
Abstract
Aging is a major risk factor for cancer, but the precise mechanism by which aging promotes carcinogenesis remains largely unknown. Here, using genetically modified mouse models, we show that p16high senescent (p16h-sn) fibroblasts accumulate with age, constitute inflammatory cancer-associated fibroblasts (CAFs) and promote tumor growth in bladder cancer models. Single-cell RNA sequencing of fibroblasts from aged mice revealed higher expression of the C–X–C motif chemokine 12 gene (Cxcl12) in p16h-sn fibroblasts than in p16low fibroblasts. Elimination of p16h-sn cells or inhibition of CXCL12 signaling notebly suppressed bladder tumor growth in vivo. We identified high expression levels of SMOC2, GUCY1A1 (GUCY1A3), CXCL12, CRISPLD2, GAS1 and LUM as a signature of p16h-sn CAFs in humans and mice, which was associated with age and poor prognosis in patients with advanced and nonadvanced bladder cancer. Here we show that p16h-sn fibroblasts in the aged bladder create a cancer-permissive niche and promote tumor growth by secreting CXCL12.
Similar content being viewed by others

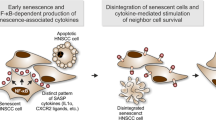
Main
Aging is a major risk factor for cancer1. The incidence of most cancers increases abruptly after the sixth decade of life1,2,3. Therefore, cancer is considered an age-related disease, although the molecular and mechanistic links between aging and cancer remain largely unknown. All cancers acquire, to a greater or lesser extent, mutations in proto-oncogenes and/or tumor suppressor genes that drive malignant transformation and cancer progression4. The critical role of the accumulation of driver gene mutations during carcinogenesis may explain why cancer incidence increases with age. As gene mutations accumulate over time, the risk of reaching the threshold sufficient for cancer to develop increases gradually with aging5. Indeed, even in physiologically normal tissues, the numbers of somatic mutations and driver-mutated clones increase with age. However, in Peto’s paradox, although the risk of developing cancer should theoretically increase with the number of cells and the lifespan of an organism, large and long-lived animals do not have a higher incidence of cancer than small and short-lived ones6. Recently, several lines of evidence have suggested that the age-related increase in cancer incidence is not simply the result of the accumulation of gene mutations but is also regulated by biological processes7.
Cancer is now known to be driven by a complex ecosystem consisting of a variety of noncancer cells, such as immune cells, fibroblasts and endothelial cells, and their myriad interactions with each other and with cancer cells8. These noncancerous cells and their environment were initially considered bystanders in carcinogenesis, but these cells and their secreted molecules were later believed to have key roles in cancer progression and pathogenesis7. In other words, biological changes in the tissue microenvironment associated with aging may promote cancer initiation, which, in turn, affects noncancerous tumor-associated stromal cells and promotes cancer progression. However, little is known about what changes occur in the tissue microenvironment during aging and what signals promote carcinogenesis. Cancer-associated fibroblasts (CAFs) are the predominant cell type of tumor-associated stromal cells in the microenvironment of solid tumors. CAFs can be derived from various cell types, such as fibroblasts, pericytes, mesenchymal stem cells and adipocytes9,10, resulting in a heterogeneous population9,11,12. However, the precise origin of CAFs and the specific CAF subset with a substantial impact on cancer initiation and progression remain elusive.
Cellular senescence is induced in previously replication-competent cells by various genotoxic and epigenetic stresses13. Senescent cells show various unique characteristics, such as permanent cessation of cell proliferation and secretion of various proinflammatory cytokines and growth factors (senescence-associated secretory phenotypes (SASPs))13. Recently, several lines of evidence have indicated that senescent or aged tumor-associated stromal cells may create a cancer growth-promoting, metastasis-promoting and chronic inflammatory environment7. However, although these stromal cells may be promising targets for cancer therapy, it remains largely unknown which types of senescent tumor-associated stromal cells have dominant effects on cancer promotion, which site these cells originate from and which signaling pathways of senescent stromal cells are involved.
Bladder cancer is a common age-related cancer, ranking fourth in estimated new cases and eighth in estimated deaths among male cancers in 2022 in the United States3. Approximately 80% of patients with bladder cancer have non-muscle-invasive bladder cancer, whereas about two-thirds of patients with muscle-invasive bladder cancer develop local or systemic disease, resulting in a relatively poor prognosis despite advancements in surgical techniques and multimodal therapy14. Moreover, bladder cancer is characterized by chemoresistance and a high recurrence rate15,16. Therefore, there is a pressing need for advanced therapies to address the unsatisfactory outcomes associated with advanced bladder cancer. In this study, using mouse models in which p16high (p16h) cells are visualized by tamoxifen administration and selectively eliminated by diphtheria toxin (DT) treatment, we found that p16h senescent (p16h-sn) fibroblasts accumulate in the bladder with age, create a cancer-permissive niche by secreting C–X–C motif chemokine 12 (CXCL12) and function as cancer-promoting inflammatory CAFs (iCAFs). Importantly, we show that a transcriptomic signature of p16h-sn fibroblasts has prognostic value for patients with bladder cancer.
Results
p16h cells in the mouse bladder increase with aging
The accumulation of p16h cells in organs is known to provide an inflammatory environment that is a hallmark of the aging process and is related to tumor progression7,17. The p16-CreERT2-tdTomato mouse model was used to investigate the age-dependent alteration of p16h cell abundance in the bladder by introducing tamoxifen at different ages18. We observed a gradual increase in Tomato-positive (Tomato+) p16h nonimmune cells in the bladder of male and female mice with age. In the bladders of 22- to 25-month-old male and female mice, the number of p16h cells was approximately four times higher than that in 3-month-old mice (Fig. 1a–d). Furthermore, through section analysis, we found that the increase in these p16h cells primarily occurred in the lamina propria and muscle layer; the urothelium showed a trend of increased p16h cells but without statistical significance (Fig. 1e,f).

a, Representative scatter plots of Tomato+ (p16h) cells in the bladders of male p16-CreERT2-tdTomato mice at the indicated ages. WT, wild-type. Boxes indicate gating of Tomato+ nonimmune cells. b, Population of Tomato+ (p16h) cells in male mouse bladders at the indicated ages, as determined by fluorescence-activated cell sorting (FACS) (n = 3 for each group). c, Representative scatter plots of Tomato+ (p16h) cells in the bladders of female p16-CreERT2-tdTomato mice at the indicated ages. Boxes indicate gating of Tomato+ nonimmune cells d, Population of Tomato+ (p16h) cells in female mouse bladders at the indicated ages, as determined by FACS (3-month-old mice, n = 3; 12-month-old mice, n = 3; 22- to 25-month-old mice, n = 5). e, Representative images of IHC staining of three layers of the bladder wall in male p16-CreERT2-tdTomato mice at each age. The Tomato protein was stained using an anti-RFP antibody. The areas outlined by dashed lines are the urothelium or lamina propria regions. Scale bar, 100 μm. f, Number of Tomato+ (p16h) cells in the indicated layers of the bladder wall of male mice at each age (n = 3 for each group). g, MFI of FITC signals with or without SPiDER β-gal staining among Tomato+ (p16h) or Tomato− (p16l) cells from young (7- to 12-week-old) male p16-CreERT2-tdTomato mice. The dotted lines indicate each MFI. There is an overlap of the fluorescence spectrum between PE (Tomato) and FITC. Thus, SPiDER β-gal signals were calculated by measuring the ΔMFI of FITC signals with or without SPiDER β-gal. h, ΔMFI of FITC signals among Tomato+ (p16h) or Tomato− (p16l) cells (n = 7). One-way ANOVA with Tukey’s test was performed for b and d and with the Games–Howell test for f. A paired t test was performed for h. Data in b, d, f and h are presented as mean ± s.e.m. All t tests were two-sided.
To confirm whether Tomato+ nonimmune bladder cells had upregulated expression of p16 and other senescence-related genes, we analyzed the mRNA levels of p16, p21, p15 and Glb1 in sorted Tomato+ nonimmune bladder cells and found that these genes were significantly upregulated in the Tomato+ cells compared to Tomato− cells (Extended Data Fig. 1a,b). High activity of senescence-associated β-galactosidase (β-gal)19 was also observed in Tomato+ nonimmune bladder cells (Fig. 1g,h). We further verified by immunostaining that the Tomato+ cells were mainly composed of p16+ and p21+ cells (Extended Data Fig. 1c,d). In addition, Tomato+ cells showed reduced expression of proliferative markers, including marker of proliferation Ki-67 (MKI67) and lamin B1 (LMNB1) (Extended Data Fig. 1e,f). All these markers showed the association between Tomato+ cells and senescence characteristics in nonimmune bladder cells, further supporting the view that the Tomato+ cell population increases with age, enriching the senescent cells. Therefore, we here refer to p16h cells in the bladder as p16h-sn cells.
Upregulation of Cxcl12 in p16h-sn fibroblasts in the aged bladder
To identify the cell types and transcriptomic features of p16h-sn cells in the bladder, we performed single-cell RNA sequencing (scRNA-seq) analysis on a nonimmune population in the bladder of a 25-month-old male mouse. The scRNA libraries of Tomato+/CD45− and Tomato−/CD45− bladder cells were prepared in parallel following FACSorting (Extended Data Fig. 2a,b). We obtained 8,588 Tomato− cells and 1,527 Tomato+ cells and identified eight cell types according to previously described marker genes20 (Fig. 2a). Apart from myofibroblasts, Tomato+ cells were widely distributed among various cell types, with fibroblasts being the most abundant (Fig. 2b–d). This was further confirmed by double staining of Tomato signals and cell type-specific markers (Extended Data Fig. 2c,d).

a, Uniform Manifold Approximation and Projection (UMAP) visualization of total Tomato− (p16l) and Tomato+ (p16h-sn) bladder cells from an aged (25-month-old) male p16-CreERT2-tdTomato mouse, colored by clusters. b, Distributions of Tomato− and Tomato+ cells in the UMAP. c, Numbers of Tomato− (p16l) and Tomato+ (p16h-sn) cells among the indicated cell types. d, Cell type composition of Tomato+ (p16h-sn) cells. e, Volcano plot showing DEGs between Tomato+ (p16h-sn) and Tomato− (p16l) fibroblasts from the bladder. The red and blue dots indicate upregulated and downregulated DEGs, respectively, identified by an adjusted P value of <0.05 and a log2 fold change (log2FC) value of >0.15. This analysis was two-sided, and Benjamini–Hochberg adjustment was performed. f, Representative immunofluorescence image of bladder fibroblasts from young (7- to 12-week-old) male p16-CreERT2-tdTomato mice, using the indicated staining. White arrowheads indicate double-positive cells for both Tomato and CXCL12. Scale bar, 200 μm. g, Percentages of CXCL12+ cells in Tomato− (p16l) and Tomato+ (p16h-sn) fibroblasts (n = 3 for each group). h, Top 15 enriched GO terms derived from upregulated DEGs of Tomato+ (p16h-sn) fibroblasts. Red boxes indicate GO terms related to cancer progression. GAG, glycosaminoglycan. All terms were identified by an adjusted P value of <0.05. A paired t test was performed for g. Data in g are presented as mean ± s.e.m. All t tests were two-sided.
