
Abstract
Chronic kidney disease (CKD) is characterized by progressive attenuation of kidney functions, resulting in the accumulation of uremic toxins. Various symptoms for CKD are closely associated with the gut-kidney axis, which represents the interaction between gut microbiota and CKD. In this study, we investigated the effects of Lactobacillus acidophilus (L. acidophilus) KBL409 on uremic toxin concentrations using a multi-center, randomized, double-blind, placebo-controlled study. Participants in the L. acidophilus KBL409 group received a daily capsule containing 1 × 1010 colony-forming units of L. acidophilus KBL409 or placebo. The per protocol analysis included 34 participants in the L. acidophilus KBL409 group and 30 participants in the placebo group. After 16 weeks, the serum indoxyl sulfate (IS) concentration was significantly lower in the L. acidophilus KBL409 group than in the placebo group (p < 0.05). Additionally, significant reductions in the genera Blautia, Butyricicoccus, Lachnospiraceae UCG-004, and Megamonas were observed in the L. acidophilus KBL409 group. These bacteria exhibited positive correlations with predicted functional genes linked to uremic toxin synthesis pathways, suggesting that L. acidophilus KBL409 reduced serum IS by altering gut microbial compositions. Therefore, L. acidophilus KBL409 could be used as an effective probiotic for improving kidney health through gut microbiota modulation.
Similar content being viewed by others

Introduction
Chronic kidney disease (CKD) is a global public health concern characterized by progressive deterioration of kidney functions1,2,3. The gradual decline in kidney function due to CKD leads to the accumulation of blood urea and uremic toxins, including indoxyl sulfate (IS), p-Cresyl sulfate (PCS), and trimethylamine N-oxide (TMAO), eventually resulting in uremia4,5,6,7,8. Serum IS concentrations are strongly correlated with estimated glomerular filtration rate (eGFR) and have been associated with the progression of CKD and cardiovascular disease (CVD)8,9.
Gut microbiota play a critical role in the complex network of human organs through microbial metabolites10,11. A dramatic shift in gut microbial composition can increase both the accumulation of uremic toxins in the blood and the risk of CKD12,13. The gut-kidney axis represents the interaction between CKD and gut microbiota or intestinal physiology, such as intestinal permeability14,15. IS, one of the major uremic toxins, is originated from the gut microbiota mediated metabolism of tryptophan, an essential amino acid derived from dietary protein16. Specific gut bacteria, including the genera Alistipes, Bacteroides, Blautia, Butyricicoccus, Escherichia, and Ruminococcus, have been reported to express tryptophanase, which can metabolize tryptophan into indole, the precursor of IS17,18,19. IS acts as an aryl hydrocarbon receptor agonist, inducing pro-inflammatory and pro-fibrotic effects through nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) or mitogen-activated protein kinase (MAPK) signaling pathways20,21,22.
Despite the lack of effective ways for controlling increased uremic toxin levels in the blood, recent studies have demonstrated significant reductions in uremic toxins, including IS and PCS, through alterations in gut microbiota using probiotics or prebiotics23,24,25. Notably, the genera Bifidobacterium and Lactobacillus, which are well-known probiotic microorganisms, exhibit differing abilities to reduce IS and PCS26,27. These differences highlight the need to investigate specific probiotic strains capable of effectively reducing uremic toxins for potential applications in CKD prevention.
Previously, we confirmed that Lactobacillus acidophilus (L. acidophilus) KBL409 has the potential to reduce serum levels of several uremic toxins, including IS and PCS, improve kidney function, attenuate inflammation and fibrosis, and enhance mitochondrial function in an adenine-induced CKD mouse model28,29,30. Therefore, in this study, we investigated the effects of L. acidophilus KBL409 on the improvement of serum IS concentrations through changes in gut microbial composition using a multi-center, randomized, double-blind, placebo-controlled study.
