
Abstract
RNA-binding proteins (RBPs) regulate diverse post-transcriptional processes and play roles in adipocyte development; however, their role in white fat beiging remains unclear. Here we identify CUG-BP Elav-like family member 1 (CELF1) as a key RBP promoting beiging of inguinal white adipose tissue in response to cold. Adipocyte-specific Celf1 deficiency impairs cold-induced thermogenic gene expression and reduces energy expenditure. Mechanistically, CELF1 binds to the 3′UTR of Dio2 mRNA and enhances its stability, promoting local triiodothyronine (T3) production. Notably, CELF1 expression is significantly reduced in subcutaneous fat of individuals with obesity and negatively correlates with BMI. CELF1 enhances isoproterenol-induced beige adipocyte activation and mitochondrial respiration in vitro, and Celf1 overexpression ameliorates diet-induced obesity and metabolic dysfunction. Hence, our study identifies CELF1 as a physiological regulator of metabolic stress in activating thermogenesis and promoting energy expenditure at the post-transcriptional level, highlighting its potential as a therapeutic target for obesity and metabolic diseases.
Similar content being viewed by others

Introduction
Obesity has become a global health concern and is a significant risk factor for many metabolic diseases, including type 2 diabetes, non-alcoholic fatty liver, hypertension, and cardiovascular diseases1,2. It occurs when energy intake exceeds energy expenditure. Most current anti-obesity medications target central appetite pathways to reduce food intake and food reward, but often cause severe side effects3. Therefore, there is an urgent need to develop new therapeutic strategies to combat obesity. Adipose tissue is essential for systemic energy homeostasis and comprises functionally distinct depots, including inguinal white adipose tissue (iWAT), epididymal white adipose tissue (eWAT), and brown adipose tissue (BAT). Clinical studies have shown the presence of beige-like brown adipose tissue in the adult supraclavicular area, which correlates positively with weight loss and improved metabolic parameters4. Brown and beige adipocytes are thermogenic cells, characterized by their high mitochondrial content and the ability to uncouple heat production from ATP synthesis through uncoupling protein 1 (UCP1)5. Different from brown adipocytes, beige adipocytes are induced in iWAT by cold stress and physical exercise, in a process known as “browning” or “beiging”6,7. Given the low amount of BAT in adults and the extensive presence of white adipose tissue (WAT) throughout the body, increasing energy expenditure by inducing WAT beiging represents a promising approach to addressing obesity and metabolic disorders8. In the aspect of beige adipocyte formation, many studies have focused on the transcriptional and epigenetic levels, however, the post-transcriptional regulation of this process remains largely unknown9.
Post-transcriptional regulation is essential for maintaining cellular metabolism, as it coordinates maturation, transport, stability, and degradation of all classes of RNAs10. RBPs comprise a large group that play a central role in post-transcriptional regulation by orchestrating the production, maturation, localization, translation, and degradation of mRNAs, determining RNA fate from synthesis to decay11. Recently, it has been reported that RBPs are involved in the regulation of brown adipocyte development, thermogenesis, and white fat beiging12,13,14,15,16,17. However, the underlying mechanism remains largely unknown. CELF1, also named CUG-binding protein 1 (CUGBP1), modulates RNA processes, including alternative splicing, deadenylation, translation, and RNA stability18. CELF1 is ubiquitously expressed and plays a pivotal role in various physiological and pathological processes18,19,20,21. Accumulating evidence shows that CELF1 plays a key role in regulating adipose tissue biology function. For example, genetic variation at the CELF1 locus has been associated with obesity, and CELF1 contributes to impairing adipogenesis during aging by regulating the translation of C/EBPβ mRNA22,23. However, whether CELF1 directly regulates WAT beiging remains unknown.
The sympathetic nervous system (SNS) plays a central role in activating thermogenesis and promoting energy expenditure in the form of heat24,25. Upon exposure to cold or β3-adrenergic stimulation, the SNS releases norepinephrine, which binds to β-adrenergic receptors on adipocytes, leading to the upregulation of deiodinase 2 (DIO2)5. DIO2 is a thioredoxin-fold-containing selenoenzyme that converts the inactive form of thyroid hormone, thyroxine (T4), into the active form, triiodothyronine (T3)26. Deficiency of DIO2 impairs BAT adaptive thermogenesis and leads to cold intolerance27. In adipocytes, local T3 maintains adipocyte adrenergic responsiveness and directly enhances Ucp1 expression through its nuclear receptors (TR), contributing to fat thermogenesis28,29. Despite these critical roles, the post-transcriptional regulation of DIO2 in adipose tissue thermogenesis remains unclear.
In this study, we identify the RNA-binding protein CELF1 as a positive regulator of beige adipocyte formation in iWAT. Using both loss- and gain-of-function approaches, we demonstrate that CELF1 stabilizes the mRNA of the key thermogenic regulator Dio2 by binding to its 3′untranslated region (UTR), thereby enhancing local T3 production. Consistently, CELF1 expression is reduced in individuals with obesity compared to lean counterparts, and there is a positive correlation between CELF1 and DIO2 in human subcutaneous adipose tissue. Adipocyte-specific Celf1 deficiency exacerbates diet-induced obesity and metabolic dysfunction, accompanied by decreased energy expenditure, while local overexpression of CELF1 in iWAT resists obesity and improves metabolic disorders by promoting cold-induced beiging. Thus, our findings indicate that CELF1 orchestrates the thermogenic remodeling of iWAT at the post-transcriptional level and may serve as a potential target against obesity and related metabolic diseases.
Results
The expression of CELF1 is upregulated in iWAT in response to thermogenic activation
To identify RBPs functionally important for the beiging of iWAT, we screened the expression of 1912 canonical RBPs annotated in the RBP database following cold stimulation in iWAT30,31,32. Interestingly, we found that CELF1 responded to cold-induced adipose tissue remodeling (Fig. 1a). Further analysis of public databases31,32 showed that Celf1 mRNA expression in iWAT was upregulated upon cold exposure compared to room temperature (RT) or thermoneutrality, and positively correlated with Ucp1, a master regulator of adaptive thermogenesis (Fig. 1b–d). To confirm these results, we housed male and female mice at RT or exposed them to 4 °C for 24 h, and observed significant elevations in both Celf1 mRNA and protein levels in iWAT following cold stimulation (Fig. 1e, f, Supplementary Fig. 1a, b). Similar results were obtained in mice treated with CL316,243 (CL; a β3-adrenergic agonist for pharmacological activation of fat thermogenesis) (Fig. 1g, h). Moreover, the magnitude of CELF1 upregulation in iWAT was comparable between 24 h and 7 days of cold exposure, suggesting that CELF1 plays an important role in both acute and prolonged cold exposure (Fig. 1i, j). These data reveal that the expression of CELF1 is increased in iWAT during thermogenic activation, suggesting that CELF1 may play a direct role in promoting iWAT beiging.

a Venn analysis of overlapped genes between those significantly upregulated in response to cold stimulation from GSE148361 and GSE110420 (p < 0.01) and canonical RNA-binding proteins (RBPs) annotated in the RBP database. b Celf1 and Ucp1 expression in iWAT from wild-type (WT) mice exposed at 4 °C or RT for 4 days by microarray analysis from GSE148361 (n = 4 mice in each group). Celf1 (c) and Ucp1 expression (d) in iWAT from WT mice exposed at 5 °C (n = 6) or thermoneutrality (28 °C; n = 5) for 10 days by microarray analysis from GSE110420. e, f WT male mice housed at RT or 4 °C for 24 h. e qPCR analysis of Celf1 and Ucp1 mRNA levels in iWAT (n = 12 mice for RT group, n = 11 mice for 4 °C 24 h group). f Immunoblot analysis of the indicated protein in iWAT of mice (n = 4). g, h WT mice were injected with CL316,243 intraperitoneally twice, once per 12 h. g qPCR analysis of Celf1 and Ucp1 mRNA levels in iWAT (n = 7 biological replicates). h Immunoblot analysis of the indicated protein in iWAT (n = 4 biological replicates). i, j WT mice were housed at RT or 4 °C for 24 h or 7 days. i qPCR analysis of Celf1 and Ucp1 mRNA levels in iWAT of mice (n = 3 for RT and 4 °C 24 h group, n = 4 for 4 °C 7 d group). j Immunoblot analysis of the indicated protein in iWAT of mice (n = 3). Data are presented as mean ± SEM, statistical significance was determined by two-tailed unpaired Student’s t-test (b–e, g) or one-way ANOVA (i). Source data is provided as a Source Data file.
CELF1 is required for the beiging of iWAT
To determine the function of CELF1 in white fat beiging in vivo, we first knocked down Celf1 by injecting adenovirus carrying Celf1 shRNA in one side of iWAT, with adenovirus expressing LacZ shRNA injected into the contralateral site as a control33. As expected, following acute cold exposure, Celf1 knockdown in iWAT significantly reduced the expression of UCP1 and other key thermogenic genes as compared with the contralateral control (Supplementary Fig. 2a–e). In contrast, overexpression of Celf1 in iWAT increased the expression of thermogenic genes Ucp1 and Pgc1α (Supplementary Fig. 2f). The advantage of unilateral injection is that gene-targeting and control groups share the same biological background, minimizing variation between individuals. However, this model does not assess the effects of Celf1 knockdown on whole-body thermogenic activation. To explore the systemic effects of CELF1-regulated beiging on body temperature, we bilaterally injected shLacZ or shCelf1 adenovirus into the iWAT pads of mice. Consistent with the unilateral injection results, the Celf1 knockdown group impaired the ability of mice to maintain body temperature during cold exposure (Supplementary Fig. 2g). Conversely, overexpression of Celf1 in mice bilaterally injected with adenovirus led to higher rectal temperatures compared to controls during cold exposure (Supplementary Fig. 2h). Collectively, these results suggest that CELF1 is essential for iWAT beiging and the maintenance of body temperature during thermogenic challenges.
