
Abstract
The developing mammalian heart undergoes an important metabolic shift from glycolysis towards mitochondrial oxidation that is critical to support the increasing energetic demands of the maturing heart. Here, we describe a new mechanistic link between mitochondria and cardiac morphogenesis, uncovered by studying mitochondrial citrate carrier (SLC25A1) knockout mice. Slc25a1 null embryos displayed impaired growth, mitochondrial dysfunction and cardiac malformations that recapitulate the congenital heart defects observed in 22q11.2 deletion syndrome, a microdeletion disorder involving the SLC25A1 locus. Importantly, Slc25a1 heterozygous embryos, while overtly indistinguishable from wild type, exhibited an increased frequency of these defects, suggesting Slc25a1 haploinsuffiency and dose-dependent effects. Mechanistically, SLC25A1 may link mitochondria to transcriptional regulation of metabolism through epigenetic control of gene expression to promote metabolic remodeling in the developing heart. Collectively, this work positions SLC25A1 as a novel mitochondrial regulator of cardiac morphogenesis and metabolic maturation, and suggests a role in congenital heart disease.
Similar content being viewed by others
Introduction
In the postnatal heart, mitochondria drive metabolism, providing oxidative energy for contraction. However, the prenatal heart is a hypoxic environment1 with unresolved roles for mitochondrial oxidative energy systems. Nonetheless, mitochondrial metabolism is critical for embryogenesis, as defects in respiratory chain regulation result in lethality by embryonic (E) day 10.5 (E10.5)2,3,4,5,6. Before E11.5, cardiac mitochondria are immature and rely mainly on glycolysis for ATP. As development proceeds (E14.5), cardiac mitochondria mature, and metabolism shifts toward mitochondrial oxidation7,8. This transition may support the increasing energetic demands of the developing heart since increased oxidative metabolism is a cornerstone of cardiomyocyte maturation9.
In parallel with this embryonic metabolic shift, the developing heart undergoes ventricular wall maturation with key processes being trabeculation, septation, and compaction10,11. While metabolism and ventricular morphogenesis are seemingly disparate processes, human primary mitochondrial disorders offers insight into the connection between energetics and ventricular morphogenesis. Patients with mitochondrial disorders including Barth syndrome, MELAS (mitochondrial encephalopathy, lactic acidosis, and stroke-like episodes), and MERRF (myoclonic epilepsy with ragged-red fibers) can present with left ventricular noncompaction, a congenital heart defect caused by impaired compaction of the ventricular wall, suggesting an association between mitochondrial energetics and cardiac structural malformations12,13. However, the molecular mechanisms connecting energetics to ventricular wall morphogenesis are not fully understood. Defining these connections may inform the biology of cardiac development and highlight new targets for improved interventions for congenital heart disease (CHD).
One possible connection between mitochondrial energetics and other cellular processes is the mitochondrial citrate carrier SLC25A1. SLC25A1 is an inner membrane transporter that exports citrate out of the mitochondrial matrix14,15. While mitochondrial citrate is a tricarboxylic acid (TCA) cycle intermediate central to oxidative metabolism, cytosolic citrate is converted into acetyl-CoA which is used in biological pathways ranging from lipid biosynthesis to protein post-translational acetylation and epigenetic control of gene expression16,17,18.
Loss of SLC25A1 reduces respiratory chain function in vitro19,20,21, suggesting that SLC25A1 may regulate mitochondrial function. The SLC25A1 gene is located within a chromosome 22 region compromised in 22q11.2 deletion syndrome (22q11.2DS/DiGeorge syndrome). Hemizygous deletion of this region results in a multisyndromic disorder that includes congenital heart defects in approximately 60–80% of affected individuals22,23. We reported that the neurodevelopmental pathology of 22q11.2DS arises in part due to SLC25A1 loss, which leads to mitochondrial dysfunction and dysregulation of mitochondrial proteins24. Collectively, these data indicate SLC25A1 is a mitochondrial protein that regulates mitochondrial function and connects mitochondria to cytosolic processes. However, the roles of SLC25A1, particularly during development, are not fully delineated.
Here we investigated the role of SLC25A1 in development using systemic Slc25a1 knockout (KO) mice. Our findings implicate SLC25A1 in epigenetic regulation of metabolic reprogramming in the developing heart and positions SLC25A1 as a mitochondrial regulator of cardiac morphogenesis. Importantly, Slc25a1 KO mouse model recapitulates cardiac defects in 22q11.2DS, providing important new insights for this disorder.
Results
Systemic Slc25a1 deletion impairs embryonic growth and causes perinatal lethality
We used the Slc25a1 KO mouse line developed by the International Mouse Phenotyping Consortium to model systemic deletion of Slc25a125, and to reflect systemic gene loss that would be observed in humans with SLC25A1 deficiencies. For these mice, the Slc25a1 KO allele (Slc25a1-) was generated by insertion of an IRES:LacZ trapping cassette and a neomycin selection cassette between exons 1 and 2 of Slc25a1 (Fig. 1A). Slc25a1 KO mice (Slc25a1-/-) exhibit perinatal lethality 26. Thus, we conducted timed matings of Slc25a1 hemizygous KO mice (Slc25a1+/-) to study the effects of Slc25a1 ablation (Slc25a1-/-) and Slc25a1 hemizygous loss (Slc25a1+/-), compared with wild type (WT) littermate controls (Slc25a1+/+). We confirmed that Slc25a1-/- embryos at E18.5 were viable, and Slc25a1+/- intercrosses yielded the anticipated genotypic frequencies (Fig. 1G). However, E18.5 Slc25a1-/- embryos displayed impaired growth with reduced crown-rump lengths compared to Slc25a1+/- and Slc25a1+/+ controls (Fig. 1B, C). Additionally, while Slc25a1-/- embryos displayed microencephaly, anencephaly, encephalocele, and gastroschisis, Slc25a1+/- embryos were indistinguishable from Slc25a1+/+ controls. Analyses of SLC25A1 protein levels in E18.5 heart and brain revealed robust expression in Slc25a1+/+ embryos, reduced expression in Slc25a1+/- embryos, and absent expression from Slc25a1-/- embryos (Fig. 1D). This gene dosage-dependent SLC25A1expression was confirmed by immunofluorescence in embryonic hearts. In WT E14.5 hearts, SLC25A1 strongly colocalizes with the mitochondrial inner membrane protein ATP5A1, confirming mitochondrial localization (Supplemental Fig. 1). However, while SLC25A1 immunofluorescence was robust in Slc25a1+/+ hearts, expression was weaker in Slc25a1+/- hearts and undetectable in Slc25a1-/- hearts (Fig. 1E).

A Schematic of the Slc25a1 knockout (KO) first allele. A lacZ-neo KO cassette was inserted between exons 1 and 2 of Slc25a1. B Representative E18.5 Slc25a1+/+, Slc25a1+/-, and Slc25a1-/- embryos obtained from timed intercross of Slc25a1+/- mice. C Quantification of crown-rump lengths of E18.5 embryos of the indicated genotypes (n = 13–26/group). Values reported as mean ± SEM. One-tailed Student’s t test was used for statistical analysis. ****P < 0.0001. D Western blot of SLC25A1 expression in total protein isolated from E17.5 hearts and brains from the indicated genotypes. GAPDH or TGX protein staining was used as loading controls. E Representative immunofluorescence images of SLC25A1 expression in E14.5 hearts from the indicated genotypes. F Representative image of P0 pups obtained from timed intercross of Slc25a1+/- mice. G Genotype frequencies of E18.5 embryos and P0 pups obtained from timed intercross of Slc25a1+/- mice. Illustrations from Clkr.com.
Consistent with previous reports, we were unable to recover adult Slc25a1-/- mice from Slc25a1+/- intercrosses. However, Slc25a1-/- pups were born and obtained at the anticipated genotype frequencies (Fig. 1F, G). Further, postnatal (P) day 0 (P0) Slc25a1-/- neonates were small and cyanotic, and some displayed microencephaly and gastroschisis (Fig. 1F). Importantly, Slc25a1-/- neonates died within 6 h of birth, suggesting that systemic Slc25a1 ablation causes perinatal lethality.
Partial Slc25a1 deletion is sufficient to produce cardiac structural defects in mice
SLC25A1 is expressed in the mouse heart27,28, and given that Slc25a1-/- neonates were cyanotic, we hypothesized that SLC25A1 loss could impair cardiac contractile function. We assessed SLC25A1 expression in the developing heart by immunofluorescence in WT E10.5, E12.5, and E14.5 embryos and E18.5 hearts. Because the cardiac endocardium, myocardium, and epicardium play distinct roles in ventricular morphogenesis, we investigated expression of SLC25A1 in these different layers. Examining fields of the embryonic heart spanning the myocardium and epicardium (Fig. 2A) as well as the myocardium and endocardium (Supplemental Fig. 2), we observed strong SLC25A1 expression in the myocardium and epicardium (Fig. 2A) and minimal expression in the endocardium (Supplemental Fig. 2).

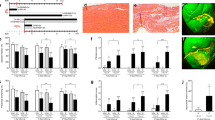
A Representative immunofluorescence images of SLC25A1 expression in wild type embryo (E10.5, E12.5, and E14.5) and heart (E18.5) sections (6 μm). DAPI was used to counterstain nuclei, and MF20 was used to counterstain cardiomyocytes. Dashed yellow line outlines myocardium from the epicardium; asterisk indicates epicardium positive for SLC25A1. B Representative images of E10.5 embryos, (C) E12.5 embryos, (D) E14.5 embryos, and (E) E18.5 hearts from the indicated genotypes stained with hematoxylin and eosin. Asterisk indicates noncompaction, arrow indicates ventricular septal defect, and triangle indicates right ventricular hypoplasia. F Quantification of relative compact myocardium thickness or (G) relative trabecular myocardium thickness in E18.5 hearts (n = 8-17/group). H Frequency of cardiac defects observed at E14.5 in embryos with the indicated genotypes (n = 27/group). Values reported as mean ± SEM. One-tailed Student’s t test was used for statistical analysis. *P < 0.05; ***P < 0.0005; ****P < 0.0001.