Results
Demographic characteristics
The per protocol set consisted of 34 participants in the L. acidophilus KBL409 group and 30 participants in the placebo group (Fig. 1). After randomization, 4 participants in the placebo group dropped out and 7 participants from each group were excluded from further analyses or withdrew from this study due to the reasons including the withdrawal of consent, the compliance score lower than 80%, the use of systemic antibiotics or the angiotensin II receptor blockers, the loss of capsules, and the adverse event. The mean ages of participants in the L. acidophilus KBL409 group and the placebo group were 55.4 ± 12.7 years and 56.4 ± 13.9 years, respectively (Table 1). The mean body mass index of the L. acidophilus KBL409 group was 26.4 ± 4.6 kg/m2, which did not show a significant difference from the placebo group (25.7 ± 3.6 kg/m2, p = 0.5260). The major lifestyle factors of both groups, including smoking, alcohol consumption, and exercise, showed no significant difference between the two groups.

Flow diagram of the participant screening process.
Dietary patterns
The L. acidophilus KBL409 group showed a significant increase in the mean protein intake in Visit 5 (16 weeks; mean change: 10.38 ± 25.10 g, p = 0.0237) (Table S1). However, participants the placebo group did not show the significant increases in the mean protein intake in Visit 5 (mean change: 3.64 ± 14.69 g). There were no significant differences in the mean dietary fiber intake of both groups in Visit 5 (Table S1). The L. acidophilus KBL409 group also showed significantly increases in the mean intake of lipids (mean change: 9.21 ± 23.68 g, p = 0.0326), moisture (mean change: 110.96 ± 233.02 g, p = 0.0101), and crude ash (mean change: 2.37 ± 6.29 g, p = 0.0378) in Visit 5 (Table S1).
Effects of L. acidophilus KBL409 on kidney function indicators and uremic toxin concentrations
After 16 weeks of L. acidophilus KBL409 administration, decreases in kidney function indicators, including urine protein (mean change: −28.42 ± 170.81 mg/dL), urine creatinine (mean change: −4.19 ± 82.55 mg/dL), and UPCR (mean change: –0.20 ± 1.20 g/g) were observed the L. acidophilus KBL409 group (Table 2). There were no significant differences in the mean eGFR of both groups after 16 weeks of administration in the per protocol set. However, in the full analysis set, a significantly lower reduction of the mean eGFR was observed in the L. acidophilus KBL409 group than the placebo group (data not shown). The mean serum IS concentration in the L. acidophilus KBL409 group was significantly reduced after 16 weeks compared to the placebo group (p = 0.0329). However, L. acidophilus KBL409 administration had no effects on serum PCS and TMAO concentrations.
Effects of L. acidophilus KBL409 on the gut microbiota
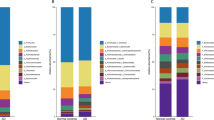
To investigate the effects of L. acidophilus KBL409 on the gut microbiota, the fecal microbiome of participants was analyzed (Fig. 2). Alpha diversities did not significantly differ between the L. acidophilus KBL409 group and the placebo group (Fig. 2A). Beta diversities of the fecal microbiome between the baseline (0 week) and after 16 weeks of administration were clustered similarly in both the L. acidophilus KBL409 group and the placebo group (Fig. 2B). After 16 weeks, the abundance of the genus Lactobacillus (p < 0.01) was significantly increased, whereas the abundances of the genera Blautia (p < 0.01), Megamonas (p < 0.05), Butyricicoccus (p < 0.01), and Lachnospiraceae UCG-004 (p < 0.05) were significantly decreased in the L. acidophilus KBL409 group (Fig. 2C). Moreover, after 16 weeks, the significant increase in the abundance of the genus Lactobacillus (p < 0.05) and significant decrease in the genera Blautia (p < 0.01) and Lachnospiraceae UCG-004 (p < 0.01) were discovered in the L. acidophilus KBL409 group compared to the placebo group. Additionally, in a subgroup of participants in the L. acidophilus KBL409 group with a mean eGFR <60 mL/min/1.73 m2, significant decreases were also observed in the abundances of the genera Butyricicoccus (p < 0.01) and Lachnospiraceae UCG-004 (p < 0.01) in the L. acidophilus KBL409 group compared to the placebo group (Fig. 2D).

A Alpha diversity indices. B Principal coordinates analysis plots based on the Bray–Curtis dissimilarity distances. C Relative abundances of specific gut bacteria in participants after 16 weeks of L. acidophilus KBL409 administration. D Relative abundances of specific gut bacteria in subgroup participants (the mean eGFR <60 mL/min/1.73 m2) after 16 weeks of L. acidophilus KBL409 administration. Asterisks indicate statistical significance in intra-group comparisons (*p < 0.05; **p < 0.01; a paired t-test). Hashtags indicate statistical significance in inter-group comparisons in changes from baseline to 16 weeks (#p < 0.05; ##p < 0.01; a two-sample t-test).
