
Abstract
Vibrant feathers from Amazonian parrots were among the most prestigious symbols of status in the pre-Hispanic Andes, yet their acquisition required traversing the formidable Andean mountain range. Here, we investigate feathers recovered from an intact, elite masonry tomb at the coastal religious center of Pachacamac, associated with the pre-Inca Ychsma culture that flourished during the Late Intermediate Period (ca. 1000–1470 CE). By integrating ancient DNA, stable isotope analysis, and spatial modeling, we reconstruct the complete journey of these feathers. Genomic data identify four distinct Amazonian parrot species (Ara macao, A. ararauna, A. chloropterus, Amazona farinosa) and reveal high genetic diversity, indicating extraction from wild populations rather than local breeding. Stable carbon and nitrogen isotope analysis reveals that these birds consumed a C4-rich coastal diet, demonstrating they were transported alive across the Andes and kept in the arid coast. Finally, we identify specific trans-Andean corridors used for this exchange by combining species distribution models of ancient habitats with landscape resistance modeling. Our findings reveal a sophisticated, managed trade network operated by the Ychsma through intermediaries, challenging views of pre-Inca regionalism and establishing a multidisciplinary toolkit for tracing ancient exchange.
Similar content being viewed by others

Introduction
The vibrant feathers of Amazonian parrots were highly prized goods in the ancient cultures of coastal Peru, essential for communicating status, power, and cosmology1,2,3,4,5,6. The reliable acquisition of these prestige goods from east of the Andes was a critical factor in the political economy of prominent coastal cultures such as Moche and Nasca as well as highland empires of the Wari and Inca7. However, the scale, organization, and specific routes of the trade networks that moved these items across one of the world’s most formidable mountain ranges before the rise of the Inca remain largely speculative. Resolving this requires identifying the geographic origins of the birds used to produce the ancient featherworks.
There is ample evidence of the long and extensive use of parrot feathers among pre-Hispanic cultures of the Andes1,2,3, Mesoamerica8, and in the American Southwest9. Within ancient Peru, parrots were represented sculpturally and/or in painting in the pottery styles ranging from Moche (Mochica; ca. 200–850 CE) and Middle Sicán (Classic Lambayeque; 900–1100 CE) on the north coast, to Paracas (ca. 800–100 BCE) and Nasca (ca. 100 BCE–650 CE) on the south coast10,11. The direct indications of the important roles and values of parrots and their feathers are their preserved remains from well-documented archaeological contexts, especially as “feathered cloth”1,2,3,6. For making these ornaments, only less than 2% of all bird species in the Amazon region were used, most commonly parrots12.
In Andean scholarship, Amazonia is indeed often considered as a source of raw materials including bird feathers7. The rise of well-organized, late pre-Hispanic Arawak exchange/trade network (perhaps starting as early as 500 CE) to collect and distribute the desired Amazonian resources could have been critical for a reliable supply of the desired bird feathers out of Amazonia. Conservative estimates describe a trapping zone of ~100,000 km2 to supply millions of Amazonian birds to the highlands for the Wari and Inca empires7. Transporting goods a great distance by land and/or sea raises the question of the high costs involved. The rarity and high esteem of exotic feathers in funerary ornaments of the central coast of Peru hint to a high value that paid off transporting from a far.
Here, we investigate a unique collection of feathered ornaments recovered from an intact, elite Ychsma culture tomb (ca. 1000–1470 CE) at the paramount religious center of Pachacamac, on the central coast of Peru13,14,15. Excavated from the Sacred Precinct in front of the Painted Temple—the same area of Max Uhle’s pioneering work that defined the Andean chronological framework16,17,18,19—the tomb contained 34 funerary bundles with five large ones adorned with bundled brilliant parrot feathers. While parrots (Psittaciformes) are a diverse tropical order20,21, and many species are native to the Neotropics, visual identification of processed feathers is often limited to broad categories like macaw (mainly Ara sp.) or Amazon parrot (Amazona sp.), precluding precise species identification and provenance. This limitation has historically hindered our ability to reconstruct ancient trade dynamics.
To overcome this challenge, we developed a cross-disciplinary approach integrating archaeological data with advanced biomolecular, geochemical, and computational modeling techniques. While ancient DNA (aDNA) can be recovered from feathers22,23, its application to fragile archaeological artifacts has been rare. Using targeted enrichment and high-throughput sequencing optimized for degraded material24,25, we successfully extracted aDNA from feather barb fragments. We combined these genetic identifications with stable isotope analysis (δ¹³C, δ¹⁵N), AMS radiocarbon dating, and spatial modeling to pinpoint the taxonomic identity, diet, and geographic origin of the birds.
This study uses a multi-proxy approach to resolve the long-standing questions of provenance and procurement for these exotic feathers. We test the central hypothesis that the Ychsma, a regional coastal polity, sourced these feathers from wild Amazonian populations, a scenario that would require a carefully coordinated, long-distance trade network. By integrating ancient DNA for species identification, stable isotopes for dietary reconstruction, and species distribution and landscape modeling to map potential routes, we aim to provide direct evidence for the scale and nature of trans-Andean trade by a non-imperial society. This framework allows us to quantitatively test the economic and social connectivity between the Andes and Amazonia before the Inca Empire.
Results
An elite Ychsma tomb at a sacred coastal center
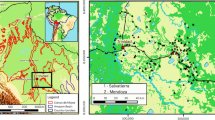
The famed religious-funerary center of Pachacamac, situated in the hyper-arid coastal desert of Peru—a region characterized by enduring aridity, punctuated by rains accompanying periodic El Niño Southern Oscillation (ENSO) events—presents a rare window into the sacred ritual practices of Andean societies. With tens of thousands of graves looted since the beginning of the Spanish colonial era and before the advent of scientific archaeology, the 2005 discovery of an intact Ychsma tomb in the Uhle Cemetery was an exceptional event (Fig. 1). The funerary bundles—varying in size and construction—were prepared to emulate the appearance of the deceased in their formal, high-status look26. The most important personages were interred in large bundles featuring a decorated “false head”, often accompanied by cinnabar-painted wooden masks and striking feather headdresses, readily recognizable markers of elite status on the pre-Inca coast27,28. These feather ornaments, the focus of our study, were attached to the back of the false heads (Fig. 1f).

a Map of Peru showing the Pachacamac Archaeological Site within the estimated territory of the Ychsma culture (gray shading) and our extended study area on the inlet map (orange shading). Map was created using QGIS v3.3 with ESRI World Topo as basemap. b Plan of the Pachacamac Sacred Precinct, with the location of Tomb 1/2 highlighted in red. c, d Drawings of the upper and lower layers of Tomb 1/2, respectively, showing the arrangement of funerary bundles. The highlighted bundles with red indicate the location of bundles mentioned in this study. e The 2005 excavation in progress in front of the Painted Temple (also known as Temple of Pachacamac). f Funerary Bundles B and C in situ in the lower level of the tomb. g A representative feather ornament from Bundle C, composed of feather bundles tied to cotton cords. Photos by I. Shimada.
To establish a firm chronology for these artifacts, we conducted accelerator mass spectrometry (AMS) ¹⁴C dating (see “Methods”). A series of feather samples from the ornaments yielded calibrated ages ranging from 1102 to 1457 CE (2σ; Supplementary Table 1). These dates, combined with previous dating of textiles and human remains from the tomb and associated ceramics, securely place the burials in the period between ca. 1000 to 1400 CE29. This timeline firmly attributes the tomb to the Ychsma, a confederation of politically autonomous chiefdoms that flourished on the central coast during the Late Intermediate Period (ca. 1000–1470 CE), after the collapse of the Wari Empire and before subjugation by the Inca30,31. The Ychsma established a distinct regional identity, with Pachacamac serving as their undisputed religious epicenter14,32.
While societies of this era are often defined by their conflicts and regionalism, there is growing evidence that they did not develop in isolation33. The presence of these vibrant feather ornaments—presumably made from tropical birds—suggests significant long-distance contact. However, identifying the species and geographic origin of these birds, and thus the trade routes they represent, is often impossible through visual or macroscopic analysis alone34. We therefore turned to archaeogenetics to unlock the history preserved within the feathers themselves.
Ancient DNA reveals a mix of local and Amazonian birds
To accurately determine the species used in the Pachacamac ornaments, we conducted genomic analysis on 25 ancient feathers sourced from eight different locations within the deposit (see “Methods” and Supplementary Data 1 for a detailed list of samples and their specific bundle provenance, whenever available). Working in the ancient DNA laboratory of the Australian Centre for Ancient DNA (ACAD, Adelaide University, Australia), we used targeted enrichment to sequence mitochondrial DNA from feather fragments (see “Methods”), which allows for robust identification even from degraded archaeological material35,36. The validated results show that the Ychsma utilized an avian assemblage sourced from two vastly different ecosystems. The colorful feathers belonged to four large parrot species (Fig. 2, Supplementary Figs. 1–7, and Supplementary Data 2–11): the Scarlet Macaw (Ara macao; haplogroup H4), Red-and-green Macaw (Ara chloropterus), Blue-and-yellow Macaw (Ara ararauna; haplogroup East = HE), and the Mealy Amazon (Amazona farinosa). In a single notable exception, one white feather was identified as a Sabine’s Gull (Xema sabini), a crow-sized seabird (Supplementary Figs. 1h and 8).

Maximum Likelihood tree based on mitochondrial genome sequences (15,455 bp), showing the placement of the six ancient feather samples from Pachacamac within the macaw genus Ara. The analysis confirms the identity of the samples as Ara ararauna, A. chloropterus, and A. macao (illustrated respectively from top to bottom, scaled relative to each other). The tree includes sequences from modern Peruvian Ara ararauna macaw samples (BYMA01, BYMA04) and other reference specimens (n = 48, including outgroup). Numbers at nodes indicate bootstrap support; circles denote 100% support. Samples from this study are highlighted. Illustrations [by Lluís Sanz] courtesy of Cornell Lab of Ornithology, from Birds of the World (https://doi.org/10.2173/bow).
Phylogenetic and haplotype analyzes provided further geographic context. The ancient macaw and Amazon parrot partial mitogenomes consistently clustered with subspecies and haplogroups found today in the eastern regions of Peru and broader South America37,38 (Fig. 3, and Supplementary Figs. 3–7). This genetic evidence creates a stark dichotomy in the origins of the birds. The Sabine’s Gull is an arctic breeder that winters along the Peruvian coast, making it a local resource for the Ychsma people within the rich Humboldt Current upwelling zone39. In sharp contrast, all four identified parrot species are native to lowland tropical rainforests and palm swamps located hundreds of kilometers to the east, on the other side of the Andes Mountains20. The unambiguous identification of this Amazonian avifauna confirms that the Ychsma society, centered on the arid coast, maintained extensive trans-Andes connections. This discovery raises a critical question regarding logistics: were these exotic birds bred locally, or were they procured from their distant, wild habitats?

a Map showing the modern distribution of the Scarlet Macaw (Ara macao)121 and the geographic locations of samples used in the network analysis (created by using QGIS v3.3 with ESRI World Topo as basemap). b Haplogroup network placing the three ancient Pachacamac samples (in black) firmly within Haplogroup 4. This haplogroup is associated with the widespread South American subspecies, A. m. macao. The network is based on mitochondrial gene sequences from 121 modern and ancient individuals, with colors corresponding to previously identified haplogroups37.
