
Abstract
Targeted immunotherapy for T cell acute lymphoblastic leukemia (T-ALL), an aggressive tumor of developing T cell progenitors, is an urgent unmet need, especially for relapsed/refractory disease. Selective T-ALL targeting is challenging due to the shared antigen expression between leukemic and normal T cells. Here we identify the pre-T cell receptor (pre-TCR), a surface receptor essential for T cell development, as a biomarker of leukemia-initiating cells (LICs) in human T-ALL. Loss-of-function genetic approaches demonstrate that pre-TCR signaling is necessary for LIC activity and tumor progression in pre-TCR+ T-ALL patient xenografts in mice. Furthermore, we demonstrate the specific therapeutic targeting of the pre-TCR with a monoclonal antibody against the invariant pTα subunit of the human pre-TCR, and validate an anti-pTα antibody–drug conjugate in vivo treatment as a potent immunotherapy for inhibiting LIC activity and tumor progression of T-ALL in mice. These findings reveal the suitability of pre-TCR targeting as a promising therapy for the treatment of individuals with relapsed/refractory T-ALL expressing the pre-TCR.
Similar content being viewed by others

Main
T-ALL is an aggressive neoplasia of developing T cells1,2, with significant rates of chemotherapy resistance and relapses, associated with fatal outcomes (20–25% in pediatric patients and over 40% in adults)3,4,5. Therefore, the development of novel therapies that selectively target the LICs of T-ALL, which are the cell type responsible for relapse6, is an urgent medical need. In recent years, the emergence of antibodies, antibody–drug conjugates (ADCs) and chimeric antigen receptor (CAR) T cells as highly potent antigen-targeted immunotherapies has demonstrated unprecedented success in treating relapsed/refractory (R/R) B cell malignancies such as B cell acute lymphoblastic leukemia (B-ALL)7,8,9. However, extending this progress to T-ALL remains challenging, mainly because sharing of target antigens between normal and neoplastic T cells precludes tumor specificity, leading to aplasia of normal T cells and increased risk of life-threatening immunodeficiency in treated patients10,11,12. Identifying specific therapeutic targets involved in T-ALL pathogenesis and relapse, but not expressed in healthy T cells, is thus a crucial yet unmet goal.
T-ALL arises from the oncogenic transformation and developmental arrest of T cell precursors at early stages of intrathymic maturation. This is a multistep process involving aberrant expression of oncogenic transcription factors and accumulating genetic alterations in key oncogenic, tumor suppressor and T cell developmental pathways, which lead to uncontrolled cell survival and proliferation capacity13. T-ALL can be classified into different subtypes based on the activation of specific T-ALL oncogenes and associated gene expression profiles that correlate with the stage of arrest in T cell development14,15.
NOTCH1 is a critical pathway involved in T cell development16. Somatic activating mutations in NOTCH1 have been identified in a high frequency of T-ALL cases (>60%) from all major molecular subtypes, resulting in overexpression of constitutively active NOTCH1 (ICN1)17. Comprehensive genomic studies have confirmed the dominant presence of NOTCH1 as a driver mutation13,18. In fact, NOTCH1 activation is an early hallmark of T cell leukemogenesis and a key regulator of LIC activity in different T-ALL subtypes19,20,21, suggesting that deregulated transcription of NOTCH1 target genes and aberrant activation of downstream proliferation pathways critically contribute to T-ALL pathogenesis13,18,22. Therefore, understanding the role of molecular effectors downstream of NOTCH1 in T-ALL must provide key information for developing specific targeted therapies for this disease.
One of the NOTCH1 targets with a prominent role in T cell development is the pre-TCR. NOTCH1 transcriptionally regulates PTCRA23, the gene encoding the invariant pTα chain that associates with the TCRβ chain and forms the pre-TCR in developing T cells24,25,26. In both mice and humans, the pre-TCR is expressed transiently in intrathymic T cell precursors, and identifies a critical developmental checkpoint, known as β-selection, which leads to the survival and expansion of pre-T cells with functional TCRβ gene (TRB) rearrangements, and finally results in their differentiation into mature T cells expressing the TCRαβ26,27,28,29. The prominent function of the pre-TCR in intrathymic cellular expansion suggests its participation in leukemogenesis. However, variable results have been generated in mice, showing that pre-TCR function can be either important or dispensable for T-ALL pathogenesis, depending of the particular driver oncogene model used30,31,32,33,34,35,36. More importantly, little is known about pre-TCR expression and function in human T-ALL. Seminal studies propose that half of the TCRαβ-lineage T-ALL cases in patients might express a pre-TCR37, and more recent reports provide emerging evidence that point to pre-TCR activation as a therapeutic target38,39. Still, the significance of pre-TCR signaling in human T-ALL has not been directly addressed, and the involvement of the pre-TCR in the maintenance and progression of T-ALL in patients remains largely undefined. Herein, we addressed such important questions based on an in vivo model of human NOTCH1-induced T-ALL pathogenesis40, in combination with pre-TCR loss-of-function approaches in T-ALL patient-derived xenograft (PDX) models, and preclinical assays of immunotherapy based on administration of a monoclonal antibody specific for the extracellular domain of the pTα chain of the human pre-TCR41. Our study provides proof-of-concept data for the relevance of pre-TCR signaling in human T-ALL LIC activity and tumor progression, uncovering the suitability of the pre-TCR as a promising therapeutic target for treatment of patients with R/R T-ALL.
Results
Human T-ALL pathogenesis induced by active NOTCH1 involves a developmental arrest at the β-selection checkpoint
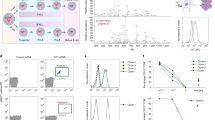
To study the multistep pathogenic process underlying T-ALL generation in patients, we recently extended the reported model of NOTCH1-induced murine T-ALL42 to humans40. To this end, immunodeficient NSG mice were intravenously (i.v.) injected with human cord blood CD34+ hematopoietic progenitor cells (HPCs) transduced with ICN1 and GFP as the cell tracer (Fig. 1a). All animals showing ICN1+ cell engraftment developed symptoms of disease and died of an aggressive leukemia that recapitulated T-ALL in patients40. Leukemic cells comprised a major (>95%) TCRβ+ clonal population of CD4+CD8+ double-positive cells with variable expression levels of surface CD3 (ref. 40 and Fig. 1b). Further phenotypic analysis revealed that a significant fraction of double-positive leukemic cells in all animals (up to 60% at 20–40 weeks after transplant) expressed low surface CD3 levels (CD3lo) and cells were negative for surface αβ and γδ TCR (TCRneg; Fig. 1b). This phenotype was compatible with a developmental arrest at the β-selection intrathymic stage where the pre-TCR is expressed25. Confirming the pathogenic nature of the arrested pre-T-like cells, we found that numbers of double-positive CD3loTCRneg cells increased progressively when infused into first and secondary recipient mice, in which they were able to serially transfer an aggressive T-ALL (Fig. 1c,d), indicating that they display a selective in vivo repopulating advantage characteristic of leukemic cells with LIC potential43. Therefore, we conclude that NOTCH1-induced human T-ALL cells arrested at the double-positive CD3loTCRneg β-selection stage include T-ALL LIC cells.

a, Schematic of the experimental design for de novo generation of human T-ALL. Sublethally irradiated NSG mice were subjected to i.v. injection with human cord blood (CB) CD34+ HPCs (1–1.5 × 105 cells per mouse) retrovirally transduced with either ICN1 along with GFP, or GFP alone as the control. b, Representative flow cytometry expression of CD4, CD8, CD3, TCRαβ and TCRγδ by ICN1+ human T-ALL cells infiltrating the bone marrow of diseased mice in a at 27 weeks after transplant. c, Representative CD3 versus TCRαβ expression of human ICN1+ T-ALL cells recovered from diseased mice in a (primary), or from NSG mice after serial transplantation with bone marrow from primary mice (1st transfer and 2nd transfer). Results correspond to T-ALL cells isolated from the bone marrow at 39, 8 and 9 weeks after transplant, respectively. d, Mean percentages ± s.e.m. of human CD8+CD4+ double-positive, CD3loTCRαβneg ICN1+ cells infiltrating the bone marrow of either primary diseased mice, or serially transplanted mice (nine mice per group from two independent experiments) as in c. Data were analyzed by one-way analysis of variance (ANOVA) with Kruskal–Wallis Dunn’s multiple-comparisons test. ****P < 0.0001.
Human T-ALL cells arrested at the β-selection checkpoint display LIC potential
To evaluate the clinical relevance of the double-positive CD3loTCRneg T-ALL population generated in the human T-ALL model, we assessed whether an equivalent population could be identified in patients. We examined a cohort of T-ALL cases characterized at the time of diagnosis (n = 37), including distinct biological subgroups according to their developmental stage, as defined by the European Group for Immunological Characterization of Leukemias (EGIL)14. We found that about half of T-ALL human samples in our cohort (22 of 37; 59%), classified as cortical (CD1a+) or pre-T (CD2+, CD5+), comprised variable proportions of CD3lo cells that lacked surface TCRαβ and TCRγδ (CD3loTCRneg), but expressed cytoplasmic TCRβ (cTCRβ), indicating that they were arrested at the β-selection stage (Table 1). To assess the pre-TCR status of the CD3loTCRneg T-ALLs, several human samples (n = 22) were propagated in NSG mice and then subjected to TRBV region sequencing and PTCRA quantitative PCR analysis. The results showed PTCRA transcription and clonal TRB rearrangements in essentially all CD3loTCRneg samples examined, with no dominant TCRVβ usage across individuals. In addition, expression of intracellular active NOTCH1 (ICN1) was a common feature of CD3loTCRneg T-ALL cells, as assessed by immunoblotting (Table 1). To further validate the results from our cohort, we took advantage of recently published RNA-sequencing and whole-genome sequencing datasets of T-ALL samples from more than 1,300 individuals15. We traced PTCRA expression and established correlations to 15 T-ALL subtypes defined in the study based on genomic drivers, gene expression patterns and developmental states15. The results indicated that 86% of T-ALL cases expressed PTCRA transcripts, as compared with public RNA-sequencing datasets of T cell, B cell and myeloid cell samples from healthy donors44 (Extended Data Fig. 1). PTCRA transcription was enriched in non-early thymic progenitor (ETP) T-ALLs (93% positive for PTCRA mRNA), as compared with near-ETP and ETP subtypes, with a maximal PTCRA mRNA expression found in essentially all cases within the TAL1 double-positive-like subtype. Notably, established correlations of PTCRA transcription to TRB, TRA, TRG and TRD gene rearrangements showed that 54% of T-ALL cases transcribed PTCRA and have rearrangements at the TRB locus but lacked TRA and either TRG or TRD rearrangements, according to a developmental arrest at the β-selection pre-TCR+ stage. Only 6.5% of T-ALL expressed PTCRA and displayed TRB and TRA rearrangements (Extended Data Fig. 1). Collectively, these data validate the results derived from our cohort, further supporting that >50% of T-ALL cases are arrested at the β-selection stage and display a CD3loTCRneg phenotype.