An analysis of differentially expressed genes (DEGs) between Tomato+ and Tomato− fibroblasts revealed several upregulated genes, including Cxcl12 (Fig. 2e). The expression of CXCL12 in bladder fibroblasts was confirmed at both the protein and mRNA levels (Fig. 2f,g and Extended Data Fig. 2e). CXCL12 is a chemokine previously described as one of the SASP factors and is secreted from senescent tumor cells21. Gene Ontology (GO) analysis of upregulated DEGs in Tomato+ fibroblasts revealed several biological processes related to cancer progression, including cell migration, protein kinase B signaling, angiogenesis and cell–cell adhesion21,22,23 (Fig. 2h and Supplementary Fig. 1a). Conversely, SASP factors and their associated functions were not evident in other cell types, such as endothelial cells, smooth muscle cells and urothelial cells (Supplementary Fig. 1b,c).
Ablation of p16h-sn cells suppresses bladder cancer growth
Given the close association between bladder cancer progression and aging3, we intended to understand whether these p16h-sn cells influence the incidence and progression of bladder cancer. In p16-CreERT2-tdTomato mice, Tomato+ noncancer cells were observed in tumors following orthotopic allograft transplantation using the MB49 mouse bladder cancer cell line (Fig. 3a and Extended Data Fig. 3a). Female mice were used for the cancer transplantation experiments because of the technical limitations of urethral transplantation in male mice. p16h-sn noncancer cells were efficiently and selectively eliminated by apoptosis24 through tamoxifen and DT administration in the allograft model using p16-CreERT2-DTR mice (Extended Data Fig. 3b,c). To prevent potential influences of tamoxifen and DT administration on bladder tumor growth, we used p16-CreERT2 mice lacking the DTR cassette gene as controls. We found that the tumor sizes and bladder weights in the p16-CreERT2-DTR group transplanted with MB49 cells were significantly lower than those in the littermate control group (Fig. 3b,c). This reduction in bladder cancer weight was not due to DT-induced elimination of p16h-sn cells per se or changes in bladder wall thickness (Extended Data Fig. 3d,e). In fact, the calculated tumor weights (transplanted bladder weights in Fig. 3c minus the average of noncancer bladder weights in Extended Data Fig. 3d) were significantly lower in the p16-CreERT2-DTR group (Extended Data Fig. 3f). Consistent with these, the p16-CreERT2-DTR group also showed a significantly improved survival rate (Fig. 3d).

a, Representative IHC images of bladder tissue from young (7- to 12-week-old) female p16-CreERT2-tdTomato mice transplanted with MB49 cells. The Tomato protein was stained using an anti-RFP antibody. In the low-magnification image, the area outlined by the dashed line indicates cancer. Low-magnification image scale bar, 200 µm; high-magnification image scale bar, 100 µm. b, Representative images of bladders from young (7- to 12-week-old) female p16-CreERT2 and p16-CreERT2-DTR mice transplanted with MB49 cells. Scale bar, 5 mm. c, Bladder weights after cancer transplantation in young (7- to 12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice (p16-CreERT2, n = 10; p16-CreERT2-DTR, n = 11). d, Survival probabilities of young (7- to 12-week-old) female p16-CreERT2 and p16-CreERT2-DTR mice (p16-CreERT2, n = 6; p16-CreERT2-DTR, n = 7). e, Images of bladders from young (7- to 12-week-old) female wild-type mice transplanted with MB49 cells and treated with ABT-263 or vehicle (vehicle, n = 12; ABT-263, n = 11). Scale bar, 1 cm. f, Bladder weights of the samples described in e (vehicle, n = 12; ABT-263, n = 11). g, Representative immunofluorescence images of bladder tissues with labeling of Tomato+ (p16h-sn) cells before (prelabeling group) or after (postlabeling group) the transplantation of MB49 cells in young (7- to 12-week-old) or middle-aged (12- to 16-month-old) female p16-CreERT2-tdTomato mice. The Tomato protein was stained with an anti-mCherry antibody. Scale bar, 100 μm. h, Number of Tomato+ (p16h-sn) cells per field in the transplanted bladder tumor in young (7- to 12-week-old) or middle-aged (12- to 16-month-old) p16-CreERT2-tdTomato mice in the pre- or postlabeling group (young, n = 3 (pre) and n = 5 (post); middle-aged, n = 4 (pre) and n = 4 (post)). i, Representative images of bladders from young (7- to 12-week-old) female p16-CreERT2 and p16-CreERT2-DTR mice with senolytic treatment followed by MB49 cell transplantation. Scale bar, 5 mm. j, Bladder weights of young (7- to 12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice with senolytic treatment followed by MB49 cell transplantation (p16-CreERT2, n = 11; p16-CreERT2-DTR, n = 11). An unpaired t test was performed for c, f, h and j. Data in h are presented as mean ± s.e.m. For all box plots, the horizontal line in the box indicates the median value. The lower and upper bounds correspond to the 25th and 75th percentiles, respectively. The cross mark indicates the mean value. Whiskers extend to the maximum and minimum values, with points outside this range drawn individually. All t tests were two-sided.
To confirm the effect of p16h-sn cells on bladder tumor growth, we treated wild-type mice bearing transplanted MB49 tumors with ABT-263, a representative senolytic drug25 (Extended Data Fig. 3g). The removal of p16h-sn cells within tumors by ABT-263 treatment led to a significant reduction in tumor weight and size compared to the vehicle group (Fig. 3e,f and Extended Data Fig. 3h,i). Double treatment with tamoxifen + DT and ABT-263 in p16-CreERT2-DTR mice did not result in an additive effect of reduced bladder weights compared to tamoxifen + DT treatment alone, suggesting that the impact of ABT-263 on tumor suppression is mainly due to the elimination of senescent cells (Extended Data Fig. 3j,k).
Interestingly, we found that the Tomato+ cells within the tumors preexisted in the bladder before tumor formation rather than being induced by the cellular microenvironment within the transplanted tumor in both young (7- to 12-week-old) and middle-aged (12- to 16-month-old) mice (Fig. 3g,h and Extended Data Fig. 4a,b). Consistent with this, we observed a greater number of Tomato+ cells in tumors (Fig. 3h), faster tumor growth and poorer survival rate in middle-aged mice than in young mice (Extended Data Fig. 4c–e). Based on these findings, we speculated that senolysis before bladder cancer transplantation might also be effective in preventing cancer progression. As expected, even in the scheme of senolysis followed by MB49 transplantation (Extended Data Fig. 4f), pretreatment with DT significantly reduced the bladder weights in the p16-CreERT2-DTR group compared to the littermate control group (Fig. 3i,j). This concept was further supported by the observation that pretreatment with ABT-263 also significantly suppressed bladder tumor growth compared to vehicle treatment (Extended Data Fig. 4g–i). Taken together, these findings indicate that p16h-sn cells preexisted in normal bladder tissue and migrated into cancerous tissue during bladder cancer progression, and prophylactic senolysis may offer protection against bladder cancer progression.
Ablation of p16h-sn cells suppresses bladder tumorigenesis
In addition to determining the influence of the elimination of p16h-sn bladder cells on cancer progression, we also investigated its impact on tumor initiation using an N-butyl-N-(4-hydroxybutyl)nitrosamine (BBN) carcinogenesis model26 (Extended Data Fig. 5a). Histopathological analysis revealed the presence of bladder cancer and Tomato+ cells within tumors in mice after 84 days of exposure to BBN water (Extended Data Fig. 5b,c). Again, Tomato+ cells were preexisting in the bladder and migrated into the tumors (Extended Data Fig. 5d,e). After the elimination of p16h-sn cells in the presence of DT, the bladder weights were significantly lower in the p16-CreERT2-DTR group than in controls (p16-CreERT2 or wild-type) (Extended Data Fig. 5f–h). The pathological analysis showed that, while there was a decrease in the total tumor area, the number of exophytic tumors in the lumen was not affected by p16h-sn cell depletion (Extended Data Fig. 5i,j). This suggests that p16h-sn cells are more likely to be involved in facilitating tumor growth and progression rather than cancer initiation.
CXCL12 from p16h-sn fibroblasts supports bladder cancer growth
The reduction in tumor size caused by either the p16-CreERT2-DTR model or the ABT-263-induced elimination of p16h-sn cells was not likely due to the induction of apoptosis or necrosis in the tumors (Supplementary Fig. 2a–c). This suggests that there might be some signal from p16h-sn cells supporting tumor growth. CXCL12 is a chemokine that promotes tumor growth, cancer cell migration, proliferation, cell adhesion, angiogenesis, epithelial–mesenchymal transition and metastasis in various cancers21,22,23,27. Among the top 10 genes upregulated in p16h-sn fibroblasts (Fig. 2e), Cxcl12 was the most enriched in 15 GO terms (Supplementary Fig. 1a). Furthermore, in human data from The Cancer Genome Atlas (TCGA), of the top 7 genes upregulated in p16h-sn fibroblasts (Extended Data Fig. 9b), CXCL12 was the most relevant to age (Extended Data Fig. 9d) and survival probability (Extended Data Fig. 10b). Therefore, we hypothesized that CXCL12 would be the most reliable candidate to reflect the characteristics of senescent cells in the age-dependent bladder tumor microenvironment. The protein level of CXCL12 in the tumor in the presence of DT was reduced in the p16-CreERT2-DTR group compared to the control (p16-CreERT2) group (Fig. 4a,b and Extended Data Fig. 6a). A decrease in the number of MKI67+ cells was observed in both the p16-CreERT2-DTR and ABT-263 treatment groups, and CD31+ and CD105+ areas were reduced in p16-CreERT2-DTR mice compared to the corresponding controls (Fig. 4c,d and Extended Data Fig. 6b,c). These data suggest that senolysis suppressed cancer cell proliferation and angiogenesis in cancer tissue and led to tumor regression, possibly by mitigating the action of CXCL12. Consistent with this, the level of phosphorylated AKT (pAKT), one of the downstream signaling pathways of CXCL12 (ref. 28), was also significantly reduced in the p16-CreERT2-DTR group after DT administration (Fig. 4e,f). To confirm the role of CXCL12 in bladder cancer progression, we knocked down two receptors of CXCL12 (CXCR4 and CXCR7) in MB49 cells (Extended Data Fig. 6d,e). We then transplanted the cells into the mouse bladder and treated the mice with a CXCL12 inhibitor (AMD3100)29 (Extended Data Fig. 6f). Knockdown of Cxcr4 and Cxcr7 in MB49 cells or treatment with AMD3100 resulted in the suppression of bladder tumor progression (Fig. 4g,h). The pAKT levels in the transplanted MB49 cells were lower in the AMD3100 treatment group than in the vehicle treatment group (Extended Data Fig. 6g). Finally, the double treatment with tamoxifen + DT and AMD3100 in p16-CreERT2 and p16-CreERT2-DTR mice did not show an additive effect on tumor suppression (Extended Data Fig. 6h,i). These results suggest that CXCL12 from p16h-sn fibroblasts is a prominent factor in bladder cancer progression by supporting cancer cell proliferation and angiogenesis in cancer tissues. However, our results do not exclude the possibility that other types of p16h-sn cells may influence bladder cancer progression.

