
Abstract
Male infertility is linked to chronic psychological stress, however, the mechanisms underlying chronic stress-induced oligoasthenozoospermia (OAZS) remain incompletely elucidated. Here, we establish a chronic forced swim stress (CFSS) rat model and identify a gut microbiota-testis/epididymis axis as the mediator. CFSS activates the HPA axis, leading to elevated corticosterone levels. The excessed glucocorticoid compromises gut barrier integrity via glucocorticoid receptor signaling, resulting in depletion of Lactobacillus and disruption of vitamins A/E metabolism. These deficits promote upregulation of Sting1. Concurrently, epididymal transcriptomics analysis reveals downregulation of Slc9c2 as the basis for impaired sperm motility. Notably, restoration of Lactobacillus and dietary supplementation with vitamins A/E ameliorate OAZS phenotypes. Genetic targeting Sting1/Slc9c2 in CFSS model rats treated with vitamins A/E, validating the involvement of vitamin A/E metabolism-Sting1/Slc9c2 axis in mediating CFSS-induced OAZS. These findings redefines OAZS as a multisystem disorder driven by gut microbial-metabolic-testis/epididymis axis dysregulation and proposes microbiota-targeted therapies for chronic stress-induced infertility.
Similar content being viewed by others


Introduction
Infertility is a prominent public health issue, affecting a substantial proportion of the global population, with male factors contributing to ~50% of cases1. It is known that oligoasthenozoospermia (OAZS), characterized by a concurrent reduction in sperm concentration (below 16 million sperm/ml) and total sperm motility (less than 42%), accounts for a substantial fraction of male infertility diagnoses2. A range of etiological factors, including genetic, environmental, and lifestyle-related factors, et al., have been implicated in OAZS; of which, chronic psychological stress has emerged as a potential key contributor to spermatogenic dysfunction3,4,5,6. Rodent models of chronic stress have successfully recapitulate key features of human OAZS, including impaired meiotic progression and disrupted spermatogenesis, these effects may extend beyond direct neuroendocrine pathways, involving systemic mediators7,8,9,10,11. However, the precise molecular mechanisms linking psychological stress perception to testicular damage remain poorly understood.
Chronic psychological stress activates the hypothalamic-pituitary-adrenal (HPA) axis, leading to elevated corticosterone (CORT) levels. This neuroendocrine response triggers inflammatory activation of enteric glia and transcriptional immaturity in enteric neurons, resulting in gastrointestinal dysmotility and creating a permissive environment for monocyte-mediated intestinal inflammation12,13. Intestinal inflammation can further disrupt the composition of the gut microbiota that is commonly referred to as dysbiosis. Emerging evidence positions the gut microbiota as a critical regulator of male fertility, orchestrating metabolic crosstalk between dietary components and spermatogenesis14,15. Stress-induced dysbiosis compromises intestinal barrier integrity, enabling microbial metabolites to translocate systemically and affect distant organs including the testes16,17,18. Notably, key microbiota-dependent metabolites such as bile acids and fat-soluble vitamins (e.g., vitamins A and E), which play essential roles in germ cell differentiation and antioxidant defense, are diminished in infertile men19,20,21,22. Despite these associations, whether stress-driven gut dysbiosis directly impairs spermatogenesis via metabolic collapse, and which specific microbial taxa may orchestrate this gut-testis axis, still remain unclear.
Chronic psychological stress has been shown to induce germ cell apoptosis and cell cycle arrest, contributing to spermatogenic failure. Conversely, supplementation with probiotics or prebiotics has been found to alleviate reproductive dysfunction in rodent models of chronic restraint stress, primarily by modulating oxidative stress and inflammation in the testes23,24. Metabolites derived from gut microbiota, such as vitamins A and E, are known to support normal spermatogenesis19,25. However, to date, no study has identified the specific genes that translate gut-derived signals into transcriptional programs governing spermatogenesis, representing a critical knowledge gap in understanding the molecular basis of the gut-testis axis.
The stimulator of interferon gene (STING) is an important innate immune mediator of the cytoplasmic DNA sensing pathway26. STING1-dependent signaling network is implicated in health and disease by regulating autophagic degradation or various cell death modalities such as apoptosis, necroptosis, pyroptosis, ferroptosis, mitotic cell death, and immunogenic cell death27,28,29,30. These findings raise the possibility of the involvement of STING1 signaling in stress-associated spermatogenesis depletion and oligozoospermia. In addition, the solute carrier family 9 (SLC9) gene family encodes Na+/H+ exchangers (NHEs), a group of ion transporters that regulate the pH of various cell compartments across an array of cell types, implicates in human physiology and disease31. Eight NHE isoforms are expressed in mammalian testis/sperm, including NHE1, NHE3, NHE5, NHE8, NHA1, NHA2, NHE10, and NHE11, of which, NHE3, NHE8, NHA1, NHA2, and NHE10 being critical for male fertility in mice and NHE10 has recently been shown to be essential for male fertility in humans32,33. SLC9C2, which encodes the NHE11 protein, has recently shown exhibits testis/sperm-restricted expression in rats and humans34. The physiological role of SLC9C2 (NHE11) has yet to be demonstrated but its predicted functional domains and unique localization suggests its potential roles in regulating sperm motility and male fertility via the modulation of intracellular pH of the sperm cells.
In this work, we characterize spermatogenic dysfunction and fecal microbiota alterations in a rat model of chronic forced swim (FS) stress (CFSS). We further profile metabolic patterns across gut, plasma, and testicular tissues to map systemic changes induced by CFSS. Our findings demonstrate the involvement of gut microbiota-vitamin A/E metabolism-Sting1/Slc9c2 (NHE11) genes axis in mediating chronic stress-induced oligoasthenozoospermia (OAZS). These results highlight a potential mechanistic pathway through which gut-derived signals influence testicular function and suggest novel therapeutic targets for male infertility driven by chronic psychological stress.
Results
Chronic stress induces anxiety- and depression-like behaviors, impairs spermatogenesis and causes oligoasthenozoospermia in male rats
To determine whether the chronic forced swim (FS) stress (CFSS) paradigm may serve as a reliable stressor in our experiments, we first assessed the activation of the HPA axis and the associated behavioral phenotypes in male rats exposed of chronic FS stress (Supplementary Fig. S1a). The results showed that CFSS significantly increased corticotropin-releasing hormone (CRH) content within the paraventricular nucleus (PVN) of hypothalamus in rats (in ng/mg, CFSS 25.64 ± 1.98 versus Sham FS 19.25 ± 1.37, p = 0.049, Supplementary Fig. S1b), accompanied by elevated plasma adrenocorticotropic hormone (ACTH) (in ng/ml, CFSS 218.60 ± 11.65 versus Sham FS 128.50 ± 16.50, p = 0.0033) and corticosterone (CORT) (in ng/ml, CFSS 351.80 ± 25.49 versus Sham FS 209.8 ± 14.70, p = 0.0037) levels compared to naïve and sham forced swim (FS) controls (Supplementary Fig. S1c, d), validating robust HPA axis activation. Next, we evaluated anxiety- and depression-like behaviors in CFSS-exposed rats. In the open field test (OFT), CFSS rats exhibited reduced exploratory activity, as evidenced by fewer entries into and shorter dwell time in the central zone, with total travelled distance not altered (Supplementary Fig. S1e–h). Similarly, in the elevated plus maze (EPM) test, CFSS rats exhibited decreased open arm entries and reduced time spent in open arms (Supplementary Fig. S1i–l), indicative of heightened anxiety-like behaviors. Besides, depression-like behaviors were also observed in rats exposure of CFSS, manifested as progressive weight loss from day 7 to day 21 (Supplementary Fig. S1m), diminished sucrose preference in the sucrose preference test (Supplementary Fig. S1n), as well as prolonged immobility in both forced swim test (Supplementary Fig. S1o) and tail suspension test (Supplementary Fig. S1p). These findings collectively affirm CFSS as an effective model for chronic psychological stress in male rats.
Given the established link between chronic stress and reproductive dysfunction, we next evaluate semen quality of CFSS-exposed male rats by using computer-assisted semen analysis (CASA)35. CFSS rats exhibited significant reductions in sperm concentration (in 10^6/ml, CFSS 9.21 ± 0.85 versus Sham FS 19.21 ± 1.24, p = 0.0096) and sperm motility, including rapid progressive motility, progressive and total motility (CFSS 31.14 ± 4.12% versus Sham FS 69.33 ± 3.80%, p < 0.0001), along with other parameters of sperm motility such as straight-line velocity (VSL), curve-line velocity (VCL), average path velocity (VAP), straightness (STR), linearity (LIN), amplitude of lateral head displacement (ALH) and wobble (WoB) (Fig. 1a–d, and Supplementary Fig. S2a–g). These results indicate the impaired reproductive function of CFSS-exposed male rats. To investigate the underlying defects in spermatogenesis of CFSS rats, we analyzed stage-specific germ cell markers in the testicular tissues via real-time quantitative polymerase chain reaction (RT-qPCR), immunofluorescence (IF) staining and Western blot techniques. Key markers including Dazl (spermatogonia), Sycp3 (spermatocytes), Tnp1 (spermatids), and Pgk2 (spermatozoa), which are essential for distinct phases of germ cell development36,37, were examined. While Dazl mRNA levels were unchanged, Sycp3, Tnp1, and Pgk2 mRNA expressions were significantly downregulated in the testicular tissues of CFSS rats (Fig. 1e–h), indicating disruptions in meiosis and spermiogenesis. IF staining revealed attenuated SYCP3 and TNP1 signals, along with reduced PGK2 expression in seminiferous tubules (Fig. 1i), consistent with Western blot data showing decreased protein level of SYCP3, TNP1, and PGK2, despite DAZL protein remained unchanged in the testicular tissues of CFSS rats (Fig. 1j–m). Functional in vivo fertility assays further confirmed reproductive impairment, e.g., female rats mated with CFSS males exhibited lower pregnancy rates, prolonged gestation period, and smaller litter sizes (in pups/litter, CFSS 6.50 ± 0.43 versus Sham FS 11.60 ± 0.50, p < 0.0001) (Supplementary Fig. S2h–k). Together, these findings demonstrate that CFSS disrupts multiple stages of spermatogenesis, leading to oligoasthenozoospermia and compromised fertility.

a–d Sperm concentration (a), rapid progressive motility (b), progressive motility (c), and total motility (d) of epididymal sperm across naïve, sham FS-, and CFSS-treated rats. (n = 7 rats in CFSS group, n = 6 in other groups). e–h Quantitative analysis of Dazl (e, n = 14), Sycp3 (f, n = 12), Tnp1 (g, n = 11), and Pgk2 (h, n = 10) mRNA expression levels in the testicular tissues. i Representative immunofluorescence images showing DAZL, SYCP3, TNP1, and PGK2 expression in seminiferous tubules of testicular sections (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 50 µm. j–m Relative protein expression levels of DAZL (j), SYCP3 (k), TNP1 (l), and PGK2 (m) in the testicular tissues (n = 4 rats per group). Data are presented as mean ± S.E.M. One-way ANOVA with Tukey’s post-hoc test for (a)–(h), and (j)–(m). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
Chronic stress induces oligoasthenozoospermia via the CORT/GR-mediated signaling pathway
Having validated the HPA axis hyperactivity in CFSS rats (Supplementary Fig. S1b–d), we next investigated whether corticosterone (CORT), the terminal effector of this axis, directly mediates testicular damage. Administration of exogeneous CORT to healthy adult male rats recapitulated the OAZS phenotype, causing significant declines in sperm concentration and sperm motility parameters (Supplementary Fig. S3a–k). Notably, co-treatment with the glucocorticoid receptor (GR) antagonist RU-48638 largely prevented these effects, implicating GR-dependent signaling in CORT-induced reproductive toxicity. At the molecular level, CORT treatment downregulated Sycp3, Tnp1, and Pgk2 mRNA expression in the testes without affecting Dazl mRNA expression (Supplementary Fig. S4a). IF staining and Western blot analyses further confirmed reduced SYCP3, TNP1, and PGK2 protein levels in the testicular tissues of CORT-treated rats, all of which were rescued by RU-486 (Supplementary Fig. S4b–d). Functionally, CORT exposure prolonged mating-to-birth intervals and reduced litter sizes, and all these effects were reversed by GR blockade (Supplementary Fig. S4e–h). These results suggest that CORT acts through GR to impair spermatogenesis and fertility.
To further determine whether inhibiting GR could ameliorate chronic stress-induced OAZS, CFSS rats were treated with RU-486. This intervention significantly improved sperm concentration and sperm motility parameters for CFSS rats (Supplementary Fig. S5a–k). Molecular analysis revealed upregulation of testicular Sycp3, Tnp1, and Pgk2 mRNA levels (Supplementary Fig. S6a), restored protein expression of testicular SYCP3, TNP1, and PGK2 (Supplementary Fig. S6b–d), and normalized fertility outcomes, including pregnancy rates, mating-to-birth intervals, and litter sizes, in CFSS rats treated with GR antagonist RU-486 (Supplementary Fig. S6e–l). Collectively, these data establish that CORT/GR signaling is a central medicator of chronic stress-induced OAZS.
Chronic stress induces oligoasthenozoospermia through gut microbiota dysbiosis
Given the high glucocorticoid sensitivity of gastrointestinal tract39,40, we examined intestinal barrier integrity in CFSS rats. Quantitative analysis revealed disrupted intestinal tight junctions, with significant reductions in mRNA and protein levels of Zo-1, Occludin, and Claudin-1 (Supplementary Fig. S7a–i). IF staining showed discontinuous localization of these proteins at epithelial junctions (Supplementary Fig. S7d–f). Importantly, RU-486 treatment reversed these impairments, indicating that corticosterone (CORT)-mediated glucocorticoid receptor (GR) activation underlies the chronic stress-induced impairment of intestinal barrier integrity. To further determine whether gut microbiota alterations contribute to the observed OAZS phenotype, we performed fecal microbiota transplantation (FMT) from CFSS- or sham FS-exposed donors into antibiotic-depleted recipients (Fig. 2a). Recipients of CFSS-derived microbiota (CFSS-FMT) exhibited significant reductions in sperm concentration (in 10^6/ml, CFSS-FMT 8.48 ± 1.94 versus Sham-FMT 14.70 ± 1.16, p = 0.025) and sperm motility, including rapid progressive motility, progressive motility, total motility (CFSS-FMT 67.20 ± 5.13% versus Sham-FMT 84.00 ± 1.38%, p = 0.0134), as well as other parameters of sperm motility such as VSL, VCL, VAP, STR, LIN and ALH (Fig. 2b–e, Supplementary Fig. S8a–f), mirroring donor phenotypes. RT-qPCR analysis revealed downregulation of Sycp3, Tnp1 and Pgk2 mRNA levels, but not Dazl mRNA in testicular tissues (Fig. 2f–i). Similarly, IF staining and Western blot analyses verified decreased protein expressions of testicular SYCP3, TNP1, and PGK2 in CFSS-FMT recipients (Fig. 2j–n). Consistently, fertility assessment revealed prolonged mating-to-birth intervals, reduced pregnancy rates, and smaller litter sizes (in pups/litter, CFSS-FMT 8.83 ± 0.60 versus Sham-FMT 12.40 ± 0.62, p = 0.0018) in CFSS-FMT pairs (Supplementary Fig. S8g–j). These findings identify the involvement of gut microbiota dysbiosis in chronic stress-induced reproductive dysfunction, suggesting the implication of gut-microbiota-dependent axis in chronic stress-induced male infertility.

