
Abstract
In male reproduction, spermatogenesis and sperm maturation are critical for the production of normal sperm and offspring, yet the underlying molecular mechanisms remain largely elusive. Tex38 is a testis-enriched gene, and its deficiency results in oligoasthenoteratospermia (OAT) with aberrant epididymis, leading to male infertility in mice. Tex38 knockout (KO) sperm primarily exhibited neck bending deformities and functional abnormalities, including impaired fertilization. Proteomic analysis identified ADAM3 and its maturation-associated chaperones (CALR3, CLGN, and PDILT) as the most significantly altered proteins among the differentially expressed proteins (DEPs) in both sperm and epididymis of Tex38 knockout mice. GO analysis revealed that DEPs were primarily involved in sperm morphogenesis, motility, and fertilization. ARRDC5 was identified as a novel interacting protein of TEX38, and its deletion resulted in similar male infertility phenotypes as Tex38 deletion. Immunoprecipitation-mass spectrometry identified TEX38 and ARRDC5 interact with CLGN and PDILT. The interactions among TEX38, ARRDC5, PDILT, and CLGN were found to affect ADAM3 maturation, resulting in the failure of both Tex38−/− and Arrdc5−/− sperm to migrate to the oviduct. Overall, these findings establish TEX38 as an essential regulator of mammalian ADAM3-related migration, sperm formation, energy metabolism (ATP generation), sperm-egg binding and fertilization. TEX38 represents a potential target for diagnosis and treatment of male infertility and male contraception.
Similar content being viewed by others
Introduction
Infertility is a prevalent health issue affecting approximately 15% of couples worldwide1,2,3. A dramatic 62.3% decline in sperm concentration and total sperm count has been observed over the past 45 years, signaling a global crisis in male fertility4. The causes of male infertility are diverse, encompassing congenital, acquired, and idiopathic factors that disrupt spermatogenesis. Spermatogenesis is a meticulously orchestrated process involving spermatogonia mitosis, spermatocyte meiosis, and spermiogenesis. Spermiogenesis, the final stage, entails a series of morphological transformations, including nuclear condensation and elongation, acrosome biogenesis, cytoplasmic elimination, and flagellum formation5,6. Mature sperm undergo capacitation to navigate the female reproductive tract and ultimately fertilize the oocyte7. Any impairments during these processes can lead to azoospermia, oligozoospermia, asthenozoospermia, or teratozoospermia, collectively resulting in male infertility. Despite the identification of over 2300 genes implicated in spermatogenesis regulation, the genetic underpinnings and pathogenesis of male infertility remain elusive1.
The regulation of spermatogenesis involves a complex interplay of genes expressed in precise cell- and stage-specific patterns. Genes specifically expressed in the testis or gonads were first coined “testis expressed” (Tex) genes by Wang et al. in 20018. To date, 69 Tex genes have been identified in human or mouse models9. The unifying characteristic of Tex genes is their predominant or exclusive expression in the testis. Numerous studies have highlighted the critical roles of Tex genes in spermatogenesis and fertilization, as most are highly conserved among mammals, and their dysfunction leads to male infertility. One or more mouse models have been generated for 47 Tex genes to investigate their functions and mechanisms underlying male fertility.
To date, transgenic male mice have been found subfertile or infertile in Tex17, Tex18, Tex11, Tex12, Tex14, Tex15, Tex19, Tex19.1, Tex38, and Tex101 knockout models9. Our literature review of Tex genes revealed that Tex38 is the only one whose function and mechanism in spermatogenesis have not been studied. In 2023, Chen et al. profiled 3696 human testicular single-cell transcriptomes from 17 idiopathic NOA (iNOA) patients and identified three significantly downregulated candidate causal genes (CD164, LELP1, and TEX38)10. Alagundagi et al. also identified TEX38 by integrated molecular-network analysis11. These findings provide strong evidence for the important roles of TEX38 in male reproduction.
TEX38, also known as THEG4, is highly conserved and predominantly expressed in the testis. In the present study, we generated Tex38 knockout (KO) mice using the CRISPR/Cas9 system. Tex38-deficient males exhibited sterility due to delayed sperm release, increased epididymis weight, oligoasthenoteratospermia (OAT), impaired sperm function, progressively increased number of bent-neck sperm during the transit of epididymis, and failed fertilization. Furthermore, dramatic expression changes of mature ADAM3 and its associated chaperones were detected in Tex38-deficient sperm and epididymis through proteomic analysis. ARRDC5, a novel TEX38-interacting protein, displayed similar male infertility phenotypes to Tex38-deficient mice. Combined with IP-MS and proteomic analysis, we determined that TEX38 likely recruits ARRDC5 and other differentially expressed proteins (DEPs) to regulate axoneme assembly, mitochondrial sheath formation, ATP production and head-tail coupling apparatus (HTCA) during spermiogenesis. Collectively, in addition to migration associated with ADAM3, our findings demonstrates that TEX38 plays critical roles in multiple processes, including sperm formation, ATP production, sperm-egg binding, and fertilization.
Results
Tex38 is an evolutionarily conserved gene predominantly expressed in the testis
To explore the evolutionary history of Tex38, we initially utilized the Ensembl comparative genomics resources (https://asia.ensembl.org/index.html) to construct the phylogenetic tree of Tex38. We observed that Tex38 first appeared in amniotes and is mainly expressed in mammals (Fig. S1A). Alignment analyses (https://www.uniprot.org/) of TEX38 orthologs from mice, rats, bovines, chimpanzees, horses, macaques, and humans revealed high homology throughout evolution (Fig. S1B). TEX38 orthologs exhibited significant sequence identity, ranging from 70.47% to 95.15% (Fig. S1C). The FANTOM5 dataset in the Human Protein Atlas (https://www.proteinatlas.org/) demonstrated that TEX38 is predominantly expressed in the testis, with a moderate expression level in the epididymis (Fig. S1D). Additionally, human single-cell RNA sequencing data revealed specific expression of TEX38 in both early and late spermatids (Fig. S1E). Furthermore, transcriptomic analysis of mouse tissues (https://www.ncbi.nlm.nih.gov/) indicated that Tex38 is predominantly expressed in the testis (Fig. S1F). Consistent with human data, single-cell sequencing data from databases (https://smart-db.cn/home) showed that Tex38 is primarily expressed in round spermatids in mouse testes (Fig. S1G). To elucidate the expression pattern of the Tex38 gene in mice, we performed reverse transcription polymerase chain reaction (RT-PCR) analysis using cDNA samples from multiple organs. Tex38 mRNA was detected only in the testis, with no evident PCR product bands observed in other tissues (Fig. 1A). TEX38 protein was detected with predominant expression in the testis (Fig. 1C). Additionally, we analyzed the expression patterns of Tex38 mRNA and protein in mouse testes at different developmental stages. Tex38 mRNA was first detected at 21 days postpartum (dpp), while TEX38 protein was first detected at 28 dpp. Both mRNA and protein remained at high levels until adulthood (Fig. 1B, D), coinciding with the appearance of elongating spermatids. These results demonstrate that Tex38 is predominantly expressed in the mouse testis, and this specific expression pattern suggests a potentially important role for TEX38 in spermatogenesis.

A RT-PCR analysis of Tex38 mRNA in mouse tissues. Gapdh was used as an endogenous control. B RT-PCR analysis of Tex38 mRNA in the testis of mice at different ages. Gapdh was used as an endogenous control. C Western blot of TEX38 protein in mouse tissues. β-actin was used as a loading control. D Western blot of TEX38 protein in the testis of mice at different ages. β-actin was used as a loading control. E Schematic diagram of generating Tex38-knockout mouse model (Tex38−/−) by CRISPR/Cas9 system. F DNA genotyping analysis of mice by RT-PCR. Homozygous (Tex38−/−) 811 bp, heterozygous (Tex38+/−) 811 bp and 469 bp, WT (Tex38+/+) 469 bp. G Western blot showed that TEX38 was deleted in 8-week-old Tex38−/− testis compared to Tex38+/+. β-actin was used as the loading control. H, I The number of litter size in Tex38−/− and Tex38+/+ females and males mated with Tex38+/+ mice (n = 8). Student’s t-test. J–L Photographs of adult mice with Tex38+/+ and Tex38−/− showing external appearance and images of testes and epididymis. M–O Body weight, testis weight and testis weight to body weight ratio of Tex38+/+ and Tex38−/− mice at 8 weeks (n = 7). Student’s t-test. P, Q Epididymis weight and epididymis weight to body weight ratio of adult Tex38+/+ and Tex38−/− mice (n = 10). Student’s t-test. All error bars show SEM.
Due to limitations in antibody availability, direct detection of TEX38 localization within testicular tissue was not feasible. To elucidate its subcellular localization, we conducted an amino acid sequence analysis of TEX38, identifying a transmembrane region spanning amino acids 4-26. Subsequently, we constructed an EGFP-TEX38 plasmid encompassing the full-length TEX38 sequence and a mutant plasmid lacking the transmembrane domain (Fig. S2A). EGFP was fused to the N-terminus of the TEX38 protein. These plasmids were transfected into HeLa cells, and the distribution of the EGFP signal was observed following protein expression. The full-length EGFP-TEX38 protein exhibited predominant perinuclear localization with a punctate pattern, whereas both EGFP alone and the EGFP-TEX38 mutant protein were uniformly distributed throughout the cell (Fig. S2B). To further define the subcellular localization of the EGFP-TEX38 fusion protein, transfected HeLa cells were stained with fluorescent probes targeting the endoplasmic reticulum (ER), Golgi apparatus, mitochondria, and lysosomes. Significant co-localization between the EGFP-TEX38 fusion protein and probes for the ER and Golgi apparatus was observed in the perinuclear region, while minimal co-localization was detected with mitochondrial and lysosomal probes (Fig. S2C). These findings indicate that TEX38 plays a functional role in multiple cytoplasmic organelles, with emphasis on the ER and Golgi apparatus.
