
Abstract
Embryoless rice is valuable for studying early seed development and has great breeding potential, however, related research remains limited. Here, we show that mutations in OsBZR4, encoding brassinazole-resistance 4, lead to 60–100% embryoless seeds across different cultivars. OsBZR4 is specifically expressed at the scutellum-endosperm interface and regulates auxin levels and distribution during early embryo development. OsBZR4 represses the expression of YUC4 and PIN5b. Exogenous auxin and overexpression of YUC4 enhance the embryoless ratio, whereas the auxin transport inhibitor N-1-naphthylphthalamic acid (NPA) decrease it. Higher temperatures increase the embryoless ratio in bzr4 mutant lines. The expression of OsPIL13 is induced by high temperature, and its overexpression enhances the embryoless ratio in both wild type and bzr4 plants. Introducing bzr4 mutant allele into some elite cultivars can improve milled rice yield and storability. Collectively, manipulating OsBZR4 can lead to the development of thermosensitive embryoless rice varieties with increased energy reserves and improved storability, potentially enabling novel rice production technologies.
Similar content being viewed by others

Introduction
Rice is a pivotal staple crop, and fluctuations in its yield and reserves have significant implications for global food security (https://doi.org/10.4060/cc3020en). Milled rice yield and storage tolerance represent two critical quality traits in post-harvest grain processing. Over the past few decades, numerous critical genes controlling yield and quality have been characterized and utilized for breeding1,2. However, limited genes have been reported on strategies to improve milled rice yield and storability3.
Rice seed development is initiated by double fertilization, wherein a haploid egg cell and two haploid central cells are fertilized by sperm cells, resulting in the formation of a diploid zygotic embryo and a triploid endosperm. The mature embryo, comprising the coleoptile, radicle, epiblast, and scutellum, is essential for seed germination and subsequent plant propagation4. The endosperm serves as the major nutrient reservoir, storing starch and proteins to support embryogenesis and germination, while also constituting a key dietary source for humans4,5. The development of the embryo and endosperm is tightly coordinated to form a viable seed, with their sizes being mutually constrained within the limited space of the glume6. Several types of mutants associated with the ratio of embryo to endosperm size have been reported in rice. Mutants endospermless (enl)7 and giant embryo (ge)8,9,10 have enlarged embryos but either lack or show significantly reduced endosperm. Conversely, embryoless (eml) and reduced embryo (re) mutants display absent or smaller embryos accompanied by an expanded endosperm11,12,13,14. Another distinct phenotype is observed in the globular embryo (gle) mutant, where embryonic growth persists but fails to undergo organ differentiation, resulting in mature seeds retaining an undifferentiated globular embryo structure15,16. To date, Giant Embryo (GE) and Globular Embryo (GLE4/OsMAPK6) genes have been cloned and functionally investigated8,9,10,15,16. However, the genetic determinants underlying embryoless seed formation have remained unidentified since its discovery three decades ago12,13, and the molecular mechanisms coordinating embryo and endosperm size regulation are elusive.
The plant hormones brassinosteroids (BRs) cooperate with other phytohormones to regulate grain size by modulating cell proliferation and expansion during the grain development process17,18. Brassinazole-resistant (BZR) members are central transcription factors and play critical roles in the BR signaling pathway by directly regulating the expression of many BR-responsive genes19. Rice BZR family has four members, but only OsBZR1 has been extensively studied17,20,21. The functions of the other BZR members and whether BR regulates early embryo development remain less studied.
Auxin plays critical roles in regulating plant embryo development, and the dynamic and polar transport of auxin within the globular embryo is essential for the initiation of cotyledon primordia22,23. The formation of embryonic and postembryonic organs is typically impaired in Arabidopsis auxin biosynthesis or signaling mutants24. Similarly, rice mutants with impaired auxin biosynthesis, metabolism, or response also exhibit aberrant embryo or endosperm development, which ultimately reduces grain yield25,26,27,28,29,30. Conversely, excess auxin can also inhibit normal embryo development. For instance, exogenous auxin treatment results in embryonic abnormalities24, and the ge mutant exhibits reduced auxin levels10. These findings collectively imply that auxin orchestrates early seed development through stage-specific and tissue-dependent mechanisms. Nevertheless, the precise role of auxin in regulating rice embryogenesis and its underlying molecular mechanism remains poorly understood.
The storage stability of rice is crucial in ensuring a nation’s food security and maintaining strategic grain reserves. According to the Food and Agriculture Organization (FAO) of the United Nations, global rice stocks reached 196.0 million metric tons at the end of the 2023 marketing year, with China accounting for 100.1 million metric tons of this total (FAO, 2023; https://doi.org/10.4060/cc3020en). In China, annual rice quality deterioration during storage results in losses exceeding 3% of the total storage capacity31,32. Despite these significant losses, genetic approaches to enhance rice storability have remained largely unexplored, and available genetic resources for this purpose are limited32.
In this study, we demonstrate that mutation of OsBZR4 results in 60–100% of seeds being embryoless, with endosperm tissue filling the embryonic cavity. We reveal that OsBZR4 regulates early embryogenesis and controls auxin levels and distribution. Importantly, introducing the bzr4 mutant allele into elite rice cultivars can simultaneously improve milled rice yield and storage stability.
Results
Characterization of embryoless mutants
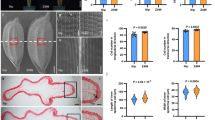
Our previous studies demonstrated that OsWRKY53 and OsMED25 regulate rice architecture and grain shape by interacting with OsBZR120,21. To systematically investigate the functions of OsBZR family genes, we generated mutants for all four members (OsBZR1–4). We occasionally found that the germination rate of bzr4 (LOC_Os02g13900) is lower than 20%. Surprisingly, observations of dehusked seeds revealed that ~80% of bzr4 seeds exhibited either an absent or a significantly reduced embryo (Fig. 1a and Supplementary Fig. 1). By conducting hand sectioning followed by aniline blue staining, we observed that a significant proportion of bzr4 seeds have either an absent or markedly reduced embryo, with the original embryonic position occupied by the endosperm (Fig. 1b). The identical morphology was observed in mature seeds via CT scanning (Fig. 1c). Furthermore, embryoless seeds can be efficiently identified under transmitted light illumination due to the absence of the characteristic embryo shadow observed in normal seeds (Supplementary Fig. 2). To verify this result, a series of allelic mutants of bzr4 were generated across multiple cultivar backgrounds, including the japonica varieties Songjing 2 (SJ2), Longjing 11 (LJ11), Hongyuandao (HYD), Suijing 18 (SJ18), and indica variety 9311 (Supplementary Fig. 1a–f). Statistical analysis showed that the embryoless ratio among various bzr4 mutants is around 60–70%, while the proportion of small or normal embryos is ~30–40% (Fig. 1f and Supplementary Fig. 1g). To further confirm this, the OsBZR4 genomic DNA was transformed into bzr4-SJ2 mutant, and the embryoless ratio was restored to that of the WT (Supplementary Fig. 3). Together, these results suggest that OsBZR4 is essential for embryo development.

a–e The embryonic morphology in SJ2 (top), small embryo (middle) and no embryo in bzr4-SJ2-2 (bottom). a Mature seed, Scale bars, 500 μm. b Hand sections and aniline blue staining of developing seeds at 20 days after pollination (DAP). Scale bars, 200 μm. c CT scan images of mature seeds. Scale bars, 1 mm. d Laser scanning confocal microscopy (LSCM) of developing embryo at 7 DAP. Scale bars, 200 μm. e Paraffin section of developing embryo at 7 DAP. Scale bars, 100 μm. f The frequency of different types of embryos in SJ2 and bzr4-SJ2 mutants. N = 5 biological replicates, around 100 seeds per replicate. Nor E normal embryo, SE small embryo, and NE no embryo. g The relative expression of marker genes related to rice embryo development in 7 DAP caryopses. n = 3 biological replicates. Data in (f, g) are presented as mean ± SD. Two-tailed unpaired Student’s t test was used for statistical analysis. Source data are provided as a Source Data file.
There are four homologs of OsBZR in the rice genome17. To determine whether the other OsBZR members participate in embryonic development, we analyzed single, double, and triple mutants of OsBZR1–4. We found that only mutants carrying disruptions in bzr4 exhibited embryoless seeds, while mutations in OsBZR1–3 did not enhance this phenotype (Supplementary Fig. 4, Supplementary Table 1). These findings demonstrate that OsBZR4 specifically regulates embryo development.