To investigate the structural consequences of SLC25A1 loss in the embryonic heart, we obtained Slc25a1-/-, Slc25a1+/-, and control Slc25a1+/+ embryos from intercrosses of Slc25a1+/- mice. Embryos were collected at E10.5, E12.5, and E14.5 and hearts were collected at E18.5 to reflect timepoints when trabeculation occurs (E10.5 and E12.5), when septation should be complete and ventricular wall compaction should be fulminant (E14.5), and when major embryonic milestones of cardiac morphogenesis should be complete (E18.5)10,11. Histological analyses revealed similar cardiac morphology in E10.5 Slc25a1+/+, Slc25a1+/-, and Slc25a1-/- embryos (Fig. 2B). At E12.5, the compact myocardium of Slc25a1-/- embryos appeared to be slightly less developed than that of Slc25a1+/+ and Slc25a1+/- embryos (Fig. 2C). However, by E14.5 and E18.5, hearts from Slc25a1-/- embryos displayed cardiac abnormalities (Fig. 2D–H) including ventricular septal defects (VSDs); ventricular noncompaction, as evidenced by decreased compact myocardium and increased trabecular myocardium (Fig. 2F, G) compared to age-matched Slc25a1+/+ controls; and right ventricular hypoplasia. As VSDs have been reported in Slc25a1-/- embryos29, our findings extend the cardiac abnormalities associated with Slc25a1 deletion to include noncompaction and right ventricular hypoplasia.
Slc25a1+/- embryos, which were morphologically indistinguishable from Slc25a1+/+ controls, unexpectedly displayed increased frequency of cardiac malformations including VSDs, ventricular noncompaction, and right ventricular hypoplasia (Fig. 2D–H). Collectively, our data demonstrate that SLC25A1 is necessary for ventricular morphogenesis and that heterozygous loss (Slc25a1 haploinsufficiency) is sufficient to cause structural defects in the developing heart.
Slc25a1 haploinsufficiency decreases neonatal survival in mice
While Slc25a1+/- embryos displayed congenital heart defects, adult Slc25a1+/- mice were fertile and did not display differences in survival with aging. This prompted us to investigate the structural and functional consequences of Slc25a1 haploinsufficiency in the postnatal heart, and specifically, if adult Slc25a1+/- mice harbor cardiac defects. Histological analyses of hearts from 2-month-old Slc25a1+/- and Slc25a1+/+ control mice (n = 10/group) revealed no structural or morphological defects (Supplemental Fig. 3A). Additionally, no changes were observed in gravimetric or echocardiographic parameters (Supplemental Fig. 3B, C), suggesting that cardiac structure and function were unaltered in adult Slc25a1+/- mice.
To understand how we could observe cardiac malformations in Slc25a1+/- embryos but not adult mice, we examined the frequencies of progeny genotypes. We used a breeding strategy of Slc25a1+/- mice crossed to Slc25a1+/+ mice, with genotyping conducted on P7 tissue biopsies. Genotype analyses of 446 pups from 80 litters revealed a slight but significant [χ2 (1, N = 446) = 6.0628; p = 0.014] increase in the frequency of P7 Slc25a1+/+ (56% observed; 50% expected) versus Slc25a1+/- pups obtained (44% observed; 50% expected; Supplemental Fig. 3D), even though Slc25a1+/- pups were recovered at expected frequencies at P0 (N = 47, Fig. 1F). These data suggest that Slc25a1 haploinsufficiency decreases neonatal survival. Moreover, as cardiac morphometry and function in Slc25a1+/- mice were normal at 2 months of age, our data suggest that Slc25a1+/- pups born with cardiac defects die in the neonatal period.
Association analysis of SLC251A variants and pediatric CHD
In humans, SLC25A1 single-gene variants underlie two rare neurometabolic and neuromuscular diseases, D-2 and L-2 aciduria (DL-2HGA; OMIM: 615182) and congenital myasthenic syndrome-23 (CMS23; OMIM: 618197)30,31,32. SLC25A1 also is among the genes lost in the chromosomal microdeletion disorder 22q11.2DS (OMIM:188400)22,33. Individuals with 22q11.2DS can present with CHD, but these defects have not been attributed to SLC25A133. Additionally, there are no human cardiac phenotypes annotated to SLC25A1 single-gene defects (https://hpo.jax.org/app/browse/gene/6576), and it is unknown if SLC25A1 variants underlie cardiac disease. Yet, our surprising findings of ventricular malformations in Slc25a1 deficient mouse embryos prompted us to investigate if SLC25A1 variants are associated with human CHD. We analyzed whole-exome sequencing data from the Pediatric Cardiac Genomics Consortium (PCGC), encompassing 3740 CHD probands with 3401 pseudo-sibship case-controlled pairs (generated from parent-proband trios)34,35. Rare SLC25A1 variants were selected using a minor allele frequency of <0.001 in the Genome Aggregation Database (gnomAD) and a combined annotation-dependent depletion (CADD) score of >20. We generated a separate set including only ultrarare (i.e., do not appear in gnomAD exomes) and loss-of-function SLC25A1 variants. While we were unable to identify an association of rare and ultrarare SLC25A1 variants with specific CHD presentations, gene burden analyses using the kernel-based adaptive cluster (KBAC) method revealed that ultrarare nonsynonymous and stop-gain SLC25A1 variants trend towards an association with CHD (p = 0.058 and Bonferroni-corrected p-value of 0.0167 ; Supplemental Data 1). Moreover, many of these variants were associated with cardiac defects including VSDs, tetralogy of Fallot, and conotruncal defects—which are observed in 22q11.2DS (Supplemental Data 2).
Slc25a1 deletion alters metabolic gene expression in mice
SLC25A1 functions as a mitochondrial solute transporter which exports mitochondrial citrate14,15. Previous studies have suggested a role for SLC25A1 beyond metabolite transport in the direct regulation of respiratory chain function, as SLC25A1 deletion in cancer and neuronal systems leads to impaired respiration19,24,36. Our work in the HAP1 human near-haploid lymphoblastoid cell line proposes a role for SLC25A1 in the regulation of mitochondrial translation, as loss of SLC25A1 is associated with reduced expression of core mitochondrial ribosome subunits21. Thus, we hypothesized that SLC25A1 regulation of mitochondrial translation and respiration would also be operant in the developing heart. We examined expression levels of key mitochondrial ribosome subunits, MRPS22 and MRPS18B, in E17.5 Slc25a1+/+, Slc25a1+/-, and Slc25a1-/- embryonic hearts. Surprisingly, MRPS22 and MRPS18B expression was unchanged with Slc25a1 deletion (Supplemental Fig. 4), suggesting that SLC25A1 does not regulate mitochondrial translation in the developing heart.
To delineate the metabolic consequences of Slc25a1 deletion, we used a NanoString mRNA quantification panel to profile expression of 768 metabolism-related genes encompassing key metabolic pathways. Profiling metabolic gene expression in Slc25a1+/- versus Slc25a1+/+, Slc25a1-/- versus Slc25a1+/+, and Slc25a1+/- versus Slc25a1-/- hearts revealed 28 distinct differentially expressed genes (DEGs) with Slc25a1 deletion at E17.5 (p < 0.05). Of these genes, 10 were differentially expressed between Slc25a1-/- versus Slc25a1+/+, two between Slc25a1+/- versus Slc25a1+/+, and 11 between Slc25a1+/- versus Slc25a1-/- hearts (fold-change = 1.5; p < 0.05; Fig. 3A). Hierarchical clustering of DEGs revealed that while Slc25a1+/+ and Slc25a1-/- hearts had distinct gene expression profiles, Slc25a1+/- hearts had an intermediate pattern (Fig. 3B), suggesting that SLC25A1 deletion-associated dysregulation of metabolic genes occurs in a dosage-dependent manner. Additionally, using Metascape37 we identified antigen processing, organic acid transport, positive regulation of cell death, AMPK signaling, and response to hormone as the most significantly altered pathways with Slc25a1 deletion (Fig. 3C).

A Volcano plots of Nanostring metabolic transcript profiling comparing gene expression of Slc25a1-/- and Slc25a1+/+, Slc25a1+/- and Slc25a1+/+, and Slc25a1-/- and Slc25a1+/- E17.5 hearts. Blue denotes downregulated transcripts. Yellow denotes upregulated transcripts. See Supplemental Data 3. B Heat map depicting transcripts differentially expressed across genotypes in A. Transcripts in rows were selected by ANOVA multiple comparisons followed by Benjamini-Hochberg correction (q < 0.05) and clustered using Kendall-Tau (see Supplemental Data 3). Each column represents an individual animal. C Gene ontologies identified with Metascape from genes differentially expressed in Slc25a1-/- and Slc25a1+/- versus Slc25a1+/+ hearts. D Geneset enrichment analysis of oxidative phosphorylation, glycolysis, hypoxia, and fatty acid metabolism genes in Slc25a1+/+ vs. Slc25a1-/- and Slc25a1+/+ vs. Slc25a1+/- E17.5 hearts. E Heatmaps visualizing gene expression of oxidative phosphorylation and glycolysis genes from Nanostring transcript profiling in E17.5 Slc25a1+/+ vs. Slc25a1-/- hearts. Blue denotes reduced expression; red denotes increased expression.
To define metabolic processes impacted by Slc25a1 deletion, we performed gene set enrichment analysis (GSEA) of REACTOME pathways using normalized gene expression from the NanoString panel as gene ranks. GSEA revealed oxidative phosphorylation, glycolysis, hypoxia, and fatty acid metabolism as the most significantly altered pathways in Slc25a1-/- versus Slc25a1+/+ and Slc25a1+/- versus Slc25a1+/+ hearts, with higher enrichment scores for these pathways in control Slc25a1+/+ hearts versus hearts with Slc25a1 deletion (Slc25a1-/- and Slc25a1+/-) (Fig. 3D, E). These results suggest that Slc25a1 deletion induces mitochondrial dysfunction and metabolic remodeling of the embryonic heart.