Deficiency of Tex38 causes male infertility
To further clarify the function of Tex38, we generated a Tex38−/− mouse model through the CRISPR-Cas9 genome editing system. Two sgRNAs were designed targeting exons 1 and 2 of Tex38 (Fig. 1E). Genotyping was performed by PCR (Fig. 1F). Western blot analysis confirmed the deletion of TEX38 protein (Fig. 1G). Fertility testing revealed that Tex38−/− male mice were completely infertile (Fig. 1H), whereas females exhibited normal fertility comparable to Tex38+/+ females (Fig. 1I). No significant differences were observed in the morphology, testis size, body weight, testis weight, or testis-to-body weight ratio between Tex38−/− and age-matched Tex38+/+ mice (Fig. 1J, K, M–O). These findings suggest that Tex38 deficiency has no effect on mouse growth and development. Interestingly, a subset of Tex38−/− mice displayed markedly enlarged epididymides compared to Tex38+/+ mice (Fig. 1L). Tex38−/− mice also exhibited a significantly increased epididymis weight/body weight ratio compared to Tex38+/+ mice (Fig. 1P, Q). Overall, these results indicate that Tex38 is essential for male fertility in mice and contributes to the regulation of epididymal weight.
Deficiency of Tex38 causes oligoasthenozoospermia in male mice
To investigate the role of Tex38 in spermatogenesis, histological examination was conducted on the caput, corpus, and cauda regions of Tex38+/+ and Tex38−/− epididymis (Fig. 2A). Hematoxylin and eosin (H&E) staining revealed that sperm within the epididymis of Tex38−/− mice exhibited abnormal morphology and reduced numbers, whereas the morphology of epithelial cells in the caput, corpus, and cauda epididymides appeared normal in Tex38−/− mice (Fig. 2A). Sperm count analysis revealed that compared to Tex38+/+, Tex38−/− mice exhibited a significant reduction in sperm abundance released from the cauda epididymis (Fig. 2B). Furthermore, reductions in both the proportion of motile spermatozoa and sperm with progressive motility were noted (Fig. 2C, D). Morphological analysis of cauda epididymal sperm revealed a significant increase in bent-neck spermatozoa in Tex38−/− mice (KO 63.3% vs WT 2.0%, P < 0.001), accompanied by other morphological abnormalities including spermatozoa with malformed heads (KO 26.8% vs WT 14.2%, P < 0.001) and multi-headed/flagellated (KO 5.5% vs WT 0.0%, P < 0.05) sperm (Fig. 2E, F). These aberrant morphological patterns were consistently observed through scanning electron microscopy (SEM) (Fig. 2G). To explore sperm defects in greater detail, sperm collected from the cauda epididymis were examined using transmission electron microscopy (TEM). In Tex38−/− spermatozoa, both the head and the midpiece of the tail were simultaneously enveloped within a membranous structure (Fig. 2H). Compared to Tex38+/+ mice, the cross-sections of sperm flagella from Tex38−/− mice exhibited clear evidence of axoneme damage, as well as disorganization of the mitochondrial sheath and outer dense fibers (Fig. 2H). Meanwhile, acrosome morphology was examined in Tex38+/+ and Tex38−/− sperm using PNA-FITC immunofluorescence staining (IF). A significant deviation from the normal acrosomal pattern was observed in Tex38−/− sperm, characterized by absent or abnormally located PNA-labeled acrosome signals. Notably, most sperm with neck bending displayed weak, diffuse acrosomal staining, with only 27.1% exhibiting normal PNA-positive signals (Fig. 2I, J). Given the abnormal head morphology (22.9%) observed in Tex38−/− sperm, DNA damage was assessed in epididymal sperm. A significantly higher proportion of comet-positive sperm was detected in Tex38−/− compared to Tex38+/+ mice (KO 27.1% vs WT 3.7%, P < 0.01), indicating increased DNA damage in Tex38−/− sperm (Fig. 2K, L). These findings collectively demonstrate that Tex38 deletion results in OAT in mice.

A H&E staining of the caput epididymis, corpus epididymis, and cauda epididymis from Tex38+/+ mice and Tex38−/− mice. B Count of the sperm from cauda epididymis (n = 3). Student’s t-test. C The percentage of motile spermatozoa in the cauda epididymis (n = 3). Student’s t-test. D The percentage of the sperm with progressive motility in the cauda epididymis (n = 3). Student’s t-test. E H&E staining of spermatozoa from the cauda epididymis. F The percentage of normal sperm, bent neck sperm, malformed head sperm, and multi-headed/flagellated sperm in Tex38+/+ and Tex38−/− cauda epididymis (n = 3). Student’s t-test. G Scanning electron microscopy (SEM) images of sperm from Tex38+/+ and Tex38−/− mice. H TEM analysis of mature sperm in the Tex38+/+ and Tex38−/− cauda epididymis. The red arrow indicates the absence and disorder of microtubules in the sperm flagella of Tex38−/− mice. I Peanut agglutinin (PNA) staining of epididymal sperm from Tex38+/+ and Tex38−/− mice. Scale bar is 25 μm. J Quantification of the percentage of PNA positive sperm in Tex38+/+ and Tex38−/− mice (n = 3, more than 200 sperm per replicate). Student’s t-test. K, L Comet assay for DNA fragmentation of epididymal sperm from Tex38+/+ and Tex38−/− adult mice (n = 9, more 500 sperm per replicate). Scale bar is 50 μm. Student’s t-test.
Tex38 deficiency affects spermiogenesis but may not affect spermatogonial proliferation or meiosis
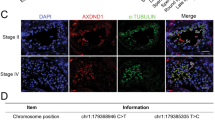
To investigate the role of Tex38 in mice, we further performed HE staining to analyze the tissue morphology of Tex38+/+ and Tex38−/− mice. No substantial differences were observed in the heart, liver, spleen, lungs, kidneys, and brain (Fig. S3A). These results demonstrate that Tex38 primarily functions within testicular tissue. To assess the cause of oligoasthenozoospermia induced by Tex38 deletion, we conducted a detailed analysis of the testicular histomorphology in Tex38−/− mice. Periodic Acid-Schiff (PAS) staining revealed that the number and diameter of seminiferous tubules in Tex38−/− mice were comparable to those in Tex38+/+ mice. Histological examination of 12 spermatogenic stages revealed elongated spermatids with delayed release during stages IX-X, along with failed individualization at the basal compartment of seminiferous tubules, which were not observed in Tex38+/+ mice. No significant abnormalities in cellular arrangement were detected in other spermatogenic stages (Fig. 3A). To clarify the impact of Tex38 on spermatogenesis, we quantified various types of germ cells at different stages. There was no significant difference in the number of germ cells in Tex38−/− testes compared to Tex38+/+ testes, except for an increase of unreleased elongated spermatids in stage IX-X seminiferous tubules (Fig. 3B).

A Different epithelium stages of seminiferous tubules in PAS-stained Tex38+/+ and Tex38−/− testes. The red dashed box indicates sperm with individualization failure, while the blue arrows indicate unreleased elongated sperm. Scale bar is 25 μm. B Spermatogenic cell count and statistical analysis of spermatogenic tubules at different stages. Leptotene (L), zygotene (Z), pachytene (P), diplotene (Di), round spermatid (Rst), elongated spermatid (Est), unreleased sperm (U). n = 4, Student’s t-test. C, D TUNEL staining of testis sections from Tex38−/− and Tex38+/+ mice (n = 3). Student’s t-test. Scale bar is 50 μm. E TEM images for testes from Tex38−/− and Tex38+/+ adult mice. Yellow arrows indicate abnormal vesicles. N nucleus, Ac acrosome, Mt manchette, MS mitochondrial sheath. Scale bar is 2 μm.
To determine the specific stage of spermatogenesis in which Tex38 functions, we further examined the expression of germ cell markers (VASA for germ cells, PCNA for spermatogonia, γ-H2AX for spermatocytes). There were no significant differences in terms of the expression and localization of these molecules in Tex38+/+ and Tex38−/− testes. Tex38−/− did not affect the number of various spermatogenic cells, the differentiation of spermatogonia, or the meiosis of spermatocytes (Fig. S3B–D). SOX9 staining and quantification also indicated no significant impact of Tex38 deletion on the number and localization of Sertoli cells (Fig. S3E, F). We performed TUNEL staining to analyze cell apoptosis in Tex38−/− testis, which revealed a significantly increased number of apoptotic cells than in Tex38+/+ testis (Fig. 3C, D). Further observation of the ultrastructure of various cells in testis tissues using transmission electron microscopy (TEM) showed that most cells in Tex38−/− testes exhibited no significant abnormalities. However, a few round spermatids with acrosome vacuolation and multinucleated cell clusters containing multiple sperm heads were observed. These clusters included sperm heads, dispersed acrosomes, sperm flagella, and multiple vacuoles (Fig. 3E). Collectively, Tex38 knockout primarily affects spermiogenesis, while potentially having no significant impact on spermatogonial proliferation and spermatocyte meiosis.
Progressive increase of bent-neck sperm during the transit in the epididymis of Tex38 −/− mice
To study the etiology of abnormal sperm in Tex38−/− males, spermatozoa were collected from the testis, caput epididymis, corpus epididymis, and cauda epididymis for analysis of morphological abnormalities. Abnormal sperm morphology was observed in Tex38−/− males, including multi-headed/flagellated, bent-neck, malformed head (Fig. 4A, B). Compared with Tex38+/+ mice, the percentages of abnormal spermatozoa from the testis, caput epididymis, corpus epididymis, and cauda epididymis of Tex38−/− mice were significantly increased (Fig. 4B). H&E staining of testicular spermatozoa within seminiferous tubules revealed no significant difference in the proportions of bent-neck spermatozoa between Tex38−/− and Tex38+/+ mice (Fig. 4C). While multi-headed/flagellated spermatozoa were absent in Tex38+/+ mice, they constituted 3.2% of spermatozoa in the testes of Tex38−/− mice, a proportion consistent with that observed in the epididymis, suggesting that these abnormal spermatozoa originated in the testis (Fig. 4C). Statistical analysis demonstrated a progressive increase in the percentage of bent-neck spermatozoa from the caput epididymis (22.9%) to the corpus epididymis (46.2%) and cauda epididymis (62.0%), indicating that normal spermatozoa released into the lumen of seminiferous tubules subsequently undergo transformation into bent-neck spermatozoa during epididymal transit from the caput to the cauda epididymis (Fig. 4C).