Effects of L. acidophilus KBL409 on profiles of functional gene alterations related to the gut microbiota
In the L. acidophilus KBL409 group, significant reductions were observed in the activities of predicted metabolic pathways for phenylalanine, shikimate, tryptophan, and tyrosine biosynthesis after 16 weeks of administration (Fig. 3). A significant decrease in the expression of tyrA2, which encodes prephenate dehydrogenase, also identified, indicating alterations in tryptophan metabolism pathways associated with gut microbial compositions.

a Phenylalanine metabolism. b Shikimate pathway. c Tryptophan metabolism/biosynthesis. d Tyrosine metabolism. Asterisks indicate statistical significance (*p < 0.05; **p < 0.01; a paired t-test).
Correlations between specific gut bacteria and predicted genes in the L. acidophilus KBL409 group
Table 3 presents the correlations between specific gut bacteria and predicted genes related to uremic toxin synthesis pathways in the L. acidophilus KBL409 group. The abundances of the genera Blautia, Butyricicoccus, Lachnospiraceae UCG-004, and Megamonas, which significantly decreased after 16 weeks of L. acidophilus KBL409 administration, were positively correlated with various predicted genes involved in the biosynthesis and degradation of aromatic amino acids. These findings suggest that L. acidophilus KBL409 downregulated uremic toxin-producing pathways by reducing the abundances of specific gut bacteria.
Safety
During the entire intervention period, a total of 11 adverse events were reported in the L. acidophilus KBL409 group, including 10 mild cases and 1 moderate case (cholecystitis acute) (Table S2). Similarly, 11 mild adverse events were reported in the placebo group. Only two gastrointestinal disorders, including constipation and gastrointestinal disorders, were identified as the adverse events probably or possibly related to L. acidophilus KBL409 administration, respectively. All adverse events were resolved during the intervention period. Blood and urine samples were collected from participants during Visit 1 and Visit 5 for safety assessments. All safety parameters did not show a significant difference between the L. acidophilus KBL409 group and the placebo group.
Discussion
The metabolism-dependent pathway of the gut-kidney axis is critical for the uremic toxin production31,32. Initially, dietary patterns high in proteins and animal fats and low in resistant starches and fibers induce dysbiosis of gut microbiota, resulting in excessive production and accumulation of uremic toxins, including IS and PCS. These uremic toxins disrupt intestinal barriers and promote kidney inflammations with endotoxins33. Our previous studies showed that L. acidophilus KBL409 significantly upregulated the expression of gut tight-junction markers, such as zonula occludens (ZO)-1 and claudin-1, increased the diversity of cecum microbiota, and reduced NLR family pyrin domain containing 3 inflammasomes and kidney fibrosis markers such as alpha-1 type I collagen and fibronectin in an adenine-induced CKD mouse model28,29,30. In the present study, L. acidophilus KBL409 administration effectively reduced serum IS concentrations, as well as other kidney function indicators, including urine protein and urine creatinine (Table 2). These findings suggest that L. acidophilus KBL409 can serve as a probiotic microorganism for improving of kidney health.
Notably, there were no significant changes in eGFR in the L. acidophilus KBL409 group, indicating that L. acidophilus KBL409 administration can reduce serum IS concentrations without altering kidney functions (Table 2). However, significant reductions of other uremic toxins, such as PCS and TMAO, were not observed during the intervention period. Previous studies have reported that different probiotic strains exhibit varying capabilities to reduce uremic toxin levels27. However, our results only evaluated the effects of L. acidophilus KBL409 over a 16-week period. Further longitudinal studies in humans, incorporating various doses are necessary to assess the potential renoprotective effects of L. acidophilus KBL409.
