
Abstract
Multi-species biofilms present a greater threat to the dairy industry than single-species biofilms due to their complex characteristics. This study examined the formation of biofilms by dairy contaminants Staphylococcus aureus and Pseudomonas fluorescens (single/co-culture) on stainless steel, and their resistance to chlorine dioxide and quaternary ammonium compounds. The findings indicated that the dual-species biofilm exhibited significantly higher biomass, cell activity, and extracellular polymeric substances production than single-species biofilms, along with a denser biofilm structure in dual-species biofilm and increased resistance to both disinfectants. Remarkably, relative quantification analysis using RT-qPCR revealed an interaction: the presence of P. fluorescens markedly upregulated the icaA and icaD genes in S. aureus. This genetic modulation was directly correlated with increased production of polysaccharide intercellular adhesion and EPS, which are crucial for biofilm maturation and structural integrity. This study offers a theoretical foundation for the hazard control of multi-species biofilms in the dairy industry.
Similar content being viewed by others

Introduction
Dairy products serve as nutrient-dense food matrices, providing bioavailable proteins, lipids, and bioactive compounds that constitute critical sources of essential nutrients for human health1. Notably, global dairy consumption has demonstrated a sustained upward trend, with the Food and Agriculture Organization (FAO) projecting a 20% growth during 2023-2032, which underscores the strong market demand for these commodities2. Bacterial biofilms, defined as structured microbial communities embedded in a self-synthesized extracellular matrix, enable bacteria to adhere to biotic/abiotic surfaces and withstand harsh environmental stresses3. In the dairy industry, these biofilms present substantial hygienic challenges: their persistent attachment to milking equipment, pipelines, and storage tanks leads to recurrent contamination of dairy products4. Staphylococcus aureus is a prevalent foodborne pathogen in the dairy industry5, primarily inducing mastitis and foodborne diseases through the production of enterotoxins6. Notably, S. aureus can form biofilms in food processing environments, allowing enterotoxin contamination to persist and escalate7. Pseudomonas fluorescences has the activity of degrading proteins and lipids in milk and can easily form biofilms on various dairy production infrastructures. The formation of biofilms intensifies its spoilage effect and highlights the crucial role of biofilm mediated contamination in the degradation of dairy products.
Studies have shown that over 70% of natural biofilms are multi-species. However, existing research on dairy biofilms has mainly focused on single-species models, which do not accurately represent the properties of multispecies biofilms8. Bacterial interactions within biofilms, including cooperative, competitive, and neutral relationships,significantly influence biofilm architecture and resistance to disinfectants9. Cooperative interactions generally promote biofilm development through mechanisms such as nutrient cross-feeding or metabolic division of labor, thus enhancing biocide resistance10. For example, It has been shown that Pseudomonas fluorescens enhanced biofilm stability in dual-species consortia with Salmonella by secreting exopolysaccharides, which increases the resistance of Salmonella to quaternary ammonium compounds11. Conversely, competitive interactions mediated by nutrient competition or the secretion of antimicrobial substances modify biofilm structure and sensitivity to disinfectants12. The findings of Zhu et al.13 indicated that dual-species biofilms of Shewanella baltica and P. fluorescens had reduced biomass and polysaccharide content compared to single-species P. fluorescens biofilms, forming fragile, and viscous spatial structures. Neutral interactions allow species to coexist without significantly altering biofilm phenotypic characteristics.
Single-species biofilm research has provided foundational insights into biofilm formation and resistance mechanisms. For example, studies on S. aureus have revealed that the agr quorum-sensing system regulates biofilm formation and dispersion at the molecular level14, P. fluorescens modulated flagellin expression to influence surface attachment during biofilm formation15. However, dual- or multi-species biofilms, which are more representative of natural dairy ecosystems, exhibit heightened disinfectant resistance compared to single-species biofilms16. This is due to complex gene regulatory networks, including inter-species signaling and horizontal transfer of resistance genes17. For instance, it was reported that co-culture of Salmonella Typhimurium and P. fluorescens downregulates genes associated with chemotaxis, epithelial invasion, and two-component systems18, Vandecandelaere et al.19 found that dual-species biofilms of Staphylococcus epidermidis and S. aureus upregulate antibiotic resistance genes in S. epidermidis.
Despite extensive research delving into disinfectant resistance mechanisms within biofilms, there are still numerous uncertainties regarding the resistance mechanisms of biofilms formed by bacterial species associated with dairy farms. The current understanding of disinfectant resistance in biofilms mainly focuses on three mechanisms: the active efflux of antimicrobials by efflux pumps20; the extracellular matrix serving as a physical barrier, where exopolysaccharides, proteins, and extracellular DNA (eDNA) sequester disinfectants21; and the survival of metabolically dormant “persister” cells during exposure to biocides22. In dual-species biofilms, interspecific interactions may enhance these mechanisms, making eradication more complex. Nevertheless, the regulatory networks that govern the formation of dual-species biofilms by S. aureus and P. fluorescens remain poorly characterized (Fig. 1).

Biofilms may form on the surfaces of pipelines and processing tanks in dairy processing scenarios (above). Detection and determination of biofilms after their formation under laboratory conditions (below).
Therefore, this study presents a comprehensive analysis of the characteristics of S. aureus and P. fluorescens biofilm, as well as their dual-species biofilm. The aim is to evaluate the efficacy of two typical commercial disinfectants against single-species and dual-species biofilms and determining the expression of biofilm-associated gene in these bacteria.
Results and discussion
Biofilm biomass
The process of biofilm formation encompasses cell adhesion, matrix secretion, biofilm formation, and bacterial dispersal23. As shown in Fig. 2 (a), the biofilm biomass of each experimental group increased with incubation time at 25 °C, peaking at 48 h after incubation and then gradually decreased, which was consistent with the growth cycle of biofilm. At 48 h, the OD 600 nm values for the single-species biofilms (SA and PF) were 1.8 and 2.9, respectively. In contrast, the dual-species biofilm exhibited a slightly higher value of 3.7. Correspondingly, the biomass accumulation in the dual-species biofilm was significantly greater than that in the single-species biofilms. This increase in biofilm formation may be attributed to cooperative interactions between the species24, which is consistent with the findings of Lin et al.25. They quantified the biofilm biomass of E. coli and S. typhimurium on SS and discovered that the biomass of dual-species biofilms increased between 2-6 fold compared to single biofilms. It can be deduced that the combination of S. aureus and P. fluorescens can promote the growth of biofilm biomass.

aureus and P. fluorescens. a Biofilm biomass; b Cell numbers. c Cell Activity Assay. d Raman spectra. e Polysaccharide content in extracellular polymers. f Protein content in extracellular polymers (SA stands for S. aureus biofilm group, PF stands for P. fluorescens biofilm group, and SP stands for biofilm formed by co-culture, SP-F stands for Cell number of P. fluorescens in co-cultured biofilms). Note: different letters (a, b, c, d, e, f) indicate significant differences over time (p < 0.05), different letters (a, b, c, d) indicate significant differences in the treatment groups (p < 0.05).