A H&E staining of sperm collected from testis, caput epididymis, corpus epididymis, and cauda epididymis of Tex38−/− and Tex38+/+ mice (n = 3). B, C Percentages of abnormal morphology sperm in testicular suspensions, caput epididymis, corpus epididymis, and cauda epididymis of Tex38−/− and Tex38+/+ mice (n = 3, more than 200 sperm per replicate). Student’s t-test. D The representative images of epididymides from Tex38+/+ and Tex38−/−mice after ligation for 4 days. The one side epididymal ducts were ligated at the corpus region, and the other epididymis was removed from the abdominal cavity and then placed back as a sham ligation control. E H&E staining was used to analyze the sperm of the caput epididymis and cauda epididymis after ligation for 4 days. EP epididymis. F The percentages of normal sperm in caput and cauda epididymis of Tex38−/− and Tex38+/+ mice after ligation for 4 days (n = 3). Student’s t-test. G The percentages of bent neck sperm in caput and cauda epididymis of Tex38−/− and Tex38+/+ mice after ligation for 4 days (n = 3). Student’s t-test. H The percentages of malformed head sperm in caput and cauda epididymis of Tex38−/− and Tex38+/+ mice after ligation for 4 days (n = 3). I The percentages of multi-headed/flagellated sperm in caput and cauda epididymis of Tex38−/− and Tex38+/+ mice after ligation for 4 days (n = 3). Student’s t-test. Data are presented as mean ± SEM. Student’s t-test. All data are mean ± SEM and dots represent values for individual mice.
To elucidate whether the observed changes were induced by an aberrant epididymal environment due to Tex38 deficiency or prolonged sperm storage, epididymal duct ligation was performed. The left corpus epididymis was ligated for four days in both Tex38−/− and Tex38+/+ mice to simulate epididymal sperm storage, while the contralateral epididymis served as a control (Fig. 4D). Four days post-ligation, both Tex38−/− and Tex38+/+ mice exhibited a substantial accumulation of spermatozoa in the caput region. Subsequent H&E staining analysis of sperm collected from the caput and cauda was conducted (Fig. 4E). Compared to the non-ligated Tex38−/− epididymis, the proportion of bent-neck sperm in the caput region of Tex38−/− mice increased significantly post-ligation, while the proportion of normal sperm was significantly decreased post-ligation, reaching a similar proportion to that observed in the cauda epididymis (Fig. 4F, G). Conversely, no significant changes were observed in the percentages of multi-headed/flagellate sperm or malformed head sperm in the Tex38−/− caput region four days post-ligation (Fig. 4H, I). These findings suggest a positive correlation between the formation of bent-neck sperm and sperm storage duration within the Tex38−/− epididymis. Consequently, we hypothesize that the presence of bent-neck sperm in Tex38−/− mice is primarily attributed to epididymal storage, regardless of storage in caput or cauda region.
Tex38 deficiency leads to sperm dysfunction and fertilization failure
To elucidate the underlying causes of male infertility in Tex38−/− mice, we comprehensively evaluated the functionality of Tex38−/− spermatozoa at various stages of fertilization, including acrosome reaction (AR), and zona pellucida (ZP) binding. To further induce the AR, capacitated sperm were exposed to the Ca2+ ionophore A23187 in vitro. Given that sperm undergoing the AR are not stained by PNA, this enabled quantification of AR occurrence. Unexpectedly, a substantial proportion of Tex38−/− sperm retained PNA staining post-ionophore treatment, with only 3.3% exhibiting the AR, significantly lower than the 70.8% observed in Tex38+/+ sperm (Fig. 5A, B). The fertilization capacity of Tex38−/− spermatozoa was assessed through in vitro fertilization assays. In contrast to Tex38+/+ sperm, which efficiently bound to the ZP, Tex38−/− sperm displayed a complete failure to bind to the ZP (Fig. 5C). Furthermore, ATP levels were markedly decreased in Tex38−/− sperm compared to Tex38+/+ controls (Fig. 5D). Consequently, Tex38−/− sperm demonstrated a notably diminished capacity to fertilize ZP-intact oocytes (Fig. 5E, F). Importantly, even after ZP removal, Tex38−/− sperm remained incapable of fertilizing oocytes, indicating a defect in sperm-oocyte fusion (Fig. 5G, H). In summary, these findings demonstrate that Tex38 deficiency results in a cascade of sperm functional abnormalities, including impaired ZP binding, acrosome reaction, and fertilization.

A Sperm from Tex38+/+ and Tex38−/− mice was induced by calcium ionophore A23187 supplementation and PNA staining was performed. Nuclei were counterstained with DAPI. Representative images were shown. Scale bar is 25 μm. B Quantification of the percentages of acrosome reaction (AR) in (A) (n = 3, more than 200 sperm per replicate). Student’s t-test. C In vitro analysis of sperm–egg binding. Sperm heads were stained with Hoechst 33342. Scale bar is 50 μm. D ATP contents in Tex38+/+ and Tex38−/− sperm (n = 3). Student’s t-test. E Zona pellucida (ZP) intact oocytes after 24 h of in vitro fertilization (IVF) with epididymal sperm from adult Tex38+/+ or Tex38−/− mice. Scale bar is 100 μm. F Quantification comparison of 2-cell rate following IVF of Tex38+/+ ZP intact oocytes with epididymal sperm from adult Tex38+/+ or Tex38−/− mice (n = 3). Student’s t-test. G ZP free oocytes after 24 h of IVF with epididymal sperm from adult Tex38+/+ or Tex38−/− mice. Scale bar is 100 μm. H Quantification comparison of 2-cell rate following IVF of Tex38+/+ ZP free oocytes with epididymal sperm from adult Tex38+/+ or Tex38−/− mice (n = 4). Student’s t-test. Data are mean ± SEM and dots represent values for individual mice.
The absence of TEX38 impairs the processing and maturation of ADAM3 and its associated chaperone proteins in epididymal sperm
To elucidate the molecular mechanisms underlying TEX38 deletion-induced male infertility, we performed proteomic analyses on proteins extracted from testis, sperm, and epididymis. The histogram illustrates the distribution of DEPs (fold change ≥1.5), revealing 1290 DEPs in Tex38−/− sperm and 445 DEPs in the epididymis, in contrast to only 37 DEPs identified in the testis (Fig. 6A and Supplementary Data 1). GO enrichment analysis revealed few enriched biological processes among testis DEPs in Tex38−/− mice, whereas DEPs in sperm and epididymis were both enriched in flagellated sperm motility, spermatogenesis, and sperm-ZP binding (Fig. 6B). Sperm DEPs were additionally enriched in other biological processes, including cilium movement and sperm axoneme assembly, while epididymis DEPs were enriched in lipid metabolic process, actin filament organization, and cilium movement involved in cell motility (Fig. 6B). Among all DEPs, CALR3 exhibited the most dramatic elevation in sperm following Tex38 deletion. As a molecular chaperone, CALR3 works synergistically with CLGN and PDILT to facilitate ADAM3 folding for sperm-ZP binding12,13,14,15,16. Consequently, we examined DEPs associated with ADAM3 folding. As depicted in Fig. 6D, F, mature ADAM3 was downregulated in Tex38−/− sperm and epididymis. Correspondingly, CALR3, CLGN, and PDILT protein expression was significantly increased in the Tex38−/− sperm and/or epididymis. Collectively, these findings suggest that TEX38 interacts with molecular chaperones to promote mature ADAM3 production for sperm-ZP binding.

Protein of adult mouse testis, sperm, and epididymis was extracted for proteomics analysis. A Histogram diagram shows the number of up-regulated and down-regulated differentially expressed proteins in the testis, epididymis, and sperm of Tex38−/− mice. B Top biological process from GO enrichment analysis of DEPs in Tex38−/− sperm, epididymis and testis. C Volcano plot of differentially expressed proteins between Tex38+/+ and Tex38−/− sperm. Red dots represent upregulated proteins, blue dots represent downregulated proteins, and gray dots represent proteins that were not differentially expressed (P < 0.05, |logFC | >1.5). D Protein expressions of ADAM3, CALR3, CLGN, PDILT in Tex38+/+ and Tex38−/− testis, epididymis, and sperm. E Protein expressions of TEX38, ARRDC5, ATPA14, DNAJB13, TEKT4, GK2, SPATA19, SUN5, and CETN1 in Tex38+/+ and Tex38−/− testis, epididymis, and sperm. F, G Quantitation of protein expressions in (D) and (E) (n = 3). Mann-Whitney U test.
Notably, flagellated sperm motility, sperm mitochondrial sheath assembly, and sperm-ZP binding were common to both sperm and epididymis DEPs. To validate these findings, Western blot analysis assessed protein expression in testis, epididymis, and sperm. Figure 6E showed that TEX38 protein expression was absent in the Tex38−/− testis, while no signaling was detected in the epididymis and sperm. Additionally, Western blot analysis revealed significant decreases in TEKT4, GK2, and SUN5 expression in both Tex38−/− epididymis and sperm, along with increased SPATA19 levels. ATP1A4 and DNAJB13 were significantly decreased in Tex38−/− sperm, while CETN1 was downregulated in KO epididymis (Fig. 6E, G). Collectively, these findings demonstrate that TEX38 deficiency not only affects sperm HTCA formation and flagellar assembly proteins but also leads to significant alterations in the maturation of ADAM3 and its associated chaperone proteins in both sperm and epididymis.