Figure 2C suggests that L. acidophilus KBL409 can affect the abundances of specific gut bacteria, including the genera Blautia, Butyricicoccus, Lachnospiraceae UCG-004, and Megamonas. Interestingly, participants with a mean eGFR <60 mL/min/1.73 m2, indicating moderate impairment of CKD34, also showed the decreases in the abundances of genera Blautia, Butyricicoccus, and Lachnospiraceae UCG-004 (Fig. 2D). These bacteria showed strong positive correlations with various predicted genes related to the biosynthesis and degradation of aromatic amino acids, including phenylalanine, tryptophan, and tyrosine (Table 3). Consequently, significant decreases in predicted metabolic pathways of aromatic amino acids were observed in the L. acidophilus KBL409 group after 16 weeks of administration (Fig. 3). Aromatic amino acids are key precursors for uremic toxins35. Changes in gut microbial composition due to L. acidophilus KBL409 administration could play crucial roles in reducing serum IS concentrations. However, to establish a clear causal relationship between L. acidophilus KBL409-mediated modulation of gut microbiota and the reduction of uremic toxins, comprehensive profiling of uremic toxin precursors in fecal, blood, and urine samples is necessary. Further studies with precise tracking of gut microbiota and metabolites related to uremic toxin production should be planned to fully elucidate the mechanisms of L. acidophilus KBL409 in preventing CKD via the gut-kidney axis.
Our findings are consistent with previous research emphasizing the important role of gut microbiota in the accumulation of uremic toxins8. However, the relatively small sample size of this study limited the statistical power to identify the effects of L. acidophilus KBL409 on major kidney function indicators, such as eGFR and UPCR. Additionally, the effects of L. acidophilus KBL409 on uremic toxins may have been underestimated because, in this study, only participants who have lower serum concentrations of uremic toxins were enrolled. Furthermore, the results of the Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt)2 analyses provided only functional predictions based on changes in gut microbiota. Further studies are necessary to evaluate the preventive or therapeutic potential and elucidate the specific mechanisms of L. acidophilus KBL409.
In conclusion, L. acidophilus KBL409 significantly reduced serum IS concentrations in study participants and altered the gut microbiota by decreasing the abundances of the genera Blautia, Butyricicoccus, Lachnospiraceae UCG-004, and Megamonas. These bacteria showed positive correlations with the activities of predicted functional genes related to uremic toxin synthesis pathways, indicating that L. acidophilus KBL409 effectively reduced serum IS within the gut-kidney axis. Especially, the effects of gut microbial alterations due to L. acidophilus KBL409 on serum IS concentrations were notable because the L. acidophilus KBL409 group showed the clear high protein-dietary pattern during this study. Our findings suggest that L. acidophilus KBL409 could be a safe and promising probiotic for improving kidney health. Further human studies involving patients with severe CKD, utilizing different dose- and time-dependent approaches, will be necessary to expand the applications of L. acidophilus KBL409 in the treatment of CKD.
Methods
Subjects
This study was conducted in accordance with Korean Good Clinical Practice (KGCP) Guidelines and the Declaration of Helsinki. All participants provided informed consent, and the study protocol was approved by the institutional review boards of Severance Hospital, Yonsei University Health System (No. 4-2019-0763), SMG-SNU Boramae Medical Center (No. 20-2019-59), and National Health Insurance Service Ilsan Hospital (No. NHIMC 2019-08-002-020).
A total of 92 individuals visited the study sites during the study period (November 2019 to November 2020) and 82 participants met the inclusion criteria (Fig. 1). The inclusion criteria for participants were: (1) aged 19 or older; (2) eGFR >30 mL/min/1.73 m2; (3) urine protein to creatinine ratio (UPCR) > 0.5 g/g; (4) voluntarily agreed to participate in this study with written informed consent provided. The major exclusion criteria were: (1) history of renal dialysis, kidney transplantation, or glomerulonephritis; (2) use of renal medications including Kremezin®, Renamezin®, and tolvaptan within 4 weeks prior to Visit 1; (3) use of immunosuppressants; (4) use of systemic antibiotics or probiotics within two weeks prior to Visit 1; (5) regular consumption of fermented milk (more than four times per week); (6) allergic reactions to functional food ingredients; (7) history of thyroid diseases; (8) levels of aspartate aminotransferase or alanine aminotransferase more than three times of the upper limit of normal (40 IU/L); (9) participation in another clinical study within one month or willingness to participation in another clinical study after this study; (10) pregnancy or breastfeeding; (11) any conditions deemed inappropriate by investigators.