The observation that CD3loTCRneg cells generated de novo in our human T-ALL model display LIC activity (Fig. 1) prompted us to investigate the LIC potential of CD3loTCRneg cells from humans. Serial transplantation of three distinct T-ALL human samples into NSG mice revealed a selective in vivo growth advantage of the CD3loTCRneg population, resulting in a progressive increase of relative CD3loTCRneg cell numbers in the bone marrow and spleen of transplanted mice (Fig. 2a,b). More importantly, limiting dilution PDX assays of two distinct T-ALL samples showed that LIC activity was significantly enriched in vivo in fluorescence-activated cell sorting (FACS)-sorted CD3loTCRαβneg cells, compared with CD3+ TCRαβ+ cells isolated from the same T-ALL sample (40-fold and 220-fold in T-ALL17 and T-ALL43, respectively; Fig. 2c,d,f,g), and LIC frequencies inversely correlated with median survival of transplanted mice (Fig. 2e,h). Together, these results indicate that about half of human T-ALL cases include variable proportions of leukemic cells with LIC potential that display the CD3loTCRneg phenotype of thymocytes at the pre-TCR+ β-selection stage.

a, CD3 versus TCRαβ expression of CD4+ CD8+ double-positive gated cells from peripheral blood samples of three individuals with T-ALL (T-ALL9, T-ALL17, T-ALL18) analyzed by flow cytometry at the time of diagnosis (patient), or after serial transplantation into consecutive NSG mice (1st and 2nd transfer). Human cells engrafting the bone marrow of transplanted mice were analyzed when mice presented advanced symptoms of disease (70–92% leukemic burden). Numbers indicate percentages of gated CD3loTCRαβneg and TCRαβ+ cells. b, Mean ± s.e.m. percentages (when n = 3) of human CD3loTCRαβneg cells from T-ALL9 and T-ALL17 human samples engrafting the bone marrow and spleen of serially transplanted mice in a. For T-ALL9 bone marrow and spleen values from the first transfer, n = 3 in both tissues; from the second transfer, n = 1 in both tissues. For T-ALL17 bone marrow and spleen values from the first transfer, n = 3 in both tissues; from the second transfer, n = 2 and n = 1, respectively. Percentages at the time of diagnosis (human, n = 1 each) are shown for comparison. c,f, Sorting strategy used to isolate CD3loTCRαβneg and CD3+ TCRαβ+ T-ALL17 (c) and T-ALL43 (f) human cells recovered from spleen xenografts of NSG mice (left histograms). CD3 versus TCRαβ and cTCRβ expression of sorted populations is shown in the middle and right histograms, respectively. d,g, LIC potential of CD3+ TCRαβ+ and CD3loTCRαβneg human T-ALL17 (d) and T-ALL43 (g) sorted populations shown in c and f and transplanted into NSG mice under limiting dilution conditions (from 105 to 10 cells per mouse, 3–4 mice per dose). LIC frequency was calculated using ELDA software by recording numbers of leukemia-free mice in the peripheral blood by 29–32 weeks after transplant. e,h, Kaplan–Meier survival curves of mice transplanted in d and g with 103 FACS-sorted CD3+ TCRαβ+ or CD3loTCRαβneg human T-ALL cells were compared using log-rank Mantel–Cox test. *P = 0.0266 and *P = 0.0339, respectively. BM, bone marrow.
Functional pre-TCR expression is a biomarker of human T-ALL cells with LIC potential
To directly assess whether surface pre-TCR expression could be used as a biomarker of CD3loTCRneg T-ALL cells with LIC potential, we took advantage of a monoclonal antibody (K5G3) generated in our laboratory against the pTα component of the human pre-TCR41. This monoclonal antibody efficiently detects by flow cytometry the pre-TCR expressed on CD3loTCRneg cells of the SupT1 human T cell line, but it was unreactive with HPB-ALL and Jurkat T-ALL cell lines expressing the TCRαβ (Extended Data Fig. 2). Flow cytometry also revealed a specific, but low pTα expression on the surface of primary double-positive CD3loTCRneg cells either isolated from fetal thymus, or developing in vitro from cord blood HPCs in OP9-Jag2 co-cultures or from ETPs in artificial thymic organoids (ATOs; Extended Data Fig. 2). However, the anti-pTα monoclonal antibody showed an extremely low reactivity with CD3loTCRneg cells from different PDX-propagated T-ALL human samples, while no reactivity with TCRαβ+ leukemic cells expressing higher surface CD3 levels was observed (Fig. 3a,b). Collectively, our data suggest that the pre-TCR is expressed on the surface of CD3loTCRneg cells from T-ALLs arrested at the β-selection stage, but at extremely low levels, as would be expected from an activated complex with high internalization rate45.

a,b, Representative pseudocolor plots of CD3 versus TCRαβ expression displayed by T-ALL3, T-ALL17, T-ALL18 (a) and T-ALL2 (b) human samples recovered from spleen xenografts of NSG mice from two independent experiments (left histograms). Representative monoparametric histograms on the right show surface pre-TCR expression levels on electronically gated CD3loTCRαβneg, TCRαβ+ and CD3neg T-ALL cells analyzed with anti-pTα monoclonal antibody with either goat anti-mouse FITC (a) or biotin-coupled anti-mouse (Fab′)2 plus PE-coupled streptavidin (b). Numbers indicate percentages of positive cells. Gray shaded histograms show background staining with an irrelevant isotype-matched antibody. c, Immunoblot analysis of ERK and S6 phosphorylation in pre-TCR+ (T-ALL3 and T-ALL17) or TCRαβ+ (T-ALL2) T-ALLs, activated either with anti-pTα or with anti-CD3ε monoclonal antibodies for the indicated times. One experiment per T-ALL sample was performed. Total ERK and S6 expression is shown as the loading control.
A key question is whether pre-TCR expression levels that are barely detectable by flow cytometry, are functionally competent in human leukemias as observed in developing thymocytes. To investigate this issue, we examined the activation of two signaling pathways downstream of the pre-TCR, ERK and mTORC1 (refs. 46,47), following pre-TCR engagement with anti-pTα or an anti-CD3ε monoclonal antibody in several T-ALL samples. Immunoblotting assays showed similar ERK phosphorylation kinetics in response to either anti-pTα or anti-CD3ε monoclonal antibodies in all T-ALL samples examined, and both monoclonal antibodies were comparably efficient for mTORC1 activation, as indicated by enhanced phosphorylation of S6 over basal levels (Fig. 3c and Extended Data Fig. 3). In contrast, only anti-CD3ε was capable of inducing ERK and mTORC1 activation in a TCRαβ+ T-ALL sample, thus confirming the specificity of the anti-pTα monoclonal antibody (Fig. 3c). These results confirm that the low levels of pre-TCR detectable by flow cytometry on the surface of CD3loTCRneg T-ALL human cells are functionally competent. Thus, we concluded that surface expression of a functional pre-TCR is a biomarker of LIC cells in about 50% of individuals with T-ALL.
Pre-TCR signaling is required for LIC activity and tumor progression of human T-ALL
Next, we sought to assess whether pre-TCR signaling participates in the proliferation and function of human T-ALL LIC cells. To directly investigate this issue, we analyzed the LIC potential and tumor progression of T-ALL samples in which pre-TCR function was abrogated by silencing the CD2AP gene encoding CMS, a functional adaptor that binds to the human pTα cytoplasmic domain and is critical for pre-TCR activation48. We first established that CMS silencing by short hairpin RNA (shRNA; shCMS) inhibited CMS expression in the pre-TCR+ SupT1 cell line, and then confirmed that lack of CMS impaired pre-TCR function in the JR.pTα cell line48, as indicated by a significant inhibition of basal and induced PLCγ activation and NFAT activity after pre-TCR engagement49,50 (Extended Data Fig. 4a–c). More importantly, CMS silencing also led to deficient pre-TCR signaling in vivo, as shown by the impaired transition through β-selection and the differentiation blockage of human thymocytes developing in Rag2−/− Il2rg (γc)−/− double knockout mice (Rag2−/− γc−/−) infused with shCMS-transduced ETPs (Extended Data Fig. 4d–f).
Having validated that CMS silencing impairs pre-TCR function, we next evaluated tumor progression in NSG mice infused with human T-ALL samples (T-ALL5 and T-ALL9; Fig. 4a) transduced with either shCMS or a scramble shRNA (shSC) plus GFP as the cell tracer (Fig. 4b,e). Kinetic studies revealed a significant dissemination of leukemic cells in the peripheral blood of control mice at distinct times after transplant (7 to 10 weeks), and mice showed clear signs of disease by weeks 11 and 13 after transplantation with T-ALL9 and T-ALL5, respectively. At this endpoint, leukemic infiltration was observed in distinct organs including bone marrow, thymus, brain, liver and spleen. In contrast, leukemic expansion was significantly impaired in the peripheral blood and peripheral organs of mice infused with shCMS-transduced T-ALL cells, in accordance with an impaired LIC potential, and cells displayed the original phenotype (Fig. 4c,d,f,g). Supporting the specificity of shCMS on pre-TCR+ T-ALL cells, human-derived B-ALL cells transduced with shCMS were capable of expanding and progressing as efficiently as shSC-transduced B-ALL controls in transplanted mice, and no significant differences were found between both groups in either the peripheral blood or the bone marrow, spleen and liver, at different times after transplant (Fig. 4h–j). Collectively, these results provide evidence that defective pre-TCR signaling selectively impairs cell proliferation and tumor progression of human pre-TCR+ T-ALL cells in vivo, highlighting a crucial role for pre-TCR function in LIC activity of human T-ALL. In addition, our data raise the possibility that the pre-TCR has also an active role in the pathogenesis of pre-TCR+ T-ALL cells. Supporting this view, the ICN1 construct shown above to induce human pre-TCR+ leukemia (Fig. 1) failed to generate mouse T-ALL when overexpressed in bone marrow HPCs from CD3ε mutant mice (C80G), which display a deficiency in pre-TCR signaling51, although C80G HPCs display multilineage potential (Extended Data Fig. 5). In contrast, NSG mice infused with ICN1-transduced wild-type HPCs developed an aggressive leukemia, leading to infiltration of peripheral organs and a marked splenomegaly, and resulting in a median survival of 5 weeks (Extended Data Fig. 5). These results suggest that the pre-TCR may be involved in T-ALL pathogenesis of human T-ALL cases arrested at the pre-TCR+ β-selection stage. Collectively, these data point to the pre-TCR as a promising therapeutic target for T-ALL.

