
Abstract
Glucosamine (GlcN) is a common supplement used to alleviate osteoarthritis, but it may dysregulate glucose tolerance and induce insulin resistance, thereby increasing metabolic burden. The liver is a vital organ that modulates the Akt/mTOR/p70S6K signaling pathway in response to growth and metabolism. Fibroblast growth factor 21 (FGF21) is a hepatokine involved in regulating glucose and lipid metabolism. Additionally, increased circulating FGF21 levels have been linked to the prediction of metabolic disorders and type 2 diabetes. However, the regulatory mechanism controlling FGF21 expression by GlcN remains unclear. In the present study, GlcN stimulation led to several outcomes, including an increase in cell content, secretion, and mRNA and protein levels of FGF21 in hepatocytes. Moreover, inhibition of the Akt/mTOR/p70S6K axis resulted in reduced FGF21 expression in response to GlcN. Importantly, GlcN-mediated expression of FGF21 relies on PGC-1α upregulation. These results suggest that GlcN increases FGF21 expression through the activation between Akt/mTOR/p70S6K pathway and PGC-1α dependent manner.
Similar content being viewed by others
Introduction
Glucosamine (GlcN) is one of the most widely used dietary supplements, known for its beneficial pharmacological effects in alleviating osteoarthritis1. Additionally, several therapeutic applications of GlcN have been reported, including its use in treating skin disorders, cardiovascular diseases, kidney toxicity, and even cancer2. However, its widespread use raises potential concerns about safety and toxicity. Clinical data suggest that GlcN may have adverse effects on glucose metabolism3,4. Further in vivo and in vitro studies have shown that GlcN induces insulin resistance and increases plasma glucose levels5,6.
Type 2 diabetes is one of the most common metabolic disorders, caused by insulin resistance and hyperglycemia. Excess glucose is converted into GlcN via the hexosamine biosynthetic pathway, leading to dysregulation of insulin signaling and promoting the development of metabolic disorders7,8. These findings suggest that GlcN synthesis may play a role in the progression of diabetes. Additionally, chronic hyperglycemia disrupts hepatic glucose metabolism, leading to non-alcoholic fatty liver disease (NAFLD). It is estimated that 50–75% of diabetic patients suffer from NAFLD, which increases the risk of cardiovascular diseases and premature mortality9.
Among the members of the fibroblast growth factor (FGF) family, FGF21 is an atypical subfamily member that can be released into circulation, acting in an endocrine manner. Generally, circulating FGF21 is primarily derived from the liver. During fasting, peroxisome proliferator-activated receptor α (PPARα) activates FGF21 expression, which stimulates fatty acid oxidation and gluconeogenesis in the liver10. Conversely, FGF21 levels are elevated in conditions such as obesity, diabetes, and NAFLD11. Thus, FGF21 is considered a crucial metabolic regulator, helping to counteract stress from pathological conditions. Indeed, FGF21 has been shown to improve insulin sensitivity and lipid profiles through various signaling pathways12,13. The Akt/mTOR signaling pathway is considered a central controller of cell growth and metabolism. This pathway mediates hypoxia-inducible factor 1α (HIF-1α) expression by phosphorylating ribosomal protein S6 kinase (p70S6K), thereby regulating glucose metabolism14. Furthermore, hyperactivation of the mTOR signaling network promotes FGF21 expression via peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α) in the liver15.
Although previous study has reported that GlcN can activate FGF21 expression both in vitro and in vivo16, the association between the Akt/mTOR signaling network and the activity of transcriptional regulators in GlcN-induced FGF21 expression remains unclear. Therefore, in the present study, we measured the effect of GlcN on FGF21 expression in hepatocytes. Additionally, we used various inhibitors to explore the potential signaling pathways involved in GlcN’s actions on hepatic FGF21 expression.
Results
GlcN increased FGF21 expression in AML12 hepatocytes in a dose- and time-dependent manner
To investigate the response of FGF21 expression to GlcN treatment, confluent AML12 cells were incubated with 5, 10, 15, 20, or 25 mM GlcN for 24 h. The results showed that both the cell content and secretion levels of FGF21 increased significantly in a dose-dependent manner (Fig. 1a, b). Similarly, GlcN treatment promoted FGF21 mRNA and protein levels in a dose-dependent manner in hepatocytes (Fig. 1c, d).

