entire murine calvarium.34 Analyzing the resulting images provides extensive quantitative data on the distribution and interactions of fluorescently stained cells or anatomic structures. Here, we used an analogous QLSM workflow to map the 3D architecture of nerves (TUBB3+, NeuF+, CGRP+, and TH+) and their spatial associations with vascular phenotypes over the entire murine calvarium over the primary lifespan (i.e. P0- 80 weeks) of C57BL/6 J mice.
We found a significant increase in nerve density from P0 to early adulthood (W4) and subsequent decreases in nerve density as the mice aged, thereby demonstrating for the first time the impact of aging on calvarial nerves. We identified elevations in unmyelinated sensory (CGRP+) and sympathetic (TH+) nerve densities during skeletal maturation of perinatal mice. For all the nerve markers analyzed, we identified their preferential spatial association with Type H blood vessels. Finally, the application of QLSM to a murine model of Apert syndrome, a disease characterized by premature suture closure and skull dysmorphogenesis, revealed that the neurovascular distribution in the calvaria was not dysregulated. We believe that these rich 3D imaging data will help to further elucidate the role of calvarial nerves in homeostasis and aging, and also aid our understanding of the changes in nerve patterning that occur following bone injury.
Results
QLSM enables a comprehensive 3D visualization of calvarial nerves
We successfully visualized the 3D distribution of beta-3 tubulin+ (TUBB3+)-stained nerves in the parietal and frontal bones and in the coronal and sagittal sutures using QLSM (Fig. 1a). Tissue clearing without decalcification (using 2’2-thiodiethanol) was effective for all murine ages studied and enabled effective visualization of 3D nerve architecture throughout the thickness of the calvarium (Fig. S1A). Due to the age-dependent background optical signal in the murine calvaria, we developed a pipeline based on the Ilastik®35 machine learning software to segment nerve structures with subsequent image analysis with Imaris® (Fig. S1B). Briefly, this pipeline followed our previous method of image conversion and stitching using Imaris® software,34 followed by cropping of the parietal bone, coronal suture, and frontal bone regions for region-specific analyses. Following export of the images into the ImageJ software36 and color channel separation, z-stacks were segmented using Ilastik®, which resulted in more accurate segmentation of nerve structure. These segmented nerve data were then imported back into Imaris® for downstream volume and spatial analyses. When the Ilastik® pipeline was compared to that from Imaris®, we observed that nerve structures were more clearly visible, background noise was reduced, and quantitative data was accurate (Fig. S1C, D). Thus, this pipeline was combined with our blood vessel workflow34 for all subsequent analyses.

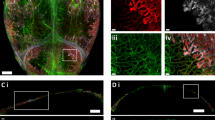
Quantitative lightsheet microscopy (QLSM) enabled 3D visualization of cranial nerves. a Maximum intensity projection (MIP) image of TUBB3+ nerves in the entire murine calvarium. Scale bar is 1 500 μm. b MIP image of the distribution of parietal nerves in the periosteum (blue) and dura mater (magenta). Scale bar is 300 μm. c 50 μm coronal cross-section of the calvarium acquired with QLSM illustrating TUBB3+ nerves. Scale bar is 1 000 μm. Inset scale bar is 300 μm
In addition to using Imaris® to compute volume and spatial association data, we also used its surface segmentation classification feature to identify nerves that were localized to either the periosteum or the dura mater (Fig. 1b, Supplementary Video 1). Next, we further explored quantitative differences in nerves within these subregions and explored the changes in their localization to these regions during aging.
In comparison to previous methods for visualizing calvarial nerves, our method enabled the 3D visualization of nerves throughout the entire murine calvarium. Although thick cryosections enable adequate visualization of nerves in 2D within specific regions, their 3D structures or interactions can be altered during sectioning. Using QLSM, we could generate cross-sectional images similar to those from thick cryosections, while also generating a 3D image of nerve distribution throughout the frontal and parietal bones without decalcification, dehydration, or geometric distortion (Fig. 1c, Fig. S2A).
To evaluate the ability of our imaging workflow for visualizing the network of nerves in the skull at high resolution, we also used confocal imaging to visualize TUBB3+ nerves at both 10× and 20× magnification (Fig. S2B, C). At 10× magnification (i.e. 1.25 μm resolution), which is similar to the resolution we employed for QLSM (i.e.1.3 μm resolution), nerves exhibited similar patterns in the parietal bone (Fig. S2B). At 20× magnification (i.e. 0.6 μm resolution), we used the DiameterJ plugin in ImageJ® and determined the average nerve diameter to be 4.013 ± 1.57 μm (n = 3) (Fig. S2C), which was in agreement with previous studies in sensory and sympathetic nerves.37,38 Assuming a normal nerve diameter distribution, this suggests that QLSM captures nearly 95% of the TUBB3+ nerves.
Calvarial nerve density and spatial distribution varies across murine lifespan
Using TUBB3 labeling, we imaged the distribution of calvarial nerves at post-natal day 0 (P0), 4 weeks, 12 weeks, 40 weeks, and 80 weeks calvaria to represent neonatal, developing, skeletally mature, middle-aged, and aged timepoints, respectively. From 4 weeks onwards, the parietal bone regions exhibited a mesh-like nerve network throughout the bone (Fig. 2a). The coronal suture region exhibited nerves that appeared to originate in the suture or along the borders of the bone and projected into the frontal and parietal bones. In aged samples, few nerves were found in the coronal suture region with one large nerve traversing the suture itself. At all ages, the frontal bone exhibited nerves that originated from nearby sutures and traveled radially into the center of the frontal bone (Fig. 2a). While we did not quantitatively analyze nerves in the sagittal and interfrontal sutures, we also observed nerves that originated from these sutures (Fig. S4D). For example, in the sagittal suture, a large nerve bundle was wrapped around the sagittal sinus and expressed high levels of TUBB3 staining at all ages.