Adipocyte Celf1 deficiency impairs cold-induced iWAT beiging and energy expenditure
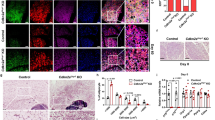
To further investigate the role of CELF1 on iWAT beiging, we generated a Celf1fl/fl mouse model. Since adipose tissue is composed of both stromal vascular fraction (SVF) cells and adipocytes, we examined CELF1 protein levels in these components. Immunoblot analysis showed that CELF1 protein levels were comparable in SVFs and adipocytes at RT (Supplementary Fig. 3a). Notably, the cold-induced increase in CELF1 protein was predominantly observed in adipocytes, but not in SVFs, suggesting that CELF1 primarily regulates white fat beiging in an adipocyte-autonomous manner (Supplementary Fig. 3b, c). To validate this hypothesis in vivo, we generated adipocyte-specific Celf1-knockout mice (Celf1AKO) by crossing Celf1fl/fl mice with Adipoq-Cre mice (Supplementary Fig. 3d), and immunoblot results confirmed selective ablation of CELF1 in adipose tissues (Supplementary Fig. 3e). The Celf1fl/fl and Celf1AKO mice were housed at RT (22° °C) or exposed to 4 °C for 6 h (acute cold exposure). When maintained at RT, Celf1AKO mice exhibited no significant differences compared to Celf1fl/fl mice in rectal temperatures, body weight, fat mass, iWAT weight, adipocyte morphology, thermogenic genes expression, or metabolic rates (Supplementary Fig. 3f–k, Fig. 2d–f, Supplementary Fig. 4c). However, upon cold exposure, Celf1AKO mice displayed apparently reduced iWAT beiging as compared with Celf1fl/fl littermates, evidenced by downregulated expression of thermogenic genes, decreased UCP1 protein level, increased adipocyte size and fat mass (Fig. 2a–c, Supplementary Fig. 4a, b). Furthermore, Celf1 deficiency impaired cold-induced whole-body energy expenditure, including reductions in oxygen consumption rates (OCRs), heat production, and CO2 production rates (Fig. 2d–f, Supplementary Fig. 4c), leading to a compromised ability to maintain body temperature during cold exposure (Fig. 2g). These results indicate that CELF1 plays a critical role in cold-responsive thermogenic remodeling. Consistent with this, RNA analysis of iWAT from cold exposure Celf1fl/fl and Celf1AKO mice showed that the cold-induced thermogenic program in iWAT was severely blunted in Celf1AKO mice. Among them, Ucp1, Dio2, Elvol3, Tbx1, Cox7a1, and Cidea were the most significantly reduced genes in iWAT of Celf1AKO mice after cold stimulation (Fig. 2h). Accordingly, Gene Ontology (GO) biological processes analysis revealed that the downregulated genes were primarily involved in adaptive thermogenesis, temperature homeostasis (response to cold), and mitochondrial-related processes, including lipid oxidation, fatty acid metabolism and so on (Fig. 2i, Supplementary Fig. 4d). KEGG pathway analysis further showed that these downregulated genes were involved in thermogenesis-related pathways, including thermogenesis, PPAR signaling, metabolic pathways, fatty acid degradation, and AMPK signaling (Fig. 2j). Similar results were observed in female mice, manifested by decreased rectal temperature, increased iWAT weight, reduced thermogenic genes expression and energy expenditure (Supplementary Fig. 5a–h). Taken together, these results indicate that adipocyte Celf1 deficiency impairs cold-induced iWAT thermogenesis and energy expenditure.

a–j 8-week-old Celf1fl/fl and Celf1AKO mice were housed at 4 °C for 6 h. a qPCR analysis of the mRNA level of thermogenic genes in iWAT (n = 10 biological replicates). b Immunoblot analysis of the indicated protein in iWAT (n = 3 biological replicates). c Representative images of H&E staining and UCP1 immunohistochemistry (IHC) staining of iWAT, scale bar, 50 μm. VO2 (d–e) and heat production (n = 6 mice for 22 °C group; n = 7 mice group for 4 °C) (f) of Celf1fl/fl and Celf1AKO mice at 22 °C and 4 °C during time (left), and average values (right) (n = 6 mice for 22 °C; n = 7 mice for 4 °C). g Rectal temperatures of Celf1fl/fl (n = 6) and Celf1AKO (n = 5) mice at different time points during acute cold stimulation for 6 h. Heat maps analysis of differentially expressed genes (h), Gene Ontology (GO) (i), and Kyoto Encyclopedia of Genes and Genomes (KEGG) (j) analysis of downregulated genes in iWAT of Celf1AKO mice according to RNA-seq results. Data are presented as mean ± SEM, statistical significance was determined by two-tailed unpaired Student’s t-test (a, e, f, g). Source data is provided as a Source Data file.
CELF1 augments isoproterenol-induced activation of beige adipocytes and mitochondrial respiration in a cell-autonomous manner
Given the complexity of adipose tissue, we sought to determine whether CELF1 promotes adipocyte beiging (activation of beige adipocytes) in a cell-autonomous manner using SVF-differentiated primary adipocytes. To mimic cold exposure, we treated primary adipocytes with isoproterenol and forskolin, both of which are known to induce beige adipocyte activation34. Consistent with our in vivo findings, isoproterenol treatment significantly increased UCP1 expression at both the mRNA and protein levels, alongside increased CELF1 expression (Fig. 3a, b). Similar results were observed in forskolin-treated primary adipocytes (Fig. 3c, d), further suggesting a positive correlation between UCP1 and CELF1 expression in vitro. Next, we explore the role of CELF1 on activation of beige adipocytes by knocking down Celf1 in combination with isoproterenol treatment. Celf1 knockdown reduced the expression of thermogenic genes and UCP1 in primary adipocytes (Fig. 3e, f, Supplementary Fig. 6a, b). Conversely, overexpression of Celf1 increased the thermogenic gene program and UCP1 expression (Fig. 3g, h). Since adipocyte thermogenesis predominantly occurs in mitochondria35, we measured the effects of CELF1 on mitochondrial respiration. OCR analysis revealed that Celf1 knockdown significantly reduced mitochondrial respiration, including basal respiration, isoproterenol-induced maximal respiration, and proton leak respiration in primary adipocytes (Fig. 3i, j). In contrast, Celf1 overexpression significantly enhanced basal respiration, isoproterenol-induced maximal respiration, and proton leak respiration, indicating robust activation of the thermogenic program (Fig. 3k). To assess the function of CELF1 in human adipocytes, we performed small interfering RNA (siRNA)-mediated knockdown of CELF1 in primary adipocytes differentiated from preadipocytes isolated from human subcutaneous WAT. Consistent with observations in mouse adipocytes, CELF1 knockdown markedly reduced the expression of thermogenic genes and UCP1 protein levels in human primary adipocytes. (Supplementary Fig. 6c, d). Collectively, these results suggest that adipocyte CELF1 promotes beige adipocyte activation in a cell-autonomous manner, enhancing both thermogenic gene expression and mitochondrial respiration.

a–d Primary preadipocytes isolated from iWAT of WT mice were differentiated into adipocytes. qPCR (n = 6 for PBS or ISO group; n = 3 for DMSO or FSK group) or immunoblot analysis (n = 3) of CELF1 and UCP1 expression in primary adipocytes treated with PBS or ISO (a, b) and DMSO or FSK (c, d). e–k Primary adipocytes were infected with control shLacZ or shCelf1 adenovirus, control Vector or Celf1-overexpressing (Celf1) adenovirus, and then treated with PBS or ISO for 10 h before harvesting. e qPCR analysis of the mRNA level of thermogenic genes in primary adipocytes infected with shLacZ or shCelf1 adenovirus (n = 4 biological replicates). f Immunoblot analysis of the indicated protein in primary adipocytes infected with shLacZ or shCelf1 adenovirus (n = 3 biological replicates). g qPCR analysis of the mRNA level of thermogenic genes in primary adipocytes infected with Vector or Celf1-overexpressing adenovirus (n = 4 biological replicates). h Immunoblot analysis of the indicated protein in primary adipocytes infected with Vector or Celf1-overexpressing adenovirus (n = 3). i Representative analysis of OCRs of primary adipocytes infected with shLacZ or shCelf1 adenovirus in the left panel; The statistical result of basal respiration is shown in the right panel (n = 4 biological replicates). j Statistical results of maximum respiration and proton leakage respiration of panels (n = 4 biological replicates). k Representative analysis of OCRs of primary adipocytes infected with Vector or Celf1-overexpressing adenovirus in the left panel; The statistical results of basal respiration, maximum respiration, and proton leakage respiration are shown in the right panel (n = 4 biological replicates). Data are presented as mean ± SEM; statistical significance was determined by two-tailed unpaired Student’s t-test (a, c, e, g, i–k). Source data is provided as a Source Data file.
CELF1 binds to mRNA 3′UTR and targets the thyroid hormone signaling pathway
As an RNA-binding protein (RBP), CELF1 likely exerts its regulatory effects on adipocyte beiging through interactions with specific mRNAs. To investigate the underlying mechanism by which CELF1 regulates beiging, we performed RNA immunoprecipitation followed by RNA sequencing (RIP-seq) in differentiated primary adipocytes treated with isoproterenol. First, we immunoprecipitated CELF1 by using an antibody against CELF1 and confirmed the success of CELF1 precipitation (Fig. 4a). Second, we analyzed the RIP-seq data using MACS2 Aligner software36 to identify candidate CELF1-bound mRNAs. Consistent with a previous report20, peak distribution density analysis suggested that the binding potential of CELF1 in 3′ UTR regions was significantly higher than in other genomic regions, such as CDS and 5’ UTR (Fig. 4b). Motif analysis of CELF1-bound mRNA regions using HOMER software37 further indicated that GC-rich and GU-rich motifs were more likely to be bound (Fig. 4c). The targets of CELF1 were involved in many RBP-general biological processes, including regulation of mRNA processing, translation, RNA stability and RNA splicing (Fig. 4d). Notably, these targets were implicated in thermogenesis-related signaling pathways such as thyroid hormone signaling, cAMP signaling, MAPK signaling, and AMPK signaling pathway, as well as metabolism-related pathway (Fig. 4e, Supplementary 7a). These results support that CELF1 may have a vital role in regulating adipocyte thermogenesis.