Importantly, these data show that loss of one copy of Slc25a1 is sufficient to induce gene expression changes. Intriguingly, comparisons of the enrichment plots for oxidative phosphorylation, glycolysis, hypoxia, and fatty acid metabolism revealed a right-ward shift of the Slc25a1+/- versus Slc25a1+/+ plots as compared to Slc25a1-/- versus Slc25a1+/+ plots (Fig. 3D). These graded differences between homozygous and heterozygous Slc25a1 knockouts suggest that full Slc25a1 ablation induces greater mitochondrial and metabolic dysregulation than partial loss. These results align with our findings that Slc25a1-/- embryos displayed a higher frequency and severity of ventricular malformations than Slc25a1+/- embryos.
Slc25a1 deletion causes mitochondrial ultrastructural defects in embryonic mouse heart
Our metabolic gene expression profiling suggested that SLC25A1 loss causes dosage-dependent mitochondrial dysfunction in the developing heart. Because mitochondrial structure and function are often linked38, we investigated the impact of SLC25A1 loss on mitochondrial ultrastructure. Transmission electron microscopy of E17.5 hearts revealed both partial and complete Slc25a1 ablation induced striking alterations in cardiac mitochondrial morphology— highly immature mitochondria with disordered cristae and open matrix voids (Fig. 4A). Quantification of mitochondrial morphology further revealed that Slc25a1+/- and Slc25a1-/- mitochondria had significantly increased area and minimal Feret’s diameter, despite similar circularity (Supplemental Fig. 5A–C), suggesting larger and swollen mitochondria. Slc25a1-/- mitochondria further displayed significantly decreased aspect ratio (Supplemental Fig. 5D) and increased roundness (Supplemental Fig. 5E), suggesting that complete SLC25A1 ablation causes greater changes in mitochondrial morphology, compared to partial loss. Our data suggest that, similar to metabolic gene expression changes, mitochondrial ultrastructural abnormalities are Slc25a1 dosage-dependent. Moreover, as cardiac mitochondria become more elongated and develop more elaborate and organized cristae during cardiac development, our data suggest that SLC25A1 loss impairs mitochondrial structural maturation.

A Representative electron micrographs of cardiac ultrastructure from the indicated genotypes at E17.5 (n = 3 Slc25a1+/+; n = 10 Slc25a1+/-; n = 3 Slc25a1-/- embryos analyzed). B Coupled and uncoupled mitochondrial oxygen consumption rates measured in lightly permeabilized whole E17.5 hearts of the indicated genotypes (n = 5 Slc25a1+/+; n = 5 Slc25a1+/-; n = 3 Slc25a1-/- hearts analyzed). C Quantification of mitochondrial DNA (mtDNA) to nuclear DNA (nDNA) ratios in E17.5 hearts of the genotypes indicated (n = 6 Slc25a1+/+; n = 6 Slc25a1+/-; n = 4 Slc25a1-/- embryos analyzed). D Western blot of respiratory chain complex subunits (SDHA, ATP5A, UQCRC2, NDUFA9, and COXIV). Coomassie staining of the gel was used as a protein loading control. E Densitometry quantification of NDUFA9, SDHA, UQCRC2, COXIV, and ATP5A expression (n = 3/group). F Respiratory chain complexes in E17.5 heart mitochondria as assessed by BN-PAGE followed by immunoblotting with an OXPHOS antibody cocktail. A western blot of VDAC was used as a protein loading control. Values presented as means ± SEM. One-way ANOVA followed by a post-hoc Dunnett’s test. ns denotes not significant; *P < 0.05; **P < 0.005.
Slc25a1 deletion impairs respiratory chain function in the developing mouse heart
Since our data suggested that Slc25a1 deletion causes mitochondrial dysfunction, we assessed the impact of Slc25a1 deletion on mitochondrial respiratory chain function. Respirometry was conducted on E17.5 Slc25a1+/-, Slc25a1-/-, and Slc25a1+/+ hearts. Coupled respiration (supported by glutamate and malate) of Slc25a1+/- and Slc25a1-/- hearts was significantly decreased compared to Slc25a1+/+ controls (Fig. 4B). These respiration deficits were also observed with chemical uncoupling by FCCP (carbonyl cyanide-p-trifluoromethoxyphenylhydrazone), indicating that Slc25a1 deletion also impairs maximal respiratory capacity. Together, these results support impaired oxidative phosphorylation with SLC25A1 loss.
Because defects in mitochondrial cristae ultrastructure have been associated with mitochondrial DNA (mtDNA) depletion39,40 and the mtDNA encodes essential respiratory chain complex subunits, Slc25a1 deletion-induced mitochondrial ultrastructural abnormalities (Fig. 4A) could lead to respiration defects by causing mtDNA loss. Thus, we measured mtDNA to nuclear DNA (nDNA) ratios (mtDNA/nDNA) in total DNA isolated from E17.5 Slc25a1+/-, Slc25a1-/-, and Slc25a1+/+ hearts. While mtDNA/nDNA ratios were not significantly different between Slc25a1+/- and Slc25a1+/+ hearts, ratios were elevated in Slc25a1-/- hearts, indicating increased mtDNA content (Fig. 4C). Thus, SLC25A1 deletion-associated respiration defects are not due to mtDNA depletion.
To confirm that SLC25A1 deletion does not alter mitochondrial content in the developing heart, we conducted immunoblotting for TOM20—a ubiquitously expressed subunit of the mitochondrial import machinery—and observed no differences between Slc25a1+/-, Slc25a1-/-, and Slc25a1+/+ control hearts (Supplemental Fig. 6A, B). Collectively, these data suggest that SLC25A1 expression does not impact cardiac mitochondrial content.
We also investigated the impact of Slc25a1 deletion on respiratory chain complex subunit expression (NDUSFA9 of complex I, SDHA of complex II, UQCRC2 of complex III, COXIV of complex IV, and ATP5A of complex V) by immunoblotting E17.5 total heart lysates (Fig. 4D). Slc25a1+/- hearts displayed a trend for decreased NDUSFA9, UQCRC2, and ATP5A as well as significantly decreased COXIV compared to controls (Fig. 4E). Slc25a1-/- hearts displayed significantly decreased expression of all subunits compared to controls (Fig. 4E). Because reduced expression of individual respiratory chain subunits can cause an overall reduction in assembled respiratory chain, we analyzed respiratory chain assembly in mitochondria isolated from E17.5 hearts using blue native polyacrylamide gel electrophoresis (BN-PAGE) and immunodetection for the respiratory chain (Fig. 4F). Cardiac mitochondria from Slc25a1+/- and Slc25a1-/- embryos displayed decreased levels of assembled complexes I, III, and V. This Slc25a1-dependent decrease in assembled respiratory chain complexes aligns with alterations in metabolic gene expression, subunit expression, mitochondrial ultrastructural abnormalities, and impaired respiration in SLC25 A1-deleted hearts.
Computational modeling predicts metabolic dysregulation and increased glucose dependence with Slc25a1 deletion
To investigate the overall metabolic consequences of Slc25a1 deletion in the E17.5 mouse heart, we performed targeted gas chromatography–mass spectrometry (GC-MS) metabolomic profiling on 25 key metabolites, encompassing TCA cycle intermediates, amino acids, and metabolic byproducts that are critical to cardiac energetics (Fig. 5A). Principal component analysis (PCA) as well as unbiased hierarchical clustering of the measured analytes separated samples according to their Slc25a1 genotype (Fig. 5B, C), with Slc25a1+/- hearts displaying an intermediate metabolite profile compared to Slc25a1+/+ and Slc25a1-/- hearts. Citrate was significantly less abundant in Slc25a1-/- compared to Slc25a1+/+ hearts, while Slc25a1+/- hearts displayed a trend for reduction (Fig. 5D), indicating that SLC25A1 deletion causes a decrease in cardiac citrate levels. Additionally, glutamate, cystine and lysine abundance increased (Fig. 5D), while threonine, tryptophan, aconitate and pyroglutamate levels were decreased with SLC25A1 deletion. Glutamine and glutamate undergo cyclization to pyroglutamate during the de novo synthesis of gamma-glutamyl cycle of glutathione (l-γ-glutamyl-l-cysteinyl-glycine; GSH). Likewise, cystine is a precursor for GSH synthesis. Hence, the reduced pyroglutamate together with increased glutamate and cystine indicates reduced GSH synthesis with Slc25a1 deletion.

A Schematic of the metabolites profiled by GC-MS. B Principal component analysis (PCA) and (C) unbiased hierarchical clustering of profiled analytes showing clear segregation of Slc25a1+/+(n = 6), Slc25a1+/- (n = 3), and Slc25a1-/- (n = 4) hearts. D Normalized abundance of selected metabolites that are significantly altered in HET and KO hearts. Values presented as means ± SEM. One-tailed Student’s t test was used for statistical analysis. P < 0.05 was considered significant, *P < 0.05 and **P < 0.005. E Malate-to-citrate ratios and (F) Fumarate-to-malate ratios in Slc25a1+/+, Slc25a1+/- and Slc25a1-/- hearts. Values presented as means ± SEM. One-way ANOVA followed by a post hoc Dunnett’s test was used for statistical analysis. P < 0.05 was considered significant, ns denotes not significant, *P < 0.05.
Because glucose-derived citrate can either be exported from the mitochondria to the cytosol via SLC25A1 or decarboxylated to succinate through the TCA cycle, we compared malate-to-citrate and fumarate-to-malate ratios to assess the incorporation and conversion of citrate-derived carbons within the TCA cycle. With Slc25a1 deletion, the malate-to-citrate ratio was significantly increased in Slc25a1-/- hearts compared to controls, while Slc25a1+/- hearts displayed a trend for an increase (Fig. 5E). However, fumarate-to-malate ratios remained unchanged (Fig. 5F). These data suggest that SLC25A1 deletion reduces citrate incorporation into the TCA cycle thereby limiting oxidative metabolism.