Changes in TUBB3+ nerve distribution over the mouse lifespan. a MIP images of the parietal bone, coronal suture, and frontal bone regions of P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks mice with TUBB3+ nerves. Scale bar is 300 μm. Volume fraction calculation for P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks TUBB3+ nerves in (b) Overall calvaria, (c) Parietal Bone Region, (d) Coronal Suture Region, and (e) Frontal Bone Region, respectively. Data are mean ± SD. Differences were detected using a two-way ANOVA with post-hoc Tukey HSD (Honestly Significant Difference) test. *P < 0.05, **P < 0.01 where designated
Following quantitative analysis, after an initial increase from P0 to 4 weeks samples, no difference in nerve volume density was seen between 4 weeks, 12 weeks, or 40 weeks samples, while there was a steep drop-off in nerve volume density in 80 weeks samples (Fig. 2a). This trend was consistent in the parietal bone and coronal suture, but not in the frontal bone (Fig. 2c–e). In the frontal bone, there was a significant decrease in nerve volume density from 12 weeks to 80 weeks, but there was also a trend towards a decrease in 40 weeks samples, as nerve volume density was not maintained throughout 40 weeks samples (Fig. 2e). This may be indicative that nerve loss with aging may occur faster in the frontal bone than in the parietal bone. We further observed this age-related loss of nerves in calvarial cryosectioned cross sections (Fig. S3). Dense TUBB3+ staining was found throughout the dura mater and periosteum of 40 weeks cross sections, however, little TUBB3+ staining was found in 80 weeks samples. Given the limited section of the calvaria that is contained within these sections, it is difficult to quantify the nerve densities, especially within the periosteal and dura mater regions alone, further emphasizing the need for 3D quantitative imaging for accurate quantification.
In QLSM cross sectional images, we visualized nerves traveling through transcortical canals with and without blood vessels (Fig. S4C, yellow and white arrowheads). Using Imaris’s® machine learning classification feature, we quantified periosteal and dural nerve populations at all ages. Overall, only 80 weeks samples demonstrated a significant difference between periosteal and dural nerves proportions (Fig. 3a–e). When comparing periosteal and dural volume densities between ages, we found an overall and parietal bone age-dependent increase in the periosteum through from P0 to 4 weeks samples, which was maintained through 40 weeks samples and followed by a steep drop off at 80 weeks (Fig. 3f, g). In comparison, in the dura mater, the increase in dural nerves from P0 to 4 weeks samples was still present, however, not as dramatic as in the periosteum (Fig. 3j, k). In the coronal suture region, no significant differences were found (Fig. 3h, l). In the frontal bone, only the periosteal samples demonstrated significant increases trends in nerve densities (Fig. 3i, m). Further, the loss of nerves in 40 weeks samples was only seen in periosteal samples, suggesting that early onset (40 weeks) frontal bone nerve loss is primarily restricted to the periosteum. Regardless, in both the periosteum and the dura mater, frontal bone nerve densities were severely diminished in 80 weeks samples.

Spatial changes in periosteum and dura mater TUBB3+ nerve distribution over the mouse lifespan. a 50 μm coronal cross sections of TUBB3+ nerves in P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks mice. Scale bar is 300 μm. Nerve fraction calculations for TUBB3+ periosteal and dural nerves at P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks in (b) Overall Calvaria, (c) Parietal Bone, (d) Coronal Suture, and (e) Frontal Bone. TUBB3+ periosteal nerve volume density calculations at P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks in (f) Overall Calvaria Periosteum, (g) Parietal Bone Periosteum, (h) Coronal Suture Periosteum, and (i) Frontal Bone Periosteum. TUBB3+ dural nerve volume density calculations at P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks in (j) Overall Calvaria Dura Mater, (k) Parietal Bone Dura Mater, (l) Coronal Suture Dura Mater, (m) Frontal Bone Dura Mater. Data are mean ± SD. Statistics were performed with a two-way ANOVA with post-hoc Tukey HSD test. *P < 0.05, **P < 0.01, ****P < 0.000 1 where designated
Nerves preferentially associate with shifting CD31hiEmcnhi vessel populations over the murine lifespan
We have previously mapped CD31hiEmcnhi blood vessels, along with CD31hiEmcn- and CD31loEmcnhi blood vessels in the young and adult murine calvaria.34 Further, we identified the spatial association between CD31hiEmcnhi vessels and osteoprogenitors. In this study, we hypothesized that nerves may demonstrate a preferential association with CD31hiEmcnhi vessels due to their known roles in bone development and healing. To first investigate the blood vessel phenotype densities with aging, P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks calvaria were stained with CD31 and Emcn (Fig. 4a, Fig. S4C). CD31+ vessels were found throughout the calvaria with similar morphologies at all ages. CD31+ vessels increased in diameter and slightly decreased in volume density with aging (Fig. 4a, Fig. S4A, D). Emcn+ vessels demonstrated drastic morphological changes with aging, as bone marrow-resident sinusoidal blood vessel signal increases with age, while the Emcnhi signal virtually disappeared (Fig. 4a, Fig. S4A, D). When quantified, the Emcnhi dramatically dropped from P0 to 4 weeks of age, and then slowly declined as aging progressed, although not significantly. Due to the poor signal-to-noise ratio (SNR) of Emcnlo vessels, we did not quantify the Type L sinusoidal (CD31loEmcnlo) phenotype.

Changes in the spatial distribution of vascular phenotypes during aging. a MIP images of the parietal bone region of P0, 4 weeks, 12 weeks, and 80 weeks calvaria with vasculature stained with CD31 (green) and Emcn (red). Scale bar is 300 μm. b Vessel volume density calculation for P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks mice. c–e Phenotypic vessel volume density calculation for (c) CD31hiEmcn-, (d) CD31hiEmcnhi, and (e) CD31loEmcnhi vessels in P0, 4 weeks 12 weeks 40 weeks, and 80 weeks mice. f–h Phenotypic vessel phenotype fraction calculation for (f) CD31hiEmcn-, (g) CD31hiEmcnhi, and (h) CD31loEmcnhi vessels in P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks mice. Data are mean ± SD. Statistics were performed with a two-way ANOVA with post-hoc Tukey HSD test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.000 1 where designated
Overall blood vessel density remained unchanged with aging (Fig. 4b). However, blood vessel phenotypes demonstrated different spatial patterns with aging (Fig. 4c–h, Fig. S5). CD31hiEmcn- vessels increased in density and phenotype fraction with aging (Fig. 4c, f), while CD31hiEmcnhi vessels significantly decreased in both density and phenotype fraction (Fig. 4d, g). No differences were found with CD31loEmcnhi blood vessels. (Fig. 4e, h). To ensure an unbiased estimate of blood vessel phenotypes, high and low measurements for CD31 and Emcn blood vessel phenotypes were generated using binary masks. Following masking, CD31hiEmcn- vessels appeared green, CD31hiEmcnhi vessels yellow, and CD31loEmcnhi vessels red (Fig. S6). Overall vessel volume and vessel phenotype volumes were then generated from the spatially down-sampled segmented images.