a Immunoblot analysis of CELF1 protein immunoprecipitated by CELF1 antibody; 10% Immunoprecipitation (IP) cell lysate was used as the input. RNA Immunoprecipitation (RIP) was conducted with differentiated primary adipocytes treated with ISO for 10 h. (Images are representative of three independent experiments) b Gene location distribution of CELF1 target according to the peak density of the RIP-Seq analysis. c Enriched sequences from the CELF1 RIP-Seq data defined by the Homer software (p < 0.05). d GO biological process analysis of CELF1 targets. e KEGG pathway enrichment analysis of CELF1 targets. f Venn analysis of overlap between differentially expressed genes from RNA-seq and enriched genes from RIP-seq. g KEGG pathway enrichment analysis of overlap genes between differentially expressed genes from RNA-seq and enriched genes from RIP-seq. h RIP-qPCR was conducted to verify target genes enriched in the thyroid hormone signaling pathway (n = 3 biological replicates). Data are presented as mean ± SEM; statistical significance was determined by two-tailed unpaired Student’s t-test (h). Source data is provided as a Source Data file.
We further analyzed the RNA-seq data from the iWAT of cold-exposed Celf1fl/fl and Celf1AKO mice and intersected the differentially expressed genes (DEGs) with the CELF1 targets identified in RIP-seq. This analysis revealed 81 overlapping genes that were both bound by CELF1 and altered in response to Celf1 deficiency under cold stimulation (Fig. 4f). Again, KEGG pathway analysis of these overlapping genes revealed enrichment in thermogenesis-related pathways, including thyroid hormone signaling, cAMP signaling, MAPK pathways and AMPK signaling pathway (Fig. 4g). Since the thyroid hormone pathway plays a key role in adipose thermogenesis, we focused on genes related to beiging and thermogenesis within the thyroid hormone signaling pathways (Supplementary Fig. 7b). RIP-qPCR verification showed that CELF1-bound mRNAs were significantly enriched for Med1, Akt2, Atp2a2, Hif1a, Foxo1, Ncoa2, and Dio2 (involved in thyroid hormone signaling) compared to the IgG control (Fig. 4h). Among these, Dio2 was the most enriched CELF1 target (Fig. 4h). Taking together, these data indicate that CELF1 mainly binds to mRNA 3′UTR and regulates thermogenic gene expression through its interaction with thyroid hormone signaling pathways.
CELF1 enhances Dio2 mRNA stability via binding to its 3′UTR
DIO2 is crucial for adaptive thermogenesis in BAT by catalyzing the conversion of T4 to the active thyroid hormone T3, and disruption of thyroid hormone activation in Dio2 knockout mice causes obesity27,38,39. Given that CELF1 can bind to Dio2 mRNA, which contains multiple GU-rich elements in its 3′ UTR (regions known for CELF1 binding), and that Dio2 mRNA expression is significantly altered in response to changes in CELF1 expression (as seen in Figs. 2a, 3e, g, Supplementary Fig. 2e), we hypothesized that Dio2 may serve as a direct target of CELF1 in promoting beiging of iWAT. To test this hypothesis, we detected DIO2 protein in Celf1-deficient tissues and cells. In line with RNA expression data, DIO2 protein levels were significantly reduced in the iWAT of Celf1AKO mice after acute cold exposure, as well as in primary adipocytes with Celf1 knockdown (Fig. 5a, b), suggesting Dio2 expression is responsive to CELF1 regulation. Notably, analysis of public datasets revealed a positive correlation between CELF1 and DIO2 expression in human subcutaneous adipose tissue (Fig. 5c), further supporting the idea that CELF1 influences Dio2 expression and plays a role in adipose tissue beiging. Second, to confirm the interaction between CELF1 and Dio2 mRNA, we performed RIP-qPCR in primary adipocytes and 3T3-L1 adipocytes, revealing a significant enrichment of Dio2 mRNA in the CELF1 immunoprecipitation compared to IgG controls (Fig. 5d, e). Additionally, when Celf1 was knocked down in primary adipocytes, the binding of Dio2 mRNA to CELF1 was reduced (Fig. 5f). To further identify CELF1 binding sites in Dio2 mRNA, we conducted an RNA pull-down assay using in vitro transcribed four sequential RNA fragments of Dio2 3′ UTR, including 4890 nucleotides. CELF1 bound strongly to the first fragment (3′ UTR-1), comprising 1245 nucleotides (Fig. 5g). As Dio2 3′ UTR-1 fragment contains multiple GU-rich sites (shown in “Methods” section), we continued to generate six small RNA segments (200–300 bp) from 3′ UTR-1 fragment for a second round of pull-down assays and found that segments 1.4, 1.5, and 1.6 fragments have strong binding ability to CELF1 (Fig. 5h). Thus, these data support that CELF1 binds to the 3′UTR of Dio2.

Immunoblot analysis (a) of the indicated protein in iWAT of Celf1fl/fl and Celf1AKO mice upon acute cold exposure and in primary adipocytes (b) of Celf1fl/fl mice treated with PBS or ISO after adenovirus infection (n = 3). c Correlation of CELF1 and DIO2 mRNA expression in human subcutaneous adipose tissue from GSE110729 database (n = 28). RIP-qPCR analysis of the Dio2 mRNA level in IgG and CELF1 IP sample in primary adipocytes (d) and in 3T3-L1 (e) treated with ISO (n = 3 biological replicates). f RIP-qPCR analysis of Dio2 mRNA levels in CELF1 IP sample in primary adipocytes of Celf1fl/fl mice treated with ISO after adenovirus infection (n = 3 biological replicates). g, h RNA pull-down assay and immunoblot analysis of Dio2 3’UTR bound to CELF1, β-actin CDS segment was used as a negative control, and 10% 3T3-L1 lysate was used as the input. 3’untranslated region (3’UTR) of Dio2 was divided into four segments (g), Dio2 3’UTR-1 was divided into six segments (h). qPCR analysis of Dio2 mRNA expression in primary adipocytes infected with shLacZ or shCelf1 adenovirus (n = 3 biological replicates) (i), Vector or Celf1-overexpressing adenovirus (j) following labeling with 4-thiouridine (4sU) at the indicated time points (n = 3 biological replicates). k Dual-luciferase reporting assay of Dio2 3’UTR-1 fluorescence intensity in HEK293T infected with adenovirus (n = 6 biological replicates). l, m Immunoblot analysis (l) of CELF1 immunoprecipitated by anti-CELF1 in primary adipocytes with PBS or ISO treatment. RIP-qPCR analysis (m) of Dio2 mRNA retrieved by anti-CELF1 in primary adipocytes with PBS or ISO treatment for 10 h (n = 3 biological replicates). n, o Primary adipocytes were infected with adenovirus, followed by treatment with negative control or siDio2. qPCR (n) and immunoblot analysis (o) of thermogenic genes expression after ISO treatment (n = 3 biological replicates). Data are presented as mean ± SEM; statistical significance was determined by Pearson correlation analysis (c), two-tailed unpaired Student’s t-test (d–f, i, j, m, n), and one-way ANOVA (k). Source data is provided as a Source Data file.
A previous study has reported that CELF1 binding to GU-rich sequences regulates mRNA stability20. Consistent with this, our results showed that Celf1 knockdown significantly reduced Dio2 mRNA levels, whereas Celf1 overexpression increased Dio2 mRNA expression in primary adipocytes (Supplementary Fig. 8a, b). To assess whether CELF1 modulates Dio2 mRNA stability, we performed Roadblock-qPCR analysis, which measures mRNA decay kinetics in living cells via 4-thiouridine (4sU) labeling of nascent transcripts. Results revealed that Celf1 knockdown impaired the stability of Dio2 mRNA in primary adipocytes, while Celf1 overexpression enhanced its stability (Fig. 5i, j). Furthermore, consistent results were obtained using transcriptional blockade with actinomycin D, confirming the regulatory role of CELF1 in Dio2 mRNA stability (Supplementary Fig. 8c, d). To determine whether CELF1 affected its mRNA stability by binding to 3′UTR-1 fragment, we inserted the intact 3′UTR-1 or the truncated 3’UTR-1 mutant without the 1.4–1.6 fragment into the psiCHECK2 plasmid and performed a dual-luciferase reporter assay. Results showed that CELF1 may enhance the expression of Dio2 by indirectly affecting mRNA stability and/or translation (Fig. 5k). Altogether, CELF1 may enhance Dio2 mRNA stability by recognizing and binding to the 3′UTR-1 fragment.
To investigate whether the interaction between CELF1 and Dio2 is enhanced during thermogenic activation, we performed a RIP assay in primary adipocytes treated with either PBS or isoproterenol. Results showed that CELF1 had a stronger binding affinity for Dio2 mRNA in isoproterenol-induced beiging adipocytes compared to control adipocytes (Fig. 5l, m). Notably, this increased binding was attributed to the upregulation of Dio2 mRNA expression rather than a change in the binding affinity of CELF1, since isoproterenol treatment did not affect the enrichment of Dio2 mRNA when primary adipocytes were treated with actinomycin D (Supplementary Fig. 8e). To further clarify the functional relationship between CELF1 and Dio2, we knocked down Dio2 in primary adipocytes overexpressing CELF1, and found that the knockdown of Dio2 significantly diminished the upregulation of thermogenic genes and UCP1 expression mediated by CELF1 overexpression (Fig. 5n, o), suggesting that Dio2 was a key effector promoting iWAT beiging downstream of CELF1. Taken together, these results indicate that thermogenic activation enhances the binding of CELF1 to Dio2 mRNA at the 3′UTR, thereby contributing to the stability and expression of Dio2 during iWAT beiging.