ARRDC5, a novel interacting protein of TEX38, deficiency of which leads to similar spermatogenesis defects to those in Tex38 −/− mouse
To investigate the mechanisms by which TEX38 functions in spermatogenesis and fertilization, we employed the STRING database (https://cn.string-db.org) to perform bioinformatic analysis on TEX38 and identify potential interacting proteins. Among these, ARRDC5 emerged as a candidate binding partner (Fig. 7A). We further validated the interaction between TEX38 and ARRDC5 using co-immunoprecipitation (Co-IP) in the HEK-293T cell line (Fig. 7B). Proteomic analysis revealed comparable ARRDC5 expression levels in the testes of Tex38−/− and Tex38+/+ mice; however, ARRDC5 was undetectable in Tex38−/− sperm. This finding was further validated by Western blot analysis (Fig. 7C). The observed interaction prompted a critical inquiry into the fundamental process governing their interplay during spermatogenesis. Accordingly, we first analyzed the expression profile of ARRDC5. Similar to TEX38, ARRDC5 exhibited predominant expression in the mice testis at both the mRNA and protein levels (Fig. 7D, E). Furthermore, the developmental stage-specific expression pattern of ARRDC5 in Arrdc5+/+ mice testis mirrored that of TEX38. Arrdc5 mRNA displayed weak expression from 7 dpp, with a significant increase observed from 28 dpp onwards. Protein expression initiated at 21 dpp, gradually increased and peaked in adulthood (Fig. 7F, G).

A Screen the interacting proteins of TEX38 in STRING database (https://cn.string-db.org). B ARRDC5 interacted with TEX38 in cultured HEK293T cells. C Protein expression of ARRDC5 in the Tex38+/+ and Tex38−/− testis, epididymis, and sperm. D RT-PCR analysis of Arrdc5 mRNA in mice tissue. Rps2 was used as loading control. E Western blot of ARRDC5 protein in mice tissue. β-actin was used as a control. F RT-PCR analysis of Arrdc5 mRNA in mice developing testis. G Western blot of ARRDC5 protein in mice developing testis. H Protein expressions of ADAM3, CALR3, CLGN, PDILT, TEX38 in Arrdc5+/+ and Arrdc5−/− testis and sperm. I Quantitation of protein expressions in (H) (n = 3). Mann-Whitney U test.
To explore the role of ARRDC5 in male reproduction, given its interaction with Tex38, we generated Arrdc5−/− mice using CRISPR/Cas-mediated genome engineering (Fig. S4A, B). A three-month fertility test revealed complete infertility in Arrdc5 −/−male mice, while female fertility remained unaffected (Fig. S4C). To explore the underlying spermatogenic defects in Arrdc5−/− mice, testes and epididymides from 8- to 10-week-old animals were collected for histological analysis. No significant differences were observed in testis size or testis-to-body weight ratio between Arrdc5−/− and age-matched Arrdc5+/+ mice (Fig. S4D, E). However, sperm counts and motility were significantly reduced in the cauda epididymis of Arrdc5−/− mice compared to Arrdc5+/+ controls (P < 0.05) (Fig. S4F, G). H&E staining of Arrdc5−/− testis sections showed apparently normal spermatogenesis (Fig. S4H), whereas sperm isolated from the cauda epididymis exhibited reduced abundance and various morphological abnormalities, including bent necks, malformed heads, and multiple heads or flagella (Fig. S4H, I). SEM confirmed the abnormal sperm morphology observed in Arrdc5−/− mice, which resembled that of Tex38−/− sperm (Fig. S4J). Additionally, Comet assay analysis revealed significant DNA damage in Arrdc5−/− sperm, indicating impaired sperm DNA integrity (Fig. S4K, L).
To further explore the underlying causes of abnormal sperm, we evaluated the 12 stages of the seminiferous cycle and observed unreleased sperm in stages IX-X of Arrdc5−/− testis (red dashed box), a phenotype similar to the delayed sperm release caused by Tex38−/− (Fig. S5A). To determine whether Arrdc5-deficient abnormal sperm originated in the epididymis, as in the case of Tex38, we compared sperm number and morphology in the testis, caput, corpus, and cauda epididymis of Arrdc5−/− and Arrdc5+/+ mice (Fig. S5B, C). Interestingly, while Arrdc5−/− testicular tissue exhibited a higher proportion of malformed heads (KO 24.8% vs WT 3.3%, P < 0.05) and multi-headed/flagellated sperm (KO 8.9% vs WT 0%, P < 0.05), there was no significant difference in bent-neck sperm compared to Arrdc5+/+ testis (KO 3.2% vs WT 4.3%, P > 0.05). However, the proportion of bent-neck sperm increased significantly upon entering the epididymis and remained elevated in the caput (KO 44.2% vs. WT 2.6%), corpus (KO 51.7% vs. WT 3.4%), and cauda (KO 49.6% vs. WT 2.0%) epididymis (P < 0.05). Similarly, the proportion of malformed heads and multi-headed/flagellated sperm showed significant increases and remained relatively stable compared to Arrdc5+/+ testis, caput, corpus, and cauda epididymis (P < 0.05). The proportion of different abnormal sperm types did not change with prolonged sperm maturation time in the epididymis.
Given the phenotypic similarities between Arrdc5−/− and Tex38−/− mice, we hypothesized that ARRDC5, analogous to TEX38, regulates ADAM3 expression and maturation. Western blot analysis of testicular and sperm samples from Arrdc5−/− mice revealed that while TEX38, ADAM3, CALR3, CLGN, and PDILT expression levels in the testes remained unchanged compared to wild-type controls, mature ADAM3 was significantly downregulated in Arrdc5−/− sperm (Fig. 7H, I). Conversely, CALR3, CLGN, and PDILT expression levels were upregulated in Arrdc5−/− sperm, mirroring the expression pattern observed in Tex38−/− mice (Fig. 7H, I). In summary, ARRDC5, an interacting protein of TEX38, contributes to spermiogenesis and ADAM3 maturation, phenotypes resembling those of Tex38−/− mice.
TEX38 and ARRDC5 interact with proteins involved in ADAM3 maturation, fertilization, protein transport, and spermiogenesis
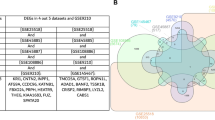
Given the similar male infertility phenotypes observed in Tex38−/− and Arrdc5−/− mice, we hypothesized that shared molecular mechanisms underlie these reproductive defects. To identify potential protein interaction partners of TEX38 and ARRDC5 involved in male infertility regulation, we performed IP-MS in WT testes. As depicted in Fig. 8A, we identified 384 binding proteins for TEX38 and 365 for ARRDC5 within the testis. Overlap analysis revealed 172 proteins shared by both TEX38 and ARRDC5 (Fig. 8A and Supplementary Data 2). Gene Ontology (GO) enrichment analysis indicated that binding proteins for both proteins were enriched in biological processes including protein transport, apoptotic process, spermatogenesis, vesicle-mediated transport, and proteolysis (Fig. 8B). The top cellular components were cytoplasm, mitochondrion, and ER (Fig. 8C). The most significant molecular functions were protein binding and identical protein binding (Fig. 8D). Meanwhile, we discovered that the ER molecular chaperones calmegin (CLGN) and protein disulfide isomerase-like, testis expressed (PDILT), which are associated with ADAM3 maturation, are also shared interacting partners of TEX38 and ARRDC5. Co-IP analysis further confirmed that CLGN and PDILT can interact with TEX38 and ARRDC5 in vitro (Fig. 8E, F). These findings reinforce our hypothesis that TEX38 and ARRDC5 may be involved in sperm maturation by influencing the maturation of ADAM3.

Protein of adult mouse testis was extracted and IP-MS was performed to identify TEX38 and ARRDC5 binding proteins. A Venn diagram shows the overlap of the TEX38 binding proteins and ARRDC5 binding proteins. B Top biological process, C top cellular component, and D top molecular function from GO enrichment analysis of TEX38 and ARRDC5 binding proteins. E, F Co-IP was performed in HEK-293T cells to validate the interactions of CLGN and PDILT with TEX38 and ARRDC5.
Specifically, these binding proteins participate in various aspects of spermatogenesis and fertilization, including: (1) flagellum assembly (e.g., ATP1A4, DNAJB13, SPACA9, and MNS1); (2) flagellar energy metabolism (e.g., TEKT4); (3) mitochondrial sheath assembly (e.g., GK2, SPATA19, and ARMC3); (4) centriole arrangement (e.g., CETN1); (5) fertilization (e.g., LYZL4, CCIN, SPACA3, and PPP3CC); (6) multinucleated giant cell or multi-headed/flagellated sperm formation (e.g., HADH and ZDHHC19); and (7) ER-to-Golgi vesicle-mediated transport (e.g., ERGIC3). Figure S6 illustrates that TEX38 and ARRDC5 interact with common binding partners critical for spermatogenesis and fertilization. Additionally, Co-IP validated the interaction between TEX38 and several of these putative binding partners (Fig. S7).
Tex38 −/− and Arrdc5 −/− spermatozoa failed to pass through the uterine-tubal junction (UTJ)
Given the impaired ADAM3 maturation in Tex38−/− and Arrdc5−/− mice, along with significant differential expression of associated ER chaperones and the shared interaction of TEX38 and ARRDC5 with CLGN and PDILT, we hypothesized that knockout of either gene would disrupt sperm migration through the UTJ. To validate this, we dissected the uterus and oviducts 3.5 h post-coitum, quantified sperm numbers in each segment, and performed histological analysis of the uterine-tubal junction (UTJ) to assess in vivo sperm migration. Approximately 2 × 10⁶ sperm were recovered from the uteri of females mated with WT males. In contrast, sperm counts in females mated with Tex38−/− or Arrdc5−/− males were significantly reduced compared to WT controls (Fig. 9B), this reduction may result from decreased sperm counts in Tex38−/− or Arrdc5−/− males. Nevertheless, 4×10⁵–8×10⁵ sperm were detected in the uteri of females mated with Tex38−/− or Arrdc5−/− males. To further examine sperm migration capacity, we quantified sperm in the oviducts. While 150–200 sperm were present in each oviduct of females mated with WT males, no sperm were detected in the oviducts of females mated with Tex38 Tex38−/− or Arrdc5−/− males (Fig. 9C). Histological analysis confirmed that Tex38−/− or Arrdc5−/− sperm were predominantly retained in the uterus post-ejaculation and rarely reached the oviducts, whereas WT sperm were primarily localized in the oviducts (Fig. 9A). These findings collectively indicate that sperm from Tex38−/− or Arrdc5−/− males exhibit defective migration from the uterus to the oviduct in vivo.