a, Schematic of the experimental design for abrogation of pre-TCR function in T-ALL human cells. T-ALL (T-ALL5 and T-ALL9) or B-ALL (B-ALL9) cells recovered from bone marrow xenografts of NSG mice were transduced with a lentiviral vector encoding an shRNA against CMS (shCMS) along with GFP as the cell tracer, or a control scramble RNA (shSC) and GFP, before infusion into sublethally irradiated NSG mice. b,e,h, Transduction efficiencies of T-ALL5 (b) T-ALL9 (e) and B-ALL9 (h) analyzed by flow cytometry. Numbers indicate percentages of transduced (GFP+) cells before transplantation. c,f,i, Percentages ± s.e.m. of CD45+ GFP+ T-ALL5 (c), T-ALL9 (f) and B-ALL9 (i) cells infiltrating the peripheral blood (PB) of transplanted mice. For shSC-transduced and shCMS-transduced T-ALL5 cells, n = 6 and n = 8 mice, respectively, from two independent experiments, at both 8 and 10 weeks after transplant (***P = 0.0007 for both times, c); for shSC-transduced and shCMS-transduced T-ALL9 cells, n = 4 and n = 3 mice, respectively, from one experiment, at both 7 and 8 weeks after transplant (*P = 0.0326 and **P = 0.0083, respectively, f); for shSC-transduced and shCMS-transduced B-ALL9 cells, n = 4 and n = 3 mice, respectively, from one experiment, at 5 weeks after transplant (i). d,g,j, Relative numbers ± s.e.m. of T-ALL5 (d), T-ALL9 (g) and B-ALL9 (j) cells transduced with shCMS or shSC, engrafting the indicated organs of NSG mice euthanized when they presented advanced symptoms of disease (13, 11 and 7 weeks after transplant for T-ALL5, T-ALL9 and B-ALL9, respectively). Data are shown as the mean ± s.e.m. percentages of CD45+ GFP+ engrafted cells normalized to percentages of GFP+ injected cells (b, e and h). For shSC-transduced and shCMS-transduced T-ALL5 cells (d), n = 4 thymus samples from one experiment, **P = 0.0075; n = 6 and n = 8, respectively, bone marrow samples from two independent experiments, **P = 0.0032; n = 2 and n = 4, respectively, brain samples, from one experiment; n = 6 and n = 8, respectively, spleen and liver samples from two independent experiments, ***P = 0.0002 for both samples. For shSC-transduced and shCMS-transduced T-ALL9 cells (g), n = 4 thymus samples from one experiment, ***P = 0.0002; n = 4 bone marrow samples, from one experiment, **P = 0.0015; n = 4 spleen and liver samples from one experiment, ****P < 0.0001, and n = 4 LN samples from one experiment, **P = 0.0056. For shSC-transduced and shCMS-transduced B-ALL9 cells (j), n = 4 and n = 3, respectively, bone marrow and liver samples from one experiment. Data were analyzed by a two-tailed unpaired t-test with Mann–Whitney correction (c, f and i) and two-tailed unpaired multiple t-test (d, g and j). LN, lymph node.
Preclinical validation of pre-TCR therapeutic targeting for T-ALL treatment
To directly evaluate the therapeutic potential of pre-TCR targeting, we used a PDX preclinical assay, in which SCID mice infused with a human T-ALL (T-ALL-3) were subjected to intraperitoneal (i.p.) injection with either the anti-pTα monoclonal antibody or an isotype-matched (IgG2a) control (10 mg per kg body weight, twice a week) for 10 weeks. Treatment was initiated at week 3 after transplant, when the disease was clearly established (>1% of total peripheral blood white cells; Fig. 5a). Sequential analysis of peripheral blood revealed that T-ALL engraftment increased dramatically to 90% during the first 8 weeks after transplant in control animals, while only 5% of leukemic cells infiltrated the peripheral blood of anti-pTα-treated mice at this time (Fig. 5b). Thereafter, T-ALL progression was significantly delayed in the peripheral blood of anti-pTα-treated mice, as compared with controls, which showed clear symptoms of disease by 11 weeks after transplant and should be euthanized. Accordingly, the median survival of anti-pTα-treated mice was significantly prolonged (>50%) when compared with that of isotype control-treated mice (Fig. 5c), validating the therapeutic potential of the anti-pre-TCR targeting strategy based on anti-pTα monoclonal antibody administration.

a,d, Schematic design of in vivo experimental treatment of human T-ALL with an anti-pTα monoclonal antibody. Sublethally irradiated SCID (a) or NSG mice (d) were subjected to i.v. injection with T-ALL3 (b; 106 cells per mouse) or T-ALL42 (e; 2 × 106 cells per mouse) samples, respectively, recovered from spleen xenografts of NSG mice. Mice were serially i.p. injected (twice weekly for 3–10 weeks) with either anti-pTα monoclonal antibody or isotype-matched control (IgG2a, 10 mg per kg body weight), starting at week 3-4 after transplant, when T-ALL engraftment in peripheral blood was >1%. b,e, Percentages of T-ALL3 (b) and T-ALL42 (e) cells infiltrating the peripheral blood of mice at the indicated days after transplant, upon treatment with anti-pTα monoclonal antibody or IgG2a control. Data from one experiment are shown as the mean ± s.e.m. cell percentages from control (n = 7) or anti-pTα-treated (n = 8) mice in b, ****P < 0.0001; or from control, or anti-pTα-treated or VxL-treated mice (n = 7 per group) in e, analyzed by a mixed-effects model (REML). c, Kaplan–Meier survival curves of mice in b were compared using a log-rank Mantel–Cox test. ***P = 0.0006).
To assess the potential mechanism responsible for the anti-pTα-mediated inhibition of T-ALL expansion in vivo, we next assayed the therapeutic activity of the anti-pTα monoclonal antibody in a PDX assay using NSG mice (Fig. 5d), which, in contrast to SCID mice, have a deficiency in functional NK cells and in complement-dependent cytotoxicity (CDC; ref. 52 and The Jackson Laboratory; JAX 005557). The results showed no significant effect of anti-pTα monoclonal antibody administration on T-ALL progression, as compared with T-ALL expansion in mice treated with an IgG2a isotype-matched control. However, treatment with three cycles of established chemotherapy based on vincristine, dexamethasone and l-asparaginase (VxL) drastically inhibited tumor growth (Fig. 5e). These results strongly suggest that NK cell-mediated antibody-dependent cellular cytotoxicity (ADCC) and CDC may contribute to anti-pTα-mediated inhibition of T-ALL growth in vivo.
Targeted immunotherapy with anti-pTα ADCs efficiently impairs tumor progression of human pre-TCR+ T-ALL in vivo
ADCs have emerged as a compelling clinical option particularly effective at targeting surface receptors endowed with dynamic internalization properties53, such as those exhibited by the pre-TCR45,48. Notably, the anti-pTα monoclonal antibody is internalized after binding to the pre-TCR and is trafficked into lysosomes48, indicating that it could be optimal for delivery of cytotoxic drugs. To maximize the efficacy of a potential pre-TCR targeted immunotherapy, the anti-pTα monoclonal antibody was subjected to drug conjugation using either non-cleavable or cleavable linkers and either mertansine (DM1) or monomethyl auristatin E (MMAE), respectively (Fig. 6a), two toxins previously used in the clinic53. The resulting anti-pTα ADCs were functionally validated in vitro for their capacity of binding to the pre-TCR expressed on SupT1 and JR.pTα cells48, as compared with the drug-free monoclonal antibody, while all anti-pTα variants were unreactive with TCRαβ+ Jurkat cells and with TCRα− Jurkat mutant (JR.MigR1) cells (Extended Data Fig. 6). Flow cytometry also confirmed that both anti-pTα ADC variants displayed optimal features for cargo intracellular delivery, as they induced a rapid internalization of surface pre-TCR (>60–70% in 5 min), with kinetics similar to those observed with the uncoupled anti-pTα monoclonal antibody (Extended Data Fig. 6). Because DM1 and MMAE efficiently disrupt the cytosolic microtubule network leading to blocked mitosis, impaired proliferation and apoptosis53, we next assessed the impact of the generated anti-pTα ADCs on the T-ALL cell cycle. In vitro functional assays showed that both ADCs impaired cell cycle progression in the pre-TCR+ cell lines, SupT1 and JR.pTα, by inducing G2/M cell cycle arrest (Fig. 6b,c), which resulted in a significant and specific decrease in cell proliferation (Fig. 6d), but they had no effect on pre-TCR-negative HPB-ALL and JR.MigR1 cell lines.

a, Illustration of the modular nature of anti-pTα-DM1 and anti-pTα-MMAE ADCs, showing non-cleavable (MCC) and cleavable (SC-VC-PABC) linkers used. b, Representative cell cycle profiles of SupT1 pre-TCR+ cells left untreated (medium) or treated with drug-free anti-pTα monoclonal antibody, anti-pTα-DM1 or anti-pTα-MMAE ADCs, or their respective isotype-matched controls for 4 days. Numbers indicate percentages of cells in the G2 and M phases of the cell cycle. c, Mean percentages ± s.e.m. of cells in the G2 and M phases of the cell cycle within the indicated pre-TCR-positive (SupT1, JR.pTα) and pre-TCR-negative (CUTLL1, HPB-ALL, JR.MigR1) cell lines, treated as in b. In the left bar plots, for SupT1 cells untreated and anti-pTα-DM1-treated, n = 8 each; IgG2a-DM1-treated, n = 6; and anti-pTα-treated, n = 7 (****P < 0.0001). For CUTLL1 cells untreated, n = 7; IgG2a-DM1-treated, n = 4; and anti-pTα- and anti-pTα-DM1-treated, n = 3 each. For HPB-ALL untreated, n = 7; IgG2a-DM1-treated, n = 4, and anti-pTα- and anti-pTα-DM1-treated, n = 3 each. In the right bar plots, n = 6 for each group of SupT1, JR.MigR1 and JR.pTα cells. d, Relative cell proliferation of the indicated leukemic cell lines recovered after in vitro treatment as in b. Data are shown as the mean ± s.e.m. percentages of recovered treated cells relative to untreated cells. In the right bar plots, for SupT1 cells untreated, n = 4; IgG2a-DM1-treated, n = 7; and anti-pTα- and anti-pTα-DM1-treated, n = 8 each (****P < 0.0001). For CUTLL1 and HPB-ALL cells untreated, n = 7 each; IgG2a-DM1- and anti-pTα-DM1-treated, n = 3 each; and anti-pTα-treated, n = 4 each (*P = 0.0213, ****P < 0.0001). Data were analyzed by two-way-ANOVA with a Tukey’s multiple-comparisons test.