CELF1 enhances the beiging of iWAT through the DIO2-T3 pathway
It is reported that DIO2 plays a key role in catalyzing the conversion of T4 to T3, and that T3 is crucial for the activation of adaptive thermogenesis28,40,41. To test whether CELF1 regulates the beiging of iWAT through DIO2-mediated T3 production, we detected T3 levels in iWAT following cold exposure. Indeed, T3 content in iWAT was significantly decreased in Celf1AKO mice as compared with Celf1fl/fl littermates after 7 days of cold stimulation in PBS-treated group, but not in T3-treated group (Fig. 6a), suggesting that the impaired beiging in Celf1AKO mice may result from the reduction of T3 production. To confirm this possibility, we supplemented T3 in vivo. Results showed that T3 was successfully supplemented as T3 levels were elevated in the T3-injected iWAT relative to the PBS-treated group (Supplementary Fig. 8f). Consistent with acute cold exposure, Celf1AKO mice exhibited lower rectal temperatures after 2 and 7 days of cold exposure compared to Celf1fl/fl littermates; however, T3 supplementation eliminated this temperature disparity (Fig. 6b). Moreover, T3 supplementation improved metabolic features in Celf1AKO mice subject to a 7-day cold exposure, as evidenced by reduced body weight and iWAT mass, along with a decrease in adipocyte size in iWAT by H&E staining (Fig. 6c–e). This metabolic improvement was likely due to enhanced iWAT beiging, as indicated by increased expression of UCP1 and other thermogenic-related genes in T3-supplemented Celf1AKO mice relative to PBS-treated counterparts (Fig. 6e–g). Consistently, T3-supplemented mice showed comparable oxygen consumption and heat production rates between Celf1AKO mice and Celf1fl/fl littermates (Fig. 6h, i). These results suggest that adipocyte-specific Celf1 deficiency reduces T3 content in iWAT and T3 supplementation rescues impaired iWAT beiging in Celf1-deficient mice.

a–i PBS or T3 was injected into Celf1fl/fl and Celf1AKO male mice intraperitoneally once a day for 7 days upon cold exposure. a T3 level in iWAT (n = 7 mice for PBS group; n = 8 mice for T3 group). b Rectal temperatures detection at 2- and 7-day of cold exposure (n = 7 mice for PBS group; n = 8 mice for T3 group). Body weight (c), and iWAT weight (d) of Celf1fl/fl and Celf1AKO mice before and after cold exposure (n = 7 for PBS group, n = 8 for T3 group). e Representative images of H&E staining and UCP1 IHC staining of iWAT. Scale bar, 50 μm. f Immunoblot analysis of UCP1 protein in iWAT (n = 3). g qPCR analysis of thermogenic genes expression in iWAT (n = 7). VO2 (h) and heat production (i) in PBS or T3-treated Celf1fl/fl and Celf1AKO mice housed at 4 °C during one 12-h light-dark cycle (left) and average values in the left panel (n = 6 for PBS group, n = 7 for T3 group). Data are presented as mean ± SEM; statistical significance was determined by two-tailed unpaired Student’s t-test (a–d, g–i). Source data is provided as a Source Data file.
Adipocyte Celf1 depletion exacerbates diet-induced obesity and metabolic dysfunction
The beiging of white adipose tissue (WAT) is closely linked to metabolic health benefits8,42,43,44. Since Celf1 deficiency impaired the beiging of iWAT under cold simulation, we wonder if Celf1 depletion in adipocytes is prone to obesity during high-fat diet (HFD) challenge. To test this, we fed both Celf1AKO mice and Celf1fl/fl littermates with a normal diet (ND) or HFD. Results showed that there were no differences in food intake between Celf1fl/fl and Celf1AKO mice under both ND and HFD feeding (Supplementary Fig 9a). Under ND feeding, Celf1fl/fl and Celf1AKO mice exhibited similar weight gain (Fig. 7a). However, when challenged with HFD, Celf1AKO mice showed a significant increase in body weight, worsened glucose intolerance, and exacerbated insulin resistance as compared with Celf1fl/fl littermates (Fig. 7a, b). Notably, in humans with obesity, CELF1 mRNA levels in subcutaneous fat were significantly lower than in non-obese individuals and were inversely correlated with BMI (Fig. 7c–e). Moreover, HFD-fed Celf1AKO mice exhibited a significant increase in iWAT mass, but not in eWAT (Fig. 7f, Supplementary Fig 9b). Although there were no observable differences in the morphology of iWAT and eWAT between Celf1fl/fl and Celf1AKO mice under the ND situation (Fig. 7g, Supplementary Fig 9c), H&E staining revealed larger adipocytes in the iWAT of HFD-fed Celf1AKO mice than Celf1fl/fl littermates, but not in eWAT (Fig. 7g, h, Supplementary Fig 9c, d). Consistently, HFD-fed Celf1AKO mice exhibited reduced oxygen consumption, lower CO2 generation, and decreased heat production compared to their Celf1fl/fl littermates, with no difference in physical activity (Supplementary Fig 9f–i). These data indicate that adipocyte Celf1 deletion reduces whole-body energy expenditure and exacerbates HFD-induced obesity and metabolic dysfunction.

Body weight curves (a), GTT and ITT (b) of Celf1fl/fl and Celf1AKO mice after ND (n = 5) or HFD (n = 6) feeding. c CELF1 mRNA expression in abdominal subcutaneous adipocytes of subjects from GSE2508 (lean, BMI = 25 ± 3; obesity, BMI = 55 ± 8; n = 9 for men, n = 10 for women). d CELF1 mRNA expression in subcutaneous adipose tissue of subjects from GSE70353 (n = 259 for BMI < 25, n = 511 for BMI > 25). e The correlation between BMI and CELF1 mRNA in subcutaneous adipose tissue of subjects from GSE70353 (n = 770). f iWAT weight of Celf1fl/fl and Celf1AKO mice on ND (n = 5) or HFD (n = 6). Representative images of H&E staining (g) and distribution (left) and quantification (right) of iWAT adipocyte size (h) of Celf1fl/fl and Celf1AKO mice under HFD (n = 6). i–s Celf1fl/fl and Celf1AKO mice were subjected to cold stimulation at 4 °C for 7 days after 12-week HFD feeding. Body weight loss (i) and iWAT weight (j) of HFD-fed Celf1fl/fl and Celf1AKO mice and after cold for 7 days (n = 9). k Representative images of H&E staining of iWAT, scale bar, 50 μm. l Rectal temperatures were monitored upon cold stimulation at the indicated time (n = 9). m qPCR analysis of thermogenic genes expression in iWAT (n = 9 biological replicates). n Immunoblot analysis of UCP1 and DIO2 protein expression in iWAT (n = 3). o T3 concentration in iWAT of HFD-fed Celf1fl/fl and Celf1AKO mice after cold stimulation (n = 10). VO2 (p), VCO2 (q, r), and heat production (s) in HFD-fed Celf1AKO mice and Celf1fl/fl housed at 4 °C during one 12-h light-dark cycle (left) and average values (right) (n = 7). Data are presented as mean ± SEM; significance was assessed by Pearson correlation analysis (e), two-way ANOVA (a, b), and two-tailed unpaired Student’s t-test (c, d, f, h–j, l, m, o, p, r, s). Source data is provided as a Source Data file.
Given that obesity arises from an imbalance between energy intake and expenditure, and that cold-induced beiging of iWAT has emerged as a promising strategy to increase energy expenditure and combat diet-induced obesity and metabolic disorders45,46, we speculated that Celf1 ablation in adipocytes might impair the therapeutic efficacy of cold exposure in obesity. Although cold stimulation effectively reduced body and iWAT weights in HFD challenged Celf1fl/fl mice, which is in line with observations in ND mice, this reduction was significantly attenuated in Celf1AKO mice (Fig. 7i, j). The Celf1AKO mice also exhibited increased adipocyte size in iWAT and worse body temperature maintainability during cold exposure (Fig. 7k, l). Consistently, cold-induced expression of thermogenic genes, including UCP1 and DIO2, was markedly reduced in Celf1AKO mice compared to their Celf1fl/fl littermates, along with downregulated T3 level in iWAT (Fig. 7m–o). Moreover, chronic cold stimulation increased oxygen consumption, carbon dioxide production, and heat generation in HFD-fed Celf1fl/fl mice, but these increases were significantly impaired in Celf1AKO mice (Fig. 7p–s). Collectively, these results indicate that adipocyte-specific Celf1 deficiency exacerbates diet-induced obesity and resists cold-induced iWAT beiging to counteract obesity.
Local overexpression of CELF1 in iWAT enhanced cold-induced metabolic benefits
Finally, we explored whether overexpression of Celf1 in adipose tissue could mitigate diet-induced obesity. To this end, we injected either Vector or Celf1 overexpression adenovirus into the bilateral inguinal fat pads of HFD-fed male mice, followed by one week of cold exposure after 10-week HFD feeding (Supplementary Fig 10a). As expected, Celf1-overexpressing mice exhibited better temperature maintainability and lower body and iWAT weights than the Vector control group during cold exposure (Supplementary Fig 10b–d), suggesting CELF1 overexpression may induce iWAT beiging to combat obesity. Indeed, H&E staining of iWAT revealed that there were more abundant smaller adipocytes in Celf1-overexpressing mice compared to the empty vector control group (Vector), coinciding with higher expression of UCP1, Dio2, and other thermogenic genes, as well as T3 levels in iWAT (Supplementary Fig 10e–g). Moreover, these enhanced iWAT beiging were accompanied by elevated oxygen consumption and heat production in Celf1-overexpressing mice (Supplementary Fig 10h, i). Notably, the reduction of body weight and increase of energy expenditure promoted the improvement of glucose intolerance and insulin resistance (Supplementary Fig 10j, k), indicating the metabolic benefits of Celf1 overexpression.
However, local overexpression of Celf1 using adenoviral vectors lacked adipocyte specificity and may influence other cell types within adipose tissue47. To overcome this limitation, we employed an adeno-associated virus (AAV) vector driven by the Adipoq promoter to achieve adipocyte-specific Celf1 overexpression in HFD-fed mice (Fig. 8a). This targeted approach also induced a pronounced beiging phenotype in iWAT, characterized by elevated rectal temperature, reduced body weight and iWAT mass, decreased adipocyte size, upregulated expression of thermogenic genes and T3 levels, increased oxygen consumption and heat production, and improved metabolic parameters in cold-exposed obese male and female mice (Fig. 8b–k and Supplementary Fig 11a–j). To further evaluate the therapeutic potential of Celf1 against obesity, we delivered the Adipoq-driven Celf1 AAV-vector directly into the iWAT of genetically obese (ob/ob) mice (Fig. 8l). In line with previous findings, Celf1-overexpressing ob/ob mice displayed reduced body weight, elevated rectal temperature, decreased iWAT mass, increased expression of thermogenic genes and T3 level, and improved metabolic outcomes following cold exposure (Fig. 8m–t). Collectively, these results indicate that both local and adipocyte-specific overexpression of Celf1 in iWAT ameliorates HFD-induced obesity and metabolic disorders by enhancing cold-induced beiging and energy expenditure.