GlcN increased FGF21 expression in a dose-dependent manner. AML12 cells were incubated with varying concentrations of GlcN (0, 5, 10, 15, 20, 25 mM) for 24 h. FGF21 expression was analyzed in terms of (a) cell content, (b) secretion, (c) mRNA, and (d) protein levels. Data are presented as mean ± SEM. *P < 0.05 compared with control (n = 3).
The effect of GlcN on FGF21 expression was evaluated at several time points (2, 4, 8, 12, and 24 h). As shown in Fig. 2a b, GlcN-treated cells (10 mM) exhibited a significant, time-dependent increase in both the cell content and secretion of FGF21 from 2 to 24 h. Consistent with these findings, GlcN treatment also significantly increased FGF21 mRNA levels, with quantitative data indicating a 1.4-fold increase at 4 h and a 3.5-fold increase at 24 h (Fig. 2c).

GlcN increased FGF21 expression in a time-dependent manner. AML12 cells were treated with GlcN (10 mM) for the indicated times (0, 2, 4, 8, 12, and 24 h). FGF21 expression was analyzed in terms of (a) cell content, (b) secretion, and (c) mRNA levels. Data are presented as mean ± SEM. *P < 0.05 compared with control (n = 3).
GlcN increased FGF21 expression through the activation of the Akt/mTOR/ p70S6K pathway
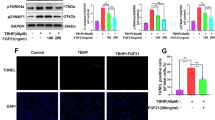
To confirm the involvement of the signaling pathway, AML12 hepatocytes were treated with LY294002 (an Akt inhibitor), rapamycin (an mTOR inhibitor), or PF4708671 (a p70S6K inhibitor) before incubation with GlcN (10 mM) for 24 h, after which FGF21 expression was measured. The results indicate that pretreatment with these inhibitors largely prevented the GlcN-induced increase in FGF21 expression, including cell content (Fig. 3a), secretion (Fig. 3b), mRNA levels (Fig. 3c), and protein levels (Fig. 3d).

GlcN induced FGF21 expression via the Akt/mTOR/p70S6K pathway. To block Akt/mTOR/p70S6K signaling, cells were pretreated with LY294002 (10− 5 M), rapamycin (10− 7 M), or PF4708671 (10− 5 M) for 1 h, followed by treatment with 10 mM GlcN for 24 h. FGF21 expression was assessed in terms of (a) cell content, (b) secretion, (c) mRNA, and (d) protein levels. Data are presented as mean ± SEM. *P < 0.05 compared with control; #P < 0.05 compared with GlcN alone (n = 3).
PGC1-α is involved in the regulation of FGF21 expression mediated by GlcN
To further elucidate the role of transcriptional regulators in GlcN-mediated FGF21 production, cells were pretreated with several inhibitors, including GW6471 (a PPARα antagonist), GW9662 (a PPARγ antagonist), KC7F2 (a HIF-1α inhibitor), and SR18292 (a PGC-1α inhibitor). Notably, SR18292-treated cells displayed reduced FGF21 protein expression upon GlcN stimulation (Fig. 4a). Additionally, an examination of PGC-1α activity confirmed that the PGC-1α activator ZLN005 significantly elevated FGF21 protein expression in a dose-dependent manner (Fig. 4b). Furthermore, results of western blot analysis showed that GlcN-treated cells exhibited increased PGC-1α protein expression compared to the vehicle control (Fig. 4c).

GlcN induced FGF21 expression mediated through PGC-1α. Cells were pretreated with various transcriptional regulator inhibitors (GW6471, GW9662, KC7F2, or SR18292; each at 10− 5 M) for 1 h, followed by exposure to GlcN (10 mM) for 24 h, or co-incubated with different concentrations of ZLN005 (0, 2, 4, 6, 8, 10 µM) for 24 h. (a, b) FGF21 protein levels were measured. Hepatocytes were incubated with GlcN (10 mM) for 24 h. (c) PGC-1α expression was assessed by western blot. Data are presented as mean ± SEM. *P < 0.05 compared with control; #P < 0.05 compared with GlcN alone (n = 3).