Study design
This study is a multi-center, randomized, double-blind, placebo-controlled study to assess the efficacy of L. acidophilus KBL409 in reducing uremic toxin levels among study participants. A total of 82 participants were assigned randomly to either the L. acidophilus KBL409 or the placebo group at a 1:1 ratio using SAS v.9.4 (SAS Institute, Cary, NC, USA) (Fig. 4). All participants received either L. acidophilus KBL409 or placebo capsules, which were labeled according to the randomization table, at 4-week intervals. To maintain the integrity of the participant allocation process, both investigators and participants were blinded to the allocation codes until the study was completed.

Study design.
Participants in the L. acidophilus KBL409 group received a capsule containing 1 × 1010 colony-forming units of L. acidophilus KBL409 daily for 30 min before a meal. Placebo capsules, mainly composed of dextrin, were prepared to match the appearance, flavor, and color of L. acidophilus KBL409 capsules. All capsules were stored at 2–8 °C until use.
The study consisted of a screening visit (Visit 1) and four additional visits (Visit 2–5) during the 16-week intervention period (Fig. 4). During Visit 3–5, a 7-day visit window was allowed for participants. Follow-up visits were performed as necessary. Participants were advised to maintain appropriate lifestyles, including a low-protein diet, moderate exercise, controlled alcohol consumption, weight management, and smoking cessation. Demographic characteristics, medical histories, clinical characteristics, and lifestyle details of participants were documented at Visit 1. Additionally, eGFR and UPCR measurements were conducted to determine the enrollment of study participants. During Visit 2–5, dietary surveys by 24-h recall method were completed for eligible participants.
Blood and urine samples were collected from participants for further analyses of biomarkers, including eGFR, UPCR, uremic toxins, and cytokines. Briefly, 17.0 mL of blood was collected from each participant using BD Vacutainer SST II Advance Tubes (BD Biosciences, San Jose, CA, USA). The collected blood samples were inverted several times, incubated at room temperature for 30 min, and centrifuged at 3000 rpm for 10 min. The supernatant, which is the separated serum, was collected into cryotubes and stored at −70 °C until use. Additionally, 1.0 mL of midstream urine was collected from each participant into cryotubes and stored at −70 °C until use.
Fecal samples were self-collected by participants using DNA/RNA Shield-Fecal Collection Tubes (Zymo Research, Irvine, CA, USA) and immediately stored at −20 °C. During Visit 2 and Visit 5, frozen fecal samples were gathered and stored at −70 °C until use.
Analysis of eGFR and UPCR
To assess eGFR, and UPCR, serum and urine samples were transferred to SCL Healthcare (Yongin-si, Gyeonggi-do, Republic of Korea), the central laboratory for this study, using a cold-chain transport route. Subsequently, these samples were analyzed in accordance with the central laboratory’s standard procedures.
Analysis of uremic toxins
Standard stock solutions of IS (Sigma-Aldrich, St. Louis, MO, USA), PCS (MedChemExpress, Monmouth Junction, NJ, USA), and TMAO (Sigma-Aldrich) were prepared in 50% methanol (Sigma-Aldrich) and stored at −70 °C until use. All calibration standards and quality control samples were freshly prepared on the day of analysis.
To prepare samples for the analysis of uremic toxins, 30 μL of the internal standard solution, 300 μL of methanol, and 30 μL of serum were mixed and incubated at room temperature for 20 min. The mixture was centrifuged at 13,523 × g at 4 °C for 5 min, and 100 μL of the supernatant was collected in an autosampler vial. Subsequently, 5 μL of the supernatant were injected into the high-performance liquid chromatography (HPLC)-tandem mass spectrometry (MS/MS) system, which consisted of an Agilent 1200 Series Analytical LC System (Agilent Technologies, Waldbronn, Germany) and an API 5000 Triple Quadrupole Mass Spectrometer (AB SCIEX, Framingham, MA, USA). Separation was preformed using a reverse-phase Gemini C18 column (5 μm; 2.0 × 150 mm2). HPLC-grade water containing 0.1% formic acid (Merck, Darmstadt, Germany) and liquid chromatography-grade acetonitrile (Sigma-Aldrich) were used as mobile phase A and mobile phase B, respectively. Isocratic conditions with 60% mobile phase B were applied. HPLC-MS/MS analyses for target uremic toxins were performed in multiple reaction monitoring mode under the following conditions: 0.2 mL/min flow rate, 15 min total running time, 40 °C column over temperature, and 8 °C autosampler temperature.