A H&E Cross sections of the UTJ of female mice mated with mice and Tex38−/− (or Arrdc5−/−) mice, respectively. The middle and right panels are rectangular areas with higher magnification for the corresponding left panel. The red arrow represents sperm. B Sperm count in the uterus after coitus. Data are presented as the mean ± SEM. (n = 4), Student’s t-test. C Sperm count in the oviducts after coitus. Data are presented as the mean ± SEM. (n = 4), Student’s t-test.
Tex38 −/− mice can be rescued by intracytoplasmic sperm injection
To probe the potential of intracytoplasmic sperm injection (ICSI) to overcome infertility in Tex38−/− mice, we performed ICSI using spermatozoa from both Tex38+/+ and Tex38−/− males. In the control group, sperm with rapid, progressive motility and normal morphology were selected for injection, while in the experimental group, spermatozoa exhibiting the typical bent-neck phenotype associated with Tex38 deletion were utilized. Three control groups, each comprising 19, 20, and 22 eggs, were injected with tailless sperm. After 24 h of culture, 19, 20, and 22 cells developed into two-cell embryos, and subsequently, 17, 18, and 21 blastocysts formed after 96 h. Similarly, three experimental groups of 17, 17, and 16 eggs were injected. Following 24 h, 12, 15, and 15 two-cell embryos were obtained, with subsequent development into 7, 13, and 12 blastocysts after 96 h (Fig. 10A). Importantly, the rates of two-cell embryo and blastocyst formation from Tex38−/− male mice were not significantly different from those derived from control mice (Fig. 10B). These findings suggest that Tex38 deletion does not overtly impact the development of early embryos produced via ICSI. To further assess the efficacy of ICSI in rescuing Tex38-associated infertility, two-cell embryos were transferred into pseudopregnant female mice, and successfully obtained the healthy developing pups (Fig. 10C). Genotyping of these offspring through PCR amplification of mouse tail DNA confirmed a heterozygous genotype in all cases (Fig. 10D). These results substantiate that ICSI can effectively overcome male infertility caused by Tex38 deficiency.

Abnormal spermatozoa from cauda epididymis of Tex38−/− mice were selected for ICSI. A Two-cell and blastocyst embryos after ICSI. The oocytes were fertilized by the sperms from Tex38−/− and Tex38+/+ mice after ICSI. B The proportion of 2-cell and blastocyst embryos after ICSI in Tex38−/− and Tex38+/+ mice. Data are mean ± SEM and dots represent values for individual mice (n = 3). C The offspring of the Tex38+/− and Tex38+/+ mice were born and grew healthy after ICSI. D Genotyping for the newborn pups of Tex38+/− and Tex38+/+ mice by RT-PCR. E Diagram of proposed action of Tex38. This figure was originally created by the authors using PowerPoint.
Discussion
Tex38 is highly conserved among mammals and is downregulated in iNOA germ cells10, suggesting its potential as a causal gene for iNOA. Recently, Kaneda et al. reported that Tex38, which is involved in the sperm head morphogenesis, is essential to male fertility in mice17. Our findings demonstrate that Tex38 is a testis-enriched gene whose deletion results in OAT accompanied by epididymal abnormalities, leading to male infertility in mice. While fertilization defects in Tex38−/− sperm can be overcome through ICSI to produce live offspring, our study provides novel evidence for the potential treatment of OAT caused by gene mutations, including those affecting TEX38.
Kaneda et al. demonstrated that ZDHHC19, an interacting partner of TEX38, palmitoylates ARRDC517. We have confirmed the interaction between ARRDC5 and TEX38 using IP-MS and Co-IP. Previous studies have identified ARRDC5 as an essential regulator of spermiogenesis18,19. In both Tex38−/− mice and Arrdc5−/− mice, no significant difference in testis weight was observed, suggesting minimal or no alteration in germ cell count within the testis20. Furthermore, no significant change in the abundance of PCNA+ cells21, γ-H2AX+ cells22, SOX9+ cells23, and VASA+ cells24 was observed in the Tex38−/− testis, indicating normal mitosis and meiosis. We found that Tex38 deletion in male mice caused OAT and defective fertilization, similar to Arrdc5 deletion. While the recent study has proved low sperm motility, increased abnormal sperm morphology and defective fertilization in mice with genetic inactivation of Tex38, the number change of sperm was not mentioned17. Taken together, interaction between TEX38 and ARRDC5 and similar phenotypes between their KO mice suggest that TEX38 and ARRDC5 may partly share the same downstream targets in the regulation of sperm morphogenesis and functions.
It is now understood that sperm produced in the testis undergo maturation during epididymal transit, acquiring motility and fertilization capacity. Epididymal dysfunction is associated with impaired sperm maturation and male infertility25. Aberrant sperm parameters and epididymal morphology prompted abnormalities in sperm and epididymis of Tex38−/− mice. While a low proportion of multi-headed/flagellate sperm was observed in the Tex38−/− testicular fluid, most sperm exhibited normal morphology. Notably, an increasing prevalence of morphological abnormalities, including bent necks, malformed heads, and multi-heads/flagella, emerged during post-testicular maturation in the Tex38−/− epididymis, resembling findings in Akap3 KO mice26. Consistent with the observation that flagella destabilization occurs upon sperm storage in the epididymis of Dnah17 mutant mice27, we found an increase in bent-neck sperm following 4 days of caput epididymis ligation. Similarly, Kaneda et al. also found gradually increased bent-neck sperm from caput to cauda epididymides of Tex38−/− mice17, suggesting defective sperm maturation within the abnormal microenvironment of the Tex38 deficiency epididymis.
We found that bent-neck sperm constituted the most prevalent abnormal sperm morphology in both Arrdc5−/− and Tex38−/− mice. Due to the abnormal cytoplasmic droplets at Tex38−/− sperm heads, Kaneda et al. anticipated that the increase of bent-neck sperm in Tex38−/− mice is attributed to the impaired cytoplasm removal17. The deletion of Zdhhc19, an interacting protein of TEX38, also showed bent-neck sperm and residual cytoplasm in the sperm heads. Moreover, Kaneda et al. proved that ZDHHC19 interacts with ARRDC5. Arrdc5−/− sperm with enlarged heads were wrapped in residual cytoplasm18. Therefore, TEX38 interacts with ZDHHC19 to form a stable complex containing ARRDC5, which may regulate proteins essential for cytoplasm removal in spermiogenesis. Differently, Liu et al. proposed that the bent-neck sperm phenotype in Arrdc5−/− mice arises from disrupted interaction between ARRDC5 and SUN5, accompanied by reduced SUN5 expression19. SUN5, a protein anchoring the sperm head to the tail, is the first identified pathogenic gene associated with acephalic spermatozoa syndrome. SUN5 deficiency causes detachment of the HTCA during spermatid elongation28,29. The HTCA is crucial for connecting the sperm head to the flagellum during rapid movement. Defects in HTCA can lead to sperm head detachment from the tail, resulting in acephalic spermatozoa30. Our findings corroborate these observations, demonstrating decreased expression of both ARRDC5 and SUN5 in the Tex38 deletion sperm. Centrioles are components of the HTCA, and can undergo organized rearrangement to facilitate the functional connection between the sperm nucleus and flagellum. Cetn1 deletion has been linked to centriole rearrangement failure in the basal-body-nucleus connection31. Our study revealed interactions between both TEX38 and ARRDC5 with CETN1 and detected CETN1 downregulation in the Tex38−/− epididymis. Collectively, these findings suggest the formation of a protein complex comprising TEX38, ARRDC5, SUN5, and CETN1, essential for HTCA structure. Tex38 deletion may disrupt protein interactions and expression within this complex, including ARRDC5, SUN5, and CETN1, leading to HTCA destabilization and subsequent bent-neck sperm formation during transit through the abnormal epididymal microenvironment.
Impaired HTCAs are consistently associated with defects in sperm tail structure, as observed in acephalic spermatozoa30, suggesting formation of HTCA and flagellum might be coupled to each other. The axoneme, a 9 + 2 arrangement of microtubule doublets, and the mitochondrial sheath are critical flagellum components essential for sperm propulsion. Our findings revealed disrupted axonemes and mitochondrial sheaths in both Tex38−/− and Arrdc5−/− sperm. However, Kaneda et al. did not observe clear abnormalities in the flagellar of Tex38−/− sperm17. Several co-binding proteins of TEX38 and ARRDC5 play crucial roles in axoneme formation, such as DNAJB13, ATP1A4, and MNS1. DNAJB13, a coupling apparatus protein located in the radial spokes of the axoneme, exhibits sperm tail defects such as coiled, short, and irregular flagella upon mutation32. Atp1a4 deficiency induces a bend at the mid-principal piece junction of the flagellum, accompanied by disrupted axoneme structure33. Mns1 KO males are sterile due to complete disruption of the 9 + 2 microtubule doublets arrangement and outer dense fibers in the flagellum34. TEX38 may recruit ARRDC5, DNAJB13, ATP1A4, and MNS1 to regulate axoneme assembly by influencing the development of radial spokes, 9 + 2 microtubule doublets, and outer dense fibers. Furthermore, co-binding proteins involved in mitochondrial sheath structure, including GK2 and SPATA19, were identified. Gk2 deletion in mice disrupts mitochondrial sheath formation and ATP production, compromising sperm motility and leading to male infertility35. Spata19 deletion results in male sterility due to damaged mitochondrial sheaths and reduced ATP production36. Tex38 deficiency may induce detachment of the GK2 and SPATA19 protein complex, leading to disorganized mitochondrial sheaths, reduced ATP production, and impaired sperm motility and fertility. Additionally, TEKT4 may contribute to the significant decrease in ATP observed in Tex38−/− sperm, as Tekt4 deletion induces increased ATP consumption due to ineffective flagellar strokes37. In addition to interacting proteins, proteomic analysis revealed that DEPs in Tex38−/− sperm were enriched in sperm axoneme assembly and flagellated sperm motility. The enriched proteins, including ATP1A4, DNAJB13, IQUB, and IFT14038 for axoneme formation, and GK2, SPATA19, ARMC339, TBC1D2140 for mitochondrial sheath development, exhibited altered expression levels in Tex38−/− sperm. These findings further support the impact of Tex38 deletion on sperm tail assembly. Collectively, these results suggest that TEX38 interacts with flagellum-associated proteins to form a functional complex that regulates flagellum development by modulating the expression of proteins involved in axoneme formation, mitochondrial sheath structure, and energy metabolism.