a Schematic diagram of the fecal microbiota transplantation (FMT) procedure, illustrating the transfer of gut microbiota from CFSS- or sham FS-donor rats to antibiotic-treated recipient rats. Created in BioRender. Zheng J. (https://BioRender.com/yms9k64). b–e Sperm concentration (b), rapid progressive motility (c), progressive motility (d), and total motility (e) of CFSS-FMT and sham FS-FMT groups (n = 5 rats per group). f–i Quantitative analysis of Dazl (f), Sycp3 (g), Tnp1 (h), and Pgk2 (i) mRNA expression levels in the testicular tissues (n = 7 rats per group). j Representative immunofluorescence images of DAZL, SYCP3, TNP1, and PGK2 signals in the seminiferous tubules of testicular sections (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 50 µm. k–n Relative protein expression levels of DAZL (k), SYCP3 (l), TNP1 (m), and PGK2 (n) in the testicular tissues (n = 4 rats per group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test for (b)–(i), and (k)–(n). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
Lactobacillus depletion contributes to chronic stress-induced oligoasthenozoospermia
To identify the key microbial taxa contributing to CFSS-induced OAZS, we performed 16S rDNA sequencing of small intestine digesta across experimental groups. Rarefaction curve indicated sufficient sequencing depth (Fig. 3a), and Bray-Curtis based principal component analysis (PCA) revealed a distinct clustering of microbiota composition between the CFSS and the sham FS group (Fig. 3b). To assess the overall composition of the bacterial community in different groups, the degree of bacterial taxonomic similarity of gut microbiota was analyzed. Microbial richness (Chao1 index) and diversity (Shannon index) of the gut microbiota were significantly reduced in the CFSS rats (Fig. 3c–e). At the phylum level, relative abundance of phylum Firmicutes was substantially decreased (Fig. 3f), and at the genus level, Lactobacillus was significantly depleted (in relative abundance * 100, CFSS 0.014 ± 0.002 versus Sham FS 0.031 ± 0.004, p = 0.0143, Fig. 3g, h). At the species level of Lactobacillus, relative abundance of Lactobacillus reuteri was significantly reduced (in relative abundance * 100, CFSS 0.085 ± 0.031 versus Sham FS 0.473 ± 0.094, p = 0.0143, Fig. 3i, j). Furthermore, Pearson correlation analysis revealed strong positive correlation between Lactobacillus abundance and sperm concentration, rapid progressive motility, and progressive motility (Fig. 3k–m), suggesting a protective role of Lactobacillus in chronic stress-induced oligoasthenozoospermia. The reliability of the animal model was confirmed by consistent microbiome profiling across two independent batches (Supplementary Fig. S9). To test this hypothesis, CFSS rats received daily oral supplementation with Lactobacillus (Lacto-suppl) during the 21-day stress protocol (Fig. 4a). Lactobacillus (Lacto) treatment restored sperm concentration (in 10^6/ml, Lacto 12.41 ± 0.58 versus Vehicle 8.49 ± 0.74, p = 0.0013) and sperm motility, including rapid progressive motility, progressive motility, total motility (Lacto 76.00 ± 4.60% versus Vehicle 58.29 ± 4.55%, p = 0.018), as well as other parameters of sperm motility including VSL, VCL, VAP, STR and LIN (Fig. 4b–e, Supplementary Fig. S10a–f). The mRNA levels of Sycp3, Tnp1 and Pgk2 in testicular tissues were upregulated, while Dazl remained unchanged in Lacto-suppl rats (Fig. 4f–i). Consistently, IF staining and Western blot analyses confirmed the recovery of SYCP3, TNP1, and PGK2 protein in testicular tissues of Lacto-suppl rats (Fig. 4j–n). Also, the in vivo fertility assays showed normalized pregnancy rates, shortened mating-to-birth intervals, and near-physiological litter sizes (in pups/litter, Lacto 10.83 ± 0.60 versus Vehicle 6.33 ± 0.88, p = 0.0037) (Supplementary Fig. S10g–j). These results position Lactobacillus as a critical modulator of chronic stress-induced reproductive decline.

a Rarefaction curves depicting α-diversity of 16S rDNA sequencing data. b Principal component analysis (PCA) score plot based on Bray–Curtis dissimilarity, showing separation of microbial communities among naïve, sham forced swim (FS)-, and chronic forced swim stress (CFSS)-treated rats. c Chao1 index of gut microbiota across the three groups (n = 6 rats in Sham FS group, n = 5 in other groups). d Shannon diversity index across the three groups (n = 6 rats in Sham FS group, n = 5 in other groups). e Rank abundance curves among the three groups. f Rank abundance at the phylum level among the three groups. g Heatmap depicting relative abundances of bacterial genera, with significant differences observed across the three groups. h Relative abundance of Lactobacillus, which was statistically reduced in CFSS rats compared to controls (n = 6 rats in Sham FS group, n = 5 in other groups). i Heatmap depicting relative abundance of different Lactobacillus species, with significant differences among the three groups. j Relative abundance of Lactobacillus reuteri, which was statistically reduced in CFSS rats compared to controls (n = 6 rats in Sham FS group, n = 5 in other groups). k–m Pearson correlation analyses showing positive associations between Lactobacillus abundance and sperm concentration (k), rapid progressive motility (l), and progressive motility (m), respectively (n = 5 rats). Data are presented as mean ± S.E.M. One-way ANOVA with Tukey’s post-hoc test for (c), (d), (h) and (j). For (c), (d), (h), and (j), the middle bar represents the median, the box represents the interquartile range, and bars extend to the minimum and maximum values. Two-tailed t test for (k)–(m). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.

a Schematic illustration of the experimental design for Lactobacillus supplementation in CFSS rats. Created in BioRender. Zheng J. (https://BioRender.com/yqasmzm). b–e Sperm concentration (b), rapid progressive motility (c), progressive motility (d), and total motility (e) of CFSS rats treated with Lactobacillus or vehicle control (n = 7 rats per group). f–i Quantitative analysis of Dazl (f, n = 8), Sycp3 (g, n = 7), Tnp1 (h, n = 7), and Pgk2 (i, n = 8) mRNA expression levels in the testicular tissues of the two groups. j Representative immunofluorescence images of DAZL, SYCP3, TNP1, and PGK2 in the seminiferous tubules testicular sections of the two groups (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 50 µm. k–n Relative protein expression of DAZL (k), SYCP3 (l), TNP1 (m), and PGK2 (n) signals in testicular tissues (n = 4 rats per group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test for (b)–(i), and (k)–(n). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
Chronic stress disrupts bile acid and fat-soluble vitamins metabolism in small intestine digesta, plasma, and testicular tissues in rats
The gut microbiota plays a vital role in regulating spermatogenesis by interfering host metabolism14. Therefore, we performed liquid chromatography–mass spectrometry (LC-MS)-based metabolomic profiling of small intestine digesta, plasma and testicular tissues in CFSS- and sham FS-treated rats. Orthogonal partial least squares-discriminant analysis (OPLS-DA) revealed a clear separation of metabolite in small intestine digesta, plasma, and testicular tissues between the two groups (Fig. 5a–c), with widespread metabolic changes (Supplementary Fig. S11a–c). Meanwhile, KEGG pathway enrichment highlighted disruptions in vitamin digestion and absorption (small intestine digesta and plasma), retinol metabolism and primary bile acid biosynthesis (plasma), and amino acid metabolism (testicular tissues) (Fig. 5d–f). Specifically, the level of 9-cis-retinol (vitamin A) and vitamin E was respectively increased and decreased in the small intestine digesta of CFSS rats (Fig. 5g–h). In line with these findings, the relative abundance of key bile acids, including cholic acid (CA), chenodeoxycholic acid (CDCA), ursodeoxycholic acid (UDCA), and gamma-muricholic acid (γMCA), as well as vitamin A and E, were significantly decreased in the plasma of CFSS rats (Fig. 5i–n). Whereas the relative abundance of other vitamin such as vitamin D and K in the small intestine digesta (Supplementary Fig. S11d–g) and plasma (Supplementary Fig. S11h–k) remained unchanged. Given the role of bile acids in fat-soluble vitamin transportation and absorption41, we examined the content of bile acids and fat-soluble vitamins in the testicular tissues and found that the relative abundance of UDCA, vitamin D, and vitamin A was reduced in CFSS rats (Fig. 5o–r). These findings highlight systemic disruption of bile acids–fat-soluble vitamin homeostasis in chronic stress-induced OAZS rats.

a–c Orthogonal partial least squares-discriminant analysis score plots (OPLS-DA) score plots showing clear separation of metabolite profiles in the gut (a), plasma (b) and testis (c) between CFSS- and sham FS-treated rats. d–f KEGG pathway enrichment analysis of differentially abundant metabolites in the gut (d), plasma (e) and testis (f), highlighting alterations in vitamin digestion and absorption, retinol metabolism, and bile acid biosynthesis pathways. g–n Violin plots depicting the relative abundance of 9-cis-retinol (vitamin A) and vitamin E nicotinate in gut contents, as well as plasma bile acids, including cholic acid (CA), chenodeoxycholic acid (CDCA), ursodeoxycholic acid (UDCA), and gamma-muricholic acid (γMCA), and as well as retinal and vitamin E in plasma, in CFSS and sham FS rats (n = 10 rats per group). o–r Violin plots showing testicular levels of UDCA, 3-epi-25-hydroxyvitamin D3, 1α,2,25-trihydroxyvitamin D3, and vitamin A in CFSS and sham FS rats (n = 4 rats per group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test with Benjamini-Hochberg correction for (d–f). Two-tailed unpaired t test for (g–r). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
The Lactobacillus–bile acids–vitamin A/E axis mediates chronic stress-induced oligoasthenozoospermia
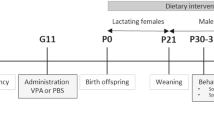
Given the above observations, we tested whether Lactobacillus supplementation in CFSS rats restores vitamin metabolism. Indeed, Lactobacillus treated-CFSS rats showed normalized vitamin A/E levels in both plasma (in ng/ml, for Vitamin A: Lactobacillus 0.56 ± 0.08 versus Vehicle 0.31 ± 0.06, p = 0.0422; for Vitamin E: Lactobacillus 6.26 ± 1.16 versus Vehicle 3.06 ± 0.59, p = 0.0391, Supplementary Fig. S11l, m) and testicular tissues (in ng/mg, for Vitamin A: Lactobacillus 156.40 ± 24.33 versus Vehicle 70.47 ± 15.89, p = 0.0182; for Vitamin E: Lactobacillus 2.92 ± 0.47 versus Vehicle 1.51 ± 0.32, p = 0.0378, Supplementary Fig. S11n, o), indicating the restoration of malabsorption. To directly evaluate the functional impact of vitamin A/E deficiency upon spermatogenesis and sperm motility in stressed rats, CFSS rats were supplemented with vitamin A palmitate and vitamin E acetate (Fig. 6a). This intervention restored systemic vitamin A and vitamin E levels in plasma (Supplementary Fig. S12a–b) and testis tissue (Supplementary Fig. S12c–d), and improved sperm concentration (in 10^6/ml, Vitamin A&E 10.97 ± 1.02 versus Vehicle 7.42 ± 1.03, p = 0.0282, Fig. 6b) and sperm motility, including rapid progressive motility, progressive motility, total motility (Vitamin A&E 76.00 ± 2.75% versus Vehicle 55.38 ± 5.57%, p = 0.0051) (Fig. 6c-e), as well as other parameters of sperm motility (Supplementary Fig. S12e–j). Molecular analyses revealed upregulated Sycp3, Tnp1, and Pgk2 mRNA expression (Fig. 6f–i), with restored SYCP3, TNP1 and PGK2 protein expression (Fig. 6j–n), while Dazl remain unaffected in the testicular tissues of vitamin A&E supplemented CFSS rats. Besides, fertility outcomes were also improved in vitamin A&E-supplemented CFSS rats, with shorter mating-to- birth intervals (Supplementary Fig. S12k), higher pregnancy rates (Supplementary Fig. S12l), and larger litter sizes (in pups/litter, Vitamin A&E 9.67 ± 0.49 versus Vehicle 6.33 ± 0.33, p = 0.0031, Supplementary Fig. S12m, n). Despite the critical role of bile acids in vitamin transport41, supplementation with glycoursodeoxycholic acid (GUDCA) did not improve sperm quality in CFSS rats (Supplementary Fig. S13a–j), suggesting that circulating bile acid levels may not be the limiting factor. Moreover, correlation analyses revealed that Lactobacillus abundance positively correlated with plasma CA, CDCA, UDCA, and γMCA (Supplementary Fig. S14a–d), and that plasma bile acids and vitamins A/E were positively associated with each other and with sperm motility parameters (Supplementary Fig. S14e–g). These results raise the possibility that Lactobacillus depletion leads to impaired bile acid–vitamin metabolism, thereby contributing to chronic stress-induced OAZS.