a, Representative IHC images of bladder tissues from young female (7- to 12-week-old) p16-CreERT2 (left) or p16-CreERT2-DTR (right) mice transplanted with MB49 cells (anti-CXCL12 antibody staining). Low-magnification image scale bar, 1 mm; high-magnification image scale bar, 200 μm. b, Percentage of CXCL12 expression values quantified in the samples described in a (p16-CreERT2, n = 11; p16-CreERT2-DTR, n = 14). c, Left: Representative IHC images of bladder tissues from young (7- to 12-week-old) female p16-CreERT2 (top) or p16-CreERT2-DTR (bottom) mice transplanted with MB49 cells (anti-MKI67 antibody staining). Scale bar, 100 μm. Right: Percentage of MKI67+ cells (p16-CreERT2, n = 4; p16-CreERT2-DTR, n = 3). d, Left: Representative IHC images of bladder tissues from young (7- to 12-week-old) female p16-CreERT2 (top) or p16-CreERT2-DTR (bottom) mice transplanted with MB49 cells (anti-CD31 antibody staining). Scale bar, 100 μm. Right: Percentage of the CD31+ area (p16-CreERT2, n = 5; p16-CreERT2-DTR, n = 3). e, Immunoblots using the indicated antibodies on sorted cancer cells from the indicated mouse groups. f, Quantified intensity ratio of pAKT/AKT in e (p16-CreERT2, n = 3; p16-CreERT2-DTR, n = 3). g, Images of bladders from young (7- to 12-week-old) female wild-type mice transplanted with the shControl or shCXCR4-1/CXCR7-1 MB49 cell line and treated with AMD3100 or vehicle (shControl, n = 10 (vehicle) and n = 9 (AMD3100); shCXCR4-1/CXCR7-1, n = 10 (vehicle) and n = 9 (AMD3100)). Scale bar, 1 cm. h, Weights of the bladder samples shown in g. A chi-squared test was performed for b. An unpaired t test was performed for c, d and f. One-way ANOVA with the Games–Howell test was performed for h. Data in c, d and f are presented as mean ± s.e.m. For all box plots, the horizontal line in the box indicates the median value. The lower and upper bounds correspond to the 25th and 75th percentiles, respectively. The cross mark indicates the mean value. Whiskers extend to the maximum and minimum values, with points outside this range drawn individually. All t tests were two-sided.
CXCL12 is predominantly secreted by p16h-sn iCAFs
We found that p16h-sn cells in bladder cancer tumors mainly migrated from the surrounding bladder tissue. Among them, fibroblasts were an abundant cell type (Fig. 2d), suggesting the possibility that p16h-sn fibroblasts might exhibit characteristics of CAFs. CAFs are a major component of the stroma and are classified into iCAFs and myofibroblastic CAFs (myCAFs) based on the expression of CD140a and α-smooth muscle actin (αSMA), respectively12,30. iCAFs are known to secrete various cytokines and chemokines, including CXCL12, in bladder cancer31,32. In CD140a+ iCAFs in bladder cancer, we found that approximately 3% of the cells were Tomato+ (Fig. 5a,b). It has been reported that immune dysfunction caused by ‘immunosenescence’ has a role in promoting tumor progression33. Surprisingly, p16h-sn immune cells were a smaller population than p16h-sn iCAFs in bladder tumors (Supplementary Fig. 3a). In addition to cellular senescence intrinsic to immune cells, we examined the potential impact of eliminating senescent cells on immune dysregulation. The populations of macrophages and T cells were not altered by DT administration in p16-CreERT2-DTR mice or by treatment with ABT-263 (Supplementary Fig. 3b–e). These findings show that, at least in the bladder cancer allograft model, there was a low abundance of senescent immune cells, and the elimination of p16h-sn cells did not substantially affect the number of immune cells within the tumor. These results suggest that the role of immune senescence and the immune system in suppressing bladder cancer during p16h-sn cell depletion may be minimal.

a, Gating strategy for sorting Tomato+ (p16h-sn) iCAFs from young (7- to 12-week-old) female p16-CreERT2-tdTomato mice. Boxes indicate gating strategy. b, Percentage of Tomato+ (p16h-sn) cells in iCAFs from young (7- to 12-week-old) female p16-CreERT2-tdTomato mice (n = 4). c, Representative immunofluorescence images of iCAFs with the indicated staining (left) and representative high-magnification images (right). White arrowheads indicate double-positive cells for both Tomato and CXCL12. Scale bars, 200 μm, 10 μm. d, Percentage of CXCL12+ cells in Tomato− (p16l) and Tomato+ (p16h-sn) iCAFs from young (7- to 12-week-old) female p16-CreERT2-tdTomato mice (n = 3). e, qPCR using the primer of Cxcl12 on the sorted iCAFs from young (7- to 12-week-old) female p16-CreERT2-tdTomato mice (n = 4 for each group). f, Relative proliferation of parental MB49 cells (n = 6 for each group). g, Relative proliferation of the indicated MB49 cell lines (n = 6 for each group). A paired t test was performed for d, e and f. One-way ANOVA with Tukey’s test was performed for g. Data in b and d–g are presented as mean ± s.e.m. All t tests were two-sided.
The scRNA-seq data of aged bladders revealed that Cxcl12 was predominantly upregulated in p16h-sn fibroblasts (Extended Data Fig. 7a,b). Indeed, p16h-sn iCAFs showed higher expression of p16, p21 and Cxcl12 than p16low (p16l) iCAFs (Fig. 5c–e and Extended Data Fig. 7c). To verify the tumor-promoting function of CXCL12 from p16h-sn iCAFs, we treated parental MB49, shControl and four knockdown (shCXCR4-1 or -2/shCXCR7-1 or -2) cell lines with conditioned medium from either the entire iCAF population or the p16l iCAFs for 48 h (Extended Data Fig. 7d,e). The proliferation of parental MB49 cells was enhanced by the treatment with conditioned medium from the total iCAFs compared to that from the p16l iCAFs (Fig. 5f). Importantly, this effect was rescued by Cxcr4/Cxcr7 knockdown (Fig. 5g).
To corroborate our findings with clinical data, we analyzed previously published scRNA-seq data obtained from six patients with bladder cancer34. The CAFs were classified into iCAFs (383 cells) and myCAFs (335 cells) based on the expression levels of PDGFRA, RGS5 and ACTA2 (ref. 32) (Extended Data Fig. 8a,b). Among them, we noticed that iCAFs were relatively more abundant in the aged group (n = 3; mean age, 75 years; 213 cells) than in the young group (n = 3; mean age, 54.3 years; 505 cells) (Extended Data Fig. 8c,d). The CXCL12-expressing fibroblasts were markedly enriched in iCAFs from the aged patients (Extended Data Fig. 8e,f), revealing that the levels of CXCL12 secreted by iCAFs increase with aging in human bladder cancer.
p16h-sn CAF signature is relevant to patient prognosis
In the publicly available clinical transcriptomic data of bladder cancer (TCGA), the pre-old (age 65–74 years) and old (age ≥75 years) groups had a significantly poorer prognosis than the young (age ≤64 years) group (Extended Data Fig. 9a). To evaluate the impact of p16h-sn CAFs on the prognosis of patients with bladder cancer, we first defined the transcriptomic signature of p16h-sn fibroblasts based on our scRNA-seq dataset. The top 7 genes (Smoc2, Gucy1a1 (Gucy1a3), Cxcl12, Matn2, Crispld2, Gas1 and Lum) with the highest fold-change values in p16h-sn fibroblasts compared to p16l fibroblasts were considered as candidates (Extended Data Fig. 9b). Among these, except for MATN2, the gene expression levels were highly positively correlated in TCGA dataset (Extended Data Fig. 9c). The expression of five genes, except for MATN2 and CRISPLD2, tended to increase in the older adult group (Extended Data Fig. 9d). Regarding cancer prognostic factors, although only GUCY1A1 (GUCY1A3), GAS1 and LUM were correlated with recurrence, six genes (except for MATN2) were associated with metastasis, pathological T stage and grade (Extended Data Fig. 10a). In addition, SMOC2, CXCL12, CRISPLD2 and LUM were correlated with survival probability, and GUCY1A1 (GUCY1A3) and GAS1 tended to be relevant to this parameter (P < 0.1) (Extended Data Fig. 10b). According to these results, we define high expression levels of SMOC2, GUCY1A1 (GUCY1A3), CXCL12, CRISPLD2, GAS1 and LUM as a signature of p16h-sn CAFs.
Subsequently, for further analysis of several clinical outcomes, we categorized patients in TCGA based on the expression levels of genes in the p16h-sn CAF transcriptomic signature. The p16h-sn CAF signature scores were correlated with age and poor prognostic outcomes (Fig. 6a–d). The hazard ratio for survival of the p16h-sn CAF signature was 1.21 and higher than that of pathological T stage (Fig. 6e). As most TCGA samples were obtained from patients with advanced bladder cancer with muscle invasion, we also attempted to examine samples from patients with nonadvanced bladder cancer to gain a better understanding of the role of p16h-sn CAFs in bladder cancer progression and to use the p16h-sn CAF signature comprehensively as a prognostic factor. We performed microarray RNA analysis on bladder cancer samples from 136 patients at Fukushima Medical University, most of whom had non-muscle-invasive bladder cancer. The p16h-sn CAF signature scores were associated with poor prognostic outcomes, including metastasis, pathological T stage, grade, muscle invasion and tumor size (Fig. 6f). Taken together, our findings suggest that p16h-sn CAFs have a critical role in age-dependent bladder cancer progression and that the p16h-sn CAF signature has prognostic value in both advanced and nonadvanced bladder cancer.