Genetic and isotopic data reveal wild origins and a coastal diet
The discovery of Amazonian parrots on the Peruvian coast begs the question of their procurement. One possibility is local captive breeding—a practice attested elsewhere in the pre-Hispanic Americas. At sites like Paquimé in Mexico40,41 and in the Mimbres region of the American Southwest42, archaeologists have found direct evidence of macaw breeding programs, including adobe cages, eggshells, and skeletons of captive birds dating to ca. 1100–1450 CE. Critically, these captive populations exhibit very low genetic diversity, a hallmark of inbred stocks43. To test this hypothesis at Pachacamac, we analyzed both the genetic diversity and the stable isotopic signatures of the ancient feathers.
The genetic evidence provides a clear verdict against local breeding. The mitochondrial genomes of the three ancient Scarlet Macaw feathers from Pachacamac were highly variable (Supplementary Table 2), defined by a high nucleotide diversity (π = 0.00223) for these individuals. The average nucleotide diversity (n = 31) reported for our H4 haplogroup in the wild is 0.00231 (SD = 0.00032), although using a smaller amount of mitochondrial data (~2200 bp)37. Furthermore, our value is also comparable to wild-sourced ancient macaws from other sites (Atacama Desert; π = 0.00272) and dramatically exceeding the near-zero (π = 0.00005) diversity found in the known pre-Hispanic captive breeding colonies in the American Southwest4,43. Based on this stark relative difference from captive and match with wild populations, the genetic pattern strongly indicates that the Pachacamac feathers were sourced not from a small, local breeding program, but from a large, wild Amazonian population.
While the genetics point to a wild origin, stable isotope analysis of carbon (δ¹³C) and nitrogen (δ¹⁵N) revealed that the birds were not living in the rainforest when these feathers grew. Five ancient Pachacamac feathers tested (Supplementary Table 1) had enriched δ¹³C and δ¹⁵N values (–10.20 to –7.64‰ and 7.87 to 13.74‰ raw values, respectively) that are distinct from modern parrots living in the Amazonian rainforest (Fig. 4, and Supplementary Data 12). This isotopic signature of the ancient feathers provides a dietary “snapshot” consistent with the consumption of C4 plants (such as maize) possibly fertilized by seabird guano, indicating the birds were sustained on a coastal diet during the specific period when these feathers grew.

Plot of carbon (δ¹³C) and nitrogen (δ¹⁵N) stable isotope values from ancient samples from Pachacamac (blue), modern wild Amazonian parrots (red), and ancient parrots from the Atacama with a known captive diet (grey). The distinct isotopic signature of the Pachacamac feathers, which differs from the wild Amazonian samples, indicates a local diet that included C4 plants possibly fertilized by seabird guano. Human data from Pachacamac are from ref. 29 and parrot data from Atacama are from ref. 4 (Supplementary Data 12). Dashed lines connect pairs of values from the same individual (see explanation in ref. 4). Values were standardized to collagen112,113.
The apparent paradox between the birds’ wild genetic origin and their coastal dietary signature is resolved by a model of capture and transport. The high genetic diversity shows they were hatched in the wild, while the coastal isotopic signature shows they were kept in captivity on the coast for an extended period. This model of capturing wild birds and transporting them live across the Andes to be reared in coastal settlements is documented nearly 1,000 km south in the Atacama Desert of Chile, where isotopic data also confirmed a local maize-based diet for exotic parrots4. This conclusion is further supported by the complete lack of archaeological evidence for breeding at Pachacamac (no cages, skeletons, or eggshells) and a coastal climate ill-suited for breeding tropical parrots26,44. The combined evidence refutes the local breeding hypothesis and points to a sophisticated system capable of capturing live birds in the Amazon and moving them across one of the world’s most formidable mountain ranges.
Modeling the trans-Andean feather trade
Having established that the Ychsma sourced wild parrots from the Amazon, we sought to identify the trade routes that made this continental-scale procurement possible. We used two computational modeling approaches. First, we applied machine-learning species distribution models (SDMs) to reconstruct the parrots’ potential distributions around 1000 CE45. Second, we used landscape resistance modeling, based on circuit theory46, to map the paths of least resistance for humans between the Ychsma heartland and 10 contemporary sites in the Andes and Amazon, selected a priori for their importance as trade hubs or as potential sourcing locations for the parrots (Table 1). Our models accounted for elevation, the navigability of rivers, and the potential for seafaring (see “Methods”).
The hindcasted SDMs confirmed that the four identified parrot species’ habitats were restricted to the eastern side of the Andes, primarily in the wet tropics of the Amazon Basin, creating a vast and formidable barrier to the coast (Fig. 5a, and Supplementary Fig. 9). The landscape resistance models then identified two primary, highly plausible corridors for traversing this barrier (Fig. 5b–d).

a Predicted habitats for the four identified Amazonian parrot species around 1000 CE. Hindcasted species distribution models show that suitable habitats were restricted to the eastern side of the Andes, far from the coastal Pachacamac site. b–d Landscape resistance models of potential trade corridors connecting key sourcing sites to the Ychsma coastal heartland. In these models, yellow indicates high conductance (easy travel) and dark blue indicates high resistance (difficult travel). The models are based on elevation alone (b); elevation plus navigable rivers as low-resistance corridors (c); and elevation, rivers, and potential sea routes (d). Red lines show the calculated least-cost paths, and the hashed area highlights the Chachapoya region150, a plausible source area for traded birds. Maps were created using QGIS v3.3.
The first was a northern route, connecting the Ychsma to the powerful north-coast polities of the Chimú (capital at Chan Chan) and Sicán (a.k.a. Lambayeque; capital at Sicán). These centers consistently appeared as low-resistance nodes in our models (Supplementary Table 3). This route is strongly supported by archaeological evidence for long-standing interaction between the north coast and the Amazon. These ties have deep historical roots, with earlier Moche iconography often depicting Amazonian species, and were later maintained by the Chimú and Sicán, who are known to have had outposts and trade relationships with the Chachapoyas people of the upper Amazonian slopes, a region inhabited by the identified parrots and whose people were known for their bird-capturing skills47,48,49,50,51,52. This suggests a sophisticated, multi-stage network: birds sourced by the Chachapoyas could have been traded to either the Sicán or Chimú (where the captive birds might have been reared at a larger scale) and then transported south to Pachacamac via established coastal routes. The documented geopolitical expansions of the Chimú polity north- and southward along the Peruvian coast during the period of ca. 1250 to 1400 CE53, however, strongly suggest that it was the expansive Chimú Empire that served as the north coast nexus in the inferred multi-stage trade network. Not only the Chimú conquered the Sicán by 1375–1400 CE and interacted with the contemporaneous Ychsma54, but their production and usage of feathered objects seems to have been unprecedented3. Max Uhle even recovered a well-preserved Chimú feathered crown at Pachacamac during his pioneering fieldwork there (Supplementary Fig. 10).
The second was a more direct central route, connecting Pachacamac eastward across the Andes to the highlands, specifically toward the region of Cerro de la Sal. This corridor was particularly favored in models that included river travel (Fig. 5c). This route aligns with ethnohistorical accounts of the Yánesha, an Arawak-speaking group who inhabited this Amazonian region and traveled along these same paths to the central coast valleys for trade and exchange55,56. This corridor essentially represents an early iteration of a route later formalized by the Inca as part of their Qhapaq Ñan imperial road system57. Of note, our models showed that maritime trade, despite its documented use for other goods, offered no significant advantage for procuring Amazonian feathers (Fig. 5d).
Discussion
By integrating ancient genomics, isotopic analysis, and spatial modeling, we have traced the journey of feathers from Amazonia to an elite tomb on the Pacific coast during the Late Intermediate Period of the Andean chronology. Our results provide direct evidence that the Ychsma culture, long before the Inca empire, participated in a sophisticated trade network that spanned the Andes. We show that they sourced live parrots from genetically diverse wild populations in the Amazon, transported them over hundreds of kilometers either through their own direct networks or via powerful intermediaries, and sustained them on a coastal diet.
Our application of spatial modeling, supported by archaeological and ethnohistoric data, identifies two independent and highly plausible trade corridors that made this possible: a northern route leveraging particularly the powerful Chimú polity and to a lesser extent, the Sicán polity, as intermediaries; and a direct central route connecting the coast to Arawak-speaking groups in the central highlands. The existence of these robust networks fundamentally challenges the view of this era as a period of frequent conflicts and regionalism, revealing instead a complex mosaic of long-distance interaction and economic integration operating outside of a pan-Andean imperial structure.
This cross-disciplinary approach provides a powerful toolkit for archaeology. It moves beyond qualitative inference to quantitatively model ancient trade routes and interaction spheres. This framework, particularly suited for organic artifacts, can be applied to enigmatic materials from archaeological contexts worldwide, holding the promise of redrawing the map of ancient economies and revealing the true extent of connectivity between the world’s past cultures. This study also provided a deep historical context for a human fascination with colorful parrots that today drives a destructive illegal trade threatening their very survival.
Methods
Study sites and sample collection
The ancient feather samples were all recovered during a 2005 excavation in the Uhle Cemetery in front of the sacred Painted Temple at the famed pre-Hispanic religious center of Pachacamac on the central coast of Peru (Fig. 1). The 2005 fieldworks by the Pachacamac Archaeological Project (PAP) co-directed by Izumi Shimada and Rafael Segura Llanos were authorized by the National Institute of Culture of Peru (Resolución Directoral Nacional No. 919). Guided by results of their prior ground-penetrating radar survey, Shimada and his team excavated a large, intact, stone-lined, chamber tomb (Fig. 1c–e), 2.25 m (N-S) × 3.27 m (E-W) in dimension in Trench 7 within the Uhle Cemetery26,29. The tomb contained 34 well-preserved funerary bundles organized into two levels, seven in the upper (Fig. 1c) and 27 in the lower levels (Fig. 1d). Isotope and mtDNA analyzes of skeletal/dental samples of the deceased inside these bundles and associated artifacts (particularly ceramic styles) revealed that the deceased came from different communities on the central coast (lower reaches of coastal valleys) and the cis-Andean slopes (higher, interior portions) of the same valleys, including the Chillón, Rimac, and Lurín26.
Strontium isotope (87Sr/86Sr) analysis of bones from 26 individuals excavated in Trench 7 at the Uhle Cemetery and the neutron activation analysis of sherds from the adjacent Pilgrims Plaza both indicate that they were derived from at least three distinct locations26. To locate these sites, a comparative study of the ceramic vessels associated with the funerary bundles in Trench 7 was conducted, providing an independent confirmation of the analytical findings. It has revealed, that in addition to Ychsma domestic vessels occurring widely on the central coast, at least two jars—with heart-shaped body, a short globular neck, a pair of symmetrically placed strap handles, and brown paste (Supplementary Fig. 11)—are most likely derived from mid to upper valleys of the central to south-central coast from Rimac to Mala (see Plates 10-4066, 4289, 4291, and p116 in ref. 58, Fig. 3 in ref. 59, and ref. 60). The bundles were most likely brought on a litter (stretcher) to Pachacamac for the honor of being interred close to the venerated deity.