To determine when the defect occurs in bzr4 mutants, we examined the morphology of ovaries and anthers, as well as pollen viability. No significant differences were observed between bzr4-SJ2-2 and SJ2 (Supplementary Fig. 5). In addition, reciprocal crosses between bzr4-SJ2-2 and SJ2 yielded normal seeds in the F1 generation, suggesting that the fertilization process in bzr4-SJ2-2 is unaffected (Supplementary Table 2). We subsequently analyzed the developing embryos using a laser scanning confocal microscope (LSCM). At 3 days after pollination (DAP), the zygote in bzr4-SJ2-2 and SJ2 formed a normal globular embryo (Supplementary Fig. 6a–d). Thereafter, the embryos in SJ2 continued to grow and differentiate, exhibiting progressively distinct development of the coleoptile, radicle, epiblast, and scutellum (Fig. 1d and Supplementary Fig. 6e, i, m, q). In contrast, in bzr4-SJ2-2 mutants, embryogenesis displays three distinct phenotypes. The majority (∼70%) of embryos arrest at the globular stage (Fig. 1d, f; Supplementary Fig. 6h, l, p, t). A subset (∼10%) show post-globular developmental arrest, forming smaller embryos with either complete or incomplete organogenesis (Fig. 1d; Supplementary Fig. 6g, k, o, s). Only a minor proportion (∼20%) maintain normal developmental potential, progressing to form fully differentiated embryos (Fig. 1d; Supplementary Fig. 6f, j, n, r). Similar results were observed and verified in the paraffin section analysis (Fig. 1e; Supplementary Fig. 7). Furthermore, the expression of several embryo development marker genes, including ROC1, ROC3, OSH1, PNH1, BBM, GE, and MPK68,16,33,34,35,36,37, was examined, show significantly reduced in bzr4-SJ2-2 relative to WT (Fig. 1g). Collectively, these results suggest that the developmental defect in bzr4 embryos occurred as early as the globular stage.
OsBZR4 specifically expressed in scutellum–endosperm interface
We investigated the spatial and temporal expression patterns of OsBZR4 by examining OsBZR4p::GUS transgenic plants. The GUS staining was observed in various tissues, including roots, stems, leaves, anthers, and seeds (Supplementary Fig. 8a–i). In the developing seeds, the GUS staining intensity was lower and mainly dispersed in the seed coat at 0–3 DAP (Supplementary Fig. 8g–i), while it became specifically concentrated in the embryo region of seeds after 5 DAP (Fig. 2a). After 7 DAP, GUS staining was gradually restricted to the scutellum-endosperm interface, which serves as the interface between embryo and endosperm (Fig. 2b, c). This expression pattern closely resembles that of GE8,9,10. Additionally, both GUS staining and RT-qPCR revealed a significant increase in OsBZR4 expression during embryo development from 3 to 10 DAP (Supplementary Fig. 8l).

a–c GUS staining of OsBZR4p::GUS caryopses at 5, 7, and 9 DAP. Scale bars, 500 μm. Experiments were repeated independently at least in triplicate with similar results. d Uniform manifold approximation and projection (UMAP) plot of unbiased reclustering of the 10,442 cells from rice caryopses of 7 DAP. Dots denote individual cells, and colors denote different cell clusters (0–14). e UMAP visualization of expression profiles of OsBZR4, GE and SWEET14. Dots denote individual cells, and color indicates the expression level in a cell. f–g RNA in situ hybridization detecting GE (f) and SWEET14 (g) expression in 7-DAP caryopses of SJ2. Scale bars, 100 μm in (f) and 200 μm in (g).
Through RNA in situ hybridization, we were unable to detect a specific signal or determine the spatiotemporal expression pattern of OsBZR4 in seeds. To overcome this difficulty, we performed single-nucleus RNA sequencing (snRNA-seq) on 7 DAP caryopses. Following quality control, we acquired a total of 10,442 nuclei, comprising 3855 and 6587 nuclei from the two biological replicates, respectively. Louvain clustering of the combined data from two biological replicates identified 15 distinct clusters (Fig. 2d). The uniform manifold approximation and projection (UMAP) algorithm was subsequently employed to visualize and explore the datasets following linear dimensionality reduction. OsBZR4 was specifically expressed in cluster 13 (Fig. 2e and Supplementary Data 1 and 2). The gene ontology (GO) enrichment analysis of marker genes in cluster 13 showed that many of them were related to transport (Supplementary Fig. 9 and Supplementary Data 3). These genes included OsSWEET14, OsSWEET15, OsPHO1;2, and OsARF1, whose homologs in maize had been reported to be expressed in the endosperm adjacent to the scutellum (EAS) region (Fig. 2e and Supplementary Data 1 and 2)38,39. GE, which is specifically expressed in the embryo scutellum and its adjacent endosperm8,9,10, was also found to be in the same cluster (Fig. 2e and Supplementary Data 1 and 2). To further validate the spatial localization of cluster 13, we confirmed that OsSWEET14 and GE exhibited specific expression at the scutellum-endosperm interface of 7-DAP caryopses by in situ hybridization (Fig. 2f, g, and Supplementary Fig. 8j, k), which is consistent with the GUS staining of OsBZR4 (Fig. 2b, c) and previous studies8,9. Together, these results suggest that OsBZR4 is specifically expressed in the scutellum-endosperm interface, consistent with its function in regulating the embryo and endosperm ratio. Double mutant ge1 bzr4 exhibits an embryoless phenotype, similar to that observed in bzr4 (Supplementary Fig. 10, Supplementary Table 3), which suggests that OsBZR4 may function at an earlier stage than GE in controlling seed development.
OsBZR4 regulates auxin content and distribution in early developing caryopses
Since OsBZR4 is a homolog of BZR1, a key transcription factor in the BR pathway, we first investigated whether OsBZR4 mediates BR signaling to regulate embryonic development. We treated the spikelets of SJ2 and bzr4-SJ2-2 with 1 μM and 10 μM Brassinolide (BL), respectively, from 1 DAP for a week, and analyzed embryo development at seed maturity. However, neither SJ2 nor bzr4-SJ2-2 exhibited significant phenotypic changes upon BL treatment compared to the control (Supplementary Fig. 11). Additionally, we conducted in vitro culture experiments using detached SJ2 caryopses at 1 DAP in the growth medium supplemented with Brassinazole (BRZ). After 20 days of growth, the embryos developed normally, with no observable differences from the control samples (Supplementary Fig. 12). These results indicate that BR may not be involved in regulating embryonic developmental processes.
Auxin plays critical roles in regulating early embryo development22,40,41. To investigate whether the OsBZR4-regulated embryoless phenotype is related to auxin, we treated the SJ2 spikelets at 1 DAP with 100 μM 2,4-D for 1 week. Upon completion of seed filling and maturation, the developmental phenotypes of the embryos were examined. Compared to the control, embryos from seeds treated with 2,4-D exhibited a significantly reduced size (Supplementary Fig. 13). Moreover, we performed an in vitro culture assay on detached caryopses of SJ2. Following a 20-day culture period on medium containing 50 μM 2,4-D, we observed that ~45% of the caryopses developed globular embryo-like structures resembling those in bzr4 mutants, while the remaining 55% formed smaller embryos (Fig. 3a, b). In addition, the severity of the embryonic defects increased with higher concentrations of 2,4-D (Fig. 3b). In contrast, control samples showed no such developmental defects (Fig. 3a, b), suggesting that auxin specifically inhibits early embryo development in rice. To verify this hypothesis, bzr4-SJ2-2 spikelets at 1 DAP were treated with 2,4-D for a week, and the results showed that 2,4-D treatment significantly increased the embryoless ratio of bzr4-SJ2-2 from 70 to 90% (Fig. 3c).

a Embryo phenotypes of detached caryopses cultured on medium containing 2,4-D for 20 days, hormone-free medium was used as control. Arrows indicate the embryo. Scale bars, 200 μm. b The frequency of embryo types in detached caryopses cultured in vitro. n = 3 biological replicates, 10 detached caryopses per replicate. c The frequency of embryo types in bzr4-SJ2-2 treated by 100 μM 2,4-D and 100 μM NPA. n = 4 biological replicates, around 100 seeds per replicate. d Free IAA levels in developing caryopses of SJ2 and bzr4-SJ2-2 at 1, 3, and 7 DAP. n = 3 biological replicates. e GUS staining of caryopses from DR5::GUS-ZH11 and DR5::GUS-bzr4-ZH11 at 7 DAP. The upper and lower images represent vertical and horizontal cross-sections of the corresponding seeds, respectively. Red arrows indicate the embryo, and yellow arrows indicate vascular tissue in the seed coat. Scale bars, 500 and 200 μm, respectively. f The DR5::GUS-bzr4-ZH11 spikelets were treated with 100 μM NPA beginning at 1 DAP for 7 days. Control plants received equivalent volumes of the solvent solution without NPA. Scale bars, 500 μm. Experiments were repeated independently at least in triplicate with similar results in (e, f). Data in (b–d) are presented as mean ± SD. Two-tailed unpaired Student’s t test was used for statistical analysis. Nor E normal embryo, SE small embryo, and NE no embryo in (b, c). Source data are provided as a Source Data file.
