
Abstract
To navigate complex environments, cells integrate chemical and mechanical cues through dynamic feedback between signaling networks and the cytoskeleton. Using synthetic tools to manipulate cytoskeletal components in Dictyostelium and human neutrophils, we uncover feedback mechanisms that regulate Ras/PI3K signaling and control front- and back-states of the cell. Increased branched actin and actin polymerization enhance Ras/PI3K activity. Similarly, decreased myosin II assembly also elevates signaling and chemotactic sensitivity. Conversely, inhibiting branched actin increases cortical actin and blocks Ras/PI3K activation—an effect lessened by decreasing filamentous actin or in myosin II-null cells. Activating RacE to increase actin crosslinking suppresses Ras activity without triggering branched actin nucleators, yet promotes spreading and protrusion. These results informed a computational model incorporating positive cytoskeletal feedback loops, which predicts shifts in polarity and migration with cytoskeletal changes. We propose that such feedback locally tunes signal network excitability, enabling cells to navigate tissues, extracellular matrix, and fluid environments.
Similar content being viewed by others


Introduction
Cell migration is a fundamental process that plays critical roles in development, immune system function, and disease progression, including metastasis. While there is a broad diversity of cell migration strategies, efficient motion usually depends on the spontaneous formation of membrane protrusions and the establishment of front-back polarity. Both protrusion formation and stable front-back polarity rely on coordination between actin cytoskeletal dynamics and signaling networks. Much is known about how signaling controls cytoskeletal activity, but very little is known about the potential feedback from changes in cytoskeletal organization and mechanics on these same signaling networks1,2.
Regulation of the actin cytoskeleton is intricately linked to signaling molecules, such as Ras GTPases and phosphoinositides. Primarily at the cell front, the activation of Ras GTPase, associated signaling molecules, and the lowering of anionic lipids correlate strongly with the formation of cell protrusions3,4,5,6,7,8. Indeed, amoeboid migrating cells rarely form stable protrusions in the absence of spontaneous or triggered signal network activation9,10,11,12. In amoeboid migrating cells, protrusion formation involves the activation of branched actin nucleator Arp2/3 and the local disassembly of myosin II (also referred to here as ‘myosin’ for simplicity) filaments and actin-actin crosslinkers13,14,15,16,17,18. Previous studies have suggested positive feedback exists between Arp2/3-mediated actin polymerization and PI3K activity at the front8,10,12,15,19,20. Synergistically, evidence suggests that negative feedback from actomyosin – typically enriched at the cell back – inhibits front signaling. For example, cells lacking myosin have elevated Ras activation, and retracting pseudopods are insensitive to chemotactic stimulation21,22,23. Together, these findings suggest that branched actin and actomyosin networks may differentially feedback on signaling networks. However, because the network governing cell migration is complex and adaptable, local and acute perturbations are critical to dissect the cause and nature of cytoskeletal feedback.
Both signal transduction networks and actin cytoskeletal networks exhibit properties of excitability, including an activation threshold and propagating waves9,24,25,26,27,28,29,30. Waves of Ras-GTP, phosphatidylinositol 3,4,5-trisphosphate (PIP3), actin polymerization, and Arp 2/3 traverse the ventral cell membrane and define the “front” state of the cortex. Complementary zones (“shadow waves”) of PI(4,5)P2, cortexillin, and myosin II depletion are also associated with this Ras-GTP/PIP3 state of the cortex. While waves of signaling and cytoskeletal activity are coupled, they are morphologically distinct and independent25. In the absence of the cytoskeleton, signaling waves still propagate but cannot create protrusions, and in the absence of signaling activity, cytoskeletal activity exhibits excitable properties but does not propagate in broad waves that move cells9,28. These networks have been referred to as the Signal Transduction Excitable Network (STEN) and the Cytoskeletal Excitable Network (CEN), respectively. Previous studies suggest that modifying the threshold of STEN tunes the size and speed of these waves and ultimately the nature of actin-based protrusions3,25. Changes in wave behavior are therefore a surrogate for alterations in the STEN and CEN and can predict effects on cell behavior.
The properties of the signaling and cytoskeletal networks can be captured in computational models3,25,31,32,33,34. Excitable systems require both fast positive and delayed negative feedback35. In the models, we previously reported mutual inhibition between front and back activities, which leads to a positive feedback loop, and molecules at the cell front produced their own inhibitor on longer timescales3,9. These models predicted the behavior of coupled STEN-CEN waves and the response of cells to chemotactic stimulus. However, these models oversimplified the behavior of the cytoskeleton in this process. CEN activity represented the activation of Arp2/3 by Rac at the cell front and did not consider regulation to and feedback from the morphologically distinct actomyosin network at the cell back25. Furthermore, they have not been able to easily explain experimental observations about front-back polarity, which was shown to be dependent on cytoskeletal architecture36.
We sought to understand the impact of modulating the F-actin or actomyosin cytoskeletal structures on the STEN. First, prior reports indicated a positive effect of cytoskeletal activity on PIP3 production. Here, we show that increasing the pool of available actin monomers and promoting the formation of branched actin networks leads to the activation of the Ras. Second, it was shown previously that increasing Ras activity can lower myosin assembly16,37. Here, by designing precise chemically induced dimerization (CID) and optogenetic tools to control myosin assembly in real time, we found that disassembly of the actomyosin cortex increases STEN activity. Together, these mutually negative effects constitute a positive loop that augments the back state. Consistently, increasing the abundance of linear actin and cross-linked actin inhibits STEN activation. Moreover, inhibiting branched actin nucleation further enhances the overall effect. These findings were incorporated into a computational model, which predicted the observed effects of altering cytoskeletal feedback on signaling and migration. Our study here delineates the reciprocal relationship between cytoskeletal dynamics and signaling networks that takes place during cell migration. We discuss how the cross-interaction of molecular mechanisms that we show may allow cells to integrate signal transduction and mechanical cues and successfully navigate complex three-dimensional environments.
Results
Positive and negative feedback loops from the cytoskeleton control signaling networks
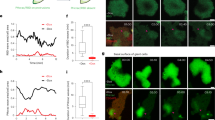
To investigate the effects of altering the cytoskeleton on signaling, we first examined the impact of increasing the amount of actin monomers available to assemble in Dictyostelium, a classic model of amoeboid migration. While there are differences in the networks that drive cell migration in these cells and other amoeboid cells like human neutrophils (e,g., the lack of clear homologs for Rock, RhoA, and CDC42), most molecules, network topologies, and cell morphology and migration behavior are quite similar38. Previously, we found that mutating actobindins, which bind and sequester free G-actin, increases actin polymerization at the cell front39,40. Here, to determine the effects of increased actin assembly on signaling, we measured Ras activation levels using the Ras-Binding Domain (RBD) of Raf41 in actobindin triple knockout (ABN ABC-) cells to examine the effect of increased “front” actin network formation on STEN activity (Fig. 1a–c, Supplementary Movie 1). Significantly, the ABN ABC- cells exhibited elevated levels of Ras activation, suggesting that increases in the abundance of front actin activates STEN. Compared to wild-type cells, actobindin mutants show increased localization of Arp2 protein ArpB and WAVE complex subunit HSPC300, previously shown to directly precede actin polymerization9, to the cell front (Fig. S1a–e). Differences in Ras activation or ArpB localization are not correlated to differences in expression level or fluorescent protein tag (Figs. 1d, S1c, S1f, g) and no change in membrane morphology was observed (Fig. S1h). Because previous reports suggested that lowering Arp2/3 activity reduces PIP3 levels, we sought to specifically reduce Arp2/3 activity by overexpressing Arp2/3 inhibitor protein Arpin and assess the effect on Ras activity. Arpin overexpression reduced Ras activation levels compare AX3 controls (Fig. 1a–c). To confirm that Ras activation and Arp2/3 activity are directly related, we performed a biochemical RBD pulldown assay in AX3 cells, ABN ABC- cells, and AX3 cells treated with Arp2/3 inhibitor CK666 (Fig. S2). Immunoblotting the products of this pulldown with Pan-Ras antibody reveals that ABN ABC- cells and AX3 CK666-treated cells exhibit approximately 2-fold and 0.66-fold activation, respectively, relative to AX3 controls, in close agreement with imaging data (Fig. S2). This result indicates the presence of a positive feedback loop from branched actin to STEN activities.

a Spinning disk confocal imaging of Ras activation (RBD-EGFP/RBD-mCherry) in wild-type (AX3) and actobindin ABC triple knockout (abnABC−) Dictyostelium cells. Arpin-OE cells are AX3 cells expressing Arp2/3-inhibitor Arpin-EGFP. b Membrane kymographs (left) of RBD corresponding to the movies in a. Scale bars = 2 minutes (horizontal) and 5 µm (vertical). Diagram (left) depicts the method of creating a membrane kymograph with 2 Fluorescent Proteins (FPs). c, d Individual (dots) and average (lines) percentages of the membrane periphery with RBD localization significantly above background (C) and average total cell intensity in AX3, abnABC−, and Arpin-OE cells. n=cells, *=p < 0.05, ***= p < 0.005, 2-sided Mann-Whitney U test. e Scanning confocal imaging of mCherry-FRB-MHCKC membrane recruitment and cell shape (brightfield) in AX3 cells. Cells are also expressing an unlabeled membrane-localized FKBP domain (cAR1-2xFKBP, see methods). t = 00:00 indicates rapamycin addition. f Examples (left) and diagram (right) of temporal color projections of cell outlines corresponding to the movie in (D). Blue and yellow times indicate the first and last images in the projection, respectively. g Average (line) and SEM (shaded area) of normalized (top) or absolute (bottom) cell area before and after rapamycin addition in cells expressing FRB-MHCKC (cyan) or FRB alone (orange). (dashed line, t = 0). n = 21 and 17 cells, respectively. h Scanning confocal imaging of RBD and MHCKC membrane recruitment in AX3 cells. t = 00:00 indicates rapamycin addition. i Membrane kymograph of Ras activation corresponding to the movie in (g). Dashed line indicates rapamycin addition, scale bars = 2 minutes (horizontal) and 5 µm (vertical). j Average and SEM of normalized percentages of the membrane periphery with RBD localization significantly above background before and after rapamycin addition (dashed line, t = 0) in AX3 cells expressing FRB-MHCKC (cyan) and FRB alone (orange) and in myosin II-null (myoII-) cells expressing FRB-MHCKC (magenta). n = 20,32, and 31, respectively. Time is in min:sec; scale bars = 5 µm unless otherwise noted. Source data are provided as a Source Data file.
While previous reports suggest that myosin constrains protrusions22,42,43,44,45,46, it is not clear whether this inhibition is purely mechanical or functions through signaling networks, or both. We sought to determine whether specifically and acutely lowering Myosin altered spontaneous signaling activity. We utilized myosin heavy chain kinase C (MHCKC) to control bipolar thick filament (BTF) assembly of myosin. MHCKC is recruited to the cell cortex after chemoattractant stimulation, leading to myosin phosphorylation and disassembly at specific locations47,48. However, since chemoattractant addition triggers multiple events simultaneously, we aimed to develop a method for specifically controlling myosin assembly. To achieve this, we designed a chemically induced dimerization system (CID) utilizing a MHCKC-FRB fusion protein, which is expected to dimerize with cAR1-2xFKBP (cAR1 is a uniformly distributed membrane protein), upon addition of rapamycin (Fig. 1e, Supplementary Movie 2).
To confirm BTF disassembly, we engineered the CID system into myosin II-null (mhcA null) cells rescued with GFP-myosin II (mhaA::GFP-myosin II) and imaged with total internal reflection fluorescence (TIRF) microscopy. Adding rapamycin led to uniform recruitment of MHCKC to the membrane and an approximately 40% decrease in the intensity of GFP-myosin in the cortex as viewed via TIRF imaging (Fig. S3a, b). Imaging a cytosolic protein using confocal microscopy revealed that MHCKC recruitment to the cell periphery induced cell spreading, with an approximately 20% increase in cell area, and triggered dynamic changes in cell shape that persisted for at least 20 minutes (Fig. 1e–g, Supplementary Movie 2). Recruiting FRB alone to the membrane with rapamycin had no effect on cell area (Fig. 1g).