The functional validation of the generated ADCs prompted us to investigate the in vivo efficacy of a specific immunotherapy based on anti-pTα ADC administration. To this end, NSG mice infused with a human pre-TCR+ T-ALL (T-ALL42) were subjected to i.p. injection twice a week for 3 weeks with anti-pTα-MMAE or with an irrelevant IgG2a-MMAE, starting when the disease was clearly established (>1% T-ALL cells in peripheral blood by 5 weeks after transplant; Fig. 7a). Peripheral blood examination of control mice revealed a massive expansion (up to 90%) of leukemic cells from weeks 8 to 10 after transplant. Despite the severity of the disease, administration of anti-pTα-MMAE ADCs resulted in a significant inhibition of leukemia burden in the peripheral blood that was maintained for several weeks after the end of treatment (Fig. 7b). Of note, median survival of treated mice was significantly prolonged (>60%) when compared to that of control animals, (Fig. 7c), indicating that pre-TCR ADC targeting is an efficient T-ALL immunotherapy.

a, Timeline of in vivo pre-TCR targeting immunotherapy based on administration of anti-pTα-MMAE or its corresponding isotype-matched control to NSG mice i.v. injected with T-ALL42 human cells (2 × 106 cells per mouse) recovered from spleen xenografts of NSG mice. b, Mean ± s.e.m. percentages of T-ALL42 cells recovered at the indicated days from the peripheral blood of NSG mice (n = 6) treated as indicated in a; ****P < 0.0001. c, Kaplan–Meier survival curves of NSG mice in b were compared using a log-rank Mantel–Cox test, ****P < 0.0001. d, Timeline of in vivo pre-TCR targeting immunotherapy based on administration of drug-free anti-pTα monoclonal antibody, anti-pTα-DM1 or anti-pTα-MMAE ADCs, or their respective isotype-matched controls, or VxL chemotherapy, to SCID mice i.v. injected with T-ALL3 human cells (106 cells per mouse) recovered from spleen xenografts of NSG mice. Doses of anti-pTα-DM1 and anti-pTα-MMAE ADCs were based on the drug-to-antibody ratio (DAR), which was three and six molecules of DM1 and MMAE per antibody molecule, respectively. e, Mean ± s.e.m. percentages of T-ALL3 cells recovered at the indicated days from the peripheral blood of SCID mice treated as indicated in d. For VxL treatment, n = 6; for IgG2a and anti-pTα treatment, n = 7 each; for IgG2a-DM1, IgG2a-MMAE, anti-pTα-DM1 and anti-pTα-MMAE, n = 6 each, from one experiment. f, Mean ± s.e.m. percentages of electronically gated CD3loTCRαβneg cells infiltrating the peripheral blood of SCID mice in e at day 91 after transplant. For IgG2a and anti-pTα treatment, n = 7 each; for IgG2a-DM1, IgG2a-MMAE and anti-pTα-DM1 treatment, n = 6 each, and for anti-pTα-MMAE treatment, n = 4. Data were analyzed by a mixed-effects model (REML; b) or by one-way (f) or two-way (e) ANOVA with Tukey’s multiple-comparisons test. ***P = 0.0001; ****P < 0.0001.
Next, we sought to directly compare the therapeutic efficacy of the anti-pTα ADCs to that of the unconjugated monoclonal antibody. Thus, SCID mice infused with a human-derived T-ALL (T-ALL3) were subjected to i.p. injection (twice a week for 3 weeks, starting at week 4 after transplant), with either the unconjugated anti-pTα monoclonal antibody or the DM1-coupled or MMAE-coupled ADCs, or their corresponding isotypic controls, also including VxL treatment for comparison (Fig. 7d). We observed rapid tumor progression in control mice, with >90% of T-ALL infiltration in peripheral blood by week 11 after transplant. Of note, unconjugated anti-pTα monoclonal antibody as well as MMAE-coupled and DM1-coupled ADCs displayed a strong anti-leukemic effect, resulting in a significantly delayed T-ALL dissemination in the peripheral blood. However, ADCs, particularly anti-pTα-MMAE, were more efficient than the unconjugated monoclonal antibody in suppressing T-ALL growth, as indicated by a longer-lasting effect, and maximal efficacy was observed with VxL treatment (Fig. 7e). Supporting the specificity of the therapy, the anti-leukemic effect of both anti-pTα monoclonal antibody and ADCs correlated with decreased proportions of CD3loTCRneg cells, as observed in the peripheral blood of late-stage treated mice (13 weeks after transplant), compared with control mice (Fig. 7f).
Finally, although no off-target effects derived from the pre-TCR targeted therapy are expected, we sought to investigate its potential deleterious effect on normal T cell development. To this end, human cord blood HPCs (105) were i.v. injected into NSG mice, which were treated with either the anti-pTα-DM1 ADC or an IgG2a-DM1 isotypic control for 3 weeks, starting at 9 weeks after transplant, when reconstitution with human cells was confirmed in the peripheral blood of all transplanted mice, but <1% of T cells were detected. Three weeks later, we assessed the impact of the treatment on the human T cell populations generated in the thymus and periphery of treated mice (Extended Data Fig. 7a). We found no significant differences in the relative kinetics of human T cell reconstitution in the peripheral blood of mice treated with anti-pTα-DM1, compared with IgG2a-DM1-treated controls, although a trend toward lower proportions was observed in the former, and human B cell reconstitution was similar in both groups, as expected (Extended Data Fig. 7b). Likewise, no significant changes were observed in the relative numbers of human T cells repopulating the spleen in the two groups of mice (Extended Data Fig. 7c). Regarding human T cell development, we found no significant differences in the proportions of human cells placed upstream or downstream of pre-TCR expression recovered from thymi of anti-pTα-DM1-treated mice, as compared to IgG2a-DM1-treated controls, and neither significant differences were observed in pre-TCR-expressing pre-T cells (Extended Data Fig. 7d). These data indicate that generation of new human T cells is not severely affected by the proposed ADC therapy. Moreover, similar proportions of peripheral T (and B) cells repopulating the peripheral blood and spleen were recovered from mice treated with anti-pTα-DM1 or IgG2a-DM1, starting at 21 weeks after transplant, when a significant T cell reconstitution (25–30%) was evident in the periphery, thus indicating no effect of ADC treatment on normal mature T cells (Extended Data Fig. 7a,e,f). In the thymus, a slight, but not significant, decrease of thymic pre-T cells was observed in ADC-treated mice (Extended Data Fig. 7g). Therefore, no severe adverse effects on normal T cell development are expected from the application of the proposed therapy. Collectively, our results provide evidence for the efficacy and safety of an anti-pTα ADC targeted immunotherapy against pre-TCR+ T-ALL cells with LIC potential, offering a promising strategy to prevent and treat T-ALL relapses.
Discussion
In this study, we have identified the pre-TCR as a targetable pathway exhibiting key molecular and functional features optimal for developing a selective T-ALL immunotherapy. This finding represents an important advance in the field, as T-ALL targeted therapies have remained underdeveloped owing to challenges in identifying targetable pathways restricted to malignant T cells. Critically, the pre-TCR is not expressed on healthy mature T cells, but it was previously proposed to be present in about half of TCRαβ-lineage T-ALL cases that display the TCR status of pre-TCR+ thymocytes arrested at the β-selection developmental stage37. Here we show that such T-ALL cases can be identified by a selective CD3loTCRneg phenotype that associates with PTCRA transcription and clonal TRB, but lack of TRA rearrangements, and confirm that they represent ~50% of T-ALL cases. Moreover, functional studies formally demonstrate that human T-ALLs with a ‘β-selection’ phenotype are reactive with a specific anti-human pTα monoclonal antibody41 that induces pre-TCR activation, indicating that they express a surface pre-TCR endowed with signaling properties. This discovery provided the molecular basis for developing an anti-pre-TCR ADC targeted therapy, which efficiently impairs T-ALL progression and increases survival in preclinical assays, as proof of concept of a promising immunotherapy for individuals with T-ALL.
An important aspect uncovered by this study is that functional pre-TCR is a biomarker of T-ALL cells with LIC potential. The therapeutic relevance of this finding is strengthened by the observation that pre-TCR signaling is required for LIC function, a feature that may prevent the loss of cell-surface pre-TCR from T-ALL in response to therapeutic pressure. Accordingly, we found that late-stage (>3 months) tumor progression in our treated mice is associated with the emergence of leukemic cells with the CD3loTCRneg pre-TCR phenotype. The pre-TCR dependency of malignant LIC growth is likely to result from the physiologic proliferative signals induced by the pre-TCR, rather than from a specific oncogenic effect, suggesting that signaling pathways downstream of the pre-TCR may be key therapeutic targets, as has been recently proposed for the LCK pathway38,39. Importantly, here we provide direct biochemical and functional evidence that several pre-TCR signaling pathways activated at the β-selection checkpoint, including ERK, mTORC1 and PLCγ46,47,48,49,50, are linked to pre-TCR signaling also in T-ALL cells. Although further studies are required to confirm that pre-TCR activation works similarly in leukemias and developing pre-T cells, it is worth noting that human T-ALLs recapitulate physiological pre-TCR expression features, with limited numbers of pre-TCR complexes reaching the leukemic cell surface45,54. Of note, such a low pre-TCR expression was sufficient to induce a selective growth advantage to T-ALL cells in vivo, a finding with important therapeutic implications regarding the functional efficacy of pre-TCR targeting with specific monoclonal antibodies. Therefore, while weak pre-TCR expression makes particularly challenging the implementation of flow cytometry assays for assessment of pre-TCR expression in humans, it does not preclude the viability of pre-TCR targeting strategies against T-ALL cases displaying a CD3loTCRneg phenotype.
Regarding the role of the pre-TCR in T-ALL pathogenesis, studies in mice produced variable results depending on the particular driving oncogenes involved30,31,32,33,34,35,36. However, it is well known that pre-TCR function commonly accelerates leukemia onset, increases leukemia aggressiveness and supports tumor survival in different T-ALL mouse models32,33,36. These findings concur with the observation that human progenitors undergoing pre-TCR-mediated β-selection are particularly prone to malignant transformation37. Therefore, the pre-TCR may play a cooperative role in distinct human T-ALL oncogenic settings. This is particularly exemplified by oncogenic NOTCH1, as ICN1 constructs with different activation potential can lead to either pre-TCR-dependent (ref. 30; Extended Data Fig. 5) or pre-TCR-independent35 T-ALL generation in mice, although pre-TCR signaling largely facilitates NOTCH1-induced leukemogenesis35, as also does NOTCH3 (ref. 31). Given that PTCRA is a NOTCH1 target23, and NOTCH1 activating mutations are found in most molecular T-ALL subgroups17,18, it is possible that the pre-TCR has a common role facilitating leukemogenesis in distinct human T-ALL subtypes with active NOTCH1. Accordingly, pre-TCR expression and NOTCH1 activation are coincident events in our T-ALL human cohort (Table 1).