Analysis of gut microbiota
The gut microbiota of participants was analyzed using fecal samples, as previously described with some modifications36. Initially, total bacterial DNA was extracted using a FastDNA Stool Mini Kit (Qiagen, Hilden, Germany) in accordance with the manufacturer’s instructions. The V3-V4 region of the 16S ribosomal RNA (16S rRNA)-coding gene was amplified using the following primers: 341 F (5′-CCTACGGGNGGCWGCAG-3′) and 805 R (5′- GACTACHVGGGTATCTAATCC-3′)37. An Illumina MiSeq platform (Illumina, San Diego, CA, USA) and a MiSeq Reagent Kit v. 3 (Illumina) were used for the sequencing process of barcoded amplicons. FastQC v. 0.11.9 (Bioinformatics Group of Babraham Institute; https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and Trimmomatic v. 0.38 (USEDEL lab; https://github.com/usadellab/Trimmomatic)38 were applied for the manipulations of raw reads, including the removal of primer sequences and quality control. Reads below a Q33 quality score were excluded from analyses. Quantitative Insights into Microbial Ecology (QIIME) 2 v. 2024.5 (QIIME 2 Development Team; https://qiime2.org)39, using the Divisive Amplicon Denoising Algorithm 2, was applied for demultiplexing, denoising, and chimera checking of reads. Subsequently, amplicon sequence variants (ASVs) were generated and classified using the SILVA v. 138.1 database (SILVA rRNA database project; https://www.arb-silva.de)40. ASVs with fewer than 500 reads or a minimum frequency <20% were excluded to reduce noise. Alpha diversity indices, including Faith’s phylogenetic diversity and Shannon diversity, were calculated using the q2-diversity function in QIIME 2 v. 2024.5. Principal coordinates analysis (PCoA) plots with the Bray–Curtis dissimilarity distances for beta diversities were also suggested using the phyloseq R package in Bioconductor v. 3.20 (The Bioconductor Project; https://www.bioconductor.org/packages/release/bioc/html/phyloseq.html)41. Profiles of functional gene alterations related to gut microbiota were analyzed using PICRUSt 2 v. 2.5.3 (The Huttenhower Lab; https://huttenhower.sph.harvard.edu/picrust/)42 with the Kyoto Encyclopedia of Genes and Genomes (KEGG) orthologous gene family database (Kanehisa Laboratories, Kyoto University; https://www.genome.jp/kegg/)43.
Participant size and statistical analysis
The appropriate participant size for was calculated based on the results of a previous study44. UPCR was used as the variable to calculate the number of participants. To achieve 80% power with an alpha error probability of 0.05 and a beta error probability of 0.2, a minimum of 32 participants were required in each group. The target enrollment was set at 40 participants per group to adjust for a potential 20% drop-out rate.
All statistical analyses for the per protocol set were performed using SAS v. 9.4 (SAS Institute). Categorical and continuous variables are presented as frequencies with proportions or as means ± standard deviations (SDs), respectively. The chi-square test or Fisher’s exact test were used for inter-group comparisons of categorical variables. For continuous variables, a paired t-test for intra-group comparisons or a two-sample t-test or the Wilcoxon rank-sum test for inter-group comparisons were performed, as appropriate. A P-value (p) <0.05 was considered statistically significant. The centered log-ratio transformation was applied to normalize gut microbiota abundances. Spearman correlation coefficients among specific gut bacteria and predicted genes related to uremic toxin synthesis pathways were calculated using the rcorr function in R v. 4.1.2 (R Core Team; https://www.r-project.org).
Data availability
All data supporting the findings of this study are available from the corresponding author upon reasonable request.
References
Krukowski, H. et al. Gut microbiome studies in CKD: opportunities, pitfalls and therapeutic potential. Nat. Rev. Nephrol. 19, 87–101 (2023).
Chesnaye, N. C., Ortiz, A., Zoccali, C., Stel, V. S. & Jager, K. J. The impact of population ageing on the burden of chronic kidney disease. Nat. Rev. Nephrol. 2, 569–585 (2024).
Francis, A. et al. Chronic kidney disease and the global public health agenda: an international consensus. Nat. Rev. Nephrol. 20, 473–485 (2024).