a Schematic diagram depicting the experimental design of Vitamin A&E supplementation in CFSS rats. Created in BioRender. Zheng J. (https://BioRender.com/2x9nxx2). b–e Sperm concentration (b), rapid progressive motility (c), progressive motility (d), and total motility (e) of CFSS rats supplemented with vitamin A&E or vehicle control (n = 8 rats per group). f–i Quantitative analysis of Dazl (f, n = 7), Sycp3 (g, n = 7), Tnp1 (h, n = 8), and Pgk2 (i, n = 7) mRNA expression levels in the testicular tissues. j Representative immunofluorescence images of DAZL, SYCP3, TNP1, and PGK2 signals in seminiferous tubules of testicular sections (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 50 µm. k–n Relative protein expression levels of DAZL (k), SYCP3 (l), TNP1 (m), and PGK2 (n) in the testicular tissues of CFSS rats supplemented with vitamin A&E or vehicle control (n = 4 rats per group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test for (b)–(i), and (k)–(n). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
The Lactobacillus–bile acids–vitamin A/E–Sting1 axis mediates chronic stress-induced oligozoospermia
To clarify the molecular mechanisms underlying the downstream targets of Lactobacillus-bile acids-vitamin A/E axis in chronic stress-induced oligozoospermia, we performed transcriptome sequencing (RNA-sequencing, RNA-seq) on testicular tissues from sham FS- and CFSS-treated rats. Principal component analysis (PCA) revealed clear transcriptomic divergence between the two groups (PC1: 42.57%, Fig. 7a), with 33 upregulated and 49 downregulated genes (Supplementary Fig. S15a). Hierarchical clustering heatmap revealed distinct transcriptional profiles (Fig. 7b). Among differentially expressed genes (DEGs), stimulator of interferon genes (Sting1) stood out due to its marked upregulation in Hierarchical clustering heatmap and volcano plot analyses (Fig. 7b, c). Gene Ontology (GO) enrichment analysis demonstrated that DEGs were significantly enriched in retinol metabolism and apoptotic signaling (Supplementary Fig. S15b). KEGG pathway analysis further associated these genes with TNF signaling, NOD-like receptor signaling, and JAK-STAT signaling (Supplementary Fig. S15c)42,43,44. Validation analyses showed increased Sting1 mRNA (CFSS 1.33 ± 0.16 versus Sham FS 0.66 ± 0.04, p = 0.0023, Fig. 7d), elevated STING1 protein expression (CFSS 1.26 ± 0.05 versus Sham FS 0.72 ± 0.03, p < 0.0001, Fig. 7e), and intensified immunofluorescence signal in testicular tissues of CFSS rats (Fig. 7f). Functional validation confirmed its causal roles in CFSS-induced spermatogenesis depletion and its associated oligozoospermia, e.g., overexpression of Sting1 using lentivirus expressing Sting1 (LV-Sting1) in the testicular tissues of naïve rats induced significant spermatogenesis depletion and oligozoospermia (Supplementary Fig. S16), whereas knocking-down Sting1 using lentivirus expressing Sting1 short hairpin RNA (LV-shSting1) in the testicular tissues of CFSS rats partially restored the oligozoospermia phenotype (Supplementary Fig. S17). Importantly, both Lactobacillus and vitamin A/E supplementation restored the CFSS-induced upregulation of Sting1 expression (Fig. 7g–l) at mRNA (for Lactobacillus: Lacto 0.35 ± 0.03 versus Vehicle 1.62 ± 0.20, p = 0.0008; for Vitamin A&E: Vitamin A&E 0.65 ± 0.15 versus Vehicle 1.60 ± 0.33, p = 0.0412, Fig. 7g, j) and protein levels (for Lactobacillus: Lacto 0.64 ± 0.05 versus Vehicle 1.33 ± 0.15, p = 0.0057; for Vitamin A&E: Vitamin A&E 0.66 ± 0.10 versus Vehicle 1.28 ± 0.10, p = 0.0049, Fig. 7h, k), linking microbial and metabolic rescue to transcriptional normalization.

a Two-dimensional principal component analysis (PCA) score plot showing transcriptomic separation between the CFSS and sham FS groups based on testicular RNA-seq data. b Heatmap showing hierarchical clustering of differentially expressed genes (DEGs) in the testis tissues between the CFSS and sham FS rats. c Volcano plot illustrating the DEGs between the two groups, with Sting1 highlighted as significantly upregulated in CFSS rats. d Increased expression of Sting1 mRNA level in the testicular tissues of CFSS rats (n = 6 rats per group). e Elevated expression of STING1 protein in the testicular tissues of CFSS rats (n = 4 rats per group). f Representative immunofluorescence images showing enhanced STING1 signal in the seminiferous tubules of CFSS rats (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars=50 µm. g Reduced Sting1 mRNA expression in testicular tissues of Lactobacillus-treated CFSS rats compared to vehicle-treated counterparts (n = 4 per group). h Decreased STING1 protein levels in the testes of Lactobacillus-supplemented CFSS rats (n = 4 per group). i Representative immunofluorescence images revealing attenuated STING1 staining in seminiferous tubules of Lactobacillus-treated CFSS rats (n = 3 biological replicates per group). j Downregulation of Sting1 mRNA in testicular tissues of vitamin A/E-supplemented CFSS rats (n = 6 per group). k Reduced STING1 protein expression in the testes of vitamin A/E-treated CFSS rats (n = 4 per group). l Representative immunofluorescence images showing diminished STING1 signal in seminiferous tubules of vitamin A/E-treated CFSS rats (n = 3 biological replicates per group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test with Benjamini-Hochberg correction for (c).Two-tailed unpaired t test for (d), (e), (g), (h), (j) and (k). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
To further establish a direct link across plasma vitamin A/E, testicular Sting1, and spermatogenesis, we first employed Sting1 overexpression in the testis of CFSS rats supplemented with vitamin A/E (Fig. 8a, hereafter referred as LV-Sting1-treated rats). Viral transduction efficiency was validated by increased Sting1 mRNA and protein levels (Fig. 8b–c), along with disrupted seminiferous tubule architecture and enhanced protein level of STING1 (Fig. 8d) in testicular tissues of LV-Sting1-treated rats. Notably, overexpression of Sting1 in the testicular tissues effectively counteracted the improved effects of vitamin A/E supplementation upon CFSS-induced spermatogenesis depletion and oligozoospermia. For example, Sting1 overexpression downregulated Dazl and Tnp1 expression at both mRNA and protein levels in the testicular tissues (Fig. 8e–g), and disrupted spermatogenesis characterized by a reduction of sperm concentration in vitamin A/E supplemented CFSS rats (in 10^6/ml, LV-Sting1 21.55 ± 0.57 versus LV-EGFP 27.09 ± 0.73, p = 0.0003, Fig. 8h). Nevertheless, LV-Sting1 did not affect the sperm motility of CFSS rats supplemented with vitamin A/E, including rapid progressive motility, progressive motility, and total motility (Fig. 8i–k), indicating the specific role of Sting1 in testicular spermatogenesis. Consistently, LV-Sting1-treated males displayed decreased fertility outcomes including decreased pregnancy rates and litter sizes (in pups/litter, LV-Sting1 11.80 ± 1.11 versus LV-EGFP 15.83 ± 0.48, p = 0.0062), although mating-to-birth intervals was not altered (Fig. 8l–o). Additionally, negative correlations between plasma vitamin A/E and testicular Sting1 mRNA, as well as between Sting1 and sperm concentration, further support the negative regulatory role of Sting1, activated by plasma vitamin A/E deficiency, in testicular spermatogenesis (Supplementary Fig. S14h–i). Together with the aforementioned evidence of the correlation between gut Lactobacillus and plasma bile acids–vitamin A/E, these data identify the Lactobacillus–bile acids–vitamin A/E-Sting1 axis as a key regulator of chronic stress-induced spermatogenesis depletion and oligozoospermia.

a Schematic illustration of the experimental strategy for testicular Sting1 overexpression using lentiviral vectors expressing Sting1 (LV-Sting1) in vitamin A/E-treated CFSS rats. LV-EGFP was used as a control. Created in BioRender. Zheng J. (https://BioRender.com/ujfo5sg). b, c Increased expression of Sting1 mRNA (b, n = 4) and protein (c, n = 5) levels in the testicular tissues of LV-Sting1 group. d Representative immunofluorescence images of STING1 and EGFP signals in the seminiferous tubules in the LV-Sting1 and LV-EGFP groups (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 50 µm. e Quantitative analysis of Dazl, Sycp3, Tnp1, and Pgk2 mRNA expression levels in testicular tissues of the two groups (n = 4 rats per group). f–g Relative expression of DAZL, SYCP3, TNP1, and PGK2 protein levels in testicular tissues of the two groups (n = 4 rats per group). h–k Sperm concentration (h), rapid progressive motility (i), progressive motility (j), and total motility (k) of rats between the groups (n = 5 rats per group). l Days to birth of female rats mated to male rats between the two groups (n = 5 female rats in LV-EGFP group, n = 6 female rats in LV-Sting1 group). m Pie charts illustrating pregnancy rates in female rats across groups. n, o Representative image (n) and summary plot (o) of litter size (number of pups per litter, n = 5 female rats in LV-EGFP group, n = 6 female rats in LV-Sting1 group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test for (b), (c), (e), (g)–(l), and (o). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
The Lactobacillus–bile acids–vitamin A/E–Slc9c2 (NHE11) axis governs chronic stress-induced asthenozoospermia
Since Sting1 overexpression did not affect the improved sperm motility of CFSS rats supplemented with vitamin A/E, and no sperm motility-related DEGs were detected in testicular tissues RNA-seq, we turned to examine epididymal sperm transcriptome. RNA-seq of corpus epididymis cells revealed DEGs linked to sperm motility (Fig. 9). PCA revealed pronounced transcriptional divergence between the CFSS and sham FS groups along PC1 (Fig. 9a), with 2,395 upregulated and 841 downregulated genes (Supplementary Fig. S15d). Hierarchical clustering heatmap revealed distinct expression profiles (Fig. 9b), and the volcano plot highlighted Slc9c2, a gene encodes the sperm-specific sodium/hydrogen exchanger NHE11, which is essential for sperm pH and calcium homeostasis36, positioning among the significantly downregulated gene participated in sperm function (Fig. 9c). Functional enrichment analysis further linked these transcriptional changes to sperm motility defects. GO analysis demonstrated enrichment in regulation of calcium ion transport, cation transmembrane transport, flagellar dynamics, calcium ion binding, voltage-gated calcium channel activity, and microtubule binding (Supplementary Fig. S15e). KEGG pathway analysis validated these findings, revealing perturbations in calcium, cAMP, and cGMP-PKG signaling pathways (Supplementary Fig. S15f), the key pathways governing sperm hyperactivation and motility45,46,47. Furthermore, IF staining revealed species-specific localization of NHE11 expression, head-specific in human sperm (Supplementary Fig. S18a), flagellar-restricted in mouse sperm (Supplementary Fig. S18b), and dual head and flagellum expression in rat sperm (Supplementary Fig. S18c), consistent with roles in acrosome reaction and sperm motility. In epididymal sperm of CFSS rats, Slc9c2 mRNA (CFSS 0.61 ± 0.06 versus Sham FS: 1.24 ± 0.14, p = 0.0025, Fig. 9d) and protein (NHE11, CFSS 0.55 ± 0.05 versus Sham FS 1.09 ± 0.05, p = 0.0003, Fig. 9e) levels were significantly reduced (Fig. 9d–f), accompanied by impaired hyperactivation (Supplementary Fig. S18d) and acrosome reaction (Supplementary Fig. S18e–g). With respect to its functional role in sperm motility, knockdown of Slc9c2 in naïve rats via intratesticular injection of lentivirus expressing Slc9c2 short hairpin RNA (LV-shSlc9c2) recapitulated the sperm motility defects seen in CFSS rats (Supplementary Fig. S19), whereas overexpression of Slc9c2 in CFSS rats via lentivirus expressing Slc9c2 (LV-Slc9c2) restored the CFSS-induced impairment of progressive motility and fertility (Supplementary Fig. S20). Furthermore, the expression of NHE11 was investigated in Lactobacillus- and vitamin A/E-supplemented CFSS rats (Fig. 9g–j). Both Lactobacillus and vitamin A/E supplementation restored the CFSS-induced reduction of NHE11 protein levels in epididymal sperm of CFSS rats (for Lactobacillus: Lacto 1.49 ± 0.22 versus Vehicle 0.51 ± 0.11, p = 0.0165, Fig. 9g; for Vitamin A&E: Vitamin A&E 1.45 ± 0.16 versus Vehicle 0.51 ± 0.03, p = 0.0045, Fig. 9i). Positive correlations between plasma vitamin A/E and epididymal Slc9c2 expression, as well as between Slc9c2 expression and sperm motility, further support the positive regulatory role of Lactobacillus-vitamin A/E-Slc9c2 (NHE11) axis in sperm motility (Supplementary Fig. S14j–l). Taken together, these data suggest that the dysfunction of Lactobacillus-vitamin A/E-Slc9c2 (NHE11) axis likely impairs epididymal sperm motility, thereby playing an important role in chronic stress-induced asthenozoospermia.