a Schematic of the experimental design. b–k Bilateral iWAT of male WT mice fed on high-fat diet for 7 weeks were injected with Adipoq-promoter driven control AAV virus (AAV-Vector) and Celf1-overexpression AAV virus (AAV-Celf1) for four weeks, followed by cold stimulation for one week. b Rectal temperatures of AAV-Vector and AAV-Celf1 mice at different time points during cold stimulation (n = 6). c Body weight of AAV-Vector and AAV-Celf1 mice before and after cold stimulation (n = 6). d iWAT weight of AAV-Vector and AAV-Celf1 mice (n = 6). e Representative images of H&E staining and UCP1 IHC staining of iWAT from AAV-Vector and AAV-Celf1 mice, scale bar, 100 μm. qPCR analysis of thermogenic genes expression (f), T3 levels (g), VO2 (h), and heat production (i) of AAV-Vector and AAV-Celf1 mice housed at 4 °C during one 12-h light-dark cycle (left) and average data (right) (n = 6). GTT (j) and ITT (k) of AAV-Vector and AAV-Celf1 mice (n = 10). l Schematic of the experimental design. m–t Bilateral iWAT of 7-week-old ob/ob male mice was injected with Adipoq-promoter driven Celf1 overexpression and control AAV virus, after 4 weeks, the mice were exposed to cold stimulation at 15 °C for a week (n = 6). Body weight curve (m) and weight loss (n) of ob/ob mice after AAV-Vector and AAV-Celf1 infection. o Rectal temperatures of AAV-Vector and AAV-Celf1 ob/ob mice at different time points during cold stimulation (n = 6). p iWAT weight of AAV-Vector and AAV-Celf1 ob/ob mice (n = 6). q qPCR analysis of thermogenic genes expression in iWAT of AAV-Vector and AAV-Celf1 ob/ob mice (n = 5 biological replicates). T3 concentration in iWAT (r), GTT (s), and ITT (t) of AAV-Vector and AAV-Celf1 ob/ob mice (n = 6). Data are presented as mean ± SEM; statistical significance was determined by two-tailed unpaired Student’s t-test. Source data is provided as a Source Data file.
Discussion
CELF1 has been implicated in various metabolic disorders, including insulin resistance, diabetic cardiomyopathy, and obesity21,22,48,49. However, its role in the beiging of iWAT remained unclear. In this study, we show that the RBP CELF1 is a regulator of iWAT beiging, playing a critical role in maintaining body temperature by activating the DIO2-T3 pathway and resisting obesity through enhanced energy expenditure. Our findings identify CELF1 as a key post-transcriptional regulator of thermogenesis and suggest its potential as a therapeutic target for combating obesity.
RBPs play a central role in post-transcriptional gene regulation, including RNA splicing, localization, stability, and translation50. Emerging evidence highlights the significant role of RBPs in regulating gene expression programs and physiological processes, including the control of adaptive thermogenesis12,14,51. For instance, the deletion of RBP Human antigen R (HuR) impairs acute adaptive thermogenesis by regulating calcium transport and signaling, while RBP YBX2 activates brown adipose tissue by targeting thermogenic genes such as Pgc1α12,51. In this study, we uncovered the critical role of CELF1 in regulating the beiging of iWAT and provided insights into metabolic regulation, thereby offering potential therapeutic targets for metabolic diseases through RBP-mediated post-transcriptional regulation.
CELF1 is a well-characterized RBP that regulates mRNA stability by binding to the 3'UTR of target transcripts18,52. Although previous studies have linked Celf1 genetic variations to obesity, and CELF1 has been shown to impair adipogenesis by regulating the translation of C/EBPβ, the role of CELF1 in iWAT remained undefined22,23,53. Our study demonstrates that CELF1 responds to cold stimulation or β-adrenergic agonists by binding to Dio2 mRNA, thereby stabilizing it and promoting the production of T3, which facilitates the beiging of iWAT. This effect was particularly prominent in iWAT, with no significant changes observed in eWAT. The stimulation of Celf1 expression upon cold exposure appears to be more pronounced at the protein level than at the mRNA level. This suggests that Celf1 may be regulated predominantly at the translational or post-transcriptional level, rather than at the transcriptional level, which warrants further investigation. Furthermore, adipocyte-specific ablation of Celf1 did not affect the expression of thermogenic genes at room temperature but significantly impaired cold-induced iWAT beiging. Different from a previous study, it reports that BAT-enriched long noncoding RNAs 10 (lncBATE10) can compete with Pgc1α mRNA for Celf1 binding, protecting Pgc1α mRNA from CELF1-mediated repression in brown adipocytes and indicating the suppressive role of CELF1 in brown adipocyte adaptive thermogenesis54. This phenotype discrepancy suggests the fine-tuning of CELF1 on target mRNA in different fat depots. While CELF1 is often associated with mRNA destabilization by binding to GU-rich sequences52,55,56,57, our study shows that CELF1 enhances Dio2 mRNA stability, consistent with other reports indicating that CELF1 stabilizes transcripts such as Survivin and DEK in cancer models20,58. This suggests a tissue-selective and context-dependent role for CELF1 in regulating target mRNAs.
T3 regulates Ucp1 transcription via TRβ, while TRα1 maintains adrenergic responsiveness in brown adipocytes29. Our findings support the role of adipocyte CELF1 in activating beige adipocytes through DIO2-mediated T3 production at both cellular and animal levels. These results align with previous studies showing that Dio2 induction in brown adipose tissue (BAT) upon cold exposure increases T3 production, enhancing the sensitivity of BAT to β-adrenergic stimulation28. Although T3 is essential for BAT thermogenesis, there are limited reports on its role in iWAT beiging. In the initial stages of cold exposure, iWAT lacks direct innervation, making it necessary to amplify sympathetic signaling. Previous studies have shown that M2 macrophages in iWAT generate catecholamines to promote the beiging of iWAT59,60. Our work identifies a new mechanism in which CELF1 stabilizes Dio2 mRNA in response to cold, rapidly converting circulating T4 into T3, which amplifies β-adrenergic signaling and enhances cold adaptation.
It’s well known that the activation of the SNS system and T3 production are critical for adaptive energy expenditure40,61. T3 stimulates mitochondrial activity, increasing oxygen consumption and resting energy expenditure29. However, the systemic elevation of T3 has adverse effects, including increased hepatic gluconeogenesis, elevated heart rate, and heightened neuronal sensitivity, which can lead to elevated blood pressure, palpitations, anxiety, and so on62. Therefore, selectively increasing T3 production in iWAT represents a promising strategy to combat obesity without the harmful side effects of systemic T3 elevation. In this study, we overexpressed CELF1 in iWAT, resulting in localized T3 elevation via Dio2 upregulation and cold-induced beiging of iWAT, which improved diet-induced obesity and associated metabolic dysfunction. Localized T3 production via CELF1 overexpression avoided the detrimental effects of systemic T3 elevation on the heart and other organs. Thus, specific overexpression or pharmacological activation of CELF1 in iWAT could provide a therapeutic approach for treating obesity and related metabolic disorders, warranting further investigation.
Our RIP-seq analysis revealed that CELF1 targets are enriched in several key pathways, including cAMP, MAPK, and AMPK signaling pathways. Additionally, some CELF1 targets are located in mitochondria and are involved in oxidative phosphorylation. This suggests that CELF1 may regulate iWAT beiging through other pathways beyond the DIO2-T3 axis, indicating a more complex regulatory network in adaptive thermogenesis that requires further exploration.
In conclusion, our study identifies CELF1 as a regulator that facilitates the beiging of white adipose tissue by stabilizing Dio2 mRNA. This highlights the critical role of post-transcriptional regulation in adipocyte thermogenic remodeling. These findings suggest that adipocyte CELF1 could be a promising target for managing obesity-associated metabolic diseases.
Methods
Ethics statement
Human subcutaneous adipose tissue sampling procedures and operations were approved by the Ethical Committee of Chengdu No. 3 Hospital (Permit Number: No. 20231S-177). Written informed consent was obtained from all participants. All animal studies used the protocol that has been approved by the Institutional Animal Care and Use Commission of Chongqing Medical University (Permit Number: IACUC-CQMU-2023-0156).
Animal studies
All animal experiments were approved by the Institutional Animal Care and Use Committee of Chongqing Medical University. Celf1fl/fl mice were obtained by inserting loxP between intron 5–6 and 8–9 and generated by Biocytogen (Beijing, China). Adipoq-Cre was generated by the Jackson Laboratory and purchased from Cyagen company (Shanghai, China). The Celf1fl/fl mice were bred with Adipoq-Cre mice to obtain Celf1AKO mice. The primers that were used for genotyping are listed in Table S1. All transgenic mice were on a C57BL/6J background, and experiments were conducted with littermate controls. All mice used in vivo experiments were age-matched males or females between 8- and 10 weeks old. Mice were maintained at 22 °C unless otherwise indicated and in specific pathogen-free conditions.