To assess how SLC25A1 deletion impacts metabolic flux in the embryonic heart, we integrated our data using CardioNet41, a genome-scale metabolic network of cardiac metabolism. To infer flux distributions, we used flux balance analysis and constrained internal metabolite fluxes based on our GC-MS-based metabolomics data (Fig. 5), and mitochondrial respiration rates (Fig. 4B). Linear programming was used to calculate solutions that satisfy mass balance and flux constraints. The objective function was defined as a maximization-minimization problem to reflect the requirements of the embryonic heart to maximize ATP provision and biomass synthesis (e.g., protein and membrane lipid synthesis) while minimizing utilization of extracellular substrates and oxygen from the plasma. Flux distributions were analyzed by PCA and among genotype-dependent separation, 80.6% of the total variance was captured in the first two dimensions (Fig. 6A). CardioNet simulations predict increased exogenous glucose utilization with SLC25A1 loss (Fig. 6B, C), which corresponds with increased glycolytic flux and significantly less glucose-derived pyruvate entry into the TCA cycle, which corresponds with reduced TCA cycle flux and ATP provision (Supplemental Fig. 7A, Fig. 6B), as well as reduced oxidative phosphorylation (Fig. 6D). Correspondingly, we identified reduced glutamine-pyruvate transaminase flux compared to increased glutamic-oxaloacetic transaminase flux (Supplemental Fig. 7B), suggesting that SLC25A1 loss increases glutamine anaplerosis. Ultimately, as embryonic cardiac metabolism matures from dependence on glycolysis to mitochondrial oxidation-derived energy, our simulations suggest that SLC25A1 loss impairs metabolic reprogramming and maturation in the developing heart.

A PCA of the CardioNet simulation revealed a clear clustering of Slc25a1+/+, Slc25a1+/-, and Slc25a1-/- hearts by Slc25a1 genotype. B Unsupervised hierarchical clustering of significantly altered flux distributions. Two-way ANOVA with post hoc Tukey’s test followed by multiple comparison analysis (FDR < 1% after Benjamini, Krieger and Yekuteli). C Predicted flux rates for glucose uptake, and D oxidative phosphorylation in response to SLC25A1 deletion. Box plot parameters are defined by the minimum value, the first quartile, the median, the third quartile, and the maximum value. *P < 0.05; ****P < 0.00005.
Slc25a1 deletion reduces histone acetylation in developing mouse hearts
Our combined transcriptomics, biochemical, and metabolomics analyses revealed defects in mitochondrial oxidative metabolism with Slc25a1 deletion. We next assessed potential mechanisms that control Slc25a1 deletion-associated metabolic dysregulation. SLC25A1 is the main mitochondrial transporter for citrate export10,11. In the cytosol, citrate is converted to acetyl-CoA, a substrate for epigenetic modifications like histone acetylation17,18. As acetyl-CoA abundance modulates histone acetylation, SLC25A1 could connect mitochondrial metabolism to epigenetic gene regulation via modulating citrate. To determine whether the decreased citrate levels in Slc25a1+/- and Slc25a1-/- hearts (Fig. 5D), alters histone acetylation, we assessed acetylation of histone H3 at lysine 9 (H3K9), which is often enriched at promoters of actively transcribed genes42, by immunoblots of total extracts from E17.5 hearts (Fig. 7A). The ratio of acetylated H3K9 (H3K9ac) to total histone H3 was reduced in Slc25a1+/- and Slc25a1-/- hearts compared to Slc25a1+/+ controls (Fig. 7B), suggesting that SLC25A1 regulates histone acetylation in the developing heart.

A Representative western blot of histone H3 acetylated at K9 (H3K9ac) and total histone H3 in total heart protein from E17.5 embryos of the indicated genotypes. B Densitometry quantification of H3K9ac to H3 levels in E17.5 hearts (n = 3/group). One-way ANOVA followed by post hoc Dunnett’s test was used for statistical analysis. *P < 0.05. C H3K9ac ChIP qPCR of H3K9ac enrichment at the promoters of Ndufa1, Ndufb2, Ndufb4, Ndufb8, Ndufs8, Cox4i1, Cox5b, Uqcrq, Pdha1 and Pparg in E17.5 hearts (n = 4-5/group). ChIP with anti-rabbit IgG was used as a negative control. Data were normalized using input DNA. Values presented as means ± SE. One-way ANOVA followed by post hoc Dunnett’s test was used for statistical analysis. ns denotes not significant; *P < 0.05, **P < 0.005, and ***P < 0.0005. D Proposed model of SLC25A1-mediated citrate export and its role in regulation of metabolic gene expression in the developing heart. Citrate in the mitochondria is generated from acetyl-CoA and oxaloacetate in the TCA cycle. SLC25A1 mediates mitochondrial citrate export. Citrate in the cytosol is converted to acetyl-CoA, which is used for histone acetylation (H3K9ac) and impacts gene expression necessary for proper heart development. Diagram created using BioRender.
We next postulated that SLC25A1-related histone acetylation changes could affect gene expression. Given technical challenges in profiling gene expression in large numbers of embryonic hearts, we chose a candidate gene approach as a proof of concept. Because SLC25A1 deletion caused a striking decrease in respiration, we focused on mitochondrial metabolism genes that were both dysregulated in the NanoString panel (Ndufa1, Ndufb4, Ndufs8, Ndufb2, Ndufb8, Uqcrq, Cox4i1, Cox5b, and Pdha) and had H3K9ac promoter enrichment assessed by analyses of publicly available H3K9ac chromatin immunoprecipitation-sequencing (ChIP-seq) datasets42,43 (Supplemental Figure 8A). We additionally prioritized Pparg for analysis because it is a transcription factor that regulates metabolic gene expression44 that is significantly dysregulated in our transcriptomics analysis, has H3K9ac promoter enrichment (Supplemental Fig. 8A), and cardiomyocyte-specific deletion phenocopies the VSDs45 observed in Slc25a1 deleted embryos. Moreover, analysis of the SLC25A1 deletion-dysregulated metabolism genes identified Pparg as a top potential transcriptional regulator (Supplemental Fig. 8B). Thus, to test the possibility that SLC25A1 impacts the expression of these candidate genes by regulating H3K9ac promoter enrichment, we performed H3K9ac ChIP followed by quantitative PCR (qPCR) to assess H3K9ac promoter abundance in E17.5 hearts. Our results revealed significantly reduced H3K9ac at the promoters of Ndufa1, Ndufb4, Ndufs8, Ndufb2, Ndufb8, Uqcrq, Cox5b, Pdha1, and Pparg in Slc25a1+/- and Slc25a1-/- hearts compared to Slc25a1+/+ controls, and a trend for decreased H3K9ac at the Cox4i1 promoter (Fig. 7C). These results support an epigenetic role for SLC25A1, in which SLC25A1 alters histone acetylation to regulate gene expression in the developing heart.
Discussion
Previous investigations have associated SLC25A1 with several diseases24,26,30,31,32,36. Our results here provide new evidence that SLC25A1 also contributes to metabolic reprogramming during cardiac development. Our mouse model results show that systemic loss of SLC25A1 led to stunted embryonic growth, perinatal lethality, and developmental abnormalities including cardiac structural defects. Moreover, SLC25A1 loss resulted in mitochondrial dysfunction and perturbations in metabolic gene expression, and derangements in cardiac mitochondrial ultrastructure and function that collectively point to a role for SLC25A1 in metabolic reprogramming during mouse heart development. Importantly, Slc25a1 haploinsufficiency was sufficient to produce cardiac developmental defects, which may underscore the clinical relevance of our findings.
Our initial efforts to identify the mechanisms by which Slc25a1 deletion reduces respiratory chain function in the embryonic mouse heart led us to examine the relationship between SLC25A1 and the mitochondrial ribosome, as SLC25A1 regulates mitochondrial ribosome subunit expression in vitro21. Our analyses of E17.5 hearts did not reveal any alterations in protein expression of key mitochondrial ribosome subunits with SLC25A1 loss, indicating that Slc25a1 deletion-induced mitochondrial dysfunction is not due to impaired mitochondrial translation. Because the mitochondrial proteome and function can differ between different tissues and cell types46,47, our data point to possible cell type- and tissue-specific effects of SLC25A1.
Our studies of metabolic gene expression revealed intriguing consequences of SLC25A1 deletion. First, hierarchical analysis of differentially expressed genes revealed a Slc25a1 dosage-dependent dysregulation of metabolic genes that parallels the dosage-dependent frequency/degree of severity of cardiac malformations observed in vivo. Moreover, while gene ontology analysis revealed organic acid transport as a significant pathway impacted by SLC25A1 deletion—anticipated given that SLC25A1 itself is an organic acid transporter—AMPK signaling and positive regulation of cell death unexpectedly also emerged as key pathways. AMPK signaling is a critical metabolic pathway that mediates a coordinated response to energy dysregulation48, and the identification of this pathway reinforces the dysregulation of pathways like OXPHOS and glycolysis that we identified by GSEA, as well as mitochondrial dysfunction we observed by respirometry. Notably, increased cell death during cardiac morphogenesis can perturb cardiac structural maturation49,50,51, suggesting that SLC25A1 deletion-associated upregulation of cell death may contribute to observed cardiac malformations.
Integrating our mitochondrial functional studies, transcriptomics, metabolomics, and in silico studies to understand the metabolic ramifications of SLC25A1 deletion confirmed dosage dependency of metabolic dysregulation and highlighted the critical role of SLC25A1 in metabolic reprogramming in the developing mouse heart. We found that SLC25A1 loss causes dysregulation of cardiac metabolism by increasing glucose utilization and glutamine anaplerosis while impairing TCA cycle flux and oxidative metabolism. The immature developing heart is mainly fueled by glycolytic ATP, while the maturing heart incorporates a wide-range of substrates including fatty acids, ketone bodies and amino acids. Our multi-omics analysis indicates that SLC25A1 loss leads to an ‘immature’ metabolic profile, whereby SLC25A1 deleted hearts remain reliant on glucose with impaired oxidative metabolism.
How can the focal loss of a mitochondrial transporter result in such widespread changes to the metabolic landscape of the developing heart? SLC25A1 transports mitochondria-produced citrate to the cytosol14,15. As part of the TCA cycle, mitochondrial citrate is formed from condensation of acetyl-CoA and oxaloacetate and is further oxidized to enable oxidative energy production52. In the cytosol, citrate can be converted back into acetyl-CoA by ATP-citrate lyase53. Acetyl-CoA is an essential substrate for protein post-translational acetylation and epigenetic regulation of gene expression via histone acetylation and cellular acetyl-CoA levels modulate histone acetylation status18,53. As acetyl-CoA cannot cross the inner membrane, SLC25A1 may link mitochondrial metabolism to epigenetic regulation of gene expression by modulating acetyl-CoA compartmentalization via citrate.