a PCA score plot showing transcriptomic separation between the CFSS and sham FS groups based on RNA-seq data from corpus epididymal tissues. b Heatmap depicting hierarchical clustering of DEGs in epididymal tissues from CFSS and sham FS rats. c Volcano plot showing DEGs between the two groups. d Relative expression of Slc9c2 mRNA in epididymal sperm of the two groups (n = 6 rats per group). e Relative expression of NHE11 protein in epididymal sperm of the two groups (n = 4 rats per group). f Representative immunofluorescence images of NHE11 and peanut agglutinin (PNA) staining in epididymal sperm of CFSS and sham FS rats (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 20 µm. g, h Relative expression of NHE11 protein (g) and representative images showing the immunofluorescence staining with NHE11 (h) in epididymal sperm of vehicle- and Lactobacillus-treated CFFS rats (n = 3 rats or 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 20 µm. i, j Relative expression of NHE11 protein (i) and representative images showing the immunofluorescence staining with NHE11 (j) in epididymal sperm of vehicle- and vitamin A/E-treated CFFS rats (n = 3 rats or 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 20 µm. Data are presented as mean ± S.E.M. Two-tailed unpaired t test with Benjamini-Hochberg correction for (c). Two-tailed unpaired t test for (d), (e), (g) and (i). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
To establish a direct link across plasma vitamin A/E, epididymal sperm Slc9c2, and chronic stress-induced asthenozoospermia, we knocked down Slc9c2 in vitamin A/E-supplemented CFSS rats (hereafter referred as LV-shSlc9c2-treated rats, Fig. 10a). Viral transduction efficiency was validated by reduced Slc9c2 mRNA and NHE11 protein levels (Fig. 10b–c, Supplementary Fig. S19b–d) in the epididymal sperm of LV-shSlc9c2-infected rats. Notably, knockdown of Slc9c2 effectively counteracted the improved effects of vitamin A/E supplementation upon CFSS-induced sperm motility impairment. For instance, Slc9c2 knockdown led to reduced sperm motility, including rapid progressive motility, progressive motility, and total motility (LV-shSlc9c2 53.46 ± 1.45% versus LV-EGFP 77.88 ± 1.24%, p < 0.0001), but not affected sperm concentration (in 10^6/ml, LV-shSlc9c2 23.80 ± 2.11 versus LV-EGFP 26.82 ± 2.30, p = 0.3713) in vitamin A/E-supplemented CFSS rats (Fig. 10d–g), indicating the specific role of Slc9c2 in sperm motility. Unexpectedly, fertility assays revealed that the mating-to-birth intervals, pregnancy rates, and litter sizes (in pups/litter, LV-shSlc9c2 15.40 ± 0.93 versus LV-EGFP 16.40 ± 0.51, p = 0.3724) were not different between the two groups (Fig. 10h–k), probably due to other compensatory mechanisms. In view of the fact that single supplementation of Lactobacillus or vitamin A/E only partially rescued the CFSS-induced sperm impairment and fertility deficiency in CFSS rats (Supplementary Fig. S10 and Fig. S12), we performed additional experiments to investigate whether combination therapies with Lactobacillus plus vitamin A/E supplementation have a complete rescue effect upon CFSS-induced sperm impairment and fertility deficiency. The results showed that combination therapies with Lactobacillus plus vitamin A/E supplementation still partially restored the CFSS-induced sperm impairment and fertility deficiency, and failed to fully normalize fertility (Supplementary Fig. S21). We speculate that either insufficient restoration of key pathways or the existence of alternative mechanisms contributing to chronic stress-induced infertility. Collectively, our findings delineate two parallel axes: the Lactobacillus–bile acids–vitamin A/E–Sting1 pathway regulating testicular spermatogenesis, and the Lactobacillus–bile acids–vitamin A/E–Slc9c2 (NHE11) axis governing epididymal sperm motility, both of which are disrupted by chronic psychological stress, thereby leading to oligozoospermia and asthenozoospermia, respectively.

a Schematic illustration of the experimental approach for Slc9c2 knockdown in testicular tissues of vitamin A/E-treated CFSS rats using lentiviral vectors expressing Slc9c2-targeting short hairpin RNA (LV-shSlc9c2). Created in BioRender. Zheng J. (https://BioRender.com/43l8ygf). b Relative expression of NHE11 protein in epididymal sperm of rats across the two groups (n = 3 rats per group). c Representative immunofluorescence images of NHE11 and EGFP signals in epididymal sperm of rats in the two groups (n = 3 biological replicates per group). Nuclei were counterstained with DAPI (blue). Scale bars = 20 µm. d–g Sperm concentration (d), rapid progressive motility (e), progressive motility (f), and total motility across the two groups (n = 4 rats per group). h Days to birth of female rats mated to male rats across the two groups (n = 5 female rats per group). (i) Pie charts illustrating pregnancy rates. j, k Representative image (j) and summary plot (k) of litter size (number of pups per litter, n = 5 female rats per group). Data are presented as mean ± S.E.M. Two-tailed unpaired t test for (b), (d)–(h), and (k). The exact p values are presented in the figures as indicated. Source data are provided in the Source Data file.
Discussion
Male infertility is an emerging global health concern, with stress-associated oligoasthenozoospermia (OAZS) frequently comorbid with anxiety and depression in industrialized populations48. Although this psychosomatic link is clinically recognized, the mechanisms connecting chronic stress to spermatogenic failure remains unclear. Current treatments often target hormone imbalances symptomatically, yielding limited efficacy due to their failure to address underlying neuroendocrine and molecular disruptions49,50,51,52. Here, we identify a gut-testis axis in which chronic stress disrupts bile acid and vitamin homeostasis via corticosterone-induced intestinal barrier dysfunction and Lactobacillus depletion. This disruption compromises spermatogenesis through Sting1 activation and reduces sperm motility via Slc9c2 suppression (Supplementary Fig. S22).
Chronic restraint stress (CRS) can lead to male reproductive deficits by activating the HPA axis and causing oxidative stress23. Chronic stress also disrupts intestinal homeostasis, as demonstrated by a reduction of intestinal tight junction proteins in CFSS rats, which are consistent with mechanistic evidence indicating that corticotropin-releasing factor (CRF)-mediated activation of the glucocorticoid receptor (GR) directly compromises intestinal barrier integrity53,54. This pathway appears to play critical roles in chronic stress-induced epithelial barrier dysfunction. Supportingly, RU-486 (an inhibitor of GR signaling) ameliorated intestinal barrier defects, underscoring the role of corticosterone (CORT)-GR axis in the pathogenesis of gut hyperpermeability. This axis is also associated with increased bacterial translocation and subsequent systemic inflammation in irritable bowel syndrome (IBS) model mice55, suggesting that dysregulated neuroendocrine signaling contributes to the disruption of mucosal barrier function. CORT concentration is a well-established biomarker of stress in capercaillies (Tetrao urogallus), a physiological parameter known to influence male reproductive function in this species56. Moreover, chronic stress-induced elevation of CORT secretion and GR signaling activation may impair spermatogenesis and reduce testosterone biosynthesis, through the induction of apoptosis and cell cycle arrest in germ cells and Leydig cells in murine models24,57. Administration of CORT in neonate mice induces p27 (a cyclin-dependent kinase inhibitor) expression in Sertoli cells and terminate Sertoli cell proliferation, leading to decreased Sertoli cell number and spermatozoa count in the testes of prepubescent mice58. Collectively, these findings indicate that chronic stress may adversely affect spermatogenesis in male rodents via the CORT/GR signaling pathway-mediated intestinal barrier integrity disruption and gut microbiota dysbiosis.
Gut microbiota is a key regulator of male reproductive health, since its dysbiosis may result in chronic inflammation, oxidative stress, and spermatozoa structural impairment, thus impairing spermatogenesis and sperm quality59,60,61,62. Key mechanisms linking microbial dysbiosis to infertility include inflammation, oxidative stress, and sperm structural deterioration. Emerging strategies such as targeted antimicrobial therapies, probiotics, prebiotics, and fecal microbiota transplantation (FMT) have shown promise in restoring microbial homeostasis and improving male fertility63. In the present study, the FMT experiments have established a causal link between CFSS-induced gut dysbiosis and oligoasthenozoospermia. Recipient rats colonized with CFSS microbiota recapitulated the spermatogenic defects of donors. Moreover, Lactobacillus abundance was highly correlated with semen quality parameters, suggesting a potential direct interaction between specific gut microbial taxa and male reproductive function. Indeed, alterations of the relative abundance were observed in multiple Lactobacillus species, including Lactobacillus reuteri, Lactobacillus mucosae, Lactobacillus salivarius, Lactobacillus intestinalis, Lactobacillus murinus, Lactobacillus coleohominis, Lactobacillus ruminis and Lactobacilus iohnsoni, of which only Lactobacillus reuteri showed significantly different. These findings suggest a potential direct interaction between specific gut microbial taxa and male reproductive function. For instance, a reduction in Lactobacilli spp. particularly Lactobacillus crispatus, clearly serves as a marker of low semen quality, whereas an increase in Lactobacillus iners is considered a risk factor for a reduced sperm concentration64. In mice model of CRS-induced male reproductive deficits, oral administration of Lactobacillus rhamnosus NCDC-610 or Lactobacillus fermentum NCDC-400, both of prebiotics can prevent CRS-induced activation of the HPA axis, protect against CRS-induced oxidative stress and sperm deficits, including sperm count, motility, morphology, and histopathology of testes, and improve intestinal health23. Similar protective effects of probiotics were also observed on chronic unpredictable stress (CUS)-induced male reproductive deficits65. A systematic review of randomized clinical trials on the administration of Lactobacilli in males with idiopathic infertility, reveals that probiotics supplementation reduce levels of reactive oxygen species (ROS) in semen and significantly improve sperm motility, and is indicated as a therapy for these patients66. Altogether, these findings underscore the critical role of microbial balance within the gut-genital axis in maintaining optimal male reproductive health.
The gut microbiota can ameliorate spermatogenic impairments by modulating host metabolism67,68. For instance, A10-fecal microbiota transplantation (FMT) improves the gut microbiota disrupted by type 1 diabetes (T1D), especially the increases in small intestinal Lactobacillus, and blood and testicular metabolome to produce n-3 polyunsaturated fatty acid (PUFA) docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA), which contribute to the improvement of spermatogenesis and semen quality37. KEGG analysis in our current study reveals aberrant bile acid metabolism and vitamin absorption pathways in CFSS rats, and these findings are supported by diabetic infertility models where microbiota-derived metabolites rescue steroidogenesis37,69. Specifically, CFSS-induced depletion of Lactobacillus disrupted the enterohepatic circulation of bile acids, impairing fat-soluble vitamin absorption70. This leads to gut luminal accumulation of vitamin A alongside systemic deficiency, indicating defective microbiota-dependent transport. Notably, Lactobacillus supplementation restored plasma and testicular levels of vitamins A and E, establishing a direct link between gut microbes and gonadal vitamin bioavailability. This functional axis was further validated by co-administration experiments: dietary vitamins A and E supplementation rescued meiotic progression and spermiogenesis, phenocopying the effects of Lactobacillus treatment. Vitamin A is essential for initiating meiosis in germ cells, a process of spermatogenesis regulated by gut microbiota, with specific microbial species capable of promoting the synthesis of retinoic acid, an active metabolite of vitamin A20,37. Retinoic acid regulates multiple stages of spermatogenesis, including spermatogonial differentiation, meiotic initiation, and sperm maturation71. The production of gut microbes-induced retinoic acid/vitamin A synthesis results in the upregulation of spermatogonial cell marker genes, specifically promyelocytic leukemia zinc-finger (PLZF) and deleted in azoospermia-like (DAZL), which signifies the commencement of spermatogenesis37,71. Also, Vitamin E has been demonstrated to improve spermatogenesis by regulating key proteins such as filamin A (FLNA), signal peptide peptidase like 3 (SPCS3), Y-box binding protein 3 (YBX3), and arginyl-tRNA synthetase (RARS), which are associated with the plasma membranes and protamine synthesis process in spermatozoa25. Together, these findings highlight the critical role of the Lactobacillus–vitamin A/E axis in male germ cell development.
Moreover, our current study by transcriptomic profiling of CFSS rats reveal systemic dysregulation of testicular spermatogenic programs, with marked enrichment in inflammatory pathways. Gut microbiota-derived lipopolysaccharide (LPS) impairs testicular function by activating inflammatory pathways, suppressing hypothalamic-pituitary activity, and reducing testosterone production72. Systemic endotoxemia and elevated LPS levels also compromise the blood-testis barrier, disrupting cellular junctions and inciting inflammatory damage72. In the epididymis, endotoxin upregulates inflammatory and monocyte chemotactic factors, which suppress meiosis-related gene expression in the testes and reduce sperm count73. Psychological and physiological stress can provoke endotoxemia and low-grade inflammation by enhancing barrier permeability74. Microbial-associated molecular patterns (MAMPs) such as LPS, bacterial lipoproteins, and peptidoglycans, infiltrate the testis via the vas deferens and testicular artery, inflicting potential harm, where MAMPs are recognized by pattern recognition receptors (PRRs) on resident immune cells, triggering innate immune responses and local inflammation74. Mechanically, LPS activates the NF-κB-MyD88/TRAM pathways, subsequently promotes the expression of inflammatory cytokines including interleukins (ILs) and tumor necrosis factors (TNFs)74,75, probably a crucial role in gut microbiota-associated local inflammation and functional impairment of the testis. These findings offer alternative mechanisms for chronic stress-induced male infertility and highlight the contribution of gut microbiota-mediated immuno-inflammatory responses.
STING is a widely identified intracellular sensor of foreign and endogenous DNA29. Activation of the STING1-dependent signaling network is implicated in the process of cell death, such as apoptosis, pyroptosis, necroptosis, and autophagy27,76. Acting as a key cytosolic DNA sensor, the cyclic GMP-AMP synthase (cGAS)/STING pathway orchestrates inflammatory responses triggered by microbial infections or endogenous cellular stressors such as autophagy and apoptosis77. The cGAS-STING1 signaling pathway is involved in regulating various pathological conditions such as cancer, autoimmune and inflammatory diseases, microbial and parasitic infections, as well as in regulating various cellular physiological processes, including cytokine production, autophagy, protein synthesis, metabolism, aging, and different types of cell death78,79. Emerging evidence underscores the dynamic interplay between cGAS-STING signaling and cell death, highlighting their relevance in disease pathogenesis and progression76,80, and the cGAS-STING1-NF-κB signaling pathway is shown to form an important pathological chain of “DNA stress-inflammation-metabolic abnormality” in metabolic-associated fatty liver disease81. In the present study, transcriptomic profiling analysis reveals widespread disruption of spermatogenic programs in CFSS rats, with testicular tissues showing enrichment in apoptosis, aberrant differentiation, and inflammatory pathways. Additionally, among differentially expressed genes (DEGs), Sting1 stood out due to its marked upregulation in Hierarchical clustering heatmap and volcano plot analyses, consistent with environmental toxin models where mitochondrial DNA leakage activates cGAS-STING1 signaling to drive germ cell apoptosis82. We therefore speculate that Sting1 activation likely mediates the CFSS-induced spermatogenesis impairment and oligozoospermia. Indeed, functional validation confirmed its causal role, e.g., lentiviral overexpression of Sting1 in the testicular tissues of naïve rats suppressed meiotic markers and induced severe oligospermia, whereas Sting1 knockdown in the testicular tissues of CFSS rats partially restored spermatid counts. Importantly, STING1-mediated apoptosis exhibited stage specificity, and Sting1 knockdown rescues spermatogonial differentiation but not sperm motility, suggesting distinct regulatory mechanisms for germ cell survival and function. These findings align with previous reports in bone/gastric cancer models, where STING1 activation promotes apoptosis via NF-κB/JAK-STAT crosstalk28,83, indicating conserved signaling logic across tissues. Moreover, in both in vivo (rat) and in vitro (GC-2 cells) models of testicular injury, increased inflammation levels are linked to elevated expression of STING gene in the testis, and STING-mediated AMPK/mTOR/P70S6K autophagy pathway participates in cyfluthrin-induced testicular injury84. Together, these findings suggest that chronic psychological stress impairs male fertility by disrupting testicular gene networks, in particular those governing immune and inflammatory responses, of which, the STING1-mediated apoptosis may be an important contributor of chronic stress-induced spermatogenesis depletion and oligozoospermia.