Increased area and shape change suggested that MHCKC recruitment increased the cells deformability, most likely by reducing its elastic modulus, which is a short time-scale mechanical parameter49. This increased deformability then likely makes protrusion formation more likely. To analyze the impact of MHCKC recruitment on STEN activity specifically, we imaged RBD-GFP, an active Ras biosensor as one of the indicators of STEN activation50. MHCKC recruitment resulted in a 50% increase in Ras activity at the periphery where protrusions formed (Fig. 1h–j, Supplementary movie 3). This increase was not observed when recruiting the FRB domain alone or when recruiting MHCKC in myosin II-null cells (Figs. 1h–j, S3c, d). In fact, Ras activation slightly decreased in these cells, indicating that in the absence of lowered Myosin activity rapamycin may have a minor negative effect on basal STEN activation.
Previously, we found that reducing PI(4,5)P2 levels lowers the STEN threshold, increasing the size and number of Ras and PIP3 membrane patches3. We sought to determine whether inhibiting myosin would similarly lower this threshold. To address this, we prepared sets of cells having MHCKC recruited to the membrane by pretreatment with rapamycin with untreated cells having no recruitment. We exposed each set to two doses of cAMP (1 nM and 100 nM). At 1 nM cAMP, the pretreated set exhibited a higher average peak response, as measured by depletion of RBD-GFP from the cytoplasm, compared to the untreated set, while at 100 nM cAMP, the two sets responded similarly (Fig. S4a, b). Altogether, these findings reveal a negative feedback loop from myosin, which increases the STEN activation threshold and inhibits front state formation.
The balance of branched and cortical actin controls STEN activity
To further investigate how cytoskeletal feedback loops cooperate to tune signaling, we inhibited branched actin nucleation with the Arp2/3 inhibitor CK66651 and examined changes in actin network morphology and STEN activation. LimEΔCoil, a biosensor for polymerizing actin, is enriched in growing pseudopods and macropinosomes in randomly migrating Dictyostelium cells52. As expected, within 2 minutes of CK666 treatment, the formation of new pseudopods was significantly inhibited. Interestingly, instead of leaving the cell periphery, LimE redistributed from transient patches and ruffles into a more homogenous and uniform, cortical cytoskeletal structure, which began to form after 4 minutes and reached completion after 12 minutes (Fig. 2a, b, Supplementary Movie 4).

a Scanning confocal imaging of polymerizing actin (LimEΔCoil-RFP) and PIP3 (PHCRAC-YFP) in wild type (AX3) Dictyostelium cells before and after treatment with the Arp2/3 inhibitor CK666 (CK). t = 00:00 indicates CK666 addition. b Average (line) and SEM (shaded area) of the total normalized intensity and the variance of the peripheral intensity of LimE (magenta) and PHCRAC (green) patches before and after CK666 addition (dashed line, t = 0). n = 27 cells. c TIRF imaging of activated Ras (RBD-mCherry) in electrofused (“giant”) AX3 cells before and after CK666 and latrunculin addition. Cells are incubated in caffeine to raise basal activity levels. t = 00:00 indicates the addition of CK666 or, in parentheses, latrunculin. d Average and SEM of average RBD membrane intensity in giant AX3 cells before and after CK666 addition (left, dashed line, t = 0) or in CK666 before and after latrunculin addition (right, dashed line, t = 0). n = 22 cells. e Individual traces from d. t = 00:00 indicates CK666 addition; dashed lines correspond to drug addition. f Spinning disk confocal imaging of PIP3 (RFP-PHAKT) in human neutrophil-like (dHL60) cells before and after CK666 and latrunculin addition. t = 00:00 indicates the addition of CK666 or, in parentheses, latrunculin. g Average and SEM of the normalized PHAKT membrane-to-cytosol ratio before and after CK666 (left, n = 51 cells) and subsequently latrunculin (right, n = 65 cells) addition in dHL60 cells. h Individual (dots) and average (lines) PHAKT membrane-to-cytosol ratio normalized to the average level before CK666 (left, n = 51 cells) or latrunculin (right, n = 65 cells) addition. ***=P < 0.005, 2-sided Wilcoxon signed-rank test. i Spinning disk confocal imaging of Ras activation levels (RBD-EGFP) in developed AX3 Dictyostelium cells treated with CK666 (CK), latrunculin (Lat), or buffer (Buff) before and 6 seconds after the addition of 0.1 nM, 0.5 nM, and 10 nM cAMP. j Normalized RBD membrane-to-cytosol ratio before and after cAMP addition (dashed lines) in cells treated with CK666 (purple), latrunculin (yellow) or buffer (cyan). n = 31, 40, and 22 cells. k Individual and average maximum values of normalized RBD membrane-to-cytosol ratio in the 42 seconds after addition of cAMP. ***=p < 0.005, 2-sided Mann-Whitney U test. Time=min:sec; scale bars = 5 µm. Source data are provided as a Source Data file.
Because this uniform actin organization resembled proteins in the actomyosin cortex, we further investigated the effect of CK666 on the localization of cortexillin I. Cortexillin I is an actin-actin crosslinker, actin-membrane crosslinker, and myosin-binding protein that, as noted earlier, enriches sites of high actomyosin, including the cell back, the outside of ventral waves, and the cleavage furrow of dividing cells27,53,54,55,56,57. Treatment with CK666 led to an increase of approximately 30% in the membrane to cytosol ratio of cortexillin I (Fig. S5a, b). These results indicate that CK666 treatment not only inhibits branched actin nucleation, but also increases the level of cortical actomyosin. To further confirm this, we performed a cytoskeletal fractionation assay where membrane and cytosolic proteins are separated from a triton-insoluble pellet previously shown to be enriched with actomyosin cortex-associated proteins57. Treating cells with CK666 led to an ~2-fold enrichment of proteins in the insoluble fraction, indicating increased cortical assembly (Fig. S5c, d). The amount of GFP-Cortexillin I in the pellet increased 4-fold, 2-fold more than total protein (Fig. S5e, f). Similarly, CK666 treatment increases the amount of endogenous Cortexillin I and actin in the insoluble fraction in cells either with or without GFP-Cortexillin I expression (Fig. S5g, h).
To assess the impact of CK666 treatment on signal transduction network (STEN) activity, we employed the biosensor PHCRAC, which detects PI(3,4,5)P358,59,60. In single cells, PHCRAC overlaps with RBD and LimE in growing pseudopods and macropinosomes. Similar to actin, PIP3 disappeared from these patches following CK666 treatment. Unlike actin, PIP3 did not relocalize elsewhere (Fig. 2a, b), consistent with previous reports and our observations in Arpin-overexpression cells. To further examine how CK666 treatment affects STEN, we measured changes in STEN waves on the bottom cell surface of electrofused giant cells61. Initially, these cells displayed propagating waves of RBD along the bottom membrane; however, upon addition of CK666, these waves disappeared within two minutes (Fig. 2c–e, Supplementary Movie 5). These experiments reveal that both PIP3 production, Ras activation, and likely many other STEN activities, are reduced when branched actin is removed and cortical assembly is increased.
To find out if the drop in STEN activation after CK666 was only because of the loss of branched actin, or if the increase in cortical actin directly suppresses STEN, we introduced the actin depolymerizing drug latrunculin to CK666-treated cells. Strikingly, removal of cortical actin by latrunculin led to the recovery of RBD waves within two minutes (Fig. 2c–e, Supplementary Movie 5). No increase in STEN activity was observed after latrunculin treatment in cells not pre-treated with CK666 (Fig. S5i, j). In fact, latrunculin alone slightly lowers Ras activation. This inhibitory role of cortical actin networks on STEN activity was not limited to Dictyostelium. We conducted the same experiment using human neutrophil-like HL60 cells expressing the PIP3 biosensor PH-AKT. Consistent with previous reports, PIP3 was observed in patches at the front of migrating HL60 cells. Upon CK666 addition, these cells rounded up and lost their pseudopods within 2 minutes. The elimination of branched actin in neutrophils also resulted in a decrease in PIP3 levels on the membrane. Subsequently, the addition of latrunculin, which removed the remaining actin networks, partially restored PIP3 levels (Fig. 2f–h, Supplementary Movie 6). These findings suggest that the actomyosin cortex suppresses STEN activity and that this phenomenon is evolutionarily conserved.
To test if the inhibition of STEN by the actomyosin cortex was due to an increase in the STEN threshold, we created a dose-response curve of Ras activation in response to increasing doses of cAMP stimuli in Dictyostelium cells treated with CK666 or latrunculin (Figs. 2i–k, S6a–c). The EC50 of buffer- and latrunculin treated cells were both approximately 0.1 nM while CK666-treated cells was 1 nM, a 10-fold decrease in sensitivity (Fig. S6c). No significant difference was detectable between the three populations when stimulated with high cAMP (Figs. 2i–k, S6a–c). This relative inhibition indicates that the threshold for STEN was raised, but the system is still functional. By lowering STEN excitability, the actomyosin cortex both inhibits spontaneous activity (Fig. 2c–h) and tunes response to receptor inputs (Fig. 2i–k).
Myosin II is a critical contributor to negative feedback from the cortex to signaling networks
Because myosin is a critical component of the cortex, we investigated whether removing myosin ameliorates the effect of CK666 on the STEN. CK666 treatment in myosin-null cells prevented protrusions and decreased PIP3 levels similar to its effects in wild-type cells. However, unlike the near absence of PIP3 in wild-type cells, myosin-null cells treated with CK666 periodically exhibited bright, transient PIP3 patches (Fig. 3a–d, Supplementary Movie 7). Interestingly, slightly fewer overall patches form in myosin-null cells as compared to the AX3 strain (Fig. 3c, d). Because strain-to-strain variation exists, to directly assess the effects of myosin II on PIP3 patches, we compared the myoII null cells to myosin-null cells rescued with GFP-Myosin II. These rescued cells, in fact, had fewer patches than myosin-null cells and did not display bright patches after CK666 treatment (Fig. 3a–d). Differences in PHCRAC expression do not influence changes in patch number after CK666 treatment (Fig. S7). To further elucidate the role of the actomyosin cortex, we applied our method to rapidly dissemble myosin in cells treated with CK666. First, cells were inhibited with CK666, which reduced the number of Ras patches by 95% and immobilized the cell. Then, MHCKC was recruited to the membrane using rapamycin. While disassembling myosin filaments did not restore motility, it caused Ras activation to recover to 40% of its wild type value (Fig. 3e–g, Supplementary Movie 8). Taken together, these results show that myosin II is a major contributor to cortical feedback on the STEN.

a Scanning and spinning disk confocal images of PIP3 levels (PHCRAC-YFP/ PHCRAC-mScarlet3) in wild type (AX3), myosin II-null (myoII-) Dictyostelium cells, and myosin II-null Dictyostelium cells expressing GFP-MyosinII (myoII- rescue) before and after CK666 addition. t = 00:00 indicates CK666 addition. b Heatmap of the number of PHCRAC patches in individual AX3, myoII-, and rescue cells. Every row is a single cell and every column is a timepoint. c Average (lines) and individual (dots) mean number of PHCRAC patches over time before and after CK666 addition in AX3 and myoII−, and rescue cells. n = cells, ***=p < 0.005, 2-sided Mann-Whitney U test. d Average and individual percent decrease in the average number of PHCRAC patches after CK666 addition in AX3 and myoII−, and rescue cells. Data are from c, *** = p < 0.005 2-sided Mann-Whitney U test. e Scanning confocal images of Ras activation (RBD-EGFP) before and after CK666 addition and subsequently mCherry-FRB-MHCKC membrane recruitment in AX3 cells. Cells are also expressing an unlabeled membrane-localized FKBP domain (cAR1-2xFKBP). t = 00:00 indicates the addition of CK666 or, in parentheses, rapamycin. f Membrane kymograph of Ras activation corresponding to the movie in c. Dashed lines represent the addition of specified drug, scale bars = 2 minutes (horizontal) and 5 µm (vertical). g Average and individual mean number of RBD patches over time before and after CK666 addition and subsequently MHCKC recruitment by rapamycin addition. n = cells, *** = p < 0.005, 2-sided Mann-Whitney U test. Source data are provided as a Source Data file.