Cell numbers and cell activity assay
To visualise the specific relationship between S. aureus and P. fluorescens in the dual-species biofilm, cell counts and cell activity assays were performed on the bacterial cells of the biofilm. P. fluorescens selective medium and total colony counting medium were used to count the colonies in the biofilm. As shown in Fig. 2 (b), the cell count of P. fluorescens reached 6.8 ± 0.21 log CFU/mL at 25 °C after 48 h. Conversely, the cell count of P. fluorescens in dual-species biofilms significantly declined to 5.9 ± 0.08 log CFU/mL (p < 0.05). These results indicated that the growth of P. fluorescens was inhibited in the dual-species biofilms. Similarly, Wang et al.26 co-cultured S. aureus and Streptococcus parahaemolyticus for 24 h to form biofilms and counted the S. parahaemolyticus cells in the dual-species biofilms. The findings showed that the number of S. parahaemolyticus cells decreased significantly, which aligns with the results of this study. Furthermore, Fig. 2 (b) showed that under both single-culture and co-culture conditions, the colony counts of S. aureus and P. fluorescens exhibited similar temporal trends: they initially increase, peaked at 48 hours, and then show a declining trend. Meanwhile, the study also compared the colony counts of single- and dual-species biofilms at 24, 48, and 72 h. It was found that at 25 °C, the number of bacteria in the dual-species biofilms was significantly higher than that in the single-species biofilms, which is consistent with the study by Zarei et al.27. They found a significant increase in cell counts after co- culture of P. fluorescens and S. aureus. This indicated that there appears to be a cooperative relationship between S. aureus and P. fluorescens.
In addition, the MTT assay was employed to measure cell activity. This assay can detects metabolically active cells that reduce MTT to purple formazan. By detecting the amount of formazan produced in biofilms, the metabolic activity of viable bacteria within biofilms can be quantitatively evaluated, and the growth status of biofilms can be determined28. The high cell activity values of the bacteria indicates that the bacteria are well adapted to their environment, possibly due to the production of extracellular polymeric substances (EPS) for protection29. As shown in Fig. 2 (c), cell activity in all experimental groups (S. aureus biofilm, P. fluorescens biofilm, dual-species biofilm) exhibited time-dependent dynamics, peaking at 48 h of incubation at 25 °C before gradually declining. The cell activity of all three experimental groups was also found to decrease with the extension of incubation time. Notably, at 25 °C, metabolic activity of the dual-species biofilm formed by S. aureus and P. fluorescens was consistently higher than in single-species at matched time points, suggesting that the co-culture enhanced the activity of the cells in the biofilm. This result is consistent with the study of Ceresa et al.30 and further confirmed that co-culture of S. aureus and P. fluorescens promoted higher metabolic activity. It is worth noting that during the cultivation of the bacteria, cells often reduce their metabolic activity to survive31. These metabolically inactive cells, regardless of whether they are growing slowly or not, are typically located in the deeper layers of the biofilm. Such cells may exist in various physiological states, including viable but non-culturable (VBNC) state, persister cells, or dormant cells32. These cells cannot be captured by MTT, so this part must be taken into account when using MTT to study disinfectant resistance.
Extracellular polymeric substance (EPS) analysis
Raman spectroscopy in the 400–2600 cm−1 range was used to monitor changes in EPS in S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm groups. Table 1 presented the surface-enhanced raman spectroscopy (SERS) peak assignments for biofilm-forming bacterial strains. As shown in Fig. 2 (d), the S. aureus biofilm and dual-species biofilm groups exhibited similar spectral characteristics and chemical compositions, suggesting that the EPS compositions of S. aureus biofilm group and the dual-species biofilm group were similar. To be specific, both S. aureus and dual-species biofilms enhanced the peak intensities of the spectral bands in the range of 560–582 cm−¹. These bands were attributed to polysaccharide biomolecules and correlates with - COH, HC- C deformation of polysaccharide33. Peak intensities in the spectral bands at 1337 cm−1 and 1588 cm−1 were associated to nucleic acid34. Additionally, peak intensities in the spectral bands of 854–856 cm−1 and 1200–1280 cm−1 were detected, which were related to proteins35. The typical band at 856 cm−¹ was associated with CCH deformation in the ring breathing of tyrosine36. It indicated that after the co-culture of S. aureus and P. fluorescens, the protein and polysaccharide contents in the dual-species biofilm increased. Furthermore, in the P. fluorescens biofilm, specific peaks were observed in the regions of 780–785 cm–¹ and 1130 cm–¹ (orange line). These two peaks were attributed to the C-C torsion vibrations of cytosine (C) and uracil (U) in DNA/RNA37, and the CC, CO, and -COH bonds in carbohydrates38, respectively. The disappearance of these two peaks suggested a reduction in the of EPS contents of P. fluorescens after the formation of dual-species biofilms.
EPS, mainly composed of polysaccharides and proteins, preserve the structural integrity of biofilms and facilitate bacterial adhesion, proliferation, and protection, as well as microbial survival in the biofilm. These processes enhance microbial pathogenicity and endow resistance to antimicrobial agents39. As shown in Fig. 2 (e) and (f), representing the changes in the extracellular polymeric polysaccharide and protein content of S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm groups at three points in time. When left for 48 h at 25 °C., extracellular polysaccharide content peaked in both single- and dual-species biofilms (p < 0.05), with significantly higher levels observed in the dual-species biofilm group compared to single-species biofilm (Fig. 2 (e)). Similarly, extracellular proteins were also higher at 48 h and the protein levels were remarkably upper in the dual-species biofilm group in comparison with S. aureus biofilm and P. fluorescens biofilm groups (Fig. 2 (f)). Which is in line with the findings of Kim et al.40. Thus, co-culture of S. aureus with P. fluorescens at 25 °C to form a dual-species biofilm would increase the polysaccharides and proteins in the biofilm.
Micromorphology of biofilm
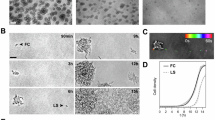
Field Emission Scanning Electron Microscope (FESEM) was used to observe the morphology and attachment patterns of cells in the S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm groups formed on SS after 48 h of incubation. The presence of only rod-shaped (P. fluorescens) and spherical (S. aureus) cells in the dual-species co-culture confirmed the absence of external bacterial contamination throughout the experiment41. Figure 3 showed that the bacterial cell on the SS remained across all groups after 48 h of incubation, it indicated that S. aureus and P. fluorescens are capable of surviving on the surface of the food processing equipment, resulting in persistent contamination. FESEM images revealed that after 48 h of incubation, bacterial cells in the dual-species biofilm group exhibited close interbacterial contact and remarkable adhesion. The overall cell density was significantly higher compared to that of the single-species biofilm group. It indicated that S. aureus and P. fluorescens can form structurally mature dual-species biofilms. Notably, after 48 h of incubation at 25 °C, the cell abundance of P. fluorescens in the dual-species biofilm group was significantly lower than that of S. aureus, which was consistent with the colony count data (Fig. 2 (b)). S. aureus was observed to grow while enclosing P. fluorescens, suggesting that P. fluorescens was under competitive inhibition in the presence of S. aureus. The increase in dual-species biofilm biomass, cell viability, and cell counts compared to the single-species biofilms indicated a cooperative interaction between the two bacterial species. From this, it can be inferred that S. aureus utilizes P. fluorescens to facilitate its own growth in dual-species biofilms.