After traversing the entire epididymal duct, immature sperm produced in the testis acquire full fertilization competence. They undergo capacitation within the female reproductive tract, a process essential for subsequent ZP binding, acrosome reaction (AR), and sperm-egg fusion. Our findings revealed that TEX38 absence induces defective ZP binding, impaired AR, and failure of in vitro fertilization (IVF), which is consistent with the results of the recent study17. Notably, similar defects were observed in Arrdc5−/− mice, although some sperm could successfully fertilize oocytes following ZP removal18. Given the highly condensed sperm nuclei and removal of excess cytoplasm, the epididymal sperm is transcriptionally and translationally inert, requiring proteins secreted by the epididymal epithelium for maturation41. Proteomic data indicated that DEPs of Tex38−/− sperm and epididymis were both enriched in sperm-ZP binding. Previous studies have demonstrated that SPA17 expression is crucial for AR, with increased translational levels observed in patients with high sperm DNA fragmentation index42. Our study revealed decreased SPA17 expression in both Tex38−/− sperm and epididymis, potentially contributing to defective AR. ARSA, a lysosomal enzyme secreted by epididymal principal cells into the lumen, binds to the sperm head for sperm-ZP binding43. Caballero et al. reported that epididymosomes transfer GLIPR1L1 to spermatozoa in the epididymis, facilitating sperm-ZP binding44. We observed downregulated ARSA and GLIPR1L1 expression in both Tex38−/− sperm and epididymis. Additionally, TEX38 and ARRDC5 interact with LYZL4, SPACA3, and CCIN, proteins critical for sperm-ZP binding. These findings suggest that TEX38 regulates sperm-ZP binding by influencing the supply of ZP-binding associated proteins from the epididymal epithelium and their interactions with TEX38. ACTL7A, an acrosomal protein, is essential for normal acrosome formation. ACTL7A mutations result in malformed acrosomes, leading to defective ZP binding, fertilization failure during IVF, and ICSI45. We detected downregulated ACTL7A expression in both sperm and epididymis, potentially explaining the malformed acrosomes in Tex38−/− mice. However, the successful rescue of defective fertilization in TEX38-deficient mice through ICSI suggests that aberrant ACTL7A expression may not affect the normal haploid genome of Tex38−/− sperm. Epididymosomes, membrane-bound vesicles produced by the epithelium, transfer and insert proteins into the sperm membrane46,47. Our identification of certain DEPs on the sperm surface supports the hypothesis that Tex38−/− epididymis exhibits disordered protein secretion and transfer to the sperm surface, hindering the acquisition of capacitation, ZP binding, AR, and fertilization, ultimately leading to male infertility.
Sperm plasma membrane undergoes significant modification during epididymal transit. Post-translational modifications of sperm plasma membrane proteins, such as proteolytic processing, are essential for sperm maturation48. ADAM3, a glycosylated membrane protein located in spermatids and sperm, is initially synthesized as a 110 kDa precursor in round spermatids and subsequently processed into a mature 42 kDa form during epididymal transit. The mature ADAM3 resides on the sperm head surface and is removed during the AR. Disruption of Adam3 impairs sperm-ZP binding and sperm migration from the uterus to the oviduct12,49,50. Previous studies have identified mutations in 16 genes and gene clusters (Ace, Adam1a, Adam2, Calr3, Clgn, Cmtm2a/Cmtm2b, Cst family, Pate8-Pate10, Pdilt, Pmis2, Rnase10, Prss37, Prss55, Tex101, Tmprss12, Tpst2) that result in male infertility phenotypes similar to those observed in Adam3 deletion51. Our findings revealed normal ADAM3 precursor expression in the testis but reduced mature ADAM3 in Tex38−/− sperm, suggesting that Tex38 deficiency may prevent ADAM3 maturation, leading to impaired sperm-ZP binding, akin to the effects of deficiencies in the aforementioned 16 genes and gene clusters. ADAM3 is co-translationally translocated into the ER of round spermatids, where molecular chaperones facilitate glycoprotein folding and transfer to the sperm membrane. CALR3, CLGN, and PDILT are testis-specific ER chaperones crucial for ADAM3 maturation. They form a molecular chaperone complex that folds ADAM3 in the ER and transfers the mature protein to the sperm surface for ZP binding and oviduct migration14,16. Our observation that TEX38 localizes to the ER in Hela cells and interacts with CALR3 suggests its potential role as a molecular chaperone within this complex. Interestingly, dramatic increase of CALR3, CLGN, and PDILT expression in Tex38−/− sperm indicates a potential compensatory mechanism. While TEX38 deficiency disrupts ADAM3 folding in the ER, increased activity of other chaperones, including CALR3, CLGN, and PDILT, may counteract this effect, preserving proper sperm-ZP binding and oviduct migration. However, despite elevated levels of these chaperones, defective fertilization and failed migration of sperm into oviduct persists in Tex38−/− mice, suggesting that TEX38 may play a dominant role in the chaperone complex regulating glycoprotein folding essential for fertilization and migration.
Understanding how genes regulate sperm production is vital to our understanding of spermatogenesis and male infertility. We show Tex38 is required for normal sperm structure and function in mice. TEX38 interacts with functional proteins, including ARRDC5, to form a protein complex that may influence sperm HTCA formation, flagellum assembly, and fertilization (Fig. 10E). TEX38 may disrupt ADAM3 maturation during sperm maturation by up-regulating ER chaperones. Taken together, we identify TEX38 as an essential regulator of sperm morphogenesis and diverse functions including the migration of sperm into oviduct, which suggests it as a potential target for diagnosis and treatment of male infertility, and male contraceptive drug development.
Methods
Ethics statement
All animal experiments were conducted in accordance with the guidelines established by the Yangzhou University Animal Ethics Committee and were approved by the Jiangsu Administrative Committee of Laboratory Animals (permit number SYXK (SU) 2017-0044).
Generation of a knockout mouse model
Knockout mouse models of C57BL/6J background were generated by Cyagen Biosciences (CA, USA) using CRISPR/Cas-mediated genome engineering. Briefly, single guide RNAs were designed to target exons 1 and 2 of Tex38 (NM_029196.1) and exons 1 and 3 of Arrdc5 (NM_029799). Genotype was determined using specific primers by RT-PCR (Table S2). The animals were maintained at a constant temperature of 25 ± 1 °C under a 12-hour light/dark regimen, with illumination occurring between 06:00 and 18:00. During the entire experimental duration, they had unrestricted access to standard laboratory feed and drinking water. Euthanasia was performed using carbon dioxide asphyxiation, subsequently followed by neck dislocation.
Antibody preparation
The preparation of antibodies was performed according to previously established methods. Specifically, the full-length open reading frames of mouse Tex38 and Arrdc5 genes were amplified by PCR using specific primers (Table S2) and cloned into the pET30a (+) expression vector. Prokaryotic protein expressions were induced in Escherichia coli using Isopropyl beta-D-thiogalactopyranoside (IPTG) (Solarbio, China). These recombinant proteins were purified, refolded, and used to immunize New Zealand White rabbits, leading to the generation of polyclonal anti-Tex38 and anti-Arrdc5 antibodies.
RNA isolation, RT-PCR, and qPCR
Total RNA was extracted from tissues using TRIzol® Reagent (Invitrogen, USA). For each sample, 1 µg of RNA was reverse transcribed into cDNA using a cDNA Reverse Transcription Kit (Takara, Japan). Specific primers and Taq polymerase were employed for PCR amplification of cDNA. Quantitative real-time PCR (qPCR) was performed on a QuantStudio™ 3 Real-Time PCR System (Applied Biosystems, USA) using TB Green® Premix Ex Taq™ II (Takara, Japan). Gapdh/Rps2 served as an endogenous control. Primer sequences are listed in Table S2.
Fertility assessment
To assess male fertility, 8- to 10-week-old Tex38−/− and Tex38+/+ males were mated with WT females at a ratio of 1:2 for three months (n = 8). For female fertility assessment, one Tex38−/− or Tex38+/+ female was housed with one WT male for three months (n = 8). Mouse fertility was determined by calculating the average number of pups per litter.
Preparation of testicular cells and histological analysis
The testes were removed from adult mice and incubated for 32 min at 32 °C in a 15 ml centrifuge tube with 5 ml DMEM containing 0.5 mg/mL collagenase IV and 1.0 μg/mL DNase I (Sigma Aldrich). Then, after 5 min at 1000 rpm, the testes were washed once with phosphate-buffered saline (PBS), and then the supernatant was discarded. The pellet was resuspended with PBS and then mounted on positively charged glass slides, and the cells were fixed with 4% PFA.