Expression of hepatic FGF21 and FASN in GlcN-infused mice
To verify the in vitro findings, we examined changes in FGF21 levels in plasma and liver tissues from mice infused with GlcN. After 2 weeks of GlcN administration, a significant increase in plasma and hepatic FGF21 levels was observed in GlcN-infused mice compared to saline-infused controls (Fig. 5a, b). In addition to inducing FGF21 expression, GlcN upregulated hepatic FASN expression (Fig. 5c). Furthermore, we assessed the effect of GlcN on intracellular lipid accumulation in AML12 cells. Oil Red O staining revealed that GlcN further enhanced OA-induced lipid accumulation in hepatocytes (Fig. 5d). These findings suggest that GlcN may potentiate lipid accumulation by upregulating FASN expression in hepatocytes.

GlcN infusion increased plasma levels and hepatic expression of FGF21, as well as lipogenesis-related genes in mice. C57BL/6J mice were infused with GlcN or saline for 2 weeks. (a, b) FGF21 levels in plasma and liver were measured. (c) Hepatic expression of FASN was assessed (n = 6). (d) AML12 cells were co-treated with OA and GlcN for 24 h, followed by quantification of Oil Red O staining (n = 3). Scale bar: 100 μm. Data are presented as mean ± SEM. *P < 0.05 compared with control.
Discussion
In the present study, we demonstrate that GlcN induces FGF21 expression in a dose- and time-dependent manner. FGF21 is a key regulator in improving lipid homeostasis and insulin sensitivity17. Interestingly, systemic inflammation triggered by obesity and insulin resistance also promotes FGF21 expression in patients with type 2 diabetes18. However, caution is advised when using GlcN in patients with type 2 diabetes, as studies have reported that GlcN can disrupt insulin secretion by pancreatic β-cells19. Additionally, GlcN administration may impair glucose tolerance in both rodents and humans3,4,6. Therefore, our findings suggest that FGF21 upregulation under GlcN stimulation may play a role in protecting against the burden of increased metabolic demands.
The Akt/mTOR/p70S6K signaling network plays an essential regulatory role in protein synthesis and cellular metabolism. However, dysregulation of mTOR signaling has been implicated in obesity and type 2 diabetes20. To gain insight into how GlcN regulates FGF21 expression, we demonstrated that the upregulation of FGF21 by GlcN depends on the activation of the Akt/mTOR/p70S6K axis in hepatocytes. These findings align with a previous study by Cornu et al., which indicated that ectopic activation of mTOR promotes FGF21 expression in the liver. This study also reported that FGF21 expression is associated with an increase in PGC-1α levels15.
We then investigated whether GlcN regulates FGF21 production through the activity of transcriptional regulators. The GlcN-induced activation of FGF21 expression was significantly reduced with PGC-1α inhibition, whereas the activation of PGC-1α led to a marked increase in FGF21 expression. As expected, GlcN significantly elevated PGC-1α levels, aligning with previous study showing that GlcN stimulation enhances PGC-1α expression in hepatocytes7. This raises the question, how does the Akt/mTOR/p70S6K axis activate PGC-1α, thereby promoting FGF21 production? Previous study has reported that ectopic activation of hepatic mTOR leads to increased PGC-1α expression, and that PGC-1α deficiency downregulates FGF21 production15. These findings suggest that GlcN induces an interaction between the Akt/mTOR/p70S6K network and PGC-1α, a key transcription factor regulating FGF21. Therefore, using genetic knockdown of PGC-1α would further strengthen our findings.
Consistent with our in vitro observations, both blood levels and hepatic expression of FGF21 were specifically increased in GlcN-infused mice. Interestingly, GlcN infusion also led to the upregulation of hepatic FASN. Furthermore, the increased lipid accumulation in hepatocytes can be directly attributed to GlcN administration. FGF21 is known to play a regulatory role in hepatic lipid metabolism21. Previous studies have shown that FGF21 induces reductions in triglyceride levels and lipogenesis-related gene expression, while promoting lipid oxidation and transport in hepatocytes22. Among the mechanisms involved in GlcN-induced FGF21 expression, the Akt/mTOR/p70S6K axis appears to be a key regulatory pathway. Additionally, in macrophages, GlcN-mediated mTOR/p70S6K activation has been shown to regulate acetyl-CoA and FASN expression, promoting lipid accumulation23. Collectively, these findings suggest that GlcN stimulates the Akt/mTOR/p70S6K axis and PGC-1α activity, ultimately promoting FGF21 expression and modulating lipogenesis.