Our proof-of-concept ChIP of metabolism-related genes suggest a critical role for SLC25A1 in regulating gene expression in the developing heart through H3K9 acetylation (Fig. 7D). We found that SLC25A1 loss results in decreased total H3K9ac in the developing heart, leading to reduced H3K9ac at the promoters of mitochondrial metabolism-associated genes, which aligns with our observed reduction in metabolic gene expression and mitochondrial respiration of SLC25A1 deleted hearts. As embryos with cardiomyocyte-specific deletion of PPARγ phenocopy the VSDs we observed in mouse embryos with systemic Slc25a1 deletion45, our finding of reduced H3K9ac at the PPARγ promoter further support a link between SLC25A1 and PPARγ in the developing heart. Collectively, our findings indicate that SLC25A1 is a mitochondrial regulator of genetic reprogramming required for oxidative metabolism in the maturing heart.
Our discovery that Slc25a1 KO embryos display cardiac malformations and mitochondrial dysfunction is notable but could be expected. However, one striking finding of our study is that Slc25a1 haploinsufficiency in mice also produces cardiac malformations and mitochondrial dysfunction, generating a clinical connection to CHD. In humans, SLC25A1 is located on chromosome 22q11.2 within the critical deletion region associated with 22q11.2DS22. Cardiac presentations of 22q11.DS commonly include conotruncal defects like VSDs, tetralogy of Fallot23, and left ventricular noncompaction54,55. Cardiac defects associated with 22q11.2DS have largely been ascribed to TBX1, a transcription factor located in the critical deletion region that plays important roles in cardiovascular development56. Yet, 22q11.2DS is a syndrome of haploinsufficiency and Tbx1 haploinsufficiency does not reproduce the full spectrum of cardiac defects associated with 22q11.2DS; thus, additional factors must contribute to its cardiac phenotypes. Because partial loss of Slc25a1 in mice is sufficient to produce CHD observed in 22q11.2DS, our data position SLC25A1 as a novel 22q11.2 gene that contributes to the cardiac presentation of 22q11.2DS.
Additionally, SLC25A1 variants underlie two rare autosomal recessive neurometabolic/neuromuscular disorders, DL-2HGA and CMS2330,31,32. While cardiac disease is not part of the OMIM characterization of these disorders, there have been hints that SLC25A1 variants may also impact the heart. VSDs, atrial septal defects, patent ductus arteriosus, bicommissural aortic valve, and patent foramen ovale were reported in isolated cases of DL-2HGA due to SLC25A1 variants57,58,59. Our analyses of the PCGC dataset demonstrates a trend towards an association of ultrarare SLC25A1 variants (a limitation being one-stop only stop-gain variant alongside missense variants of uncertain significance identified) with pediatric CHD, and may suffer from type II error. While our SLC25A1 variant burden testing did not reach statistical significance, it is important to note that our studies of the Slc25a1 KO mouse model support neonatal lethality of Slc25a1+/- pups before P7. As such, predicted damaging SLC25A1 variants may be underrepresented in the PCGC cohort due to lethality of undiagnosed neonates. Further, the CHDs associated with ultrarare damaging SLC25A1 variants (VSDs, tetralogy of Fallot, and conotruncal defects) align with the malformations associated with 22q11.2DS. We also previously discovered that SLC25A1 affects neurodevelopment in 22q11.2DS24, and some DL-2HGA patients exhibit microcephaly57, a feature we also observed in Slc25a1-/- mouse embryos. Together, our work suggests that SLC25A1 is a nuclear DNA-encoded mitochondrial gene that can connect the neurological and cardiac pathologies of 22q11.2DS and DL-2HGA.
We have uncovered a novel pathway of metabolic regulation in which SLC25A1 regulates gene expression in the developing heart. We demonstrated that Slc25a1 is a dosage-dependent regulator of ventricular morphogenesis in the developing mouse embryo. Critically, our work in Slc25a1 KO mice has clinical relevance, as we identified a strong trend for pathogenic variants of human SLC25A1 to associate with CHD. Collectively, our work positions SLC25A1 as a mitochondrial transporter that mediates epigenetic regulation of gene expression in the developing heart and highlights a role for SLC25A1 in metabolic maturation in heart development. As SLC25A1-dependent regulation of gene expression may extend beyond metabolism, our work opens the door to a potential broader role for SLC25A1 in mitochondrial control of developmental programs. Finally, as SLC25A1 is a 22q11.2 gene, and our studies point to a role for SLC25A1 as novel CHD risk factor, this study may be an important first step in defining SLC25A1 as a connection between cardiac and neurodevelopmental pathologies in 22q11.2DS.
Methods
Animals
Slc25a1tm1a(EUCOMM)Wtsi KO-first mutant mice (Slc25a1+/-) were obtained from the Mutant Mouse Resource and Research Center at the University of California, Davis and maintained on a C57BL6/J background. Timed matings were established using 2–3-month-old Slc25a1+/- male and female mice, with the morning of an observed copulation plug established as E0.5. Experiments were conducted on male and female embryos collected at E10.5, E12.5, E14.5, E17.5, and E18.5. Genotyping of Slc25a1+/- mice as well as embryos from Slc25a1+/- intercrosses was conducted on genomic DNA isolated from embryonic yolk sacs or tail biopsies. All animal experiments were carried out with experimental protocols reviewed and approved by Emory University’s Institutional Animal Care and Use Committee in compliance with ethical regulations for animal use.
Histology and immunostaining
Hearts were fixed in 10% formalin and embedded in paraffin. Paraffin-embedded tissues were sectioned (6 µm), deparaffinized, rehydrated, stained with hematoxylin and eosin (H&E), and imaged using a Nanozoomer 2.0-HT whole-slide scanner (Hamamatsu). Myocardial trabecular and compact layer thicknesses were measured on H&E-stained sections using QuPath image analysis software60.
For immunofluorescence, paraffin-embedded sections were deparaffinized and rehydrated, and antigen retrieval was performed with citrate buffer (pH 6.0). The following primary antibodies were used: anti-myosin heavy chain (Developmental Studies Hybridoma Bank, MF20, 1:20), anti-SLC25A1 (Proteintech, 15235-1-AP, 1:100), and anti-ATP5A (Abcam, ab14748, 1:200). Secondary antibodies used were: donkey anti-mouse 488, and donkey anti-rabbit 594 (1:200, Cell Signaling Technologies). Slides were mounted with ProLong Gold Antifade Mountant with DAPI (Thermo Fisher Scientific) to counterstain nuclei or ProLong Gold Antifade Mountant (Thermo Fisher Scientific). Fluorescence imaging was conducted using a Keyence BZ-X810 system or a Leica Stellaris confocal microscope.
Whole exome sequencing and burden testing
Genomic analysis using the Pediatric Cardiac Genomic Consortium (PCGC) whole exome cohort followed published protocols61,62,63. Briefly, whole-exome DNA from blood or salivary samples was captured using the Nimblegen v.2 exome capture reagent (Roche) or Nimblegen SeqxCap EZ MedExome Target Enrichment Kit (Roche), followed by Illumina DNA sequencing as previously described61,62,63. The resulting sequencing data were processed at Yale University School of Medicine, and reads were mapped to the hg19 reference genome. Mapped reads were further processed using the GATK Best Practices workflows64, as previously described61. Single nucleotide variants and small indels were called with GATK HaplotypeCaller65. Further data filtering was performed using plink66, including removing individuals with low call rates, outlying heterozygosity rates, outlying relatedness rates, and sex discrepancy. A total of 3740 probands passed individual filtering, and there were a remaining 3401 complete trios. A case-control cohort was generated for all complete trios using pseudo-sibling controls. With this method, a proband’s genotypes are taken as the case, and a pseudo-control sample is created with all untransmitted alleles between the parents and the proband using plink67. Variants were filtered on call rate, Hardy-Weinberg equilibrium, and a high number of Mendel errors (≥3). Variants within SLC25A1 were extracted using the longest known canonical hg19 SLC25A1 variant bed file downloaded from UCSC TableBrowser on 4/5/2368. The remaining variants were annotated using wANNOVAR69. Loss of function (frameshift, splice site, start lost, stop gained, and stop loss) and missense variants with combined annotation-dependent depletion score >2070 and minor allele frequency <0.001 in all gnomAD71 populations were kept for burden analysis. An additional “ultrarare” variant set was created by keeping SLC25A1 variants that do not occur in the gnomAD exome database (v2.1.1, as of 4/12/23). Burden testing using the pseudo-sibling-controlled cohort was performed using kernel based adaptive cluster methods using WISARD72. The Bonferroni correction for multiple testing was used, establishing an alpha level of 0.0167.
Echocardiography
Mice were anesthetized with isoflurane (1.5%), and body temperature was maintained at 37°C. Echocardiography measurements were performed using a Vevo 2100 Imaging System (VisualSonics) equipped with a MS-400 transducer. M-mode and B-mode measurements were taken of the parasternal short- and long-axis views. Systolic and diastolic ventricular chamber dimensions as well as left and right ventricular wall thicknesses were assessed. Ejection fraction, fractional shortening, and stroke volume were calculated using VevoLab software.
Metabolic gene expression profiling
Transcriptomic profiling of expression of 768 metabolism-related genes was conducted using the NanoString nCounter Mouse Metabolic Pathways Panel. Briefly, total RNA was isolated from E17.5 hearts (n = 4/genotype) using a RNeasy Micro Kit (Qiagen). NanoString processing was completed by the Emory Integrated Genomics Core. RNA quality was assessed with the NanoString protocol. mRNA counts were normalized by expression of the housekeeping gene Tbp. Data were analyzed using Qlucore Omics Explorer Version 3.7(24) or GSEA v4.3.2. Differentially expressed mRNAs from QLUCORE were analyzed with the Metascape engine to determine ontology enrichment as well as representation of transcription factors using TRRUST. GSEA analysis was used to query the REACTOME database73.