(SA stands for S. aureus biofilm group, PF stands for P. fluorescens biofilm group, and SP stands for biofilm formed by co-culture).
To gain a deeper understanding of the biofilm morphology and structures of the S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm groups. Confocal laser scanning microscopy (CLSM) was employed to observe and acquire 2D and 3D images. Key parameters, including biofilm roughness, thickness, Biofilm volume, and porosity, were quantified and analyzed quantitatively using using ISA-2 image analysis software on the 3D images. Specifically, Roughness assesses surface irregularity, biofilm volume quantifies three-dimensional architecture, porosity characterizes the density of void spaces within the biofilm matrix, thickness determines vertical exten42.
In Fig. 4, blue fluorescence represents the distribution of viable cells in the biofilm of each treatment group at 48 h, while red fluorescence represents dead cells43. The red fluorescence distribution was minimal, indicating that dead cells were rarely distributed when the dual-species biofilm was cultured for 48 h. The 3D images showed uniform component distribution in the dual-species biofilm formed by S. aureus and P. fluorescens, suggesting that the two bacteria adapted to form a stable biofilm structure. Figure 5 showed that green fluorescence represented the overall biofilm distribution of each treatment group. The fluorescence intensity was higher at 48 h. The 3D images revealed uniform distribution of biofilm components in the dual-species biofilm.

DAPI and PI were employed to stain viable cells and dead cells. Scale bars represent 100 μm. (SA stands for S. aureus biofilm group, PF stands for P. fluorescens biofilm group, and SP stands for biofilm formed by co-culture).

The 3D CLSM images of dual-species biofilms cultured for 48 h. SYBR Green was employed to stain biofilm distribution. Scale bars represent 100 μm. (SA stands for S. aureus biofilm group, PF stands for P. fluorescens biofilm group, and SP stands for biofilm formed by co-culture).
The CLSM parameters of the biofilm were further analysed. Specifically, the biofilm thicknesses of S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm groups were presented in Fig. 6 (b), it can be observed that the biofilm thickness of the single-species cultures was low, while the thickness of P. fluorescens and S. aureus dual-species biofilm was significantly increased. The biofilm volumes of the formed single- and dual-species biofilms were shown in Fig. 6 (c), where the biofilm volume of dual-species biofilms was significantly increased (p < 0.05). It was noteworthy that the biofilm roughness and biofilm porosity had a similar tendency to form when the biofilms were incubated up to 48 h (Fig. 6 (a) and (d)). The roughness and porosity of the dual-species biofilm group was smaller than those of the single-species biofilms, indicating that the dual-species biofilms were denser than the single-species biofilms. It indicated that after 48 h of incubation at 25 °C, the dual-species biofilm formed by S. aureus and P. fluorescens was characterized by lower roughness, greater thickness, larger volume, and lower porosity. Single-species biofilms were sparser, whereas the dual-species biofilm formed by the two bacteria exhibited a more complex structure.

a Biofilm roughness. b Biofilm thickness. c Biofilm volume; d Porosity. Analysis of the inhibition rate of CD and QAC on biofilms formed by S. aureus and P. fluorescens for 48 h: e the activity of cell, f the biomass of biofilm (Control Check (CK) refers to the group that has not been treated with disinfectants, SA stands for S. aureus biofilm group, PF stands for P. fluorescens biofilm group, and SP stands for biofilm formed by co-culture). Note: different letters (a, b, c) indicate significant differences in the treatment groups (p < 0.05).
Biofilm resistance to disinfectants
This study evaluated the effectiveness of two commonly used industrial disinfectants—QAC and CD—on the formation of S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm groups. Figure 6 (e) and (f) showed that the effectiveness of CD and QAC in removing single and dual-species biofilms of S. aureus and P. fluorescens formed on SS, as well as their ability to inhibit the activity of cells in the biofilm. The ability of disinfectants to eliminate viable cells in biofilms was shown in Fig. 6 (e). The inhibitory capacities of the two disinfectants, QAC and CD, against viable bacteria in single-species biofilms of S. aureus and P. fluorescens were comparable. Nevertheless, their efficiencies significantly decreased when inhibiting viable bacteria in co-cultured biofilms, reaching only 9.3% and 11.7%, respectively. The findings indicated that both disinfectants were effective in eradicating live bacteria in the biofilm formed by the isolated cultures. The data showed that the disinfectant was significantly less potent in inhibiting the bacteria in the dual-species biofilm group than in the other two groups (p < 0.05), verifying that live bacteria in dual-species biofilms were not easy to inhibit by these two disinfectants. This further suggested that the biofilm formed by the dual-species biofilm group encapsulateed S. aureus and P. fluorescens more effectively, enabling the cells to withstand unfavourable factors in the external environment22. This was consistent with the findings of Wang et al.44, they found that cells in dual-species biofilms formed by S. aureus and P. fluorescens showed increased resistance to carvacrol compared to single-species cultures.
Consequently, the ability of the two disinfectants to eliminate biofilm biomass was further investigated. Specifically, as shown in Fig. 6 (f), CD removed 37.0% and 48.8% of the biomass from S. aureus and P. fluorescens biofilms, respectively. In contrast, QAC removed 22.8% and 35.9% of the biomass from S. aureus and P. fluorescens biofilms, respectively. However, both disinfectants were less effective (p < 0.05) against dual-species biofilms compared to single-species biofilms, with CD and QAC removing 22.4% and 13.1%, respectively. The results indicated that dual-species biofilms showed increased resistance to disinfectants. This finding is in line with the observations reported by Lin et al.25, they conducted a study on the chlorine disinfection of single and dual-species biofilms of E. coli O45 and O121, as well as S. typhi, on SS. The reason for this was the presence of an extracellular matrix and the structural complexity of pathogenic organism structures in the dual-species biofilm, which led to greater resistance22. Evidently, both disinfectants were less effective in removing biofilms. It indicated the limitations of chemical methods such as detergents and disinfectants in completely eliminating biofilms. Therefore, to ensure the continued health and safety of the food industry, it is crucial to take measures to enhance the removal of biofilms.