Notably, the failure of Sting1 knockdown to improve asthenospermia highlights pathway selectivity, inferring epididymal ion homeostasis as a key regulator of sperm motility. While Sting1 knockdown rescued oligospermia by reducing germ cell apoptosis, it failed to ameliorate sperm motility deficits in CFSS rats, suggesting distinct mechanisms governing sperm quantity versus quality. We thus performed RNA-seq on epididymal sperm to avoid confounding signals from heterogeneous testicular cells85,86, and identify a downregulation of Slc9c2 gene in epididymal of CFSS rats. Slc9c2, a gene encoding the sperm-specific sodium/hydrogen exchanger NHE11, is essential for sperm pH and calcium homeostasis36. Sodium/hydrogen exchangers (NHEs) are a family of ion transporters that regulate the pH of various cell compartments across an array of cell types34. We propose that Slc9c2 deficiency may disrupt intracellular pH homeostasis, leading to the dysfunction of CatSper (cation channel of sperm) channels, a family of sperm-specific, pH-sensitive calcium channels that serves as a key contributor of sperm mobility, hyperactivation and acrosome reaction87,88,89. Therefore, the Slc9c2 deficiency-mediated CatSper channels dysfunction ultimately results in sperm mobility impairment and asthenospermia phenotype. Of course, the direct evidence need be further studied.
The Slc9c2 gene product (NHE11) is localized to the head of mature mammalian sperm and is believed to play a crucial role in male fertility34. Among eight NHE isoforms (NHE1, NHE3, NHE5, NHE8, NHA1, NHA2, NHE10, and NHE11) expressed in testis and sperm, each one plays stage- and site-specific roles in sperm development, motility, capacitation, and the acrosome reaction32. Consistent with its functional importance, Slc9c2 knockdown in naïve rats recapitulated sperm motility defects seen in CFSS rats, while Slc9c2 overexpression restored progressive motility and fertility. Mechanistically, NHE11 likely extrudes protons to alkalinize sperm cytoplasm, a prerequisite for CatSper channels activation. This mechanism is evolutionarily conserved, mutations of Slc9c1 cause human asthenospermia90. Intriguingly, species-specific NHE11 localization (human: head; rat: head/flagella) suggests divergent pH buffering strategies. The dual localization in rat sperm may enable broader spatial pH control, potentially compensating for chronic stress-induced epididymal dysfunction. These findings position Slc9c2 as a stress-responsive regulator of sperm motility, operating independently of testicular gene networks. Our study expands the therapeutic options for idiopathic asthenospermia and identifies epididymis as a node in the gut-testis axis.
In conclusion, this study demonstrates the involvement of gut microbiota–vitamin A/E metabolism–Sting1/Slc9c2 (NHE11) genes axis in mediating chronic stress-induced oligoasthenozoospermia (OAZS). Our findings reframe chronic stress-induced infertility as a systemic, multi-organ disorder and support microbiome-targeted interventions as a viable therapeutic strategy. Combined supplementation with Lactobacillus and vitamins A/E may offer a promising complementary approach for managing chronic stress-related oligoasthenozoospermia, in particular for those patient populations of chronic psychological stress-induced male subfertility.
Several limitations merit consideration. First, species differences in stress responses, gut microbiota composition, and reproductive physiology may limit the CFSS rat model for fully replicating human chronic psychological stress and OAZS, thereby limiting the direct translation from the CFSS rat model to humans with psychological stress-induced OAZS. Second, key clinical parameters, such as optimal probiotic strains, vitamin dosages, and treatment duration, remain undefined. Thus, well-designed clinical trials, in particular human validation studies for confirming the translational relevance of our identified gut microbial-metabolic-testis/epididymis axis, are needed to validate the translational potential of the Lactobacillus–bile acid–vitamin A/E–Sting1/Slc9c2 (NHE11) pathways in the therapy of male subfertility.
Methods
Ethics
All experimental procedures involving animals were performed in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and approved by the Animal Care and Use Committee of Peking University Health Science Center (approval no. J2024179).
Animals
Sexually mature male Wistar rats (Crl:Wl;Outbred; 200–220 g at the beginning of the experiment) and 6-week-old female Wistar rats (Crl:Wl;Outbred) were provided by the Department of Experimental Animal Sciences, Peking University Health Science Center. For the study of male fertility, we focused solely on male rats; while female rats were used only for the in vivo fertility assay. The rats were housed in separated cages with free access to standard rodent chow (SPF-F01-001, SPF Biotech, Beijing, China) and water. The room temperature was kept at 24 ± 1 °C and 40–60% humidity under a 12-h light/12-h dark cycle91. Animals are randomly allocated into treatment groups. At the end of the experiments, animals were euthanized via gradual displacement of cage air with CO2, in accordance with institutional animal welfare guidelines.
Rat model of chronic stress
Chronic forced swim (FS) is a well-established stress model that is widely employed to assess the therapeutic efficacy of antidepressant agents92. This model has been extensively utilized in neuroscience research to investigate the neurobiological mechanisms underlying depression and related disorders93,94. Prolonged exposure to the FS protocol has been demonstrated to induce cognitive impairments that closely mirror those observed in major depressive disorder (MDD)95,96. The behavioral phenotypes associated with this model, such as behavioral despair, anhedonia, and reduced motivational drive, alongside the concomitant neurochemical and endocrine alterations, are consistent with core features of depressive pathology. These changes emerge during the development of learned helplessness following repeated, inescapable stress exposure, supporting the validity of the chronic FS-stressed rat as a preclinical model of depression92,97. Moreover, chronic FS stress elicits robust neurochemical (e.g., alterations in monoaminergic, glutamatergic, and neurotrophic systems) and endocrine responses (notably activation of the hypothalamic-pituitary-adrenal axis), thereby serving as a potent psychological and physiological stressor in experimental studies98,99. In this study, chronic forced swim (FS) was used as a stressor100. Briefly, rats were placed in a glass cylinder (45 cm in height, 20 cm in diameter) filled with 30 cm depth of ice water (0–2 °C). The FS procedure was conducted to chronic FS (CFSS) group rats individually twice per day in 15-min sessions, continued for 21 consecutive days. According to the methods described elsewhere101, control rats were subjected to a sham swimming (sham FS) session by allowing them to wade in the cylinder that contained 2–4 cm of warm water (24–26 °C). Here, we used sham FS animals rather than naïve animals as control for the reason to exclude the factor of habituation for rats to the water. Rats were allowed to dry in a warm environment (30–33 °C) after swimming. The water was changed and the container was thoroughly cleaned for each rat.
Enzyme-linked immunosorbent assay (ELISA)
Blood samples were collected from the tail vein for plasma analysis of adrenocorticotropic hormone (ACTH), corticosterone (CORT), and vitamins A/E. All sampling was completed within a 2-h window (10:00 am-12:00 pm) at 24 h post-CFSS to minimize circadian interference. Plasma was separated by centrifugation (2000 × g, 20 min) after 30 min clotting at room temperature, aliquoted, and stored at −20 °C. Plasma vitamin A and E levels were detected by a rat vitamin A ELISA Kit (MERDA, Cat#M053968-48T) and rat vitamin E ELISA Kit (MERDA, Cat#M053967-48T), respectively. Rat CORT ELISA Kit (Elabscience, Cat#E-OSEL-R0002) and rat ACTH ELISA Kit (Elabscience, Cat#E-EL-R0048c) were used for plasma CORT and ACTH detection, respectively102. Hypothalamic paraventricular nucleus (PVN) tissues were homogenized in RIPA buffer with protease inhibitors. Testes were homogenized in PBS containing 0.1% butylated hydroxytoluene. After centrifugation (12,000 × g, 10 min), supernatants were analyzed using a rat CRH ELISA Kit (Elabscience, Cat#E-EL-R0270c), a rat vitamin A ELISA Kit (MERDA, Cat#M053968-48T), or a rat vitamin E ELISA Kit (MERDA, Cat#M053967-48T), with CRH and vitamin levels normalized to total protein (BCA assay, Thermo Fisher).
Open field test (OFT)
The open field test was conducted 24 h after the final chronic stress exposure. A square arena (100 × 100 × 50 cm) was placed in a dark, quiet room. Each rat was positioned at the center and allowed to freely explore for 5 min under video recording. The arena was thoroughly cleaned with 75% ethanol and dried between trials to eliminate odor cues. Behavioral parameters including total travel distance, entries into the central zone (50 × 50 cm), and time spent in the central zone were analyzed using SMART software (Panlab)103,104.
Elevated plus maze (EPM)
The elevated plus maze consisted of two open arms (50 × 10 cm), two enclosed arms (50 × 10 × 35 cm), and a central platform (10 × 10 cm) elevated 50 cm above the floor. Testing was performed 48 h post-stress. Rats were placed on the central platform facing an open arm and allowed to explore freely for 5 min. The maze was cleaned with 75% ethanol after each session. SMART software quantified total distance moved, open arm entries, and time spent in open arms103.
Sucrose preference test (SPT)
Rats were first acclimated with a 1% sucrose solution for 48 h, then subjected to a 12-hour water deprivation period before undergoing the SPT. During the test, the rats were given unrestricted access to two bottles, one containing 1% sucrose solution and the other containing tap water, for a duration of one hour. The position of the two bottles was switched midway through the test. Thereafter, consumption was measured and sucrose preference (%) was calculated. The SPT was performed both on the day prior to the CFSS treatment and 24 h after the final exposure of rats to the CFSS104.
Forced swim test (FST)
24 h after the final exposure to CFSS, rats were subjected to a 6-minute forced swimming session, during which their behavior was captured on video for subsequent analyses. The rats were considered immobile when they ceased to struggle and remained suspended in the water, making only necessary movements to keep their heads above water. The duration of immobility was recorded by an experimenter who was blinded to the experimental conditions during the last 4 minutes of the 6-minute testing session. This was done to eliminate any potential ‘first-two-minute bias’, as most animals tend to be highly active at the beginning of the FST104.
Tail suspension test (TST)
The TST was performed according to previously published methods104. In brief, a wooden box painted gray, with dimensions of 54 × 30 × 52 cm, was used as the apparatus. The rats were suspended 50 cm above the floor using adhesive tape placed ~1 cm from the tip of their tails, and the session was videotaped. Immobility was defined as complete absence of limb or body movement, except for those required for respiration, while the rat remained passively and completely still. The duration of immobility during a 6-minute test period was recorded while the rats were isolated from each other to eliminate any potential visual or acoustic associations. Observers scoring the test were blinded to the experimental conditions under which the rats were tested104.
Sperm concentration, viability and motility
Cauda epididymal spermatozoa of rats were collected and prepared as previously described105. In brief, caudal epididymis were placed in high-saline (HS) solution containing 135 mM NaCl, 5 mM KCl, 2 mM CaCl2·2H2O, 1 mM MgSO4·7H2O, 20 mM HEPES, 5 mM glucose, 10 mM sodium lactate, 1 mM sodium pyruvate, adjusted to pH 7.4 with NaOH. Then, the cauda epididymis was slightly cut into three pieces, incubated at 37 °C for 10 min in a 5% CO2 incubator and the sperm was gently filtered through nylon gauze, centrifuged, and resuspended in 1 ml HS solution. A drop of the sperm suspension was used for assessment of sperm concentration and motility by CASA.
In vivo fertility assay
In vivo fertility assay was conducted following previously described methods105. Briefly, male rats were mated with 6-week-old female rats in a 1:2 ratio, where one male rat was housed overnight with two female rats. The following morning, vaginal swabs were taken from the female rats and examined under a microscope. Female rats with swabs positive for sperm were considered successfully mated and were then housed individually. After birth, the number of offspring and the pregnancy rate of these female rats were calculated and recorded.
RNA extraction and RT-qPCR
Total RNA was extracted from rat testis tissues using Trizol reagent (Life Technologies). Reverse transcription coupled with polymerase chain reaction (PCR) was executed using oligo deoxythymidine (oligo-dT) primers, in conjunction with moloney murine leukemia virus reverse transcriptase (Promega), following the manufacturer’s protocol. NanoDrop spectrophotometer was used to measure samples, recording nucleic acid concentration and A260/A280 purity ratio for rapid quantification and purity assessment. PCR primer sequences are listed in Supplementary Tables S1 and S2. Real-time quantitative PCR (RT-qPCR) assay was conducted on an ABI 7500 Fast Real-Time PCR Detection System (Applied Biosystems) employing GoTaq qPCR Master Mix (Promega). In brief, a 20 μL PCR reaction was established in strict accordance with the manufacturer’s protocol, which contains 1 μL of complementary DNA, 0.2 μM of each primer, and 10 μL of GoTaq qPCR Master Mix, with double distilled H2O (ddH2O) to adjust the final volume. Gene expression was normalized to β-actin. PCR conditions was performed with an initial 3-min incubation at 95 °C, followed by 40 thermal cycles (95 °C for 10 s, 58 °C for 20 s, and 72 °C for 10 s). The relative expression ratio of mRNA was determined using the 2−ΔΔCt method105.