Examination of the 20 bundles—specifically those unfolded or partially exposed due to degradation—identified three infants, while the rest were adults aged ~22–61 years. The adult mortuary bundles (e.g., Bundles B, C, and W) were constructed using a complex, multi-layered technique. This process began by wrapping the flexed body in a cotton shroud (Fig. 1c, d). In the most elaborate examples (Bundles B and C; Fig. 1f), the deceased was clothed in a high-quality tunic woven from camelid fiber, with the cranium secured in decorated textiles. This primary wrapping was subsequently enclosed within a coarse netting made of braided junco or Scirpus sp. reeds. To create volume and shape, the spaces within and around this netting were packed with raw cotton bolls, bundled reeds, and other vegetative matter. Successive layers of cotton bags or large textiles were applied, repeating the stuffing and wrapping process to achieve the final dimensions. Structural stability was provided by inserting vertical cane or wooden supports along the flanks of the bundle after the formation of the third or fourth layer. The exterior was finished with a large textile sack simulating a tunic. A “false head” (constructed from a cloth bag filled with reed bundles and vegetation) was often stitched onto the top of the bundle. Due to the relatively poor state of preservation and ethical consideration of preserving some for future studies, not all bundles were opened. Consequently, Bundles A and C were neither unbundled nor radiocarbon-dated.
The site of Pachacamac being the preeminent religious-funerary center close to the city of Lima was subjected to intense grave looting—particularly targeting bundles of elite/wealthy individuals—since early Colonial era until the 20th century. Uhle15, who conducted the first scientific excavation of Pachacamac and often regarded as the father of the scientific archaeology in the Andes, considered that the great majority of an estimated 60,000 to 80,000 pre-Hispanic burials in six cemeteries that once existed at the site had been looted prior to his work26,61,62. In the Sacred Precinct toward the southern corner of the site, Uhle15 (see Figs. 2–5 in ref. 15) uncovered at least 50 relatively well-preserved funerary bundles (dating from Late Lima, ca. 600–650 CE to the Late Horizon, ca. 1470–1533 CE) in his excavations in areas (including cemeteries) around the sacred temples (Painted and Sun Temples). Many of these bundles did not preserve well and generally speaking, Uhle did not detail how these bundles were prepared61. He did not illustrate any of the excavated bundles, but there is a photograph (The Uhle Pachacamac Collections at the University of Pennsylvania Museum) that presumedly he took of the back of a relatively well-preserved bundle, apparently taken in the field soon after its excavation, that is dressed in a striking feather-covered cloak. A bundle of large feathers is tied to the back of its false head. Subsequent archaeological projects largely focused on clearing the top of these and other temples in the same zone, finding a handful of offering burials without any feather ornaments. The long-standing Ychsma Project directed by P. Eeckhout, on the other hand, has focused on the history, organization, and functions of the inferred Ychsma royal palaces (“Pyramids with Ramps”) in the central sector of the site, and since 1993 has “uncovered several hundred burials” of diverse age groups, both sexes, and those of unknown age and sex of predominantly Late Intermediate and Late Horizon date (ca. 1000–1500 CE)63,64,65,66,67. There are no mentions of any feather ornaments associated with these late pre-Hispanic individuals. They have been interpreted as either sacrificial victims to the Ychsma/Pachacamac deity or those who long suffered from health problems and were brought to the site to be divinely healed64,67. In addition, there is a well-preserved funerary bundle attributed to have been found (by looters) at Pachacamac and currently at the Ethnological Museum in Berlin, Germany (Inventory No. Berlin-VA 60382) that has a false head decorated with large yellow feathers on its back. It appears to be late pre-Hispanic in date.
The funerary bundles in these tombs belonged to the Ychsma culture, which is widely regarded as a late pre-Hispanic confederation of ethnic chiefdoms or kingdoms (known ethnohistorically señoríos or curacazgos) on the central coast. They were politically autonomous but acted as a religious community through the shared veneration of Ychsma (also spelled Ichma, Ichmay, or Ishma)30,31 that was renamed as Pachacamac during the Inca domination of the site68,69,70,71,72,73. Its political composition probably accounted for the variability in funerary bundle construction74,75. Ychsma became recognizable ca. 1000 CE or somewhat earlier and was later subjugated by the Inca Empire around 1460 CE29. Its territory covered the lower and partially middle portions of the valleys of Chillón, Rimac, and Lurín (Fig. 1a) with the sites of Maranga (Rimac Valley) and Pachacamac (Lurín Valley) serving the political and religious epicenters, respectively14,32,76,77. This time period—after the Wari and before the Inca Empire—is known as the Late Intermediate Period (1000–1476 CE) or Period of Regional States, in which Andean cultures develop a strong self-identity in their respective territories. These regionalized cultures somewhat also expand their territories but incomparable to the pan-Andean integration processes led by the Wari or the Incas.
Visual and macroscopic identification of feathers is often challenging, especially with old and/or poorly preserved samples78. Even with extensive ornithological expertise, especially in parrots, we were only able to identify some of the samples as macaws (Ara sp.) and Amazon parrots (Amazona sp.) but given the condition of the feathers and their similarity within each genus, explicit species identification was not always possible.
A subset of 25 ancient feathers from ornaments (Fig. 1g) in funerary bundles (see details in Supplementary Data 1) from the Ychsma period (1000–1470 cal. CE) were selected for the genetic analysis29 and sampled. To contribute to the preservation of the funerary bundles, feathers were not obtained directly from them, except samples #11938–11946 from Bundle C (Fig. 1f, and Supplementary Fig. 12). Whereas all feathers certainly came from the excavated funerary bundles, source of some feathers (Supplementary Data 1) could be identified and inferred according to their position on tomb floors and the immediate proximity to the bundles (e.g., samples #11947–11948 from Bundle W and samples #11949–11951 from Bundle A). Samples #11960 and #11961 were obtained from an excavation cut (Cut 1) into Layer 1 (near the modern surface) of the NE–NW sector in the large Trench T7, suggesting that these samples came from a looted burial or funerary bundle with a feather head ornament. The feather samples were placed into zip-lock bags or Eppendorf tubes, marked with a unique identifier, and sent to the paleogenomics laboratory of the ACAD, Adelaide University (Australia). Exportation of the feather samples used in this study was granted by the Ministry of Culture (Resolución Viceministerial No. 703-2011-VMPCIC-MC and 147-2024-VMPCIC-MC) and the Servicio Nacional Forestal y de Fauna Silvestre (SERFOR) in Peru (CITES permit No. 24-PE-005991-SP).
Blood samples from macaw nestlings were collected in 2010, using jugular venipuncture in the Bahuaja-Sonene National Park, southeastern Peru, following procedures approved by the Institutional Animal Care and Use Committee of the Texas A&M University (Animal Use Permit No. AUP 2009-215), and stored in 70% ethanol78,79. The sample S1109 from a female Scarlet Macaw A. macao nestling was used to generate mtDNA baits for capturing DNA sequences from the ancient feathers having been visually identified as belonging to macaws and Amazon parrots. Samples BYMA01 and BYMA04 from two Blue-and-yellow Macaw Ara ararauna nestlings were used for reference of the Peruvian haplogroup. Molted feather samples from parrots and macaws were also collected from the same region and in the Tambopata National Reserve, and used as reference for stable isotope analysis.
All modern samples were collected under research permits from the Servicio Nacional de Areas Naturales Protegidas (SERNANP) in Peru (Resolución Jefatural No. 001-2010-SERNANP-DGANP-PNBS and Resolución del Jefe de la Reserva Nacional Tambopata No. 001-2010-SERNANP-DGANP-JEF). For the transportation of modern blood and feather samples, CITES permits were provided by SERFOR in Peru (export permit No. 15-PE-000448-SP) and the Wildlife Trade and Biosecurity Branch of the Department of Environment in Australia (import permit No. PWS2015-AU-001508). Genetic access to the samples was granted by SERFOR in Peru (Resolución de Direccion General No. 013-2016-SERFOR-DGGSPFFS).
DNA extraction, sequencing, and blasting
Modern DNA
Fresh DNA from the modern blood samples was extracted using general salting out protocol and DNAzol (Invitrogen) at the genetic laboratories of The Australian National University in Canberra, Australia79. TrueSeq Nano Double-Stranded libraries (DSL) of samples BYMA01 and BYMA04 were constructed at Macrogen (South Korea) and shotgun sequenced in a NovaSeqX (150 bp paired-end). Adapter sequences from sequencing results were trimmed using “fastp” v0.20.180 and paired-end reads were mapped to the A. ararauna mitochondrial genome (GenBank accession number NC_029319) using BWA v0.7.17 “mem” algorithm81 with stringent mapping parameters (options -B 8 -O 10,10 -E 4,4 -T 80 -k 50). Duplicated reads were filtered out using samtools “rmdup”82. A mitochondrial genome 75% majority consensus for each sample was generated using Geneious v7.1.783, only calling nucleotides for positions with a coverage depth ≥3× (other positions were called as N; see Supplementary Data 5). The complete mitochondrial genome sequences for BYMA01 (coverage depth 23.2×) and BYMA04 (coverage depth 10.5×) were deposited to GenBank under accession numbers PV816315 and PV816316, respectively.
Ancient DNA
All ancient DNA extractions and library constructions were performed at the dedicated ancient DNA laboratory at ACAD, Australia.
Fragments from the 25 feathers from the Ychsma tomb were cut into tiny pieces using scissors (amount of feathers used ranged between 10 and 210 mg; see details in Supplementary Data 1). DNA from these feather samples was extracted following ref. 35. Shortly, digestion step was performed for each feather using 4 ml of a buffer containing 75 mM Tris pH 8.0, 50 mM NaCl (Sigma-Aldrich), 2 mg Proteinase K (Life Technologies), 50 mM DTT (Promega), and 0.75% SDS (Life Technologies), and digested for 1 h under constant rotation at 55 °C. Samples were then centrifuged at 3085 × g (4600 r.p.m.) for 1 min and the supernatant was pipetted into 16 ml of a modified QG buffer containing 91% QG buffer (Qiagen), 1.3% Triton X-100 (Sigma-Aldrich), 25 mM NaCl (Sigma-Aldrich), 0.17 M Sodium Acetate (Sigma-Aldrich), and 100 µl of silicon dioxide solution84, and left for 1 h at room temperature under constant rotation. Samples were centrifuged at 3085 × g (4600 r.p.m.) for 5 min to pellet the silica suspension, discarding supernatant. This silica pellet was washed three times in 80% ethanol and centrifuged at 18,215 × g (13,000 r.p.m.) for 1 min, and finally air dried for 30 min and resuspended in 120 μl of a pre-warmed (at 50 °C) mix of EB buffer (Qiagen) and 0.05% Tween 20, and incubated for 10 min. A final volume of 120 μl of DNA extract was obtained after centrifugation at 18,215 × g (13,000 r.p.m.) for 1 min. A negative control was included for all extractions.