a, Heat map showing the expression levels of LUM, CRISPLD2, CXCL12, SMOC2, GUCY1A3 and GAS1 in TCGA bladder cancer data with clinical information (n = 436). b, Quantitative score of the p16h-sn CAF signature across different age groups of patients from TCGA dataset (age ≤64 years, n = 138; age 65–74 years, n = 133; age ≥75 years, n = 155). c, Overall survival probability of patients from TCGA. The patients were evenly divided into two groups based on the p16h-sn CAF signature score (low, n = 212; high, n = 213). d, Scores of the p16h-sn CAF signature in relation to the indicated clinical predictive factors among patients from TCGA dataset (metastasis, n = 139 (negative), n = 140 (positive); recurrence, n = 264 (negative), n = 129 (positive); pathological T stage, n = 127 (≤pT2), n = 204 (pT3), n = 61 (pT4); grade, n = 21 (low), n = 402 (high)). e, Hazard ratios and 95% confidence intervals of the p16h-sn CAF signature and the four clinical outcomes for survival. This analysis was two-sided. f, Scores of the p16h-sn CAF signature in relation to the indicated clinical predictive factors from the microarray data of patients with bladder cancer at Fukushima Medical University (metastasis, n = 114 (negative), n = 13 (positive); recurrence, n = 90 (negative), n = 31 (positive); pathological T stage, n = 64 (pTa), n = 45 (pT1), n = 21 (≥pT2); grade, n = 38 (low), n = 61 (high); muscle invasion, n = 97 (negative), n = 39 (positive); tumor size, n = 86 (≤3 cm), n = 48 (≥3 cm); number of tumors, n = 56 (single), n = 80 (multiple)). An unpaired t test was performed for d and f. One-way ANOVA with the Games–Howell test was performed for b and with Tukey’s test for d and f. Cox proportional hazards analysis was performed for e. Data in e are presented as hazard ratios with 95% confidence intervals. For all box plots, the horizontal line in the box indicates the median value. The lower and upper bounds correspond to the 25th and 75th percentiles, respectively. The cross mark indicates the mean value. Whiskers extend to the maximum and minimum values, with points outside this range drawn individually.
Discussion
Aging is one of the major risk factors for cancer, with the incidence of cancer being higher in the older than in the younger population. Epidemiological statistics and the overlap of biological processes underlying the hallmarks of aging and cancer indicate their strong correlation1,7. Several mechanisms have been proposed to explain the link between aging and cancer. For example, an accumulation of oxidative stress and DNA damage with age causes mutations in certain proto-oncogenes and tumor suppressor genes, ultimately promoting carcinogenesis35,36. The progressive decline in immune function in older individuals also has a role in this association, as effective antitumor immunity may be compromised33. However, none of these mechanisms fully explain the molecular basis of the age-related exponential increase in cancer incidence.
Our current study shows that the accumulation of p16h-sn fibroblasts creates a cancer-permissive niche in the bladder and then forms cancer-promoting iCAFs that secrete CXCL12; these are likely part of the mechanisms underlying the age-related increase in cancer incidence. Interestingly, although previous studies have suggested that senescence of stromal cells is induced by interaction with cancer cells and molecules secreted from cancer cells37,38, our results showed that p16h-sn fibroblasts were already present in the aged bladder. This is intriguing because the aged tissue microenvironment promotes bladder cancer progression.
Recently, there has been overwhelming evidence for a degree of specialization among CAFs, such as myCAFs and iCAFs, suggesting that some specific subpopulations of CAFs exert dominant effects on cancer progression34,39,40. CXCL12 was previously reported as a SASP factor in senescent tumor cells of papillary thyroid carcinoma21. CXCL12 is upregulated by hypoxic conditions, and its interaction with its receptor (CXCR4) promotes tumor growth, adhesion, vascularization and metastasis41. In the bladder cancer microenvironment, CXCL12 was reported to be secreted by stromal iCAFs31,32. In this study, we found that the signaling pathway involving CXCL12 secreted by p16h-sn iCAFs and CXCR4/7 expressed on tumor cells is important for tumor growth. However, the molecular mechanisms through which this cytokine affects bladder cancer cells require further comprehensive analysis in future studies. These findings indicate that, although p16h-sn iCAFs are a small population, targeting them may show clinical benefit. Indeed, we observed that the elimination of p16h-sn tumor-associated stromal cells suppressed bladder cancer initiation and progression.
Bladder cancer is one of the most challenging cancers to treat and has a poor prognosis. In the systemic therapy of metastatic urothelial carcinoma, in addition to typical cisplatin-based combination regimens or immune checkpoint inhibitors, the practicality of integrating advanced agents, such as fibroblast growth factor receptor tyrosine kinase inhibitors and enfortumab vedotin, has been identified42,43,44. However, the 5-year survival rates of patients with muscle-invasive bladder cancer are still unsatisfactory14. Therefore, the identification of the prognostic value of high expression levels of SMOC2, GUCY1A1 (GUCY1A3), CXCL12, CRISPLD2, GAS1 and LUM in both nonadvanced and advanced bladder cancer is useful for diagnosing bladder cancer that needs aggressive therapies.
Methods
Mice
Mice were housed in groups of two to five per cage at an ambient temperature of 23–25 °C in a humidity-controlled room. They were maintained on a 12-h light/dark cycle (08:00 to 20:00 light on) with standard food (CA-1, CLEA) and water provided ad libitum and with access to environmental enrichment. All animals were handled according to the Guidelines for Animal Experiments of the Institute of Medical Science of the University of Tokyo (IMSUT) and the Institutional Laboratory Animal Care. All animal experiments were approved by the Animal Experiment Committee at IMSUT (A16-33, A21-26). All p16-CreERT2-tdTomato or p16-CreERT2-DTR mice were heterozygous and generated by crossing p16INK4A-CreERT2 mice18 with Rosa26-CAG-LSL-tdTomato or Rosa26-SA-LSL-DTR-IRES-tdTomato mice (The Jackson Laboratory). For labeling of p16h-sn cells in the p16-CreERT2-tdTomato mouse model, intraperitoneal (i.p.) injections of tamoxifen (80 mg kg−1 body weight (BW)) (#T5648-5G, Sigma-Aldrich or #13258, Cayman Chemical) dissolved in sunflower oil (#196-15265, Wako) were performed for five consecutive days using a 25-gauge needle. Mice were killed 2 weeks after the first day of tamoxifen injection. We used age-matched and same-strain mice in all groups of the same experiments.
Orthotopic allograft transplantation of mouse bladder carcinoma cells
The mouse bladder carcinoma cell line MB49 (#SCC148, Sigma-Aldrich) was cultured in RPMI 1640 medium (#30264-56, Nacalai Tesque) supplemented with 10% FBS (#173012, Sigma-Aldrich) and 1× penicillin/streptomycin/amphotericin B solution (#09366-44, Nacalai Tesque). Female mice were anesthetized using isoflurane and placed in the dorsal position. The bladder was emptied by applying gentle manual pressure on the abdomen. A polyethylene catheter with a 0.28-mm inner diameter and a 0.61-mm outer diameter (#427400, BD) was inserted into the urethra. On the opposite end of the catheter, a 28-gauge needle connected to a 1-ml injection syringe was attached. Before the instillation of MB49 cells, 100 µl poly(l-lysine) hydrobromide solution (#4832, Sigma-Aldrich) was instilled through the urethra. After 30 min, the bladder was emptied again, and 1.0 × 105 MB49 cells in 100 µl PBS were administered. Mice were monitored and killed with CO2 when the bladder weight reached 10% of the BW, ulcers developed or the BW loss exceeded 20%. The maximum bladder weight, including the tumor, was 427.6 mg.
Treatments for senolysis
In p16-CreERT2-DTR, p16-CreERT2 or wild-type mice, i.p. injections of tamoxifen (80 mg kg−1 BW) dissolved in sunflower oil and DT (25 µg kg−1 BW) (#048-34371, Wako) diluted in PBS (#14249-95, Nacalai Tesque) were performed alternately for 14 consecutive days. For ABT-263 (#S1001, Selleck) treatment, ABT-263 was dissolved in ethanol/polyethylene glycol 400 (#91893, Sigma-Aldrich)/Phosal 50 PG (#GC36897, GlpBio) at 10:30:60 and orally administered (50 mg kg−1 BW) for seven consecutive days per cycle. Following the transplantation of bladder cancer cells, a total of two cycles were conducted with a 1-week interval.
CXCL12 inhibitor treatment
AMD3100 (#S3013, Selleck) was dissolved in normal saline and administered by intravesical installation through the urethra at a dose of 3.5 mg kg−1 BW. The administration was performed for five consecutive days per cycle under isoflurane anesthesia. Following the transplantation of bladder cancer cells, a total of two cycles were conducted with a 2-day interval. Considering cases in which transplanted cancer cells flowed out during treatment, bladders without cancer cells under the microscope were excluded from the analysis.
Bladder cancer model using BBN
We used BBN (#B0938, TCI America) to establish a bladder carcinogenesis model. Male wild-type mice aged 4–8 weeks (as young mice) were included in the study and provided with drinking water containing 0.05% BBN. The BBN-containing water was refreshed at least once a week to ensure its potency. The administration of BBN continued for 12 weeks, after which the mice were killed for further analysis.
scRNA-seq preparation
The bladder from a 25-month-old male p16-CreERT2-tdTomato mouse was dissected 2 weeks after i.p. injections of tamoxifen for five consecutive days. The dissected bladder tissue was minced using a blade and then subjected to agitation two times using 18- and 21-gauge needles during 1-h incubation at 37 °C in 2 ml PBS containing 5 mg collagenase type IV (#9001-12-1, Gibco) and 5,000 U ml−1 DNase I (#D4263, Sigma-Aldrich). The cell suspension was filtered through a 70-µm strainer (#352350, Falcon), followed by centrifugation (800g, 4 °C, 3 min). The pellet was then resuspended in wash buffer (4% FBS in PBS). After pelleting the cells, they were resuspended in wash buffer containing 10% FcR blocking reagent (#130-092-575, MACS) for 15 min. Subsequently, the cells were stained with an anti-CD45 antibody (#103133, BioLegend, 1:200 dilution) for 30 min at 4 °C in the absence of light. After being pelleted, cells were resuspended in wash buffer containing 0.1 µg ml−1 DAPI (#4083S, Cell Signaling Technology) to exclude dead cells during sorting. The CD45−/DAPI−/Tomato+ and CD45−/DAPI−/Tomato− cells were sorted using BD FACSAria SORP (BD Biosciences).
scRNA-seq data analysis
Using the 10x Genomics Chromium Single Cell 3′ reagent kit (version 3.1), we constructed the scRNA-seq libraries of Tomato− and Tomato+ bladder cells. The libraries were sequenced using the DNBSEQ-G400RS sequencer (MGI Tech), and approximately 2,000 and 500 million reads were obtained for Tomato− and Tomato+ cells, respectively. For sequence alignment, unique molecular identifiers and barcodes were recognized, and transcripts were mapped to a customized genome reference including mm10 with tdTomato sequences18 through the Cell Ranger package (version 7.0.0). The Seurat (version 4.3.0), dplyr (version 1.1.1), circlize (version 0.4.15), ComplexHeatmap (version 2.10.0), ggplot2 (version 3.4.2) and ggbeeswarm (version 0.7.2) packages in R (version 4.1.3) were used to process scRNA-seq data45,46. GO analysis was performed using Metascape47. Genes related to red blood cells (Hbb-bs, Hba-a1, Hba-a2 and Hbb-bt) were excluded from the analysis.