To visualize the spatial relationship of nerves with blood vessel phenotypes, CD31+ and Emcnhi blood vessel staining was combined with TUBB3+ nerve staining (Fig. 5a). While some nerves were seen traveling adjacent to larger vessels, most nerves did not directly track alongside vessels in the calvaria (Fig. 5a). Overall, we observed that the network of nerves resided in the same plane as the blood vessel network but did not exhibit the same spatial patterns. For spatial association analysis, blood vessels and nerves were separated into 10 μm volumes using the Imaris® down-sampled segmented images to calculate the Euclidean distance for each individual section of the nerve segmentation to that of the blood vessels (Fig. S6).

Spatial correlation between vessel phenotypes and TUBB3+ nerves. a Masked MIP images of the parietal bone, coronal suture, and frontal bone regions of P0, 4 weeks, 12 weeks, 40 weeks, and 80 weeks mice with TUBB3+ nerves (blue) and vasculature stained with CD31 (green) and Emcn (red). Blood vessel phenotypes have been segmented based on binary masking to visualize CD31hiEmcn-, CD31hiEmcnhi, and CD31loEmcnhi blood vessles. Scale bar is 300 μm. b The fraction of nerves within 10 μm of each vessel phenotype is displayed for (b) P0, (c) 4 weeks, (d) 12 weeks, (e) 40 weeks, and (f) 80 weeks. Data are mean ± SD. Statistics were performed with a two-way ANOVA with post-hoc Tukey HSD test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.000 1 where designated
Following classification of vessel phenotypes, nerves were found to spatially associate with CD31hiEmcnhi vessels for all ages studied, although the degree of association significantly reduced with aging (Fig. 5b–f, Fig. S4E–I). At all adult ages, this trend was significantly higher than for CD31hiEmcn- vessels. This trend was not exclusively due to the prevalence of each vessel phenotype, as CD31hiEmcn- vessels were equally or even more prevalent than CD31hiEmcnhi vessels in 40 weeks and 80 weeks calvaria, respectively (Fig. 4c, d).
Small, unmyelinated nerve subtypes increase with postnatal development, while large, myelinated nerves remain unaffected
While TUBB3+ nerves were impacted by aging, we did not find any substantial differences in TUBB3+ nerve distributions or spatial distribution during skeletal maturation that occurs in early adulthood. Murine bones continue to develop after birth and are considered skeletally mature around 10 weeks of age.39 Furthermore, previous studies have determined that nerves are essential to proper bone development.1 Thus, we explored changes in sensory markers, neurofilament (NeuF) and calcitonin gene-related peptide (CGRP), and sympathetic marker, tyrosine hydroxylase (TH), during (i.e., 4 weeks) and after (i.e., 12 weeks) postnatal development and skeletal maturation. CGRP+ and TH+ nerves demonstrated a higher density following postnatal development, while NeuF+ nerves were unchanged, suggesting that some nerve subtypes may expand, mature, and specialize along with bone maturation (Fig. 6a, b, f, g, k–l, Fig. S7). Overall, increases in CGRP+ and TH+ nerves seemed to be consistent throughout the calvaria with slight increases found in the parietal bone, coronal suture, and frontal bone for both markers (Fig. S7).

Changes in nerve subtype distribution and neurovascular interactions during postnatal development. a MIP images of the parietal bone region of 4 weeks and 12 weeks mice with NeuF+ nerves (blue) and vasculature stained with CD31 (green) and Emcn (red). Scale bar is 300 μm. b Nerve volume fraction calculation for 4 weeks and 12 weeks NeuF+ nerves. c Periosteal and dural nerve fraction calculations for NeuF+ nerves in 4 weeks and 12 weeks mice. d, e Periosteal and dural nerve volume density calculations for NeuF+ nerves in 4 weeks and 12 weeks samples in (d) Overall Calvaria Periosteum, e Overall Calvaria Dura Mater. f Maximum intensity projections of the parietal bone region of 4 weeks and 12 weeks mice with CGRP+ nerves (blue) and vasculature stained with CD31 (green) and Emcn (red). Scale bar is 300 μm. g Nerve volume fraction calculation for 4 weeks and 12 weeks CGRP+ nerves. h Periosteal and dural nerve fraction calculations for CGRP+ nerves in 4 weeks and 12 weeks mice. i, j Periosteal and dural nerve volume density calculations for CGRP+ nerves in 4 weeks and 12 weeks samples in (i) Overall Calvaria Periosteum, j Overall Calvaria Dura Mater. k Maximum intensity projections of the parietal bone region of 4 weeks and 12 weeks mice with TH+ nerves (blue) and vasculature stained with CD31 (green) and Emcn (red). Scale bar is 300 μm. l Nerve volume fraction calculation for 4 weeks and 12 weeks TH+ nerves. m Periosteal and dural nerve fraction calculations for TH+ nerves in 4 weeks and 12 weeks. n,o Periosteal and dural nerve volume density calculations for TH+ nerves in 4 weeks and 12 weeks samples in (n) Overall Calvaria Periosteum, o Overall Calvaria Dura Mater. p–r Spatial association calculations for nerves and vessels phenotypes for (p) NeuF+, (q) CGRP+, and (r) TH+ nerves. The fraction of nerves within 10 μm is displayed for each vessel phenotype. Data are mean ± SD. Statistics were performed with a two-way ANOVA with post-hoc Tukey HSD test and a two-tailed t-test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.000 1 where designated
In terms of their distribution in the periosteum and dura mater, only NeuF+ and TH+ nerves demonstrated a preferential localization to the periosteum at 4 weeks; however, this localization was lost after postnatal development in both cases (Fig. 6c, h, m, Fig. S8). Interestingly, this trend was consistent in the parietal bone for both NeuF+ and TH+ nerves and in the frontal bone for NeuF+ nerves, but no changes were seen in the other regions (Fig. S8). When periosteum and dura mater nerve volumes were considered, we found an increase in nerve volume density in both the periosteum and dura mater for CGRP+ nerves, but only an increase in the dura mater for TH+ nerves (Fig. 6d, e, i, j). This trend was consistent throughout the regions of the calvaria. CGRP+ nerves increased in every region with postnatal development, while TH+ nerves exhibited only regional increases in the dura mater (Fig. S9). Thus, the unmyelinated nerve growth after postnatal development preferentially occurred in the dura mater for TH+ nerves and throughout the periosteum and dura mater for CGRP+ nerves. No differences were observed for dura and periosteum densities of NeuF+ nerves (Fig. 6c). For these nerve subtypes, their spatial associations with blood vessel phenotypes were virtually unchanged with postnatal development. Similar to TUBB3+ nerves, NeuF+ , CGRP+ , and TH+ nerves all maintained their association with CD31hiEmcnhi vessels within 10 μm, yet slightly decreased with age (Fig. 6p–r).