Immunofluorescence staining
For the immunofluorescence staining of epididymal sperm, 20 μL of the sperm suspension was combined with an equal volume of 4% paraformaldehyde (0.1 M PB, pH 7.4). One drop of the mixture was applied to clean glass slides to create smears and then allowed to air-dry. For the immunofluorescence staining of small intestine tissues or testis tissues, rats were deeply anesthetized and underwent transcardial perfusion with 300 mL of phosphate buffer (PB, 0.1 M) followed by an equivalent volume of 4% paraformaldehyde for fixation. The removed specimens underwent post-fixation in 4% paraformaldehyde (0.1 M PB, pH 7.4) for 6 h at 4 °C, followed by cryoprotection using a 30% sucrose solution (in 0.1 M PB) at 4 °C. Several days later, 20-μm-thick sections were prepared from the tissues using a cryostat and subsequently collected onto gelatin-coated slides for immunostaining105. To perform immunostaining, small intestine, testis tissues or sperm were washed in PB (3 times × 5 min), followed by a 1-h blocking step in 10% goat or rabbit serum (0.1 M PBST with 0.3% Triton X-100) at room temperature. Intestinal tissues were incubated with rabbit anti-ZO-1 antibody (1:200, Invitrogen, Cat#40-2200, RRID: AB_2533456), mouse anti-Claudin-1 antibody (1:200, Invitrogen, Cat#37-4900, RRID: AB_2533323) or rabbit anti-Occludin antibody (1:200, Abcam, Cat#33-1500, RRID: AB_2533101) in PBST at 4 °C overnight. Testicular tissues were incubated with the primary antibody in PBST at 4 °C overnight, including rabbit anti-DAZL antibody (1:200, Abcam, Cat#ab34139, RRID: AB_731849), rabbit anti-SYCP3 antibody (1:100, Abcam, Cat#ab15093, RRID: AB_301639), rabbit anti-TNP1 antibody (1:100, Abcam, Cat#ab73135, RRID: AB_10714560), rabbit anti-PGK2 antibody (1:200, Sangon biotech, Cat#D121803, RRID: AB_3717431) or rabbit anti-STING1 antibody (1:200, Abcam, Cat#ab288157, RRID: AB_3086730). Rat spermatozoa were incubated with rabbit anti-Slc9c2 antibody (1:100, Proteintech, Cat#17398-1-AP, RRID: AB_2191353). Then, after three washes in PBS, tissues or sperm were incubated with the following appropriate secondary antibodies at room temperature for 1 h, including Alexa Fluor®488-Conjugated Donkey anti-Rabbit IgG (1:1000, Jackson Immuno Research, Cat#711-547-003, RRID: AB_2340620), Alexa Fluor®488-Conjugated Donkey anti-mouse IgG (1:1000, Jackson Immuno Research, Cat#115-545-003, RRID: AB_2338840) or Alexa Fluor®647-Conjugated Donkey anti-rabbit IgG (1:1000, Jackson Immuno Research, Cat#711-605-152, RRID: AB_2492288). The tissues or sperm were counterstained with the nuclear marker DAPI (100 ng/mL) carrying blue fluorescence for 10 min at room temperature. The slides were mounted in Gel-Mount medium. Visualization of fluorescence signal was performed by Leica Stellaris 5 microscopy (Leica Microsystems, Wetzlar, Germany) at excitation wavelengths of 488 nm (green), 647 nm (red) or 405 nm (blue), respectively. At least three fields per slide were analyzed to establish reproducibility105.
Western blot
Rat testicular tissues, small intestine tissues or sperm cells were homogenized in ice-cold lysis buffer (50 mM Tris pH 8.0, 150 mM NaCl, 1% NP40, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM PMSF). Homogenates were centrifuged (12,000 × g, 10 min, 4 °C), and supernatants were analyzed with BCA assay (Pierce). Equal protein amounts (40 μg) were separated on 10% SDS-PAGE gels, transferred to PVDF membranes (Bio-Rad), and blocked with 5% nonfat milk/BSA in TBST. Membranes were incubated overnight at 4 °C with primary antibodies, including rabbit anti-DAZL antibody (1:1000, Abcam, Cat#ab34139, RRID: AB_731849), rabbit anti-SYCP3 antibody (1:1000, Abcam, Cat#ab15093, RRID: AB_301639), rabbit anti-TNP1 antibody (1:500, Abcam, Cat#ab73135, RRID: AB_10714560), rabbit anti-PGK2 antibody (1:2000, Sangon biotech, Cat#D121803, RRID: AB_3717431), rabbit anti-STING1 antibody (1:1000, Abcam, Cat#ab288157, RRID: AB_3086730), or mouse anti-β-actin antibody (1:2000, Immunoway, Cat#YM3028, RRID: AB_2629465) for rat testis tissues. Rabbit anti-ZO-1 antibody (1:1000, Invitrogen, Cat#40-2200, RRID: AB_2533456), mouse anti-Claudin-1 antibody (1:1000, Invitrogen, Cat#37-4900, RRID: AB_2533323), rabbit anti-Occludin antibody (1:1000, Abcam, Cat#33-1500, RRID: AB_2533101), or mouse anti-β-actin antibody (1:2000, Immunoway, Cat#YM3028, RRID: AB_2629465) for rat small intestine tissues. Rabbit anti-Slc9c2 antibody (1:500, Proteintech, Cat#17398-1-AP, RRID: AB_2191353) and mouse anti-β-actin antibody (1:2000, Immunoway, Cat#YM3028, RRID: AB_2629465) for rat spermatozoa. The blots were washed in TBST and then were incubated in horseradish peroxidase-conjugated secondary antibody, including goat anti-rabbit IgG-HRP (1:2000, Santa Cruz, Cat# sc-2004, RRID: AB_631746) or goat anti-mouse IgG-HRP (1:2000, Santa Cruz, Cat# sc-2005, RRID: AB_631736). Protein bands were visualized using an enhanced chemiluminescence detection kit (Pierce) followed by autoradiography using Hyperfilm MP (Santa Cruz). The bands, including all the target protein and the internal loading control (β-actin), were quantified with a computer-assisted imaging analysis system (Image J, NIH), and the normalized ratio of the target protein to β-actin band density was used to calculate the alteration of corresponding protein expression105.
Exogenous CORT and RU-486 administration
CORT (Twbio, Cat#DA88478) was dissolved in absolute ethanol and diluted with saline (1:19 ratio) to a final concentration of 2 mg/kg106. The vehicle control comprised 5% ethanol in saline. RU-486 (KIRGEN, Cat#IM320) was suspended in 5% DMSO, 5% Cremophor EL, and 90% ddH2O, with its vehicle containing the same solvent mixture38. For exogenous CORT administration in normal rats, adult male Wistar rats were randomly divided into four groups as follows: vehicle group received 1 mL of CORT vehicle (5% ethanol in saline) daily; CORT group was administered 2 mg/kg CORT solution daily; CORT + RU-486 group was co-treated with 2 mg/kg CORT and 3 mg/kg RU-486 suspension daily; CORT + RU-vehicle group received 2 mg/kg CORT plus RU-486 vehicle daily. For exogenous RU-486 administration to CFSS model rats, adult male Wistar rats undergoing CFSS were divided into three groups as follows: CFSS+vehicle received RU-486 vehicle (5% DMSO, 5% Cremophor EL, 90% ddH2O) daily; CFSS + RU-486 was administered 2 mg/kg RU-486 suspension via oral gavage daily; CFSS + RU-vehicle was treated with RU-486 vehicle daily. All treatments were delivered via oral gavage once daily for 21 consecutive days.
ABX cocktail treatment and FMT
Male Wistar rats (recipient rats) were given a sterile-water drink containing: 1 g/L ampicillin (Songon, Cat#A610028), 1 g/L neomycin (Songon, Cat#A610366), 1 g/L metronidazole (Songon, A600633), and vancomycin 0.5 g/L (Songon, Cat#A600983) for seven days starting from the initiation of the experiment. Fresh fecal pellets (200 mg/day) from sham FS and CFSS donor rats were homogenized in sterile saline (1:10 w/v), allowed to sediment for 15 min at room temperature, and filtered through sterile gauze. Sham-FMT and CFSS-FMT recipient rats received 1 mL of respective donor fecal suspensions via oral gavage once per day for 21 days91.
Lactobacillus supplementation
For Lactobacillus supplement to CFSS group rats, Lactobacillus (BNCC, Cat#223759), which composition is Lactobacillus sp. enrichment culture clone MP30 16S ribosomal RNA gene, were cultured in MRS medium supplemented with 5% FPS under anaerobic conditions at 37%. Perform serial dilutions of the bacterial culture, spread onto agar plates, incubate, then count colonies on plates containing 30-300 colonies, and calculate the number of viable bacteria per milliliter (CFU/mL) based on the dilution factor. CFSS group rats were colonized with 200 μL indicated bacteria consortia at a dose of 2 × 108 colony-forming units per 200 μL suspended in the MRS medium or the same volume of MRS medium by oral gavage once per day for 21 days107.
16S rDNA amplicon sequencing and bioinformatics analysis
Small intestine digesta samples were immediately snap-frozen and stored at −80 °C. Bacterial DNA was extracted using the MagPure Soil DNA LQ Kit (Magen, China), with concentration and integrity assessed by NanoDrop 2000 spectrophotometry and agarose gel electrophoresis. The V3-V4 hypervariable regions of the 16S rRNA gene were amplified via PCR (25 μL reaction volume) using universal primers 343 F (5’-TACGGRAGGCAGCAG-3’) and 798 R (5’-AGGGTATCTAATCCT-3’), incorporating Illumina adapters and sample-specific barcodes. Amplified fragments were purified with Agencourt AMPure XP beads (Beckman Coulter), quantified via Qubit dsDNA assay, and sequenced on an Illumina NovaSeq6000 platform (250 bp paired-end reads; OE Biotech Co., China). Raw reads were preprocessed with Trimmomatic to remove low-quality sequences (Q20 threshold) and assembled using FLASH (10–200 bp overlap; ≤20% mismatch). Chimeric sequences were filtered by VSEARCH, followed by clustering into operational taxonomic units (OTUs) at 97% similarity via QIIME (v1.8.0). Representative OTU sequences were taxonomically annotated against the SILVA database (v132) using the RDP classifier (70% confidence). Alpha diversity (Chao1 and Shannon indices) and beta diversity (unweighted UniFrac PCoA) analyses were performed in QIIME, with phylogenetic trees generated from UniFrac distance matrices91. Sequencing and bioinformatics were conducted by OE Biotech Co., Ltd. (Shanghai, China).
Metabolomics profiling and bioinformatics analysis
Small intestine digesta (n = 10 biological replicates per group), plasma (n = 10 biological replicates per group), and testis (n = 4 biological replicates per group) samples stored at −80 °C were thawed on ice. Plasma was separated by centrifugation (2000 × g, 20 min) after 30 min clotting at room temperature. For solid samples, 20 mg of material was homogenized in 400 μL methanol:water (7:3, v/v) containing internal standards, vortexed, sonicated (ice bath, 10 min), and centrifuged (12000 × g, 4 °C). For liquid samples, 50 μL was mixed with 300 μL ACN:methanol (1:4, v/v) with internal standards, followed by vortexing and centrifugation. Supernatants from both sample types were filtered (0.22 μm) and analyzed via LC-MS using a Shimadzu LC20 UPLC system coupled with an AB Sciex Triple TOF-6600 mass spectrometer (Waters ACQUITY UPLC HSS T3 C18 column, 1.8 μm, 2.1 × 100 mm; 40 °C; 0.4 ml/min flow rate). The mobile phase consisted of 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B), with a gradient from 5% to 90% B over 11 min, held for 1 minute, and then come back to 5% mobile phase B within 0.1 minute, held for 1.9 minutes. Key settings included: IonSpray Voltage, 5500 V (ESI + )/ -4500 V (ESI-); Ion Source Gas 1 and 2, 50/60 psi; Curtain Gas, 35 psi; Source Temperature, 550 °C; and Collision Energy, 30 eV/ -30 eV. Raw data (mzML format, converted by ProteoWizard v3.0) underwent peak extraction and alignment using XCMS, with SVR-based area correction and removal of low-frequency peaks ( < 50% detection rate). Metabolites were identified against in-house and public databases (SILVA, AI, MetDNA). For bioinformatics, PCA and hierarchical clustering (HCA, heatmaps with dendrograms) were performed in R v3.5.1 (unit variance scaling). Differential metabolites were defined by VIP > 1 (OPLS-DA, MetaboAnalystR v1.0.1) and P < 0.05 (Student’s t-test), validated via permutation testing (200 cycles). KEGG pathway enrichment was assessed using a hypergeometric test on annotated metabolites91. Representative total ion chromatograms (TICs) or Base Peak Chromatogram (BPC) for the methods are provided in Supplementary Fig. S23.
Vitamin A and Vitamin E supplementation
The procedure of vitamin A and vitamin E supplementation was performed as described in previous studies108,109. In brief, vitamin A palmitate and vitamin E acetate were dissolved in corn oil, supplemented with 0.3% Tween 80 (v/v) and 0.5% sodium carboxymethyl cellulose (CMC-Na, w/v). For vitamin A/E supplement, CFSS model rats received daily oral gavage of 1 mL emulsion containing corn oil, ddH2O, 0.3% Tween 80, and 0.5% CMC-Na (without vitamins) for 21 consecutive days, while the vitamin A/E supplementation group was administered 1 mL emulsion containing the same solvent components plus 2000 IU/kg vitamin A palmitate108 and 50 mg/kg vitamin E acetate109 for the same duration. Here, the duration of 21-days treatment was used for the reason that the total duration of the spermatogenesis is 50 days for rats, and 3 weeks treatment was almost covered half of one spermatogenesis duration110.
Glycoursodeoxycholic acid (GUDCA) supplementation
For GUDCA intervention, CFSS rats in the treatment group received daily oral gavage of 1 mL phosphate-buffered saline (PBS) containing 60 mg/kg glycoursodeoxycholic acid (GUDCA, Acmec, T91840)107 for 21 consecutive days. In parallel, CFSS rats in the control group received daily oral gavage of 1 mL PBS alone over the same period.
Comprehensive supplementation of Lactobacillus and vitamin A/E
CFSS rats in the comprehensive therapy group received daily oral gavage of 200 μL containing a defined bacterial consortium (Lactobacillus), along with 1 mL of vitamin A palmitate (2000 IU/kg) and vitamin E acetate (50 mg/kg) for 21 consecutive days. In contrast, CFSS rats in the control group received daily gavage of 200 μL MRS medium and 1 mL vitamin-free emulsion over the same period.
RNA sequencing and data analysis
Total RNA was extracted from testis or epididymis tissues of male rats after CFSS treatment. The Beijing Genomics Institute (BGI) conducted the RNA sequencing (RNA-seq) for this study. In briefly, total RNA was extracted using TRIzol reagent as per the manufacturer’s instructions following the dissociation of testis or epididymis tissues. Isolation of total RNA was achieved using the RNeasy mini kit (Qiagen, Germany). Subsequently, TruSeq RNA Sample Preparation Kit (Illumina, USA) was employed to prepare the RNA-seq library from 1 µg of total RNA. Briefly, following the purification of poly A-containing mRNA, the molecules were cleaved into small fragments via divalent cations at 94 °C for 8 min. Subsequently, library concentration was determined with a Qubit 2.0 Fluorometer (Invitrogen, USA) and verified via an Agilent 2100 bioanalyzer (Agilent Technologies, USA) to assess the insert size and to establish the mole concentration. To identify differentially expressed genes (DEGs) across groups, gene expression levels were normalized to TPM (Transcript per kilobase per million mapped reads). For inter-group comparisons, the binary logarithm of the fold change (log2(FC)) was calculated based on these TPM values. Significant differences in gene expression were defined using a log2(FC) cutoff of ≥ 1.0, which corresponds to a minimum of two-fold change. Following transcriptome assembly, we determined the expression levels for all identified genes and transcripts. Functional enrichment was ultimately conducted using Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) databases to elucidate the biological roles of the DEGs. GO analysis categorized these genes into biological processes, molecular functions, and cellular components. Simultaneously, KEGG pathway analysis identified critical signal transduction and disease-related mechanisms, providing a comprehensive functional framework for the identified genes and their associated pathways91.