To further confirm that auxin might be involved in OsBZR4-regulated embryo formation, we examined the auxin levels in developing caryopses. Our results demonstrated that the auxin levels were very low and comparable in 1 DAP caryopses between SJ2 and bzr4-SJ2-2 (Fig. 3d). Subsequently, auxin levels increased dramatically during caryopsis development. Specifically, at 3 and 7 DAP, the auxin levels in bzr4-SJ2-2 were ~3- and 1.2-fold higher than those in SJ2, respectively (Fig. 3d). These findings suggest that OsBZR4 negatively regulates auxin levels. In addition, GUS staining was detected in caryopses of DR5::GUS lines at 1–7 DAP in both ZH11 and bzr4-ZH11 backgrounds. At 1 DAP, GUS staining in ZH11 caryopses was distributed throughout the seed coat, vascular bundles, and caryopsis-glume junction. In bzr4-ZH11, however, the signal was largely confined to the vascular bundles and caryopsis-glume junction, with minimal expression in the seed coat (Supplementary Fig. 14a, b). By 3 and 5 DAP, GUS staining in both genotypes was largely confined to the vascular bundles and the caryopsis-glume junction (Supplementary Fig. 14c-f). Analyzing auxin distribution within caryopses at 1–5 DAP was technically challenging due to their small size and the absence of a fully formed endosperm (the endosperm chamber remained fluid-filled). At 7 DAP, cross sections revealed that GUS staining in ZH11 was primarily localized to the vascular tissue and the embryo (Fig. 3e). In contrast, bzr4-ZH11 showed intense GUS staining on the ventral side of the endosperm, a pattern absent in ZH11 (Fig. 3e). These results suggest that in early developing caryopses, OsBZR4 not only regulates auxin levels but also modulates its spatial distribution as early as 1 DAP. To verify this, we treated DR5::GUS-bzr4-ZH11 plants with the polar auxin transport inhibitor N-1-naphthylphthalamic acid (NPA). Intriguingly, NPA treatment drastically reduced the auxin signal in the ventral endosperm region, while GUS activity remained detectable in the dorsal vascular bundle and the embryo (Fig. 3f, Supplementary Fig. 15). Given that the embryoless phenotype of bzr4-SJ2-2 might be attributed to both elevated auxin levels and altered auxin distribution, we suspected that inhibiting auxin transport could rescue the defect. Consistent with this hypothesis, exogenous NPA significantly reduced the embryoless ratio of bzr4-SJ2-2 from 70 to 30% (Fig. 3c). Together, these results suggest that OsBZR4 regulates both auxin levels and distribution during caryopsis development, and that auxin signaling influences embryo development.
OsBZR4 suppresses the expression of YUC4 and PIN5b
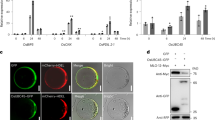
To investigate whether OsBZR4 regulates auxin synthesis, we examined the expression of auxin biosynthesis genes in 3-DAP caryopses. Among the 7 rice YUC genes42, we found that the expression levels of YUC4 and YUC11 were significantly increased in bzr4-SJ2-2 compared to SJ2, suggesting that OsBZR4 suppresses the expression of YUC4 and YUC11 (Fig. 4a). YUC4 and YUC11 promoter regions contains like BRRE-box and E-box motifs (Fig. 4c, Supplementary Fig. 16), which have been identified as recognition elements of BZR1/BES1 in Arabidopsis43. Electrophoretic mobility shift assays (EMSA) demonstrated that the OsBZR4 protein can specifically bind to the BRRE-box and E-box in the promoter regions of YUC4 and YUC11, respectively (Fig. 4b, Supplementary Fig. 16a). The ChIP assay conducted with Flag-OsBZR4 OE plants demonstrated that OsBZR4 binds to the promoters of YUC4 and YUC11 in vivo (Fig. 4c; Supplementary Figs. 16b and 17). Since YUC4 is mainly expressed at the interface between the embryo and endosperm (Supplementary Fig. 18), a pattern very similar to that of OsBZR4 (Fig. 2b, c), while YUC11 mainly functions in endosperm25, we then focused on YUC4 in subsequent studies. To investigate the role of YUC4 in early embryo development, we generated the yuc4 and yuc4 yuc5 mutants in both SJ2 and bzr4-SJ2-2 backgrounds (Supplementary Table 4). The yuc4 and yuc4 yuc5 mutants develop normal embryos, whereas the bzr4 yuc4 and bzr4 yuc4 yuc5 mutants exhibit similar embryoless ratios and smaller embryos, as observed in the bzr4-SJ2-2 background (Supplementary Fig. 19). We suspected that the YUC family may possess redundant functions in regulating auxin biosynthesis. To test this hypothesis, we introduced OsBZR4p::YUC4 into SJ2 and found that the proportions of embryoless and small embryos in OsBZR4p::YUC4 seeds were ~1% and 20%, respectively, significantly higher than those in SJ2 seeds (Fig. 4d). In addition, we introduced OsBZR4p::YUC4 into the bzr4-SJ2-2 mutant and found that the embryoless ratio increased to 90%, compared to 70% in the bzr4-SJ2-2 under identical growing conditions (Fig. 4e). Collectively, these results demonstrate that OsBZR4 directly binds to and suppresses the YUC4 expression, suggesting that the function of OsBZR4 is, at least partially, dependent on YUC4.

a Relative expression of YUC genes in 3-DAP caryopses. n = 3 biological replicates. b EMSA confirming that the His-OsBZR4 protein binds to the YUC4 promoter region containing like BRRE-box. Unlabeled probes were used as competitors. Experiments were repeated independently at least in triplicate with similar results. c ChIP assays showing that OsBZR4 binds to the promoters of YUC4 in vivo. Immunoprecipitation was performed with anti-Flag antibody. Immunoprecipitated chromatin was analyzed by qPCR. n = 3 biological replicates. d–e The frequency of OsBZR4p:YUC4 embryo types under SJ2 (d) and bzr4-SJ2-2 (e) background. n = 5 biological replicates, around 100 seeds per replicate. f Relative expression of PIN genes in 3-DAP caryopses. n = 3 biological replicates. g ChIP assays showing that OsBZR4 binds to the promoters of PIN5b in vivo. Immunoprecipitation was performed with anti-Flag antibody. Immunoprecipitated chromatin was analyzed by qPCR. n = 3 biological replicates. Data in (a, c–g) are presented as mean ± SD. Two-tailed unpaired Student’s t test was used for statistical analysis. Nor E normal embryo, SE small embryo, and NE no embryo in (d, e). Source data are provided as a Source Data file.
Similarly, to investigate how OsBZR4 regulates auxin distribution, we examined the expression levels of 11 PIN family genes, which encode auxin efflux carriers44. Our analysis revealed that the expression levels of PIN5b and PIN5c were significantly higher in bzr4-SJ2-2 compared to SJ2, while no significant differences were observed in other PIN genes (Fig. 4f). We identified two E-box motifs in the PIN5b promoter region. ChIP-qPCR assays revealed substantial enrichment of OsBZR4 on these E-box-containing fragments, demonstrating direct binding of OsBZR4 to the PIN5b promoter in vivo (Fig. 4g). Furthermore, we examined PIN5b-overexpressing plants45. No embryo developmental defects were observed (Supplementary Fig. 20). Collectively, our findings suggest that OsBZR4 regulates auxin biosynthesis and transport by modulating the expression of YUC4 and PIN5b.
The embryoless phenotype in bzr4 is temperature dependent
Around 30 years ago, an embryoless mutant eml1 was described, but the causal gene remained unknown13,14. It was also found that the embryoless phenotype exhibited temperature sensitivity12. To examine whether the embryoless phenotype of bzr4 in this study is temperature-dependent, three independent bzr4-SJ2 lines at the heading stage were transferred to three chambers set at different temperatures for further growth until seed maturity. The results demonstrated that the average embryoless ratios of bzr4-SJ2 at lower (22 °C), normal (27 °C), and higher (32 °C) temperatures were ~42%, 70%, and over 90%, respectively (Fig. 5a, Supplementary Fig. 21). In addition, the average embryoless ratios of bzr4-SJ2-2 seeds in Sanya, Hainan Province (18°30’N) exceeded 95%, which is substantially higher than the ~70% observed in Harbin, Heilongjiang Province (45°30’N) (Fig. 5b). To determine the temperature-sensitive stage of bzr4, bzr4-SJ2-2 plants were exposed to a temperature shift from 32 °C to 22 °C for seven consecutive days, either before or after pollination. The average frequency of embryoless seeds was ~40% when the temperature shift occurred prior to pollination, but sharply increased to nearly 90% when the shift was imposed at or after 1 DAP (Supplementary Fig. 22). These results demonstrate that the occurrence of embryoless seeds in bzr4-SJ2-2 is highly temperature-dependent, with the most critical sensitivity window coinciding with 1 DAP.

a The frequency of embryo types in bzr4-SJ2-2 seeds produced at different temperatures: 22 °C (LT), 27 °C (NT), and 32 °C (HT). n = 5 biological replicates, around 100 seeds per replicate. b The frequency of embryo types in bzr4-SJ2-2 seeds produced from Harbin and Sanya. SJ2-H and bzr4-H represent plants grown in Harbin, whereas SJ2-S and bzr4-S were cultivated in Sanya. n = 5 biological replicates, around 100 seeds per replicate. c Free IAA levels in bzr4-SJ2-2 caryopses at 3 DAP under LT and HT. n = 3 biological replicates. d The relative expression level of YUC4 in bzr4-SJ2-2 caryopses at 3 DAP under LT and HT. n = 3 biological replicates. e–f The frequency of embryo types in OsPIL13 OE seeds under SJ2 (e) and bzr4-SJ2-2 (f) background, respectively. n = 5 biological replicates, around 100 seeds per replicate. g The time course expression of OsPIL13 under NT and HT. n = 3 biological replicates. h Transient transcriptional activity assay showing OsPIL13 activates YUC4 expression. Relative REN (Renilla luciferase) activity was used as an internal control, and the relative LUC (Firefly luciferase)/REN ratios are shown. n = 3 biological replicates. i The relative expression level of YUC4 in 3-DAP caryopses of SJ2 and OsPIL13 OE lines. n = 3 biological replicates. Data in (a–i) are presented as mean ± SD. Two-tailed unpaired Student’s t test was used for statistical analysis. Nor E normal embryo, SE small embryo, and NE no embryo in (a, b, e, f). Source data are provided as a Source Data file.