For H&E staining, tissue sections were stained with hematoxylin and eosin procured from Beyotime (China). For PAS staining, a Schiff’s kit obtained from Solarbio (China) was employed according to the manufacturer’s instructions. Sections were stained with Schiff’s solution under dark conditions, followed by hematoxylin nuclear counterstaining. Images were captured using an 80i microscope (Nikon, Japan).
Scanning electron microscopy and transmission electron microscopy
For SEM, sperm was extracted from the epididymis and washed twice with phosphate buffer before preservation in 2.5% glutaraldehyde overnight. Samples were dehydrated in a graded ethanol series, dried, and gold-coated. Imaging was performed using a GeminiSEM 300 scanning electron microscope (Carl Zeiss, USA). For TEM, testes and sperm were dissected and fixed in 2.5% (v/v) glutaraldehyde in 0.1 M cacodylate buffer at 4 °C overnight. Following cleaning with 0.1 M cacodylate buffer, samples were immersed in 1% OsO4 at 4 °C for one hour. After dehydration in a graded acetone series, samples were embedded in resin. Ultrathin sections (60 nm) were cut and stained with lead citrate and uranyl acetate. Images were acquired using an HT7800 transmission electron microscope (HITACHI, Japan).
Immunofluorescence staining
For IF staining of paraffin sections, antigen retrieval was performed using sodium citrate recovery solution. For IF staining of cells, HeLa cells were fixed with 4% paraformaldehyde and permeabilized with 0.3% Triton X-100. Following blocking with 1% bovine serum albumin, samples were incubated with primary antibodies (Table S1) at 4 °C overnight, followed by incubation with a secondary antibody at room temperature for 1 hour. To assess TUNEL staining, the TUNEL BrightRed Apoptosis Detection Kit (Vazyme, China) was utilized according to the manufacturer’s instructions to identify apoptotic cells with double-stranded DNA breaks. Images were captured instantaneously using a Scope AI microscope (Carl Zeiss, USA) and an AXR confocal microscope (Nikon, Japan).
Western blot
Sperm-removed epididymis and other protein samples were extracted using RIPA lysis buffer. Protein concentrations were determined using the BCA protein assay kit (Solarbio, Beijing, China). Proteins were separated by SDS-PAGE and subsequently transferred to polyvinylidene fluoride membranes. Membranes were blocked with 5% non-fat milk and incubated overnight at 4 °C with primary antibodies (Table S1). Secondary antibodies were incubated at room temperature for 1.5 h. Protein detection was performed using the ECL system (P2300, NCM Biotech, China) and visualized with the Tanon imaging system (5200, Tanon, China).
Sperm DNA integrity analysis
Sperm DNA damage was assessed using a comet assay kit (R&D, USA) according to the manufacturer’s protocol. Fresh sperm at a concentration of 2-3 million cells per milliliter were combined with molten agarose and pipetted onto CometSlides. Following the appearance of a clear ring, the slides were immersed in lysis solution at 4 °C for 1 hour and subsequently incubated with 500 μg/mL proteinase K for 18 h. The slides were then positioned equidistant from the electrodes in a large electrophoresis chamber and subjected to a voltage of 1 volt per centimeter (measured electrode-to-electrode). After treatment with DNA precipitation solution and 70% ethanol, the samples were dried at 37 °C for 15 min. 100 μL diluted SYBR Gold was applied to each circle of dried agarose and stained for 30 min in the dark. Comet signals were visualized using a Scope AI microscope (Carl Zeiss, USA).
Intracytoplasmic sperm injection
Four-week-old female mice were injected intraperitoneally with 5 international units (IU) of pregnant mare serum gonadotropin (PMSG). Forty-eight h later, the same females received a subsequent intraperitoneal injection of 5 IU human chorionic gonadotropin (hCG). Cumulus-oocyte complexes (COCs) were collected from the ampulla of the oviduct 13 to 15 h post-hCG injection. For ICSI, sperm heads obtained from the caudal epididymis were injected into the mouse oocytes. The injected oocytes were cultured in a CBZ medium at 37 °C for 24 h. The development of two-cell embryos and blastocysts was assessed at 24 and 96 h, respectively. Blastocysts were subsequently transferred to recipient mice.
Sperm-egg binding and in vitro fertilization
The superovulation regimen employed for female mice was identical to that used for ICSI. COCs were extracted from the oviductal ampulla and treated with 330 µg/mL hyaluronidase to remove cumulus cells, followed by 1 mg/mL collagenase to remove the zona pellucida. For sperm-egg binding assays, cumulus cell-free COCs were incubated with capacitated sperm for 30 min. Sperm-egg complexes were fixed on ice in HTF medium drops containing 0.25% glutaraldehyde for 15 min and stained with Hoechst to visualize bound sperm. Images were captured using a Zeiss Axio Vert A1 microscope (Carl Zeiss, USA). For IVF, both ZP-intact and ZP-free eggs were transferred into an HTF medium containing sperm. After a 4-6 h incubation period, the eggs were washed with HTF drops and cultured in KSOM medium (Aibei Biotechnology, China) at 37 °C with 5% CO2. The two-cell embryo rate was assessed 24 h post-insemination.
Acrosome reaction analysis
To induce capacitation, sperm were collected from the cauda epididymis of adult male mice and incubated in HTF medium (Aibei Biotechnology, China) for 90 min. To induce the acrosome reaction, capacitated sperm were treated with the calcium ionophore A23187 at a final concentration of 10 μM. Following a 30-minute incubation, sperm slides were prepared, and PNA (SIGMA, L7381, USA) was employed to assess acrosomal status.
Sperm migration in the female reproductive tract
Sperm migration from the uterus to the oviduct was quantitatively evaluated through both oviductal sperm counting and histological analysis of the uterine-tubal junction (UTJ). Eight-week-old female mice were superovulated and subsequently paired with age-matched male mice of three genotypes: wild-type (WT), Tex38−/−, and Arrdc5−/−. Successful mating was confirmed by examining vaginal plug formation. Uterus and oviducts (excluding UTJ) were excised 3 h post-coitus, flushed with HTF medium, and centrifuged for sperm counting. For histology, oviduct-UTJ complexes were fixed in 4% PFA solution. Tissue sections were stained with hematoxylin and eosin (H&E) for microscopic examination.
Epididymal duct ligation
Adult Tex38+/+ and Tex38−/− mice underwent epididymal duct ligation following intraperitoneal administration of tribromoethanol. The morphology of the testis, epididymis, and fat pad was examined for abnormalities. One side of the epididymis was ligated with a surgical suture at corpus region. Four days post-treatment, sperm samples were collected from the caput or cauda epididymis for H&E staining analysis.
Co-immunoprecipitation
Full-length cDNA was acquired from Unibio. The open reading frame was amplified via RT-PCR and cloned into pCMV-Myc and pCMV-HA vectors. Lipofectamine 2000 (Invitrogen, USA) facilitated plasmid transfection into HEK-293T cells. Protein-protein interactions were assessed using the Pierce Crosslink IP kit (Thermo Scientific, USA). Briefly, IP Lysis/Wash Buffer was employed for protein extraction. A 500 μL aliquot of protein lysate (1 mg protein) was incubated with antibody-crosslinked resin overnight at 4 °C. Following extensive washing, 60 μL elution buffer was added to release the pull-down protein complexes from the resin. The eluted proteins were subsequently subjected to Western blot analysis.
Immunoprecipitation-mass spectrometry
For IP-MS, adult WT testes were lysed using IP Lysis Buffer. Sixty microliters of antibody were incubated with 10 mg of protein at 4 °C overnight. Protein A beads were added to the mixture and incubated for 4 h at 4 °C. The beads were washed four times with phosphate-buffered saline (PBS), and the supernatant was discarded. Mass spectrometry analysis was performed by Genechem Company (China). To identify proteins, the acquired raw data were loaded into Proteome Discoverer 2.2 (Thermo Fisher Scientific). A protein was considered positively identified when the peptide score of a particular peptide surpassed the significance criterion of false discovery rate (FDR) = 0.01.
Label-free proteomics analysis
Proteins from the testis, sperm collected from the cauda epididymis, and sperm-free epididymis were extracted for proteomic analysis (Genechem, China). Briefly, a nanoElute (Bruker, Bremen, Germany) system was employed to analyze samples, coupled to a timsTOF Pro (Bruker, Bremen, Germany) mass spectrometer equipped with a CaptiveSpray source. Mass spectrometry (MS) data were analyzed using MaxQuant software version 1.6.17.0. Protein abundance was quantified based on normalized spectral protein intensity (LFQ intensity). Proteins exhibiting a fold change ≥ 1.5 and a P-value < 0.05 (Student’s t-test) were considered to be differentially expressed proteins. Heatmap was plotted by https://www.bioinformatics.com.cn (last accessed on 10 Dec 2024), an online platform for data analysis and visualization.
Statistical analysis and reproducibility
SPSS 27.0 was utilized for the analysis of all statistical data. Each experiment was repeated at least three times, and the data are presented as mean ± SEM. When the data followed a normal distribution, Student’s t-test was used to compare the two groups; when it did not, the Mann-Whitney U test was utilized. p < 0.05 was considered statistically significant.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data are available from the corresponding author upon reasonable request. The source data for all statistical graphs can be found in Supplementary Data 3. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium (https://proteomecentral.proteomexchange.org) via the iProX partner repository with the dataset identifier PXD066187.
References
Agarwal, A., Mulgund, A., Hamada, A. & Chyatte, M. R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 13, 37 (2015).
Agarwal, A. et al. Male infertility. Lancet 397, 319–333 (2021).
Bhattacharya, I., Sharma, S. S. & Majumdar, S. S. Etiology of male infertility: an update. Reprod. Sci. 31, 942–965 (2024).
Levine, H. et al. Temporal trends in sperm count: a systematic review and meta-regression analysis of samples collected globally in the 20th and 21st centuries. Hum. Reprod. Update 29, 157–176 (2023).
O’Donnell, L. Mechanisms of spermiogenesis and spermiation and how they are disturbed. Spermatogenesis 4, e979623 (2014).