Overall, the dependency of T-ALL pathophysiology on pre-TCR function provides compelling evidence that the pre-TCR may be an ideal targetable pathway for fighting relapse of distinct T-ALL subtypes. Two additional molecules, CD1a55 and CCR9 (ref. 56), have recently been proposed as suitable therapeutic targets for treatment of R/R T-ALL with CAR-T cells; but, in contrast to the pre-TCR, no evidence supports the requirement of these molecules in T-ALL initiation or progression. Although comprehensive phenotypic and molecular studies in relapsed individuals are needed to confirm the CD3loTCRneg pre-TCR+ phenotype in R/R T-ALLs, our preclinical data provide evidence that targeting the pre-TCR with an anti-pTα monoclonal antibody is a valid therapeutic strategy against LIC-mediated T-ALL relapse. This opens new avenues for the development of additional targeting therapies such as BiTEs and CAR-T cells. In vivo administration of the anti-pTα monoclonal antibody markedly impairs T-ALL progression, likely through induction of CDC and/or ADCC, although we cannot formally exclude a role for monocytes/macrophages in mediating ADCC or antibody-dependent cellular phagocytosis (ADCP). However, a direct pro-apoptotic effect of anti-pTα, similar to that induced by persistent TCR/CD3 engagement of TCRαβ+ T-ALLs57, seems unlikely, considering that T-ALL growth was not impaired in NSG mice devoid of CDC and ADCC functions52, and no effect was found in vitro on T-ALL cell lines. Whatever mechanism mediates the therapeutic function of the anti-pTα monoclonal antibody, the particular endocytosis and internalization features of the pre-TCR45,54 suggested that anti-pTα coupled to cytotoxic drugs (ADCs) could provide a more potent therapy53. Supporting this hypothesis, comparative experiments directly demonstrated the superior therapeutic efficacy of ADCs as compared with unconjugated antibodies, with particularly significant results for the MMAE-coupled ADCs. Importantly, the safety and efficacy of the ADC strategy for various hematologic tumors have been established in clinical trials53 and, recently, preclinical data support the ability of anti-TRBC1 ADCs to cure disseminated T cell cancers58. Other safety aspects to be considered rely on the possible off-target effects of the proposed therapy. Because the pre-TCR is exclusively expressed within the body in a transient population of thymus-restricted T cell progenitors25,29, no severe or irreversible toxicities are expected in young infants, in whom pre-TCR− progenitors upstream of β-selection could eventually regenerate the pre-TCR+ cell pool, which will ultimately mature into functional T cells. Consistent with this, we show that anti-pTα ADC treatment in humanized mice has no significant impact on the generation of new human T cells. Furthermore, as no deleterious effects on peripheral mature T cells were observed in treated mice, no severe defects are anticipated in pediatric and adult individuals, who already display a peripheral long-lasting mature T cell repertoire established early in life59. Therefore, we conclude that the anti-pTα ADC targeting strategy validated in this study holds strong therapeutic promise for individuals with R/R T-ALL.
Methods
Human and mouse cell samples
Human samples were obtained according to the Declaration of Helsinki principles and to the procedures approved by Local Bioethics Committees (Hospital Infantil Universitario Niño Jesús, UAM, Madrid, Spain; Centro de Transfusión de la Comunidad de Madrid, Spain; and Hospital Universitario 12 de Octubre, Madrid, Spain), after written informed consent was provided. Sex information was unknown and, therefore, was not taken into account in the study design for human samples. Human postnatal thymus samples were removed during corrective cardiac surgery of individuals aged 1 month to 4 years. Fetal thymus samples were obtained from volunteers after legal termination of pregnancy. ETPs were isolated by inmunomagnetic sorting from postnatal thymocyte suspensions obtained by centrifugation on Ficoll-Hypaque (Lymphoprep, Axis-Shield PoC AS) using the Dynal CD34 Progenitor Cell Selection System (Life Technologies). HPCs were obtained from Ficoll-Hypaque-purified cord blood samples using the CD34 Progenitor Cell Isolation Kit (Miltenyi Biotec)40. Sorted populations were at least 99% CD34+ and negative for CD3, CD4, CD8, CD13, CD14, CD19 and CD56 lineage markers on reanalysis (Lin−). Primary T-ALL cells were Ficoll-Hypaque-purified from peripheral blood or bone marrow pediatric (<18 years old) human samples obtained at the time of diagnosis. Mouse HPCs were isolated by cell sorting (FACSAria, BD Biosciences) of cells labeled with APC-coupled anti-CD117 (c-kit; BD Biosciences), from Lin− bone marrow samples from 6- to 10-week-old C57BL/6 mice (The Jackson Laboratory) or CD3ε-mutant (C80G) mice51. Lin− cells were obtained by immunomagnetic sorting using biotin-coupled monoclonal antibodies against B220/CD45R, Gr1.1, CD11b, CD3 or Ter-119 (BD Biosciences) and streptavidin-coupled magnetic spheres (Miltenyi Biotec)60.
Flow cytometry
All antibodies and staining reagents used in the study are outlined in supplementary Table 1.
Flow cytometry was performed on a FACSCanto II (BD Biosciences) or on an AURORA 4L (Cytek) cytometer. Data were collected using BD FACSDiva software version 8.0.1 and SpectroFlo software version 3.3.1. Data analysis was performed using FlowJo software version 10 (BD Biosciences).
Cell lines, OP9-Jag2 cultures and ATOs
Human T-ALL cell lines SupT1 (CRL-1942) and Jurkat (TIB-152) were obtained from the American Type Culture Collection and HPB-ALL from DSMZ (ACC 483), and CUTLL1 was kindly provided by A. Ferrando61. To verify the identity of HPB-ALL and Jurkat cell lines, both listed in cross-contamination databases (https://iclac.org/databases/cross-contaminations/), a flow cytometry panel was used to analyze the expression of CD8, CD4, CD1a and CD44 surface markers. This strategy confirmed that HPB-ALL cells are CD4+CD8+CD1a+CD44+, whereas Jurkat cells are CD4+CD8−CD1a−CD44−. JR.pTα and JR.MigR1 were generated by lentiviral transduction of TCRα-deficient Jurkat cells (JR3.11) with MigR1-pTα or MigR1-control lentiviral vectors, respectively48. T-ALL cell lines were cultured in RPMI 1640 medium (Lonza) supplemented with 10% FBS (Gibco). For in vitro T cell differentiation, HPCs isolated from cord blood were seeded (105 per ml) on semiconfluent monolayers of OP9-Jag2 stromal cells generated by retroviral transduction of OP9 cells with Jag2 ligand62. Cells were cultured in 24-well plates with α-MEM medium (Gibco) supplemented with 20% FBS, 200 IU per ml of recombinant human (rh)IL-7, 100 IU per ml of rhSCF (National Institute of Biological Standards and Controls) and 50 ng ml−1 of rhFlt3-L (PeproTech). T cell development was analyzed from human ETPs cocultured in ATOs generated with the stromal cell line MS5 transduced with the Notch ligand DLL4 (MS5-hDLL4), kindly provided by G. Crooks63. ATOs were plated in a 0.4 μM Millicell Transwell (Thermo Fisher Scientific) insert and placed in one well of a six-well plate in 1 ml serum free culture medium (RB27) composed of RPMI 1640 (Lonza), 4% B27 supplement (Thermo Fisher Scientific), 30 μM l-ascorbic acid 2-phosphate sesquimagnesium salt hydrate (Sigma-Aldrich), 1% Glutamax (Thermo Fisher Scientific), 200 IU per ml rhIL-7 and 5 ng ml−1 rhFlt3-L.
T-ALL generation, xenotransplantion and LIC activity assay
All animal studies were approved by the Animal Experimentation Ethics Committee of the Comunidad de Madrid Animal Research Review Board (PROEX002621). All results presented include both female and male animals. No sex-specific effects were observed, and the study did not consider sex as a biological variable. Human T-ALL was generated as described40 in sublethally irradiated (1.5 Gy) 6- to 10-week-old NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ (NSG; The Jackson Laboratory) mice subjected to i.v. injection with human cord blood HPCs retrovirally transduced with the ICN1 construct previously described64. Mouse T-ALL was generated following a modification of the original model42, in sublethally irradiated (1.5 Gy) 6- to 10-week-old NSG mice subjected to i.v. injection with mouse (C57BL/6) bone marrow Lin− c-kit+ HPCs transduced with ICN1, as reported60. Human in vivo T cell development was analyzed in sublethally irradiated (3.5 Gy) 3- to 5-day-old Rag2−/− γc−/− mice65 subjected to intrahepatic injection with shCMS-transduced or shSC-transduced human ETPs (105), or in sublethally irradiated (1.5 Gy) 6- to 10-week-old NSG mice subjected to i.v. injection with human cord blood HPCs. At 3 and 5 weeks after transplant, thymi from host mice were collected and analyzed for reconstitution with human developing T cells by flow cytometry. For leukemia xenotransplantation assays, sublethally irradiated (1.5 Gy) 6- to 10-week-old NSG or CB17/lcr-Prkdscid/lcrlcoCrl (SCID; The Jackson Laboratory) mice were subjected to i.v. injection with T-ALL or B-ALL human samples or with human T-ALLs generated de novo in host mice (first grafts). When indicated, first grafts from host mice were i.v. injected into secondary recipients (second grafts). LIC activity was analyzed by transplantation into sublethally irradiated (1.5 Gy) 6- to 10-week-old NSG mice under limiting dilution conditions (105 to 10 cells per mouse, 1–4 mice per dose)60. Transplanted mice were analyzed by flow cytometry for the presence of human CD7+CD45+ blasts in peripheral blood from 6 to 32 weeks after transplant, and the frequency of leukemia-free mice was calculated using ELDA software (http://bioinf.wehi.edu.au/software/elda/)66. In all experiments, mice were euthanized when they presented advanced symptoms of disease.
shRNA lentiviral constructs and transductions
For silencing endogenous CMS expression, five distinct shRNAs directed against CD2AP encoding human CMS (shCMS) (Sigma-Aldrich Mission TRCN shRNA Target set, TRCN0000119097, TRCN0000119098, TRCN0000119099, TRCN0000119100, TRCN0000119101) were assayed by transfection and puromycin selection (1 μg ml−1; Sigma-Aldrich) of SupT1 cells, then analyzed for CMS expression by immunoblotting. The best-performing shCMS or a scrambled shRNA (shSC), used as control, were then cloned under the U6 promoter into the pHRSIN-GFP lentiviral vector. T-ALL and B-ALL cell transduction was performed as described60 in the presence of rhIL-7 (200 IU per ml).