Using RacE as a tool to probe the role of cortical actin in STEN regulation
To further probe the role of the actomyosin cortex in signaling, we turned to RacE, a member of the Rac/Rho subfamily which has been shown to regulate linear actin nucleation, actin crosslinking, myosin II assembly and contraction, and cortical viscoelasticity and tension62,63,64,65,66. To abruptly increase RacE activity in migrating cells and assess signaling impacts, we designed a CID system to recruit the RacE GEF, GXCT67, to the cell membrane. Recruiting the GEF domain of GXCT to the cell membrane by adding rapamycin led to a phenotype with cells dramatically flattening and undulating rapidly around the perimeter, while losing well-defined protrusions and motility (Fig. 4a–d, Supplementary Movie 9). LimE localization shifted from a few broad patches to a uniform distribution along the cell perimeter. Quantitative analysis showed a 40% increase in F-actin periphery localization within 8 minutes after rapamycin addition (Fig. 4e–g, Supplementary Movie 10). Similarly, the amount of actin crosslinker dynacortin increased significantly upon GXCT recruitment, consistent with the role of RacE in the recruitment this cytoskeletal protein to the cortex62 (Fig. S8a–c). Furthermore, we confirmed that the changes in actin were not due to an unknown role of RacE in activating Arp2/3 by repeating the experiment in cells treated with CK666. When we recruited GXCT to the membrane following CK666 treatment, actin polymerization still significantly increased, indicating that RacE activates actin polymerization independently of Arp2/3 (Fig. S8d–f).

a Scanning confocal imaging of RacE-GEF (mCherry-FRB-GXCTΔNT) membrane recruitment and cell shape in wild type (AX3) Dictyostelium cells. Cells are also expressing an unlabeled membrane-localized FKBP domain (cAR1-2xFKBP). t = 00:00 indicates rapamycin addition. b Temporal color projections of cell outlines corresponding to the movie in a. Blue and yellow times indicate the first and last images in the projection, respectively. c Average (line) and SEM (shaded area) of cell area in AX3 cells before and after GXCT recruitment (dashed line, t = 0). n = 35 cells. d Traces of cell movement in AX3 cells 200 seconds before and after GXCT recruitment. n = 35 cells. e Scanning Confocal imaging of GXCT recruitment and polymerizing actin (LimEΔCoil-EGFP) in AX3 cells. t = 00:00 indicates rapamycin addition. f Membrane kymograph of LimE from the movie in e. Dashed line indicates rapamycin addition, scale bars = 2 minutes (horizontal) and 5 µm (vertical). g Average and SEM membrane-to-cytosol ratio of LimE before and after GXCT recruitment by rapamycin addition (dashed line, t = 0). n = 8 cells. h Scanning Confocal imaging of GXCT recruitment and Ras activation (RBD-EGFP) in AX3 cells. t = 00:00 indicates rapamycin addition. i Membrane kymograph of RBD from the movie in h. Dashed line indicates rapamycin addition, scale bars = 2 minutes (horizontal) and 5 µm (vertical). j Average and SEM membrane-to-cytosol ratio of RBD before and after GXCT recruitment by rapamycin addition (dashed line, t = 0). n = 9 cells. Time is in min:sec. Scale bars = 5 µm unless otherwise noted. Source data are provided as a Source Data file.
To determine the effects of RacE activation on STEN, we observed changes in the localization of the active Ras biosensor, RBD, after GXCT recruitment. Before recruitment, cells displayed several large, intense, and long-lived RBD patches along their periphery. However, upon activating RacE, the RBD patches became smaller, weaker, and shorter-lived, with a potential increase in the total number of patches. The amount of Ras activation on the cell periphery dropped by 30% within 8 minutes (Fig. 4h–j, Supplementary Movie 11). This reduction in Ras activation after GXCT recruitment did not occur in cells treated with latrunculin (Fig. S8g–i). Taken together, these results suggest that RacE activation is able to increase actin polymerization in an Arp2/3-independent manner and inhibit STEN activation, most likely through the nucleation of linear actin networks and increased actin-actin crosslinking in the actomyosin cortex.
RacE reversibly and locally inhibits STEN and triggers Arp2/3-independent protrusions
To reversibly and locally control RacE activity, we designed an iLID-based optogenetic system68 to recruit GXCT to the membrane by applying blue light to cells. In this system, the GEF domain of GXCT was fused to a SSPB domain which binds to a membrane-localized iLid domain (see methods) when exposed to blue light. Similar to the CID system, uniform light stimuli led to rapid cell flattening and cessation of movement, which rapidly reversed upon removal of illumination (Fig. 5a–c, Supplementary Movie 12). We then examined how transiently increasing RacE activity affected propagating ventral RBD waves. Recruiting GXCT caused a sudden decrease in RBD wave activity, which was quickly reversed upon removal of blue light and dissociation of GXCT (Fig. 5d–f, Supplementary Movie 13).

a Scanning confocal imaging of RacE-GEF (tagRFP-SSPB-GXCTΔNT) optical membrane recruitment in wild type (AX3) Dictyostelium cells. Cells also express a membrane-localized iLID domain (N150-ILID). t = 00:00 indicates blue light exposure or blue light loss (parentheses). b Average (lines) and SEM (shaded area) of cell area before and after GXCT recruitment (left, dashed line) or dissociation (right, dashed line). n = 17 cells (left) and 6 cells (right). c Individual traces from (b). d TIRF imaging of Ras activation (RBD-emiRFP670) before, during, and after GXCT recruitment in electrofused (“giant”) AX3 cells. Cells are treated with 50 µg/ml Biliverdin to activate emiRFP670 fluorescence. t = 00:00 indicates blue light exposure. e Average and SEM of the percent of cell membrane with RBD recruitment before and after GXCT recruitment (left, dashed line) or GXCT dissociation (right, dashed line). n = 7 cells. f Individual traces from e. g Scanning confocal imaging of cell protrusion after recruitment of SSPB-GXCT or SSPB alone. +CK indicates cells treated with CK666. Boxes indicate the blue light area; arrows form a line between the center of the protrusion after stimulation and the center of the cell. t = 00:00 indicates the last timepoint before blue light exposure. h Angular histograms of the angle formed between the protrusion and GXCT recruitment relative to the cell center. n = 26 cells, SSPB-GXCT and SSPB-GXCT + CK666. n = 15 cells, SSPB. i TIRF imaging of RBD localization before and after local GXCT recruitment in giant AX3 cells. The yellow box indicates blue light area and the blue arrow is the location of the linear kymograph. t = 00:00 indicates the last timepoint before blue light exposure. j Linear kymograph of GXCT recruitment and RBD intensity from i. Yellow box indicates blue light area. k Quantification of RBD wave motion away from GXCT recruitment (see methods). This measurement is a composite of RBD translocation and disappearance. Kymograph scale bars = 20 seconds (horizontal) and 2 µm (vertical). n=cells, ***=p = 6.31*10−5, 2-sided Mann-Whitney U test. Time is in min:sec; scale bars = 5 µm unless otherwise noted. Source data are provided as a Source Data file.
Next, we took advantage of our optogenetic system to assess the morphological and signaling effects of local increases in RacE activity. Interestingly, despite the ability of RacE to turn off Ras activity, the localized recruitment of GXCT induced the formation of protrusions expanding from the recruitment site towards the source of the blue light. These protrusions were observed to form even in the presence of CK666, suggesting that RacE-triggered actin polymerization can occur independently of Arp 2/3 (Fig. 5g, h, Supplementary Movie 14). However, the lifetime and length of these protrusions is lower than typical chemoattractant-induced pseudopods. Moreover, recruiting GXCT to specific regions on the ventral surface of the cell led to the local extinguishment or redirection of propagating RBD waves (Fig. 5i–k, Supplementary Movie 15).
Incorporating cytoskeletal feedback into a core model of the STEN
Our results suggest the presence of two different cytoskeletal feedback loops. To determine how these loops could affect excitability and cell polarity, we turned to a computational model. The core of the model is an activator-inhibitor system (Fig. 6a) in which the activator positive feedback is implemented as double negative feedback between Ras and anionic lipids, including PIP2, and the slow inhibition is between Ras and PKB25. Simulations of this core system show occasional firings randomly distributed around a cell perimeter (Fig. 6b). These elevated regions of activity propagate for several seconds before they extinguish. The complementary pattern between Ras and PIP2 was used as a readout.

a Schematic showing the signal transduction network involving Ras/ PIP2 and PKB and how they couple to the two types of actin feedback loops. b Kymographs showing activity around the cell perimeter as a function of time. PIP2 and Ras are shown in green and magenta, respectively. The top and bottom correspond to simulations without and with the feedback loops, respectively. c Simulated cell trajectories of 10 cells each with feedback loops turned off (top) and on (bottom). d Kymographs showing wave activity across the cell perimeter for varying strengths of the branched actin feedback. e Total Ras activity around the cell perimeter with respect to the strength of the branched actin feedback. Wildtype corresponds to a strength of 1. The black bar denotes the mean of 10 simulations per strength. Simulations with total Ras activity less than 500 showed no firings. f Kymographs showing wave activity across the cell perimeter for varying strengths of the actomyosin feedback. g Total Ras activity around the cell perimeter with respect to the strength of the actomyosin feedback. Wildtype corresponds to a strength of 0.4. The black bar denotes the mean of 10 simulations per strength. Simulations with total Ras activity less than 200 showed no firings. h Frames from a 2D simulation of the effects of adding CK666 and then Latrunculin to wild-type cells. The three rows represent waves in the wild-type cell, waves after CK666 addition, and subsequent Latrunculin treatment. i The total Ras activity for simulations as in h. The solid line and the shaded area represent the mean ± 1 standard deviation. In all the simulations, the CK666 effect is incorporated at the 200 s and the additional Latrunculin effect is added at 400 s. Scale bars = 10 microns. Source data are provided as a Source Data file.
To the basic network, we add two complementary feedbacks (Black lines in Fig. 6a). The first implements the feedback between branched actin and Ras at the cell front while the second implements feedback between actomyosin and Ras at the rear. While previous models contained local positive feedback from branched actin, most of those systems also incorporated negative cytoskeletal feedback as a global inhibitor generated by Arp2/3-nucleated actin25,26,33,69,70. In contrast, negative feedback in this model is generated at the cell back and decreases at the site of activation. This assumption, based on the data in this paper, distinguishes this model from our previous models as well as most other polarity models. Details of the mathematical model are given in the Methods. Simulations of the system with added cytoskeletal feedback have the same complementarity as before, but Ras shows increased persistence, characterized by localized long-lived regions of elevated Ras with minimal lateral propagation (Fig. 6b).
To illustrate the effect of these feedbacks on cell movement, we first plotted the trajectories of simulated cells (see Methods) migrating randomly in the absence of branched actin and actomyosin feedback (Fig. 6c, top) and compared it to those in the presence of these loops (Fig. 6c, bottom). Cells without feedback remained near their starting point, while the addition of feedback enabled cells to move farther away. To characterize the effect of the two feedbacks separately, we altered the strength of each individually while holding the other constant and quantified the total Ras activity. Increases in branched actin feedback led to elevated Ras-GTP levels, greater persistence, and wider patches (Fig. 6d, e). Increases in actomyosin feedback had the opposite effect (Fig. 6f, g).
Finally, we carried out simulations in a two-dimensional surface to recapitulate ventral waves and recreated the experiments of Fig. 2c, d. Initially, waves are seen, but after simulating the effect of CK666 addition through the removal of the branched actin feedback, they disappear. The subsequent removal of actomyosin feedback, mimicking latrunculin addition, leads to a partial recovery of the Ras waves (Fig. 6h; Supplementary Movie 17). Quantitating the effect over multiple simulations shows that total Ras activity decreases greatly when CK666 is added and partial recovery upon latrunculin treatment (Fig. 6i).
The improved model predicts migratory behavior of cells with altered cytoskeletal activity
One prediction of this model is that the effect of altering the strength of cytoskeletal feedback on cell trajectory depends on initial feedback strength. To Illustrate this, we simulated the effect of raising positive feedback or lowering negative feedback where the initial setpoint corresponded to higher or lower polarity. At a low polarity starting point, raising the strength of branched actin feedback or lowering the strength of myosin feedback increased the distance covered by trajectories (Fig. 7a, b). However, the same changes decreased the area covered by trajectories of highly polarized cells by making fronts too large to maintain front-rear polarity (Fig. 7c, d). It is worth noting that a loss of polarity from increased branched actin activity would not be predicted in a global inhibitor model, as increased actin nucleation also increases global inhibitor production.