Transmission electron microscopy
Mouse heart samples were collected from E17.5 embryos and fixed in 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.4). Hearts were subsequently postfixed in 1% osmium tetroxide and embedded in epoxy resin. Ultrathin sections (80–90 nm) were cut with a Leica EM CU6 microtome and counterstained with uranyl acetate and lead citrate. Sections were imaged on a JEOL JEM-1400 transmission electron microscope (Tokyo, Japan) equipped with a Gatan US1000 CCD camera (Pleasanton, CA). Mitochondrial morphology was quantified as previously described in ref. 74 using ImageJ software (NIH).
Electrophoresis and immunoblotting
For western blots, total protein extracts were prepared from embryonic hearts solubilized in radioimmunoprecipitation assay buffer supplemented with combined protease/phosphatase inhibitor cocktail (Thermo Fisher Scientific). Proteins were resolved on 10% SDS-PAGE gels (TGX stain free gels, BioRad), transferred to PVDF membranes, immunodetected with antibodies, and imaged using a ChemiDoc XRS+ system (BioRad). Primary antibodies used were: anti-SLC25A1 (Proteintech, 15235-1-AP, 1:1000), anti-TOM20 (Proteintech, 11802-1-AP, 1:1000), OXPHOS Blue Native WB Antibody Cocktail (contains antibodies against SDHA ATP5A, UQCRC2, NDUFA9, and COXIV; Abcam, ab110412, 1:500), anti-H3K9ac (Cell Signaling Technologies, 9649, 1:1000), anti-H3 (Cell Signaling Technologies, 4499, 1:1000), anti-VDAC (Proteintech, 55259-1-AP, 1:1000) and anti-GAPDH (Fitzgerald, 10 R-G109A, 1:10,000). Secondary antibodies used were alkaline phosphatase-linked goat anti-rabbit IgG (Cell Signaling Technologies, 7054), alkaline phosphatase-linked goat anti-mouse IgG (Cell Signaling Technologies, 7056), horseradish peroxidase-linked goat anti-mouse IgG (Cell Signaling Technologies, 7076), and horseradish peroxidase-linked goat anti-rabbit IgG (Cell Signaling Technologies, 7074). Blots were imaged using a ChemiDoc XRS+ System (BioRad) and quantified using Image Lab software (BioRad).
Blue native polyacrylamide gel electrophoresis
BN-PAGE and immunoblotting for mitochondrial respiratory complexes were performed using pooled E17.5 hearts (n = 6/genotype), as previously described75. 50ug mitochondrial extracts from the pooled hearts were solubilized with digitonin (Sigma Aldrich, 4 g/g digitonin/protein ratio) and separated on a NativePAGE 3–12% Bis-Tris polyacrylamide gradient gel (Invitrogen). Proteins were transferred to a PDVF membrane and immunoblotting for respiratory complexes was conducted with the Total OXPHOS Blue Native WB Antibody Cocktail (Abcam, ab110412, 1:250) at 4 °C overnight. The membrane was washed in TBST and incubated in horseradish peroxidase-linked anti-mouse IgG (Cell Signaling Technologies, 7076, 1:5000). The blot was developed using Clarity Max Western ECL Substrate (BioRad) and imaged using the BioRad Chemidoc XRS+ System.
Mitochondrial oxygen consumption
Mitochondrial oxygen consumption was conducted on embryonic hearts as previously described76. Briefly, hearts were harvested from E17.5 embryos. Hearts were gently homogenized on ice in isolation buffer (225 mM mannitol, 10 mM sucrose, 0.5 mM EGTA, 10 mM HEPES, pH 7.4). The heart homogenate was diluted in respiration buffer (70 mM mannitol, 25 mM sucrose, 20 mM HEPES, 120 mM KCl, 5 mM KH2PO4, 3 mM MgCl2, pH 7.4) containing 5 mM glutamate and 3 mM malate. Oxygen consumption was measured using an Oxytherm System (Hansatech). Basal ADP-stimulated respiration was initiated with the addition of 1 mM ADP, uncoupled respiration was induced with the addition of 0.3 µM FCCP, and nonmitochondrial oxygen consumption was assessed with the addition of KCN. Basal and maximal uncoupled mitochondrial oxygen consumption rates were obtained by subtracting KCN-nonmitochondrial respiration, followed by normalization to protein concentration measured by Bradford assay (BioRad).
mtDNA copy number analysis
Total DNA was prepared from snap-frozen embryonic heart tissue using a DNAeasy Kit (Qiagen). Four E17.5 hearts per genotype were pooled for each replicate (n = 4 replicates/genotype). Quantification of relative mtDNA copy number was conducted by measuring mtDNA/nDNA ratio using qPCR. Primers for mouse mtDNA (forward, CTAGAAACCCCGAAACCAAA; reverse, CCAGCTATCACCAAGCTCGT) and the β-2-microglobulin nuclear DNA (forward, 5’ ATGGGAAGCCGAACATACTG 3’; reverse, 5’ CAGTCTCAGTGGGGGTGAAT 3’) were used as previously described in ref. 77. qPCR was performed using LightCycler 480 SYBR Green I Master (Roche) on an Applied Biosystems QuantStudio 6 Flex Real-Time PCR system. Relative mtDNA copy number was calculated using the ΔΔC(t) method.
Gas chromatography and mass spectrometry (GC/MS)-based targeted metabolomics
Metabolites were isolated as described previously78. Briefly, frozen heart tissue sample were homogenized in 0.4 mL −80 °C cold MS-grade methanol, acetonitrile, water at a volume ratio of 40:40:20 (v/v/v/). Further disruption of tissue sample was achieved by freeze-thawing sample three-times. Extracts were centrifuged at 13,000 rpm for 5 min at 4 °C. The supernatant was transferred to clean glass tubes and D27-myristic acid (0.15 µg/µL, Cat No 366889, Sigma- Aldrich; St Louis, MO) was added as internal run standard. Vials were covered with a breathable membrane and metabolites were evaporated to dryness under vacuum. Dried samples were stored at −80 °C for further GC-MS analysis. For derivatization of polar metabolites frozen and dried metabolites from cell extractions were dissolved in 20 µL of Methoxamine (MOX) Reagent (2% solution of methoxyamine-hydrogen chloride in pyridine; Cat. No. 89803 and 270970; Sigma- Aldrich; St Louis, MO) and incubated at 30 °C for 90 min. Subsequently, 90 µL of N-tert-Butyldimethylsilyl-N-methyltrifluoroacetamide with 1% tert-Butyldimethyl-chlorosilane (TBDMS; Cat. No. 375934, Sigma- Aldrich; St Louis, MO.) was added and samples were incubated under shaking at 37 °C for 30 min. GC-MS analysis was conducted using an Agilent 8890 GC coupled with a 5977-mass selective detector. Metabolites were separated on an Agilent HP-5MS Ultra Inert capillary column (Cat.No. 19091S-433UI). For each sample, 1 µL was injected at 250 °C using helium gas as a carrier with a flow rate of 1.1064 mL/min. For the measurement of polar metabolites, the GC oven temperature was kept at 60 °C and increased to 325 °C at a rate of 10 °C/min (10 min hold) followed by a postrun temperature at 325 °C for 1 min. The total run time was 37 min. The MS source and quadrupole were kept at 230 °C and 150 °C, respectively, and the detector was run in scanning mode, recording ion abundance within 50 to 650 m/z.
Mathematical modeling of the myocardial metabolic adaptations in the embryonal heart
In silico simulations were performed using the metabolic network of the cardiomyocyte CardioNet41,78,79. Mathematical modeling can accurately predict the dynamics of cardiac metabolism in response to stress, and CardioNet can identify metabolic profiles and flux distributions. Flux balance analysis (FBA) allows to estimate flux rates by integrating experimental constraints on metabolites or proteins within a defined biochemical network. We specified flux constraints for extracellular metabolites reflecting the circulating metabolite composition in fetal plasma. Intracellular metabolites were constrained using experimentally determined metabolite abundance (GC-MS). Based on these constraints we first determined flux distributions (\({v}_{m}\)) under Slc25a1+/+ (control) conditions. We then calculated fold-changes (FC) for experimentally measured metabolite concentrations between control and experimental groups (Slc25a1+-+ and Slc25a1-/-) and used these fold-changes to further constrain fluxes (\({v}_{m}\)) for the synthesis and/or degradation of intracellular metabolites. We included fold-changes (FC) based on the assumption that changes in metabolite concentrations under experimental conditions are accompanied by a proportional increase or decrease in the respective flux for the metabolite pool. By using metabolite level changes (fold changes) to estimate flux rate changes (vFC), we imply that the altered steady-state concentrations of metabolites are reflected in the newly evolved flux state and potentially limit metabolic functions.
To reflect the unique growth requirements in the fetal heart, we defined a biomass function including the synthesis of structural proteins, membrane lipids and ATPase activity41. Linear problems were defined for experimental group to identify steady-state flux distributions that fulfill the applied constraints (e.g., substrate uptake and release rates), as well as changes in the metabolite pools. Regular FBA seeks to minimize the sum of all internal fluxes to fulfill applied constraints and the steady conditions41. However, in the context of a growing heart this condition may not yield correct flux estimations, because the maximization of biomass is not reflected in a regular minimization optimization problem. Instead, we defined the objective as a maximin problem, which maximizes the minimum objective (e.g., flux) for all a given set of internal constraints41
\({{{\rm{maximize}}}} \, z\)
subject to
where \({v}_{i}\) denotes the flux rate change through reaction \(i\), \({v}_{j}\) denotes the measured uptake or secretion rate through reaction \(j\), S is the stoichiometric matrix, \({v}_{i}^{(-)}\) and \({v}_{i}^{(\mp )}\) are flux constraints, and z that is an upper bound for each of the individual variables within the biomass function. Linear problems were implemented in perl and solved using the GUROBI LP solver80. The resulting fluxes were compared between groups using a two-way ANOVA followed by multiple comparisons analysis. Adjusted q-values were computed using the Bonferroni, Krieger and Yekuteli two-step method.