Zoccali, C. et al. The systemic nature of CKD. Nat. Rev. Nephrol. 13, 344–358 (2017).
Hung, S. C., Kuo, K. L., Wu, C. C. & Tarng, D. C. Indoxyl sulfate: a novel cardiovascular risk factor in chronic kidney disease. J. Am. Heart Assoc. 6, e005022 (2017).
Wang, M. et al. The gut microbial metabolite trimethylamine N-oxide, incident CKD, and kidney function decline. J. Am. Soc. Nephrol. 35, 749–760 (2024).
Rosner, M. H. et al. Classification of uremic toxins and their role in kidney failure. Clin. J. Am. Soc. Nephrol. 16, 1918–1928 (2021).
Gryp, T. et al. Gut microbiota generation of protein-bound uremic toxins and related metabolites is not altered at different stages of chronic kidney disease. Kidney Int. 97, 1230–1242 (2020).
Vanholder, R. et al. Metabolomics to identify unclassified uremic toxins: a comprehensive literature review. Kidney Med. 7, 100955 (2024).
Chakaroun, R. M., Olsson, L. M. & Bäckhed, F. The potential of tailoring the gut microbiome to prevent and treat cardiometabolic disease. Nat. Rev. Cardiol. 20, 217–235 (2023).
Koh, A. & Bäckhed, F. From association to causality: the role of the gut microbiota and its functional products on host metabolism. Mol. Cell 78, 584–596 (2020).
Fan, Y. & Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 19, 55–71 (2021).
Mirmiran, P., Yuzbashian, E., Asghari, G., Sarverzadeh, S. & Azizi, F. Dietary fibre intake in relation to the risk of incident chronic kidney disease. Br. J. Nutr. 119, 479–485 (2018).
Stavropoulou, E. et al. Focus on the gut-kidney axis in health and disease. Front. Med. 7, 620102 (2021).
Yang, T., Richards, E. M., Pepine, C. J. & Raizada, M. K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 14, 442–456 (2018).
Roager, H. M. & Licht, T. R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 9, 3294 (2018).
Graboski, A. L. et al. Mechanism-based inhibition of gut microbial tryptophanases reduces serum indoxyl sulfate. Cell. Chem. Biol. 30, 1402–1413 (2023).
Wang, X. et al. Aberrant gut microbiota alters host metabolome and impacts renal failure in humans and rodents. Gut 69, 2131–2142 (2020).
Devlin, A. S. et al. Modulation of a circulating uremic solute via rational genetic manipulation of the gut microbiota. Cell Host Microbe 20, 709–715 (2016).
Kim, H. Y. et al. Uremic toxin indoxyl sulfate induces trained immunity via the AhR-dependent arachidonic acid pathway in end-stage renal disease (ESRD). Elife 12, RP87316 (2024).
Barisione, C. et al. Indoxyl sulphate-initiated activation of cardiac fibroblasts is modulated by aryl hydrocarbon receptor and nuclear factor-erythroid-2-related factor 2. J. Cell. Mol. Med. 28, e18192 (2024).
Lekawanvijit, S. et al. Does indoxyl sulfate, a uraemic toxin, have direct effects on cardiac fibroblasts and myocytes? Eur. Heart. J. 31, 1771–1779 (2010).
Rossi, M. et al. Synbiotics easing renal failure by improving gut microbiology (SYNERGY): a randomized trial. Clin. J. Am. Soc. Nephrol. 11, 223–231 (2016).
Ranganathan, N. et al. Probiotic dietary supplementation in patients with stage 3 and 4 chronic kidney disease: a 6-month pilot scale trial in Canada. Curr. Med. Res. Opin. 25, 1919–1930 (2009).
Sohn, M. B. et al. Pilot studies in CKD consortium. targeting gut microbiome with prebiotic in patients with CKD: The TarGut-CKD study. Kidney Int. Rep. 9, 671–685 (2023).
Fang, C. Y. et al. Selection of uremic toxin-reducing probiotics in vitro and in vivo. J. Funct. Foods 7, 407–415 (2014).
Vacca, M. et al. In vitro selection of probiotics, prebiotics, and antioxidants to develop an innovative synbiotic (NatuREN G) and resting its effect in reducing uremic toxins in fecal batches from CKD patients. Microorganisms 9, 1316 (2021).