For acute cold exposure experiments, 8–10-week-old male mice were placed in a 4 °C incubator for 6 h. Core body temperature was monitored using the BAT-12 Microprobe Thermometer (Physitemp, USA). For CL treatment, wild-type mice were intraperitoneally (i.p.) injected with 1 mg/kg CL316,243 (MCE, HY-116771A) twice, once every 12 h. Transgenic mice were i.p. injected with CL316,243 (1 mg/kg) daily for three consecutive days. For unilateral adenovirus injection, purified adenoviruses of Celf1 shRNA or shLacZ (control), Vector, or Celf1 were respectively injected into the left and right iWAT of eight-week-old C57BL/6 male mice daily for three consecutive days. For bilateral adenovirus injections under normal diet, purified adenoviruses of Celf1 shRNA or shLacZ (control), Vector or Celf1 were respectively injected into both sides of iWAT daily for three consecutive days. Adenoviruses were injected at 100 µL per inguinal adipose depot in a multiposition way (109-10 pfu/mL). Animals recovered for 48 h at RT after injection and then were exposed to 4 °C for the indicated time. For the HFD-induced obesity study, 8-week-old male mice were fed ND or HFD (60% kcal fat, Research Diets, D12492) for 10 or 12 weeks. For bilateral adenovirus injection in HFD-fed mice, the Vector or Celf1-overexpressing adenovirus was injected into iWAT every 3 days, and one week after injection, the mice were exposed to 4 °C for 7 days, during which time adenovirus was also injected every 3 days. For bilateral AAV injection in HFD-fed and ob/ob mice, 2.0 × 1011 vg of AAV8-adipoq Celf1 overexpression virus or control AAV8-adipoq empty vector virus (Shanghai Obio Technology) was injected into iWAT at multiple sites14,63,64. The body weights of mice were recorded weekly. After 4 weeks, the mice were exposed to 4 °C for one week. For the T3 supplementation assay, the Celf1fl/fl and Celf1AKO mice were intraperitoneally injected with 200 µg/kg T3 (MCE, HY-A0070A) every day for 7 consecutive days upon 4 °C cold stimulation. T3 concentration was detected by an ELISA kit (Abcam, ab285259).
Mice were euthanized by carbon dioxide (CO2) asphyxiation inhalation, and cervical dislocation was performed as a secondary euthanasia procedure; then, the tissues were isolated.
Metabolic parameters detection
The energy consumption and expenditure of mice were measured using an OxyMax Comprehensive Laboratory Animal Monitoring System (Columbus Instruments; CLAMS) via indirect calorimetry measures according to the manufacturer’s protocols. Briefly, mice were singly housed and acclimatized in metabolic chambers for 24 h before data collection. Mice had free access to food and water. Oxygen consumption, carbon dioxide production, physical activity, and food intake were monitored for 24 h, with heat production and RER calculated. Data were calculated by normalization with the body weight of each mouse.
Glucose tolerance test (GTT) and insulin tolerance test (ITT)
For the GTT, mice were i.p. injected with D-glucose (2 mg/g body weight, Sigma–Aldrich) after fasting for 12–14 h. Blood glucose levels were measured at 0, 30, 60, 90, and 120 min using a blood glucose meter and strips after glucose injection.
For the ITT, mice were i.p. injected with human insulin (0.75 mU/g body weight, Novo Nordisk) after fasting for 4 h. Blood glucose levels were measured at 0, 30, 60, and 90 min using a blood glucose meter and strips after insulin injection.
Primary preadipocyte isolation and in vitro differentiation
For isolation of mice SVF cells, iWAT from 6–8-week-old mice was pooled together, minced, and digested in 0.75% collagenase (Sigma–Aldrich, C2139) and 0.6 U/mL dispase II (Sigma–Aldrich, D4693) for 30–40 min. Tissue homogenates were filtered through a 100 µm cell strainer (Corning, CLS431752) and followed by centrifugation at 1500 rpm for 5 min, then the upper liquid was discarded to collect SVF cells at the bottom. SVF cells were washed with DMEM/F12 (Gibco, 11330032), followed by centrifuging, and then resuspended in the basic media containing DMEM/F12 supplemented with 10% fetal bovine serum (Gibco, A3161002C), 100 U/mL penicillin, and 100 mg/mL streptomycin (Thermo Fisher, 15140122) and cultured at 37 °C in a 5% CO2 humidified incubator. To induce SVFs differentiate to adipocytes, confluent SVF cells (Day 0) were cultured in the induction media containing 0.5 mM 3-isobutyl-1-methylxanthine (Sigma–Aldrich, I5879), 1 µM dexamethasone (Sigma–Aldrich, D4902), 850 nM insulin (Solarbio, I8830) and 1 µM rosiglitazone (Sigma–Aldrich, R2408) for 2 days (Day 2), then switched to the media containing 850 nM insulin, and 1 µM rosiglitazone for 2 days (Day 4), lastly switched to the basic media for 2 days (Day 6). The cells to be infected were incubated with the adenovirus-containing supernatant with 5 µg/mL polybrene (Santa Cruz Biotechnology, SC-134220) for 48 h. To activate thermogenic gene expression, fully differentiated adipocytes (Day 6) were treated with 10 µM isoproterenol (Sigma–Aldrich, I5627) or 10 µM forskolin (Sigma–Aldrich, F6886) for 10 h before the cells were harvested.
For human primary adipocyte differentiation, single-cell suspensions of SVF were prepared from human subcutaneous WAT by enzymatic digestion with 1.5% collagenase (Sigma–Aldrich, C8140). Cells were then cultured to enrich for PDGFRα⁺ adipocyte progenitors65. Preadipocytes were maintained and differentiated with the MSC (Mesenchymal stem cells) adipogenic differentiation medium at 37 °C in a humidified incubator with 5% CO2. The adipogenic induction cocktail used for differentiation was identical to that applied for mouse preadipocytes.
siRNA transfection
Two siRNAs targeting Dio2 were designed and synthesized by Sangon Biotech (Shanghai, China). The sequences of the two siRNAs targeting Dio2 are: GCGCUCUAUGACUCGGUCAUU and ACCUACAAGAAGUCC
GAAGUU. Two siRNAs targeting CELF1 were designed and synthesized by Tsingke Biotech (Shanghai, China). The sequences of the two siRNAs targeting CELF1 are: CCTAGCTCTAGCAGCAGTAAT and CGAGTCATGTTCTCTTCGTTT. siRNAs were transfected into differentiated adipocytes using RNAimax Transfection Reagent (Thermo Fisher, 13778100) according to the manufacturer’s protocol. The gene expression analysis was determined by RT-qPCR after 48 h.
Mitochondrial respiration assays
The OCRs were determined using the XF24 Extracellular Flux Analyzer (Agilent Technologies, USA) and Agilent Seahorse XF24 Flux Assay Kit. Briefly, SVF cells isolated from iWAT were plated in XF24 cell culture microplates and differentiated to adipocytes as described above. On day 5 of differentiation, the cells were infected with adenovirus for 48 h to knock down or overexpress Celf1. Cells were then washed and incubated in a pre-warmed assay medium (XF basal medium supplemented with 1 mM pyruvate, 2 mM glutamine, and 10 mM glucose) at 37 °C in an incubator for 1 h. The drug-injection ports of the sensor cartridges were loaded with the assay reagents. The respiration rates of adenovirus-infected primary adipocytes were measured for three cycles for basal respiration, three cycles for ISO-stimulated respiration, three cycles for ATP-linked respiration with oligomycin (1.5 μM) treatment, three cycles for maximum respiration with FCCP (0.5 μM) treatment, and three cycles for non-mitochondrial respiration with rotenone (0.5 μM) and antimycin-A (0.5 μM) treatment. The OCR values were automatically calculated by the Seahorse XF24 software (Wave, Agilent). Immediately after measurement, total protein levels were measured with the BCA Protein Assay Kit (Beyotime, P0010) for data correction.
Adenovirus construction and purification
The Celf1 knockdown adenovirus was constructed by using a BLOCK-iTTM-U6-Entry Vector (Thermo Fisher, K494500) according to the manufacturer’s instructions. The Celf1 overexpression adenovirus was constructed by using the pHBAD-EF1-MCS-3falg-CMV-EGFP-loxp vector purchased from Haiheng Science and Technology Company (Shanghai, China) according to the manufacturer’s instructions. The inserted oligos sequences of Celf1 shRNA and Primer Sequences for Celf1 overexpression are listed in Table S2. The adenovirus was generated and amplified in HEK293A cells. The collected adenovirus was purified by Adenovirus Purification Kits (Sartorius, VS-AVPQ022) according to the manufacturer’s instructions.
RNA extraction, reverse transcription, and quantitative PCR
Total RNA was extracted from cultured cells or tissues with TRIzolTM Reagent (Thermo Fisher, 15596026), and 1 μg total RNA was reverse transcribed to cDNA using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher, K1622) according to the manufacturer’s instructions. Quantitative Real-time PCR was performed using PowerUp™ SYBR™ Green Mix (Thermo Fisher, A25743) with the ABI Prism 7500 quantitative machine (Applied Biosystems, USA). Relative expression of target genes was calculated using the comparative method by normalization to the internal control 18S. Sequences of primers used for real-time PCR are listed in Table S3.
RNA-Seq analysis
Total RNA was extracted from the iWAT of mice using the TRIzol reagent (Thermo Fisher,15596026) according to the manufacturer’s protocol. RNA was subjected to commercial RNA-seq analysis using the DNBSED platform at the Beijing Genomics Institution. Three independent samples per group were analyzed. Briefly, mRNA was enriched by oligo (dT)-attached magnetic beads, and further subjected to mRNA fragmentation, cDNA synthesis, end repair, and A and adapter ligation. PCR products were heat-denatured, and ssDNA was cyclized using a cyclization reaction system and procedure. The uncyclized linear DNA was digested, and finally, the prepared library was sequenced. The sequencing data was filtered with SOAPnuke, and HISAT was applied to align the clean reads to the mouse genome (GRCm38.p6) with 94.43% alignment. Expression levels of genes were calculated by RSEM. The criteria for considering differentially expressed genes (DEGs) were a log2FoldChange > 0.5 (either direction) and a significant p-value (< 0.05) versus control. The heatmap of DEGs was drawn by the pheatmap package. Gene Ontology enrichment analysis and KEGG pathway enrichment analysis of DEGs were performed using R based on clusterprofiler and ggplot2, respectively. The significant levels of terms and pathways were corrected by p-value with a rigorous threshold (p < 0.05).
Immunoblotting
Cells and tissues were lysed in lysis buffer containing 50 mM Tris-HCl, 2% SDS, 1 mM PMSF (phenyl methane sulfonyl fluoride) (Solarbio, China), and protease inhibitors (Thermo Fisher, A32963). The collected proteins were diluted in loading sample buffer, heated at 100 °C for 10 min, and loaded on a 10–12% polyacrylamide gel. Subsequently, separated proteins were transferred onto PVDF membranes. PVDF membrane blots were blocked in TBST containing 5% skimmed milk for 1 h at RT, and incubated overnight at 4 °C with primary anti-UCP1(Abcam, ab209483), anti-CELF1 (Abcam, ab129115), anti-DIO2 (Invitrogen, PA5-49631), anti-β-actin (CST, 4970S), and anti-HSP90 (CST, 4877S). All primary antibodies are diluted at 1:1000 for use. After 3 washes with TBST, the membrane was incubated with a secondary antibody (Jackson ImmunoResearch). After 3 washes with TBST again, the membranes were incubated with universal and enhanced chemiluminescence (ECL) substrate (Share-bio, China). Bands were visualized and recorded with an Amersham Imager 600 (GE Healthcare Life Sciences).