a, b 10 simulated tracks of cell motion before (left) and after (right) increasing the strength of the branched actin loop (a) or decreasing the strength of the actomyosin loop (b). Cells begin with low feedback strength for both branched actin and actomyosin, creating low polarity. 10 simulated tracks of cell motion before (left) and after (right) increasing the strength of the branched actin loop (c) or decreasing the strength of the actomyosin loop (d). Cells begin with high feedback strength for both branched actin and actomyosin, creating high polarity. e Tracks of vegetative AX3 and abnABC− cells migrating for 10 minutes. f Tracks of vegetative AX3 cells expressing mCherry-FRB-MHCKC and an unlabeled membrane-localized FKBP domain (cAR1-2xFKBP) migrating for 10 minutes before rapamycin addition to recruit MHCKC and for 10 minutes after 10 minutes of incubation in rapamycin. g Tracks of developed, polarized AX3 and abnABC− cells migrating for 10 minutes. h Tracks of developed, polarized AX3 cells expressing mCherry-FRB-MHCKC and an unlabeled membrane-localized FKBP domain (cAR1-2xFKBP) migrating for 10 minutes without rapamycin or after 10 minutes of incubation in rapamycin. i Average (lines) and individual (dots) estimated 2D diffusion coefficients of cells in each indicated condition from E-F. All cells are vegetative. j Average and individual estimated 2D diffusion coefficients of cells in each indicated condition from G-H. All cells are developed. *=p = 0.041, **=p = 0.0057, ***=p < 0.005, 2-sided Mann-Whitney U test. Source data are provided as a Source Data file.
To test whether dependence on initial conditions occurs in real cells, we decided to quantify cell motility in either vegetative or developed Dictyostelium cells, which have very different initial polarity setpoints. In vegetative cells, increasing branched actin feedback strength by knocking out actobindins made many cells move in a keratocyte-like manner which led to an increase both in speed and persistence (Fig. 7e). Similarly, lowering the strength of myosin feedback by recruiting MHCKC led to a smaller but significant speed increase (Fig. 7f). Recruiting the FRB domain alone had no significant effect on migration (Fig. S9a–c). Repeating these same perturbations in developed cells produced different results: both actobindin-null cells and cells with MHCKC recruited migrated slower than controls (Fig. 7g, h). This is due to an increase in Ras activation and a resulting decrease in polarity (Fig. S9d, Movie S17). Analyzing the estimated diffusion coefficients of cells further substantiated the effects of changing feedback strength on speed and directionality (Figs. 7i, j, S9e–h).
Discussion
Much previous work has focused on control of the cytoskeleton by Ras/PI3K signal transduction networks, particularly in the context of chemotaxis, and many studies have focused on the reaction of cytoskeletal elements to mechanical forces. However, feedback from the cytoskeleton to “upstream” signal transduction events has only occasionally been considered. Using synthetic biological techniques, we have uncovered two crucial feedback mechanisms from different actin networks that characterize the front and back regions of the cell cortex. A front-promoting mechanism is supported by the observation that increases in Arp2/3-nucleated actin strongly boost Ras/PI3K activation, while reduction of F-actin overall decreases the activation. A back-promoting mechanism is indicated by multiple observations. First, acute reduction of myosin assembly enhances Ras/PI3K activation and heightens sensitivity to chemotactic stimuli, indicating that the actomyosin cortex suppresses Ras activation. Second, selective inhibition of branched actin nucleation nearly eliminates Ras activity; this inhibition is mitigated by depolymerizing the bulk of actin and decreasing myosin assembly, as well as in cells lacking myosin. Collectively, these observations indicate that the inhibition depends on the cortical actomyosin network. Third, recruitment of RacE GEF, which promotes actin crosslinking and formin nucleation, substantially reduces Ras/PI3K activation and yet can still induce protrusions. This indicates that cells can suppress Ras/PI3K signaling locally or globally by increasing the abundance of the actomyosin cortex.
When taken together with interactions discovered previously, the mechanisms we have delineated create two positive feedback loops between signal transduction and cytoskeletal networks. A front-promoting loop becomes clear by merging earlier findings that Ras/PI3K activation promotes Arp2/3 nucleation with observations here and elsewhere that branched actin activates these signal transduction networks71. A back-promoting loop emerges by bringing together our studies here that myosin assembly and strengthening the actomyosin cortex inhibits Ras/PI3K activation with prior work that Ras/PI3K activation leads to activation of myosin heavy chain kinase and consequent disassembly of myosin16,37. In human neutrophil-like cells, Ras has been found to reverse polarity through its downstream target AKT1, indicating a similar relationship may exist11. The reciprocal interactions between Ras/PI3K signal transduction and the branched actin and actomyosin cytoskeletal networks further highlight the crosstalk between these systems. These interconnected networks provide a molecular scheme for generating cell polarity and for integrating the chemical and mechanical cues that a cell would encounter in navigating a complex environment.
Previous investigators have speculated that polarity is established by multiple feedback loops, but these findings differ from ours in several ways. First, we show here that the actomyosin cortex is a direct negative regulator of spontaneous Ras/PI3K signaling, even in cells without Arp2/3 activity. Second, other models focused on direct feedback between different cytoskeletal regulators such as Rac and Rho, or competition between F-actin networks, and did not consider a role for upstream signaling events like Ras/PI3K activation46,72. For the sake of simplicity, we have not included direct interactions between cytoskeletal populations in our model. However, these possibilities are not inconsistent and may cooperate. Third, while many other models portray negative feedback as a global inhibitor created at the front, in ours a local inhibitor is generated at the cell back. Our model correctly predicts that this negative regulator alters polarity differently depending on the initial setpoint of the cell. However, it does not fully account for polarity due to a lack of communication between the front and the back.
The mechanism of negative feedback from the actomyosin cortex to signaling is not yet clear, but there are several attractive possibilities. One idea is that the integrity of the underlying cortex influences the fluidity of the membrane. Lower fluidity in regions with high cortical density may slow the diffusion of signaling activators, locally lowering excitability. Supporting this, multiple studies have reported lower membrane fluidity in back regions where actomyosin density should be high73,74,75. However, our previous studies have reported that myosin II promotes cortical fluidity in Dictyostelium and mammalian cells76,77. Thus, an equally likely possibility is that myosin II may reduce the cortical lifetime of front signaling activators (e.g. Ras GEFs or PI3K). Supporting this, Myosin has been found to reduce the binding lifetime of other cortical proteins78. Another possibility is related to the finding that anionic lipids such as PI(4,5)P2 on the inner leaflet of the membrane inhibit Ras/PI3K signaling6. Electrostatic interactions between molecules in the actomyosin cortex and the lipids, such as through cortexillin and PI(4,5)P279, or the enzymes that regulate them, may contribute to the enrichment of these inhibitory lipids at the cell back. Recent work has indicated that actin and the membrane are more tightly associated at the cell back, indicating a stronger interaction between the membrane and the actomyosin cortex than branched actin75. Finally, force generation by the actomyosin cortex could alter cortical tension, a parameter that includes contributions from the membrane and the underlying cytoskeleton, or membrane curvature, leading to the recruitment or activation of negative regulators specifically at the cell back69,80,81,82,83,84.
A cell migrating through its natural environment towards a chemical cue encounters a complex mix of other cells, the extracellular matrix, and externally imposed forces, such as shear stresses. These physical features are usually integrated with chemical cues to determine the optimal path towards a destination. One might imagine that the cell is like a self-driving car, following a GPS signal to a destination: if the car merely followed a pre-planned route, it would soon collide with an unmarked obstacle. To safely reach its goal, the car must use sensors to detect other cars or road work. Sensing biochemical signals using receptors is fairly straightforward, but the mechanisms cells use to sense mechanical information are diverse and complicated. Recent studies have suggested various physical properties, like nuclear stiffness and membrane curvature, may help steer cells in crowded environments83,85,86. Our findings, combined with long-standing evidence that physical forces alter the properties of the cytoskeleton, suggest an attractive hypothesis. These physical cues could locally alter the balance between actomyosin and branched actin. Indeed, myosin II itself is a key sensor of mechanical stresses along with other actin-crosslinking proteins such as filamins and alpha-actinins66,77. These proteins accumulate locally in response to mechanical stresses. As we have shown here, this accumulation of cortical components has quite significant effects on the same signaling nodes that are also controlled by chemical stimuli. Therefore, the core excitable system we and others have described likely serves as an integrator of diverse environmental signals to bring about successful navigation87.
Our results also have important implications for current and future therapeutic interventions. Currently, many cancer therapies target specific signaling molecules involved in cell growth and migration. Layers of redundancy and feedback from the local environment may compensate for inhibition at any single node. Targeting the cytoskeleton in combination with molecules like Ras and PI3K may be critical for preventing tumor growth and metastasis88,89,90.
Methods
Cells and plasmids
Dictyostelium discoideum cells were cultured in HL5 media91 for a maximum of 2 months after thawing from frozen stock. AX3 cells were obtained from the R. Kay laboratory (MRC Laboratory of Molecular Biology, UK), abnABC− cells were generated with homologous recombination previously in the Devreotes Lab. mhcA− cells were obtained from the Robinson Lab. Female human neutrophil-like HL-60 cells stably expressing RFP-PHAKT were previously created in the Devreotes lab11. Cells were cultured in RPMI medium 1640 with L-glutamine and 25 mM HEPES (Gibco; 22400-089) supplemented with 15% heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific; 16140071) and 1% penicillin-streptomycin (Thermo Fisher Scientific; 15140122). Cells were split at a density of 0.15 million cells/ml and every 3 days. 1.3% DMSO was added to cells at a density of 0.15 million cells/ml to trigger differentiation 5-7 days before imaging.
Plasmids were introduced into Dictyostelium cells using electroporation92. To improve efficiency, heat-killed Klebsiella aerogenes was added after transformation. In chemically-induced dimerization (CID) experiments (Figs. 1e–j, 3e–g, 4, 7f, g, S3, S4, S8, S9) all cells were transformed with an unlabeled cAR1-tandem FKBP (cAR1-FKBP-FKBP) in a pCV5 vector as well as an FRB-tagged protein in a separate pCV5 vector. cAR1 was selected because it is a uniform membrane protein93 and will therefore recruit FRB-tagged proteins to the membrane evenly upon rapamycin addition (Fig. 1e). mCherry-FRB-MHCKC was created using standard restriction digest cloning starting with a GFP-MHCKC construct from the Robinson lab. mCherry-FRB-GXCTΔNT was made by inserting the PH and RhoGEF (DH) domains of GXCT67 from a GFP-GXCT construct (gift from the M. Iijima lab, JHU) into a pCV5 plasmid containing mCherry and FRB. FRB controls used this initial construct. While both cAR1-2xFKBP and FRB vectors contain G418 resistance markers, a significant population of cells simultaneously transformed with both have significant FRB protein expression and membrane recruitment.
For optogenetic experiments (Fig. 5), cells were transformed with hygromycin-resistant vector pDM35894 containing an iLID68 domain linked to the myristoylation domain of PKBR1 (n150-ILID) which was previously shown to localize to the membrane3. Cells were subsequently transformed with SSPB(R73Q)-tagRFP-GXCTΔNT in a pCV5 vector (Fig. 5a–c, g, h) created by In-Fusion cloning or a dual-expression G418 resistance vector (O1N) containing mCherry-SSPB(R73Q)- GXCTΔNT and RBD-emiRFP670 (Fig. 5d–f, i–k) created by GoldenBraid cloning95.
pDM358 RBD-EGFP, based on the 51–220 amino acids of Raf41 (Figs. 1a, g–j, 2i–k, 3e–g, 4h–j, S3c, d, S4, S5g, h, S8g–i, S9d), LimEΔCoil-eGFP (Figs. 4e–h, S5d–f), pDM358 PHCRAC-YFP/LimEΔCoil-RFP (Fig. 2a, b), and pDM358 PHCRAC-YFP (Fig. 3a–d), and Arpin-EGFP (Fig. 1a–d) were previously created in the Devreotes lab3. RBD-mCherry (Fig. 1a–d, Arpin-eGFP cells) was assembled into a hygromycin resistance vector (A2H) using GoldenBraid assembly. PHCRAC-mScarlet3 (Fig. 3a–d) was assembled into a G418 resistance vector (A1N) using GoldenBraid assembly. arpB cDNA (Fig. S1a–c) was synthesized (Twist Bioscience, South San Francisco) and then inserted into a pCV5 vector using In-Fusion cloning. Hygromycin-resistant vector pDRH eGFP-Myosin II (Figs. 3a–d, S2a, b), pDRH-Cortexillin I (Fig. S5a–f), and pDRH-Dynacortin (Fig. S8a–c) were previously created in the Robinson lab. pDM358 eGFP-HSPC300 (Fig. S1d, e) was a gift from the C. Huang lab (JHU).