Real-time fluorescence quantitative PCR
RT-qPCR, a technique that integrates reverse transcription with real-time quantitative PCR, offers high sensitivity and specificity, enabling precise quantification of RNA expression levels. This method is extensively utilized in gene expression analysis due to its accuracy and reliability45. The biofilm-related genes of S. aureus and P. fluorescens were presented in Table 2. As shown in Fig. 7 (a), the expression of flgA gene in P. fluorescens was significantly down-regulated by 2.3-fold when the bacterium was co-cultured with S. aureus (p < 0.05). The flgA gene is a key regulatory gene for cell adhesion in P. fluorescens, which has a role in encoding flagella. The flagella is necessary for the bacteria to approach and form a durable bond with the target surface, and the mediated movement is essential for biofilm formation, which increases the likelihood that the bacteria will interact with the contacting surface and contributes to the establishment of a physical framework in the biofilm matrix46. The observed downregulation of the flgA gene in P. fluorescens during co-culture with S. aureus suggested an altered flagellum-associated capacity that may affect biofilm formation or structural stability.

a flgA; b pgaA; c pgaC; d pgaD; e lapD. Effect of co-culture on the expression of regulatory genes in S. aureus: f icaA; g icaB; h icaD. Note: **significant difference between different tissues treatment (p < 0.05); ***very significant difference between different tissues (p < 0.01) (SA stands for S. aureus biofilm group, PF stands for P. fluorescens biofilm group, and SP stands for biofilm formed by co-culture).
Similarly, as shown in Fig. 7 (b), (c), and (d), the co-culture with S. aureus significantly altered the gene expression profile of P. fluorescens. Specifically, the pgaC gene of P. fluorescens was down-regulated by 1.46-fold, and the pgaD gene expression was also down-regulated by 4.3-fold (p < 0.01). These genes constitute the pgaABCD operon, encoding enzymes responsible for synthesizing and exporting poly- β- 1,6-N- acetyl- D- glucosamine (PGA), a biofilm exopolysaccharide critical for structural integrity and microbial adhesion47. Specifically, pgaC plays a pivotal role in the biosynthesis of adhesins by catalyzing the polymerization of UDP- N-acetylglucosamine monomers to produce PGA. PgaA is responsible for exporting these biofilm adhesins to the outer layer of the biofilm, while PgaD assists glycosyltransferase pgaC in the polymerization process. Overall, pgaA, pgaC and pgaD are required for adhesion formation in P. fluorescens biofilms48. Downregulation of adhesion-related genes in P. fluorescens suggested impaired adherence to SS under co-culture conditions with S. aureus. Additionally, the lapD gene contributed to biofilm matrix and surface attachment in P. fluorescens. After co-culture with S. aureus, lapD gene expression was down-regulated by 2.4-fold. LapD regulates surface attachment, which is necessary for the formation of biofilms as a biofilm protein49. Zemke et al.50 found that the absence of lapD resulted in biofilm dispersal across surfaces, emphasizing the critical role of the lapD gene in maintaining biofilm structural integrity and adhesion. Collectively, these findings indicated that the biofilm-forming capacity of P. fluorescens was compromised in the presence of S. aureus. The observed gene downregulation likely revealed an inhibitory effect of S. aureus on P. fluorescens, thereby disrupting the genetic machinery required for biofilm development and surface adhesion. This finding aligns with the report by Frozi et al.51, they demonstrated that when S. aureus and Escherichia coli were co-cultured for 8 h, S. aureus dominated in the dual-species biofilm.
As shown in Fig. 7 (f), the gene expression of icaA in S. aureus was 0.3 ± 0.04. Following co-culture with P. fluorescens, the gene expression of icaA was significantly upregulated (0.4 ± 0.01; p < 0.05). EPS, including polysaccharide intercellular adhesion (PIA), extracellular DNA, proteins, and amyloid fibrils, play a crucial role in the structure of the S. aureus biofilm. The icaA gene encodes N- acetylglucosaminyltransferase, which catalyzes the polymerization of UDP-N-acetylglucosamine into PIA oligomers, a critical component of biofilm matrix52. Thus, the upregulation of icaA gene expression was associated with enhanced PIA synthesis, a key factor for the integrity of the S. aureus biofilm matrix integrity and intercellular adhesion. Furthermore, icaB encodes an N-deacetylase responsible for the partial deacetylation of PIA53. This partial deacetylation of PIA stabilizes the biofilm structure through charge cross-linking and resistance to degradation, promoting cell adhesion and aggregation and enhancing antibiotic resistance54. Figure 7 (g) indicated that the gene expression of icaB in S. aureus cells was 0.9 ± 0.09. When co-culture with P. fluorescens, the gene expression of icaB was up-regulated, and the gene expression level was 1.2 ± 0.07. Evidently, the increased expression of the icaB gene further demonstrated that interspecies interaction promoted the formation of the S. aureus biofilm.
Figure 7 (h) indicated that after co-culturing with P. fluorescens, the expression level of the icaD gene in S. aureus increased significantly, with a 2.7-fold up-regulation. IcaD is the product that optimises the efficiency of icaA55, icaD can function as an auxiliary protein and synergise with icaA to maximise the catalytic efficiency of icaA, enhance its enzyme activity and stability. Consequently, its effectively synthesized intercellular polysaccharides, promoted the formation of S. aureus biofilms, and enhanced bacterial adhesion and drug resistance56. This suggested that P. fluorescens improved adhesion of S. aureus on the SS, leading to greater biofilm biomass. These findings further confirmed that S. aureus inhibited P. fluorescens while promoted its own growth, as well as increased biomass, cell activity, extracellular polysaccharide, and protein content. Therefore, the dominance of S. aureus in the dual-species biofilm group led to a remarkably increase in the expression of icaA and icaD genes. The combined effect of icaA and icaD led to an increase in PIA, which in turn increased polysaccharide content and enhanced disinfectant resistance. Consistent with these results, Xiang et al.57 found that under cadmium (Cd) stress, both single- and co-cultured species exhibited significantly higher surface polysaccharide content compared to the Cd-free control, suggesting that bacteria can resist external stress by increasing cell surface polysaccharides.
Biofilms in dairy processing environments pose significant risks, including product contamination, spoilage, and safety hazards, ultimately endangering consumer health and product quality. These risks are further exacerbated by the fact that multi-species biofilms exhibit greater structural and functional resilience compared to single-species biofilms. This study aimed to investigate in depth the biofilm formation characteristics of S. aureus and P. fluorescens in both single-species and dual-species biofilm. Firstly, a systematic quantitative analysis system was established. The CV assay was used to quantify the biofilm-forming capacity of single-species biofilms of the two strains and their dual-species biofilm. It was found that the biomass significantly increased when the two bacteria were co-cultured (Fig. 2(a)). Meanwhile, the MTT assay combined with cell counting was employed to reflect the number of viable cells within the biofilms, preliminarily indicating a synergistic effect between S. aureus and P. fluorescens in the dual-species biofilm. Secondly, counting of P. fluorescens in the dual-species biofilm revealed that its cell number was significantly lower compared to that in single-species culture. This observation was further validated by SEM images (Fig. 3), leading to the speculation that P. fluorescens is inhibited in the dual-species biofilm.