H&E and immunohistochemistry (IHC)
Adipose tissues collected from 3 individual mice in each group, fixed with 4% formaldehyde and embedded in paraffin, were sectioned into 5 μm slides, followed by deparaffinization, rehydration, and H&E staining. Adipocyte size was analyzed by AdipoCount software. For IHC, antigen retrieval was conducted with Citrate Antigen Retrieval solution at 95 °C for 20 min after rehydration. Tissue sections were incubated with 3% H2O2 for blocking endogenous peroxidases, and incubated with UCP1 antibody (1:500, Abcam, ab209483) at 4 °C overnight. Subsequently, adipocytes with UCP1 antibody labeling were detected by goat anti-rabbit IgG conjugated with HRP, followed by detection using DAB Color Development Kit (ZSGB-BIO, ZLI-9018, China). Images were acquired by using the Olympus imaging system (Olympus).
RIP and RIP-seq
Primary adipocytes and differentiated 3T3-L1 cells were used to RIP using the Magna RIP kit (Merck Millipore,17-701) according to the manufacturer’s instructions. The input RNA sample of isoprenaline-treated primary adipocytes and RNA samples retrieved from anti-CELF1 (Merck Millipore,03-104) with the Magna RIP kit were used for RNA sequencing (Nonogene, China). The quality statistics of raw reads and clean reads after trimming were performed by using FastQC software. The reads were compared to the reference genome using BWA. Peak calling analysis was performed by MACS2 software. Motifs enriched in the peaks were assessed with the Homer software. FoldEnrichment > 1.5 and p < 0.05 were used to screen CELF1 binding targets. Gene Ontology enrichment analysis and KEGG pathway enrichment analysis of CELF1 targets were performed using R based on clusterprofiler and ggplot2, respectively.
The biotinylated RNA-probe preparation and RNA pull-down
To obtain the biotinylated RNA probe, Specific primers containing the T7 promoter were designed to amplify each fragment of Dio2 by PCR and are listed in Table S4. Four sequential RNA fragments of Dio2 3'UTR (3'UTR-1, 3'UTR2, 3'UTR3, 3'UTR4) and the sequences of six sequential RNA fragments of Dio2 3'UTR-1 (3'UTR-1-1, 3'UTR-1-2, 3'UTR-1-3, 3'UTR-1-4, 3'UTR-1-5, 3'UTR-1-6) are listed in Table S5. Subsequently, PCR products were transcribed in vitro into biotin-labeled RNA with TranscriptAid T7 High Yield Transcription Kit (Thermo Fisher, K0441) and Biotin-14-CTP (Thermo Fisher, 19519016). To purify the transcription products, which were extracted by chloroform and isopropyl alcohol in a ratio of 24:1 and precipitated by glycogen and 70% ethanol, were finally dissolved in DEPC (diethylpyrocarbonate)-treated water. For RNA pull-down assay, 3T3-L1 cells were lysed in IP lysis buffer (Beyotime, P0013) supplemented with protease inhibitors and Rnase inhibitors (Thermo Fisher, AM2694). Purified RNA probes were incubated with 100 µL 3T3-L1 cell lysates at 4°C for 3 h, then conjugated to Streptavidin Magnetic Beads (Thermo Fisher, 88817) for 6 h at 4°C. After washing 5 times with IP lysis buffer, RNA-Protein complexes were eluted from beads by heating at 95 °C for 5 min. An immunoblot assay was conducted to detect the level of CELF1 protein.
Measuring mRNA decay with roadblock-qPCR
Primary adipocytes infected with adenovirus were treated with isoproterenol and then incubated with 400 µM 4-thiouridine (4sU, Sigma–Aldrich, T4509), which is readily incorporated into nascent mRNAs during transcription66. Cells were collected at 0, 4, 8, and 12 h after 4sU treatment, and total RNA was extracted using the Eastep® Super Total RNA Extraction Kit (Promega, LS1040). 1 µg of RNA was transferred into a PCR tube and denatured at 65 °C for 5 min. Subsequently, N-ethylmaleimide (NEM, Sigma–Aldrich, 04259) and NEM buffer were added to the RNA, along with 0.2 ng of spike-in mRNA (renilla luciferase mRNA) for normalization. The mixture was then incubated at 42 °C for 1.5 h. NEM-treated RNA was purified using the Zymo RNA Clean and Concentrator Kit (Zymo Research, cat no. R1013), and then reverse transcribed using oligo(dT) primers. The levels of target mRNA (Dio2) and the spike-in reference mRNA were quantified at each time point using qPCR.
Luciferase reporter assay
The Dio2 3’UTR-1 fragment was amplified by PCR and inserted into the 3’ ends (the enzyme cleavage sites of Xhol and NotI) of the psiCHECK2 plasmid using seamless cloning, and then a mutant primer of the 3’UTR-1 fragment was designed for obtaining a truncated 3’UTR-1 fragment (fragment containing only 3’UTR-1-1, 3’UTR-1-2, and 3’UTR-1-3) by PCR. The plasmid template was removed by adding the DpnI enzyme. Then, the plasmids inserted Dio2 3’UTR-1 WT and mutant were transfected into HEK293 T cells, respectively. After 10 h of plasmid transfection, HEK293 T cells were infected with Vector or Celf1-overexpressing adenovirus. Finally, Firefly and Renilla luciferase activities were detected after 24 h of adenovirus treatment. Primers used for amplifying each fragment of Dio2 are listed in Table S6.
Statistical analyses
All data are presented as mean ± SEM. All analyses were performed with GraphPad Prism 08 (GraphPad). Comparisons between two groups were performed using an unpaired two-tailed Student’s t-test. Multiple-group comparisons were performed using one-way ANOVA or two-way ANOVA by Tukey’s multiple comparisons test. A p-value < 0.05 was considered statistically significant. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data supporting the findings described in this manuscript are available in the article and in the Supplementary Information and from the corresponding author upon request. The sequencing data of RNA-seq generated in this study are available in Gene Expression Omnibus under accession code GSE281771. The sequencing data of RIP-seq data generated in this study are available in Gene Expression Omnibus under accession code GSE297248. The thermogenesis-related sequencing data reused in this study are available in Gene Expression Omnibus under accession codes GSE148361, GSE110420, and GSE86338. The human sequencing data of RNA-seq data reused in this study are available in Gene Expression Omnibus under accession codes GSE2508 and GSE70353. Source data are provided with this paper.
Change history
18 September 2025
In the version of Supplementary Information initially published with this article, due to a figure preparation error, in Supplementary Fig. 5e the panel labeled as the Celf1fl/fl female mouse was an inadvertent duplicate of the Celf1fl/fl male mouse in Fig. 2c. The supplementary figure and associated source data have been updated in the HTML version of the article.
References
Hagberg, C. E. & Spalding, K. L. White adipocyte dysfunction and obesity-associated pathologies in humans. Nat. Rev. Mol. Cell Biol. 25, 270–289 (2024).
Bluher, M. Obesity: global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 15, 288–298 (2019).
Perdomo, C. M., Cohen, R. V., Sumithran, P., Clement, K. & Fruhbeck, G. Contemporary medical, device, and surgical therapies for obesity in adults. Lancet 401, 1116–1130 (2023).
Sakers, A., De Siqueira, M. K., Seale, P. & Villanueva, C. J. Adipose-tissue plasticity in health and disease. Cell 185, 419–446 (2022).
Cohen, P. & Kajimura, S. The cellular and functional complexity of thermogenic fat. Nat. Rev. Mol. Cell Biol. 22, 393–409 (2021).
Bartelt, A. & Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 10, 24–36 (2014).
Wang, W. & Seale, P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 17, 691–702 (2016).
Lizcano, F. The beige adipocyte as a therapy for metabolic diseases. Int. J. Mol. Sci. 20, 5058 (2019).
Inagaki, T., Sakai, J. & Kajimura, S. Transcriptional and epigenetic control of brown and beige adipose cell fate and function. Nat. Rev. Mol. Cell Biol. 17, 480–495 (2016).
Gerstberger, S., Hafner, M. & Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet 15, 829–845 (2014).
Kelaini, S., Chan, C., Cornelius, V. A. & Margariti, A. RNA-binding proteins hold key roles in function, dysfunction, and disease. Biology 10, 366 (2021).
Xu, D. et al. RNA binding protein Ybx2 Regulates RNA stability during cold-induced brown fat activation. Diabetes 66, 2987–3000 (2017).
Wu, R. et al. RNA-binding protein YBX1 promotes brown adipogenesis and thermogenesis via PINK1/PRKN-mediated mitophagy. FASEB J. 36, e22219 (2022).
Yan, S. et al. Adipocyte YTH N(6)-methyladenosine RNA-binding protein 1 protects against obesity by promoting white adipose tissue beiging in male mice. Nat. Commun. 14, 1379 (2023).
Chi, Y. L. & Lin, J. C. RBM4a modulates the impact of PRDM16 on development of brown adipocytes through an alternative splicing mechanism. Biochim Biophys. Acta Mol. Cell Res. 1865, 1515–1525 (2018).
Dai, N. et al. IGF2BP2/IMP2-Deficient mice resist obesity through enhanced translation of Ucp1 mRNA and Other mRNAs encoding mitochondrial proteins. Cell Metab. 21, 609–621 (2015).
Chou, C. F. et al. KSRP ablation enhances brown fat gene program in white adipose tissue through reduced miR-150 expression. Diabetes 63, 2949–2961 (2014).
Dasgupta, T. & Ladd, A. N. The importance of CELF control: molecular and biological roles of the CUG-BP, Elav-like family of RNA-binding proteins. Wiley Interdiscip. Rev. RNA 3, 104–121 (2012).