As we have shown that OsBZR4 may regulate auxin levels and distribution (Fig. 3), and the ratio of embryoless seeds in bzr4-SJ2-2 increases under higher temperature conditions (Fig. 5a, b). Previous studies have reported that elevated temperatures can induce auxin biosynthesis46. Consequently, we measured the auxin levels of bzr4-SJ2-2 under 22 °C and 32 °C temperature treatments. We found that high temperature significantly increased the auxin levels in bzr4-SJ2-2 (Fig. 5c). Consistently, increased temperature also promoted the expression of YUC4 in bzr4-SJ2-2 (Fig. 5d). Collectively, these results imply that the higher temperature-increased embryoless ratio in bzr4 may be attributed to elevated auxin levels.
Arabidopsis phytochrome interacting factor 4 (PIF4) is a well-known key regulator of thermomorphogenesis. Elevated temperatures induce the expression of PIF4, which directly activates the expression of auxin biosynthetic genes to promote hypocotyl growth46,47,48. To investigate whether rice homologs of PIF4 are involved in the temperature-dependent embryoless ratio in bzr4, we individually overexpressed five members of the PIF-Like (PIL) family, PIL11 to PIL15 (Supplementary Fig. 23a, b)49. Phenotypic analysis revealed that PIL13 OE seeds exhibited 2% embryoless and 5% small embryo compared to WT and other PILs OE plants (Fig. 5e and Supplementary Fig. 23c). Therefore, PIL13 was further overexpressed in the bzr4-SJ2-2 mutant background (Supplementary Fig. 24). We found that the embryoless ratio in PIL13 OE bzr4-SJ2-2 is 90%, which is significantly higher than 70% in bzr4-SJ2-2 observed (Fig. 5f). Moreover, we showed that elevated temperatures markedly enhanced the expression of PIL13 (Fig. 5g), which in turn dramatically activated the expression of YUC4 (Fig. 5h, i). However, we observed that the mutants of pil13 and pil11 pil13 pil14 do not result in embryoless seeds and fail to enhance the embryoless ratio of bzr4-SJ2-2 at either higher or lower temperatures (Supplementary Fig. 25, Supplementary Table 5). This phenomenon may be attributed to functional redundancy within the PIL family, and need further investigation.
Manipulating bzr4 has the potential to enhance the milled rice yield
To evaluate the application value of bzr4, we assessed the yield and quality characteristics of bzr4-SJ2-2 and demonstrated that the yield traits of bzr4-SJ2-2, including tiller number, grain number per panicle, 1000-grain weight, and yield per plant, are comparable to those of SJ2 (Supplementary Fig. 26a). Additionally, compared to the SJ2, the bzr4-SJ2-2 brown rice showed a 33.03% decrease in lipid content, a 10.22% increase in amylose content, and comparable protein content (Supplementary Fig. 26b). These results are consistent with the embryoless phenotype of bzr4-SJ2-2, as the lipids is abundant in embryos and starch is primarily located in the endosperm50,51.
In the process of rice milling, the embryos are usually shed passively. Considering that the endosperm of bzr4 occupies the position of the embryo, we hypothesized that bzr4 would yield a higher percentage of milled rice compared to the WT. Contrary to our expectations, bzr4-SJ2-2 demonstrated a slightly lower milled rice yield than SJ2, which might be attributed to a significantly higher chalkiness in bzr4-SJ2-2 compared to SJ2 (Supplementary Fig. 26c, Supplementary Table 6). Given that chalkiness and appearance quality are closely associated with grain shape and amylose content52, we knocked out OsBZR4 in Zhongjia 10 (ZJ10) and Zhongjia 11 (ZJ11), two cultivars characterized by lower amylose content and slender grains, which exhibit superior appearance and taste quality (Supplementary Fig. 27a–c). The bzr4-ZJ10 and bzr4-ZJ11 mutants were grown in Sanya, Hainan Province, where the embryoless ratio exceeded 90% (Supplementary Fig. 27d). The yield-related traits of bzr4-ZJ10 and bzr4-ZJ11 are comparable to those of ZJ10 and ZJ11 (Supplementary Fig. 28a). Fortunately, bzr4-ZJ10 and bzr4-ZJ11 show similar chalky grains and chalkiness degree to their respective WT, although the starch content in bzr4 lines is increased compared to their respective wild types. (Fig. 6a–c, and Supplementary Fig. 28b, c, Supplementary Table 7). Most importantly, bzr4-ZJ10 and bzr4-ZJ11 demonstrate a 2.52 and 2.57% increase in milled rice yield compared to their corresponding WT levels (Fig. 6d and Supplementary Table 7).

a–b The milled rice phenotypes of bzr4 mutants in ZJ10 (a) and ZJ11 (b). The upper is the wild type, the bottom is the corresponding bzr4 mutant. Scale bars, 1 mm. c The appearance quality of ZJ10 and bzr4-ZH10. Scale bars, 5 mm. d Milled rice yield of bzr4 mutants in ZJ10 and ZJ11. n = 3 biological replicates. e Lipoxygenase enzyme (LOX) activity of brown rice before or after 30-day accelerated aging treatment. 1U = Δ234/min/mg protein. n = 3 biological replicates. f The relative expression of LOX3 in brown rice before or after the 30-day accelerated aging treatment. n = 3 biological replicates. g–h The fatty acid values of brown rice (g) and unhusked rice (h) during accelerated aging treatment. The values were expressed by the mass (mg) of KOH required to neutralize the free fatty acids contained in 100 g of rice flour. n = 3 biological replicates. Data in (d–h) are presented as mean ± SD. Two-tailed unpaired Student’s t test was used for statistical analysis. Source data are provided as a Source Data file.
The bzr4 significantly enhanced the storage capability of rice
Lipids in rice seeds are mainly stored in the embryo, where oxidative rancidity causes seed aging through lipid hydrolysis and oxygenation, generating volatile carbonyl compounds that are responsible for rancid odors53. We hypothesized that bzr4 may possess enhanced anti-aging properties due to the reduced lipid content (Supplementary Figs. 26b and 28b). To this end, brown rice and unhusked rice were subjected to accelerated aging treatment in an incubator maintained at 42 °C and 85% relative humidity for 30 and 60 days, respectively. Lipoxygenase (LOX) catalyzes the initial step of lipid peroxidation, leading to the formation of hydroperoxides54. The absence of LOX in rice can reduce oxidative spoilage55,56,57. Consistent with this finding, bzr4-SJ2-2 showed a 19.64% and 31.76% decrease in LOX enzyme activity compared to SJ2 before and after the aging treatment (Fig. 6e). Additionally, the expression level of the embryo-specific LOX3 was significantly decreased in bzr4-SJ2-2 relative to SJ2 (Fig. 6f). Lipid oxidation during grain storage generates free fatty acids, consequently increasing acidity levels. Our findings reveal that the fatty acid value increased with the storage duration (Fig. 6g, h). However, compared to SJ2, the rate of increase in fatty acid value in bzr4-SJ2-2 is significantly slower (Fig. 6g, h). Specifically, after 30 days of storage, the fatty acid value in bzr4-SJ2-2 brown rice was 34.34 mg/100 g, which is comparable to the value of SJ2 after 15 days storage (37.32 mg/100 g) (Fig. 6g). Similarly, the fatty acid value in the unhusked bzr4-SJ2-2 after 60 days of storage was 28.87 mg/100 g, closely matching the value of SJ2 after 30 days of storage (28.12 mg/100 g) (Fig. 6h). Consistent results were observed in accelerated aging tests using bzr4 brown rice from both ZJ10 and ZJ11 genetic backgrounds (Supplementary Fig. 29). These results suggest that the bzr4 seeds can effectively extend the storage time of rice by ~2-fold.
During rice storage, off-flavor development is primarily caused by the accumulation of volatile aldehydes, which typically increase with storage duration58. We found that the volatile aldehydes exhibited a significant increase after storage. However, the increase in bzr4-SJ2-2 was smaller than that in SJ2, indicating that the flavor quality decline was lower in bzr4-SJ2-2 rice compared to SJ2 (Supplementary Fig. 30). Collectively, these results suggest that bzr4 possesses the potential to enhance rice storability.