ChIP-qPCR and analyses of ChIP-seq data
ChIP experiments were conducted using the Magna ChIP HiSens Chromatin Immunoprecipitation Kit (EMD Millipore) with modifications. Briefly, E17.5 hearts were collected and pulverized with a mortar and pestle in liquid nitrogen. DNA–protein interactions were preserved with crosslinking buffer at continuous rotation for 20 min. The crosslinking reaction was stopped with addition of 2.5 M glycine. Following centrifugation (2000 g for 15 min) and washing with cold 1X PBS, samples were centrifuged again (2500 g for 10 min). Nuclei isolation buffer and sonication/ChIP/wash buffer were added to each sample for cell lysis and release of crosslinked protein–DNA. Samples were sonicated on ice with Misonix XL-2000 output 3 for 50 cycles of 15 s with a 45 s refractory period. ChIP was performed according to the manufacturer’s instructions. Antibodies used were: anti-H3K9ac (Sigma Aldrich, 17-658), and normal rabbit IgG (Sigma Aldrich, 12-370). Immunoprecipitated and input DNA was subjected to qPCR on a CFX96 Touch Real-Time PCR Detection System (BioRad) using SsoAdvanced Universal SYBR Green Supermix (Bio Rad 172-5270). The following primer sets were used to amplify targeted promoter regions: Pparg forward, 5’ CTGCGTAACTGACAGCCTAACC 3’ reverse, 5’ CTTGTGTGACTTCTCCTCAGCC 3’; Ndufa1 forward, 5’ CCCCGTTGGTGAATTTGTGG 3’ reverse, 5’ GCGGAGATGTGGTTCGAGAT 3’; Pdha1 forward, 5’ AGAACTAGGCCCTCAGACGA 3’, reverse, 5’CATGAGGAAGATGCTTGC CG 3’; Ndufs8 forward, 5’ AACCTGCAGAGTGACCTTGG 3’, reverse,5’ TTGAACCGCATC TGGGCC 3’; Cox4i1 forward, 5’ TCTGAAACGGACGGGCTTG 3’, reverse,5’ GACCGCCCTCTA CACCCT 3’; Ndufb2 forward, 5’ CGGGACTCGGGGAAGTGA 3’, reverse,5’ CTGGACCCGTCT TGTCTCTG 3’; Ndufb8 forward, 5’ CACTCACCCCATGCCATCAT 3’, reverse,5’ CAGAGTGA ACCGCGGAGAAG 3’; Cox5, forward, 5’ GCGTTGTTAGACTCCCACCA 3’, reverse, 5’CCACT CCGCGAAGTAACCTT 3’; Ndufb4 forward, 5’ GGGGTCGTTGTACTGAAGCA3’, reverse,5’ GC ACTTCCTGAGCCTGAAGG 3’; Uqcrq forward, 5’ GTCTTTCAGAGGCAGGGGAC 3’, reverse,5’ GCAACCAGGAGTTTGAGCAG 3’. Percent input was calculated using the ΔΔC(t) method81.
Statistics and reproducibility
The number of replicates for each experiment, as well as the numbers of mice/embryos used are denoted in figure legends. Unless stated otherwise, data are expressed as mean ± standard error of the mean. One-tailed Student’s t-test was used for comparisons of two groups and one-way ANOVA followed by post-hoc Dunnett’s test was used for multiple group comparisons tests. P < 0.05 were considered statistically significant and the following statistical significance indicators are used: ns denotes not significant, *P < 0.05; **P < 0.005.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Source data for all graphs and charts are provided in the figshare repository, https://doi.org/10.6084/m9.figshare.26455591.v1. NanoString data can be accessed through the NCBI Gene Express Omnibus repository with the accession number: GSE279295. Model figure created in BioRender. Kwong, J. (2023) BioRender.com/p34q043. Any remaining information can be obtained from the corresponding author upon request.
References
Patterson, A. J. & Zhang, L. Hypoxia and fetal heart development. Curr. Mol. Med. 10, 653–666 (2010).
Vempati, U. D., Torraco, A. & Moraes, C. T. Mouse models of oxidative phosphorylation dysfunction and disease. Methods 46, 241–247 (2008).
Piruat, J. I., Pintado, C. O., Ortega-Saenz, P., Roche, M. & Lopez-Barneo, J. The mitochondrial SDHD gene is required for early embryogenesis, and its partial deficiency results in persistent carotid body glomus cell activation with full responsiveness to hypoxia. Mol. Cell Biol. 24, 10933–10940 (2004).
Li, K. et al. Cytochrome c deficiency causes embryonic lethality and attenuates stress-induced apoptosis. Cell 101, 389–399 (2000).
Larsson, N. G. et al. Mitochondrial transcription factor A is necessary for mtDNA maintenance and embryogenesis in mice. Nat. Genet.18, 231–236 (1998).
Hance, N., Ekstrand, M. I. & Trifunovic, A. Mitochondrial DNA polymerase gamma is essential for mammalian embryogenesis. Hum. Mol. Genet. 14, 1775–1783 (2005).
Zhao, Q., Sun, Q., Zhou, L., Liu, K. & Jiao, K. Complex Regulation of Mitochondrial Function During Cardiac Development. J. Am. Heart Assoc. 8, e012731 (2019).
Baker, C. N. & Ebert, S. Development of aerobic metabolism in utero: requirement for mitochondrial function during embryonic and foetal periods. OA Biotechnol. 2, 16–22 (2013).
Persad, K. L. & Lopaschuk, G. D. Energy Metabolism on Mitochondrial Maturation and Its Effects on Cardiomyocyte Cell Fate. Front. Cell Dev. Biol. 10, 886393 (2022).
Liu, Y., Chen, H. & Shou, W. Potential Common Pathogenic Pathways for the Left Ventricular Noncompaction Cardiomyopathy (LVNC). Pediatr. Cardiol. 39, 1099–1106 (2018).
Choquet, C., Kelly, R. G. & Miquerol, L. Defects in Trabecular Development Contribute to Left Ventricular Noncompaction. Pediatr. Cardiol. 40, 1331–1338 (2019).
Towbin, J. A. & Jefferies, J. L. Cardiomyopathies Due to Left Ventricular Noncompaction, Mitochondrial and Storage Diseases, and Inborn Errors of Metabolism. Circ. Res 121, 838–854 (2017).
Finsterer, J. Noncompaction in Mitochondrial Disorders. Circ. Res. 121, e88–e89 (2017).
Kunji, E. R. S., King, M. S., Ruprecht, J. J. & Thangaratnarajah, C. The SLC25 Carrier Family: Important Transport Proteins in Mitochondrial Physiology and Pathology. Physiol. (Bethesda) 35, 302–327 (2020).
Iacobazzi, V., Lauria, G. & Palmieri, F. Organization and sequence of the human gene for the mitochondrial citrate transport protein. DNA Seq. 7, 127–139 (1997).
Majd, H., King, M. S., Smith, A. C. & Kunji, E. R. S. Pathogenic mutations of the human mitochondrial citrate carrier SLC25A1 lead to impaired citrate export required for lipid, dolichol, ubiquinone and sterol synthesis. Biochim Biophys. Acta Bioenerg. 1859, 1–7 (2018).
Morciano, P. et al. A conserved role for the mitochondrial citrate transporter Sea/SLC25A1 in the maintenance of chromosome integrity. Hum. Mol. Genet. 18, 4180–4188 (2009).
Sivanand, S., Viney, I. & Wellen, K. E. Spatiotemporal Control of Acetyl-CoA Metabolism in Chromatin Regulation. Trends Biochem Sci. 43, 61–74 (2018).
Fernandez, H. R. et al. The mitochondrial citrate carrier, SLC25A1, drives stemness and therapy resistance in non-small cell lung cancer. Cell Death Differ. 25, 1239–1258 (2018).
Wynne, M. E. et al. APOE expression and secretion are modulated by mitochondrial dysfunction. Elife 12, https://doi.org/10.7554/eLife.85779 (2023).
Gokhale, A. et al. Mitochondrial Proteostasis Requires Genes Encoded in a Neurodevelopmental Syndrome Locus. J. Neurosci. 41, 6596–6616 (2021).
McDonald-McGinn, D. M. et al. 22q11.2 deletion syndrome. Nat. Rev. Dis. Prim. 1, 15071 (2015).
Unolt, M. et al. Congenital heart diseases and cardiovascular abnormalities in 22q11.2 deletion syndrome: From well-established knowledge to new frontiers. Am. J. Med. Genet. A 176, 2087–2098 (2018).
Gokhale, A. et al. Systems Analysis of the 22q11.2 Microdeletion Syndrome Converges on a Mitochondrial Interactome Necessary for Synapse Function and Behavior. J. Neurosci. 39, 3561–3581 (2019).
Dickinson, M. E. et al. High-throughput discovery of novel developmental phenotypes. Nature 537, 508–514 (2016).
Tan, M. et al. Inhibition of the mitochondrial citrate carrier, Slc25a1, reverts steatosis, glucose intolerance, and inflammation in preclinical models of NAFLD/NASH. Cell Death Differ. 27, 2143–2157 (2020).
Geiger, T. et al. Initial quantitative proteomic map of 28 mouse tissues using the SILAC mouse. Mol. Cell Proteom. 12, 1709–1722 (2013).
Wang, M., Herrmann, C. J., Simonovic, M., Szklarczyk, D. & von Mering, C. Version 4.0 of PaxDb: Protein abundance data, integrated across model organisms, tissues, and cell-lines. Proteomics 15, 3163–3168 (2015).
Spielmann, N. et al. Extensive identification of genes involved in congenital and structural heart disorders and cardiomyopathy. Nat. Cardiovasc. Res. 1, 157–173 (2022).
Nota, B. et al. Deficiency in SLC25A1, encoding the mitochondrial citrate carrier, causes combined D-2- and L-2-hydroxyglutaric aciduria. Am. J. Hum. Genet. 92, 627–631 (2013).
Balaraju, S. et al. Congenital myasthenic syndrome with mild intellectual disability caused by a recurrent SLC25A1 variant. Eur. J. Hum. Genet. 28, 373–377 (2020).
Alzahrani, A. Y. B., Alghamdi, L. S. A., Alghamdi, H. A. M., Hassan, A. F. & Alsehemi, M. A. Congenital Myasthenic Syndrome Associated With SLC25A1 Gene Variant: The First Reported Case in Saudi Arabia. Cureus 15, e35808 (2023).
Du, Q., de la Morena, M. T. & van Oers, N. S. C. The Genetics and Epigenetics of 22q11.2 Deletion Syndrome. Front. Genet. 10, 1365 (2019).