Partial-UDG treated85 DSL were constructed following the protocol by ref. 86 using modifications in ref.87, which uses truncated Illumina adapters with a 7-mer barcode on both P5 and P7, and using AmpliTaq Gold (Invitrogen) for final library amplification. In-house synthesized RNA captures for mitochondrial enrichment were constructed using genomic DNA from the Ara macao macao sample S1109 following ref. 36, and libraries were enriched for mitochondrial DNA following ref. 88. Enriched libraries were finally sequenced in an Illumina NextSeq platform (mid-output, 150 bp paired-end).
Resulting sequencing reads were demultiplexed according to both P5 and P7 barcode sequences using Sabre v1.0 (https://github.com/najoshi/sabre), adapter sequences were removed using AdapterRemoval v2.3.089, and paired reads were collapsed (merged) into a single read when overlapping by 11 nucleotides, discarding reads shorter than 30 bp.
Collapsed reads of each sample were initially mapped to the A. macao mitochondrial genome (GenBank accession number CM002021) using BWA v0.7.17 backtrack algorithm81 and using mapping parameters for ancient samples (options: -l 1024, -n 0.01, -o 2). Duplicated reads were filtered out using FilterUniqueSAMCons.py90. A 75% majority consensus was then generated using Geneious v7.1.783, only calling nucleotides for positions with a coverage depth ≥3× (other positions were called as N). Longest contigs (>50 bp) obtained for each sample with at least 50 reads mapped (Supplementary Data 2) were BLASTn91 analyzed to identify them taxonomically. Thus, a subsequent mapping to mitochondrial genome of Amazona aestiva (NC_033336), Ara ararauna (NC_029319), A. macao (NC_045076), A. chloropterus (NC_047199), and Larus brunnicephalus (JX155863) was performed for those samples assigned to Amazona ventralis, Ara ararauna, A. macao, A. chloropterus, and Larus crassirostris by BLASTn, respectively. Iterative mapping to those references was then performed generating an intermediate 75% majority consensus sequence in Geneious retaining the reference allele at positions with coverage depth <3×. The new consensus sequences were then used as the references for a new round of mapping following the process described above. Once no more reads were mapped, a final 75% majority consensus was then generated using Geneious, only calling nucleotides for positions with a coverage depth ≥ 3× (other positions were called as N). Ancient DNA damage pattern was assessed using mapDamage v2.2.192.
Archaeogenetics results indicate that from the 25 feathers analyzed, eight provided enough endogenous DNA for taxonomic identification (Supplementary Fig. 1, and Supplementary Data 2–4). Ancient DNA damage analyzes for these samples display the expected patterns for sequencing libraries partially repaired with UDG (Supplementary Fig. 2).
Phylogenetic and haplogroup network analyses
To construct the different datasets for the species identification of the samples, just the samples covering at least 50% of the reference in the BWA mapping (independently of the coverage depth of the final consensus sequence) were used, except in the case of sample #11942 (~30%), identified as Amazona sp., as it was the only sample from this species with more than 50 reads mapped in the preliminary mapping to Ara macao. The sequence datasets were constructed to both verify the species identification of the samples furnishing enough DNA and to identify haplogroup for samples from which haplotype data were available (i.e., Ara macao and A. ararauna). Datasets were generated by aligning the corresponding sequences using MUSCLE93 implemented in Geneious v7.1.783. Maximum Likelihood (ML) phylogenetic analyzes for Ara and Laridae datasets were performed using IQTREE294, implementing substitution models as estimated by ModelFinder in IQTREE2 in both the partitioned (Ara mitochondrial genome dataset and A. macao network dataset, see below) and unpartitioned data (using option -m TEST, in Laridae and Ara ararauna phylogenetic trees), with node support values estimated by performing 1000 ultrafast bootstrap replicates95. Maximum Likelihood analyzes for Amazona unpartitioned datasets were performed using RAxML 8.2.1196 with node support values estimated by performing 100 bootstrap replicates. Haplogroup networks for Ara macao and A. ararauna were estimated using Fitchi97.
Ara species identification
Macaws are a polyphyletic group (consisting of six genera) and the largest sized parrots in the Neotropics with 16 species still extant in the wild, although many of them threatened by extinction today98. Samples from Pachacamac initially identified as belonging to Ara spp. provided partial mitochondrial genome sequences ranging from 12.3 to 96.8% of reference coverage and with a coverage depth ranging from 0.2 to 11.4×. A dataset was constructed using available Ara spp. mitochondrial genomes in GenBank (Supplementary Data 6) and partial mitogenomes from feather samples providing a minimum reference coverage of 50% (i.e., samples #11954 and #11957 after mapping to A. ararauna, sample #11951 after mapping to A. chloropterus, and samples #11947, #11948, and #11959 after mapping to A. macao; Supplementary Data 3). The 39 mitochondrial genome sequences of Ara species (available in GenBank) and our newly generated Ara sequences (six ancient individuals and the two modern A. ararauna BYMA01 and BYMA04) were aligned using MUSCLE in Geneious, using Amazona ventralis as outgroup (Supplementary Data 6), and the Control Region gene was removed. The dataset was divided in Protein Coding Genes (PCGs), 12S_rRNA, 16S_rRNA, and tRNAs. PCGs were further partitioned in codon positions (Supplementary Data 7) using DAMBE v7.0.599. Final dataset contained 48 individuals and was 15,455 bp long.
The ML phylogenetic analysis using mitochondrial genomes of Ara spp. (Fig. 2) confirms the preliminary BLASTn identification of feather samples, with unambiguous assignment [i.e., Maximum Likelihood Bootstrap (MLB) = 100] of samples #11954 and #11957 as A. ararauna, sample #11951 as A. chloropterus, and samples #11947, #11948, and #11959 as A. macao.
Ara macao and A. ararauna haplogroup networks
The concatenated alignment of mitochondrial genes 12S_rRNA, 16S_rRNA, COXI, and CYTB, and the corresponding data from complete and partial mitochondrial genomes from A. macao specimens available in GenBank (Supplementary Data 8) was used to construct a network to identify the haplogroup of samples #11947, #11948, and #11959. This dataset was also partitioned in 12S_rRNA, 16S_rRNA, and PCGs (further divided in codon positions using DAMBE; Supplementary Data 7). Haplogroups within A. macao were used following refs. 37, 43. The dataset contained 125 individuals of length 2243 bp.
Haplogroup network analysis of A. macao (Fig. 3, and Supplementary Fig. 3), as well as the mitochondrial genome phylogeny, confirmed the three ancient A. macao feathers belong to haplogroup H4 (but MLB = 86), which is one of two haplogroups characteristic of A. macao macao.
An alignment of 1290 bp and 55 individuals using the available A. ararauna mitochondrial CR longer than 500 bp and corresponding CR sequences from mitochondrial genomes available in GenBank (Supplementary Data 9), as well as the two sequences of modern individuals (BYMA01 and BYMA04) obtained here and partial sequences from samples #11954 and #11957, was used to generate a haplogroup network for the species. The Eastern/Western haplogroups (HE/HW) are based on ref. 100.
The A. ararauna haplogroup network analysis using partial sequences of the mitochondrial control region (Supplementary Figs. 4 and 5) showed that the two ancient feathers (#11954 and #11957) belong to the same haplogroup, which is related to the HE haplogroup. Modern sample BYMA01 is closely related to Eastern h6 and h23 haplotypes, and BYMA04 forms a single haplotype also within the HE haplogroup related to the other Peruvian samples. However, all the currently available Control Region gene information is from Brazilian individuals or of unknown origin (except the four Peruvian individuals analyzed here), so it is not possible to establish more accurate relationships with populations from other South American countries.
Ara macao nucleotide diversity
Nucleotide diversity was calculated for A. macao using DnaSP v5.10.1101 and the complete mitochondrial genomes divided in different sets based on haplogroup identification and specimen availability. The analysis was performed for all individuals of H4 (n = 9, comprising both ancient and modern samples) and H6 haplogroups (n = 15 with just ancient individuals from putative breeding colonies43). Three different subsets from the H4 haplogroup were analyzed: a subset including the Pachacamac samples (n = 3), another one including ancient samples from the Atacama Desert sourced from wild populations (n = 4)4, and a third one including modern samples (n = 2). An additional analysis was performed on H6 haplogroup removing sequence MH400234 (i.e., n = 14), which includes about 35% of unknown positions (Ns).
The nucleotide diversity (π) estimated for the A. macao mitochondrial genomes (Supplementary Table 2) is higher in the H4 (~0.002) than in H6 haplogroup individuals (~0.00005) coming from captive breeding colonies according to ref. 43. As expected, both nucleotide diversity and haplotype diversity (Hd) are lower in captive breeding individuals (Hd = 0.371 when using all H6 individuals, and Hd = 0.495 when removing the H6 individual with 35% Ns). In contrast, in the H4 sets of in individuals, the Hd = 1 (i.e., one haplotype for each individual)4, suggesting that the four individuals studied from the Atacama Desert came from traded animals from different wild populations. The very similar nucleotide diversity of these Atacama individuals (0.00272) and the three Pachacamac individuals (0.00223), and the remarkably lower nucleotide diversity of the individuals coming from breeding colonies in the American Southwest (0.00005), suggest that the Peruvian individuals would have also come from wild populations.
Laridae species identification
A single dataset concatenating available sequences for mitochondrial genes COXI, CYTB, and ND2 was generated to identify the Laridae species for sample #11960 by aligning selected genes obtained for the feather sample and the available gull CYTB, COXI, and ND2 gene sequences, and corresponding sequences from mitochondrial genomes available in GenBank (Supplementary Data 10). One individual per species was selected, and when multiple sequences per gene were available, the longest one was selected. Specific names in this Laridae group follow the proposed taxonomy by ref.102. Three tern species were also used (Gelochelidon nilotica, Sterna hirundo, and Sternula albifrons), as well as Haematopus ater and Calidris pugnax as outgroups. The final dataset contained 62 sequences of a total length of 3736 bp.
The white feather sample #11960 (Supplementary Fig. 1h) was identified as belonging to a Laridae species (Supplementary Data 1). The phylogenetic analyzes, which included most species of Laridae, place the feather within Sabine’s Gull Xema sabini with high confidence (MLB = 100; Supplementary Fig. 8).
Amazona species identification
Although the amount of data retrieved for sample #11942 is very limited (see below), three different datasets were generated to infer the species identification of this sample. As no CYTB data were recovered from the ancient sample, the datasets used were obtained by aligning the sample #11942 sequences to Amazona sp. gene sequences for mitochondrial COXI [334 individuals (using Ara macao as outgroup), 1548 bp)], 12S_rRNA [74 individuals (Ara ararauna as outgroup), 391 bp], and 16S_rRNA [72 individuals (A. ararauna as outgroup), 542 bp] genes (Supplementary Data 11).
When analyzing the feather preliminarily identified as Amazona sp. (sample #11942), just 9, 15.3, and 17.3% of the final alignments for COXI, 12S_rRNA, and 16S_rRNA Amazona datasets, respectively, were non-degenerate nucleotides. However, phylogenetic analyzes strongly support [MLB = 94 for COXI analysis (Supplementary Fig. 6), 94 for 12S_rRNA, and 100 for 16S_rRNA (Supplementary Fig. 7)] the attribution of this sample to the Mealy Amazon Amazona farinosa and probably to subspecies A. f. farinosa, inornata, or chapmani.