Discussion
The embryo and endosperm in rice play crucial roles in propagation and calorie storage, respectively. Despite the identification of several mutants affecting either the embryo or endosperm, the causal genes and underlying mechanisms remain largely unknown, impeding our understanding and utilization of these germplasms8,9,11,12,13,15,50,51. In this study, we demonstrated that the embryoless ratios in various bzr4 mutants range from 60% to 100%, enabling an increase in endosperm size while maintaining generational transmission integrity. Moreover, our results indicate that moderate levels and proper distribution of auxin are crucial for normal embryo development. In WT plants, OsBZR4 maintains auxin homeostasis by suppressing both auxin biosynthesis and transport (Fig. 7, top). However, in the bzr4 mutant, the auxin homeostasis is disrupted, causing excessive auxin accumulation and abnormal distribution, which ultimately suppresses embryonic growth and development (Fig. 7, middle). Elevated temperature promotes PIL13 and YUC4 expression and increases auxin accumulation, thereby could exacerbate the embryoless ratio in bzr4 (Fig. 7, bottom). Next, we showcased the immense potential of bzr4 in generating superior varieties with enhanced milled rice yield and storability. Together, our study exemplifies the successful characterization of rare rice germplasm, elucidates the underlying molecular mechanisms, and demonstrates their potential in developing superior rice strains.

Top: In wild-type rice, OsBZR4 maintains auxin homeostasis by suppressing both auxin biosynthesis and transport in seeds. Middle: in bzr4 mutant, the auxin homeostasis is disrupted, causing excessive auxin accumulation and abnormal distribution that ultimately suppresses embryonic growth and development. Bottom: high temperature promotes expression of OsPIL13 and YUC4 and increases auxin accumulation in bzr4 seeds, which could exacerbate the proportion of embryoless seeds. Created in BioRender. Tu, D. (2025) https://BioRender.com/woufypt.
Specifically, the embryoless mutant eml1 was described around 30 years ago, but the causal gene remained unknown13,14. Currently, studies on embryo lethal or defective mutations in rice remain substantially limited, and the relative mutants exhibit distinct phenotypes from bzr4. For example, the embryo of mpk6 mutant (also called gle) is totally lethal, embryo development was arrested at the globular embryo stage, and the embryonic organs did not differentiate normally. Simultaneously, unlike the bzr4 mutant, the endosperm of mpk6 did not occupy the original embryo position; the embryo cavity was visible in mature seeds15,16. In addition, mutants of three rice WUSCHEL HOMEOBOX9 (WOX9) genes produce a proportion of seeds exhibiting abnormal embryo development, including embryos with an inverted apical-basal axis or lacking distinct organ structure. These seeds exhibit delayed germination and distorted outgrowth59. In contrast, unlike the aforementioned mutants, bzr4 showed an embryoless ratio ranging from 60 to 100% across different cultivars, which is very similar to the reported thermosensitive embryoless (eml) mutant described 30 years ago12. We believe that OsBZR4 should be the causal gene of embryoless. In the bzr4 embryoless seed, the endosperm fully occupies the embryo cavity, and the seed is lethal (Fig. 1 and Supplementary Fig. 1). However, a partial bzr4 has normal or small embryos, which can germinate and produce offspring. Therefore, bzr4 shows partial embryonic lethality.
Our findings indicate that OsBZR4 modulates auxin contents and distribution possibly through repressing the expression of auxin biosynthesis genes and auxin transporter genes (Fig. 4). Transgenic plants with OsBZR4pro::YUC4 in the WT produced a small number of embryoless seeds, while in the bzr4 background, there was a significant enhancement the embryoless seed ratio (Fig. 4d, e). However, the embryoless phenotype of bzr4 was not rescued by yuc4 or yuc4 yuc5 (Supplementary Fig. 19), and no abnormal embryo development phenotypes were observed in PIN5b overexpression lines (Supplementary Fig. 20). We speculate that OsBZR4 plays a critical role in maintaining auxin homeostasis in developing seeds by simultaneously regulating auxin biosynthesis and distribution. Mutation in partial auxin synthesis or transport genes alone might be insufficient to completely disrupt auxin homeostasis.
In maize, the interface between the endosperm and the embryo is developmentally dynamic. During early development, the embryo is completely surrounded by cells of the embryo-surrounding region (ESR)60. As embryonic development progresses, the ESR gradually degenerates and eventually disappears61. Subsequently, a new interface is formed, which consists of the scutellum and the adjacent endosperm region known as EAS39. These interfaces play a crucial role in the material transportation between the embryo and endosperm, as well as in embryo growth and development39. Whether a similar region exists in rice is largely unknown. Through single-nucleus RNA sequencing, we found that OsBZR4 is specifically localized at the interface between the embryo and endosperm. Sweet transporters SWEET14 and SWEET1562, GE8,10, ABC transporter OsABCC963, auxin transporter OsAUX364, and a rice grain-filling-controlling PHO1-type Pi transporter OsPHO1;265 were in the same cluster as well (Supplementary Data 1 and 2). This finding suggests that rice may also possess a specific interface region between the embryo and the endosperm, which functions in molecular communication and nutrient transport and is required for the normal development of both the embryo and the endosperm. Further investigation is needed to understand how OsBZR4 and co-localized genes modulate the formation and function of this specific interface.
In recent years, perennial rice varieties capable of producing eight cultivation cycles within 4 years have been achieved, demonstrating various advantages in low-latitude regions66. This system provides significant economic advantages through reduced labor requirements and seed costs66. To integrate the embryoless and perennial traits, we developed the bzr4-YD25 and bzr4-YD26 mutants in the perennial rice varieties Yunda 25 and Yunda 26 (glutinous rice), respectively (Supplementary Fig. 31a). The bzr4-YD25 exhibits an average embryoless ratio of ~70%, similar to that observed in bzr4-SJ2-2 (Supplementary Fig. 31b–d). In contrast, the embryoless ratio of bzr4-YD26 reaches nearly 100% under the same conditions (Supplementary Fig. 31d). The development of bzr4 mutants in perennial rice enables the synergistic combination of desirable traits from both systems. The perennial growth habit bypasses bzr4’s germination defect, simultaneously improving milled rice production and the storability of the ratoon crop.
Although the introduction of bzr4 into several cultivars resulted in an embryoless ratio ranging from 60 to 100%, the embryoless ratio reached 100% in glutinous rice (Supplementary Fig. 31). Chalkiness increased in some, but not all, bzr4-containing cultivars (Supplementary Figs. 26c and 28c, Supplementary Tables 6 and 7). Considering that the embryoless ratio is influenced by factors such as genetic background and environmental conditions, it is crucial to extensively study the effectiveness of bzr4 across diverse contexts. Moreover, the embryoless ratio is 40% at lower temperatures and exceeds 70% under normal conditions (Fig. 5a, Supplementary Fig. 21), introducing innovative concepts for designing temperature-regulated embryoless germplasm, like thermosensitive male sterility. In this system, the ideal bzr4 mutant can generate normal embryos for reproduction and embryoless seeds for rice production in diverse circumstances, potentially leading to the development of a novel rice production technology.
Methods
Plant materials and growth conditions
Rice varieties in this study include japonica cultivars Songjing 2 (SJ2), Zhongjia 10 (ZJ10) and Zhongjia 11 (ZJ11), Longjing 11 (LJ11), Suijing 18 (SJ18), Hongyuandao 1 (HYD1), Zhonghua 11 (ZH11), Yunda 25 (YD25), Yunda26 (YD26), and indica cultivar 9311. The rice plants were cultivated in experimental fields at Harbin (45°30’N, Heilongjiang province) from April to October and at Sanya (18°30’N, Hainan province) from November to March. For plants grown in a controlled greenhouse, the conditions are 14 h of daylight at 27 °C and 10 h of nighttime at 24 °C with a relative humidity of 60%. For the temperature sensitivity experiment, rice plants were grown in a field and then transferred to three growth chambers with different temperatures during the heading stage: 22 °C (lower temperature, LT), 27 °C (normal temperature, NT), and 32 °C (higher temperature, HT).
Vector construction, genome editing, and generation of transgenic plants
For generating bzr4, yuc4, yuc4 yuc5, pil13, pil11 pil13 pli14 mutants, the CRISPR/Cas9 editing method was used for generating mutant67. The sgRNA target sequences of genes are shown in Supplementary Data 4. For generating the ge1 bzr4 double mutant, the bzr4 mutant was generated in ge1 background. For functional complementation of bzr4, a 5.6 kb genomic fragment DNA containing the promoter region and the entire coding sequence of OsBZR4 was subcloned into the binary vector pCAMBIA2300. To obtain DR5::GUS-bzr4, we crossed bzr4-ZH11 with DR5::GUS line under ZH11 background, and homozygous offspring were chosen. To construct the OsBZR4p::GUS and YUC4p::GUS vectors, 2032-bp and 1715-bp fragments upstream from ATG were amplified and cloned into pENTR/D TOPO (Invitrogen), and cloned into the binary vector pHGWFS7 by LR reaction. To construct the over-expression vectors of OsBZR4 and PIL13, their cDNA sequences were amplified and cloned into pCAMBIA1300-Flag and pCAMBIA2300, driven by the 35S and UBI promoter, respectively. To construct the OsBZR4p::YUC4 vector, 2032-bp OsBZR4 promoter fragment was cloned into pCAMBIA2300, and then the YUC4 cDNA was amplified and inserted downstream of OsBZR4 promoter. The primers for creating the above constructs are listed in Supplementary Data 4.