On the basis of quantitative analysis, Raman spectroscopy was used to conduct in-depth analysis of the component distribution in single-species and dual-species biofilms. Raman spectroscopic analysis demonstrated that the component distribution of S. aureus single-species biofilm was highly similar to that of the formed dual-species biofilm, further supporting the inference that S. aureus plays a predominant role in the dual-species biofilm. In addition, this study detected the content of EPS in the biofilms. By determining the contents of major EPS components (polysaccharides and proteins), it was found that the extracellular polysaccharide and protein contents in the dual-species biofilm were significantly higher than those in single-species biofilms (Fig. 2 (e) and (f)). From this, it can be inferred that in the dual-species biofilm, S. aureus may inhibit the growth of P. fluorescens to promote its own proliferation, thereby contributing to the overall growth of the dual-species biofilm.
To further reveal the spatial structure of the biofilms, CLSM was used for three-dimensional structural observation. After processing these three-dimensional images with ISA-2 software, it was found that during the formation of the dual-species biofilm, the interaction between S. aureus and P. fluorescens was intense, which promoted the overall growth of the dual-species biofilm and led to the formation of larger and more compact aggregates58. The increase in the number and density of these aggregates, combined with the verification from MTT assay and EPS content determination, indicated active cell proliferation and vigorous extracellular matrix synthesis59. Such structural maturity is a hallmark of well-developed biofilms, where cells are embedded in a self-produced EPS matrix. This matrix not only facilitates nutrient exchange but also provides structural stability and protects the biofilm from environmental stresses (e.g., disinfectants)60. These findings are of great significance for understanding the disinfectant resistance of dual-species biofilms.
Our results showed that the dual-species biofilm formed by these two common bacteria in the dairy industry exhibited stronger resistance to commercial disinfectants compared to single-species biofilms (Fig. 6 (e) and (f)). Meanwhile, gene regulation analysis using RT-qPCR revealed that the biofilm-related genes icaA and icaD were significantly up-regulated. Studies have shown that icaD can enhance the activity of icaA and maintain its stability by promoting its structural conformation, thereby increasing polysaccharide content. This process promotes biofilm formation and enhances resistance to disinfectants. These findings provide a suitable model for managing potential biofilm hazards in dairy products. A key challenge facing the dairy industry is the ineffectiveness of sanitizers in removing biofilms. This issue can be addressed by combining sanitizers with bioactive ingredients, antimicrobial peptides, and other components to achieve effective biofilm eradication.
Methods
Materials
Tryptic Soy Broth (TSB), Luria-Bertani Broth (LB), King B Medium (KB) were purchased from Qingdao Hi-Tech Haibo Biotechnology Co., Ltd. (Qingdao, China); Labgic Technology Co., Ltd. (Beijing, China) offered 24-well plates; 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenylam-monium tetrabromide (MTT), 4% glutaraldehyde, and SYBR Green I were obtained from Macklin Biochemical Technology Co., Ltd. (ShangHai, China); chlorine dioxide (CD) was purchased from Foshan Green Companion E-commerce Co., Ltd. (Foshan, China), quaternary ammonium compounds (QAC) were acquired from Shanghai yuanye Bio- Technology Co., Ltd (ShangHai, China).
Strain information and culture conditions
S. aureus ATCC 6538 was storaged at Laboratory for quality and safety risk assessment of aquatic products in storage and preservation of Ministry of Agriculture and Rural Affairs, P. fluorescens CICC 23250 was isolated from spoiled dairy products and purchased from China Centre for the Preservation and Management of Industrial Microbial Strains, Beijing. S. aureus ATCC 6538, P. fluorescens CICC 23250 were incubated in LB at 30 °C for 12 h, and then 200 μL bacterial solution were resuspended in fresh LB at 30 °C for 12 h, respectively. The absorbance of the solution was controlled at 0.8–1.0 (106 CFU/mL) by serial dilution and set aside for use.
Biofilm formation
The stainless steel (SS, AISI304) (12 × 12 mm, Wuxi Safeway Trading Co., Ltd., China) were immersed in 95% ethanol for 30 min to remove grease from the SS, then washed with 75% ethanol and rinsed with water before being sterilised. The treated SS was then first laid flat in a 24-well plate with 1 mL of LB liquid medium and 20 μL of bacterial solution. There were three treatment groups, S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm, respectively. The initial ratio of S. aureus and P. fluorescens was 1:1 in dual-species biofilm at the time of inoculation. The three treatment groups were cultured at 25 °C for 3 days, with sampling conducted at 12-h intervals (12 h, 24 h, 36 h, 60 h, and 72 h). At each sampling point, the samples were removed from the culture medium using sterile forceps, followed by three successive rinses with 0.01 M PBS to remove non-adherent cells. The rinsed samples were then transferred to a sterile 24-well plate for subsequent analysis60.
Crystal violet assay
The crystal violet (CV) assay was used to quantify biofilms, with a slight modification of the method assay in reference to Guo et al.61. Briefly, 1 ml 0.1% CV solution was added to each well and then stained with CV for 20 min. Wash three times with purified water to remove excess CV solution, followed by drying at 60 °C for 10 min, and elute with 1 mL of 95% ethanol for 5 min to dissolve. The absorbance value was determined at 600 nm by transferring 200 μL to an enzyme plate.
Cell numbers of biofilms
The counting of viable cells in biofilms was modelled on the method of Wang et al.26 with slight modifications. Briefly, S. aureus, P. fluorescens and both strains were incubated on SS to form biofilms. The biofilm samples from the three time points (24, 48 and 72 h) were selected and planktonic cells were removed by rinsing with 0.01 M PBS. The samples were then transferred to new 24-well plates. Biofilms were disrupted by repeated pipetting with a 1 mL of physiological saline and ultrasonic treatment (40 KHZ) for 20 s to create mixed biofilm samples, which were then dispersed into KB and TSB media by gradient dilution. Select 2-3 appropriate dilution ratios, take 100 μL of each and spread on the surface of the plate, incubate at 37 °C for 24–48 h, select plates with colony counts between 30 and 300 CFU, count the colonies, multiply by the dilution ratio and then by 10 to obtain the cell count. Moreover, KB medium was used to count P. fluorescens in the mixed biofilm.
Cell activity assay
Cell activity was appraised by using MTT assay62. To assess metabolic activity, SS were immersed in 24-well plates containing 1 mL of fresh tryptic soy broth (TSB) supplemented with 100 μL of 5 mg/mL MTT solution, and incubated at 25 °C for 4 h. After aspirating the liquid, 1 mL of dimethylsulfoxide (DMSO) was added to dissolve formazan crystals, and the samples were allowed to stand for 30 min. Then the absorbance of the samples was measured at optical density 570.
Analysis of EPS
Raman spectroscopy was used to analyze EPS, according to Liu et al.63. Biofilm cells were suspended by adding 0.01 M potassium chloride solution. The suspension was sonicated for 5 s at a time at 5 s intervals for a total of 1 min. After centrifugation at 4,000 rpm for 20 min at 4 °C, the supernatant was filtered through a 0.22 μm membrane to sterilize it and remove residual particulates. The filtered solution was pre-frozen at −80 °C and then lyophilized at low temperature to obtain powders. Six positions in the EPS were randomly spoted and analysed by plotting the average of the peak intensities. Raman spectra were recorded with a 633 nm semiconductor laser and a 50 × objective lens (laser power of 3 mw) with an integration time of 1 min. The average Raman spectra were analysed and plotted by Origin 2021 (OriginLab Corp., Northampton, USA). Polysaccharide and protein concentrations were determined by sulphuric acid-phenol method (Polysaccharide content determination kit, Shanghai Shangbao Biotechnology Co., Ltd., Shanghai) and lowry method, respectively.