Microscopy
For Dictyostelium imaging experiments, cells were seeded in 8-well Lab-Tek chambers (Thermo-Fisher, 155409) and left to settle for ten minutes before the media was gently aspirated and replaced with Development Buffer (DB) (5 mM NA2HPO4, 5 mM KH2PO4, 1 mM CaCl2, 2 mM MgCl2). In experiments with non-electrofused vegetative cells, cells were allowed to sit for an hour starving in DB prior to imaging to decrease photosensitivity. For experiments requiring cAMP stimulation (Fig. S4, 2i–k, S6), growth-phase cells were washed and suspended in DB at a density of 2 × 107 cells/ml. Cells were then developed by being shaken for 1 hour and subsequently pulsed with 50–100 nM cAMP every 6 minutes and shaken for 4 hours.
For dHL60 cells, cells were seeded in Lab-Tek 8-well chambers coated with fibronectin at approximately 35 µg/cm and allowed to adhere for one hour prior to imaging. Directly prior to imaging, cells were treated with 200 nM N-Formyl-Met-Leu-Phe (FMLP) to encourage migration.
Laser scanning confocal imaging was carried out on two microscopes: a Zeiss AxioObserver inverted microscope with an LSM800 confocal module and a Zeiss AxioObserver with 880-Quasar confocal module & Airyscan FAST module. On the LSM800 microscope, GFP and YFP proteins were excited with a solid-state 488 nm laser and on the 880 with an argon laser. mCherry and other red fluorescent proteins were excited with a solid-state 561 nm laser on both systems. Emission wavelengths collected were chosen to avoid overlap between GFP and mCherry emission profiles. All imaging was done with 63X/1.4 PlanApo oil DIC objectives and appropriate raster zoom. Brightfield images acquired using a transmitted-photomultiplier tube (T-PMT) detector.
Spinning Disk confocal imaging was performed on a Nikon TiE2 CSU W1 SoRA microscope, solid-state 488 and 561 nm lasers, a 60x/1.49 Apo TIRF objective and a Hamamatsu FusionBT camera. Camera binning was adjusted to optimize for signal-to-noise, resolution, and speed of acquisition. All images for a given experiment were acquired at the same binning.
For Total Internal Reflection Fluorescence (TIRF) imaging, experiments measuring the response of RBD to CK666 and latrunculin (Fig. 2c–e) were performed on a Nikon TiE microscope with a solid state 561 nM laser for mCherry excitation, a 100x/1.49 Apo TIRF objective, an RFP emission filter set, and a Photometrics Evolve 512 EMCCD camera. Experiments measuring the response of RBD to RacE GEF recruitment and RBD wave response to latrunculin alone (Figs. 5d–f, i–k, S5g, h) were performed on a Nikon Ti2-E microscope with an iLas2 Ring-TIRF and optogenetics module (GATACA Systems). Red fluorescent proteins were excited with a 561 nM solid-state laser and far-red proteins were excited with a 647 nm solid-state laser. Global and local recruitment were accomplished using a 488 nM solid-state laser connected to the iLas2 system. Cells were imaged using a 60x/1.49 Apo TIRF objective and a Hamamatsu FusionBT camera.
Cell fusion
Protocol adapted from (Miao et al., 2019).Growth‐phase cells were washed twice with and then resuspended in SB (17 mM Sorensen buffer containing 15 mM KH2PO4 and 2 mM Na2HPO4, pH 6.0) at a density of 1.5 × 107 cells/ml. 3 ml of cells were put into a 15‐ml conical tube and rolled gently for ~ 30 min, to promote cell clustering. 800 μl of rolled cells was transferred to a 4‐mm‐gap electroporation cuvette (Bio-Rad, 1652081), using pipette tips whose edges were cut off to avoid breaking clusters. Cells were then electroporated using the following settings: 1,000 V, 1 μF three times, with 1–2 s between pulses. Then, 30 μl of cells was transferred to the center of a well in an 8‐well chamber and was left still for 5 min. 370 (Fig. 2c–e) or 470 (Fig. 5d–f, i, j) μl of SB containing 2 mM CaCl2 and 2 mM MgCl2 was added to the well and was pipetted briefly to suspend the cells evenly. After allowing cells to settle for 10 minutes, all media together with excess floating cells were removed and 450 μl of new SB plus 2 mM CaCl2 and 2 mM MgCl2 was gently added to the well against the wall. Cells were then allowed to recover for 1 h before imaging. 50 µg/ml Biliverdin was added to cells expressing RBD-emiRFP670 (Fig. 5d–f, i, j) during the rolling stage to activate emiRFP970 fluorescence.
Preparation of Reagents and inhibitors
Rapamycin (Millipore Sigma, 553210) was dissolved in DMSO to a concentration of 10 mM. Then, 1 µl aliquots were diluted 1:200 in the imaging buffer of the experiment (DB or SB plus 2 mM CaCl2 and 2 mM MgCl2) to a 10x concentration (50 µM). CK666 (Millipore Sigma, 182515) was dissolved in DMSO to a concentration of 100 mM. Then, 0.5 µl aliquots were diluted 1:100 in warm imaging buffer of the experiment (DB, SB plus 2 mM CaCl2 and 2 mM MgCl2, or RPMI) to a 10x concentration (1 mM). 2.5 µl aliquots of 1 mM latrunculin in DMSO (Millipore Sigma, 428026) were diluted 1:20 in the imaging buffer of the experiment (DB, SB plus 2 mM CaCl2 and 2 mM MgCl2, or RPMI). cAMP (Millipore-Sigma, A6885) was dissolved to 1 mM in water and then diluted to a 10x working concentration (1 µM or 10 nM) in DB. Caffeine (Millipore-Sigma, 1085003) was dissolved to 1 M in water and then diluted to a 10x concentration (30 µM) in SB plus 2 mM CaCl2 and 2 mM MgCl2. FMLP was dissolved in DMSO to a concentration of 10 mM, and then individual aliquots were diluted in RPMI to a 10X concentration (2 µM). FM4-64 (Thermo Fisher, T13320) was diluted in DB to a 10X working concentration of 50 µg/ml and then added to cells for final concentration of 5 µg/ml. Cells were then incubated for two minutes and immediately imaged.
Chemically induced dimerization and inhibitor experiments
When imaging FRB protein recruitment without any additional inhibitors (Figs. 1e–j, 4, 7f–j, S3, S8a–c, S9) 50 µl of 50 µM rapamycin was added to a well containing 450 µl of buffer at the indicated time for a final concentration of 5 µM. Similarly, in experiments with CK666 alone (Figs. 2a, b, S5a, b, 3a–d) 50 µl of 1 mM CK666 was added to a well containing 450 µl of buffer at the indicated time for a final concentration of 100 µM. For experiments with latrunculin alone (Fig S5i, j) 50 µl of 50 µM latrunculin was added to a well containing 450 µl of buffer at the indicated time for a final concentration of 5 µM. For experiments with rapamycin and CK666 or latrunculin (Figs. 3e–g, S8d–i) or CK666 and latrunculin (Fig. 2c–g) 50 µl of each of the drugs was added to a well containing 400 µl of buffer at the indicated times for a final concentration of 5 µM Rapamycin, 100 µM CK666, and 5 µM latrunculin. In Fig. 2c–e, cells were pre-incubated with 3 mM caffeine and all drugs were diluted in buffer containing 3 mM Caffeine.
For cAMP stimulation experiments with 2 stimulations (Fig. S4), cells were pre-treated in 400 µl DB containing rapamycin, CK666, or CK666 and latrunculin at the concentrations indicated above for 30 minutes. Then, 50 µl of 10 nM cAMP was added to the well during imaging followed at least a minute later by 50 µl of 1 µM cAMP for final concentrations of 1 nM and 100 nM, respectively. The + rap population was treated with 5 µM rapamycin at least 30 minutes prior to imaging.
For cAMP stimulation experiments with 3 stimulations (Figs. 2i–k, S6), cells were left in 300 µl DB with nothing, CK666, or latrunculin for 10 minutes prior to imaging. A pre-calculated amount of cAMP was added to well 2 or 3 separate times, with at least 3 minutes between each stimulation. The volume and concentration of the cAMP added were adjusted to account for changes in volume and cAMP concentration between steps. For cells treated with CK666 or latrunculin, the cAMP solution also contained 100 µM CK666 or 5 µM latrunculin, respectively.
Optogenetics
In Fig. 5a–c and g, h, experiments were performed on the Zeiss 800 LSM. For global activation (Fig. 5a–c), the 488 laser was switched on and scanned across the entire field of imaging for the indicated time. For local activation (Fig. 5h, g), the microscope’s bleaching tool was used to scan the 488 laser in an approximately 1 µm square 1 µm away from the cell edge.
In Fig. 5d–f and i–k, experiments were performed on the iLas TIRF system. For global activation (Fig. 5d–f), the cells were exposed to 488 laser light at every time point for the indicated time. For local activation (Fig. 5i–k), the microscope’s targeted illumination tool was used to scan the 488 laser in an approximately 300 µm square on the cell membrane at the indicated location.
Image analysis
In order to quantify biosensor intensity on the membrane in confocal images, single cells were selected for analysis using cell health, biosensor expression, and FRB/SSPB protein recruitment (if applicable). Cells were then tracked manually using the Manual Tracking plugin in FIJI96. After manual tracking, masks of individual cells were created in MATLAB (Mathworks). Background was subtracted from the image by applying a 2D Gaussian blur filter with a high σ value (100) to the original image and then subtracting the resulting image from the original. Then, a threshold intensity value was calculated on the background-subtracted image using Otsu’s method97 and applied to a smoothed version of the background-subtracted image created using a 2D Gaussian blur filter with a low σ value (2). Masks were then modified by performing a morphological closing operation followed by a hole-filling operation. The selected cell was then identified by comparing the centroid of every object in the binary mask to the manual tracking coordinates and removing every object but the object closest to the tracking coordinate. Finally, a watershed filter was applied to remove cells which transiently collided with the selected cell. If the selected cell could not be satisfactorily separated from neighbors it was discarded in the dataset. All shape and motion descriptors were created using the properties extract from this centroid using Matlab’s regionprops function. For area calculation (Fig. 1g), all segmentation was done on the RBD channel to avoid artifacts from the shifting localization of FRB proteins.
These binary masks were then used to create kymographs of membrane intensity over time. The boundary of the binary mask was identified and then smoothed using a moving average filter. Then, the membrane intensity (IM) at equally spaced points along the membrane was determined by taking a 3 µm-long, 1 pixel-wide linescan perpendicular to the boundary and finding the maximum intensity along that boundary. IM was equal to the sum of that brightest pixel along and any pixel 250 nm in either direction of the pixel along the linescan. The number of pixels (npix) summed was saved for later. Then, cytosolic intensity (IC) was calculated by performing a binary erosion to original image mask to remove the membrane and then taking the average intensity inside that eroded mask. A background intensity (Ibg) was acquired by manually selecting a 1 µm box outside of a cell and taking the average intensity in that box. Finally, the background-corrected membrane-to-cytosol ratio (M/C) at each point on the membrane was acquired using this formula: \(\frac{{I}_{M}-{I}_{{bg}}\,\times {n}_{{pix}}}{({I}_{C}-{I}_{{bg}})\times {n}_{{pix}}}\). This value was then used for kymograph display. To obtain the total cellular intensity, the sum of IM at each point and the total cytosolic intensity (\({I}_{{C}}\times {{Area}}_{C}\)) were background-corrected, added together, and then divided by the cell area.