Plasmid construction and lentivirus infection
Lentivirus-expressing Sting1 (LV-Sting1) and lentivirus-expressing shRNA targeting rat Sting1 (LV-shSting1), both linked with an EGFP tag, using either the pcSLenti-EF1-EGFP-CMV-MCS-WPRE or pCLenti-U6-shRNA-CMV-EGFP-WPRE vector were produced by OBiO technology (Shanghai, China). For Sting1 silencing, three shRNA sequences were designed (Supplementary Data 1), with preliminary experiments demonstrating that shRNA (Sting1)-1 effectively suppressed Sting1 expression in testis tissues, leading to its selection for further study. Lentivirus-expressing Slc9c2 (LV-Slc9c2) and lentivirus-expressing shRNA targeting rat Slc9c2 (LV-shSlc9c2) fused to an EGFP tag, using pcSLenti-EF1-EGFP-CMV-MCS-WPRE and pCLenti-U6-shRNA-CMV-EGFP-WPRE vector, were provided by BrainVTA technology (Wuhan, China). Lentiviral titer was determined by qPCR quantification of viral DNA copies integrated into the host genome. Genomic DNA was extracted from 293 T cells (ATCC, Cat# CRL-3216) 72 h post-infection, and qPCR was performed targeting a viral-specific sequence (e.g., WPRE). The copy number was calculated using a standard curve and converted to transducing units per milliliter (TU/mL) based on the number of infected cells, volume of virus used, and gDNA parameters. Three shRNA sequences for Slc9c2 were evaluated (Supplementary Data 1), and shRNA (Slc9c2)-3 was selected based on its superior silencing efficacy in preliminary experiments. For testis Sting1 overexpression or spermatozoa Slc9c2 knockdown in naïve rats, LV-Sting1 lentivirus or LV-shSlc9c2 lentivirus were injected through the efferent ductule of testis at a final titer of 5 × 108 transducing units/ml. The procedure of intratesticular virus injections were performed as described elwhere with minor modification111,112,113. Briefly, rats were anesthetized via intraperitoneal injection of pentobarbital sodium (50 mg/kg). The surgical site was prepared aseptically using sequential disinfection with 70% ethanol and povidone-iodine solution. A midline ventral incision, ~1.5 cm anterior to the genital apparatus, was made through the skin and abdominal wall using sterile surgical scissors under aseptic conditions. The testes were exteriorized by gentle traction on the epididymal fat pad. Each testis was stabilized with fine forceps, and lentiviral particles were delivered directly into the seminiferous tubules via the efferent ducts using a 33-gauge microsyringe (Hamilton, Switzerland). A volume of 50 μL containing 0.04% Trypan blue, used as a tracer to visualize distribution, was injected into the interstitial compartment of both left and right testes, to ensure consistent delivery volume and location within the testis or efferent ductule. Following successful delivery, the abdominal wall and skin incisions were sutured in layers using sutures, respectively111,112. 21 days after viral infection, a transfection efficiency of lentivirus was routinely achieved as observed under fluorescence microscopy. For testis Sting1 knockdown or spermatozoa Slc9c2 overexpression of CFSS-treated rats, LV-shSting1 lentivirus or LV-Slc9c2 lentivirus was injected through the efferent ductule of testis at a final titer of 5 × 108 transducing units/ml (total volume of 50 μL in both left and right side of the testis tissues). The CFSS procedure was initiated 2–3 days after lentiviral injection to allow initial recovery from surgical trauma while avoiding interference with viral transduction.
Statistical analysis
The sample size was determined based on previously described guidelines114,115. In brief, the total number of animals was selected according to the number of experimental groups and the specific requirements of each assay. For instance, n = 6–8 rats per group were used for qPCR and RNA-seq analyses, whereas n = 3–6 rats per group were sufficient for Western blot experiments, in accordance with established standards for statistical power and experimental reproducibility. Statistical analyses were performed with GraphPad Prism 9.0 for Windows (GraphPad Software, La Jolla, CA). The number of independent biological replicates are provided in the figures and figure legends. All quantitative biochemical data and immunofluorescence staining are representative of at least three independent biological replicates. Shapiro–Wilk tests were used to assess normality in the distribution (Gaussian distribution, p > 0.05, Levene’s post hoc test p > 0.05) for each group, and only the data were normally distributed and variances were similar between groups to be compared were subjected to parametric statistical tests. Two-tailed unpaired Student’s t test was used for the comparison of the mean values between two groups. One-way analysis of variance (ANOVA) with Tukey’s post hoc test was used for multiple comparison. All data are reported as means ± SEM, and differences with p < 0.05 were considered statistically significant. The investigators were blinded to group allocation during data collection. All statistical data are presented in Supplementary Data 2.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All relevant data supporting the key findings of this study are provided in the Supplementary Information/Source Data files, or in public repositories. All the 16S rDNA sequencing data for intestinal contents and the RNA-seq data of testis and epididymis generated in this study have been deposited in the NCBI’s Sequence Read Archive (SRA) under accession code as follows: PRJNA1416019. PRJNA1416363. PRJNA1416387. The metabolomic data generated in this study have been deposited in the OMIX database under accession code OMIX015806. Source data are provided with this paper.
Code availability
No custom software code was used in this research.
References
Agarwal, A. et al. Male infertility. Lancet 397, 319–333 (2021).
Boitrelle, F. et al. The sixth edition of the WHO manual for human semen analysis: a critical review and SWOT analysis. Life (Basel) 11, 1368 (2021).
Wyrwoll, M. J. et al. Improved phenotypic classification of male infertility to promote discovery of genetic causes. Nat. Rev. Urol. 21, 91–101 (2024).
Selvaraju, V., Baskaran, S., Agarwal, A. & Henkel, R. Environmental contaminants and male infertility: Effects and mechanisms. Andrologia 53, e13646 (2021).
Methorst, C., Perrin, J., Faix, A. & Huyghe, E. Male infertility, environment and lifestyle]. Prog. Urol. 33, 613–623 (2023).
Bräuner, E. V. et al. Psychological stress, stressful life events, male factor infertility, and testicular function: a cross-sectional study. Fertil. Steril. 113, 865–875 (2020).
Kolbasi, B. et al. Chronic unpredictable stress disturbs the blood-testis barrier affecting sperm parameters in mice. Reprod. Biomed. Online 42, 983–995 (2021).
Mohamed, Z. I., Sivalingam, M., Radhakrishnan, A. K., Jaafar, F. & Zainal Abidin, S. A. Chronic unpredictable stress (CUS) reduced phoenixin expression, induced abnormal sperm and testis morphology in male rats. Neuropeptides 107, 102447 (2024).
Li, T. et al. Chronic stress impairs male spermatogenesis function and Nectin-3 protein expression in the testis. Physiol. Res 69, 297–306 (2020).
Li, L. et al. Hormone regulation in testicular development and function. Int J. Mol. Sci. 25, 5805 (2024).
Wang, X., Liu, Q., & Li, J. Dynamic transcriptional regulation of the hypothalamic-pituitary-testis axis in the seasonally breeding teleost Sebastes schlegelii. Int. J. Mol. Sci. 26, 2048 (2025).
Schneider, K. M. et al. The enteric nervous system relays psychological stress to intestinal inflammation. Cell 186, 2823–2838.e2820 (2023).
Sidik, S. Chronic stress can inflame the gut - now scientists know why. Nature 618, 221–222 (2023).
Zhou, Y. et al. Gut microbiota combined with metabolome dissects long-term nanoplastics exposure-induced disturbed spermatogenesis. Ecotoxicol. Environ. Saf. 267, 115626 (2023).
Chen, T. et al. Gut-derived exosomes mediate the microbiota dysbiosis-induced spermatogenesis impairment by targeting meioc in mice. Adv. Sci. (Weinh.) 11, e2310110 (2024).
Dodiya, H. B. et al. Chronic stress-induced gut dysfunction exacerbates Parkinson’s disease phenotype and pathology in a rotenone-induced mouse model of Parkinson’s disease. Neurobiol. Dis. 135, 104352 (2020).
Doney, E. et al. Chronic stress exposure alters the gut barrier: sex-specific effects on microbiota and jejunum tight junctions. Biol. Psychiatry Glob. Open Sci. 4, 213–228 (2024).
Hao, Y. et al. Gut microbiota-testis axis: FMT mitigates high-fat diet-diminished male fertility via improving systemic and testicular metabolome. Microbiol Spectr. 10, e0002822 (2022).
Zhang, T. et al. Disrupted spermatogenesis in a metabolic syndrome model: the role of vitamin A metabolism in the gut-testis axis. Gut 71, 78–87 (2022).
Li, H., Palczewski, K., Baehr, W. & Clagett-Dame, M. Vitamin A deficiency results in meiotic failure and accumulation of undifferentiated spermatogonia in prepubertal mouse testis. Biol. Reprod. 84, 336–341 (2011).
Zhao, Q. et al. Polyamine metabolism links gut microbiota and testicular dysfunction. Microbiome 9, 224 (2021).
Zhang, Z. et al. Differential gene expression and gut microbiota composition in low-altitude and high-altitude goats. Genomics 116, 110890 (2024).
Akram, M., Ali, S. A. & Kaul, G. Probiotic and prebiotic supplementation ameliorates chronic restraint stress-induced male reproductive dysfunction. Food Funct. 14, 8558–8574 (2023).
Zou, P. et al. Mechanisms of stress-induced spermatogenesis impairment in male rats following unpredictable chronic mild Stress (uCMS). Int. J. Mol. Sci. 20, 4470 (2019).
Gao, Y. et al. Vitamin E can promote spermatogenesis by regulating the expression of proteins associated with the plasma membranes and protamine biosynthesis. Gene 773, 145364 (2021).
Song, K., Heng, L. & Yan, N. STING: a multifaced player in cellular homeostasis. Hum. Mol. Genet 34, R68–r74 (2025).
Zhang, R., Kang, R. & Tang, D. The STING1 network regulates autophagy and cell death. Signal Transduct. Target Ther. 6, 208 (2021).
Guo, Q. et al. STING promotes senescence, apoptosis, and extracellular matrix degradation in osteoarthritis via the NF-κB signaling pathway. Cell Death Dis. 12, 13 (2021).
Liu, S. & Guan, W. STING Signaling Promotes Apoptosis, Necrosis, and Cell Death: An Overview and Update. Mediators Inflamm. 2018, 1202797 (2018).
Sun, Y. et al. STING directly interacts with PAR to promote apoptosis upon acute ionizing radiation-mediated DNA damage. Cell Death Differ. 32, 1167–1179 (2025).
Fuster, D. G. & Alexander, R. T. Traditional and emerging roles for the SLC9 Na+/H+ exchangers. Pflug. Arch. 466, 61–76 (2014).
Gardner, C. C., & James, P. F. Na(+)/H(+) Exchangers (NHEs) in mammalian sperm: essential contributors to male fertility. Int. J. Mol. Sci. 24, 14981 (2023).
Martins, A. D. et al. Physiology of na+/h+ exchangers in the male reproductive tract: relevance for male fertility. Biol. Reprod. 91, 11 (2014).
Gardner, C. C. James, P. F. The SLC9C2 gene product (Na(+)/H(+) exchanger isoform 11; NHE11) is a testis-specific protein localized to the head of mature mammalian sperm. Int. J. Mol. Sci. 24, 5329 (2023).
van der Horst, G. Computer Aided Sperm Analysis (CASA) in domestic animals: Current status, three D tracking and flagellar analysis. Anim. Reprod. Sci. 220, 106350 (2020).
Zhao, Y. et al. Alginate oligosaccharides improve germ cell development and testicular microenvironment to rescue busulfan disrupted spermatogenesis. Theranostics 10, 3308–3324 (2020).
Hao, Y. et al. Gut microbiota-testis axis: FMT improves systemic and testicular micro-environment to increase semen quality in type 1 diabetes. Mol. Med 28, 45 (2022).
Bradesi, S., Eutamene, H., Fioramonti, J. & Bueno, L. Acute restraint stress activates functional NK1 receptor in the colon of female rats: involvement of steroids. Gut 50, 349–354 (2002).
Feng, B. et al. Glucocorticoid induced group 2 innate lymphoid cell overactivation exacerbates experimental colitis. Front Immunol. 13, 863034 (2022).
Lin, R. et al. Role of melatonin in murine “restraint stress”-induced dysfunction of colonic microbiota. J. Microbiol 59, 500–512 (2021).
Li, A. et al. Gut microbiota-bile acid-vitamin D axis plays an important role in determining oocyte quality and embryonic development. Clin. Transl. Med 13, e1236 (2023).
Tegtmeyer, P. K. et al. STING induces early IFN-β in the liver and constrains myeloid cell-mediated dissemination of murine cytomegalovirus. Nat. Commun. 10, 2830 (2019).
Willemsen, J. et al. TNF leads to mtDNA release and cGAS/STING-dependent interferon responses that support inflammatory arthritis. Cell Rep. 37, 109977 (2021).
Kessler. N et al. Monocyte-derived macrophages aggravate pulmonary vasculitis via cGAS/STING/IFN-mediated nucleic acid sensing. J. Exp. Med. 219, e20220759 (2022).
Hwang, J. Y. et al. Dual Sensing of Physiologic pH and Calcium by EFCAB9 Regulates Sperm Motility. Cell 177, 1480–1494.e1419 (2019).
Li, X. et al. Calcium regulates motility and protein phosphorylation by changing cAMP and ATP concentrations in boar sperm in vitro. Anim. Reprod. Sci. 172, 39–51 (2016).
Miraglia, E. et al. Nitric oxide stimulates human sperm motility via activation of the cyclic GMP/protein kinase G signaling pathway. Reproduction 141, 47–54 (2011).
Braverman, A. M., Davoudian, T., Levin, I. K., Bocage, A. & Wodoslawsky, S. Depression, anxiety, quality of life, and infertility: a global lens on the last decade of research. Fertil. Steril. 121, 379–383 (2024).