FESEM
The biofilms were analysed by FESEM (Tescan Mira 3 XH, TESCAN Trading (Shanghai) Co., Ltd., Shanghai) in accordance with Yu et al.64. Take samples cultured on SS for 48 h. Fix with 2.5% glutaraldehyde at 4 °C for 4 h. After removing the fixative, dehydrate for 15 min with a series of gradually increasing ethanol solutions (30%, 50%, 70%, 80%), respectively. Then, the sample surface was treated with low-temperature freeze drying. A uniform 5 nm-thick film of nano-gold particles was sprayed onto the surface of the sample. The sample was then placed into the sample chamber, and the chamber was evacuated to create a vacuum. The voltage was adjusted to 5.0 kV. Images were captured at magnifications of 2000 x, 5000 x, and 10,000 x, with more than three points selected at each magnification, resulting in a total of more than 12 images taken.
CLSM
Biofilms were fixed with 1 mL 4% (w/v) glutaraldehyde at 4 °C for 30 min. Excess fixative was rinsed with 0.01 M PBS, and samples were stained with DAPI, PI, and SYBR Green (1× concentration) for 30 min under light-protected conditions. After three 0.01 M PBS washed to remove unbound dye, biofilm samples were visualized using a CLSM (FV3000, Olympus Corp., Janpan). Three random fields of view per sample were imaged, and structural parameters-including roughness, thickness, biofilm volume, and porosity-were quantified using ISA-2 software. The fluorescences were detected using the following combination of laser excitation and emission band- pass wavelengths: 358/461 nm for DAPI, 535/617 nm for PI, and 497/520 nm for SYBR Green35.
Biofilm resistance to disinfectants
The resistance of biofilm to disinfectants was determined using the method of Chaves et al.65 with minor modification. SS with biofilm formation after 48 h were taken and washed with 0.1 M PBS buffer, then treated with chlorine dioxide (CD), quaternary ammonium compounds (QAC) at 100 ppm for 5 min and immersed in sterile 0.1 M PBS solution as a control. The surfaces where gently washed and then transferred to new 24-well plates. Biomass and cell activity of disinfectant-treated biofilm surfaces were quantified by CV and MTT assays, respectively.
RT-qPCR
RT-qPCR was employed to investigate the genetic regulation of dual-species biofilm formation by S. aureus and P. fluorescens, following the methodological framework proposed by Song et al.41. The procedures were as follows:
Sequencing and Data Processing. Bacterial Sequencing: Both bacterial strains were subjected to genome sequencing. Data Filtering: Raw sequencing data were processed using fastp software to remove low-quality reads and joint sequences, yielding clean reads. Gene Identification: The clean reads and provided reference sequences were analyzed via blastn (-evalue 0.00001) to obtain biofilm-related gene sequences (Table 2).
Experimental Incubation and Sample Preparation. Treatment Groups: Three groups were incubated at 25 °C for 48 h. Cell Harvesting: Cells were pelleted by centrifugation at 12,000 rpm for 10 min at 4 °C. The supernatant was discarded, and the pellet was stored at −20 °C for subsequent RNA extraction.
RNA Extraction and Reverse Transcription. RNA Extraction: Total RNA was extracted using the Rapid Cellular RNA Extraction Kit (Shandong Scitech Biotechnology Co., Ltd., China), and the quality of extracted RNA samples was assessed. cDNA Synthesis: Reverse transcription was performed using the following system: 5 μL RNA, 1 μL gDNA Eraser, 10 μL 2× SPARKscript II All-in-one qRT SuperMix, RNase-Free H2O up to 20 μL. Reaction conditions: 50 °C for 15 min, followed by 85 °C for 30 s in a PCR machine. Concentration Normalization: cDNA concentrations from S. aureus biofilm, P. fluorescens biofilm, and dual-species biofilm treatments were normalized using a fully automated protein nucleic acid analyzer (Veriti™ 96-Well Thermal Cycler, Thermo Fisher (Shanghai) Instruments Co., Ltd., China), then stored at −20 °C.
RT-qPCR Analysis. Housekeeping Gene and Primers: 16S rRNA served as the housekeeping gene, and biofilm-related gene primers were designed and synthesized for RT-qPCR. Amplification System: 2X SGExcel FastSYBR Mixture 10 μL, Forward and reverse primers (10 μmol/L) 0.4 μL each, 2 μL gradient-diluted cDNA template, ddH2O up to 20 μL total volume. Reaction Protocol: Preincubation: 95 °C for 3 min, 40 cycles: 95 °C for 5 s (denaturation), 60 °C for 20 s (annealing/extension), Cooling: 37 °C for 30 s. Data Collection: Cycling threshold (CT) values were recorded with 3 replicates per treatment. Relative gene expression was calculated using the 2-ΔΔCt method, and dissolution curves were analyzed using a LightCycler® 96 Instrument (Shanghai Roche Pharmaceutical Co., Ltd., China).
Statistical analysis
Unless otherwise stated, all experiments were conducted in triplicate. Statistical significance was analysed using one-way ANOVA followed by Duncan’s test, with significance expressed as p < 0.05. Data were analysed using SPSS 26.0 (SPSS- Statistical Package for the Social Sciences, Inc.) and results are expressed as mean ± standard deviation (SD), different letters (a, b, c, d, e, f) indicate significant differences (p < 0.05). By calculating the skewness coefficient and kurtosis coefficient, the experimental data all follows a normal distribution. Utilizing Origin 2021 (OriginLab Corp., Northampton, USA) to generated figures and graphs.
Data availability
Data are provided within the manuscript or supplementary information files.
References
Wu, H., Li, S., Chen, L., Xia, Y. & Tan, X. Intake of the different types of dairy products, genetic predisposition, and the risks of nonalcoholic fatty liver disease and cirrhosis: a prospective cohort study. Food Funct. 15, 5050–5062 (2024).
OECD, Food & Nations, A.O.o.t.U. OECD-FAO Agricultural Outlook 2023-2032 (2023).
Das, S. Genetic regulation, biosynthesis and applications of extracellular polysaccharides of the biofilm matrix of bacteria. Carbohydr. Polym. 291, 119536 (2022).
Chowdhury, M. A. H., Chowdhury, S. A. R., Rahman, S. M. & Sarkar, F. Comprehensive Approaches for Ensuring Microbial Safety in the Dairy Industry: Monitoring Systems, Inhibitory Strategies, and Future Prospects. Food Control, 110894 (2024).