The Fgfr2 P253R mutation associated with Apert syndrome demonstrates limited impact on neurovascular architecture
Considering that we found a large density of nerves within the coronal suture regions, and that previous studies have identified correlations between TrkA inhibition and suture fusion,24 we wanted to apply our comprehensive calvarial visualization platform to investigate whether neurovascular distributions were impact by suture-related diseases, such as craniosynostosis. Nearly 100% of all cases of Apert syndrome are caused by one of two FGFR2 mutations and result in coronal suture fusion prior to birth. The Fgfr2+/P253R Apert syndrome mouse is widely validated25,40 and characterized by early mortality with variations in syndrome prevalence. Mice with more moderate phenotypes tend to live up to three weeks, while mice with severe phenotypes live up to only a few days. Thus, for this study, we explored the impact of Apert syndrome on neonatal mice as compared to wild-type (WT) non-mutant litter mates. Calvaria of P0 Fgfr2+/P253R Apert syndrome mice and their unaffected littermates (Fgfr2+/+) were stained with TUBB3, CD31, and Emcn and imaged with our QLSM workflow. Qualitatively, the neurovascular architecture was maintained between Fgfr2+/+ and Fgfr2+/P253R samples (Fig. 7a). A network of vessels was identified throughout for both groups and nerves were clearly identifiable in all regions. Specifically, in the coronal suture, nerves were found traversing the suture region in both groups. When quantified, there were no discernable differences in nerve distributions between Fgfr2+/+ and Fgfr2+/P253R groups (Fig. 7b–d). Overall and regional nerve volumes remained unchanged (Fig. 7b, c), When the fraction and volume density of periosteal nerves and dural nerves was compared between groups, no difference was seen between periosteal or dural volumes (Fig. 7e, f).

Changes in neurovascular distributions with Apert’s syndrome. a MIP images of the parietal bone, coronal suture, and frontal bone regions of P0 WT and Apert mice with TUBB3+ nerves (blue) and vasculature stained with CD31 (green) and Emcn (red). Scale bar is 300 μm. b Overall nerve volume fraction calculation for WT and Apert TUBB3+ nerves. c Regional nerve volume fraction calculations in the parietal bone, coronal suture, and frontal bone regions for WT and Apert mice. d Periosteal and dural nerve fraction calculations for TUBB3+ nerves in WT and Apert mice. e, f Periosteal and dural nerve volume density calculations for TUBB3+ nerves in WT and Apert samples in (e) Overall Calvaria Periosteum, (f) Overall Calvaria Dura Mater. g Total vessel volume fraction calculation for WT and Apert mice. h Phenotypic vessel volume density calculation for CD31hiEmcn-, CD31hiEmcnhi, and CD31loEmcnhi vessels in WT and Apert mice. I Phenotypic vessel phenotype fraction calculation for CD31hiEmcn-, CD31hiEmcnhi, and CD31loEmcnhi vessels in WT and Apert mice. j Spatial association calculations for TUBB3+ nerves and vessels phenotypes. The fraction of nerves within 10 μm is displayed for each vessel phenotype
Similarly, the vasculature of Fgfr2+/P253R Apert syndrome mice was not impacted. Overall vessel volumes were unchanged between groups (Fig. 7G), while no significant differences were observed between vessel phenotypes or spatial association (Fig. 7h–j). While no major differences were found between the neurovasculature of WT and Apert samples, this platform can be applied to additional calvarial-based diseases to further investigate changes in the calvarial microenvironment with disease.
Discussion
Here, we present the first quantitative assessment of nerve subtype distributions and their association with vascular phenotypes in the calvaria during postnatal development. Compared to previous studies that explored neonatal and adult calvaria,21,33 we demonstrated distinct association of sensory and sympathetic nerves with blood vessels in the skulls of 4 weeks and 12 weeks mice. Other than in the transcortical canals, we did not observe nerves in cortical bone. Following postnatal development, CGRP+ and TH+ nerve volume fractions increased with skeletal maturation, suggesting that nerves may further specialize as the skeleton matures. This increase in CGRP+ and TH+ nerves was similar to the increase in the percentage of CGRP+ nerves found in the dorsal root ganglia (DRG) from P12 to adulthood.41 CGRP is known to stimulate osteoblast proliferation and differentiation, while inhibiting osteoclast formation.42,43 Thus, CGRP+ nerves likely play a role in maintaining bone turnover and structure in mature bone. As for TH+ nerves, expression of TH increased in the superior cervical ganglion, olfactory bulb, and brain following postnatal development.44,45,46 The increase in TH+ expression was associated with tissue maturation and was likely similar in the calvarium. Further studies will be required to address calvarium-specific CGRP+ and TH+ neurons that develop with postnatal development, as they may play significant roles in the response to environmental stressors and bone maintenance. Interestingly, we did not find any differences in NeuF+ nerves with postnatal development. Previous studies in the trigeminal ganglia have suggested that large diameter neurons mature prior to small diameter neurons,47 which may explain why the small, unmyelinated CRGP+ and TH+ nerves continue to increase as the skeleton matures, while large, myelinated NeuF+ nerves do not.
Age-related changes in nerves have been characterized in long bones. Chartier et al. reported a decrease in the number of nerves in the periosteum of murine femurs, but no decrease in nerve density, due to the thinning of the periosteum with age.48 Using multivariate analysis, Steverink et al. (2021) found an age-related decrease in nerves in human bones that was most significant in the periosteum.10 Despite differences in embryonic origin and developmental pathways of calvarial and post-cranial bone, the calvaria also exhibited decreased nerve density with aging, specifically in parietal and frontal bone regions, but not in the coronal suture region. Due to the ability of QLSM to provide ‘global’ 3D readouts of the murine calvaria, we could investigate differences throughout the full calvaria and quantitatively characterize specific regions. Thus, we obtained unique insight into age-related nerve loss in the calvaria, which occurred earlier in the frontal bone compared to the parietal bone. Since nerves have been shown to be necessary for the maintenance of bone homeostasis and skeletal stem cell proliferation,24,49 this nerve loss may be associated with age-related bone loss and enhanced bone fragility. In contrast, nerves may also become less necessary for aged bone homeostasis, since Wu et al. (2016) identified a reduced trabecular and cortical thinning after inferior alveolar nerve transection in aged mice as compared to young and middle-aged mice.50 Thus, nerves may not mediate homeostasis to the same extent in aged individuals as compared to highly regenerative young or densely innervated middle-aged individuals. Previous studies have identified nerves in both the periosteum16,33,51,52,53 and dura mater,21,33,51,52 with one study suggesting that there is a higher density of CGRP staining in the dura mater.33 Therefore, we further stratified nerve populations as belonging to the periosteum or dura mater. Conversely, we identified a no differences in CGRP+ nerve populations in the periosteum compared to the dura mater in young mice (4 weeks). Nevertheless, the spatial distribution of nerves in the periosteum and dura mater varied depending on their location (i.e. parietal bone, coronal suture, and frontal bones). This might explain some of the discrepancies with prior findings.