To calculate the average M/C ratio at each timepoint, each line in a kymograph, excluding black space, was averaged. Similarly, intensity variance on the membrane (Fig. 2b) was measured on each line in a kymograph, excluding black space. For dim biosensors with variable expression levels, (Figs. 1c, j, S3c, d), the percentage of pixels in each line above an intensity threshold was calculated. This threshold was determined by hand-identifying 3 separate patches in the image and finding the value 10 percentile points lower than the average of those three pixels in the distribution of all kymograph intensities. To average over an entire movie (Fig. 1c), the intensity at each timepoint recorded was simply averaged. To create a plot of average intensity or area over time, all traces were synchronized to the specified perturbation and then all values were placed in time “bins” relative to 0 where 0 is the rightmost edge of a bin and then averaged. Bin width was determined by the rate of acquisition but was generally 30 seconds. In all cases except Fig. 2i–k, intensities were then normalized to the average intensity from −1 minute to 0. In Fig. 2i–k, intensity was normalized to the average value of all frames greater than 2 minutes away from a stimulus.
To calculate the integrated intensity of PHCRAC and LimEΔcoil (Fig. 2b, a smoothed image of a cell was created from an original image using a 2D gaussian blur filter with a low σ value (8). This blurred image was subtracted from the original image to highlight the bright spots in the cell. Then, a threshold intensity value was calculated on the background-subtracted image using Otsu’s method. The binary mask created from the threshold was then modified by a morphological closing operation and all shapes four pixels and smaller were removed. A background intensity was acquired by manually selecting a 1 µm box outside of a cell and taking the average intensity in that box. After subtracting the background value from the original image, the total patch intensity was acquired by summing the intensity of all pixels inside of the binary mask.
The 2D diffusion coefficients of migrating cells (Figs. 7i, j, S9C) was obtained by applying Scikit’s linear regression function to the MSD vs Δt curve of each cell. All data for Figs. 7 and S8 was acquired at a rate of 30 s/frame.
The calculate the membrane intensity in TIRF images (Figs. 2d, e, 5e, f, S1e, S3b, S5g, h), a mask of the entire cell was first created at every timepoint using Otsu’s method on a background-subtracted image similar to confocal images. For the average intensity (Figs. 2d, e, S2b, S5g, h), all pixels in the original image within this mask were averaged and then background-corrected by subtracting the average intensity of a 1 µm box outside the cell. For data with multiple timepoints (Figs. 2d, e, S5g, h) intensities were then normalized to the average intensity from −1 minute to 0 prior to CK666/latrunculin addition. To obtain the percentage of area occupied by a wave (Figs. S1e, 5e, f), a wave mask was created using a threshold calculated from only intensities inside the cell mask on a frame in the movie where there is a clear active wave. The percentage was calculated by dividing the area of the wave mask by the area of the cell mask and multiplying by 100. All cells were selected based on criteria of cell health, biosensor expression, SSPB recruitment (where applicable) and wave formation prior to perturbation.
To calculate the average biosensor patch number (Fig. 3b–d) the number of patches was identified manually at every timepoint. To be counted as a patch, the bright spot had to be larger than a single pixel and located on the cell periphery. The total number of patches over an interval (for example, all timepoints prior to CK666 addition) was then divided by the total time of that interval. Total cellular intensity for these cells (Fig. S7) was obtained by outlining each cell with the freehand drawing tool in imageJ and measuring the average intensity. The intensity was then background-corrected by subtracting the average intensity of a large square of noncellular background in the same movie.
To calculate the angle of protrusion formation relative to SSPB recruitment (Fig. 5h), in FIJI a line was drawn from the center of the cell to the center of the stimulation region and then another from the center of the cell to center of the first protrusion formed after stimulation. The angle between these two lines was calculated using each of their angles relative to the x axis of the image.
To calculate the RBD extinction from light (Fig. 5k), a linear kymograph was created by drawing a line through the region of light stimulation and through the wave targeted by the stimulation in FIJI and then using the reslice tool. The extinction rate is calculated by drawing one line along the border of the RBD region 30 s (10 timepoints) before stimulation and another along the border 30 s after. The height of this line (µm traveled) divided by the width (time) indicates the rate the RBD patch is displacing from the region of stimulation.
To calculate the cytoplasmic intensity of RBD in cAMP MHCKC expressing cells with or without rapamycin (Fig. S4b), the cytoplasm of a cell was identified manually using the freehand selection tool in FIJI. The intensity in this selection was averaged and then background-corrected by subtracting the average intensity of a 1 µm box outside the cell. Intensities were then normalized to the average intensity at 0.
To calculate the protein and western blot band intensity, The integrated intensity of each band was taken in FIJI using the same size rectangle for all bands. Intensities were then corrected by subtracting the integrated intensity of the background. To correct for total protein, western blot intensities were divided by the total amount of Ras in the initial lysate (Fig. S2) or the total protein intensity of the given sample (Fig. S5).
RBD pulldown assay
To biochemically determine the level of Ras activation in different cell lines with or without CK666 (Fig. S2), growth-phase AX3 and abnABC− cells were suspended then washed twice with nutrient-free DB. After washing and determining density via hemocytometer, 1.5 × 107 cells per reaction were separated, spun down, and resuspended in DB to a density of 1.5 × 107 cells/ml. At this point, 1 µl DMSO was added to allow the cells to adapt to DMSO addition. Once added, cells were starved and shaken for 1 hour.
After starvation, lysis and RBD pulldown were performed using a modified version of a Ras Pull-down Activation Assay Biochem Kit protocol (Cytoskeleton inc, #BK008-S). Cells were spun down at room temperature for 5 minutes at 500xG, the media was removed, and the pellet was resuspended in 1 ml DB + 1 µl DMSO (AX3-GTPγS, AX3-DB, abnABC−) or + 1 µl 100 mM CK666 (AX3-CK666) and incubated at room temperature for 5 minutes. After incubation, cells were spun down at room temperature for 5 minutes at 500xG, the media was removed, and then washed once in ice-cold DB. After resuspension, cells were spun down at 4 °C for 5 minutes at 500xG. To lyse the cells, media was removed and then replaced with 400 µl lysis buffer (50 mM Tris pH 7.5, 10 mM MgCl2. 0.5 M NaCl, and 2% IGEPAL) supplemented with a protease inhibitor cocktail (0.62 µg/ml Leupeptin, 0.62 µg/ml Pepstatin A, 0.14 mg/ml Benzamidine, and 0.12 ml/ml tosyl arginine methyl ester). After 3 minutes in lysis buffer, 30 µl per tube was removed for protein concentration determination and the rest was snap-frozen in liquid nitrogen and stored at −80 °C until use.
Protein concentration of the lysates was determined using a standard Bradford assay protocol. In short, protein standards ranging from 0–2 mg/ml BSA were created using 2 mg/ml BSA (Thermo Scientific, #23209) and Deionized water. 5 µl of each standard was added to wells on a 96-well plate (Millipore-Sigma, #M0812) in triplicate. For the unknown lysates, 5 µl of each lysate was added to wells undiluted, diluted 1:2, 1:5, 1:10, and 1:50 in duplicate. Finally, 250 µl of Coomassie Plus Protein Assay Reagent (Thermo Scientific, # 1856210) was added to each well and mixed. After incubating for 10 minutes, the A595 for each sample was determined using a FLUOstar Omega plate reader. Protein concentration of a given lysate was determined using the average of all A595 values within the linear range of the standard curve. All lysate concentrations fell between 3177-3251 µg/ml.
For a positive control for RBD-pulldown efficiency, AX3 lysate was loaded with GTPγS. AX3 lysate was rapidly thawed in room temperature water, and then 750 µg of lysate was mixed with lysate buffer to a final concentration of 750 µg/ml. The lysate was then mixed with loading buffer (15 mM EDTA) followed by GTPγS (200 µM). The tube was then shook at 37 °C for 30 minutes. During this incubation, all other lysates to be used were similarly thawed, diluted to 1 ml of a 750 µg/ml solution, and kept on ice for no more than 5 minutes prior to pulldown. 30 µg of each lysate was also saved to determine total Ras expression. At the end of the incubation, the GTPγS reaction was stopped by adding STOP buffer (60 mM MgCl2). All lysates were then immediately used for a pulldown reaction.
To probe for Ras activation, all lysates were mixed with 30 µl (100 µg) of Raf-RBD beads and gently rotated for 1 hour at 4 °C. Once done, beads were pelleted and then washed once in wash buffer (25 mM Tris pH 7.5, 30 mM MgCl2, 40 mM NaCl). After pelleting and removing supernatant, beads were resuspended in 20 µl 2X Laemmli sample buffer (125 mM Tris pH 6.8, 20% glycerol, 4% SDS, 0.005% Bromophenol blue, 5% beta-mercaptoethanol) and boiled at 98 °C for 2 minutes. After boiling, all tubes were spun down to pellet the beads and then all supernatant was loaded onto a gel for western blot analysis. The first time the experiment was run, 10 µl of 2 ng/µl recombinant His-Ras protein in 1X Laemmli sample buffer was loaded to confirm antibody function.
Cytoskeleton fractionation assay
To biochemically isolate assembled actin cytoskeleton from cells (Fig. S5c–h), a modified version of a well-characterized cytoskeletal sedimentation assay89 was used. AX3 cells with or without GFP-CortexillinI were grown on plates, washed once with DB, and then resuspended at a concentration of 1.5 × 106 cells/ml. 1 ml was then transferred to a 1.5 ml tube for each reaction. Cells were treated with 1 µl DMSO (-CK) or 1 µl 100 mM CK666, mixed by inverted, and then incubated gently rolling at room temperature for 5 minutes.
After incubation, cells were pelleted by centrifuging for 5 minutes at 400 x g and then resuspended in cold 1 mL MES starvation buffer (50 mM MES pH 6.8, 0.2 M CaCl2, 2 mM MgCl2). Cell were then pelleted by centrifuging for 5 minutes at 400 x g at 4 °C and resupended in 50 µl Buffer A (0.2 M MES pH 6.8, 2.5 mM EGTA, 5 mM MgCl2, 0.5 mM ATP) and incubated on ice for 5 minutes. Cells were then lysed by adding 50 µl of Buffer B (Buffer A + 1% Triton X-100, 100 µM PMSF, 150 µM TLCK, 1 µg/ml aprotinin, 5 µg/ml Leupeptin, 1 µg/ml Benzamidine, 2 µg/ml Pepstatin A, 100 µg/ml TAME, and 80 µg/ml TPCK) and incubating on ice for 5 minutes.
After removing 20 µl of total lysate and mixing with 20 µl 2x Laemmli sample buffer (2X SB), the samples were spun down at 10,000 x g for 2 minutes at 4 °C. The supernatant was then separated from the pellet and mixed with 80 µl 2X SB. The pellet was resuspended in 160 µl 1X SB. The pellet and supernatant tubes were then visually inspected to insure volumes were equal. Any small differences were adjusted by adding 1X SB to the tube with less buffer. All tubes were then boiled at 95 °C for 5 minutes. After boiling, 15 µl of each lysate per gel was loaded for western blot and Coomassie anaylsis.
Immunoblotting
Proteins were loaded into wells of pre-cast 4–15% or 12% polyacrylamide gels (Bio-Rad; #5671085 and #5671043) at 100 V for 1.5 hours. For total protein stain (Fig. S5c), gels were washed once in H20 and then incubated for 1 hour in Coomassie Protein assay solution (Thermo Scientific, #1856209). Gels were then destained in water overnight.
Gels were transferred onto nitrocellulose membranes (Thermo Scientific; 77010) for 10 mins using a Trans-Blot Turbo semi-dry transfer apparatus (Bio-Rad; 1704150EDU). For RBD pulldown (Fig. S2) the membrane was stained for total protein using Ponceau S solution (Sigma Aldrich, #P7170). Because of a large nonspecific band at 30 kD (see Fig. S2B) which interfered with staining, these membranes were trimmed at 25 kD. All membranes were then blocked using Intercept Blocking buffer (Li-Cor; #927–60001) in TBST (20 mM Tris, 150 mM NaCl, 0.1% Tween-20) for 1 hour. Membranes were incubated overnight at 4 °C with primary antibody (1:250 mouse anti-Pan Ras antibody Cytoskeleton Inc #AESA02 (Fig. S2), 1:10,000 mouse anti-GFP Invitrogen #33-2600 (Fig. S5f), 1:1000 anti-CortI Developmental Studies Hybridoma Bank # 241-438-1) in TBST (anti-Ras) or blocking buffer (all others).