Embryo size measurement
The rice seeds were photographed, and the area in the embryo and whole seed was analyzed using Image J software (version 1.38; http://rsbweb.nih.gov/ij/). The area ratio of embryo to whole seed was calculated to determine the embryo size. The definition of the embryo type is as follows: normal embryo, the area ratio of embryo to whole seeds in mutants is identical to that in the corresponding WT; small embryo, the area ratio is less than that in WT; no embryo, there is no visible embryo in seeds to the naked eye. For data statistics, individual means of five independent biological replicates are shown. Each replicate contained around 100 seeds from one plant.
Pollen viability assessment
For pollen viability assays, anthers were fixed in a formalin-acetic acid-alcohol solution (5% formaldehyde, 5% glacial acetic acid, 63% ethanol, and 27% water, 5/5/63/27) Fixed anthers were ground to release pollen grains, stained with 1% (w/v) I2-KI Lugol’s iodine solution, then observed under a light microscope (Olympus BX53). Pollen grains that were stained blue were counted to score pollen fertility68.
Laser scanning confocal microscope (LSCM)
LSCM observation was performed according to a previous publication with some modifications69. Briefly, developing caryopses of 3–7 DAP were collected and fixed in FAA (3.7% formaldehyde, 5% acetic acid, and 50% ethanol) for at least 24 h, rehydrated with a graded ethanol series and distilled water. Then, the samples were mordanted with aluminum potassium sulfate (2%) for 20 min and stained with eosin Y (10 mg/L) for 12 h. The samples were washed with distilled water, dehydrated using an ethanol series, and infiltrated with an ethanol-methyl salicylate [1:1(vol/vol)] overnight, then incubated in methyl salicylate. The infiltrated caryopses were observed using excitation/emission wavelengths of 543 nm with an LSM 800 confocal microscope (Carl Zeiss, Oberkochen, Germany).
Histochemical analyses
Developing caryopses of 3–7 DAP were fixed overnight at 4 °C in 3% (w/v) paraformaldehyde and dehydrated in an ethanol series70. Following substitution with xylene, the samples were embedded in paraplast, then sectioned to an 8 μm thickness with a rotary microtome (2165, Leica Microsystems, Germany). After staining with 0.05% toluidine blue, sections were observed with a light microscope (BX63, Olympus, Japan).
GUS assay
The plant samples were collected and infiltrated with staining solution [100 mM sodium phosphate, 5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, 0.5% Triton X-100, 10 mM EDTA (pH 8.0), 0.1% 5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid/cyclohexylammonium salt (X-gluc), 2% DMSO, 5% methanol]. After vacuum infiltration for 60 min, the samples were incubated for 8 h at 37 °C71. To remove chlorophyll, the samples were incubated sequentially in 70% then 95% ethanol at 60 °C. GUS staining was observed using a stereoscopic microscope (SZX16, Olympus, Japan).
Single-nucleus sequencing of developing caryopses in rice
The 7-DAP caryopses were collected and frozen in liquid nitrogen in a 2.0 mL safe-lock EP tube. The nucleus isolation and single-nucleus sequencing of the samples were performed by Personalbio (Shanghai, China). The simple process is as follows. Cell lysis and nucleus isolation were performed in accordance with the instructions provided in CelLyticTM PN Isolation/Extraction Kit (Sigma-Aldrich, USA). The crude suspension nuclei were stained with 5 μg/mL DAPI and analyzed the overall nuclei integrity and concentration using the laser excitation lines of 405 and 488 in a Leica TCS SP5 confocal microscope and a cell counting chamber (INCYTO C-Chip, catalog number: DHC-B02-5). Simultaneously, the remaining volume of the sample was used for the fluorescence-activated nuclei sorting step. A BD FACSAriaTM IIU/III upgraded cell sorter with the following laser configuration was used to sort 40,000 nuclei at a final concentration of ~500 nuclei/μL. After the sorting, the nuclei integrity was examined under the scope as described above.
A total of 20,000 nuclei were loaded onto the 10 × Chromium Single Cell Platform (10 × Genomics) at a concentration of 700–1200 cells/μL (Single Cell 3’ library and Gel Bead Kit v.3, USA) as described in the manufacturer’s protocol. Generation of gel beads in emulsion (GEM), barcoding, GEM-RT clean-up, complementary DNA amplification, and library construction were all performed as per the manufacturer’s protocol. Qubit was used for library quantification before pooling. The final library pool was sequenced on the Illumina Nova6000 instrument by Shanghai Personalbio (Shanghai, China) using 150-base-pair paired-end reads.
Following sequencing, low-quality nuclei for which over 20% of their reads were mapped to plastids or mitochondria were removed. The nuclei with gene numbers detected out of the range of 400–5000 were also removed. Data were collected from 2 independent biological batches. R package Seurat (V4.1.1) was used for dimension-reduction and clustering. NormalizeData and ScaleData were used to normalize and scale all gene expression, and the top 2000 variable genes with FindVariableFeautres were selected for PCA analysis. FindClusters was used to separate cells into multiple clusters using the top 20 principal components. Batch effect between samples was removed by Harmony. Cells were separated into multiple clusters by using the Louvain algorithm and setting the resolution parameter at 0.8. Cell clusters were visualized by using UMAP.
To identify differentially expressed genes (DEGs), we used the Seurat FindMarkers function based on the Wilcox likelihood-ratio test with default parameters, and selected the genes expressed in more than 25% of the cells in a cluster and with an average log (Fold Change) value greater than 1 as DEGs. The specifically expressed genes in a cluster refer to the DEGs that are exclusively expressed in a single cluster and exhibit an average log (Fold Change) value less than 1 in other clusters. To investigate the potential functions of DEGs, the GO analysis was used with the “clusterProfiler” R package. Pathways with p-adj value less than 0.05 were considered as significantly enriched.
RNA in situ hybridization
Specific fragments of OsBZR4, GE and SWEET14 were amplified using the primers listed in Supplementary Data 4, and fused with the T7 or SP6 promoter. Subsequently, these fragments were synthesized and labeled using the DIG RNA Labeling Kit (Roche). Developing caryopses of 7 DAP were fixed in 3% (w/v) paraformaldehyde and dehydrated in an ethanol series and embedded in Paraplast Plus (Sigma-Aldrich, St. Louis)72. Following tissue sectioning using a microtome (2165; Leica Microsystems) and digested with Proteinase K (Roche), hybridized with corresponding probes, and incubated with anti-digoxigenin-AP Fab fragments (Roche). After washing, the signals were detected with NBT/BCIP stock solution (Roche).
Quantification of endogenous auxin content
The developing caryopses were collected and immediately placed in liquid nitrogen, and a sample of at least 0.2 g was taken from each. Endogenous IAA content was determined by UPLC-MS/MS analysis at Wuhan Greensword Creation Technology Co., Ltd.
The chemical treatment
For the BL, 2,4-D, and NPA treatments, the panicles were infiltrated with BL, 2,4-D, or NPA daily from 1 DAP for a week. A total of 10 mL of solution was used per plant in each treatment. The hormones were applied between 16:00 and 18:00 h to avoid photolysis of the chemicals. Ethanol and DMSO were used to aid in the dissolution of chemicals, while distilled water containing the same concentration of ethanol and DMSO was used as a control. Twenty plants were used in each treatment, and the embryo phenotypes were observed after the seeds matured.
In vitro, culture of detached caryopses was performed as described by Uchiumi and Okamoto73. Briefly, SJ2 spikelet taken at 10–12 h after pollination were cultured on medium containing different chemicals, including 1 μM or 50 μM 2,4-D, and 2 μM BRZ. The hormone-free medium was used as a control. After 20 days of culturing, the embryo morphology in the developing caryopses was examined with a dissecting microscope.
Gene expression analysis
Total RNA from various rice tissues was isolated using RNA extraction kit (Invitrogen) and was reverse transcribed with the PrimeScriptTM RT reagent Kit with gDNA Eraser (Perfect Real Time) from TaKaRa according to the manufacturer’s instructions. Quantitative real-time PCR reactions were done on the LightCycler 96 Instrument (Roche, Germany) with three biological replicates. The expression of Ubiquitin was used as an internal control. The primers for quantitative real-time PCR are summarized in Supplementary Data 5. Analysis of protein expression levels in transgenic plants was performed using Western blot. Total protein was extracted from the transgenic plants. The target protein was detected using an anti-Flag antibody (Abmart, M20008, 1:3000), with an anti-Actin antibody (Abmart, M20009, 1:3000) serving as the internal control. The secondary antibody used was goat anti-mouse IgG (H + L) secondary antibody, HRP (Thermo Fisher, 31430, 1:10000).
Electrophoretic mobility shift assay
The N-terminal (1–324 bp) of OsBZR4 was cloned into the vector pET-32a to generate the His-nOsBZR4 recombinant proteins. The recombinant protein was expressed in Escherichia coli Rosetta and purified by High Affinity Ni-Charged Resin FF (GenScript). Oligonucleotide probes of YUC4 promoter containing the wild-type and mutant BRRE-box motif were synthesized and labeled with biotin using the EMSA Probe Biotin Labeling Kit (Beyotime). For unlabeled probe competition, an unlabeled probe was added to the reactions. EMSA was performed using a Chemiluminescent EMSA kit (Beyotime). Probe sequences are shown in Supplementary Data 4.