Immunoblotting
Activation of signaling pathways downstream of the pre-TCR was analyzed by immunoblotting of cells incubated with 10 μg ml−1 of anti-CD3ε (UCHT-1; BD Biosciences) or anti-pTα (K5G3)41 monoclonal antibodies at 4 °C for 30 min, followed by crosslinking with 10 μg ml−1 of anti-mouse IgG (Sigma-Aldrich) at 37 °C for the indicated incubation times. Whole-cell lysates (RIPA buffer) separated on 10% SDS–PAGE (Bio-Rad) were transferred to polyvinylidene difluoride membranes (Merck Life Science) that were incubated with antibodies against phospho 44/42 MAPK (ERK1/ERK2), p44/42 MAPK ERK, pS6 ribosomal (pS6), S6 (Cell Signaling) or CMS (Santa Cruz Biotechnology). α-tubulin expression (Sigma-Aldrich) was analyzed as the loading control. For PLCγ activation analysis, cells were lysed in 1 ml Brij96 lysis buffer containing protease and phosphatase inhibitors, and subjected to immunoprecipitation with anti-PLCγ1 monoclonal antibody (BD Bioscience) and protein G-sepharose CL-4B beads (Sigma-Aldrich). After SDS–PAGE and immunoblotting, membranes were probed with anti-phospho-Y783-PLCγ1 antibody or anti-PLCγ1 monoclonal antibody (Cell Signaling). Probed membranes were washed and incubated with horseradish peroxidase-conjugated anti-mouse or anti-rabbit antibodies for 1 h and developed with Lumi-LightPLUS Western Blotting Substrate (Roche) on Lumi-Light autoradiography films or using an ImageQuant LAS 4000 Mini (GE Healthcare/Fujifilm). Quantification was performed using ImageJ software version 2.
NFAT transcriptional activation assay
For NFAT activation assays, JR.pTα cells48 were transfected with a luciferase reporter plasmid containing three tandem copies of the composite NFAT/AP1-response element from the human IL2 gene promoter, together with a pRL-CMV Renilla reporter construct (Promega). After 24 h, cells were cultured for 6 h with or without plate-bound anti-CD3ε (OKT3, Sigma-Aldrich) and lysed using the Dual luciferase assay Kit (Promega). Luciferase activity was normalized to the Renilla luciferase activity and expressed as fold-induction relative to the basal activity of non-stimulated cells48.
Anti-pTα monoclonal antibody and ADC in vivo administration
For in vivo administration of anti-pTα, SCID or NSG immunodeficient mice were i.v. injected with T-ALL3 (106 cells per mouse) or T-ALL42 (2 × 106 cells per mouse) cell samples, respectively, that were recovered from bone marrow or spleen xenografts of NSG mice. Mice were then serially i.p. injected (twice weekly) for 3–10 weeks with 200 μl of PBS containing either anti-pTα monoclonal antibody or IgG2a isotypic control (10 mg per kg body weight) starting when T-ALL engraftment in peripheral blood was >1% (week 3–4). Anti-pTα ADCs were generated with the support of the U2 Custom Antibody Service (CAbS) of the ICTS ‘NANBIOSIS’. Conjugation of the anti-pTα monoclonal antibody to DM1 was performed via a non-cleavable MCC linker, and to MMAE via a valine-citrulline protease-cleavable linker (SC-VC-PABC). The bioconjugation process involves the binding of the drug-linker complex to the lysines of the antibody through the succinimidyl group incorporated at the beginning of both linkers. Briefly, after stirring for 2 h at room temperature and overnight at 4 °C, the resulting mixture was desalted using a compact liquid chromatography system to purify the bioconjugate by using 10 mM PBS as elution buffer (10 mM sodium phosphate, 137 mM sodium chloride, 3.2 mM potassium chloride, 2 mM potassium dihydrogen phosphate, pH 7.4). For anti-pTα-MMAE treatment, xenotransplanted NSG mice (2 × 106 T-ALL42 cells per mouse) were i.p. injected with either anti-pTα-MMAE or IgG2a-MMAE (6 mg per kg body weight, twice weekly, for 2 weeks, and 3 mg per kg body weight for an additional week), starting when T-ALL engraftment in peripheral blood was >1% (week 5). For comparative treatments, xenotransplanted SCID mice (106 T-ALL3 cells per mouse) were i.p. injected with anti-pTα monoclonal antibody (10 mg per kg body weight), anti-pTα-DM1 (6 mg per kg body weight) or anti-pTα-MMAE (3 mg per kg body weight) or their corresponding isotype-matched controls (twice weekly, for 3 weeks), starting when T-ALL engraftment in peripheral blood was >1% (week 4). When indicated, a group of mice received a standard VxL induction treatment consisting of vincristine (Selleckchem; 0.15 mg per kg body weight, weekly for 3 weeks), plus dexamethasone (Sigma-Aldrich; 5 mg per kg body weight) and l-asparaginase (Kidrolase; EuSA Pharma; 1,000 U per kg body weight), during 5 days, for 3 weeks. For evaluation of treatment safety, NSG mice reconstituted with human cord blood HPCs were i.p. injected with anti-pTα-DM1 or IgG2a isotypic control (6 mg per kg body weight), twice a week for 3 weeks, starting at 9 or 21 weeks after transplant. Treated mice were analyzed at 2–3 weeks after treatment.
Pre-TCR labeling and internalization of pTα ADCs
SupT1 cells were incubated with 1 µg of anti-pTα monoclonal antibody or anti-pTα ADCs (anti-pTα-DM1 or anti-pTα-MMAE) at 4 °C for 30 min, washed in ice-cold PBS and incubated at 37 °C for the indicated times. Then, cells were washed with ice-cold PBS, and the primary antibody was detected using FITC-conjugated goat anti-mouse IgG (H + L) antibody (Invitrogen). Relative surface pre-TCR expression at each time point was calculated as the percentage of positive cells normalized to time 0.
Cell proliferation and cell cycle analysis
Cells were seeded at 2 × 105 cells per ml per well in 48-well plates in the absence or presence of 5 μg ml−1 of anti-pTα monoclonal antibody, anti-pTα-DM1, anti-pTα-MMAE or isotype-matched controls at 37 °C in 5% CO2. After 4 days, cells were collected, washed and counted. Cell proliferation was calculated as the percentage of total cells at the end of culture relative to input cell number. For cell cycle analysis, 4 × 105 cells were incubated with 5 μg ml−1 of Hoechst 33342 (Sigma) at 37 °C for 45 min. Then, cells were washed with ice-cold PBS and analyzed by flow cytometry.
Statistical analysis
Statistical analyses were performed with GraphPad Prism version 8. Statistical tests used for each experiment are indicated in the corresponding figure legends. In all cases, the confidence interval was 95%.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Source data are provided with this paper.
References
Pui, C. H., Robison, L. L. & Look, A. T. Acute lymphoblastic leukaemia. Lancet 371, 1030–1043 (2008).
Pui, C. -H., Carroll, W. L., Meshinchi, S. & Arceci, R. J. Biology, risk stratification, and therapy of pediatric acute leukemias: an update. J. Clin. Oncol. 29, 551–565 (2011).
Sutton, R. et al. Persistent MRD before and after allogeneic BMT predicts relapse in children with acute lymphoblastic leukaemia. Br. J. Haematol. 168, 395–404 (2015).
Litzow, M. R. & Ferrando, A. A. How I treat T-cell acute lymphoblastic leukemia in adults. Blood 126, 833–841 (2015).
Fielding, A. K. et al. Outcome of 609 adults after relapse of acute lymphoblastic leukemia (ALL); an MRC UKALL12/ECOG 2993 study. Blood 109, 944–950 (2006).
Clappier, E. et al. Clonal selection in xenografted human T cell acute lymphoblastic leukemia recapitulates gain of malignancy at relapse. J. Exp. Med. 208, 653–661 (2011).
Rafei, H., Kantarjian, H. M. & Jabbour, E. J. Recent advances in the treatment of acute lymphoblastic leukemia. Leuk. Lymphoma 60, 2606–2621 (2019).
Kantarjian, H. M. et al. Inotuzumab ozogamicin versus standard therapy for acute lymphoblastic leukemia. New Engl. J. Med. 375, 740–753 (2016).
Maude, S. L. et al. Chimeric antigen receptor T cells for sustained remissions in leukemia. N. Engl. J. Med. 371, 1507–1517 (2014).
Scherer, L. D., Brenner, M. K. & Mamonkin, M. Chimeric antigen receptors for T-cell malignancies. Front. Oncol. 9, 126 (2019).
Bayón-Calderón, F., Toribio, M. L. & González-García, S. Facts and challenges in immunotherapy for T-cell acute lymphoblastic leukemia. Int. J. Mol. Sci. 21, 7685 (2020).
Notarangelo, L. D., Kim, M. -S., Walter, J. E. & Lee, Y. N. Human RAG mutations: biochemistry and clinical implications. Nat. Rev. Immunol. 16, 234–246 (2016).
Belver, L. & Ferrando, A. The genetics and mechanisms of T cell acute lymphoblastic leukaemia. Nat. Rev. Cancer 16, 494–507 (2016).
Bene, M. C. et al. Proposals for the immunological classification of acute leukemias. European Group for the Immunological Characterization of Leukemias (EGIL). Leukemia 9, 1783–1786 (1995).
Pölönen, P. et al. The genomic basis of childhood T-lineage acute lymphoblastic leukaemia. Nature 632, 1082–1091 (2024).
Radtke, F. et al. Deficient T cell fate specification in mice with an induced inactivation of Notch1. Immunity 10, 547–558 (1999).
Weng, A. P. et al. Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia. Science 306, 269–271 (2004).
Grabher, C., von Boehmer, H. & Look, A. T. Notch 1 activation in the molecular pathogenesis of T-cell acute lymphoblastic leukaemia. Nat. Rev. Cancer 6, 347–359 (2006).
Kindler, T. et al. K-RasG12D-induced T-cell lymphoblastic lymphoma/ leukemias harbor Notch1 mutations and are sensitive to gamma-secretase inhibitors. Blood 112, 3373–3382 (2008).
Armstrong, F. et al. NOTCH is a key regulator of human T-cell acute leukemia initiating cell activity. Blood 113, 1730–1740 (2009).
Tatarek, J. et al. Notch1 inhibition targets the leukemia-initiating cells in a Tal1/Lmo2 mouse model of T-ALL. Blood 118, 1579–1590 (2011).
Toribio, M. L. & González-García, S. Notch partners in the long journey of T-ALL pathogenesis. Int. J. Mol. Sci. 24, 1383 (2023).
Reizis, B. & Leder, P. Direct induction of T lymphocyte-specific gene expression by the mammalian Notch signaling pathway. Genes Dev. 16, 295–300 (2002).
Saint-Ruf, C. et al. Analysis and expression of a cloned pre-T cell receptor gene. Science 266, 1208–1212 (1994).