Lentivirus-mediated gene transfer of Venus into MB49
The generation of lentiviruses and their infection into cells were performed as described previously48. Lentiviruses expressing the respective genes were generated by cotransfection of HEK293T cells with the pCMV-VSV-G-RSV-RevB, pCAG-HIVgp and CSII-EF-MCS-IRES2-Venus plasmids using the calcium phosphate coprecipitation method. The lentiviruses were infected into MB49 cells, and Venus+ cells were sorted using BD FACSAria SORP (BD Biosciences).
shRNA constructs and lentivirus-mediated gene transfer into MB49
pLKO.1-puro, pLKO.1-puro shRNA scramble, pMD2.G and psPAX2 were purchased from Addgene. We used the In-Fusion HD cloning kit (Clontech) to insert the PCR-amplified fragment of BamHI-tRFP657-KpnI into BamHI/KpnI-digested pLKO.1-puro or pLKO.1-puro shRNA scramble to replace the puromycin-resistant cassette with the tRFP657 gene, generating the pLKO.1-tRFP657 and pLKO.1-tRFP657 shRNA scramble plasmids. pLKO.1-puro and pLKO.1-tRFP657 were digested by EcoRI/AgeI and ligated with the annealing products of shCXCR4 and shCXCR7 oligonucleotides, respectively. Oligonucleotide sequences are shown in Supplementary Table 1. pLKO.1, pMD2.G, psPAX2 and PEI-MAX (Polysciences) were mixed at a ratio of 2:1:1:16 and transfected into HEK293T cells to produce lentiviruses. After infecting MB49 cells with the lentivirus, we used 2 µg ml−1 of puromycin for selection and sorted tRFP657+ cells using BD FACSAria SORP (BD Biosciences).
SPiDER β-gal staining
The bladders from young male p16-CreERT2-tdTomato mice were dissected 2 weeks after five consecutive days of i.p. injections of tamoxifen. The cell suspension was prepared following the same method as described above. After blocking, SPiDER β-gal staining (#SG03, Dojindo) was performed according to the manufacturer’s instructions. For other staining procedures, anti-CD45 antibody, anti-Ter119 antibody (#116233, BioLegend, 1:200 dilution) and DAPI were used. Using BD FACSAria III (BD Biosciences), we calculated the change in the median fluorescence intensity (ΔMFI) of FITC signals as the difference in the median FITC (β-gal) intensities between CD45−/Ter119−/DAPI−/Tomato− or Tomato+ cell populations with and without SPiDER β-gal treatment.
Detection of immune cells in bladder tumors
The bladders of young female p16-CreERT2-tdTomato mice transplanted with MB49 cells were prepared as described above. Bladder tumors were divided into two parts, and two types of antibody combinations—(1) CD45, CD11b (#11-0112-82, eBioscience, 1:50 dilution), F4/80 (#123128, BioLegend, 1:50 dilution) and Ly6G (#127609, BioLegend, 1:400 dilution) or (2) CD45, CD3 (#100217, BioLegend, 1:50 dilution), B220 (#103205, BioLegend, 1:100 dilution) and NK.1.1 (#108719, BioLegend, 1:1,000 dilution)—were used to detect immune cells. Fixable Viability Dye eFluor 780 (#65-0865-14, Thermo Fisher Scientific) was used to exclude dead cells. Using BD FACSAria, we identified Tomato− or Tomato+ immune cells as neutrophils (CD45+CD11b+Ly6G+F4/80mid), monocytes (CD45+CD11b+Ly6G−F4/80−), macrophages (CD45+CD11b+F4/80high), T cells (CD45+CD3+), natural killer cells (CD45+NK.1.1+) or B cells (CD45+B220+).
Immunohistochemistry
For histological evaluation, sections (4–6 µm in thickness) of paraffin-embedded bladder tissue were used. In immunohistochemistry (IHC) staining, sections were stained with an anti-RFP (#PM005, MBL, 1:500 dilution), anti-CXCL12 (#25117, Abcam, 1:1,000 dilution), anti-MKI67 (#16667, Abcam, 1:100 dilution), anti-CD31 (#28364, Abcam, 1:50 dilution), anti-cleaved caspase-3 (#9664S, Cell Signaling Technology, 1:250 dilution), anti-CD3 (#135372, Abcam, 1:150 dilution), anti-F4/80 (#6640, Abcam, 1:250 dilution) or anti-CD105 (#221675, Abcam, 1:50 dilution) antibody by conventional immunostaining using DAB (#415171, Nichirei Biosciences). For immunofluorescence staining, an anti-mCherry (#205402, Abcam, 1:1,000 dilution), anti-MKI67 (1:100 dilution), anti-vimentin (#5741S, Cell Signaling Technology, 1:50 dilution), anti-CK5 (#64081, Abcam, 1:100 dilution), anti-αSMA (#19245, Cell Signaling Technology, 1:50 dilution) or anti-CD31 antibody was used as a primary antibody and AF647 anti-chicken IgY (#A32933, Thermo Fisher Scientific, 1:1,000 dilution) or AF488 anti-rabbit IgG (#11034, Thermo Fisher Scientific, 1:1,000 dilution) antibody was used as a secondary antibody. Hoechst (#62249, Thermo Fisher Scientific, 1:1,000 dilution) was used to stain nuclei. An IN Cell Analyzer 2500 HS system (Cytiva) or a BZ-X810 fluorescence microscope (Keyence) was used to image fluorescence signals. A single-blind design was used to assess the levels of CXCL12 expression in the entire bladder tissue. The expression was categorized into ‘none’, ‘low’, ‘intermediate’ or ‘high’. The percentages of MKI67+ cells and the CD31+ area were calculated using ImageJ software.
Immunocytochemistry
For the isolation and immunocytochemistry of fibroblasts, the bladders from young male p16-CreERT2-tdTomato mice were dissected 2 weeks after five consecutive days of i.p. injections of tamoxifen. For iCAFs, the bladders from young female p16-CreERT2-tdTomato mice treated with tamoxifen and transplanted with cancer cells were dissected 28 days after transplantation. The cell suspension was prepared following the same method as described above. For cell staining, anti-CD45, anti-Ter119 and anti-CD140a (#135907, BioLegend, 1:50 dilution) antibodies were used. CD45−/Ter119−/DAPI−/CD140a+ fibroblasts or iCAFs (0.7–3.0 × 104) were sorted using FACS SORP Aria and dropped into high-glucose DMEM (#08458-16, Nakalai Tesque) with 10% FBS and 1× penicillin/streptomycin/amphotericin B solution. Sorted fibroblasts or iCAFs were incubated with brefeldin A (#00-4506-51, Thermo Fisher Scientific, 1:1,000 dilution) for 1 h at 37 °C and collected on a glass plate by centrifugation at 1,200 rpm for 5 min using CytoSpin 4 (Thermo Fisher Scientific). Fixation was performed using 4% paraformaldehyde in PBS (#9154-56, Nakalai Tesque) for 10 min at room temperature (RT). After three washes with 0.5% FBS in PBS, permeabilization was performed using 0.1% Triton X-100 (#35501-15, Nakalai Tesque) in PBS for 5 min at RT. After washing, blocking was performed using 5% BSA (#A2153, Sigma-Aldrich) in PBS, followed by incubation with an anti-CXCL12 antibody (1:100 dilution) for 2 h at RT. After washing, cells were incubated with AF488 anti-rabbit IgG antibody (#150117, Abcam, 1:500 dilution) in 0.5% FBS in PBS for 1 h at RT without light. After washing, cells were mounted and evaluated using fluorescence microscopy (BZ-X810). For p16 or p21 staining, CD45−/Ter119−/DAPI− cells from the bladders of young male p16-CreERT2-tdTomato mice were sorted, and a p16 antibody (#211542, Abcam, 1:100 dilution) or a p21 antibody (#188224, Abcam, 1:100 dilution) was used following the same method as described above. For LMNB1 staining, CD45−/Ter119−/DAPI−cells from the bladders of middle-aged male p16-CreERT2-tdTomato mice were sorted, and an anti-LMNB1 antibody (#16048, Abcam, 1:500 dilution) was used following the same method as described above.
Culture of MB49 lines using conditioned medium from iCAFs
For the conditioned medium experiment, 2.0–7.0 × 104 iCAFs were sorted and cultured in 600 µl high-glucose DMEM with 1× penicillin/streptomycin/amphotericin B solution (without 10% FBS) on a 24-well plate for 48 h at 37 °C, 5% CO2 and 5% O2. For each experiment, the numbers of Tomato− iCAFs and total iCAFs were aligned. MB49 cells were precultured with high-glucose DMEM with 1× penicillin/streptomycin/amphotericin B solution (without 10% FBS) for 48 h as a starvation condition. A 100-µl volume of the medium was collected and used to treat 1.0 × 104 cells of each MB49 line at 37 °C and 5% CO2 for 48 h. Cell counting was performed using CCK-8 (#CK04, Dojindo) according to the manufacturer’s instructions, and absorbance was measured using CLARIOstar Plus (BMG Labtech). Relative proliferation (%) was calculated as follows: (absorbance of MB49 cells treated with conditioned medium from total iCAFs)/(absorbance of MB49 cells treated with conditioned medium from Tomato− iCAFs) × 100.
Immunoblotting
Protein was extracted from homogenized cancer tissue or sorted cells from cancer tissue using Laemmli buffer (10% glycerol, 2% SDS, 50 mM Tris at pH 6.8, 0.01% bromophenol blue, 1% 2-mercaptoethanol). As primary antibodies, anti-AKT (#9272S, Cell Signaling Technology; 1:1,000 dilution), anti-pAKT (#4060S, Cell Signaling Technology, 1:2,000 dilution) and anti-ACTB (#69879, Santa Cruz, 1:2,000 dilution) antibodies were used. A peroxidase-conjugated secondary antibody (#NA931V or #NA934V, Cytiva, both 1:3,000 dilution) was used, and the antigen–antibody reaction was visualized using Amersham ECL detection reagents (#RPN2106, Cytiva). Images were captured using Amersham Imager 680 (Cytiva) or DIP500A (Daito Mitech). The integrated pixel intensities of pAKT/AKT were calculated using ImageJ software. For the experiment using intact MB49, shControl and shCXCR4-1/CXCR7-1 lines (Extended Data Fig. 9d), all cell lines were pretreated with high-glucose DMEM with 1× penicillin/streptomycin/amphotericin B solution (without 10% FBS) for 48 h as a starvation condition before protein extraction. Recombinant mouse CXCL12 (100 ng ml−1) (#460-SD-010/CF, RSD) was used for treatment.