ChIP-qPCR analysis
Two grams of 2-week-old shoot tissue was harvested from Flag-OsBZR4 overexpressing plants and then crosslinked in 1% (v/v) formaldehyde for 30 min at room temperature. The cross-linking reaction was then stopped by the addition of glycine (final concentration, 0.125 M). Chromatin was extracted and fragmented using sonication with an ultrasonic disruptor (BC100, Diagenode Bioruptor, China). 2.5 μg anti-Flag antibody (Abcam, M20008) and 20 μl protein A/G Magbeads (Genscript, L00277) were used for immunoprecipitations. The precipitated DNA was recovered using the QIAGEN DNA purification kit (QIAGEN, 28104) for the subsequent qRT-PCR. All primers used for ChIP-qPCR assays are listed in Supplementary Data 4.
Transient transcriptional activity assay
For transient transcriptional activity analysis, the YUC4 promoter regions (upstream of the ATG) were cloned into pGreenII0800-luc double reporter vector and used as a reporter. The whole length CDS of PIL13 was cloned into PRT107 to generate 35Sp::PIL13, which was used as an effector. The reporter and effector constructs were co-transformed into rice leaf protoplasts by the PEG method74. Signals of Firefly Luciferase (LUC) and Renilla Luciferase (REN) were collected with a Dual-LUC reporter assay kit (Beyotime; RG027). The 35S::GFP was used as a control. Relative LUC activity was calculated by normalizing LUC activity to REN activity. The sequences of the specific primers are listed in Supplementary Data 4.
Calculation of milled rice yield
Head rice refers to the rice that has been husked into brown rice by a huller, and then ground into national standard Grade 3 (GB1354, China) rice by a rice mill, with its length reaching three-quarters or more of the average length of the whole rice grains. The milled rice yield is the percentage of milled rice to the total weight of unhusked rice.
Determination of fatty acid value in rice
The fatty acid value of the samples was measured according to a previous publication with some modifications75. Briefly, 10 g rice powder was extracted with 50 mL of absolute ethyl alcohol for 10 min, and then filtered. 50 mL distilled water was added to 25 mL filtrate and immediately titrated with potassium hydroxide (0.01 M) to determine the end-point using an automatic potentiometric titrator (T9616, Hanon, China). The values were expressed by the mass (mg) of KOH required to neutralize the free fatty acids contained in 100 g of rice flour.
Determination of seed lipoxygenase (LOX) activity
Total protein from rice seeds was extracted with 0.1 M sodium phosphate buffer (pH 6.3), and LOX activity was measured using sodium linolenate as substrate76. Absorbance at 234 nm of the enzyme reaction solution was measured (e = 25,000), and an amount of the product was calculated. One unit of LOX was defined as producing 1 nmol of peroxide per min. Protein concentrations were determined using a BCA protein assay kit (Beyotime) with bovine serum albumin as a standard.
Statistics and reproducibility
All data are shown as the mean ± standard deviation (SD), shown by error bars. Unless indicated otherwise. Comparisons were made by two-tailed Student’s t test or one-way ANOVA and Tukey’s multiple comparisons test (P < 0.05) in statistical software GraphPad Prism 8.0.2 and IBM SPSS Statistics 25. No statistical method was used to predetermine sample size.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data and materials are available from the corresponding authors upon request. The raw data for snRNA-seq generated in this study have been deposited in the National Genomics Data Center (NGDC), China National Center for Bioinformation under accession OMIX010934 and BioProject accession PRJCA042164. Genes sequence data from this study can be accessed from the Rice Genome Annotation Project website (https://rice.uga.edu/) under the following accession numbers: OsBZR4 (LOC_Os02g13900 [https://rice.uga.edu/cgi-bin/ORF_infopage.cgi?orf=LOC_Os02g13900.1]), OsYUC4 (LOC_Os01g12490 [https://rice.uga.edu/cgi-bin/ORF_infopage.cgi?orf=LOC_Os01g12490.1]), and OsPIL13 (LOC_Os03g56950 [https://rice.uga.edu/cgi-bin/ORF_infopage.cgi?orf=LOC_Os03g56950.2]). Source data are provided with this paper.
References
Ren, D., Ding, C. & Qian, Q. Molecular bases of rice grain size and quality for optimized productivity. Sci. Bull. 68, 314–350 (2023).
Sen, P. et al. Genetic and functional mechanisms of yield-related genes in rice. Acta Physiol. Plant. 46, 29 (2024).
Li, Y. et al. Chalk5 encodes a vacuolar H(+)-translocating pyrophosphatase influencing grain chalkiness in rice. Nat. Genet. 46, 398–404 (2014).
Itoh, J. et al. Rice plant development: from zygote to spikelet. Plant Cell Physiol. 46, 23–47 (2005).
Wu, X., Liu, J., Li, D. & Liu, C. M. Rice caryopsis development II: dynamic changes in the endosperm. J. Integr. Plant Biol. 58, 786–798 (2016).
Doll, N. M. et al. A two-way molecular dialogue between embryo and endosperm is required for seed development. Science 367, 431–435 (2020).
Hara, T. et al. Rice SNF2 family helicase ENL1 is essential for syncytial endosperm development. Plant J. 81, 1–12 (2015).
Nagasawa, N. et al. GIANT EMBRYO encodes CYP78A13, required for proper size balance between embryo and endosperm in rice. Plant J. 75, 592–605 (2013).
Yang, W. et al. Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome p450. Mol. Plant 6, 1945–1960 (2013).
Chen, Y. et al. Loss of function of the cytochrome P450 gene CYP78B5 causes giant embryos in rice. Plant Mol. Biol. Rep. 33, 69–83 (2015).
Nagato, Y., Kitano, H., Kamijima, O., Kikuchi, S. & Satoh, H. Developmental mutants showing abnormal organ differentiation in rice embryos. Theor. Appl. Genet. 78, 11–15 (1989).
Hong, S. K., Aoki, T., Kitano, H., Satoh, H. & Nagato, Y. Temperature-sensitive mutation, embryoless 1, affects both embryo and endosperm development in rice. Plant Sci. 108, 165–172 (1995).
Hong, S.-K., Aoki, T., Kitano, H., Satoh, H. & Nagato, Y. Phenotypic diversity of 188 rice embryo mutants. Dev. Genet. 16, 298–310 (1995).
Hong, S.-K., Kitano, H., Satoh, H. & Nagato, Y. J. D. How is embryo size genetically regulated in rice? Development 122 7, 2051–2058 (1996).
Kamiya, N. et al. Rice globular embryo 4 (gle4) mutant is defective in radial pattern formation during embryogenesis. Plant Cell Physiol. 44, 875–883 (2003).
Yi, J. et al. OsMPK6 plays a critical role in cell differentiation during early embryogenesis in Oryza sativa. J. Exp. Bot. 67, 2425–2437 (2016).
Liu, D. et al. Diversification of plant agronomic traits by genome editing of brassinosteroid signaling family genes in rice. Plant Physiol. 187, 2563–2576 (2021).
Yin, W. et al. Understanding brassinosteroid-centric phytohormone interactions for crop improvement. J. Integr. Plant Biol. 67, 563–581 (2025).
Shi, H., Li, X., Lv, M. & Li, J. BES1/BZR1 family transcription factors regulate plant development via brassinosteroid-dependent and independent pathways. Int. J. Mol. Sci. 23, 10149 (2022).
Tian, X. et al. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. Plant Cell 33, 2753–2775 (2021).
Ren, Y. et al. Oryza sativa mediator subunit OsMED25 interacts with OsBZR1 to regulate brassinosteroid signaling and plant architecture in rice. J. Integr. Plant Biol. 62, 793–811 (2020).
Moller, B. & Weijers, D. Auxin control of embryo patterning. Cold Spring Harb. Perspect. Biol. 1, a001545 (2009).
Liu, C., Xu, Z. & Chua, N. H. Auxin polar transport is essential for the establishment of bilateral symmetry during early plant embryogenesis. Plant Cell 5, 621–630 (1993).
Hadfi, K., Speth, V. & Neuhaus, G. Auxin-induced developmental patterns in Brassica juncea embryos. Development 125, 879–887 (1998).
Xu, X. et al. OsYUC11-mediated auxin biosynthesis is essential for endosperm development of rice. Plant Physiol. 185, 934–950 (2021).
Ishimaru, K. et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat. Genet. 45, 707–711 (2013).
Liu, L. et al. Activation of Big Grain1 significantly improves grain size by regulating auxin transport in rice. Proc. Natl. Acad. Sci. USA 112, 11102–11107 (2015).
Zhao, Z. et al. A role for a dioxygenase in auxin metabolism and reproductive development in rice. Dev. Cell 27, 113–122 (2013).
Sazuka, T. et al. A rice tryptophan-deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos. Plant J. 60, 227–241 (2009).
Inukai, Y. et al. Crown rootless1, which is essential for crown root formation in rice, is a target of an auxin response factor in auxin signaling. Plant Cell 17, 1387–1396 (2005).
Luo, Y., Huang, D., Wu, L. & Zhu, J. The impact of metal silos on rice storage and storage losses in China. Food Secur. 14, 81–92 https://doi.org/10.1007/s12571-021-01194-4 (2021).