Ramiro, A. R., Trigueros, C., Marquez, C., San Millan, J. L. & Toribio, M. L. Regulation of pre-T cell receptor (pT alpha-TCR beta) gene expression during human thymic development. J. Exp. Med. 184, 519–530 (1996).
von Boehmer, H. & Fehling, H. J. Structure and function of the pre-T cell receptor. Annu. Rev. Immunol. 15, 433–452 (1997).
Dudley, E. C., Petrie, H. T., Shah, L. M., Owen, M. J. & Hayday, A. C. T cell receptor β chain gene rearrangement and selection during thymocyte development in adult mice. Immunity 1, 83–93 (1994).
Fehling, H. J., Krotkova, A., Saint-Ruf, C. & von Boehmer, H. Crucial role of the pre-T-cell receptor α gene in development of αβ but not γδ T cells. Nature 375, 795–798 (1995).
Trigueros, C. et al. Identification of a late stage of small noncycling pTalpha- pre-T cells as immediate precursors of T cell receptor alpha/beta+ thymocytes. J. Exp. Med. 188, 1401–1412 (1998).
Allman, D. et al. Separation of Notch1 promoted lineage commitment and expansion/transformation in developing T cells. J. Exp. Med. 194, 99–106 (2001).
Bellavia, D. et al. Combined expression of pTα and Notch3 in T cell leukemia identifies the requirement of preTCR for leukemogenesis. Proc. Natl Acad. Sci. USA 99, 3788–3793 (2002).
Tremblay, M. et al. Modeling T-cell acute lymphoblastic leukemia induced by the SCL and LMO1 oncogenes. Genes Dev. 24, 1093–1105 (2010).
Dos Santos, N. R. et al. Pre-TCR expression cooperates with TEL-JAK2 to transform immature thymocytes and induce T-cell leukemia. Blood 109, 3972–3981 (2007).
Fasseu, M. et al. p16INK4A tumor suppressor gene expression and CD3ϵ deficiency but not pre-TCR deficiency inhibit TAL1-linked T-lineage leukemogenesis. Blood 110, 2610–2619 (2007).
Campese, A. F. et al. Notch1-dependent lymphomagenesis is assisted by but does not essentially require pre-TCR signaling. Blood 108, 305–310 (2006).
Reschly, E. J. et al. Notch1 promotes survival of E2A-deficient T cell lymphomas through pre-T cell receptor-dependent and -independent mechanisms. Blood 107, 4115–4121 (2006).
Asnafi, V. et al. Analysis of TCR, pTα, and RAG-1 in T-acute lymphoblastic leukemias improves understanding of early human T-lymphoid lineage commitment. Blood 101, 2693–2703 (2003).
Gocho, Y. et al. Network-based systems pharmacology reveals heterogeneity in LCK and BCL2 signaling and therapeutic sensitivity of T-cell acute lymphoblastic leukemia. Nat Cancer 2, 284–299 (2021).
Courtois L. et al. Surface pTα expression predicts LCK activation and preclinical synergy of LCK and JAK coinhibition in adult T-ALL. Blood 145, 2903–2913 (2025).
García-Peydró, M. et al. The NOTCH1/CD44 axis drives pathogenesis in a T cell acute lymphoblastic leukemia model. J. Clin. Invest. 128, 2802–2818 (2018).
Ramiro, A. R. et al. Differential developmental regulation and functional effects on pre-TCR surface expression of human pTalpha(a) and pTalpha(b) spliced isoforms. J. Immunol. 167, 5106–5114 (2001).
Pear, W. S. et al. Exclusive development of T cell neoplasms in mice transplanted with bone marrow expressing activated Notch alleles. J. Exp. Med. 183, 2283–2291 (1996).
Lapidot, T. et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 367, 645–648 (1994).
Monaco, G. et al. RNA-seq signatures normalized by mRNA abundance allow absolute deconvolution of human immune cell types. Cell Rep. 26, 1627–1640 (2019).
Carrasco, Y. R., Navarro, M. N. & Toribio, M. L. A role for the cytoplasmic tail of the pre-T cell receptor (TCR) alpha chain in promoting constitutive internalization and degradation of the pre-TCR. J. Biol. Chem. 278, 14507–14513 (2003).
Michie, A. M., Trop, S., Wiest, D. L. & Zúñiga-Pflücker, J. C. Extracellular signal–regulated kinase (Erk) activation by the pre-T cell receptor in developing thymocytes in vivo. J. Exp. Med. 190, 1647–1656 (1999).
Hinton, H. J., Clarke, R. G. & Cantrell, D. A. Antigen receptor regulation of phosphoinositide-dependent kinase 1 pathways during thymocyte development. FEBS Lett. 580, 5845–5850 (2006).
Navarro, M. N. et al. Identification of CMS as a cytosolic adaptor of the human pTalpha chain involved in pre-TCR function. Blood 110, 4331–4340 (2007).
Aifantis, I., Gounari, F., Scorrano, L., Borowski, C. & von Boehmer, H. Constitutive pre-TCR signaling promotes differentiation through Ca2+ mobilization and activation of NF-κB and NFAT. Nat. Immunol. 2, 403–409 (2001).
Fu, G. et al. Phospholipase Cγ1 is required for pre-TCR signal transduction and pre-T cell development. Eur. J. Immunol. 47, 74–83 (2017).
Blanco, R., Borroto, A., Schamel, W., Pereira, P. & Alarcon, B. Conformational changes in the T cell receptor differentially determine T cell subset development in mice. Sci. Signal. 7, ra115 (2014).
Verma, M. K. et al. A novel hemolytic complement-sufficient NSG mouse model supports studies of complement-mediated antitumor activity in vivo. J. Immunol. Methods 446, 47–53 (2017).
Drago, J. Z., Modi, S. & Chandarlapaty, S. Unlocking the potential of antibody–drug conjugates for cancer therapy. Nat. Rev. Clin. Oncol. 18, 327–344 (2021).
Carrasco, Y. R. et al. An endoplasmic reticulum retention function for the cytoplasmic tail of the human pre-T cell receptor (TCR) alpha chain: potential role in the regulation of cell surface pre-TCR expression levels. J. Exp. Med. 193, 1045–1058 (2001).
Sánchez-Martínez, D. et al. Fratricide-resistant CD1a-specific CAR T cells for the treatment of cortical T-cell acute lymphoblastic leukemia. Blood 133, 2291–2304 (2019).
Maciocia, P. M. et al. Anti-CCR9 chimeric antigen receptor T cells for T-cell acute lymphoblastic leukemia. Blood 140, 25–37 (2022).
Trinquand, A. et al. Triggering the TCR developmental checkpoint activates a therapeutically targetable tumor suppressive pathway in T-cell leukemia. Cancer Discov. 6, 972–985 (2016).
Nichakawade, T. D. et al. TRBC1-targeting antibody–drug conjugates for the treatment of T cell cancers. Nature 628, 416–423 (2024).
Haynes, B. F. et al. Analysis of the adult thymus in reconstitution of T lymphocytes in HIV-1 infection. J. Clin. Invest. 103, 453–460 (1999).
González-García, S. et al. IL-7R is essential for leukemia-initiating cell activity of T-cell acute lymphoblastic leukemia. Blood 134, 2171–2182 (2019).
Palomero, T. et al. CUTLL1, a novel human T-cell lymphoma cell line with t(7;9) rearrangement, aberrant NOTCH1 activation and high sensitivity to gamma-secretase inhibitors. Leukemia 20, 1279–1287 (2006).
Ogando, J. et al. Notch-regulated miR-223 targets the aryl hydrocarbon receptor pathway and increases cytokine production in macrophages from rheumatoid arthritis patients. Sci. Rep. 6, 20223 (2016).
Seet, C. S. et al. Generation of mature T cells from human hematopoietic stem and progenitor cells in artificial thymic organoids. Nat. Methods 14, 521–530 (2017).
Aster, J. C. et al. Essential roles for ankyrin repeat and transactivation domains in induction of T-cell leukemia by Notch1. Mol. Cell. Biol. 20, 7505–7515 (2000).
van Rijn, R. S. et al. A new xenograft model for graft-versus-host disease by intravenous transfer of human peripheral blood mononuclear cells in RAG2−/− γc−/− double-mutant mice. Blood 102, 2522–2531 (2003).
Hu, Y. & Smyth, G. K. ELDA: Extreme limiting dilution analysis for comparing depleted and enriched populations in stem cell and other assays. J. Immunol. Methods 347, 70–78 (2009).
Acknowledgements
We thank J. C. Aster (Department of Pathology, Brigham and Women’s Hospital), K. Weijer and H. Spits (Department of Experimental Immunology, Academic Medical Center of the University of Amsterdam), Gay M. Crooks (Molecular Biology Institute, UCLA); A. Ferrando (Institute for Cancer Genetics, Columbia University), J. C. Zúñiga-Pflücker (Department of Immunology, University of Toronto & Sunnybrook Research Institute); and P. Menéndez and C. Bueno (Josep Carreras Leukemia Research Institute) for providing useful reagents, cell samples and mouse strains. We thank E. Castillo for technical support, the Animal Facility and the Flow Cytometry Core at the Centro de Biología Molecular Severo Ochoa and the Mouse Embryo Cryopreservation Facility at CNB-CSIC for their invaluable assistance; and the Centro de Transfusión de la Comunidad de Madrid and the Pediatric Cardiosurgery Unit, Hospital Universitario 12 de Octubre (Madrid) for cord blood and thymus samples, respectively. This research was supported in part by grants PID2019-105623RB-I00, PID2022-138880OB-I00 and PDC2021-121238-I00 funded by Ministerio de Ciencia, Innovación y Universidades. Agencia Estatal de Investigación (MCIN/AEI/ 10.13039/501100011033/European Regional Development Fund, European Union), and by grants from Fundación Unoentrecienmil and Fundación Inocente Inocente to M.L.T. Institutional grants from the Fundación Ramón Areces and Banco de Santander to the Centro de Biología Molecular Severo Ochoa are also acknowledged. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
M.L.T. and P.F. conceived and designed the project, supervised experiments and wrote the paper. P.F., M.G.-P., J.A., M.M., C. Cela, B.A. and C. Cifuentes performed and analyzed experiments; M.C., M.T., I.I. and M.R. performed clinical evaluations of individuals and M.R. provided human samples.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks Iannis Aifantis, Laura Belver, Paul Love and Vinothkumar Rajan for their contribution to the peer review of this work. Primary Handling Editor: P. Jauregui, in collaboration with the Nature Immunology team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Expression of pre-TCR in human T-ALLs.