Quantitative real-time PCR
Total RNA was extracted using an RNeasy mini kit (#74104, Qiagen) according to the manufacturer’s instructions. cDNAs were synthesized using the ReverTra Ace qPCR reverse transcription kit (#FSQ-101, Toyobo). For RNA extraction from sorted cells, the QuantAccuracy RT-RamDA cDNA synthesis kit (#RMQ-101, Toyobo) was used according to the manufacturer’s instructions. Real-time PCR amplifications were performed in 96-well optical reaction plates with THUNDERBIRD SYBR qPCR mix (#QPS-201, Toyobo). The relative expression of each gene was determined by normalization to Actb expression for each sample. p16: forward primer, CGTACCCCGATTCAGGTGAT; reverse primer, TTGAGCAGAAGAGCTGCTACGT. p21: forward primer, CCTGGTGATGTCCGACCTG; reverse primer, CCATGAGCGCATCGCAATC. p15: forward primer, GACCCTGCCACCCTTACCAGACC; reverse primer, CAGGCGTCACACACATCCAGCC. Glb1: forward primer, CTTCCCACTGAACACTGAGGC; reverse primer, TTGGCACGAACAAGGTCTTTT. Cxcl12: forward primer, TGCATCAGTGACGGTAAACCA; reverse primer, CACAGTTTGGAGTGTT. Actb: forward primer, GGCTGTATTCCCCTCCATCG; reverse primer, CCAGTTGGTAACAATGCCATGT. Cxcr4: forward primer, AAGCACGTTCTTGTTAGGCAT; reverse primer, AGAAGGGGAGTGTGATGACAAA. Cxcr7: forward primer, GCCTGGCAACTACTCTGACAT; reverse primer, GACTGGCATAGTCGGCAATG.
Cytokine detection in transplanted bladder cancer
Using the mouse cytokine antibody array kit panel A (#ARY006, R&D Systems), we evaluated cytokines in transplanted bladder cancer. Tissue lysates from transplanted bladder cancer were prepared, and procedures were performed according to the manufacturer’s instructions. Spot images were detected with the Amersham Imager 680 system (Cytiva). Data were analyzed using ImageJ software, and the normalized mean pixel densities of each spot were calculated.
Human data analysis
The data of 436 patients with bladder cancer from TCGA (Urothelial Bladder Carcinoma collection) were extracted through the University of California Santa Cruz Xena platform49. Regarding mRNA microarray data, the samples were from 136 patients with bladder cancer who had undergone transurethral resection of the bladder tumor for the first time from May 2008 to January 2020 at Fukushima Medical University. The procedure from RNA extraction to data normalization was performed as described previously50. Clinical information was extracted from the electronic health record system. The approval numbers from the ethics committee of Fukushima Medical University were 1953 and 2097, and the procedures conformed to the ethical guidelines of the 1975 Declaration of Helsinki. Informed consent was obtained from all participants, and participants did not receive compensation for their participation. Information on the participants’ sex and age is shown in Supplementary Table 2. For the p16h-sn CAF signature, the z scores of each gene were calculated, and the average of the z scores of the six genes were evaluated as the ‘p16h-sn CAF signature’ for each patient.
Statistics and reproducibility
For animal experiments, all mice with the same genotype were randomly assigned to each group, and the same age-dependent schedule was followed in each experimental design. No statistical methods were used to predetermine sample sizes, but our sample sizes are similar to those reported in previous publications18,51. The blind design was adopted only for the evaluation of CXCL12 expression in bladder cancer between the senolysis and control groups (single-blind). For all the other evaluations, data collection and analysis were not performed blind to the conditions of the experiments. For the quantification of images, more than five fields were randomly selected to prevent cell composition bias, although fewer than five fields were selected when the suitable area in IHC or the number of sorted cells in immunocytochemistry was too small. Data are presented as mean ± s.e.m. unless otherwise noted. Data distribution was assumed to be normal, but this was not formally tested. Comparisons between two groups were made by a paired-sample two-tailed t test or an unpaired two-tailed Student’s or Welch’s t test. Multiple comparisons of one-variable data were carried out by one-way analysis of variance (ANOVA) followed by a post hoc Tukey’s test or the Games–Howell test. Overall survival was calculated using the Kaplan–Meier method, and comparisons were made using the log-rank test. The raw lifespan data of mice and humans are provided in Supplementary Table 3. The graphs and the statistical analysis results provided in Extended Data Fig. 10b were directly extracted from the Xena platform49, and the raw data are not provided.
For the evaluation of the correlation coefficient among genes, the Pearson product-moment correlation test was performed. Cox proportional hazards analysis was performed to calculate hazard ratios. For preclinical data, outlier values were excluded from data analysis according to the 1.5 × interquartile range rule in case the number of samples was ten or more. For clinical data, outlier values were excluded from data analysis according to the 1.5 × interquartile range rule. Statistical analyses were performed using the SPSS software package (version 26, IBM) or Microsoft Excel (Microsoft). For consistency in comparisons, significance in all figures is denoted as follows: *P < 0.05, **P < 0.01, ***P < 0.005, ****P < 0.001. For all representative findings, triplicate or multiple independent experiments were performed, and similar results were obtained.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The scRNA-seq data described in this article are available in the Gene Expression Omnibus (GEO) with accession no. GSE253338. The construction of the artificial tdTomato reference genome was previously described18. All other data needed to evaluate the conclusions of the study are present in the article and/or supplementary materials. Source data are provided with this paper. All other data are available from the corresponding authors upon reasonable request.
References
Lenis, A. T., Lec, P. M., Chamie, K. & Mshs, M. D. Bladder cancer: a review. JAMA 324, 1980–1991 (2020).
Shariat, S. F. et al. The effect of age and gender on bladder cancer: a critical review of the literature. BJU Int. 105, 300–308 (2010).
Siegel, R. L., Miller, K. D., Fuchs, H. E. & Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 72, 7–33 (2022).
Martínez-Jiménez, F. et al. A compendium of mutational cancer driver genes. Nat. Rev. Cancer 20, 555–572 (2020).
Takeshima, H. & Ushijima, T. Accumulation of genetic and epigenetic alterations in normal cells and cancer risk. NPJ Precis. Oncol. 3, 7 (2019).
Peto, R., Roe, F. J., Lee, P. N., Levy, L. & Clack, J. Cancer and ageing in mice and men. Br. J. Cancer 32, 411–426 (1975).
Fane, M. & Weeraratna, A. T. How the ageing microenvironment influences tumour progression. Nat. Rev. Cancer 20, 89–106 (2020).
Quail, D. F. & Joyce, J. A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 19, 1423–1437 (2013).
Sahai, E. et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 20, 174–186 (2020).
Chen, X. & Song, E. Turning foes to friends: targeting cancer-associated fibroblasts. Nat. Rev. Drug Discov. 18, 99–115 (2019).
Öhlund, D., Elyada, E. & Tuveson, D. Fibroblast heterogeneity in the cancer wound. J. Exp. Med. 211, 1503–1523 (2014).
Biffi, G. & Tuveson, D. A. Diversity and biology of cancer-associated fibroblasts. Physiol. Rev. 101, 147–176 (2021).
Di Micco, R., Krizhanovsky, V., Baker, D. & d’Adda di Fagagna, F. Cellular senescence in ageing: from mechanisms to therapeutic opportunities. Nat. Rev. Mol. Cell Biol. 22, 75–95 (2021).
Sadeghi, S. et al. Phase II California cancer consortium trial of gemcitabine–eribulin combination in cisplatin-ineligible patients with metastatic urothelial carcinoma: final report (NCI-9653). J. Clin. Oncol. 37, 2682–2688 (2019).
Messing, E. M. et al. Effect of intravesical instillation of gemcitabine vs saline immediately following resection of suspected low-grade non-muscle-invasive bladder cancer on tumor recurrence: SWOG S0337 randomized clinical trial. JAMA 319, 1880–1888 (2018).
Kamat, A. M. et al. Bladder cancer. Lancet 388, 2796–2810 (2016).
Yasuda, T. et al. Inflammation-driven senescence-associated secretory phenotype in cancer-associated fibroblasts enhances peritoneal dissemination. Cell Rep. 34, 108779 (2021).
Omori, S. et al. Generation of a p16 reporter mouse and its use to characterize and target p16high cells in vivo. Cell Metab. 32, 814–828 (2020).
Dimri, G. P. et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl Acad. Sci. USA 92, 9363–9367 (1995).
Yu, Z. et al. Single-cell transcriptomic map of the human and mouse bladders. J. Am. Soc. Nephrol. 30, 2159–2176 (2019).
Kim, Y. H. et al. Senescent tumor cells lead the collective invasion in thyroid cancer. Nat. Commun. 8, 15208 (2017).
Du, Y. et al. Comprehensive analysis of CXCL12 expression reveals the significance of inflammatory fibroblasts in bladder cancer carcinogenesis and progression. Cancer Cell Int. 21, 613 (2021).
López-Gil, J. C., Martin-Hijano, L., Hermann, P. C. & Sainz, B.Jr. The CXCL12 crossroads in cancer stem cells and their niche. Cancers 13, 469 (2021).
Buch, T. et al. A Cre-inducible diphtheria toxin receptor mediates cell lineage ablation after toxin administration. Nat. Methods 2, 419–426 (2005).
Chang, J. et al. Clearance of senescent cells by ABT263 rejuvenates aged hematopoietic stem cells in mice. Nat. Med. 22, 78–83 (2016).
Nagao, M., Suzuki, E., Yasuo, K., Yahagi, T. & Seino, Y. Mutagenicity of N-butyl-N-(4-hydroxybutyl)nitrosamine, a bladder carcinogen, and related compounds. Cancer Res. 37, 399–407 (1977).
Barbero, S. et al. Stromal cell-derived factor 1α stimulates human glioblastoma cell growth through the activation of both extracellular signal-regulated kinases 1/2 and Akt. Cancer Res. 63, 1969–1974 (2003).
Majka, M., Ratajczak, J., Kowalska, M. A. & Ratajczak, M. Z. Binding of stromal derived factor-1α (SDF-1α) to CXCR4 chemokine receptor in normal human megakaryoblasts but not in platelets induces phosphorylation of mitogen-activated protein kinase p42/44 (MAPK), ELK-1 transcription factor and serine/threonine kinase AKT. Eur. J. Haematol. 64, 164–172 (2000).
Fricker, S. P. Physiology and pharmacology of plerixafor. Transfus. Med. Hemother. 40, 237–245 (2013).
Pereira, B. A. et al. CAF subpopulations: a new reservoir of stromal targets in pancreatic cancer. Trends Cancer 5, 724–741 (2019).
Chen, Z. et al. Single-cell RNA sequencing highlights the role of inflammatory cancer-associated fibroblasts in bladder urothelial carcinoma. Nat. Commun. 11, 5077 (2020).