Xu, H. et al. Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity. Plant Biotechnol. J. 13, 526–539 (2015).
Ito, M. et al. Position-dependent expression of GL2-type homeobox gene, Roc1: significance for protoderm differentiation and radial pattern formation in early rice embryogenesis. Plant J. 29, 497–507 (2002).
Ito, M. et al. Roles of Rice GL2-type homeobox genes in epidermis differentiation. Breed. Sci. 53, 245–253 (2003).
Sato, Y. H. S. et al. A rice homeobox gene, OSH1, is expressed before organ differentiation in a specific region during early embryogenesis. PNAS 93, 8117–8122 (1996).
Nishimura, A., Ito, M., Kamiya, N., Sato, Y. & Matsuoka, M. OsPNH1 regulates leaf development and maintenance of the shoot apical meristem in rice. Plant J. 30, 189–201 (2002).
Khanday, I., Skinner, D., Yang, B., Mercier, R. & Sundaresan, V. A male-expressed rice embryogenic trigger redirected for asexual propagation through seeds. Nature 565, 91–95 (2019).
Doll, N. M. et al. Transcriptomics at maize embryo/endosperm interfaces identifies a transcriptionally distinct endosperm subdomain adjacent to the embryo scutellum. Plant Cell 32, 833–852 (2020).
Fu, Y. et al. Spatial transcriptomics uncover sucrose post-phloem transport during maize kernel development. Nat. Commun. 14, 7191 (2023).
Robert, H. S. et al. Maternal auxin supply contributes to early embryo patterning in Arabidopsis. Nat. Plants 4, 548–553 (2018).
Figueiredo, D. D. & Kohler, C. Auxin: a molecular trigger of seed development. Genes Dev. 32, 479–490 (2018).
Yamamoto, Y., Kamiya, N., Morinaka, Y., Matsuoka, M. & Sazuka, T. Auxin biosynthesis by the YUCCA genes in rice. Plant Physiol. 143, 1362–1371 (2007).
Martinez, C. et al. PIF4-induced BR synthesis is critical to diurnal and thermomorphogenic growth. EMBO J. 37, e99552 (2018).
Liu, S. et al. Adventitious root formation in rice requires OsGNOM1 and is mediated by the OsPINs family. Cell Res. 19, 1110–1119 (2009).
Wu, W. et al. Auxin-responsive OsMADS60 negatively mediates rice tillering and grain yield by modulating OsPIN5b expression. Plant J. 121, e70107 (2025).
Franklin, K. A. et al. Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. USA 108, 20231–20235 (2011).
Leivar, P. & Monte, E. PIFs: systems integrators in plant development. Plant Cell 26, 56–78 (2014).
Zhao, H. & Bao, Y. PIF4: integrator of light and temperature cues in plant growth. Plant Sci. 313, 111086 (2021).
Mo, W. et al. Phytochrome-interacting factor-like14 and slender rice1 interaction controls seedling growth under salt stress. Plant Physiol. 184, 506–517 (2020).
Lee, G. et al. Identification and characterization of large embryo, a new gene controlling embryo size in rice (Oryza sativa L.). Rice 12, 22 (2019).
Hu, Z. et al. Identification of a new giant emrbryo allele, and integrated transcriptomics and metabolomics analysis of giant embryo development in rice. Front. Plant Sci. 12, 697889 (2021).
Zhao, D., Zhang, C., Li, Q. & Liu, Q. Genetic control of grain appearance quality in rice. Biotechnol. Adv. 60, 108014 (2022).
Saikrishna, A., Dutta, S., Subramanian, V., Moses, J. A. & Anandharamakrishnan, C. Ageing of rice: a review. J. Cereal Sci. 81, 161–170 (2018).
Feussner, I. & Wasternack, C. The lipoxygenase pathway. Annu. Rev. Plant Biol. 53, 275–297 (2002).
Zhang, Y. et al. Effect of the absence of lipoxygenase isoenzymes on the storage characteristics of rice grains. J. Stored Prod. Res. 43, 87–91 (2007).
Long, Q. et al. Molecular genetic characterization of rice seed lipoxygenase 3 and assessment of its effects on seed longevity. J. Plant Biol. 56, 232–242 (2013).
Suzuki, Y. et al. Volatile components in stored rice [Oryza sativa (L.)] of varieties with and without lipoxygenase-3 in seeds. J. Agric. Food Chem. 47, 1119–1124 (1999).
Hu, X. et al. Change in volatiles, soluble sugars and fatty acids of glutinous rice, japonica rice and indica rice during storage. LWT 174, 114416 (2023).
Tang, J., Huang, X., Sun, M. & Liang, W. DWARF TILLER1 regulates apical-basal pattern formation and proper orientation of rice embryos. Plant Physiol. 196, 309–322 (2024).
Opsahl-Ferstad, H. G., Le Deunff, E., Dumas, C. & Rogowsky, P. M. ZmEsr, a novel endosperm-specific gene expressed in a restricted region around the maize embryo. Plant J. 12, 235–246 (1997).
Giuliani, C., Consonni, G., Gavazzi, G., Colombo, M. & Dolfini, S. Programmed cell death during embryogenesis in maize. Ann. Bot. 90, 287–292 (2002).
Yang, J., Luo, D., Yang, B., Frommer, W. B. & Eom, J. S. SWEET11 and 15 as key players in seed filling in rice. N. Phytol. 218, 604–615 (2018).
Yang, G. et al. The tonoplast-localized transporter OsABCC9 is involved in cadmium tolerance and accumulation in rice. Plant Sci. 307, 110894 (2021).
Qiao, J. et al. A novel miR167a-OsARF6-OsAUX3 module regulates grain length and weight in rice. Mol. Plant 14, 1683–1698 (2021).
Ma, B. et al. A plasma membrane transporter coordinates phosphate reallocation and grain filling in cereals. Nat. Genet 53, 906–915 (2021).
Zhang, S. et al. Sustained productivity and agronomic potential of perennial rice. Nat. Sustain. 6, 28–38 (2023).
Ma, X. et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 8, 1274–1284 (2015).
Tang, J. et al. WRKY53 negatively regulates rice cold tolerance at the booting stage by fine-tuning anther gibberellin levels. Plant Cell 34, 4495–4515 (2022).
Huang, X., Peng, X. & Sun, M. X. OsGCD1 is essential for rice fertility and required for embryo dorsal-ventral pattern formation and endosperm development. N. Phytol. 215, 1039–1058 (2017).
Yoon, J. et al. The BEL1-type homeobox gene SH5 induces seed shattering by enhancing abscission-zone development and inhibiting lignin biosynthesis. Plant J. 79, 717–728 (2014).
Nakata, M. et al. High temperature-induced expression of rice alpha-amylases in developing endosperm produces chalky grains. Front. Plant Sci. 8, 2089 (2017).
Yi, J. et al. The rice gene defective tapetum and meiocytes 1 (DTM1) is required for early tapetum development and meiosis. Plant J. 70, 256–270 (2012).
Uchiumi, T. & Okamoto, T. Rice fruit development is associated with an increased IAA content in pollinated ovaries. Planta 232, 579–592 (2010).
Tian, X. et al. Transcription Factor OsWRKY53 positively regulates brassinosteroid signaling and plant architecture. Plant Physiol. 175, 1337–1349 (2017).
Biao, Y. et al. Influence of gene regulation on rice quality: impact of storage temperature and humidity on flavor profile. Food Chem. 283, 141–147 (2019).
Mizuno, K., Iida, T., Takano, A., Yokoyama, M. & Fujimura, T. A new 9-lipoxygenase cDNA from developing rice seeds. Plant Cell Physiol. 44, 1168–1175 (2003).
Acknowledgements
We thank Prof. J. Tang (Anhui Agricultural University) for providing bzr mutants, Prof. R. Lin (Xianghu Laboratory) for providing OsPIL-OE lines, Prof. L. Xiong (Huazhong Agricultural University) for providing DR5-GUS line, and Prof. Z. Fang (Guizhou University) for providing PIN5b OE line. This work was supported by the National Key Research and Development Program of China (2024YFD1201001-3 to Q.B.), the National Natural Science Foundation of China-Heilongjiang Joint Fund (U23A20193, to Q.B.), the National Natural Science Foundation of China (32171988, to Q.B.), the National Natural Science Foundation of China (32171989, Z.W.), and the Heilongjiang Provincial Key Research and Development Program Innovation Base Project (JD24A011, Z.W.).
Author information
Authors and Affiliations
Contributions
Q.B., Z.W. and Q.Q. conceived and designed the study. Z.W., M.X. and Y.L. performed experiments. X.Z., Z.H., M.H., X.J., J.T., X.L. and X.T. provided assistance. Q.B., Z.W., M.X. and Q.Q. analyzed the data and wrote the manuscript. Q.B. and Q.Q. supervised the study.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Duarte Figueired, Yutaka Sato and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, Z., Xu, M., Liu, Y. et al. OsBZR4 regulates temperature-dependent embryogenesis in rice. Nat Commun 16, 6893 (2025). https://doi.org/10.1038/s41467-025-62262-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-62262-3