Emiliano, J. V.dS. et al. Staphylococcus aureus in Dairy Industry: Enterotoxin Production, Biofilm Formation, and Use of Lactic Acid Bacteria for Its Biocontrol. Foodborne Pathog. Dis. 21, 601–616 (2024).
Kümmel, J. et al. Staphylococcus aureus Entrance into the Dairy Chain: Tracking S. aureus from Dairy Cow to Cheese. Front. Microbiol. 7 (2016).
Shen, J. et al. Effect of biofilm on the survival of Staphylococcus aureus isolated from raw milk in high temperature and drying environment. Food Res Int. 149, 110672 (2021).
Diarra, C. et al. Biofilm formation by heat-resistant dairy bacteria: multispecies biofilm model under static and dynamic conditions. Appl. Environ. Microbiol. 89, e00713-23 (2023).
Zeng, X. et al. Quorum sensing-mediated microbial interactions: Mechanisms, applications, challenges and perspectives. Microbiological Res. 273, 127414 (2023).
Wang, L.; Wang, X.; Wu, H.; Wang, H.; Lu, Z. Interspecies synergistic interactions mediated by cofactor exchange enhance stress tolerance by inducing biofilm formation. mSystems 9, (2024)
Pang, X., Chen, L. & Yuk, H.-G. Stress response and survival of Salmonella Enteritidis in single and dual species biofilms with Pseudomonas fluorescens following repeated exposure to quaternary ammonium compounds. Int. J. Food Microbiol. 325, 108643 (2020).
De Wit, G., Svet, L., Lories, B. & Steenackers, H. P. Microbial Interspecies Interactions and Their Impact on the Emergence and Spread of Antimicrobial Resistance. Annu. Rev. Microbiol. 76, 179–192 (2022).
Zhu, J., Yan, Y., Wang, Y. & Qu, D. Competitive interaction on dual-species biofilm formation by spoilage bacteria, Shewanella baltica and Pseudomonas fluorescens. J. Appl. Microbiol. 126, 1175–1186 (2019).
Derakhshan, S., Navidinia, M. & Haghi, F. Antibiotic susceptibility of human-associated Staphylococcus aureus and its relation to agr typing, virulence genes, and biofilm formation. BMC Infect. Dis. 21, 1–10 (2021).
Jung, Y.-C., Lee, M.-A. & Lee, K.-H. Role of flagellin-homologous proteins in biofilm formation by pathogenic Vibrio species. MBio 10, 4 (2019).
Li, H. et al. Microbial biodiversity of raw milk collected from Yangzhou and the heterogeneous biofilm-forming ability of Pseudomonas. Int. J. Dairy Technol. 76, 51–62 (2022).
Feng, Y. et al. Microplastics exhibit accumulation and horizontal transfer of antibiotic resistance genes. J. Environ. Manag. 336, 117632 (2023).
Yuan, L. et al. Dual-species biofilm and other profiles altered by interactions between Salmonella Typhimurium and Pseudomonas fluorescens isolated from meat. Food Res. Int. 203, 115914 (2025).
Vandecandelaere, I., Van Nieuwerburgh, F., Deforce, D. & Coenye, T. Metabolic activity, urease production, antibiotic resistance and virulence in dual species biofilms of Staphylococcus epidermidis and Staphylococcus aureus. PLoS ONE 12, e0172700 (2017).
Tong, C. et al. Disinfectant resistance in bacteria: Mechanisms, spread, and resolution strategies. Environ. Res. 195, 110897 (2021).
Li, Y. et al. From “resistance genes expression” to “horizontal migration” as well as over secretion of Extracellular Polymeric Substances: Sludge microorganism’s response to the increasing of long-term disinfectant stress. J. Hazard. Mater. 469, 133940 (2024).
Maillard, J.-Y. & Pascoe, M. Disinfectants and antiseptics: mechanisms of action and resistance. Nat. Rev. Microbiol. 22, 4–17 (2024).
Alonso, V. P. & Kabuki, D. Y. Formation and dispersal of biofilms in dairy substrates. Int. J. Dairy Technol. 72, 472–478 (2019).
Alonso, V. P. P. Harada, A. M. M. Kabuki, D. Y. Competitive and/or Cooperative Interactions of Listeria monocytogenes With Bacillus cereus in Dual-Species Biofilm Formation. Front. Microbiol. 11 (2020)
Lin, Z. et al. Dual-Species Biofilms Formed by Escherichia coli and Salmonella Enhance Chlorine Tolerance. Appl. Environ. Microbiol. 88, e01482-22 (2022).
Wang, L. et al. Formation of a biofilm matrix network shapes polymicrobial interactions. ISME J. 17, 467–477 (2023).
Zarei, M., Rahimi, S., Saris, P. E. J. & Yousefvand, A. Pseudomonas fluorescens group bacterial strains interact differently with pathogens during dual-species biofilm formation on stainless steel surfaces in milk. Front Microbiol 13, 1053239 (2022).
Oh, Y. J. & Hong, J. Application of the MTT-based colorimetric method for evaluating bacterial growth using different solvent systems. Lwt 153, 112565 (2022).
Constante, C. K., Rodríguez, J., Sonnenholzner, S. & Domínguez-Borbor, C. Adaptation of the methyl thiazole tetrazolium (MTT) reduction assay to measure cell viability in Vibrio spp. Aquaculture 560, 738568 (2022).
Ceresa, C. et al. Inhibitory Effects of Lipopeptides and Glycolipids on C. albicans-Staphylococcus spp. Dual-Species Biofilms. Front. Microbiol. 11 (2021).
Braissant, O., Astasov-Frauenhoffer, M., Waltimo, T. & Bonkat, G. A review of methods to determine viability, vitality, and metabolic rates in microbiology. Front. Microbiol. 11, 547458 (2020).
Alonso, V. P. P. et al. Microbial resistance to sanitizers in the food industry: review. Crit. Rev. Food Sci. Nutr. 64, 654–669 (2022).
De Gussem, K., Vandenabeele, P., Verbeken, A. & Moens, L. Raman spectroscopic study of Lactarius spores (Russulales, Fungi). Spectrochimica Acta Part A: Mol. Biomolecular Spectrosc. 61, 2896–2908 (2005).
Shakeel, M. et al. Surface-enhanced Raman spectroscopy for the characterization of pellets of biofilm forming bacterial strains of Staphylococcus epidermidis. Photodiagnosis Photodyn. Ther. 40, 103145 (2022).
Tan, L. et al. Dual-species biofilms formation of Vibrio parahaemolyticus and Shewanella putrefaciens and their tolerance to photodynamic inactivation. Food Control 125, 107983 (2021).
de Siqueira, E. et al. Biochemical characterization of pathogenic bacterial species using Raman spectroscopy and discrimination model based on selected spectral features. Lasers Med. Sci. 36, 289–302 (2021).
Chen, B. et al. Eradication of planktonic Vibrio parahaemolyticus and its sessile biofilm by curcumin-mediated photodynamic inactivation. Food Control 113, 107181 (2020).