Zhou, L. et al. Structures of sperm flagellar doublet microtubules expand the genetic spectrum of male infertility. Cell 186, 2897–2910.e2819 (2023).
Krawetz, S. A. Paternal contribution: new insights and future challenges. Nat. Rev. Genet. 6, 633–642 (2005).
Wang, P. J., McCarrey, J. R., Yang, F. & Page, D. C. An abundance of X-linked genes expressed in spermatogonia. Nat. Genet. 27, 422–426 (2001).
Bellil, H., Ghieh, F., Hermel, E., Mandon-Pepin, B. & Vialard, F. Human testis-expressed (TEX) genes: a review focused on spermatogenesis and male fertility. Basic Clin. Androl. 31, 9 (2021).
Chen, Y. et al. Deciphering the molecular characteristics of human idiopathic nonobstructive azoospermia from the perspective of germ cells. Adv. Sci.10, e2206852 (2023).
Alagundagi, D. B. et al. Integrated molecular-network analysis reveals infertility-associated key genes and transcription factors in the non-obstructive azoospermia. Eur. J. Obstet. Gynecol. Reprod. Biol. 288, 183–190 (2023).
Yamaguchi, R. et al. Disruption of ADAM3 impairs the migration of sperm into oviduct in mouse. Biol. Reprod. 81, 142–146 (2009).
Ikawa, M. et al. Calsperin is a testis-specific chaperone required for sperm fertility. J. Biol. Chem. 286, 5639–5646 (2011).
Tokuhiro, K., Ikawa, M., Benham, A. M. & Okabe, M. Protein disulfide isomerase homolog PDILT is required for quality control of sperm membrane protein ADAM3 and male fertility [corrected]. Proc. Natl. Acad. Sci. USA 109, 3850–3855 (2012).
Ikawa, M. et al. The putative chaperone calmegin is required for sperm fertility. Nature 387, 607–611 (1997).
Muro, Y. & Okabe, M. Mechanisms of fertilization-a view from the study of gene-manipulated mice. J. Androl. 32, 218–225 (2011).
Kaneda, Y. et al. TEX38 localizes ZDHHC19 to the plasma membrane and regulates sperm head morphogenesis in mice. Proc. Natl. Acad. Sci. USA 122, e2417943122 (2025).
Giassetti, M. I. et al. ARRDC5 expression is conserved in mammalian testes and required for normal sperm morphogenesis. Nat. Commun. 14, 2111 (2023).
Liu, R. et al. ARRDC5 deficiency impairs spermatogenesis by affecting SUN5 and NDC1. Development 150, dev201959 (2023).
Russell, L. D. Histological and Histopathological Evaluation of the Testis 1st edn (Cache River Press, 1990).
Chapman, D. L. & Wolgemuth, D. J. Expression of proliferating cell nuclear antigen in the mouse germ line and surrounding somatic cells suggests both proliferation-dependent and -independent modes of function. Int. J. Dev. Biol. 38, 491–497 (1994).
Hamer, G. et al. DNA double-strand breaks and gamma-H2AX signaling in the testis. Biol. Reprod. 68, 628–634 (2003).
Hemendinger, R. A., Gores, P., Blacksten, L., Harley, V. & Halberstadt, C. Identification of a specific Sertoli cell marker, Sox9, for use in transplantation. Cell Transpl. 11, 499–505 (2002).
Raz, E. The function and regulation of vasa-like genes in germ-cell development. Genome Biol. 1, REVIEWS1017 (2000).
Elbashir, S., Magdi, Y., Rashed, A., Henkel, R. & Agarwal, A. Epididymal contribution to male infertility: an overlooked problem. Andrologia 53, e13721 (2021).
Xu, K., Yang, L., Zhang, L. & Qi, H. Lack of AKAP3 disrupts integrity of the subcellular structure and proteome of mouse sperm and causes male sterility. Development 147, dev181057 (2020).
Zhang, B. et al. A DNAH17 missense variant causes flagella destabilization and asthenozoospermia. J. Exp. Med. 217, e20182365 (2020).
Shang, Y. et al. Essential role for SUN5 in anchoring sperm head to the tail. Elife 6, e28199 (2017).
Zhang, Y. et al. The missing linker between SUN5 and PMFBP1 in sperm head-tail coupling apparatus. Nat. Commun. 12, 4926 (2021).
Wu, B., Gao, H., Liu, C. & Li, W. The coupling apparatus of the sperm head and tail†. Biol. Reprod. 102, 988–998 (2020).
Avasthi, P. et al. Germline deletion of Cetn1 causes infertility in male mice. J. Cell Sci. 126, 3204–3213 (2013).
Liu, M. et al. A novel homozygous mutation in DNAJB13-a gene associated with the sperm axoneme-leads to teratozoospermia. J. Assist. Reprod. Genet. 39, 757–764 (2022).
McDermott, J. P., Numata, S. & Blanco, G. Na,K-ATPase Atp1a4 isoform is important for maintaining sperm flagellar shape. J. Assist. Reprod. Genet. 38, 1493–1505 (2021).
Zhou, J., Yang, F., Leu, N. A. & Wang, P. J. MNS1 is essential for spermiogenesis and motile ciliary functions in mice. PLoS Genet. 8, e1002516 (2012).
Shimada, K., Kato, H., Miyata, H. & Ikawa, M. Glycerol kinase 2 is essential for proper arrangement of crescent-like mitochondria to form the mitochondrial sheath during mouse spermatogenesis. J. Reprod. Dev. 65, 155–162 (2019).
Mi, Y., Shi, Z. & Li, J. Spata19 is critical for sperm mitochondrial function and male fertility. Mol. Reprod. Dev. 82, 907–913 (2015).
Roy, A., Lin, Y. N., Agno, J. E., DeMayo, F. J. & Matzuk, M. M. Absence of tektin 4 causes asthenozoospermia and subfertility in male mice. FASEB J. 21, 1013–1025 (2007).
Zhang, Y. et al. Intraflagellar transporter protein 140 (IFT140), a component of IFT-A complex, is essential for male fertility and spermiogenesis in mice. Cytoskeleton75, 70–84 (2018).
Lei, Y. et al. Autophagic elimination of ribosomes during spermiogenesis provides energy for flagellar motility. Dev. Cell 56, 2313–2328.e2317 (2021).
Wang, Y. Y. et al. Deficiency of the Tbc1d21 gene causes male infertility with morphological abnormalities of the sperm mitochondria and flagellum in mice. PLoS Genet. 16, e1009020 (2020).
Dacheux, J. L. et al. The contribution of proteomics to understanding epididymal maturation of mammalian spermatozoa. Syst. Biol. Reprod. Med. 58, 197–210 (2012).
Zhang, K. et al. Decreased AKAP4/PKA signaling pathway in high DFI sperm affects sperm capacitation. Asian J. Androl. 26, 25–33 (2024).
Xu, H. et al. Arylsulfatase A deficiency causes seminolipid accumulation and a lysosomal storage disorder in Sertoli cells. J. Lipid Res. 52, 2187–2197 (2011).
Caballero, J. N., Frenette, G., Belleannee, C. & Sullivan, R. CD9-positive microvesicles mediate the transfer of molecules to Bovine Spermatozoa during epididymal maturation. PLoS ONE 8, e65364 (2013).
Ferrer, P., Upadhyay, S., Ikawa, M. & Clement, T. M. Testis-specific actin-like 7A (ACTL7A) is an indispensable protein for subacrosomal-associated F-actin formation, acrosomal anchoring, and male fertility. Mol. Hum. Reprod. 29, gaad005 (2023).
Skerget, S., Rosenow, M. A., Petritis, K. & Karr, T. L. Sperm proteome maturation in the mouse epididymis. PLoS ONE 10, e0140650 (2015).
Barrachina, F. et al. Sperm acquire epididymis-derived proteins through epididymosomes. Hum. Reprod. 37, 651–668 (2022).
Kiyozumi, D. & Ikawa, M. Proteolysis in reproduction: lessons from gene-modified organism studies. Front. Endocrinol.13, 876370 (2022).
Nishimura, H., Myles, D. G. & Primakoff, P. Identification of an ADAM2-ADAM3 complex on the surface of mouse testicular germ cells and cauda epididymal sperm. J. Biol. Chem. 282, 17900–17907 (2007).
Xiong, W. et al. Dissecting the PRSS37 interactome and potential mechanisms leading to ADAM3 loss in PRSS37-null sperm. J. Cell Sci. 134, jcs258426 (2021).
Larasati, T. et al. Tmprss12 is required for sperm motility and uterotubal junction migration in mice†. Biol. Reprod. 103, 254–263 (2020).
Acknowledgements
This work was supported by National Natural Science Foundation of China (82371614 and 82071696 to Ying Zheng, 82101674 to Changmin Niu, 82301799 to Fan Yang), Major Project of the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (20KJA310002 to Ying Zheng), and Practice Innovation Program of Jiangsu Province (KYCX233618 to Tingting Ge).
Author information
Authors and Affiliations
Contributions
Project administration: Ying Zheng; Conceived and designed the experiments: Ying Zheng, Changmin Niu, and Fan Yang; Performed the experiments: Lu Yuan, Tingting Ge, Ling Yang, Changmin Niu, Wenhua Xu, Guanghua Li, Linwei Xu, Wenting Lu, Shiqi Meng, and Jieyu Zhao; Analyzed the data: Lu Yuan, Tingting Ge, Ying Zheng, Fan Yang, Changmin Niu, Wenhua Xu, Yichun Zhao, and Xu Cheng; Wrote the paper: Lu Yuan, Tingting Ge, Fan Yang, and Changmin Niu; All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Mamiko Maekawa and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editors: Rupinder Kaur and Kaliya Georgieva.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yuan, L., Ge, T., Yang, L. et al. Disruption of TEX38 impairs sperm morphogenesis and the migration of sperm into the oviduct. Commun Biol 8, 1191 (2025). https://doi.org/10.1038/s42003-025-08644-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s42003-025-08644-1