Caramelo, B., Zagorac, S., Corral, S., Marqués, M. & Real, F. X. Cancer-associated fibroblasts in bladder cancer: origin, biology, and therapeutic opportunities. Eur. Urol. Oncol. 6, 366–375 (2023).
Fulop, T. et al. Immunosenescence and inflamm-aging as two sides of the same coin: friends or foes? Front. Immunol. 8, 1960 (2018).
Ma, Z. et al. Interferon-dependent SLC14A1+ cancer-associated fibroblasts promote cancer stemness via WNT5A in bladder cancer. Cancer Cell 40, 1550–1565 (2022).
Aunan, J. R., Cho, W. C. & Søreide, K. The biology of aging and cancer: a brief overview of shared and divergent molecular hallmarks. Aging Dis. 8, 628–642 (2017).
Zinger, A., Cho, W. C. & Ben-Yehuda, A. Cancer and aging—the inflammatory connection. Aging Dis. 8, 611–627 (2017).
Lee, S. & Schmitt, C. A. The dynamic nature of senescence in cancer. Nat. Cell Biol. 21, 94–101 (2019).
Gabai, Y., Assouline, B. & Ben-Porath, I. Senescent stromal cells: roles in the tumor microenvironment. Trends Cancer 9, 28–41 (2023).
Affo, S. et al. Promotion of cholangiocarcinoma growth by diverse cancer-associated fibroblast subpopulations. Cancer Cell 39, 866–882 (2021).
Lavie, D., Ben-Shmuel, A., Erez, N. & Scherz-Shouval, R. Cancer-associated fibroblasts in the single-cell era. Nat. Cancer 3, 793–807 (2022).
Nagarsheth, N., Wicha, M. S. & Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 17, 559–572 (2017).
Witjes, J. A. et al. Updated 2016 EAU guidelines on muscle-invasive and metastatic bladder cancer. Eur. Urol. 71, 462–475 (2017).
Loriot, Y. et al. Erdafitinib in locally advanced or metastatic urothelial carcinoma. N. Engl. J. Med. 381, 338–348 (2019).
Rosenberg, J. E. et al. Pivotal trial of enfortumab vedotin in urothelial carcinoma after platinum and anti-programmed death 1/programmed death ligand 1 therapy. J. Clin. Oncol. 37, 2592–2600 (2019).
Satija, R., Farrell, J. A., Gennert, D., Schier, A. F. & Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 33, 495–502 (2015).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587 (2021).
Zhou, Y. et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 10, 1523 (2019).
Johmura, Y. et al. SCFFbxo22-KDM4A targets methylated p53 for degradation and regulates senescence. Nat. Commun. 7, 10574 (2016).
Goldman, M. J. et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nat. Biotechnol. 38, 675–678 (2020).
Watanabe, T. et al. Prediction of lymphovascular space invasion in endometrial cancer using the 55-gene signature selected by DNA microarray analysis. PLoS ONE 14, e0223178 (2019).
Wang, T.-W. et al. Blocking PD-L1–PD-1 improves senescence surveillance and ageing phenotypes. Nature 611, 358–364 (2022).
Acknowledgements
We thank C. Konishi, Y. Chiba, M. Ishiura, T. Ando, M. Takagi and R. Honma for providing technical assistance. The supercomputing resource was supplied by the Human Genome Center (University of Tokyo). This study was supported by the Pathology Core Laboratory and FACS Core Laboratory, Institute of Medical Science, University of Tokyo; the Japan Agency for Medical Research and Development (AMED) under grant nos. JP23zf0127003h (M.N.), JP23gm1410013h (M.N.), JP20gm5010001s (M.N.), JP20ck010655h (M.N.) and JP21gm6410014h (Y.J.); the MEXT/Japan Society for the Promotion of Science KAKENHI under grant nos. JP20H00514 (M.N.), JP20K21497 (M.N.), JP19H05740 (M.N.), JP19H03431 (Y.J.) and JP20H04940 (Y.J.); and the Princess Takamatsu Cancer Research Fund (M.N.).
Author information
Authors and Affiliations
Contributions
M.N. and Y.J. conceived the project. S.M., Y.J., T.-W.W., S.K. and M.N. planned the experiments. S.M., S.T. and S.O. performed the experiments. S.M., Y.J., T.-W.W., S.K., Y.T.O., S. Hoshi, E.K., K.Y., S. Hatakeyama, S.Y., E.S., S.I., Y.F., Y.K. and M.N. analyzed the results. M.N., S.M. and T.-W.W. wrote the paper, with editing by all the other authors.
Corresponding authors
Ethics declarations
Competing interests
M.N. is a scientific advisor and shareholder at reverSASP Therapeutics. S.Y. is a co-founder of Celaid Therapeutics. The other authors declare no competing interests.
Peer review
Peer review information
Nature Aging thanks Valery Krizhanovsky, Juan Pedro Martínez-Barbera and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Characteristics of p16h cells as senescent cells.
a, qPCR using the indicated primers on the sorted cells from young (7–12-week-old) male p16-CreERT2-Tomato mice. (p16, n = 9; p21, n = 10; p15, n = 11). b, qPCR using the primer of Glb1 on the sorted cells of young (7–12-week-old) male p16-CreERT2-Tomato mice. (n = 6). c, Representative Immunofluorescence image of sorted cells from young (7–12-week-old) male p16-CreERT2-Tomato mice as the indicated staining (left) and the percentages of p16+ cells in Tomato− (p16l) and Tomato+ (p16h) cells (right). The arrows indicate the representative Tomato+ cells and the arrowheads indicate the representative p16+ cells. Scale bar, 100 μm. (n = 4). d, Representative Immunofluorescence image of sorted cells from young (7–12-week-old) male p16-CreERT2-Tomato mice as the indicated staining (left) and the percentages of p21+ cells in Tomato− (p16l) and Tomato+ (p16h) cells (right). Scale bar, 50 μm. (n = 4). e, Representative immunofluorescence image of the bladder as the indicated staining (left). The samples were from 24–28-month-old male p16-CreERT2-Tomato mice. The Tomato proteins were stained by using an anti-mCherry antibody. The arrows indicate the representative Tomato+ cells and the arrowheads indicate the representative MKI67+cells. Scale bar, 50 µm. The percentages of MKI67+cells in Tomato− (p16l) and Tomato+ (p16h) cells from the mouse bladders (right). (n = 4). f, Representative immunofluorescent image of non-immune cells sorted from the bladders as the indicated staining (left). The samples were from 12–16-month-old male p16-CreERT2-Tomato mice. The arrows indicate the representative Tomato+ cells and the arrowheads indicate the representative LMNB1+cells. Scale bar, 100 µm. The percentages of LMNB1+ cells in Tomato− (p16l) and Tomato+ (p16h) cells from the mouse bladders (right). (n = 4). Paired t-test was performed for a–f. Data in a–f are presented as mean values ± SEM. All the t-test were two−side.
Extended Data Fig. 2 Related Fig. 2; Single-cell RNA sequence analysis of p16h cells in aged male mouse bladder.
a, Scheme of scRNA libraries preparation using non-immune Tomato− (p16l) and Tomato+ (p16h-sn) bladder cells from a 25-month-old male p16-CreERT2-tdTomato mouse. b, Gating strategy for sorting non-immune Tomato− (p16l) and Tomato+ (p16h-sn) bladder cells. c, Representative Immunofluorescence images of aged (22–26-month-old) male mouse bladder as the indicated staining. White arrowheads indicate double positive cells for both Tomato and each indicated protein. Scale bar, 100 μm. d, Percentage of cell type among Tomato+ (p16h-sn) cells. (n = 8 for each cell type). e, qPCR using the primer of Cxcl12 on sorted fibroblasts from young (7–12-week-old) male p16-CreERT2-Tomato mice. (n = 6). Paired t-test was performed for e. Data in d, e are presented as mean values ± SEM. All the t-test were two−side.
Extended Data Fig. 3 Related Fig. 3; Elimination of p16h-sn cells suppresses bladder cancer growth.
a, Experimental design of bladder allograft model using young (7–12-week-old) female p16-CreERT2-Tomato mice. MB49 cells were transplanted intravesically through the urethra. b, Experimental design of the bladder allograft model using young (7–12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice. c, Representative immunofluorescence images of transplanted bladder tumors treated with tamoxifen alone or tamoxifen+DT in young (7–12-week-old) female p16-CreERT2-DTR mice (left). Tomato protein was stained with an anti-mCherry antibody. Scale bar, 100 μm. The number of Tomato+ (p16h-sn) cells/field in the transplanted bladder tumor under tamoxifen alone or tamoxifen+DT (right). (n = 3 for each group. d, Representative images of bladder without MB49 cell transplantation using young (7–12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice (left) and bladder weights (right). Scale bar, 5 mm. (n = 3 for each group). e, Representative HE staining images of the bladders without MB49 cell transplantation in young (7–12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice (left). Bladder wall thickness is shown on the right panel. Scale bar, 400 μm. (n = 3 for each group). f, Weights of bladder+tumor or tumor only. Weights of “tumor” were calculated as: transplanted bladder weights in Fig. 3c minus the average of bladder weights without cancer in Extended Data Fig. 3d. p16-CreERT2, n = 10; p16-CreERT2-DTR, n = 11. g, Experimental design of the bladder allograft model with ABT-263 or vehicle administration using young (7–12-week-old) female wild-type mice. h, Experimental design of bladder allograft after tamoxifen treatment, followed by ABT-263 or vehicle administration in young (7–12-week-old) female p16-CreERT2-Tomato mice. i, Representative immunofluorescence images of transplanted bladder tumor on the design of h (left) and the number of Tomato+ (p16h-sn) cells/field in transplanted bladder tumors among ABT-263 or vehicle group (right). Tomato protein was stained with an anti-mCherry antibody. Scale bar, 200 μm. (Vehicle, n = 4; ABT-263, n = 3). j, Experimental design of bladder allograft with double treatment of tamoxifen+DT and ABT-263 or vehicle using young (7–14-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice. k, Bladder weights on the design of j. tamoxifen+DT+vehicle: p16-CreERT2, n = 11; p16-CreERT2-DTR, n = 8. tamoxifen+DT + ABT-263: p16-CreERT2, n = 11; p16-CreERT2-DTR, n = 7. Unpaired t-test was performed for c, d, e, f and i. One-way ANOVA with Games-Howell’s test was performed for k. Data in c, d, e, and i are presented as mean values ± SEM. For all box plots, notch of box blots indicates median value. Lower and upper bounds correspond to 25th and 75th percentiles, respectively. Cross mark indicates mean value. Whiskers extend to maximum or minimum value, with points outside this range drawn individually. All the t-test were two-side.