(A) qPCR analysis of PTCRA mRNA expression normalized to GAPDH expression of T-ALL patient samples recovered from spleen xenografts of NSG mice (n = 22, Table 1). Data are shown as mean of technical triplicates relative to values derived from healthy PB T cell samples (n = 2). Dashed line shows relative PTCRA expression in the pre-TCR+ SupT1 cell line included as positive control. (B-E) Normalized PTCRA expression values derived from public RNA-seq raw data of a cohort of >1300 T-ALL patient samples15 and >100 B, T and myeloid PB cells from healthy donors44. Data are shown as normalized counts +1 either from healthy B, T, and myeloid peripheral blood cells, and total T-ALL samples (B) or from T-ALL samples (C-E) defined by ETP status (C, ****P < 0.0001), genetic subtypes grouped by their similarity to the expression signature of control haematopoietic cells (D, ****P < 0.0001), and TRA, TRB, TRG and TRD gene rearrangements (E, ****P < 0.0001). Dashed line shows cut-off value for PTCRA expression. Median and quartiles values within violin plots are indicated by solid and dashed lines, respectively. Data were analysed by one-way ANOVA with Dunn’s multiple comparison test.
Extended Data Fig. 2 Expression of surface pre-TCR in T-ALL cell lines and primary samples.
(A) Representative pseudocolour plots of CD3 vs pTα expression in pre-TCR-expressing SupT1, and TCRαβ-expressing HPB-ALL and Jurkat human cell lines from three independent experiments, as assessed by reactivity with anti-CD3ε and anti-pTα monoclonal antibodies (left biparametric histograms). Representative monoparametric histograms on the right show surface pre-TCR expression levels analysed with anti-pTα monoclonal antibody plus goat anti-mouse FITC. Numbers indicate percentages of positive cells. Shaded histograms show background staining with an irrelevant isotype-matched antibody. (B) Representative CD3 vs TCRαβ expression of either primary human thymocytes from a single 17-weeks-old fetal thymus sample (upper left), or developing T cells derived either from two independent human CB HPC samples after 70 days of culture onto OP9-Jag2 cells (middle left), or from a single CD34+ human ETP sample after 18 days of culture in ATOs (bottom left). Pre-TCR expression levels of electronically gated CD3lo TCRαβneg, TCRαβ+ and CD3neg cells analysed with anti-pTα mAb plus biotin-coupled anti-mouse (Fab’)2 plus BV421-coupled streptavidin are shown in the monoparametric histograms on the right. Numbers indicate percentages of positive cells relative to background staining with an isotype-matched mAb.
Extended Data Fig. 3 T-ALL patient cells arrested at the β-selection checkpoint express barely detectable levels of a functional pre-TCR.
(A) Flow cytometry of CD3 vs TCRαβ expression of T-ALL patient samples representative of developmental arrest at the β-selection checkpoint (T-ALL42, T-ALL43), recovered from spleen xenografts of NSG mice from two independent experiments (left biparametric histograms). Monoparemetric histograms on the right show pre-TCR surface expression on electronically gated CD3lo TCRneg cells analysed with anti-pTα monoclonal antibody plus biotin-coupled anti-mouse (Fab’)2 plus PE-coupled streptavidin. Shaded histograms represent background staining with an irrelevant isotype-matched control. (B) Immunoblot analysis of ERK and S6 phosphorylation in pre-TCR+ T-ALLs (T-ALL42, T-ALL43) stimulated either with anti-pTα or with anti-CD3ε monoclonal antibodies for the indicated times from a single experiment. One experiment per T-ALL sample was performed. Total ERK and S6 expression is shown as loading control.
Extended Data Fig. 4 In vitro and in vivo abrogation of pre-TCR function by silencing of CD2AP gene encoding the CMS functional adaptor of pTα.
(A) Immunoblot analysis of CMS expression in SupT1 cells that were either transfected with several shRNAs (shCMS1-5) specific for CD2AP, the gene encoding CMS, or left untransfected (NT) (left panel). Relative CMS expression normalized to α-tubulin expression used as loading control is shown in the right panel. (B) Immunoblot analysis of CMS expression (upper) and PLCγ tyrosine (Tyr) phosphorylation (bottom) in JR.pTα pre-TCR+ cells transduced either with shCMS4 or with shSC as control, prior to activation with an anti-CD3ε monoclonal antibody for the indicated times. PLCγ Tyr-phosphorylation was analysed after immunoprecipitation with anti-PLCγ1 monoclonal antibody by probing with an anti-Y783-PLCγ1 antibody. Tubulin expression was analysed as loading control. (C) Relative NFAT activity of JR.pTα cells transduced with either shCMS4 or shSC as control, analysed upon stimulation with an anti-CD3ε monoclonal antibody by a luciferase assay. Data are shown as mean percentages ± SEM of NFAT activity relative to activated shSC cells analysed by two-tailed unpaired t test (n = 3). ****P < 0.0001. (D) Immunoblot analysis of CMS in human CD34+ early thymic progenitors (ETPs) transduced with a lentivirus encoding either shCMS4 and GFP or shSC and GFP as control. (E) Absolute numbers of shCMS- or shSC-transduced ETP-derived human cells reconstituting the thymus of RAG-2−/−γc−/− mice at the indicated weeks post-transplant (n = 6 week 3, n = 4 week 5; ***P = 0.0002). ETP transduction efficiencies with shCMS and shSC were 24,5% ± 4,5% and 21,5% ± 3,5%, respectively. (F) Absolute numbers of shCMS- or shSC-transduced human thymocytes in (E) expressing the double positive CD3lo TCRαβ− phenotype at 3- and 5-weeks post-transplant (left panel; *P = 0.0222) or the post-β selected DP CD3+ TCRαβ+ phenotype at 5-weeks post-transplant (right panel; *P = 0.0146). Data in (E, F) are shown as mean numbers ± s.e.m. of transduced (GFP+) cells normalized to 105 transduced input cells from two independent experiments, analysed by two-tailed unpaired t test.
Extended Data Fig. 5 ICN1 is unable to induce T-ALL generation in C80G mutant mice deficient in pre-TCR signalling.
(A) Schematic diagram of experimental design for mouse T-ALL generation. Sublethally irradiated NSG (2H-Kd+) mice were subjected to i.v. injection with bone marrow lin− c-kit+ HPCs isolated from wild type (wt) or CD3ε C80G mutant C57BL/6 (H2-Kb+) mice and transduced with a lentiviral vector encoding either ICN1 and GFP or GFP alone as control. (B) Representative flow cytometry analysis of transduction efficiencies of wt (upper) and mutant (bottom) HPCs in (A). Numbers indicate percentages of transduced (GFP+) cells. (C) Representative CD8 and CD4 expression of electronically-gated mock-transduced (gate I; ICN1−) and ICN1-transduced (gate II; ICN1+) cells, derived from donor wt (upper) or C80G mutant (bottom) bone marrow HPCs engrafting the bone marrow of NSG mice at 5 weeks post-transplant. Results are representative of one out of at least 9 mice per group from two independent experiments. (D) Mean ± s.e.m. percentages of myeloid-lineage (CD11b+ or Gr1.1+), B-lineage (B220+), T-lineage (double positive; CD4+ CD8− and/or CD8+ CD4−) and Lin− (CD11b−, Gr1.1−, B220−, CD4−, CD8−) cells derived from ICN1+ (n = 5) and ICN1− (n = 4) wt or C80G (n = 5, each) mutant mouse bone marrow HPCs engrafting the bone marrow of NSG mice in (C). (E) Image of representative spleens obtained from mice shown in (C, D). (F) Mean percentages ± s.e.m. of ICN1+ cells derived from wt or C80G mutant HPCs engrafting the indicated organs of NSG mice (9 mice per group) at sacrifice, analysed by two-tailed unpaired t test. **P = 0.0022 (PB), ***P = 0.0005 (BM), ***P = 0.0003 (spleen), ****P < 0.0001 (liver), and *P = 0.0106 (thymus). (G) Kaplan-Meier survival curves of transplanted NSG mice in (F) were compared using Log-rank Mantle-Cox test. ****P < 0.0001.
Extended Data Fig. 6 Anti-pTα ADCs induce pre-TCR internalization.
(A) Surface pre-TCR expression levels detected on SupT1 and JR.pTα pre-TCR+ cell lines by flow cytometry, using anti-CD3ε in combination with either drug-free anti-pTα monoclonal antibody, or anti-pTα-DM1 or anti-pTα-MMAE ADCs plus biotin-coupled anti-mouse (Fab’)2 plus PECy7-coupled streptavidin. Pre-TCR-negative Jurkat (TCRαβ+) and mock-transduced JR.MigR1 (TCRα−) cell lines were assayed as negative controls. (B) Pre-TCR internalization in SupT1 cells pulsed with drug-free anti-pTα monoclonal antibody or anti-pTα ADCs at 4 °C for 30 min and incubated for the indicated times at 37 °C. Results are shown as mean percentages ±s.e.m. of pre-TCR+ cells detected by flow cytometry with a secondary anti-mouse IgG-FITC antibody normalized to time 0, (n = 3).
Extended Data Fig. 7 Anti-pre-TCR ADCs have no deleterious effect on normal T cell development.
(A) Schematic diagram of experimental design for anti-pTα-ADC treatment of humanized mice. NSG mice transplanted with human CB HPCs received i.p. injections of either DM1-conjugated anti-pTα monoclonal antibody or a DM1-conjugated isotype-matched control, twice a week for 3 weeks, starting at 9 (B-D; n = 5 mice per group) or 21 (E-G; n = 3 mice per group) weeks post-transplant. (B, E) Kinetics of human T (left panels) and B cell (right panels) reconstitution of peripheral blood of mice treated as shown in (A). (C, F). Percentages of T and B cells among human CD45+ cells recovered from the spleens of treated mice at week 2 (C) or week 3 (F) post-treatment. (D, G) Percentages of human T-cell progenitors expressing the pre-TCR, or placed up (pre-pre-TCR) or downstream (post-pre-TCR) of pre-TCR expression, recovered from the thymus of treated mice at week 2 (D) or week 3 (G) post-treatment. Data are shown as mean ± s.e.m. cell percentages analysed by two-way ANOVA with Sidak’s multiple comparison test: ordinary (C, D, F, G) or with Geisser-Greenhouse correction (B and E).
Supplementary information
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Unmodified gels
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 3
Unmodified gels.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 4
Unmodified gels.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fuentes, P., García-Peydró, M., Alcain, J. et al. Pre-TCR-targeted immunotherapy for T cell acute lymphoblastic leukemia. Nat Immunol 26, 1712–1725 (2025). https://doi.org/10.1038/s41590-025-02265-w
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41590-025-02265-w
This article is cited by
-
The pre-T cell receptor as a tumor immunotherapy target in T cell leukemia
Nature Immunology (2025)
-
Slaying the pre-TCR beast
Nature Reviews Cancer (2025)