In conclusion, our study is the first to elucidate the molecular mechanism underlying the effect of GlcN on FGF21 expression, as summarized in Fig. 6. We observed that GlcN administration activates the Akt/mTOR/p70S6K axis in hepatocytes, which in turn activates the transcriptional coactivator PGC-1α, ultimately leading to an increase in the expression of the stress-responsive hormone FGF21.

Proposed mechanism illustrating the effect of GlcN on hepatic FGF21 expression. GlcN induces activation of the Akt/mTOR/p70S6K axis and stimulates PGC-1α activity, leading to upregulated hepatic FGF21 expression and increased circulating FGF21 levels. (Created in BioRender.com).
Limitations
In the present study, we demonstrated that exogenous infusion of GlcN led to a significant increase in hepatic FGF21 expression and lipid accumulation. It has been reported that chronic metabolic disorders and stress-related burdens can provoke FGF21 resistance, thereby disrupting the healthy aging process24. However, our use of young and healthy mice does not reflect age-related metabolic changes. Additionally, the absence of diet-induced obesity models limits our ability to fully understand FGF21’s role within the context of obesity. Therefore, to achieve a more comprehensive understanding, future experimental designs should incorporate assessments of GlcN’s effects in aging and obesity models, which more accurately mimic human aging and metabolic conditions.
Materials and methods
Materials
Dulbecco’s modified Eagle medium (DMEM), fetal bovine serum (FBS), penicillin, and streptomycin were obtained from GIBCO BRL, Life Technologies Inc. (Grand Island, NY, USA). LY294002 was purchased from Tocris Bioscience (1130, Minneapolis, MN, USA). GW6471 (11697), GW9662 (70785), KC7F2 (14123), PF4708671 (15018), rapamycin (13346), SR18292 (22084), and ZLN005 (14121) were obtained from Cayman Chemicals (Ann Arbor, MI, USA). The FGF21 antibody was purchased from Abcam (Cambridge, UK), while the antibodies for fatty acid synthase (FASN) and GAPDH were purchased from Cell Signaling Technology, Inc. (St. Louis, MO, USA), and the PGC-1α antibody from Merck Millipore (Burlington, MA, USA). Glucosamine hydrochloride and other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Experimental design
To investigate the dose-dependent effect of GlcN on FGF21 expression in AML12 hepatocytes, cells were treated with varying concentrations of GlcN (0–25 mM) for 24 h, and FGF21 levels were measured in terms of cell content, secretion, mRNA, and protein expression. To further explore the time-dependent effect of GlcN on FGF21 expression, hepatocytes were treated with a fixed concentration of GlcN (10 mM) for different time intervals (0–24 h). Additionally, to gain insight into the mechanisms involved, the following agents were used: Akt inhibitor LY294002 (10− 5 M), mTOR inhibitor rapamycin (10− 7 M), p70S6K inhibitor PF4708671 (10− 5 M), PPARα antagonist GW6471 (10− 5 M), PPARγ antagonist GW9662 (10− 5 M), HIF-1α inhibitor KC7F2 (10− 5 M), PGC-1α inhibitor SR18292 (10− 5 M), and PGC-1α activator ZLN005 (0–10 µM).
AML12 hepatocyte culture and processing
The AML12 (alpha mouse liver 12) cell line (ATCC; Manassas, VA, USA) was cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 100 U/mL penicillin, 100 µg/mL streptomycin, and 10% fetal bovine serum (FBS) at 37 °C in a 5% CO2 atmosphere. The hepatocytes were grown to 100% confluence before being treated with GlcN and various inhibitors for subsequent experiments.
Animals
Eight-week-old male C57BL/6 mice were obtained from the animal center at National Yang Ming Chiao Tung University. Mice were housed under a 12-hour light/dark cycle in a temperature-controlled environment (20–22 °C) with unrestricted access to food and water. Glucosamine hydrochloride was dissolved in phosphate-buffered saline (PBS). For the experiment, animals were randomly assigned to two groups: a PBS-infused control group and a GlcN-infused group. Mice were anesthetized with 2% isoflurane via a nosecone and implanted subcutaneously with mini-osmotic pumps (Alza, Palo Alto, CA, USA) containing either GlcN (1 M, 0.25 µL/h) or PBS. After two weeks of GlcN administration, the mice were euthanized via CO2 exposure, and liver tissue was collected. The study protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of National Yang Ming Chiao Tung University (IACUC number: 1081005). All methods were performed in accordance with ARRIVE guidelines and relevant regulations. Animal experiments were performed at National Yang Ming Chiao Tung University.