We also identified preferential spatial association of nerves with CD31hiEmcnhi vessels in orthotopic bone which is maintained throughout murine aging. In a previous study by Qin et al., the number of CD31hiEmcnhi vessels was reduced following TrkA inhibition during heterotopic ossification, suggesting that neuron-to-vessel signaling may mediate CD31hiEmcnhi vessel development in ectopic bone formation and may also be important in orthotopic bone development, homeostasis, and healing. In general, it is well known that nerves control vessel patterning, specifically for arteries and arterioles.11,12,13 Further, NGF/TrkA signaling in sensory neurons resulted in VEGF secretion and angiogenesis.54,55 Previous studies have identified a role for TrkA+ nerves in controlling vascularization during development, response to mechanical loading, and bone healing.1,49,52,56 To the best of our knowledge, there have been no studies that have explored the importance of vessel phenotype in these interactions. However, as CD31hiEmcnhi vessels are also positively associated with bone development and healing,22 they are likely the forerunners in neurovascular signaling. Although further studies are necessary to discern the interactions between nerves and CD31hiEmcnhi vessels in the calvarium, this study provides strong justification for the importance of these interactions. Interestingly, although CD31hiEmcnhi blood vessels were reduced in aged mice, their preferential spatial association was maintained throughout all ages studied, albeit at a lower level, emphasizing the importance of this interaction.
In contrast to our findings, a prior study57 was unable to identify a network of blood vessels throughout the full parietal and frontal bones for adult ages (10–14 weeks), whereas we found a dense network of vessels throughout the entire tissue. This discrepancy may be due to their focus on the bone marrow region while excluding the periosteum and dura mater. In fact, we found that CD31hiEmcn- blood vessels increased in density and fraction with age, while CD31hiEmcnhi blood vessels decreased in fraction with age, which was in agreement with previous studies in long bone.6 Loss of CD31hiEmcnhi blood vessels is associated with reduced osteogenic-angiogenic coupling and regenerative capacity. Differences in methodology may account for some of the differences in overall blood vessel volume found between our studies. The Adams et al. study segmented both Emcnhi and Emcnlo blood vessels, whereas we exclusively segmented Emcnhi blood vessels and did not take the CD31loEmcnlo phenotype into account due to its low SNR.
In addition to aging, we also investigated the impact of craniofacial disease associated genetic mutations on neurovascular architecture in a model of Apert syndrome, which demonstrates early coronal suture fusion in utero. In a study of Crouzon syndrome, another FGFR-related craniosynostosis syndrome that demonstrates premature suture fusion, an increase in blood vessel diameter was found in the cranial vault of four human patients.58 In comparison, in a study of TWIST1 mutations, which also demonstrates craniosynostosis, normal arterial development was identified, which more closely aligns with our findings where we found no differences in arteriole-like CD31hiEmcn- blood vessels.59 While Apert syndrome patients demonstrate abnormalities in the central nervous system,60,61 little is known its impact on peripheral nerves in the calvaria, specifically those in the impacted calvaria. In this study, we did not identify an overall change to calvarial nerves with Apert syndrome. In previous studies, TrkA inhibition resulted in premature sagittal suture fusion, suggesting that a loss of nerve signaling may lead to suture closure.24 In comparison, in this study, following coronal suture fusion, nerves were still maintained in all three regions of the calvaria, including the coronal suture. Thus, while TrkA inhibition may lead to suture fusion, suture fusion does not lead to nerve loss within the suture regions.
Given the close spatial association of the skull and brain along with the direct physical connections between the two, understanding nerve distribution and their vascular association within the skull may provide important insights into developmental abnormalities, age-related cognitive disorders, migraines, or simply, regeneration of injuries. Developing methods that impact the neurovascular architecture can be informative in the design of therapies that promote bone healing. In this study, we acquired the 3D architecture and distribution of nerves throughout the parietal and frontal bones of C57BL/6 J mice and quantified changes that occurred over their lifespan (Fig. 8). Additionally, we characterized vascular changes throughout the lifespan and the spatial associations of calvarial nerves with specific vascular phenotypes. Notably, we identified varying levels of age-related nerve growth or loss in the parietal and frontal bones, as well as in the periosteum and dura mater. Thus, for studies that target neurovascular infiltration following injury or disease induction, it is important to consider the distribution and densities of nerves and blood vessels in each specific bone or region of interest because they could impact study success and expected outcomes.

Schematic representing the spatial distribution of nerves and vessel phenotypes in the calvarium. Illustrations showing the relative locations of nerves (cyan) and vessel phenotypes (CD31hiEmcn- – Green, CD31hiEmcnhi – Yellow, and CD31loEmcnhi – Red) in the murine calvaria. All nerve subtypes are primarily found in the periosteum and dura mater and preferentially associate with CD31hiEmcnhi blood vessels
Materials and methods
Materials
All antibodies and reagents used in this study can be found in Table S1.
Animal models
All animal experiments were approved by the Johns Hopkins University Institutional Animal Care and Use Committee (Protocol No. MO21M146). Animals were housed and cared for in Johns Hopkins’ Research Animal Resources central housing facilities. Young (4 weeks), adult (12 weeks), and middle aged (40 weeks) mice used in this study were C57BL/6J (The Jackson Laboratory, Stock No: 000664). Aged (80 weeks) mice were a generous gift from Dr. Jeremy Walston. Fgfr2+/P253R Apert syndrome mice and their non-mutant littermates were a generous gift from Dr. Joan Richtsmeier and were generated at Icahn School of Medicine at Mount Sinai and the Pennsylvania State University.25,40 Newborn mice (P0) were euthanized by inhalation anesthetics and fixed in 4% paraformaldehyde. Gestation time was 19.0 ± 0.5 days. Genotyping of tail DNA by PCR was performed to distinguish mutant from non-mutant littermates.25,40 Mouse litters were produced, sacrificed, and processed in compliance with animal welfare guidelines approved by the Icahn School of Medicine at Mount Sinai and the Pennsylvania State University Animal Care and Use Committees.