Wu, X. et al. CUG-binding protein 1 regulates HSC activation and liver fibrogenesis. Nat. Commun. 7, 13498 (2016).
Cifdaloz, M. et al. Systems analysis identifies melanoma-enriched pro-oncogenic networks controlled by the RNA binding protein CELF1. Nat. Commun. 8, 2249 (2017).
Zhai, K. et al. RNA-binding protein CUGBP1 regulates insulin secretion via activation of phosphodiesterase 3B in mice. Diabetologia 59, 1959–1967 (2016).
Hinney, A. et al. Genetic variation at the CELF1 (CUGBP, elav-like family member 1 gene) locus is genome-wide associated with Alzheimer’s disease and obesity. Am. J. Med. Genet. B Neuropsychiatr. Genet. 165B, 283–293 (2014).
Karagiannides, I. et al. Increased CUG triplet repeat-binding protein-1 predisposes to impaired adipogenesis with aging. J. Biol. Chem. 281, 23025–23033 (2006).
Mori, S. et al. Sympathetic innervation of the supraclavicular brown adipose tissue: A detailed anatomical study. PLoS One 18, e0290455 (2023).
Cao, Q., Jing, J., Cui, X., Shi, H. & Xue, B. Sympathetic nerve innervation is required for beigeing in white fat. Physiol. Rep. 7, e14031 (2019).
Bianco, A. C. & McAninch, E. A. The role of thyroid hormone and brown adipose tissue in energy homoeostasis. Lancet Diabetes Endocrinol. 1, 250–258 (2013).
de Jesus, L. A. et al. The type 2 iodothyronine deiodinase is essential for adaptive thermogenesis in brown adipose tissue. J. Clin. Investig. 108, 1379–1385 (2001).
Zekri, Y., Flamant, F. & Gauthier, K. Central vs. peripheral action of thyroid hormone in adaptive thermogenesis: a burning topic. Cells 10, 1327 (2021).
Yau, W. W. & Yen, P. M. Thermogenesis in adipose tissue activated by thyroid hormone. Int. J. Mol. Sci. 21, 3020 (2020).
Liao, J. Y. et al. EuRBPDB: a comprehensive resource for annotation, functional and oncological investigation of eukaryotic RNA binding proteins (RBPs). Nucleic Acids Res. 48, D307–D313 (2020).
Rodo, J. et al. Integrated gene expression profiles reveal a transcriptomic network underlying the thermogenic response in adipose tissue. Sci. Rep. 13, 7266 (2023).
Defour, M. et al. The peroxisome proliferator-activated receptor alpha is dispensable for cold-induced adipose tissue browning in mice. Mol. Metab. 10, 39–54 (2018).
Qian, S. W. et al. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc. Natl. Acad. Sci. USA 110, E798–E807 (2013).
Guo, X. et al. cAMP-MicroRNA-203-IFNgamma network regulates subcutaneous white fat browning and glucose tolerance. Mol. Metab. 28, 36–47 (2019).
Bertholet, A. M. & Kirichok, Y. Mitochondrial H(+) leak and thermogenesis. Annu. Rev. Physiol. 84, 381–407 (2022).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Heinz, S. et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 38, 576–589 (2010).
Marsili, A. et al. Mice with a targeted deletion of the type 2 deiodinase are insulin resistant and susceptible to diet induced obesity. PLoS ONE 6, e20832 (2011).
Castillo, M. et al. Disruption of thyroid hormone activation in type 2 deiodinase knockout mice causes obesity with glucose intolerance and liver steatosis only at thermoneutrality. Diabetes 60, 1082–1089 (2011).
Zekri, Y. et al. Brown adipocytes local response to thyroid hormone is required for adaptive thermogenesis in adult male mice. eLife 11, e81996 (2022).
Shu, L. et al. A-FABP mediates adaptive thermogenesis by promoting intracellular activation of thyroid hormones in brown adipocytes. Nat. Commun. 8, 14147 (2017).
Xue, S., Lee, D. & Berry, D. C. Thermogenic adipose tissue in energy regulation and metabolic health. Front Endocrinol.14, 1150059 (2023).
Min, S. Y. et al. Human ‘brite/beige’ adipocytes develop from capillary networks, and their implantation improves metabolic homeostasis in mice. Nat. Med. 22, 312–318 (2016).
Cohen, P. et al. Ablation of PRDM16 and beige adipose causes metabolic dysfunction and a subcutaneous to visceral fat switch. Cell 156, 304–316 (2014).
Hanssen, M. J. et al. Short-term cold acclimation improves insulin sensitivity in patients with type 2 diabetes mellitus. Nat. Med 21, 863–865 (2015).
Yoneshiro, T. et al. Recruited brown adipose tissue as an antiobesity agent in humans. J. Clin. Invest. 123, 3404–3408 (2013).
Meng, M. et al. Zinc finger protein ZNF638 regulates triglyceride metabolism via ANGPTL8 in an estrogen dependent manner. Metab. Clin. Exp. 152, 155784 (2024).
Verma, S. K. et al. Reactivation of fetal splicing programs in diabetic hearts is mediated by protein kinase C signaling. J. Biol. Chem. 288, 35372–35386 (2013).
Savkur, R. S., Philips, A. V. & Cooper, T. A. Aberrant regulation of insulin receptor alternative splicing is associated with insulin resistance in myotonic dystrophy. Nat. Genet. 29, 40–47 (2001).
Glisovic, T., Bachorik, J. L., Yong, J. & Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 582, 1977–1986 (2008).
Anthony, S. R. et al. HuR expression in adipose tissue mediates energy expenditure and acute thermogenesis independent of UCP1 expression. Adipocyte 9, 335–345 (2020).
Vlasova-St Louis, I. & Bohjanen, P. R. Coordinate regulation of mRNA decay networks by GU-rich elements and CELF1. Curr. Opin. Genet. Dev. 21, 444–451 (2011).
Bae, E. J. & Kim, S. G. Enhanced CCAAT/enhancer-binding protein beta-liver-enriched inhibitory protein production by Oltipraz, which accompanies CUG repeat-binding protein-1 (CUGBP1) RNA-binding protein activation, leads to inhibition of preadipocyte differentiation. Mol. Pharm. 68, 660–669 (2005).
Bai, Z. et al. Dynamic transcriptome changes during adipose tissue energy expenditure reveal critical roles for long noncoding RNA regulators. PLoS Biol. 15, e2002176 (2017).
Masuda, A. et al. CUGBP1 and MBNL1 preferentially bind to 3’ UTRs and facilitate mRNA decay. Sci. Rep. 2, 209 (2012).
Rattenbacher, B. et al. Analysis of CUGBP1 targets identifies GU-repeat sequences that mediate rapid mRNA decay. Mol. Cell Biol. 30, 3970–3980 (2010).
Chang, K. T., Cheng, C. F., King, P. C., Liu, S. Y. & Wang, G. S. CELF1 mediates connexin 43 mRNA degradation in dilated cardiomyopathy. Circ. Res 121, 1140–1152 (2017).
Chang, E. lizabethT. et al. The RNA-binding protein CUG-BP1 increases survivin expression in oesophageal cancer cells through enhanced mRNA stability. Biochem. J. 446, 113–123 (2012).
Villarroya, F., Cereijo, R., Villarroya, J., Gavalda-Navarro, A. & Giralt, M. Toward an understanding of how immune cells control brown and beige adipobiology. Cell Metab. 27, 954–961 (2018).
Pirzgalska, R. M. et al. Sympathetic neuron-associated macrophages contribute to obesity by importing and metabolizing norepinephrine. Nat. Med. 23, 1309–1318 (2017).
Guilherme, A. et al. Control of adipocyte thermogenesis and lipogenesis through beta3-adrenergic and thyroid hormone signal integration. Cell Rep. 31, 107598 (2020).
Razvi, S. et al. Thyroid hormones and cardiovascular function and diseases. J. Am. Coll. Cardiol. 71, 1781–1796 (2018).
Wang, H. et al. SLC35D3 promotes white adipose tissue browning to ameliorate obesity by NOTCH signaling. Nat. Commun. 14, 7643 (2023).
Fu, T. et al. Proteolytic rewiring of mitochondria by LONP1 directs cell identity switching of adipocytes. Nat. Cell Biol. 25, 848–864 (2023).
Lin, Y. et al. The chemerin-CMKLR1 axis limits thermogenesis by controlling a beige adipocyte/IL-33/type 2 innate immunity circuit. Sci. Immunol. 6, eabg9698 (2021).
Watson, M. J. & Thoreen, C. C. Measuring mRNA Decay with Roadblock-qPCR. Curr. Protoc. 2, e344 (2022).
Acknowledgements
We thank Dr. Zhichun Zhang for his guidance and support throughout this project. We are also grateful to Professor Lei Sun from Duke-NUS Medical School for instructing the RIP and RNA pull-down experiments. This research was supported by the National Key R&D Program of China (2018YFA0800401 to X.L.); the National Natural Science Foundation of China (No. 82070899 to X.L., 82000744 to X.L., 82303241 to L.X.); the general project of Natural Sciences Foundation of Chongqing (CSTB2022NSCQ-MSX0827 to X.L.); Talent Recruitment Program of Chongqing (CQYC20230A04321 to L.X.); the Program for Youth Innovation in Future Medicine, Chongqing Medical University (W0046 to F.Z.); Chongqing Medical University Start-up Fund (R2057 to L.X., R2019 to T.Z.).
Author information
Authors and Affiliations
Contributions
X.L. conceived and directed the study. T.Z. and L.X. designed experiments. T.Z. and J.L. performed experiments and analyzed the data. H.W., X.G., F.Z., X.Y., Y.W., T.Z., W.H., and H.Z. assisted in animal experiments. H.W., Y.C., and X.Z. helped with the bioinformatics analysis. T.Z. L.X., J.L., and X.L. wrote and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Jose Maria Moreno-Navarrete and Luc Paillard for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zeng, T., Xiao, L., Li, J. et al. Adipocyte RNA-binding protein CELF1 promotes beiging of white fat through stabilizing Dio2 mRNA. Nat Commun 16, 7414 (2025). https://doi.org/10.1038/s41467-025-62740-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-62740-8