Human contamination
The BLASTn analysis of the longest contig after mapping for sample #11938 suggested possible contamination with human DNA (Supplementary Data 2). In order to verify if the DNA was potentially coming from the human buried in the funerary bundle, we remapped the sequencing reads for this sample to the human RSRS mitochondrial genome103. Up to 1077 unique reads (covering 66% of the reference at a mean coverage depth of 5.3× and with a mean mapped read length of 81.1 bp) were mapped, especially in those mitochondrial genome regions highly conserved across vertebrates (i.e., 12S_RNA and 16S_RNA genes). Analysis of the consensus sequence using the tool Haplocheck v1.3.1104 from the Mitoverse platform (https://mitoverse.readthedocs.io/) identified the human haplogroup as H2a1, and there was no other source of human contamination (i.e., no ambiguity about H2a1 and no sequences that could come from another individual). H2a1 is a pan-Eurasian haplogroup found from Siberia to southern Spain105,106,107,108, indicating that the source of human DNA of that sample is a recent contamination that most probably occurred during excavation or sampling tasks. These results are compatible with the higher mean length of the reads mapped in comparison with the endogenous bird reads (81.1 bp versus <56 bp) and with the unclear damage pattern (Supplementary Fig. 13).
Radiometric dating and stable isotope analysis
Radiocarbon dating was performed on five feather samples from Pachacamac (#11519, #11520, #11522, #12352, #12455; Supplementary Data 1) in the Radiocarbon Laboratory (SANU), Research School of Earth Sciences, The Australian National University (Australia). During a non-routine pre-treatment, the feathers were subjected to a sequential solvent wash followed by an acid-base-acid (ABA) protocol to remove a broad range of potential contaminants. The solvent wash consisted of immersing the sample in acetone, methanol, and dichloromethane (DCM). Following the solvent wash, the ABA protocol was performed as follows: samples were treated with 1 M hydrochloric acid (HCl) for 30 min at 70 °C, then with 0.2 M sodium hydroxide (NaOH) for 1 h at room temperature, and finally with 1 M HCl for another 30 min at 70 °C. Radiocarbon dating was performed using a single-stage AMS system as described by ref. 109. This system provides high-precision measurements of radiocarbon content, ensuring reliable age determinations for the analyzed samples. We calibrated the results using the southern hemisphere calibration curve SHCal20110 in Oxcal v4.4.4111. Calibrated current era (CE) years are presented in 95.4% (2σ) ranges.
Carbon (δ13C) and nitrogen (δ15N) isotope contents were measured in five contemporary parrot feathers (collected in the Tambopata-Candamo area, Southeastern Peruvian Amazon) and the five feather samples from Pachacamac described earlier (Supplementary Data 1) in the Stable Isotope Laboratory, Research School of Biology, The Australian National University (Australia). After cleaning the feathers with three chloroform-methanol washes, the samples were dried, and their C and N contents were measured by combustion of the samples in an EA1110 elemental analyzer (CE Instruments, Milan, Italy) with the resulting gas analyzed by an Isoprime mass spectrometer (Micromass UK, Manchester, England). A mean for δ13C and δ15N values obtained for each sample was calculated. For comparative purposes, C and N isotope values for feather samples have been normalized to values obtained from bone collagen as in ref.4 following ref. 112. For this reason, we added 2.8‰ δ13C and subtracted 2.2‰ δ15N in soft tissue samples and added 1.6‰ δ13C to feather samples, which provided the data to generate a δ13C/δ15N plot using the bird samples studied here and the ones from ref. 4. For comparison with human samples, we also included data from previously analyzed human hair samples from Pachacamac29. C and N isotope values for human hair have been normalized for collagen following ref. 113, i.e., adding 1.2 ‰ to δ15N and 0.7 ‰ to δ13C values for hair keratin. Results of the five contemporary and five ancient feather samples are shown in Supplementary Table 1, Fig. 4, and Supplementary Data 12.
GIS analyses
Considering that the present distribution of all identified parrot species includes Peru and some neighboring countries, and that maritime trade might have occurred in the region and time of interest, we defined our geographical provenance investigation to a sensible catchment area around Pachacamac (82°W, 14°S, 70°E, 0°N) extending into the sea and the mainland (around 2 million km2 in total). A southernmost limit of 14°S was selected, as available archaeological evidence strongly suggests a close and probably direct contact (establishing colonies or sending emissaries) with the north as far as the Lambayeque region but no evidence is known about contact with the south114,115,116,117.
Hindcasted species distribution models
SDMs were implemented with the application Wallace EcoMod45 available from the “wallace” v2.1.0 R package in R v4.2.2118, which provides a freely available, open resource, and reproducible framework for species distribution modeling119. Occurrence data for the four parrot species identified in this study were downloaded from the Global Biodiversity Information Facility120, after the following filtering: observations were sourced from the eBird Observation Dataset, the basis of record field was “made by humans”, with coordinates without geospatial issues (e.g., invalid or zero coordinates, country-coordinate mismatch, coordinate out of range, etc.), within the distributional range of each species in Peru, as defined by the expert-derived range maps from BirdLife International121.
Environmental data were obtained with “pastclim” v1.2122 from the Krapp et al. (2021) dataset123 for 17 standard bioclimatic variables (bio02 and bio03 not being available) with a resolution of 0.5° for the present (for training) and 1000 CE (for projection). Occurrence data within the same cells of bioclimatic variables used and within 10 km of each other (spatial thinning) were also removed with “spThin” v0.2.0124.
For modeling, we first defined a training background of the country borders of Peru with a 0.5° buffer, then used “wallace” to randomly sample 532 background points. We then built SDMs using the algorithm Maxent125, which estimates the species’ responses to environmental variables126,127. We tuned the complexity of models by testing a set of feature classes (L = linear, LQ = linear-quadratic, H = hinge, LQH = linear-quadratic-hinge, and LQHP = linear-quadratic-hinge-product) with a regularization multiplier of value 1 (the default penalty against complexity in Maxent), without clamping (i.e., unconstrained extrapolation). We selected optimal model complexity settings by choosing the model with a delta AICc of 0.
We transferred the optimal models to the environmental variables for the year 1000 CE in our study area (defined above). We made model predictions with the “cloglog” transformation128, which is bounded between 0 and 1. Continuous predictions of habitat suitability were converted into binary predictions that indicate presence/absence predictions for each species using the “10 percentile” threshold, omitting all regions with habitat suitability lower than the suitability values for the lowest 10% of occurrence records within Peru. The executable R code that can reproduce the analysis with Wallace and raw data are available in Figshare (https://doi.org/10.6084/m9.figshare.24665697).
The machine-learning hindcasted species distribution reconstruction of the identified psittacine species’ distributions for 1000 CE did not find optimal climatic conditions in Pachacamac or between the Pacific coastline and the western slopes of the Andes for any of these species (Supplementary Fig. 9). The predicted distributions 1000 years ago were similar to their present-day ranges in the wet tropics, mainly in the Amazon Basin up to the eastern slopes of the Andes (Fig. 5a).
Resistance maps and least-cost paths of trade routes
Three landscape resistance models were developed to examine how topography affected potential trade routes between Pachacamac and other contemporary settlements (~1000–1500 CE) in the Peruvian Andes and Western Amazon regions. Such models are routinely used in landscape ecology studies129,130 but, to the best of our knowledge, they have not been applied to archaeology. Each map was compiled on a separate raster grid of 1-km resolution in QGIS v3.3131. The study area was defined as the valleys of Chillón, Rimac, and Lurín (Ychsma homeland) as the most probable sources of the funerary bundles26,29 with a rectangle catchment area defined above.
The resistance maps included (i) an elevation model, based on the Global Multi-resolution Terrain Elevation Data 2010132 with resistance values of meters above sea level, investigating the effect of topography on trade routes; (ii) an elevation + river model, including potentially navigable rivers for small boats (around 5–15 meters minimum extent) in South America133, with resistance values of “1” given to the cells with navigable rivers and “1000” outside of this zone, and summed with the overlapping elevation values to investigate the influence of river transport; and (iii) an elevation + river + sea model, including the sea level surface (with resistance values of “1000” to match the scale of the previous model) as potentially navigable routes for maritime trade.
Ten sites—contemporary to the Ychsma culture—were selected either within the predicted presence of the birds 1000 CE or outside their range but regarded as important trade hubs in that time (listed with details in Table 1). CIRCUITSCAPE v5.046 was used in the Julia programming language to generate landscape resistance values based on circuit theory134, between the Ychsma homeland and the 10 a priori selected sites, taking into account all possible pathways. A pairwise modeling approach was used to calculate resistance values, and cumulative current maps were generated for each resistance model to identify areas that contribute most to connectivity between sites. In this framework, “resistance” represents the difficulty of movement across a given landscape feature (e.g., high elevation poses high resistance), and “current” represents the modeled flow of movement, with high-current areas indicating the most probable corridors. Maps were visualized in QGIS v3.3, and overlaid with the potential present distribution predictions of parrot species121 identified by the archaeogenomic analyzes. To complement the connectivity maps, we also identified the single most efficient routes. While CIRCUITSCAPE excels at modeling broad corridors by considering all possible pathways simultaneously, it does not generate a single least-cost path. Therefore, to identify the single route of minimum cumulative resistance, we used the same resistance raster maps in the Least-Cost Path v1.1 plugin in QGIS v3.3 to calculate paths between Pachacamac and the a priori sites. This dual approach allowed us to visualize both the most probable regions for movement and the most efficient path within those regions.
In the first scenario, when we only accounted for elevation in the model, Chan Chan and Sicán (on the coast), and Sarayacu (in the Amazon Basin) were assigned with the lowest resistance values (Supplementary Table 3) connecting to the Ychsma homeland (Fig. 5b), while the lowest cost paths were assigned to Chan Chan, Sicán, and Manta (Supplementary Table 3), indicating a coastal transit route (Fig. 5b). Sarayacu was also located within (or at the border of) the predicted past distribution of all four parrot species (Supplementary Fig. 9).
In the second scenario, when navigable rivers were added to the elevation model as highly “conductive” pathways, Sarayacu had the lowest resistance values (Supplementary Table 3) through a trans-Andean pathway (Fig. 5c), while Chan Chan, Sicán, and Cerro de la Sal had the lowest cost path values calculated (Supplementary Table 3). Cerro de la Sal (at the foothills of the Andes on the Amazonian side) was also located within (or right at the border of) the predicted distribution of all four parrot species in 1000 CE (Supplementary Fig. 9).