Murine calvarial harvest
Mice were anesthetized with ketamine (100 mg/kg) and xylazine (20 mg/kg). Following anesthesia, mice were injected subcutaneously with 200 U of heparin prior to perfusion. An incision was first made at the xiphoid process and additional incisions were made to gain access to the rib cage and heart. Following disruption of the pleural cavity, heparinized saline (10 U/mL in 1X PBS) was perfused via a 20 G blunt needle into the heart via the left ventricle. Prior to perfusion, the right atrium was cut open to allow an open circulation. Perfusion was performed at a rate of 10 mL/min. Following perfusion, the skin was removed around the skull and the calvaria was harvested while maintaining the periosteum and the dura mater. Calvaria were fixed in 4% methanol-free paraformaldehyde overnight at 4 °C. Following fixation, calvaria were washed three times for 1 h with PBS at RT.
Whole-mount immunostaining and optical clearing
To stain and image calvaria via QLSM, we used a previously published protocol.34 Briefly, samples were blocked overnight at 4 °C in a blocking buffer containing 10% V/V normal donkey serum and wash buffer (0.1 mol/L Tris, 0.15 mol/L NaCl, 0.05% V/V Tween-20, 20% V/V dimethylsulfoxide, pH 7.5). To enhance signal in the AF555 channel, a biotin blocking kit was used. Calvaria were blocked for 8 h at RT to block endogenous biotin. Primary staining was performed with antibodies for CD31, Emcn, and TUBB3, NeuF, CGRP, or TH for 7 days at 4 °C in blocking buffer. Next, samples were stained with secondary antibodies, either fluorophore- or biotin-conjugated, for 7 days at 4 °C. Then, a streptavidin conjugate was used for 5 days to amplify Emcn signal in the AF555 channel. All antibodies and conjugates were diluted in blocking buffer. Between each staining incubation, samples were washed five times for at least 1 h in wash buffer. Following staining and immediately prior to imaging, samples were cleared in a graded series of 2,2-thiodiethanol (TDE in TBS-Tween; 25%, 50%, 75%, 90%, 100% × 2). Clearing steps were performed for a minimum of 2 h at RT or overnight at 4 °C. Samples were stored in 100% TDE prior to imaging.
Light-sheet imaging
For lightsheet imaging, we used a LaVision Biotec Ultramicroscope II that was aligned for the refractive index of TDE. Calvaria were mounted to a sample mount and immersed in a glass imaging cuvette with 100% TDE. Samples were imaged using three separate tile acquisitions: a 3 × 1 tile with double-sided illumination, a 3 × 2 tile with left-sided illumination, and a 3 × 2 tile with right sided illumination. Tiles were overlapped by 15% to facilitate stitching. The following hardware and settings were used for all scans: ×2.5 zoom with a ×2 dipping cap (×5 magnification, 1.3 μm x–y pixel size), 5.5 Megapixel sCMOS camera, 20 ms exposure time, 0.154 numerical aperture, and 2.5 μm z step size. Channels were imaged using 561, 640, and 785 nm lasers and 620/60, 680/30, and 845/55 filters, respectively. Laser intensities were set for each antibody and held consistent for each scan.
Image processing and analysis
All image processing and analysis was performed with Imaris® 9.10, Ilastik®,35 and ImageJ36 software and a Dell Precision 7820 Tower workstation. The workstation was equipped with a dual Intel Xeon Gold 6240 processor, 384 GB DDR4 SDRAM (2666 MHz speed), 512 GB and 1 TB SATA SSDs, NVIDIA Quadro RTX5000 graphics card (16 GB GDDR6 memory), and Windows 10 Pro for Workstations.
Imaris-based image conversion and stitching
To process the raw data for Imaris®, we converted LaVision Biotec raw OME-TIFF files to the Imaris® file format (.ims) using the Imaris® File Converter 9.8. Tiles were manually aligned and stitched using the Imaris® Stitcher 9.8, resulting in a final 3D image. Three pre-defined volumes-of-interest (VOI) were positioned for each data set in the parietal bone, coronal suture, and frontal bone regions. VOI dimensions were consistent between data sets. VOI were cropped and exported into ImageJ for channel separation.
Ilastik®-based image segmentation
We segmented nerves in all images using Ilastik®, a freely available machine-learning based image segmentation tool.35 Briefly, once channels were separated in ImageJ, one z-slice was selected from each region and each condition for Ilastik® training. All features were selected for Ilastik® training to account for changes in intensity, shape, and texture. Individual z-slices were trained for nerve and background signals and prediction masks exported for each region. The remaining z-slices for each condition were segmented using batch segmentation and the corresponding prediction masks. Following Ilastik® segmentation, binary segmentation images were generated in ImageJ. Next, z-stacks are imported back into Imaris® for subsequent analysis.
Imaris®-based 3D nerve analysis
Nerve channels (stained with TUBB3, NeuF, CGRP, or TH) were segmented using absolute intensity surface segmentation and a 104 μm3 volume filter to eliminate subcellular-sized segments. Blood vessels (stained with CD31 or Emcn) were segmented using surface segmentation with a 10 μm radius for background subtraction and a 104 μm3 volume filter to eliminate subcellular-sized segments. Thresholds were optimized for each stain and kept consistent between sample ages. Volume statistics for nerves and CD31 and Emcn individual stains were exported from this initial segmentation. Volume density measurements were calculated from the nerve or blood vessel volumes and normalized by the region size.
Next, images were down sampled by a factor of two in each dimension. Down sampling allowed further processing of the initial segmentation to identify vascular phenotypes and nerve localization to the periosteum and dura mater. Binary masks for all stains were generated. A second segmentation (referred to as the down-sampled segmentation) was performed on these masks which included the Imaris® “Split Objects” Surfaces function with a 10 μm seeding point diameter. CD31hiEmcn− and CD31hiEmcnhi vessels were segmented using the CD31 mask and filtered based upon the absence or presence of masked Emcn signal within each object, respectively. CD31loEmcnhi vessels were segmented based upon the Emcn mask and filtered to remove objects co-localized with masked CD31 signal. Vessel volume fraction measurements were calculated from each phenotype vessel volume and normalized by the overall vessel volume in each sample.
To quantify nerve segments in the periosteum and dura mater, the nerve down-sampled segmentation was further classified. VOIs were analyzed individually, and a training set was selected from the points within each VOI. Initial training data was selected, and iterations were made to ensure an accurate classification. Periosteal and dural nerve fractions were calculated from the nerve section count in either the periosteum or dura mater and normalized by the total nerve count in that region. Spatial association statistics were generated for each vessel phenotype by exporting the shortest distance metrics from the nerve segmentation.