Enzyme-linked immunosorbent assay
The levels of FGF21 in the culture medium or plasma samples from mice were quantified using an enzyme-linked immunosorbent assay (ELISA), following the manufacturer’s instructions (MF2100, R&D Systems, Minneapolis, MN, USA).
Western blot
For protein extraction, hepatocyte or liver tissue samples from mice were harvested using a lysis buffer containing 1% Triton X-100, 50 mM KCl, 25 mM HEPES (pH 7.8), 10 µg/mL leupeptin, 20 µg/mL aprotinin, 125 µM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride, and 1 mM sodium orthovanadate. For electrophoresis, 20 µg of protein per lane was loaded onto an SDS-PAGE gel and run for 110 min at 110 V. The gel contents were then transferred to polyvinylidene difluoride (PVDF) membranes at 300 mA for 130 min. After blocking with 5% skim milk in tris-buffered saline with Tween (TBST), the membranes were incubated overnight at 4 °C with primary antibodies: FGF21 (ab171941, Abcam), PGC-1α (ST1202, Merck Millipore), FASN (#3180, Cell Signaling), and GAPDH (#2118, Cell Signaling). This was followed by a 1 h incubation with secondary antibodies at room temperature, and band density was detected using a chemiluminescence reagent.
Oil red O staining
To assess the effect of GlcN on fatty acid accumulation, AML12 hepatocytes were exposed to 300 µM oleic acid (OA) and treated with or without GlcN (10 mM) for 24 h. Following the intervention, cells were fixed in 4% paraformaldehyde at room temperature for 10 min, then stained with a 60% Oil Red O stock solution (3 mg/mL in isopropanol) at 60 °C for 30 min. The stained cells were rinsed once with isopropanol and twice with distilled water. Lipid accumulation was observed under a microscope (Leica, Wetzlar, Germany), and the lipid droplets were subsequently dissolved in isopropanol. The absorbance of the dissolved dye was measured at 510 nm using an ELISA plate reader.
Statistical analysis
Data analysis was performed using SPSS version 22 (SPSS Inc., Chicago, IL, USA). A Student’s t-test was used to compare two groups, while one-way ANOVA was applied for comparisons among more than two groups. All values are presented as mean ± SEM, with statistical significance set at P < 0.05.
Data availability
Data is provided within the manuscript.
References
Reginster, J. Y., Neuprez, A., Lecart, M. P., Sarlet, N. & Bruyere, O. Role of glucosamine in the treatment for osteoarthritis. Rheumatol. Int. 32, 2959–2967 (2012).
Guo, Q. et al. Glucosamine induces increased musclin gene expression through Endoplasmic reticulum stress-induced unfolding protein response signaling pathways in mouse skeletal muscle cells. Food Chem. Toxicol. 125, 95–105 (2019).
Monauni, T. et al. Effects of glucosamine infusion on insulin secretion and insulin action in humans. Diabetes 49, 926–935 (2000).
Biggee, B. A., Blinn, C. M., Nuite, M., Silbert, J. E. & McAlindon, T. E. Effects of oral glucosamine sulphate on serum glucose and insulin during an oral glucose tolerance test of subjects with osteoarthritis. Ann. Rheum. Dis. 66, 260–262 (2007).
Chien, C. S., Cheng, S. C., Wu, H. T., Tsao, C. W. & Cheng, J. T. Insulin resistance induced by glucosamine in fructose-fed rats. Horm. Metab. Res. 41, 542–547 (2009).
Chen, C. H. et al. Raloxifene ameliorates Glucosamine-Induced insulin resistance in ovariectomized rats. Biomedicines 9, 1114 (2021).
Kuo, M., Zilberfarb, V., Gangneux, N., Christeff, N. & Issad, T. O-GlcNAc modification of FoxO1 increases its transcriptional activity: a role in the glucotoxicity phenomenon? Biochimie 90, 679–685 (2008).