In the third scenario, when seafaring was also considered on top of the previous models, Chan Chan, Sicán, and Sarayacu showed the lowest resistance values (Supplementary Table 3, and Fig. 5d), while Chan Chan, Sicán, and Cerro de la Sal had the lowest cost path values calculated (Supplementary Table 3). Both the river and the seafaring models predicted to connect Cerro de la Sal with Pachacamac directly through the Andes, even though it is isolated from the central coast by mountains over 5000 m above sea level.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
New sequences obtained by this study are available at the National Center for Biotechnology Information (NCBI) GenBank, Sequence Read Archive (SRA), and Figshare (see Supplementary Data 13 for accession codes and links). Previously published sequences used in this study for phylogeny reconstructions are listed in Supplementary Data 2–6 and 8–11. Sequences with more than 50% of Ns (given that GenBank does not permit the deposition of such sequences), alignments of the ten Maximum Likelihood (ML) analyzes in Phylip format, and the radiocarbon results are available in Figshare (https://doi.org/10.6084/m9.figshare.24665697). All other data are provided in the Supplementary Data sheets. Source Data for Fig. 4 can be found in Supplementary Data 12. The original archaeological samples are permanently curated and accessible at the Museo de Sitio de Pachacamac in Peru (Mg. Denise Pozzi-Escot, museopachacamac@cultura.gob.pe). Remaining samples from living birds are stored at freezers of the Research School of Biology, The Australian National University.
Code availability
Code for the hindcasted species distribution and landscape resistance models are available in Figshare at https://doi.org/10.6084/m9.figshare.24665697.
References
King, H. Peruvian Featherworks: Art of the Precolumbian Era (Metropolitan Museum of Art, New York, 2012).
Reina, R. E. & Kensinger, K. M. The Gift of Birds: Featherwork of Native South American Peoples (Penn Museum of Archaeology and Anthropology, Philadelphia, 1991).
Rowe, A. P. & O’Neill, J. P. Costumes and Featherwork of the Lords of Chimor: Textiles from Peru’s North Coast (The Textiles Museum, Washington DC, 1984).
Capriles, J. M. et al. Pre-Columbian transregional captive rearing of Amazonian parrots in the Atacama Desert. Proc. Natl. Acad. Sci. USA 118, e2020020118 (2021).
Paul, A. Paracas Ritual Attire: Symbols of Authority in Ancient Peru (University of Oklahoma Press, Norman, 1990).
Reid, J. W. Magic Feathers: Textile Art from Ancient Peru (Textile & Art Publications, London, 2005).
Wilkinson, D. The influence of Amazonia on state formation in the ancient Andes. Antiquity 92, 1362–1376 (2018).
Anawalt, P. R. Indian Clothing Before Cortéz: Mesoamerican Costumes from the Codices (University of Oklahoma Press, Norman, 1981).
Crown, P. L. Just macaws: a review for the U.S. Southwest/Mexican Northwest. KIVA 82, 331–363 (2016).
Paul, A. Paracas Necrópolis textiles: symbolic visions of coastal Peru. In The Ancient Americas: Art from Sacred Landscapes. 278–290 (Art Institute of Chicago, Chicago, 1992).
Townsend, R. F. Deciphering the Nazca world: ceramic images from ancient Peru. Art. Inst. Chic. Mus. Stud. 11, 117–139 (1985).
Kensinger, K. M. Why feathers? in The Gift of birds: Featherwork of Native South American peoples (Penn Museum of Archaeology and Anthropology, Philadelphia, 1991).
Dulanto, J. Dios de Pachacamac: El ídolo y el templo. in Los dioses del antiguo Perú (ed Makowski, K.) Vol. 2 158–181 (Banco de Crédito del Perú, Lima, 2001).
Shimada, I. Pachacamac Archaeology: Retrospect and Prospect (University Museum Press, University of Pennsylvania, Philadelphia, 1991).
Uhle, M. Pachacamac: Report of the William Pepper, M.D., LL. D., Peruvian Expedition of 1896 (Department of Archaeology, The University of Pennsylvania, Philadelphia, 1903).
Kaulicke, P., Fisher, M., Masson, P. & Wolff, G. Max Uhle (1856-1944): Evaluaciones de Sus Investigaciones y Obras (Pontificia Universidad Católica del Perú, Lima, 2010).
Menzel, D. The Archaeology of Ancient Peru and the Work of Max Uhle (UC Berkeley: Phoebe A. Hearst Museum of Anthropology, 1977).
Rowe, J. H. Max Uhle, 1856-1944: A Memoir of the Father of Peruvian Archaeology (University of California Publications in American Archaeology and Ethnology, 1954) Vol. 46.
Willey, G. R. & Sabloff, J. A. A History of American Archaeology. (W. H. Freeman, San Francisco, 1993).
Forshaw, J. M. Parrots of the World (Princeton University Press, 2006).
Olah, G. et al. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 25, 205–223 (2016).
Ellegren, H. Genomic DNA from museum bird feathers. in Ancient DNA: Recovery and Analysis of Genetic Material from Paleontological, Archaeological, Museum, Medical, and Forensic Specimens (eds Herrmann, B. & Hummel, S.) 211–217 (Springer, New York, 1994).
Rawlence, N. J., Wood, J. R., Armstrong, K. N. & Cooper, A. DNA content and distribution in ancient feathers and potential to reconstruct the plumage of extinct avian taxa. Proc. R. Soc. B Biol. Sci. 276, 3395–3402 (2009).
Bi, K. et al. Unlocking the vault: next-generation museum population genomics. Mol. Ecol. 22, 6018–6032 (2013).
Rowe, K. C. et al. Museum genomics: Low-cost and high-accuracy genetic data from historical specimens. Mol. Ecol. Resour. 11, 1082–1092 (2011).
Shimada, I. et al. Un siglo después de Uhle: Reflexiones sobre la arqueología de Pachacamac y Perú. In Max Uhle (1856-1944). Evaluaciones de sus investigaciones y obras (eds Kaulicke, P., Fischer, M., Masson, P. & Wolff, G.) 109–150 (Fondo Editorial de la Pontificia Católica del Perú: Lima, Peru, 2010).
Degano, I. & Colombini, M. P. Multi-analytical techniques for the study of pre-Columbian mummies and related funerary materials. J. Archaeological Sci. 36, 1783–1790 (2009).
Shimada, I., Segura Llanos, R., Rostworowski de Diez Canseco, M. & Watanabe, H. Una nueva evaluación de la Plaza de los Peregrinos de Pachacamac: Aportes de la primera campaña 2003 del Proyecto Arqueólogico Pachacamac. Bulletin de l’Institut Français d’Études Andines 507–538 (2004) https://doi.org/10.4000/bifea.5106.
Takigami, M. K. et al. Assessing the chronology and rewrapping of funerary bundles at the prehispanic religious center of Pachacamac, Peru. Lat. Am. Antiquity 25, 322–343 (2014).
Rostworowski de Diez Canseco, M. Breve informe sobre el señorío de Ychrna or Ychima. Boletín del. Seminario de. Arqueología, Inst. Riva Ag.üero-PUCP 13, 37–51 (1972).
Rostworowski de Diez Canseco, M. Pachacamac y el Señor de los Milagros: Una Trayectoria Milenaria (Historia Andina, 1992) Vol. 19.
Díaz Arriola, L. Reflexiones sobre el territorio y el paisaje cultural Ychsma durante el Horizonte Tardío. in Paisaje y territorio en los Andes centrales: Prácticas sociales y dinámicas regionales 145–155 (Universidad Nacional Mayor de San Marcos, Fondo Editorial, Lima, Peru, 2021).
Covey, R. A. Multiregional perspectives on the archaeology of the Andes during the Late Intermediate Period (c. A.D. 1000–1400). J. Archaeol. Res. 16, 287–338 (2008).
Whitney, B. M. Flight behaviour and other field characteristics of the genera of Neotropical parrots. Cotinga 5, 32–42 (1996).
Tobler, R. et al. Aboriginal mitogenomes reveal 50,000 years of regionalism in Australia. Nature 544, 180–184 (2017).
Richards, S. M. et al. Low-cost cross-taxon enrichment of mitochondrial DNA using in-house synthesised RNA probes. PLoS ONE 14, e0209499 (2019).
Schmidt, K. L., Aardema, M. L. & Amato, G. Genetic analysis reveals strong phylogeographical divergences within the Scarlet Macaw Ara macao. Ibis 162, 735–748 (2020).
Wenner, T. J., Russello, M. A. & Wright, T. F. Cryptic species in a Neotropical parrot: Genetic variation within the Amazona farinosa species complex and its conservation implications. Conserv. Genet. 13, 1427–1432 (2012).
Day, R. H., Stenhouse, I. J. & Gilchrist, H. G. Sabine’s Gull (Xema sabini), version 1.0. in Birds of the World (ed Billerman, S. M.) (Cornell Lab of Ornithology, Ithaca, NY, USA, 2020).
Somerville, A. D., Nelson, B. A. & Knudson, K. J. Isotopic investigation of pre-Hispanic macaw breeding in Northwest Mexico. J. Anthropol. Archaeol. 29, 125–135 (2010).
Schwartz, C. W., Somerville, A. D., Nelson, B. A. & Knudson, K. J. Investigating pre-Hispanic scarlet macaw origins through radiogenic strontium isotope analysis at Paquimé in Chihuahua, Mexico. J. Anthropol. Archaeol. 61, 101256 (2021).
Conrad, C. et al. Scarlet macaw (Ara macao) breeding at the Mimbres archaeological site of Old Town (early AD 1100s) in Southwestern New Mexico. PNAS Nexus 2, pgad138 (2023).
George, R. J. et al. Archaeogenomic evidence from the southwestern US points to a pre-Hispanic Scarlet Macaw breeding colony. Proc. Natl. Acad. Sci. USA 115, 8740–8745 (2018).
Moreno Herrera, J. L., Dueñas Olivera, K. L. & Huamán Nieto, M. Hidrogeología de la cuenca del río Lurín - [Boletín H 11]. Repositorio Institucional INGEMMET https://hdl.handle.net/20.500.12544/3297 (2021).
Kass, J. M. et al. Wallace 2: A shiny app for modeling species niches and distributions redesigned to facilitate expansion via module contributions. Ecography 2023, e06547 (2023).
Hall, K. R. et al. Circuitscape in Julia: empowering dynamic approaches to connectivity assessment. Land 10, 301 (2021).
Shimada, I. Cultura Sicán: Dios, Riqueza y Poder En La Costa Norte Del Perú (EDUBANCO, Fundación del Banco Continental para el Fomento de la Educación y la Cultura, Lima, 1995).
Horkheimer, H. Algunas consideraciones acerca de la arqueologiá en el Valle del Utcumbamba. in Actas y Trabajos del II Congreso Nacional de Historia del Perú Vol. 1 71–90 (Centro de Estudios Histórico-Militares del Perú, Lima, 1959).
Shimada, I. Detrás de la máscara de oro: La Cultura Sicán. in Cultura Sicán: Esplendor preinca de la costa norte (ed. Shimada, I.) 15–90 (Fondo Editorial del Congreso de Perú, Lima, 2014).
Thompson, D. E. Ancient highland connections with selva and coast: evidence from Uchucmarca, Peru. in Proc. 44 International Congress of Americanists (eds Browman, D. L., Burger, R. L. & Rivera, M.) 153–160 (Manchaster, 1984).
Thompson, D. E. Archaeological investigations in the eastern Andes of northern Peru. in Proc. XL Congresso Intemazionale degli Americanisti Vol. 1 363–369 (Roma-Genova, 1973).
Thompson, D. E. Prehistory of the Uchucmarca Valley in the north highlands of Peru. in Actas del XLI Congreso Internacional de Americanistas Vol. 2 99–106 (Mexico City, 1976).