Extended Data Fig. 4 Related Fig. 3; p16h-sn cancer stromal cells in tumors were present in bladder tissue before transplantation.
a, Experimental labeling design of p16h-sn cells before (pre-labeling group, top panel) or after (post-labeling group, bottom panel) the transplantation of MB49 using young (7–12-week-old) or middle-aged (12–16-month-old) female p16-CreERT2-tdTomato mice. b, Low magnification images of young mice group in Fig. 3g. The areas circled by dashed lines indicated cancer area. Tomato protein was stained with an anti-mCherry antibody. Scale bar, 500 μm. c, Experimental design of the bladder allograft model using young (7–12-week-old) or middle-aged (12–16-month-old) female wild-type mice. d, Bladder weights on the design of c. (young, n = 9; middle-aged, n = 3). e, Survival probability of the young (7–12-week-old) or middle-aged (12–16-month-old) group after cancer transplantation. (young, n = 10; middle-aged, n = 10). f, Experimental design of senolysis followed by MB49 transplantation using young (7–12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice. g, Experimental design of senolysis with ABT-263 followed by MB49 transplantation using young (7–12-week-old) female wild-type mice. h, Image of bladders on the design of g. Scale bar, 1 cm. i, Bladder weights of the samples shown in h. (n = 9 for each group). Unpaired t-test was performed for d and i. For all box plots, notch of box blots indicates median value. Lower and upper bounds correspond to 25th and 75th percentiles, respectively. Cross mark indicates mean value. Whiskers extend to maximum or minimum value, with points outside this range drawn individually. All the t-test were two-side.
Extended Data Fig. 5 Elimination of p16h–sn cells suppresses bladder tumorigenesis.
a, Experimental design of BBN-induced bladder cancer model using young (7–12-week-old) male p16-CreERT2-Tomato mice. b, Representative HE staining images of the bladder on the design of a. Scale bar: low magnification image, 1 mm; high magnification image, 200 μm. c, Representative IHC images of the bladder on the design of a. Tomato proteins were stained by using an anti-RFP antibody. The area circled by dashed lines shows the cancer region. Scale bar, 200 μm. d, Experimental design of p16h cell labelling before (pre-labeling group, top panel) or after (post-labeling group, bottom panel) BBN treatment using young (7–12-week-old) male p16-CreERT2-tdTomato mice. e, Representative immunofluorescence images of bladder tumors on the design from d (left). Tomato protein was stained with an anti-mCherry antibody. Scale bar, 100 μm. The number of Tomato+ (p16h-sn) cells/field in bladder tumor among pre- or post-labeling group (right). (n = 5 for each group). f, Experimental design of BBN-induced bladder cancer model using young (7–12-week-old) male p16-CreERT2, wild-type or p16-CreERT2 –DTR mice. g, Representative images of the bladders in young male p16-CreERT2 or p16-CreERT2 -DTR mice on the design of f. Scale bar, 5 mm. h, Bladder weights on the design of f. (p16-CreERT, n = 12; wild-type, n = 5; p16-CreERT2-DTR, n = 12). i, Tumor area in bladders on the design of f. (p16-CreERT, n = 12; wild-type, n = 5; p16-CreERT2-DTR, n = 12). j, The number of exophytic tumor in the lumen of bladders on the design of f. (p16-CreERT, n = 12; wild-type, n = 5; p16-CreERT2-DTR, n = 12). Unpaired t-test was performed for e and h–j. Data in e are presented as mean values ± SEM. For all box plots, notch of box blots indicates median value. Lower and upper bounds correspond to 25th and 75th percentiles, respectively. Cross mark indicates mean value. Whiskers extend to maximum or minimum value, with points outside this range drawn individually. All the t-test were two-side.
Extended Data Fig. 6 Related Fig. 4; Essential role of CXCL12 secreted by p16h–sn stromal cells in bladder cancer progression.
a, Relative protein amounts of CXCL12 in transplanted bladder cancer from young (7–12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice using cytokine array. (p16-CreERT2, n = 3; p16-CreERT2-DTR, n = 4). b, Representative IHC images of bladder with MB49 cells transplanted into young (7–12-week-old) female wild-type mice with vehicle (left top) or ABT-263 (left bottom) using an anti-MKI67 antibody. The percentage of MKI67+ cells is shown in the right panel. Scale bar, 100 μm. (n = 6 for each group). c, Representative IHC images of the bladders with MB49 cell transplantation in young (7–12-week-old) female p16-CreERT2 (top left) or p16-CreERT2-DTR (bottom left) mice using an anti-CD105 antibody. The percentage of CD105+ area was shown in the right panel. Scale bar, 100 μm. (n = 6 for each group). d, Immunoblotting images using the indicated antibodies on the indicated cultured cell lines with or without treatment with CXCL12 recombinant protein. e, qPCR of Cxcr4 and Cxcr7 on the indicated cell lines. (n = 3 for each group). f, Experimental design of transplantation of the indicated cell lines and AMD3100 treatment using young (7–12-week-old) female wild-type mice. g, Immunoblotting images using lysates of bladder tumor originated from transplanted parental MB49 cells in young (7–12-week-old) female wild-type mice. Treatment with vehicle or AMD3100 was performed on the design of f. h, Experimental design of bladder allografts with the double treatment of tamoxifen+DT and AMD3100 using young (7–12-week-old) female p16-CreERT2 or p16-CreERT2-DTR mice. i, Bladder weights on the design of h. (p16-CreERT2, n = 6; p16-CreERT2-DTR, n = 5). Unpaired t-test was performed for a–c and i. One-way ANOVA with Games-Howell’s test was performed for e. Data in a–c and e are presented as mean values ± SEM. For all box plots, notch of box blots indicates median value. Lower and upper bounds correspond to 25th and 75th percentiles, respectively. Cross mark indicates mean value. Whiskers extend to maximum or minimum value, with points outside this range drawn individually. All the t-test were two-side.
Extended Data Fig. 7 Related Fig. 5; CXCL12 is predominantly secreted by p16h-sn iCAFs.
a, UMAP showing the expression levels of Cxcl12 in Tomato+ (p16h-sn) cells from scRNA-seq data using the aged male mouse bladder. b, Violin plots showing the expression levels of Cxcl12 in Tomato+ (p16h-sn) cells of each cell type. c, qPCR of p16 and p21 using the sorted iCAFs from p16-CreERT2-Tomato mice. (n = 7 for each group). d, Scheme of MB49 culture using conditioned medium collected from Tomato−or whole iCAFs. e, The bright-field image of Tomato− (p16l) and Tomato+ (p16h-sn) iCAFs. The red color represented the fluorescent signals of Tomato. Scale bar, 100 μm. Paired t-test was performed for c. Data in c are presented as mean values ± SEM. All the t-test were two-side.
Extended Data Fig. 8 CXCL12 is expressed by iCAFs in aged bladder cancer patients.
a, UMAP showing the expression levels of indicated genes in fibroblasts from the scRNA-seq data using bladder cancer patient samples. The dashed lines indicates the iCAF or myCAF clusters determined by the marker genes. b, Violin plots showing the expression levels of the indicated genes in iCAFs or myCAFs. c, UMAP visualization of CAFs labeled by aged or young groups. d, Percentages of iCAF and myCAF in the aged or young group. e, UMAP showing the expression levels of CXCL12 genes in fibroblasts from the scRNA-seq data. f, Expression levels and percentages of CXCL12 in the indicated groups of CAFs. Chi-squared test was performed for d.
Extended Data Fig. 9 Genes upregulated in mouse p16h-sn fibroblasts are associated with age in bladder cancer patient TCGA dataset.
a, Overall survival probabilities of bladder cancer patients classified by age. (≦64, n = 159; 65−74, n = 135; ≧75, n = 141). b, Log2FC values of the top seven up-regulated genes in Tomato+ (p16h-sn) fibroblasts compared with Tomato− (p16l) fibroblasts from the scRNA-seq data using an aged male mouse bladder. c, Correlation coefficient matrix among mRNA expressions of gene set from TCGA bladder cancer data. P < 0.001 for all correlation coefficients. d, Expression levels of the indicated genes across different age groups of patients from the TCGA dataset. (≦64, n = 155; 65−74, n = 135; ≧75, n = 138). One-way ANOVA with Tukey’s or Games-Howell’s test was performed for d. For all box plots, notch of box blots indicates median value. Lower and upper bounds correspond to 25th and 75th percentiles, respectively. Cross mark indicates mean value. Whiskers extend to maximum or minimum value, with points outside this range drawn individually.
Extended Data Fig. 10 Association between the genes upregulated in mouse p16h-sn fibroblasts and clinical prognosis in TCGA dataset.
a, Expressions of indicated genes across different clinical predictive factors of the patients from the TCGA dataset (Metastasis: Negative, n = 139; Positive, n = 140. Recurrence: Negative, n = 264; Positive, n = 129. Pathological T stage: ≦pT2, n = 127; pT3, n = 204; pT4, n = 61. Grade. Negative, n = 21; Positive, n = 402.). b, Overall survival probabilities of the bladder cancer patients, which grouped evenly by indicated gene expression levels. (SMOC2, GUCY1A3, CXCL12, MATN2, CRISPLD2, and GAS1: Low, n = 213; High, n = 212. LUM: Low: n = 212; High, n = 213.). Unpaired t-test and one-way ANOVA with Tukey’s or Games-Howell test was performed for a. Log-rank test was performed for b. For all box plots, notch of box blots indicates median value. Lower and upper bounds correspond to 25th and 75th percentiles, respectively. Cross mark indicates mean value. Whiskers extend to maximum or minimum value, with points outside this range drawn individually.
Supplementary information
Supplementary Information
Supplementary Figs. 1–3.
Supplementary Table 1
Sequences of oligonucleotides for Cxcr4 and Cxcr7 knockdown using shRNA.
Supplementary Table 2
Sex and age of the participants described in Fig. 6f.
Supplementary Table 3
Raw lifespan data.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data and unprocessed western blots.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data and unprocessed western blots.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Meguro, S., Johmura, Y., Wang, TW. et al. Preexisting senescent fibroblasts in the aged bladder create a tumor-permissive niche through CXCL12 secretion. Nat Aging 4, 1582–1597 (2024). https://doi.org/10.1038/s43587-024-00704-1
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s43587-024-00704-1
This article is cited by
-
The relationship between CXC chemokines and cellular senescence: from mechanisms to therapy
Biogerontology (2026)
-
Unleashing endogenous regeneration by senolytics
Journal of Translational Medicine (2025)
-
Senescence-specific molecular subtypes stratify the hallmarks of the tumor microenvironment and guide precision medicine in bladder cancer
BMC Cancer (2025)
-
Extracellular vesicles from ovarian cancer cells induce senescent lipid-laden macrophages to facilitate omental metastasis
Journal of Nanobiotechnology (2025)
-
Stromal senescence contributes to age-related increases in cancer
Nature Reviews Cancer (2025)