Membranes were then washed 3 times for 5 minutes in TBST and then incubated in 1 hr in the dark with secondary antibody (1:10,000 dilution goat anti-mouse IRDye 800CW Li-Cor 925–32210). Near-infrared signal from the blots and protein gels was detected via Odyssey CLx imaging system (Li-Cor). The endogenous CortI immunoblots were then incubated in stripping buffer (200 mM Glycine, 3.5 mM SDS, 1% Tween 20, pH 2.2) for 10 minutes, washed once twice in PBS and twice in TBST, then incubated with primary antibody (1:200 anti- β-actin Santa Cruz Biotechnology #sc-47778) in blocking buffer at room temperature for 3 hours at room temperature and then in washed, stained with secondary, and imaged as previously described. Actin bands were distinguishable from cortexillin both in size and brightness.
Statistics and reproducibility
All statistical tests were performed in Python using the SciPy toolkit. Significance tests are two-sided Mann-Whitney U tests (for independent samples) and Wilcoxon rank-sum test (for the same cells before and after a treatment) as listed in the legends. NS = P > 0.05, * = P ≤ 0.05, ** = P ≤ 0.01, *** = P ≤ 0.005. Correlation tests are Spearman rank-order tests. Imaging data was drawn from movies with multiple cells analyzed in multiple stage positions. Imaging data are representative cells drawn from the dataset in their corresponding graph (e.g., Fig. 1h and j) For more information about the number of movies for each dataset, see the “Imaging data sample sizes” table in the source data file.
Simulations
The simulations are based on a model, previously described, in which three interacting species, RasGTP, PIP2, and PKB, form an excitable network25. The concentrations of each of these molecules is described by stochastic, reaction-diffusion partial differential equations:
In each of these equations, the final term represents the diffusion of the species, where \({D}_{*}\) is the respective diffusion coefficient and \({\nabla }^{2}\) is the spatial Laplacian (in one or two dimensions). The second-to-last terms represent the molecular noise. Our model assumes a Langevin approximation in which the size of the noise is based on the reaction terms98. For example, in the case of PKB, the noise is given by
where \(w(t)\) is a zero mean, unit variance Gaussian, white noise process. In the simulations, the size of this noise was adjusted with the empirical parameter \(\alpha .\)
In addition to the excitable dynamics described above, we incorporated two other terms. The first represents feedback from branched actin onto Ras. Since the cytoskeleton is not directly modeled, we model the origin of this feedback from PKB, which is upstream to the cytoskeleton25.
The second feedback loop accounts for the inhibitory regulation by actomyosin on Ras that originates from back molecules, such as PIP2.
Together, they modify the equation for Ras:
having the effect of increasing front and back contributions. In both equations, we include noise and diffusion terms.
Simulations were run on MATLAB 2023a (Mathworks) on custom-code based on the Ito solution in the Stochastic Differential Equation toolbox (http://sdetoolbox.sourceforge.net). Two-dimensional simulations were used to recreate the observed wave patterns of larger electrofused cells, and so assume a grid 40 µm × 4 µm with a spacing of 0.4 µm × 0.4 µm per grid point (i.e., 100 × 100 points) and zero flux boundary conditions. The one-dimensional simulations aim to recreate the membrane fluorescence observed in single-cell confocal images. The dimension is, therefore, smaller, assuming a cell radius of 5 µm and a spacing of 0.25 µm, resulting in \(2\pi \times 5/0.25\approx 126\) points along the perimeter, and periodic boundary conditions.
To determine the effect of these firings on cell movement, we follow an approach that simulates the movement of cells using a center-of-mass approximation. Using the spatially dependent level of activity, shown in the kymographs in Fig. 6b, we generated a series of force vectors normal to the cell surface. The vector sum of all these vectors was used to obtain a net protrusive force. After scaling this force so that it is in the range of experimentally observed protrusive pressures (0.5–5 nN/m2), we used it to push a viscoelastic model of Dictyostelium mechanics99. In this model, the net stress in the x-direction: σ (the direction of the gradient) alters the center-of-mass position (CM) through the following dynamics:
with a similar equation for the displacement in the y-direction (CM). Parameter values is this work and adopted from previous work100 are provided in Tables 1 and 2.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Raw microscope images available upon request. Source data are provided with this paper.
Code availability
Code used to create figures is available upon request. Model code (Fig. 6) available on Zenodo: 10.5281/zenodo.15800553.
References
Devreotes, P. N. et al. Excitable signal transduction networks in directed cell migration. Annu Rev. Cell Dev. Biol. 33, 103–125 (2017).
Graziano, B. R., Weiner, O. D., Huttenlocher, A. & Sahai, E. Self-organization of protrusions and polarity during eukaryotic chemotaxis This review comes from a themed issue on Cell adhesion and migration Edited by. Curr. Opin. Cell Biol. 30, 60–67 (2014).
Miao, Y. et al. Altering the threshold of an excitable signal transduction network changes cell migratory modes. Nat. Cell Biol. 19, 329–340 (2017).
Sasaki, A. T., Chun, C., Takeda, K. & Firtel, R. A. Localized Ras signaling at the leading edge regulates PI3K, cell polarity, and directional cell movement. J. Cell Biol. 167, 505–518 (2004).
Sasaki, A. T. et al. G protein–independent Ras/PI3K/F-actin circuit regulates basic cell motility. J. Cell Biol. 178, 185 (2007).
Banerjee, T. et al. Spatiotemporal dynamics of membrane surface charge regulates cell polarity and migration. Nat. Cell Biol. 24, 1499–1515 (2022).
Inoue, T. & Meyer, T. Synthetic Activation of Endogenous PI3K and Rac Identifies an AND-Gate Switch for Cell Polarization and Migration. https://doi.org/10.1371/journal.pone.0003068 (2008).
Weiner, O. D. et al. A PtdInsP3- and Rho GTPase-mediated positive feedback loop regulates neutrophil polarity. Nat. Cell Biol. 4, 509–512 (2002).
Huang, C. H., Tang, M., Shi, C., Iglesias, P. A. & Devreotes, P. N. An excitable signal integrator couples to an idling cytoskeletal oscillator to drive cell migration. Nat. Cell Biol. 15, 1307–1316 (2013).
Srinivasan, S. et al. Rac and Cdc42 play distinct roles in regulating PI(3,4,5)P3 and polarity during neutrophil chemotaxis. J. Cell Biol. 160, 375 (2003).
Pal, D. S., Banerjee, T. & Lin, Y. Actuation of single downstream nodes in growth factor network steers immune cell migration. Dev Cell 58, (2023).
Yoo, S. K. et al. Differential regulation of protrusion and polarity by PI3K during neutrophil motility in live zebrafish. Dev. Cell 18, 226–236 (2010).
Svitkina, T. M. & Borisy, G. G. Arp2/3 complex and actin depolymerizing factor/Cofilin in Dendritic organization and treadmilling of actin filament array in Lamellipodia. J. Cell Biol. 145, 1009–1026 (1999).
Theriot, J. & Mitchison, T. Actin microfilament dynamics in locomoting cells. Nature 1991 352, 126–131 (1991).
Weiner, O. D. et al. Hem-1 complexes are essential for rac activation, actin polymerization, and myosin regulation during neutrophil chemotaxis. PLoS Biol. 4, 186–199 (2006).
Jeon, T. J., Lee, D. J., Merlot, S., Weeks, G. & Firtel, R. A. Rap1 controls cell adhesion and cell motility through the regulation of myosin II. J. Cell Biol. 176, 1021 (2007).
Yumura, S. & Fukui, Y. Reversible cyclic AMP-dependent change in distribution of myosin thick filaments in Dictyostelium. Nature 1985 314, 194–196 (1985).
Cha, I. & Jeon, T. J. Dynamic localization of the actin-bundling protein Cortexillin I during cell migration. Mol. Cells 32, 281 (2011).
Sun, C. X. et al. Rac1 is the small GTPase responsible for regulating the neutrophil chemotaxis compass. Blood 104, 3758–3765 (2004).
Wang, F. et al. Lipid products of PI(3)Ks maintain persistent cell polarity and directed motility in neutrophils. Nat. Cell Biol. 4, 513–518 (2002).
Hadjitheodorou, A. et al. Leading edge competition promotes context-dependent responses to receptor inputs to resolve directional dilemmas in neutrophil migration. Cell Syst. 14, 196 (2023).
Lee, S., Shen, Z., Robinson, D. N., Briggs, S. & Firtel, R. A. Involvement of the cytoskeleton in controlling leading-edge function during chemotaxis. Mol. Biol. Cell 21, 1810–1824 (2010).
Hadjitheodorou, A. et al. Directional reorientation of migrating neutrophils is limited by suppression of receptor input signaling at the cell rear through myosin II activity. Nat. Commun. 12, 6619 (2021).
Zhan, H. et al. An excitable Ras/PI3K/ERK signaling network controls migration and oncogenic transformation in epithelial cells. Dev. Cell 54, 608 (2020).
Miao, Y. et al. Wave patterns organize cellular protrusions and control cortical dynamics. Mol. Syst. Biol. 15, e8585 (2019).
Weiner, O. D., Marganski, W. A., Wu, L. F., Altschuler, S. J. & Kirschner, M. W. An Actin-based wave generator organizes cell motility. PLoS Biol. 5, e221 (2007).
Schroth-Diez, B. et al. Propagating waves separate two states of actin organization in living cells. HFSP J. 3, 412–427 (2009).
Arai, Y. et al. Self-organization of the phosphatidylinositol lipids signaling system for random cell migration. Proc. Natl Acad. Sci. USA 107, 12399–12404 (2010).
Gerisch, G., Ecke, M., Wischnewski, D. & Schroth-Diez, B. Different modes of state transitions determine pattern in the Phosphatidylinositide-Actin system. BMC Cell Biol. 12, 1–16 (2011).
Asano, Y., Nagasaki, A. & Uyeda, T. Q. P. Correlated waves of actin filaments and PIP3 in Dictyostelium cells. Cell Motil. Cytoskelet. 65, 923–934 (2008).
Tang, M. et al. Evolutionarily conserved coupling of adaptive and excitable networks mediates eukaryotic chemotaxis. Nat. Commun. 5, 1–13 (2014).
Bhattacharya, S., Miao, Y., Devreotes, P. N. & Iglesias, P. A. A coupled excitable network model dictates cortical wave patterns and controls cellular protrusion morphology. Biophys. J. 116, 121a (2019).
Wu, Z., Su, M., Tong, C., Wu, M. & Liu, J. Membrane shape-mediated wave propagation of cortical protein dynamics. Nat. Commun. 9, 1–12 (2018).
Neilson, M. P. et al. Chemotaxis: A feedback-based computational model robustly predicts multiple aspects of real cell behaviour. PLoS Biol. 9, 1000618 (2011).
Meinhardt, H. Orientation of chemotactic cells and growth cones: models and mechanisms. J. Cell Sci. 112, 2867–2874 (1999).
Wang, M. J., Artemenko, Y., Cai, W. J., Iglesias, P. A. & Devreotes, P. N. The directional response of chemotactic cells depends on a balance between cytoskeletal architecture and the external gradient. Cell Rep. 9, 1110–1121 (2014).
Mondal, S. et al. Linking Ras to myosin function: RasGEF Q, a Dictyostelium exchange factor for RasB, affects myosin II functions. J. Cell Biol. 181, 747 (2008).
Artemenko, Y., Lampert, T. J. & Devreotes, P. N. Moving towards a paradigm: common mechanisms of chemotactic signaling in Dictyostelium and mammalian leukocytes. Cell. Mol. Life Sci. 71, 3711–3747 (2014).
Lampert, T. J. et al. Shear force-based genetic screen reveals negative regulators of cell adhesion and protrusive activity. Proc. Natl Acad. Sci. USA 114, E7727–E7736 (2017).
Hertzog, M., Yarmola, E. G., Didry, D., Bubb, M. R. & Carlier, M. F. Control of actin dynamics by proteins made of beta-thymosin repeats: the actobindin family. J. Biol. Chem. 277, 14786–14792 (2002).
Bondeva, T., Balla, A., Várnai, P. & Balla, T. Structural determinants of Ras-Raf interaction analyzed in live cells. Mol. Biol. Cell 13, 2323 (2002).