Moore, J. D. & Mackey, C. J. The Chimú Empire. in The Handbook of South American Archaeology (eds Silverman, H. & Isbell, W. H.) 783–807 (Springer New York, New York, NY, 2008). https://doi.org/10.1007/978-0-387-74907-5_39.
Eeckhout, P. Imperial Strategies in a Regional Context, Chimus and Incas at Pachacamac. in Wars and Conflicts in Prehispanic Mesoamerica and the Andes (eds Eeckhout, P. & Le Fort, G.) 110–127 (Hadrian Books, Oxford, 2005).
Smith, R. C. ¿Un sustrato arawak en los Andes centrales? La historia oral y el espacio histórico cultural Yánesha. in Por donde hay soplo (eds Chaumeil, J.-P., Espinosa de Rivero, Ó. & Cornejo C., M.) 219–254 (Instituto Francés de Estudios Andinos, Lima, 2011).
RROMAPATSRO: Mapeando el espacio histórico-cultural de los Yánesha. Vol. 1 (Instituto del Bien Comun, Lima, 2006).
Capriata Estrada, C. El tramo Xaxua-Pachacamac y su importancia dentro de la red vial andina del Qhapaq Ñan. Ministerio de Cultura http://repositorio.cultura.gob.pe/handle/CULTURA/191 (2013).
Engel, F. A. Prehistoric Andean Ecology. Man, Settlement and Environment in the Andes. Vol. 4 (Humanities Press for the Department of Anthropology, New York City, 1984).
Falcón, R. A. & Pozzi-Escot, D. Del Horizonte Medio al Horizonte Tardío en la costa sur central: El caso del valle de Asia. Bulletin de l’Institut français d’études andines 861–886 (2004) https://doi.org/10.4000/bifea.5388.
Llanos, R. S. Rito y Economía En Cajamarquilla: Investigaciones Arqueoloǵicas En El Conjunto Arquitectónico Julio C. Tello (Fondo Editorial PUCP, 2001).
Fleming, S. The mummies of Pachacamac. Expedition 28, 39–45 (1983).
Fleming, S., Miller, W. T. & Brahin, J. L. The Mummies of Pachacamac, Peru. MASCA J. 2, 137–159 (1983).
Eeckhout, P. Mummy false heads from Pachacamac and the ontology of the dead in ancient Peru. World Archaeol. 52, 685–706 (2020).
Owens, L. S. & Eeckhout, P. To the god of death, disease and healing: Social bioarchaeology of cemetery I at Pachacamac. in Funerary Practices and Models in the Ancient Andes (eds Eeckhout, P. & Owens, L. S.) 158–185 (Cambridge University Press, New York, 2015).
Owens, L. S. & Eeckhout, P. A. A computed tomography and dissection study of a unique Ychsma burial bundle from the site of Pachacamac, Peruvian Central Coast. https://doi.org/10.21203/rs.3.rs-2447960/v1 (2023).
Davies-Barrett, A. M., Owens, L. S. & Eeckhout, P. A. Paleopathology of the Ychsma: Evidence of respiratory disease during the Late Intermediate Period (AD 1000-1476) at the Central Coastal site of Pachacamac, Peru. Int. J. Paleopathol. 34, 63–75 (2021).
Eeckhout, P. & Owens, L. S. Human sacrifice at Pachacamac. Lat. Am. Antiquity 19, 375–398 (2008).
Eeckhout, P. The palaces of the Lords of Ychsma: An archaeological reappraisal of the function of pyramids with ramps at Pachacamac, Central Coast of Peru. Revista de Arqueología Americana 17–19, 217–254 (1999).
Eeckhout, P. La sombra de Ychsma. Bulletin de l’Institut français d’études andines 403–423 (2004) https://doi.org/10.4000/bifea.5047.
Rostworowski de Diez Canseco, M. Señoríos Indígenas de Lima y Canta (Instituto de Estudios Peruanos, 1978).
Segura, R. A. Andean Urban Processes and the Experience of the Environmental-social Interplay: The Case of Cajamarquilla in the Peruvian Central Coast (ca. AD 650- 1400) (Southern Illinois University, Carbondale, Illinois, 2023).
Makowski, K. Pachacamac old Wak’a or Inka syncretic deity? Imperial transformation of the sacred landscape in the lower Ychsma (Lurín) Valley. in The archaeology of Wak’as: Explorations of the sacred in the pre-Columbian Andes (ed. Bray, T.) 127–166 (University of Colorado Press, Boulder, 2015).
Patterson, T. C. Pachacamac – An Andean oracle under Inca rule. in Recent studies in Andean prehistory and protohistory (eds Kvietok, P. D. & Sandweiss, D. H.) 159–175 (Cornell Latin American Studies Program, Ithaca, New York, 1985).
Díaz Arriola, L. Identidad cultural, prácticas funerarias y territorio Ychsma. in Repensar el antiguo Perú: Aportes desde la arqueología (ed. Vega-Centeno, R. S.-L.) 341–362 (Instituto de Estudios Peruanos y Fondo Editorial PUCP, Lima, Peru, 2017).
Vallejo-Berríos, F. El estilo Ychsma: Características generales, secuencia y distribución geográfica. Bulletin de l’Institut français d’études andines 595–642 https://doi.org/10.4000/bifea.5165 (2004).
Espinoza, P. Arquitectura y procesos sociales tardíos en Maranga, Valley Bajo de Rímac, Lima. in Arqueología en el Perú: Nuevos aportes para el estudio de las sociedades andinas prehispánicas (eds Velarde, R. R. & Svendsen, T. P.) 263–309 (Anheb Impresiones, Lima, Peru, 2010).
Espinoza, P. La arquitectura de Maranga en el contexto del núcleo monumental tardío del valle bajo del Rímac. in Arqueología. Catorce años de investigaciones en Maranga (eds Carrión, L. & Narváez, J. J.) 121–149 (Municipalidad Metropolitana de Lima and Parque de Las Leyendas, Lima, Peru, 2014).
Olah, G., Heinsohn, R. G., Brightsmith, D. J., Espinoza, J. R. & Peakall, R. Validation of non-invasive genetic tagging in two large macaw species (Ara macao and A. chloropterus) of the Peruvian Amazon. Conserv. Genet. Resour. 8, 499–509 (2016).
Olah, G., Heinsohn, R. G., Espinoza, J. R., Brightsmith, D. J. & Peakall, R. An evaluation of primers for microsatellite markers in Scarlet Macaw (Ara macao) and their performance in a Peruvian wild population. Conserv. Genet. Resour. 7, 157–159 (2015).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890 (2018).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Li, H. et al. The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Kearse, M. et al. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647–1649 (2012).
Brotherton, P. et al. Neolithic mitochondrial haplogroup H genomes and the genetic origins of Europeans. Nat. Commun. 4, 1764 (2013).
Rohland, N., Harney, E., Mallick, S., Nordenfelt, S. & Reich, D. Partial uracil–DNA–glycosylase treatment for screening of ancient DNA. Philos. Trans. R. Soc. B Biol. Sci. 370, 20130624 (2015).
Meyer, M. & Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, pdb.prot5448 (2010).
Llamas, B. et al. Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas. Sci. Adv. 2, e1501385 (2016).
Mitchell, K. J. et al. Ancient mitochondrial DNA reveals convergent evolution of giant short-faced bears (Tremarctinae) in North and South America. Biol. Lett. 12, 20160062 (2016).
Schubert, M., Lindgreen, S. & Orlando, L. AdapterRemoval v2: rapid adapter trimming, identification, and read merging. BMC Res. Notes 9, 88 (2016).
Kircher, M. Analysis of high-throughput ancient DNA sequencing data. in Ancient DNA: Methods and Protocols (eds Shapiro, B. & Hofreiter, M.) 197–228 (Humana Press, Totowa, NJ, 2012). https://doi.org/10.1007/978-1-61779-516-9_23.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
Jónsson, H., Ginolhac, A., Schubert, M., Johnson, P. L. F. & Orlando, L. MapDamage2.0: Fast approximate Bayesian estimates of ancient DNA damage parameters. Bioinformatics 29, 1682–1684 (2013).
Edgar, R. C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797 (2004).
Nguyen, L.-T., Schmidt, H. A., von Haeseler, A. & Minh, B. Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 32, 268–274 (2015).
Hoang, D. T., Chernomor, O., von Haeseler, A., Minh, B. Q. & Vinh, L. S. UFBoot2: improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 35, 518–522 (2018).
Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313 (2014).
Matschiner, M. Fitchi: Haplotype genealogy graphs based on the Fitch algorithm. Bioinformatics 32, 1250–1252 (2016).
Berkunsky, I. et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 214, 278–287 (2017).
Xia, X. DAMBE6: new tools for microbial genomics, phylogenetics, and molecular evolution. J. Heredity 108, 431–437 (2017).
Caparroz, R., Miyaki, C. Y. & Baker, A. J. Contrasting phylogeographic patterns in mitochondrial DNA and microsatellites: Evidence of female philopatry and male-biased gene flow among regional populations of the Blue-and-yellow Macaw (Psittaciformes: Ara ararauna) in Brazil. Auk 126, 359–370 (2009).
Librado, P. & Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25, 1451–1452 (2009).
Pons, J.-M., Hassanin, A. & Crochet, P.-A. Phylogenetic relationships within the Laridae (Charadriiformes: Aves) inferred from mitochondrial markers. Mol. Phylogenet. Evol. 37, 686–699 (2005).
Behar, D. M. et al. A “Copernican” reassessment of the human mitochondrial DNA tree from its root. Am. J. Hum. Genet. 90, 675–684 (2012).
Weissensteiner, H. et al. Contamination detection in sequencing studies using the mitochondrial phylogeny. Genome Res. 31, 309–316 (2021).
Abu-Amero, K. K., Larruga, J. M., Cabrera, V. M. & González, A. M. Mitochondrial DNA structure in the Arabian Peninsula. BMC Evolut. Biol. 8, 45 (2008).
Brisighelli, F. et al. Uniparental markers of contemporary Italian population reveals details on its pre-Roman heritage. PLoS ONE 7, e50794 (2012).
Hernández, C. L. et al. The distribution of mitochondrial DNA haplogroup H in southern Iberia indicates ancient human genetic exchanges along the western edge of the Mediterranean. BMC Genet. 18, 46 (2017).
Översti, S. et al. Identification and analysis of mtDNA genomes attributed to Finns reveal long-stagnant demographic trends obscured in the total diversity. Sci. Rep. 7, 6193 (2017).
Fallon, S. J., Fifield, L. K. & Chappell, J. M. The next chapter in radiocarbon dating at the Australian National University: Status report on the single stage AMS. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 268, 898–901 (2010).
Hogg, A. G. et al. SHCal20 southern hemisphere calibration, 0–55,000 years cal BP. Radiocarbon 62, 759–778 (2020).
Ramsey, B. OxCal Program, Version 4.4.4 (Oxford Radiocarbon Accelerator Unit, University of Oxford, Oxford, 2021).
Thompson, D. R. & Furness, R. W. Stable-isotope ratios of Carbon and Nitrogen in feathers indicate seasonal dietary shifts in Northern Fulmars. Auk 112, 493–498 (1995).