Very, N., Vercoutter-Edouart, A. S., Lefebvre, T. & Hardivillé, S. El Yazidi-Belkoura, I. Cross-dysregulation of O-GlcNAcylation and PI3K/AKT/mTOR axis in human chronic diseases. Front. Endocrinol. 9, 602 (2018).
Li, X. et al. Liraglutide Decreases liver fat content and serum fibroblast growth factor 21 levels in newly diagnosed overweight patients with type 2 diabetes and nonalcoholic fatty liver disease. J. Diabetes Res. 3715026 (2021). (2021).
Inagaki, T. et al. Endocrine regulation of the fasting response by PPARalpha-mediated induction of fibroblast growth factor 21. Cell. Metab. 5, 415–425 (2007).
Badakhshi, Y. & Jin, T. Current Understanding and controversies on the clinical implications of fibroblast growth factor 21. Crit. Rev. Clin. Lab. Sci. 58, 311–328 (2021).
Berglund, E. D. et al. Fibroblast growth factor 21 controls glycemia via regulation of hepatic glucose flux and insulin sensitivity. Endocrinology 150, 4084–4093 (2009).
Shao, W. & Jin, T. Hepatic hormone FGF21 and its analogues in clinical trials. Chronic Dis. Transl Med. 8, 19–25 (2022).
Deng, H. et al. PI3K/AKT/mTOR pathway, hypoxia, and glucose metabolism: potential targets to overcome radioresistance in small cell lung cancer. Cancer Pathog Ther. 1, 56–66 (2022).
Cornu, M. et al. Hepatic mTORC1 controls locomotor activity, body temperature, and lipid metabolism through FGF21. Proc. Natl. Acad. Sci. USA. 111, 11592–11599 (2014).
Chen, T. Y. et al. Glucosamine regulation of fibroblast growth factor 21 expression in liver and adipose tissues. Biochem. Biophys. Res. Commun. 529, 714–719 (2020).
Wong, V. W. S. & Adams, L. A. Fibroblast growth factor 21 for non-alcoholic steatohepatitis. Lancet 392, 2658–2660 (2019).
Samms, R. J. et al. FGF21 contributes to metabolic improvements elicited by combination therapy with exenatide and Pioglitazone in patients with type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 323, E123–E132 (2022).
Kang, L. et al. 17β-estradiol protects against glucosamine-induced pancreatic β-cell dysfunction. Menopause 21, 1239–1248 (2014).
Cornu, M., Albert, V. & Hall, M. N. mTOR in aging, metabolism, and cancer. Curr. Opin. Genet. Dev. 23, 53–62 (2013).
Xi, Y. et al. +)-Dehydrovomifoliol alleviates oleic acid-Induced lipid accumulation in HepG2 cells via the PPARα-FGF21 pathway. Front. Pharmacol. 12, 750147 (2021).
Kong, Y. et al. FGF21 reduces lipid accumulation in bovine hepatocytes by enhancing lipid oxidation and reducing lipogenesis via AMPK signaling. Animals 12, 939 (2022).
Kim, S. M., Kim, D. Y., Park, J., Moon, Y. A. & Han, I. O. Glucosamine increases macrophage lipid accumulation by regulating the mammalian target of Rapamycin signaling pathway. BMB Rep. 57, 92–97 (2024).
Salminen, A., Kaarniranta, K. & Kauppinen, A. Regulation of longevity by FGF21: interaction between energy metabolism and stress responses. Ageing Res. Rev. 37, 79–93 (2017).
Acknowledgements
This study was financially supported by research grants from the FEMH-NYCU Joint Research Program (FEMH-NYCU 107DN05, 109DN07), Taiwan.
Author information
Authors and Affiliations
Contributions
S.Y.L performed the experiment, analysed the data and wrote the main manuscript; L.K.C., P.H.L., G.L.W., T.H.W. performed the experiment; Y.B.Y., H.F.L., and C.C.J. reviewed and edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Liu, SY., Chen, LK., Li, PH. et al. Glucosamine induces hepatic FGF21 expression by activating the Akt/mTOR/p70S6K axis and driving PGC-1α activity. Sci Rep 15, 13096 (2025). https://doi.org/10.1038/s41598-025-96249-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-96249-3