Wessels, D. et al. Cell motility and chemotaxis in Dictyostelium amebae lacking myosin heavy chain. Dev. Biol. 128, 164–177 (1988).
Lo, C. M. et al. Nonmuscle Myosin IIB is involved in the guidance of fibroplast migration. Mol. Biol. Cell 15, 982–989 (2004).
Lin, C. H., Espreafico, E. M. & Mooseker, M. S. Myosin drives retrograde F-actin flow in neuronal growth cones. Neuron 16, 769–782 (1996).
Ngo, K. X. et al. Allosteric regulation by cooperative conformational changes of actin filaments drives mutually exclusive binding with cofilin and myosin. Sci. Rep. 2016 6, 1–11 (2016).
Lomakin, A. J. et al. Competition for actin between two distinct F-actin networks defines a bistable switch for cell polarization. Nat. Cell Biol. 2015 17, 1435–1445 (2015).
Ravid, S. & Spudich, J. A. Myosin heavy chain kinase from developed Dictyostelium cells. J. Biol. Chem. 264, 15144–15150 (1989).
Yumura, S. et al. Multiple Myosin II heavy chain kinases: roles in filament assembly control and proper Cytokinesis in Dictyostelium. Mol. Biol. Cell 16, 4256 (2005).
Reichl, E. M. et al. Interactions between Myosin and Actin Crosslinkers Control Cytokinesis contractility dynamics and mechanics. Curr. Biol. 18, 471–480 (2008).
De Rooij, J. & Bos, J. L. Minimal Ras-binding domain of Raf1 can be used as an activation-specific probe for Ras. Oncogene 14, 623–625 (1997).
Nolen, B. J. et al. Characterization of two classes of small molecule inhibitors of Arp2/3 complex. Nature 460, 1031 (2009).
Bretschneider, T. et al. Dynamic actin patterns and Arp2/3 assembly at the substrate-attached surface of motile cells. Curr. Biol. 14, 1–10 (2004).
Simson, R. et al. Membrane bending modulus and adhesion energy of wild-type and mutant cells of dictyostelium lacking Talin or Cortexillins. Biophys. J. 74, 514–522 (1998).
Ren, Y. et al. Mechanosensing through cooperative interactions between Myosin II and the Actin Crosslinker Cortexillin I. Curr. Biol. 19, 1421–1428 (2009).
Faix, J. et al. Cortexillins, major determinants of cell shape and size, are actin-bundling proteins with a parallel coiled-coil tail. Cell 86, 631–642 (1996).
Weber, I. et al. Cytokinesis mediated through the recruitment of cortexillins into the cleavage furrow. EMBO J. 18, 586–594 (1999).
Kothari, P. et al. Contractility kits promote assembly of the mechanoresponsive cytoskeletal network. J. Cell Sci. 132, jcs226704 (2019).
Parent, C. A., Blacklock, B. J., Froehlich, W. M., Murphy, D. B. & Devreotes, P. N. G Protein signaling events are activated at the leading edge of chemotactic cells. Cell 95, 81–91 (1998).
Postma, M. et al. Uniform cAMP stimulation of Dictyostelium cells induces localized patches of signal transduction and pseudopodia. Mol. Biol. Cell 14, 5019 (2003).
Huang, Y. E. et al. Receptor-mediated regulation of PI3Ks confines PI(3,4,5)P3 to the leading edge of Chemotaxing cells. Mol. Biol. Cell 14, 1913 (2003).
Gerisch, G. et al. Membrane and actin reorganization in electropulse-induced cell fusion. J. Cell Sci. 126, 2069–2078 (2013).
Robinson, D. N. & Spudich, J. A. Dynacortin, a genetic link between equatorial contractility and global shape control discovered by library complementation of a Dictyostelium discoideum Cytokinesis mutant. J. Cell Biol. 150, 823 (2000).
Litschko, C. et al. Functional integrity of the contractile actin cortex is safeguarded by multiple Diaphanous-related formins. Proc. Natl Acad. Sci. USA 116, 3594–3603 (2019).
Zhou, Q. et al. 14-3-3 coordinates microtubules, Rac, and myosin II to control cell mechanics and cytokinesis. Curr. Biol. 20, 1881 (2010).
Gerald, N., Dai, J., Ting-Beall, H. P. & De Lozanne, A. A role for Dictyostelium RacE in Cortical tension and cleavage furrow progression. J. Cell Biol. 141, 483 (1998).
Luo, T., Mohan, K., Iglesias, P. A. & Robinson, D. N. Molecular mechanisms of cellular mechanosensing. Nat. Mater. 12, 1064–1071 (2013).
Wang, Y., Senoo, H., Sesaki, H. & Iijima, M. Rho GTPases orient directional sensing in chemotaxis. Proc. Natl Acad. Sci. USA 110, E4723 (2013).
Guntas, G. et al. Engineering an improved light-induced dimer (iLID) for controlling the localization and activity of signaling proteins. Proc. Natl Acad. Sci. USA 112, 112–117 (2015).
Houk, A. R. et al. Membrane tension maintains cell polarity by confining signals to the leading edge during neutrophil migration. https://doi.org/10.1016/j.cell.2011.10.050 (2012).
Van Haastert, P. J. M. Unified control of amoeboid pseudopod extension in multiple organisms by branched F-actin in the front and parallel F-actin/myosin in the cortex. PLoS One 15, e0243442 (2020).
Rickert, P., Weiner, O. D., Wang, F., Bourne, H. R. & Servant, G. Leukocytes navigate by compass: roles of PI3Kγ and its lipid products. Trends Cell Biol. 10, 466 (2000).
Zmurchok, C. & Holmes, W. R. Simple Rho GTPase Dynamics generate a complex regulatory landscape associated with cell shape. Biophys. J. 118, 1438–1454 (2020).
Banerjee, T. et al. A dynamic partitioning mechanism polarizes membrane protein distribution. Nat. Commun. 2023 14, 14 (2023).
Freeman, S. A. et al. Transmembrane pickets connect cyto-and pericellular-skeletons forming barriers to receptor engagement. Cell 172, 305 (2018).
Bisaria, A., Hayer, A., Garbett, D., Cohen, D. & Meyer, T. Membrane proximal F-actin restricts local membrane protrusions and directs cell migration. Science 368, 1205 (2020).
Girard, K. D., Kuo, S. C. & Robinson, D. H. Dictyostelium myosin II mechanochemistry promotes active behavior of the cortex on long time scales. Proc. Natl Acad. Sci. USA 103, 2103–2108 (2006).
Schiffhauer, E. S. S. et al. Mechanoaccumulative elements of the mammalian actin cytoskeleton. Curr. Biol. 26, 1473–1479 (2016).
Srivastava, V. & Robinson, D. N. Mechanical stress and network structure drive protein dynamics during cytokinesis. Curr. Biol. 25, 663–670 (2015).
Stock, A. et al. Domain analysis of cortexillin I: actin‐bundling, PIP2‐binding and the rescue of cytokinesis. EMBO J. 18, 5274–5284 (1999).
Diz-Muñoz, A. et al. Membrane tension acts through PLD2 and mTORC2 to Limit Actin network assembly during neutrophil migration. PLoS Biol. 14, e1002474 (2016).
De Belly, H. et al. Cell protrusions and contractions generate long-range membrane tension propagation. Cell 186, 3049–3061.e15 (2023).
Park, J. et al. Mechanochemical feedback underlies coexistence of qualitatively distinct Cell polarity patterns within diverse cell populations. Proc. Natl Acad. Sci. USA 114, E5750–E5759 (2017).
Sitarska, E. et al. Sensing their plasma membrane curvature allows migrating cells to circumvent obstacles. Nat. Commun. 14, 1–15 (2023).
Luo, T., Srivastava, V., Ren, Y. & Robinson, D. N. Mimicking the mechanical properties of the cell cortex by the self-assembly of an actin cortex in vesicles. Cit.: Appl. Phys. Lett. 104, 153701 (2014).
Renkawitz, J. et al. Nuclear positioning facilitates amoeboid migration along the path of least resistance. Nature 568, 546–550 (2019).
Brunetti, R. M. et al. WASP integrates substrate topology and cell polarity to guide neutrophil migration. J. Cell Biol. 221, e202104046 (2022).
Artemenko, Y., Axiotakis, L., Borleis, J., Iglesias, P. A. & Devreotes, P. N. Chemical and mechanical stimuli act on common signal transduction and cytoskeletal networks. Proc. Natl Acad. Sci. USA 113, E7500–E7509 (2016).
Bryan, D. S. et al. 4-Hydroxyacetophenone modulates the actomyosin cytoskeleton to reduce metastasis. Proc. Natl Acad. Sci. USA 117, 22423 (2020).
Surcel, A. et al. Pharmacological activation of myosin ii paralogs to correct cell mechanics defects. Proc. Natl Acad. Sci. USA 112, 1428–1433 (2015).
Surcel, A. et al. Targeting mechanoresponsive proteins in pancreatic cancer: 4-hydroxyacetophenone blocks dissemination and invasion by activating MYH14. Cancer Res. 79, 4665–4678 (2019).
Watts, D. J. & Ashworth, J. M. Growth of myxamoebae of the cellular slime mould Dictyostelium discoideum in axenic culture. Biochem. J. 119, 171 (1970).
Gaudet, P., Pilcher, K. E., Fey, P. & Chisholm, R. L. Transformation of Dictyostelium discoideum with plasmid DNA. Nat. Protoc. 2007 2, 1317–1324 (2007).
Xiao, Z., Zhang, N., Murphy, D. B. & Devreotes, P. N. Dynamic distribution of chemoattractant receptors in living cells during chemotaxis and persistent stimulation. J. Cell Biol. 139, 365–374 (1997).
Veltman, D. M., Akar, G., Bosgraaf, L. & Van Haastert, P. J. M. A new set of small, extrachromosomal expression vectors for Dictyostelium discoideum. Plasmid 61, 110–118 (2009).
Kundert, P. et al. A GoldenBraid cloning system for synthetic biology in social amoebae. Nucl Acids Res 48, 4139–4146 (2020).
Schindelin, J. et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Otsu, N. Threshold selection method from gray-level histograms. IEEE Trans. Syst. Man Cyber. SMC-9, 62–66 (1979).
Gillespie, D. T. The chemical Langevin equation. J. Chem. Phys. 113, 297–306 (2000).
Biswas, D., Bhattacharya, S. & Iglesias, P. A. Enhanced chemotaxis through spatially regulated absolute concentration robustness. Int. J. Robust. Nonlinear Control 33, 4923–4944 (2023).
Yang, L. et al. Modeling cellular deformations using the level set formalism. BMC Syst. Biol. 2, 68 (2008).
Acknowledgements
We thank members of the Peter Devreotes, Doug Robinson, Pablo Iglesias, and Miho Iijima labs for their helpful feedback and discussion. We thank the Iijima and Chuan-Hsiang Huang lab for supplying useful constructs. This work was supported by NIH grant R35 GM118177 (PND), DARPA HR0011-16-C-0139 (PAI/PND/DNR), AFOSR MURI FA95501610052 (PND), NIH grant GM66817 (DNR), and NIH grant S10 OD016374 (S. Kuo, JHU Microscope Facility). This manuscript is the result of funding in whole or in part by the National Institutes of Health (NIH). It is subject to the NIH Public Access Policy. Through acceptance of this federal funding, NIH has been given a right to make this manuscript publicly available in PubMed Central upon the Official Date of Publication, as defined by NIH.
Author information
Authors and Affiliations
Contributions
J.K., P.N.D., and D.N.R. conceived the experiments. JAK performed the experiments and most analysis of experimental data. A.H. and P.A.I. assisted with image analysis. P.B. and P.A. developed the computational model and performed simulations. The manuscript was primarily written by J.K. and P.N.D. with some initial writing generated by ChatGPT3 (OpenAI). Text pertaining to computational modeling was written by P.B. and P.A.I. D.N.R. assisted in editing the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Kuhn, J., Banerjee, P., Haye, A. et al. Complementary cytoskeletal feedback loops control signal transduction excitability and cell polarity. Nat Commun 16, 7482 (2025). https://doi.org/10.1038/s41467-025-62799-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-62799-3
This article is cited by
-
Establishing functional giant Dictyostelium cells reveals front–rear polarity in intracellular signaling
Communications Biology (2026)