Ye, Y. X. et al. Associations between depression, oxidative stress, and semen quality among 1,000 healthy men screened as potential sperm donors. Fertil. Steril. 117, 86–94 (2022).
Janevic, T. et al. Effects of work and life stress on semen quality. Fertil. Steril. 102, 530–538 (2014).
Wu, W., La, J., Schubach, K. M., Lantsberg, D. & Katz, D. J. Psychological, social, and sexual challenges affecting men receiving male infertility treatment: a systematic review and implications for clinical care. Asian J. Androl. 25, 448–453 (2023).
Rooney, K. L. & Domar, A. D. The impact of stress on fertility treatment. Curr. Opin. Obstet. Gynecol. 28, 198–201 (2016).
Teitelbaum, A. A., Gareau, M. G., Jury, J., Yang, P. C. & Perdue, M. H. Chronic peripheral administration of corticotropin-releasing factor causes colonic barrier dysfunction similar to psychological stress. Am. J. Physiol. Gastrointest. Liver Physiol. 295, G452–459 (2008).
Russo, S. et al. Stress-activated brain-gut circuits disrupt intestinal barrier integrity and social behaviour. Res. Sq. https://doi.org/10.21203/rs.3.rs-3459170/v1 (2023).
Xu, C., Lee, S. K., Zhang, D. & Frenette, P. S. The Gut Microbiome Regulates Psychological-Stress-Induced Inflammation. Immunity 53, 417–428.e414 (2020).
Sabater, A. S. et al. Adrenocortical activity, measured via feather corticosterone, affects semen quality but not blood plasma prolactin in the capercaillie (Tetrao urogallus L.). Eur. J. Wild. Res. 71, 23 (2025).
Chen, Y., Wang, Q., Wang, F. F., Gao, H. B. & Zhang, P. Stress induces glucocorticoid-mediated apoptosis of rat Leydig cells in vivo. Stress 15, 74–84 (2012).
Miyaso, H. et al. Neonatal corticosterone administration increases p27-positive Sertoli cell number and decreases Sertoli cell number in the testes of mice at prepuberty. Sci. Rep. 12, 19402 (2022).
Li, H. et al. Chronic alcohol-induced dysbiosis of the gut microbiota and gut metabolites impairs sperm quality in mice. Front Microbiol 13, 1042923 (2022).
Lundy, S. D. et al. Functional and Taxonomic Dysbiosis of the Gut, Urine, and Semen Microbiomes in Male Infertility. Eur. Urol. 79, 826–836 (2021).
Ramírez-Acosta, S. et al. Selenium supplementation influences mice testicular selenoproteins driven by gut microbiota. Sci. Rep. 12, 4218 (2022).
Liu, J. B., Chen, K., Li, Z. F., Wang, Z. Y. & Wang, L. Glyphosate-induced gut microbiota dysbiosis facilitates male reproductive toxicity in rats. Sci. Total Environ. 805, 150368 (2022).
Kaltsas, A. et al. Microbial dysbiosis and male infertility: understanding the impact and exploring therapeutic interventions. J. Pers. Med. 13, 1491 (2023).
Grande G, Graziani A, De Toni L, Garolla A, Ferlin A. Male Tract Microbiota and Male Infertility. Cells 13, (2024).
Guo, Y., Du, X., Bian, Y. & Wang, S. Chronic unpredictable stress-induced reproductive deficits were prevented by probiotics. Reprod. Biol. 20, 175–183 (2020).
Oliveira, L., Costa, E. C., Martins, F. D. G., Rocha, A. S. D. & Brasil, G. A. Probiotics supplementation in the treatment of male infertility: A Systematic Review. JBRA Assist Reprod. 28, 341–348 (2024).
Yan, X. et al. Gut-Testis Axis: Microbiota Prime Metabolome To Increase Sperm Quality in Young Type 2 Diabetes. Microbiol Spectr. 10, e0142322 (2022).
Han, H. et al. Alginate oligosaccharides increase boar semen quality by affecting gut microbiota and metabolites in blood and sperm. Front Microbiol 13, 982152 (2022).
Zhang, P. et al. Improvement in sperm quality and spermatogenesis following faecal microbiota transplantation from alginate oligosaccharide dosed mice. Gut 70, 222–225 (2021).
Di Ciaula, A. et al. Bile Acid Physiology. Ann. Hepatol. 16, s4–s14 (2017).
Zhao Y et al. The Role of Retinoic Acid in Spermatogenesis and Its Application in Male Reproduction. Cells 13, (2024).
Pearce, K. L., Hill, A. & Tremellen, K. P. Obesity related metabolic endotoxemia is associated with oxidative stress and impaired sperm DNA integrity. Basic Clin. Androl. 29, 6 (2019).
Wang, Z. H. & Kang, Y. F. Gut microbiota and male fertility: A two-sample Mendelian randomization study. Med. (Baltim.) 104, e41542 (2025).
de Punder, K. & Pruimboom, L. Stress induces endotoxemia and low-grade inflammation by increasing barrier permeability. Front Immunol. 6, 223 (2015).
Luo, Y. et al. MyD88 Signaling in Sertoli Cells Mediates LPS-Induced Orchitis in Mice. Reprod. Sci. 32, 3013–3024 (2025).
Sun, F., Liu, Z., Yang, Z., Liu, S. & Guan, W. The emerging role of STING-dependent signaling on cell death. Immunol. Res 67, 290–296 (2019).
Wang, W. et al. The cGAS/STING Pathway: Friend or Foe in Regulating Cardiomyopathy. Cells 14, (2025).
Zhou, J. et al. Targeting cGAS-STING signaling and cell death modes in cancer and autoimmune diseases. Cytokine Growth Factor Rev. 85, 56–80 (2025).
Ablasser, A. & Chen, Z. J. cGAS in action: expanding roles in immunity and inflammation. Science 363, 52587 (2019).
Skopelja-Gardner, S., An, J. & Elkon, K. B. Role of the cGAS-STING pathway in systemic and organ-specific diseases. Nat. Rev. Nephrol. 18, 558–572 (2022).
Tian, R. & Li, Y. Exploring the pathogenesis of MAFLD from an immunological perspective: from the perspective of the cGAS/STING/NF-κB signaling pathway. Front Immunol. 16, 1674018 (2025).
Zhu, T. et al. Aspirin alleviates particulate matter induced asymptomatic orchitis of mice via suppression of cGAS-STING signaling. Front Immunol. 12, 734546 (2021).
Miao, L. et al. Targeting the STING pathway in tumor-associated macrophages regulates innate immune sensing of gastric cancer cells. Theranostics 10, 498–515 (2020).
Zhao, J. et al. The role of stimulator of interferon genes-mediated AMPK/mTOR/P70S6K autophagy pathway in cyfluthrin-induced testicular injury. Environ. Toxicol. 38, 727–742 (2023).
Zhao, L. et al. Single-cell analysis of developing and azoospermia human testicles reveals central role of Sertoli cells. Nat. Commun. 11, 5683 (2020).
Lang, X. et al. RNA-Seq reveals the functional specificity of epididymal caput, corpus, and cauda genes of cattleyak. Anim. Sci. J. 93, e13732 (2022).
Wang, H., McGoldrick, L. L. & Chung, J. J. Sperm ion channels and transporters in male fertility and infertility. Nat. Rev. Urol. 18, 46–66 (2021).
Lin, S., Ke, M., Zhang, Y., Yan, Z. & Wu, J. Structure of a mammalian sperm cation channel complex. Nature 595, 746–750 (2021).
Ke, S. & Luo, T. The Chemosensing Role of CatSper in Mammalian Sperm: An Updated Review. Curr. Issues Mol. Biol. 45, 6995–7010 (2023).
Windler, F. et al. The solute carrier SLC9C1 is a Na(+)/H(+)-exchanger gated by an S4-type voltage-sensor and cyclic-nucleotide binding. Nat. Commun. 9, 2809 (2018).
Jin, Z. et al. The gut metabolite 3-hydroxyphenylacetic acid rejuvenates spermatogenic dysfunction in aged mice through GPX4-mediated ferroptosis. Microbiome 11, 212 (2023).
Yankelevitch-Yahav, R., Franko, M., Huly, A. & Doron, R. The forced swim test as a model of depressive-like behavior. J. Vis. Exp. 2, 52587 (2015).
Li, M. J. et al. Chronic stress exacerbates neuropathic pain via the integration of stress-affect-related information with nociceptive information in the central nucleus of the amygdala. Pain 158, 717–739 (2017).
Tian, Y. et al. Activation of CRF/CRFR1 Signaling in the Central Nucleus of the Amygdala Contributes to Chronic Stress-Induced Exacerbation of Neuropathic Pain by Enhancing GluN2B-NMDA Receptor-Mediated Synaptic Plasticity in Adult Male Rats. J. Pain. 25, 104495 (2024).
Borsoi, M. et al. Immobility behavior during the forced swim test correlates with BNDF levels in the frontal cortex, but not with cognitive impairments. Physiol. Behav. 140, 79–88 (2015).
Qi, X. et al. Fluoxetine increases the activity of the ERK-CREB signal system and alleviates the depressive-like behavior in rats exposed to chronic forced swim stress. Neurobiol. Dis. 31, 278–285 (2008).
Flemmer, D. D., Dilsaver, S. C. & Peck, J. A. Bright light blocks the capacity of inescapable swim stress to supersensitize a central muscarinic mechanism. Pharm. Biochem Behav. 36, 775–778 (1990).
Qi, X., Lin, W., Li, J., Pan, Y. & Wang, W. The depressive-like behaviors are correlated with decreased phosphorylation of mitogen-activated protein kinases in rat brain following chronic forced swim stress. Behav. Brain Res 175, 233–240 (2006).
Vega-Rivera, N. M., Fernández-Guasti, A., Ramírez-Rodríguez, G. & Estrada-Camarena, E. Acute stress further decreases the effect of ovariectomy on immobility behavior and hippocampal cell survival in rats. Psychoneuroendocrinology 38, 1407–1417 (2013).
Liu, L. Y. et al. Chronic stress increases pain sensitivity via activation of the rACC-BLA pathway in rats. Exp. Neurol. 313, 109–123 (2019).
Suarez-Roca, H., Leal, L., Silva, J. A., Pinerua-Shuhaibar, L. & Quintero, L. Reduced GABA neurotransmission underlies hyperalgesia induced by repeated forced swimming stress. Behav. Brain Res 189, 159–169 (2008).
Fang, Z. et al. Multi-omics revealed activation of TNF-α induced apoptosis signaling pathway in testis of DEHP treated prepubertal male rat. Reprod. Toxicol. 132, 108758 (2025).
Jiang, H. et al. Sensitization of neurons in the central nucleus of the amygdala via the decreased GABAergic inhibition contributes to the development of neuropathic pain-related anxiety-like behaviors in rats. Mol. Brain 7, 72 (2014).
Chen, L. et al. Activation of CRF/CRFR1 signaling in the basolateral nucleus of the amygdala contributes to chronic forced swim-induced depressive-like behaviors in rats. Behav. Brain Res 338, 134–142 (2018).
Jin, Z. R. et al. Roles of CatSper channels in the pathogenesis of asthenozoospermia and the therapeutic effects of acupuncture-like treatment on asthenozoospermia. Theranostics 11, 2822–2844 (2021).
Cao, Y. et al. Hepatic acetyl-CoA metabolism modulates neuroinflammation and depression susceptibility via acetate. Cell Metab, 37, 2185-2201 (2025).
Qi, X. et al. Gut microbiota-bile acid-interleukin-22 axis orchestrates polycystic ovary syndrome. Nat. Med 25, 1225–1233 (2019).
Zervos, I. A. et al. Effects of dietary vitamin A intake on acrosin- and plasminogen-activator activity of ram spermatozoa. Reproduction 129, 707–715 (2005).
Alsemeh, A. E. et al. Vitamin E rescues valproic acid-induced testicular injury in rats: Role of autophagy. Life Sci. 296, 120434 (2022).
Adler, I. D. Comparison of the duration of spermatogenesis between male rodents and humans. Mutat. Res 352, 169–172 (1996).
Xia, K. et al. AAV-mediated gene therapy produces fertile offspring in the Lhcgr-deficient mouse model of Leydig cell failure. Cell Rep. Med 3, 100792 (2022).
Li, K. et al. Adeno-associated-virus-mediated delivery of CRISPR-CasRx induces efficient RNA knockdown in the mouse testis. Theranostics 14, 3827–3842 (2024).
Zhang, S. et al. AAV-mediated gene therapy restores natural fertility and improves physical function in the Lhcgr-deficient mouse model of Leydig cell failure. Cell Prolif. 57, e13680 (2024).
Charan, J. & Biswas, T. How to calculate sample size for different study designs in medical research? Indian J. Psychol. Med 35, 121–126 (2013).
Zodpey, S. P. Sample size and power analysis in medical research. Indian J. Dermatol Venereol. Leprol. 70, 123–128 (2004).
Acknowledgements
This work was supported by the grants from National Natural Science Foundation of China [82371227 (G.-G.X.), 82171226 (G.-G.X.), 81974169 (G.-G.X.)], Natural Science Foundation of Beijing Municipality (7222105, G.-G.X), National Key Research and Development Program of China (2019YFC1712104, G.-G.X), and King’s College London (KCL)-Peking University Health Science Center (PKUHSC) Joint Institute for Medical Research (JI) Program (BMU2021KCL001, G.-G.X).
Author information
Authors and Affiliations
Contributions
G.-G.X. conceived the project, designed the experiments and wrote the manuscript. H.J. contributed to data interpretation and discussion. J.-C.Z. and Z.-R.J. conducted experiments, acquired data, analyzed data, and revised the manuscript. B.-H.L. conducted experiments, acquired data, analyzed data, and drafted the manuscript. Y.T. participated in experiment design, investigation, and revising the manuscript. J.-X.L. and J.C. participated in the results discussion and interpretation.
Corresponding authors
Ethics declarations
Competing interests
All authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Marco Alves, Rossella Cannarella, Sezgin Gunes, Tomas Hrncir and Kanakaraju Kaliannan for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zheng, JC., Jin, ZR., Liu, BH. et al. Chronic stress drives oligoasthenozoospermia via gut microbiota-vitamin-Sting1/Slc9c2 axis. Nat Commun 17, 4108 (2026). https://doi.org/10.1038/s41467-026-72450-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-72450-4