Statistics
GraphPad Prism was used to perform all statistics. Either a two-tailed t-test or a one-way or two-way ANOVA with post-hoc Tukey HSD test was performed. P-values < 0.05 were considered significant.
References
Tomlinson, R. E. et al. NGF-TrkA Signaling by Sensory Nerves Coordinates the Vascularization and Ossification of Developing Endochondral Bone. Cell Rep. 16, 2723–2735 (2016).
Farr, J. N. & Khosla, S. Cellular senescence in bone. Bone 121, 121–133 (2019).
Kim, H.-N. et al. DNA damage and senescence in osteoprogenitors expressing Osx1 may cause their decrease with age. Aging Cell 16, 693–703 (2017).
Halloran, B. P. et al. Changes in bone structure and mass with advancing age in the male C57BL/6J mouse. J. Bone Miner. Res. 17, 1044–1050 (2002).
Pappert, M., Khosla, S. & Doolittle, M. Influences of aged bone marrow macrophages on skeletal health and senescence. Curr. Osteoporos. Rep. 21, 771–778 (2023).
Stucker, S., Chen, J., Watt, F. E. & Kusumbe, A. P. Bone angiogenesis and vascular niche remodeling in stress, aging, and diseases. Front. Cell Dev. Biol. 8, 602269 (2020).
Jimenez-Andrade, J. M. et al. A phenotypically restricted set of primary afferent nerve fibers innervate the bone versus skin: therapeutic opportunity for treating skeletal pain. Bone 46, 306–313 (2010).
Mach, D. B. et al. Origins of skeletal pain: sensory and sympathetic innervation of the mouse femur. Neuroscience 113, 155–166 (2002).
Martin, C. D., Jimenez-Andrade, J. M., Ghilardi, J. R. & Mantyh, P. W. Organization of a unique net-like meshwork of CGRP+ sensory fibers in the mouse periosteum: implications for the generation and maintenance of bone fracture pain. Neurosci. Lett. 427, 148–152 (2007).
Steverink, J. G. et al. Sensory innervation of human bone: an immunohistochemical study to further understand bone pain. J. Pain. 22, 1385–1395 (2021).
Martin, P. & Lewis, J. Origins of the neurovascular bundle: interactions between developing nerves and blood vessels in embryonic chick skin. Int. J. Dev. Biol. 33, 379–387 (1989).
Mukouyama, Y., Shin, D., Britsch, S., Taniguchi, M. & Anderson, D. J. Sensory nerves determine the pattern of arterial differentiation and blood vessel branching in the skin. Cell 109, 693–705 (2002).
Li, W. et al. Peripheral nerve-derived CXCL12 and VEGF-A regulate the patterning of arterial vessel branching in developing limb skin. Dev. Cell 24, 359–371 (2013).
Bjurholm, A., Kreicbergs, A., Terenius, L., Goldstein, M. & Schultzberg, M. Neuropeptide Y-, tyrosine hydroxylase- and vasoactive intestinal polypeptide-immunoreactive nerves in bone and surrounding tissues. J. Auton. Nerv. Syst. 25, 119–125 (1988).
Bjurholm, A., Kreicbergs, A., Brodin, E. & Schultzberg, M. Substance P- and CGRP-immunoreactive nerves in bone. Peptides 9, 165–171 (1988).
Hill, E. L. & Elde, R. Distribution of CGRP-, VIP-, D beta H-, SP-, and NPY-immunoreactive nerves in the periosteum of the rat. Cell Tissue Res 264, 469–480 (1991).
Asmus, S. E., Parsons, S. & Landis, S. C. Developmental changes in the transmitter properties of sympathetic neurons that innervate the periosteum. J. Neurosci. 20, 1495–1504 (2000).
Coutu, D. L., Kokkaliaris, K. D., Kunz, L. & Schroeder, T. Three-dimensional map of nonhematopoietic bone and bone-marrow cells and molecules. Nat. Biotechnol. 35, 1202–1210 (2017).
Sayilekshmy, M. et al. Innervation is higher above Bone Remodeling Surfaces and in Cortical Pores in Human Bone: Lessons from patients with primary hyperparathyroidism. Sci. Rep. 9, 5361 (2019).
Li, J., Ahmad, T., Spetea, M., Ahmed, M. & Kreicbergs, A. Bone reinnervation after fracture: a study in the rat. J. Bone Miner. Res. 16, 1505–1510 (2001).
Alberius, P. & Skagerberg, G. Adrenergic innervation of the calvarium of the neonatal rat. Its relationship to the sagittal suture and developing parietal bones. Anat. Embryol. 182, 493–498 (1990).
Kusumbe, A. P., Ramasamy, S. K. & Adams, R. H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 507, 323–328 (2014).
Ramasamy, S. K., Kusumbe, A. P., Wang, L. & Adams, R. H. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone. Nature 507, 376–380 (2014).
Tower, R. J. et al. Spatial transcriptomics reveals a role for sensory nerves in preserving cranial suture patency through modulation of BMP/TGF-β signaling. Proc. Natl. Acad. Sci. USA 118, e2103087118 (2021).
Wang, Y. et al. Abnormalities in cartilage and bone development in the Apert syndrome FGFR2( + /S252W) mouse. Development 132, 3537–3548 (2005).
Aldridge, K. et al. Brain phenotypes in two FGFR2 mouse models for Apert syndrome. Dev. Dyn. 239, 987–997 (2010).
Motch Perrine, S. M. et al. Integration of brain and skull in prenatal mouse models of apert and crouzon syndromes. Front. Hum. Neurosci. 11, 369 (2017).
Holmes, G. et al. Midface and upper airway dysgenesis in FGFR2-related craniosynostosis involves multiple tissue-specific and cell cycle effects. Development 145, dev166488 (2018).
Motch Perrine, S. M. et al. Mandibular dysmorphology due to abnormal embryonic osteogenesis in FGFR2-related craniosynostosis mice. Dis. Model. Mech. 12, dmm038513 (2019).
Motch Perrine, S. M. et al. A dysmorphic mouse model reveals developmental interactions of chondrocranium and dermatocranium. eLife 11, e76653 (2022).
Martínez-Abadías, N. et al. Beyond the closed suture in apert syndrome mouse models: evidence of primary effects of FGFR2 signaling on facial shape at birth. Dev. Dyn. 239, 3058–3071 (2010).