Pediatric Cardiac Genomics, C. et al. The Congenital Heart Disease Genetic Network Study: rationale, design, and early results. Circ. Res 112, 698–706 (2013).
Hoang, T. T. et al. The Congenital Heart Disease Genetic Network Study: Cohort description. PLoS One 13, e0191319 (2018).
Hlouschek, J., Hansel, C., Jendrossek, V. & Matschke, J. The Mitochondrial Citrate Carrier (SLC25A1) Sustains Redox Homeostasis and Mitochondrial Metabolism Supporting Radioresistance of Cancer Cells With Tolerance to Cycling Severe Hypoxia. Front Oncol. 8, 170 (2018).
Zhou, Y. et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 10, 1523 (2019).
Glancy, B., Kim, Y., Katti, P. & Willingham, T. B. The Functional Impact of Mitochondrial Structure Across Subcellular Scales. Front Physiol. 11, 541040 (2020).
Kondadi, A. K., Anand, R. & Reichert, A. S. Functional Interplay between Cristae Biogenesis, Mitochondrial Dynamics and Mitochondrial DNA Integrity. Int. J. Mol. Sci. 20, https://doi.org/10.3390/ijms20174311 (2019).
Jakubke, C. et al. Cristae-dependent quality control of the mitochondrial genome. Sci. Adv. 7, eabi8886 (2021).
Karlstadt, A. et al. CardioNet: a human metabolic network suited for the study of cardiomyocyte metabolism. BMC Syst. Biol. 6, 114 (2012).
Consortium, E. P. An integrated encyclopedia of DNA elements in the human genome. Nature 489, 57–74 (2012).
Luo, Y. et al. New developments on the Encyclopedia of DNA Elements (ENCODE) data portal. Nucleic Acids Res. 48, D882–D889 (2020).
Auwerx, J., Cock, T. A. & Knouff, C. PPAR-gamma: a thrifty transcription factor. Nucl. Recept Signal 1, e006 (2003).
Zhou, L., Wang, Z. Z., Xiao, Z. C. & Tu, L. Effects of PPAR-gamma in the Myocardium on the Development of Ventricular Septation. Curr. Med Sci. 40, 313–319 (2020).
Johnson, D. T. et al. Tissue heterogeneity of the mammalian mitochondrial proteome. Am. J. Physiol. Cell Physiol. 292, C689–C697 (2007).
Johnson, D. T., Harris, R. A., Blair, P. V. & Balaban, R. S. Functional consequences of mitochondrial proteome heterogeneity. Am. J. Physiol. Cell Physiol. 292, C698–C707 (2007).
Garcia, D. & Shaw, R. J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 66, 789–800 (2017).
James, T. N. Apoptosis in congenital heart disease. Coron. Artery Dis. 8, 599–616 (1997).
Feng, Q. et al. Development of heart failure and congenital septal defects in mice lacking endothelial nitric oxide synthase. Circulation 106, 873–879 (2002).
Pexieder, T. Cell death in the morphogenesis and teratogenesis of the heart. Adv. Anat. Embryol. Cell Biol. 51, 3–99 (1975).
Martinez-Reyes, I. & Chandel, N. S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 11, 102 (2020).
Wellen, K. E. et al. ATP-citrate lyase links cellular metabolism to histone acetylation. Science 324, 1076–1080 (2009).
Stollberger, C. & Finsterer, J. Noncompaction is already known in DiGeorge anomaly from 22q11.2 deletion. Am. J. Med Genet A 155A, 662–663 (2011). author reply 664-665.
Madan, S. et al. Left ventricular non-compaction on MRI in a patient with 22q11.2 distal deletion. Am. J. Med Genet A 152A, 1295–1299 (2010).
Merscher, S. et al. TBX1 is responsible for cardiovascular defects in velo-cardio-facial/DiGeorge syndrome. Cell 104, 619–629 (2001).
Edvardson, S. et al. Agenesis of corpus callosum and optic nerve hypoplasia due to mutations in SLC25A1 encoding the mitochondrial citrate transporter. J. Med Genet 50, 240–245 (2013).
Prasun, P. et al. Expanding the Clinical Spectrum of Mitochondrial Citrate Carrier (SLC25A1) Deficiency: Facial Dysmorphism in Siblings with Epileptic Encephalopathy and Combined D,L-2-Hydroxyglutaric Aciduria. JIMD Rep. 19, 111–115 (2015).
Cohen, I. et al. A novel homozygous SLC25A1 mutation with impaired mitochondrial complex V: Possible phenotypic expansion. Am. J. Med Genet A 176, 330–336 (2018).
Bankhead, P. et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 7, 16878 (2017).
Homsy, J. et al. De novo mutations in congenital heart disease with neurodevelopmental and other congenital anomalies. Science 350, 1262–1266 (2015).
Jin, S. C. et al. Contribution of rare inherited and de novo variants in 2,871 congenital heart disease probands. Nat. Genet 49, 1593–1601 (2017).
Zaidi, S. et al. De novo mutations in histone-modifying genes in congenital heart disease. Nature 498, 220–223 (2013).
Van der Auwera, G. A. et al. From FastQ data to high confidence variant calls: the Genome Analysis Toolkit best practices pipeline. Curr. Protoc. Bioinforma. 43, 11 10 11–11 10 33 (2013).
McKenna, A. et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res 20, 1297–1303 (2010).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet 81, 559–575 (2007).
Yu, Z. & Deng, L. Pseudosibship methods in the case-parents design. Stat. Med 30, 3236–3251 (2011).
Navarro Gonzalez, J. et al. The UCSC Genome Browser database: 2021 update. Nucleic Acids Res 49, D1046–D1057 (2021).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res 38, e164 (2010).
Kircher, M. et al. A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet 46, 310–315 (2014).
Karczewski, K. J. et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 581, 434–443 (2020).
Lee, S. et al. WISARD: workbench for integrated superfast association studies for related datasets. BMC Med Genomics 11, 39 (2018).
Han, H. et al. TRRUST v2: an expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res 46, D380–D386 (2018).
Picard, M., White, K. & Turnbull, D. M. Mitochondrial morphology, topology, and membrane interactions in skeletal muscle: a quantitative three-dimensional electron microscopy study. J. Appl Physiol. (1985) 114, 161–171 (2013).
Jha, P., Wang, X. & Auwerx, J. Analysis of Mitochondrial Respiratory Chain Supercomplexes Using Blue Native Polyacrylamide Gel Electrophoresis (BN-PAGE). Curr. Protoc. Mouse Biol. 6, 1–14 (2016).
Beutner, G., Eliseev, R. A. & Porter, G. A. Jr. Initiation of electron transport chain activity in the embryonic heart coincides with the activation of mitochondrial complex 1 and the formation of supercomplexes. PLoS One 9, e113330 (2014).
Ghazal, N., Peoples, J. N., Mohiuddin, T. A. & Kwong, J. Q. Mitochondrial functional resilience after TFAM ablation in the adult heart. Am. J. Physiol. Cell Physiol. 320, C929–C942 (2021).
Karlstaedt, A. et al. Oncometabolite d-2-hydroxyglutarate impairs alpha-ketoglutarate dehydrogenase and contractile function in rodent heart. Proc. Natl Acad. Sci. USA 113, 10436–10441 (2016).
Aksentijevic, D. et al. Cardiac dysfunction and peri-weaning mortality in malonyl-coenzyme A decarboxylase (MCD) knockout mice as a consequence of restricting substrate plasticity. J. Mol. Cell Cardiol. 75, 76–87 (2014).
Gurobi Optimization, L. Gurobi Optimizer Reference Manual. (2023).
Lin, X., Tirichine, L. & Bowler, C. Protocol: Chromatin immunoprecipitation (ChIP) methodology to investigate histone modifications in two model diatom species. Plant Methods 8, 48 (2012).
Acknowledgements
J.Q.K. was supported by the Additional Ventures Single Ventricle Research Fund, the Department of Defense (0000063651), and the National Institutes of Health (R01-GM-144729). V.F. was supported by the National Institutes of Health (1RF1AG060285). A.K. was supported by the National Institutes of Health (R00-HL-141702). G.A.P. was supported by the National Institutes of Health (R01-HL144776). Transcriptomics for this study was supported in part by the Emory Integrated Genomics Core shared resource of Winship Cancer Institute of Emory University and the National Institutes of Health (NIH)/National Cancer Institute (P30CA138292). Electron microscopy for this study was supported by the Robert P. Apkarian Integrated Electron Microscopy Core, which is subsidized by the Emory University School of Medicine and the Emory College of Arts and Sciences. Additional support for electron microscopy was provided by the Georgia Clinical and Translational Science Alliance of NIH (UL1TR000454). Echocardiography for this study was supported by the Animal Physiology Core, which is subsidized by Emory University and Children’s Healthcare of Atlanta. Additional support was provided by the NIH Office of the Director (S10OD021748). Microscopy and imaging for this study was supported in part by the Emory University Integrated Cellular Imaging Core and Children’s Healthcare of Atlanta. The content is solely the responsibility of the authors and does not necessarily reflect the official views of funding sources.
Author information
Authors and Affiliations
Contributions
J.Q.K. and C.O. wrote the manuscript. J.N.P., C.O., N.G., A.G., F.S., J.T.G., M.E.B., and J.Q.K. performed experiments. A.K. conducted computational flux analysis and CardioNet modeling. J.Q.K., V.F., A.K., M.E.D., J.N.P., C.O., J.T.G., and G.A.P. analyzed the data. J.Q.K., J.N.P., and C.O. designed the study. J.Q.K. conducted experimental oversight.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Carrie Welch and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Dario Ummarino.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ohanele, C., Peoples, J.N., Karlstaedt, A. et al. The mitochondrial citrate carrier SLC25A1 regulates metabolic reprogramming and morphogenesis in the developing heart. Commun Biol 7, 1422 (2024). https://doi.org/10.1038/s42003-024-07110-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-024-07110-8
This article is cited by
-
Targeting mitochondrial transporters and metabolic reprogramming for disease treatment
Journal of Translational Medicine (2025)
-
The integral membrane protein smim4 modulates redox balance via malate compartmentalization in pancreatic cancer
Nature Communications (2025)