Naman, A. et al. Surface-enhanced Raman spectroscopy for characterization of supernatant samples of biofilm forming bacterial strains. Spectrochimica Acta Part A: Mol. Biomolecular Spectrosc. 305, 123414 (2024).
Yang, X., Lan, W. & Sun, X. Effects of chlorogenic acid-grafted-chitosan on biofilms, oxidative stress, quorum sensing and c-di-GMP in Pseudomonas fluorescens. Int. J. Biol. Macromolecules 273, 133029 (2024).
Kim, U., Kim, J.-H., Lee, S.-Y. & Oh, S.-W. Detection of Bacillus cereus and Pseudomonas fluorescens in dual-species biofilm via real-time PCR and eradication using grapefruit seed extract. LWT 165, 113679 (2022).
Song, D. et al. Co-culture of Cronobacter sakazakii and Staphylococcus aureus: Explore the influence of mixed biofilm formation and regulation of biofilm formation genes. Food Res. Int. 173, 113457 (2023).
Wang, Z. et al. Characterization of Listeria monocytogenes biofilm formation kinetics and biofilm transfer to cantaloupe surfaces. Food Res. Int. 161, 111839 (2022).
Arjharn, W. et al. The detection of Escherichia coli biofilm in green oak with spectroscopic imaging techniques based on confocal laser scanning microscope. E3S Web of Conferences 187, (2020).
Wang, Y., Hong, X., Liu, J., Zhu, J. & Chen, J. Interactions between fish isolates Pseudomonas fluorescens and Staphylococcus aureus in dual-species biofilms and sensitivity to carvacrol. Food Microbiol. 91, 103506 (2020).
Hou, Q. et al. Identification and validation of reference genes for normalization of gene expression analysis using qRT-PCR in Megalurothrips usitatus (thysanoptera: thripidae). Front. Physiol. 14, 1161680 (2023).
Myszka, K. et al. Role of gallic and p-coumaric acids in the AHL- dependent expression of flgA gene and in the process of biofilm formation in food-associated Pseudomonas fluorescens KM120. J. Sci. Food Agric 96, 4037–4047 (2016).
Kang, J. et al. The specific effect of gallic acid on Escherichia coli biofilm formation by regulating pgaABCD genes expression. Appl. Microbiol. Biotechnol. 102, 1837–1846 (2018).
Poulin, M. B. & Kuperman, L. L. Regulation of biofilm exopolysaccharide production by cyclic di-guanosine monophosphate. Front. Microbiol. 12, 730980 (2021).
Newell, P. D., Monds, R. D. & O’Toole, G. A. LapD is a bis-(3′,5′)-cyclic dimeric GMP- binding protein that regulates surface attachment by Pseudomonas fluorescens Pf0-1. Proc. Natl Acad. Sci. 106, 3461–3466 (2009).
Zemke, A. C. et al. Bacterial respiratory inhibition triggers dispersal of Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 89, e01101–e01123 (2023).
Frozi, J. B., Esper, L. M. R. & Franco, R. M. Single- and Multispecies Biofilms by Escherichia coli, Staphylococcus aureus, and Salmonella spp. Isolated from Raw Fish and a Fish Processing Unit. Ciência Rural 47 (2017).
Kutsuno, S. et al. Non- deacetylated poly- N-acetylglucosamine- hyperproducing Staphylococcus aureus undergoes immediate autoaggregation upon vortexing. Front. Microbiol. 13, 1101545 (2023).
Yanagihara, A. et al. Polysaccharide intercellular adhesin and proper phospholipid composition are important for aggregation in Tetragenococcus halophilus SL10. Appl. Environ. Microbiol. 90, e00334–00324 (2024).
Pugazhendhi, A. S., Wei, F., Hughes, M. & Coathup, M. Bacterial Adhesion, Virulence, and Biofilm Formation. In: Coathup, M. (eds) Musculoskeletal Infection, 19-64 (2022).
El-Sawaf, G. et al. Identifying biofilm-forming strains of Staphylococcus epidermidis isolated from intravascular-catheterized patients by icaA and icaD genes. Chiang Mai Univ. J. Nat. Sci. 21, 1–10 (e2022024) (2022).
Zhang, T., Luo, X., Xu, K. & Zhong, W. Peptide- containing nanoformulations: Skin barrier penetration and activity contribution. Advanced Drug Delivery Reviews, 115139 (2023).
Xiong, X., Othmer, H. G. & Harcombe, W. R. Emergent antibiotic persistence in a spatially structured synthetic microbial mutualism. ISME J. 18, wrae075 (2024).
Akter, S., Rahman, M. A., Ashrafudoulla, M. & Ha, S.-D. Biofilm formation and analysis of EPS architecture comprising polysaccharides and lipids by Pseudomonas aeruginosa and Escherichia coli on food processing surfaces. Food Res. Int. 209, 116274 (2025).
Flemming, H.-C. et al. Microbial extracellular polymeric substances in the environment, technology and medicine. Nat. Rev. Microbiol. 23, 87–105 (2025).
Flemming, H.-C. et al. The biofilm matrix: multitasking in a shared space. Nat. Rev. Microbiol. 21, 70–86 (2023).
Onuoha, C., Stephenson-Clarke, S., Thoms-Rodriguez, C. A. & Brown, P. MIXED-SPECIES BIOFILMS AND RESISTANCE TO HEAVY METALS AND DISINFECTANTS: IMPLICATIONS FOR UPEC AND UTIS. Int. J. Infect. Dis. 130, S29 (2023).
Guo, Z., Xiong, R., Yan, J. & Xie, J. Role of bpfA in adhesion and biofilm formation of Shewanella putrefaciens CN32 under cold stress: a comprehensive transcriptomic analysis. Food Science and Human Wellness (2024).
Tan, L. J. et al. Potent eradication of mixed-species biofilms using photodynamic inactivation coupled with slightly alkaline electrolyzed water. 155 (LWT-FOOD SCIENCE AND TECHNOLOGY, 2022).
Liu, H. et al. Punicalagin inhibits biofilm formation and virulence gene expression of Vibrio parahaemolyticus. Food Control 139 (2022).
Chaves, R. D. et al. Comparing the susceptibility to sanitizers, biofilm-forming ability, and biofilm resistance to quaternary ammonium and chlorine dioxide of 43 Salmonella enterica and Listeria monocytogenes strains. Food Microbiol. 117, 104380 (2024).
Acknowledgements
This work was supported by the Open Project Program of State Key Laboratory of Dairy Biotechnology (No. SKLDB2023-007).
Author information
Authors and Affiliations
Contributions
All authors did a investigation and provided the methodology, Jiaqi Yuan wrote the original draft, prepared Figures 1-7 and Tables 1-2, Zhenmin Liu and Jun Yan offered resources, Jing Xie and Jun Yan served as supervision and reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yuan, J., Liu, Z., Xie, J. et al. The dual-species biofilm formed by Staphylococcus aureus and Pseudomonas fluorescens exhibited enhanced resistance to disinfectants. npj Sci Food 9, 220 (2025). https://doi.org/10.1038/s41538-025-00581-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41538-025-00581-x