Lorenz, M. R., Brazill, J. M., Beeve, A. T., Shen, I. & Scheller, E. L. A neuroskeletal atlas: spatial mapping and contextualization of axon subtypes innervating the long bones of C3H and B6 mice. J. Bone Miner. Res. 36, 1012–1025 (2021).
Kosaras, B., Jakubowski, M., Kainz, V. & Burstein, R. Sensory innervation of the calvarial bones of the mouse. J. Comp. Neurol. 515, 331–348 (2009).
Rindone, A. N. et al. Quantitative 3D imaging of the cranial microvascular environment at single-cell resolution. Nat. Commun. 12, 6219 (2021).
Berg, S. et al. ilastik: interactive machine learning for (bio)image analysis. Nat. Methods 16, 1226–1232 (2019).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Little, G. J. & Heath, J. W. Morphometric analysis of axons myelinated during adult life in the mouse superior cervical ganglion. J. Anat. 184, 387–398 (1994).
Topley, M. et al. Evaluation of motor and sensory neuron populations in a mouse median nerve injury model. J. Neurosci. Methods 396, 109937 (2023).
Dutta, S. & Sengupta, P. Men and mice: Relating their ages. Life Sci. 152, 244–248 (2016).
Wang, Y. et al. Activation of p38 MAPK pathway in the skull abnormalities of Apert syndrome Fgfr2(+P253R) mice. BMC Dev. Biol. 10, 22 (2010).
Staaf, S., Franck, M. C. M., Marmigère, F., Mattsson, J. P. & Ernfors, P. Dynamic expression of the TRPM subgroup of ion channels in developing mouse sensory neurons. Gene Expr. Patterns 10, 65–74 (2010).
He, H. et al. CGRP may regulate bone metabolism through stimulating osteoblast differentiation and inhibiting osteoclast formation. Mol. Med. Report. 13, 3977–3984 (2016).
Wee, N. K. Y. et al. Inhibition of CGRP signaling impairs fracture healing in mice. J. Orthop. Res. 41, 1228–1239 (2023).
McLean, J. H. & Shipley, M. T. Postmitotic, postmigrational expression of tyrosine hydroxylase in olfactory bulb dopaminergic neurons. J. Neurosci. 8, 3658–3669 (1988).
Willing, J., Cortes, L. R., Brodsky, J. M., Kim, T. & Juraska, J. M. Innervation of the medial prefrontal cortex by tyrosine hydroxylase immunoreactive fibers during adolescence in male and female rats. Dev. Psychobiol. 59, 583–589 (2017).
Kedzierski, W. & Porter, J. C. Quantitative study of tyrosine hydroxylase mRNA in catecholaminergic neurons and adrenals during development and aging. Brain Res. Mol. Brain Res. 7, 45–51 (1990).
Sharma, N. et al. The emergence of transcriptional identity in somatosensory neurons. Nature 577, 392–398 (2020).
Chartier, S. R., Mitchell, S. A. T., Majuta, L. A. & Mantyh, P. W. The changing sensory and sympathetic innervation of the young, adult and aging mouse femur. Neuroscience 387, 178–190 (2018).
Tomlinson, R. E. et al. NGF-TrkA signaling in sensory nerves is required for skeletal adaptation to mechanical loads in mice. Proc. Natl Acad. Sci. USA 114, E3632–E3641 (2017).
Wu, Q., Yang, B., Cao, C., Guang, M. & Gong, P. Age-dependent impact of inferior alveolar nerve transection on mandibular bone metabolism and the underlying mechanisms. J. Mol. Histol. 47, 579–586 (2016).
Zhao, J. & Levy, D. The sensory innervation of the calvarial periosteum is nociceptive and contributes to headache-like behavior. Pain 155, 1392–1400 (2014).
Meyers, C. A. et al. A neurotrophic mechanism directs sensory nerve transit in cranial bone. Cell Rep. 31, 107696 (2020).
Fujita, S. et al. Quantitative analysis of sympathetic and nociceptive innervation across bone marrow regions in mice. Exp. Hematol. 112–113, 44–59.e6 (2022).
Cantarella, G. et al. Nerve growth factor-endothelial cell interaction leads to angiogenesis in vitro and in vivo. FASEB J. 16, 1307–1309 (2002).
Nakamura, K., Tan, F., Li, Z. & Thiele, C. J. NGF activation of TrkA induces vascular endothelial growth factor expression via induction of hypoxia-inducible factor-1α. Mol. Cell. Neurosci. 46, 498–506 (2011).
Li, Z. et al. Fracture repair requires TrkA signaling by skeletal sensory nerves. J. Clin. Invest. 129, 5137–5150 (2019).
Koh, B. I. et al. Adult skull bone marrow is an expanding and resilient haematopoietic reservoir. Nature 8041, 172–181 (2024).
Tholpady, S. S. et al. Aberrant bony vasculature associated with activating fibroblast growth factor receptor mutations accompanying Crouzon syndrome. J. Craniofac. Surg. 15, 431–435 (2004).
Tischfield, M. A. et al. Cerebral Vein Malformations Result from Loss of Twist1 Expression and BMP Signaling from Skull Progenitor Cells and Dura. Dev. Cell 42, 445–461.e5 (2017).
Breik, O. et al. Central nervous system and cervical spine abnormalities in Apert syndrome. Childs Nerv. Syst. 32, 833–838 (2016).
Yeh, E. et al. Novel molecular pathways elicited by mutant FGFR2 may account for brain abnormalities in Apert syndrome. PLoS ONE 8, e60439 (2013).
Acknowledgements
This work was supported by funding from NIDCR (1R01DE027957), Maryland Stem Cell Research Fund (2022-MSCRFV-5782), the NSF GRFP and NCI (5R01CA237597-05, 2R01CA196701-06A1). Lightsheet imaging was performed at JHU’s Integrated Imaging Center. We thank Joan Richtsmeier for comments on the manuscript. We also thank Eileen Su and Shawna Snyder for their help with the illustration in Fig. 8.
Author information
Authors and Affiliations
Contributions
A.L.H. and W.L.G. conceived the study. A.L.H. performed all experiments, imaging, and image analysis. Y.R., A.N.R., and A.P. helped design the staining and image analysis protocols. A.L.H. wrote the manuscript. All authors reviewed the manuscript and discussed the work.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Horenberg, A.L., Ren, Y., Zeng, E.Z. et al. 3D imaging reveals changes in the neurovascular architecture of the murine calvarium with aging. Bone Res 13, 24 (2025). https://doi.org/10.1038/s41413-025-00401-8
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41413-025-00401-8