Drtikolová Kaupová, S., Velemínský, P., Grossová, I., Půtová, L. & Cvrček, J. Stable isotope values of carbon and nitrogen in hair compared to bone collagen from individuals with known medical histories (Bohemia, 19th–21st centuries). Int. J. Osteoarchaeol. 32, 996–1010 (2022).
Kaulicke, P. Contextos Funerarios de Ancón: Esbozo de Una Síntesis Analítica (Fondo Editorial de la Pontificia Universidad Católica del Perú, Lima, 1997).
Rowe, J. H. The Kingdom of Chimor. Acta Am. 6, 27–59 (1948).
Segura, R. A. & Shimada, I. La interacción Sicán Medio-Costa central, hacia 1000 d.C. in Cultura Sicán: Esplendor preinca de la costa norte (ed. Shimada, I.) 303–322 (Editorial del Congreso de Perú, Lima, 2014).
Stumer, L. Contactos foráneos en la arquitectura de la costa central. Rev. del. Mus. Nacional 27, 11–30 (1958).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2025).
Kass, J. M. et al. Wallace: A flexible platform for reproducible modeling of species niches and distributions built for community expansion. Methods Ecol. Evol. 9, 1151–1156 (2018).
GBIF.org. GBIF Occurrence Download. The Global Biodiversity Information Facility https://doi.org/10.15468/dl.up84d4 (2023).
BirdLife International and HBW. Bird Species Distribution Maps of the World. http://datazone.birdlife.org/species/requestdis. (2021).
Leonardi, M., Hallett, E. Y., Beyer, R., Krapp, M. & Manica, A. pastclim 1.2: An R package to easily access and use paleoclimatic reconstructions. Ecography 2023, e06481 (2023).
Krapp, M., Beyer, R. M., Edmundson, S. L., Valdes, P. J. & Manica, A. A statistics-based reconstruction of high-resolution global terrestrial climate for the last 800,000 years. Sci. Data 8, 228 (2021).
Aiello-Lammens, M. E., Boria, R. A., Radosavljevic, A., Vilela, B. & Anderson, R. P. spThin: an R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 38, 541–545 (2015).
Phillips, S. Maxnet: Fitting’Maxent’ Species Distribution Models with’Glmnet’ (version R package 0.1.4). (2021).
Phillips, S. J., Anderson, R. P. & Schapire, R. E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 190, 231–259 (2006).
Elith, J. et al. A statistical explanation of MaxEnt for ecologists. Diversity Distrib. 17, 43–57 (2011).
Phillips, S. J., Anderson, R. P., Dudík, M., Schapire, R. E. & Blair, M. E. Opening the black box: an open-source release of Maxent. Ecography 40, 887–893 (2017).
Keighley, M. V., Langmore, N. E., Peñalba, J. V. & Heinsohn, R. Modelling dispersal in a large parrot: a comparison of landscape resistance models with population genetics and vocal dialect patterns. Landsc. Ecol. 35, 129–144 (2020).
Olah, G. et al. Exploring dispersal barriers using landscape genetic resistance modelling in Scarlet Macaws of the Peruvian Amazon. Landsc. Ecol. 32, 445–456 (2017).
QGIS Development Team. QGIS Geographic Information System (Open Source Geospatial Foundation, 2023).
EROS. Earth Resources Observation and Science Center: Global Multi-resolution Terrain Elevation Data 2010 (GMTED2010). https://doi.org/10.5066/F7J38R2N (2010).
Schielein, J. Potentially Navigable Rivers in South America. Harvard Dataverse https://doi.org/10.7910/DVN/1G8PZI (2017).
McRae, B. H. & Beier, P. Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. USA 104, 19885–19890 (2007).
Burger, R. L. Chavín and the Origins of Andean Civilization (Thames and Hudson, New York, 1992).
Watanabe, S. Continuidad cultural y elementos foráneos en Cajamarca, sierra norte del Perú: El caso del Horizonte Medio. Boletín de Arqueología PUCP 221–238 (2010) https://doi.org/10.18800/boletindearqueologiapucp.201001.011.
Disselhoff, H. D. Cajamarca-Keramik von der Pampa von San José de Moro (Prov. Pacasmayo). Baessler-Arch. 6, 181–194 (1958).
Nolan, J. L. Pre-Hispanic Irrigation and Polity in the Lambayeque Sphere, Peru (Department of Anthropology, Columbia University, Ann Arbor, 1980).
Brochado, J. J. P. & Lathrap, D. W. Chronologies in the New World: Amazonia. Unpublished manuscript. On file at the Department of Anthropology, University of Illinois at Urbana-Champaign. Revised by authors in 1984. (1982).
Bodley, J. H. Deferred exchange among the Campa: a reconsideration. in Networks of the past: Regional interaction in archaeology (eds Francis, P. D., Kense, F. J. & Duke, P. G.) 49–60 (Archaeology Association of the University of Calgary, Calgary, 1981).
Martin, Aj. J. The Dynamics of Pre-Columbian Spondylus Trade Across the South American Central Pacific Coast (Florida Atlantic University, Boca Raton, Florida, 2001).
Camino, A. Trueque, correrías e intercambios entre los Quechuas andinos y los Piro y Machiguenga de la montaña peruana. Amazon.ía Peru. 1, 123–140 (1977).
Santos-Granero, F. Etnohistoria de La Alta Amazonia: Siglos XV–XVIII. (AbyaYala, Quito, 1992).
Murra, J. V. The historical tribes of Ecuador. in Handbook of South American Indians. Vol. 2. The Andean Civilizations (ed. Steward, J. H.) 785–821 (Smithsonian Institution, Washington D.C., 1946).
Eriksen, L. Nature and culture in prehistoric Amazonia: Using G.I.S. to reconstruct ancient ethnogenetic processes from archaeology, linguistics, geography, and ethnohistory. in Lund Studies in Human Ecology Vol. 12 (Lund University, 2011).
Bonavia, D. The role of the ceja de selva in the cultural development of pre-Columbian Peru. in The Inca World: The Development of Pre-Columbian Peru, A.D. 1000–1534 (ed Minelli, L. L.) 121–131 (University of Oklahoma Press, Norman, 2000).
Church, W. B. Early occupations at Gran Pajatén, Peru. Andean 4, 281–318 (1994).
Guengerich, A. Settlement organization and architecture in Late Intermediate Period Chachapoyas, northeastern Peru. Lat. Am. Antiquity 26, 362–381 (2015).
Kauffmann Doig, F. Constructores de Kuelap y Pajaten: Los Chachapoya(s) (Derrama Magisterial, Lima, 2009).
Church, W. & Guengerich, A. La (re)construcción de Chachapoyas a través de la historia e histografía. Boletín de Arqueología PUCP 5–38 (2017) https://doi.org/10.18800/boletindearqueologiapucp.201702.000.
Schjellerup, I. Reflexiones sobre los chachapoya en el Chinchaysuyu. Boletín de Arqueología PUCP 43–56 (2002) https://doi.org/10.18800/boletindearqueologiapucp.200201.003.
Schjellerup, I. 1470 Inca expansion into the land of the Chachapoya: Archaeological research in the cloud forest on the eastern slopes of the Andes Mountains in Peru. Acta Archaeol. 90, 135–154 (2019).
Bradley, R. The Architecture of Kuelap (Columbia University Press, New York, 2005).
Narváez Vargas, A. Kuelap: Centro del poder político y religioso de los chachapoyas. in Los Chachapoyas (ed Kauffmann, F.) 87–160 (Banco de Crédito del Perú, Lima, 2013).
Carter, B. P. Technology, society and change: Shell artifact production among the Manteño (A.D. 800–1532) of coastal Ecuador. ProQuest Dissertations and Theses (Washington University in St. Louis, United States – Missouri, 2008).
Shimada, I. & Merkel, J. F. Naipes: Functions and significance of middle Sicán standardized sheetmetal artifacts. Ñawpa Pacha 41, 211–249 (2021).
Reeve, M. E. Regional interaction in the Western Amazon: The early colonial encounter and the Jesuit years: 1538-1767. Ethnohistory 41, 106–138 (1994).
Salomon, F. Native Lords of Quito in the Age of the Incas. The Political Economy of North Andean Chiefdoms (Cambridge University Press, Cambridge, UK, 1986).
Salomon, F. A North Andean Status Trader Complex under Inka Rule. Ethnohistory 34, 63–77 (1987).
Steward, J. H. & Métraux, A. Tribes of the Peruvian and Ecuadorian Montaña. in Handbook of South American Indians (ed. Steward, J. H.) Vol. 3. The Tropical Forest Tribes 535–656 (Smithsonian Institution, Washington D.C., 1948).
Shimada, I. The late prehispanic coastal states. in The Inca World: The Development of Pre-Columbian Peru, A.D. 1000–1534 (ed. Minelli, L. L.) 49–110 (University of Oklahoma Press, Norman, 2000).
Raymond, S. J. A view from the tropical forest. in Peruvian prehistory: An overview of pre-Inca and Inca society (ed Keatinge, R. W.) 279–300 (Cambridge University Press, Cambridge, 1988).
Acknowledgements
The 2005 fieldworks at Pachacamac by the Pachacamac Archaeological Project were supported by grants to Izumi Shimada from the National Science Foundation (BCS-0411625) and the National Geographic Society (7724-04). George Olah was funded by Australian Research Council DECRA Fellowship (DE230100085) and the Loro Parque Fundación (PP-250-2024). Pere Bover was funded by project PID2022-140091NB-I00 (Spanish Ministerio de Ciencia, Innovación y Universidades [MICIU/AEI/10.13039/501100011033] and FEDER, UE). We thank Hilary Stuart-Williams and Stewart Fallon for conducting and interpreting stable isotope testing and radiocarbon dating, respectively. We appreciate the help of Donald Brightsmith, Gabriela Vigo, Robert Heinsohn, Rod Peakall, Braulio Poje Mishaja, many other team members, and Rainforest Expeditions for their help with research administration and contemporary sample collection in Tambopata. We thank Go Matsumoto for the map of the site of Pachacamac (Fig. 1b), the Cornell Lab of Ornithology for permission of using their illustrations (Fig. 2), and Antonia Uguet for comments on the design of several graphics.
Author information
Authors and Affiliations
Contributions
G.O. and I.S. conceived the work. I.S. and R.S.L. conducted the archaeological research and collected ancient feathers. G.O. collected and extracted DNA from modern samples, and sampled feathers for aDNA analysis. P.B., H.H., and B.L. extracted and sequenced aDNA. G.O. performed spatial modeling, P.B. analyzed DNA results. G.O. and P.B. analyzed stable isotope results and prepared all visualization. I.S., R.S.L., and G.O. administered the project and acquired permits. I.S., G.O., and P.B. acquired fundings. I.S. and B.L. performed supervisory roles. All authors contributed to the writing of the paper and reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Aleksa Alaica and the other anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Olah, G., Bover, P., Llamas, B. et al. Ancient DNA and spatial modeling reveal a pre-Inca trans-Andean parrot trade. Nat Commun 17, 2117 (2026). https://doi.org/10.1038/s41467-026-69167-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-69167-9