Kim, H. et al. Lactobacillus acidophilus KBL409 reduces kidney fibrosis via immune modulatory effects in mice with chronic kidney disease. Mol. Nutr. Food Res. 66, e2101105 (2021).
Kim, M. J. et al. Effects of Lactobacillus acidophilus KBL409 administration on uremic toxins, Korean. J. Microbiol. 60, 161–168 (2024).
Park, J. et al. Lactobacillus acidophilus KBL409 protects against kidney injury via improving mitochondrial function with chronic kidney disease. Eur. J. Nutr. 63, 2121–2135 (2024).
Glorieux, G., Nigam, S. K., Vanholder, R. & Verbeke, F. Role of the microbiome in gut-heart-kidney cross talk. Circul. Res. 132, 1064–1083 (2023).
Evenepoel, P., Meijers, B. K., Bammens, B. R. & Verbeke, K. Uremic toxins originating from colonic microbial metabolism. Kidney Int. Suppl. 114, S12–S19 (2009).
Lau, W. L., Kalantar-Zadeh, K. & Vaziri, N. D. The gut as a source of inflammation in chronic kidney disease. Nephron 130, 92–98 (2015).
Levin, A. & Stevens, P. E. Summary of KDIGO 2012 CKD Guideline: behind the scenes, need for guidance, and a framework for moving forward. Kidney Int. 85, 49–61 (2014).
Barba, C. et al. A low aromatic amino-acid diet improves renal function and prevent kidney fibrosis in mice with chronic kidney disease. Sci. Rep. 11, 19184 (2021).
Yoon, H. S. et al. Akkermansia muciniphila secretes a glucagon-like peptide-1-inducing protein that improves glucose homeostasis and ameliorates metabolic disease in mice. Nat. Microbiol. 6, 563–573 (2021).
Singer, E. et al. High-resolution phylogenetic microbial community profiling. ISME J. 10, 2020–2032 (2016).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Bolyen, E. et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857 (2019).
Quast, C. et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596 (2013).
McMurdie, P. J. & Holmes, S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS ONE 8, e61217 (2013).
Douglas, G. M. et al. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688 (2020).
Ogata, H. et al. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 27, 29–34 (1999).
Oblak, M., Mlinšek, G., Kandus, A., Buturović-Ponikvar, J. & Arnol, M. Paricalcitol versus placebo for reduction of proteinuria in kidney transplant recipients: a double-blind, randomized controlled trial. Transpl. Int. 31, 1391–1404 (2018).
Acknowledgements
This research was supported by the Technology Innovation Program (No. 10076742) funded by the Ministry of Trade, Industry & Energy (MOTIE), Republic of Korea and the Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT) (NRF-2022M3A9F3017371). The authors thank Prof. Seung Hyeok Han from the Department of Internal Medicine, Yonsei University, Prof. Jung Pyo Lee from the Department of Internal Medicine, SMG-SNU Boramae Medical Center, and Prof. Kyoung Sook Park from the Department of Internal Medicine, National Health Insurance Service Ilsan Hospital for their contributions to the successful completion of this investigation.
Author information
Authors and Affiliations
Contributions
S.J.J.: Conceptualization, formal analysis, investigation, methodology, visualization, writing—original draft. S.P.: Conceptualization, investigation, methodology, supervision, writing—original draft, writing—review and editing. K.L.: Formal analysis, investigation, methodology. W.-K.K.: Investigation, methodology. S.H.M.: Investigation, methodology. B.-R.C.: Investigation, methodology. C.L.: Methodology, Writing—original draft, writing – review & editing. H.L.: Conceptualization, funding acquisition. T.-W.N.: Conceptualization, investigation, funding acquisition, project administration. G.K.: Conceptualization, funding acquisition, resources, supervision.
Corresponding authors
Ethics declarations
Competing interests
G.K. is the chief executive officer of KoBioLabs, Inc. K.L., S.H.M., and B-R.C. are employees of KoBioLabs, Inc. H.L. is the chief executive officer of weBiom Inc. S.J.J. and S.P. are employees of weBiom Inc. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Jang, S.J., Park, S., Lee, K. et al. Lactobacillus acidophilus KBL409 improves serum indoxyl sulfate via gut microbial changes in a human study. npj Sci Food 10, 108 (2026). https://doi.org/10.1038/s41538-026-00755-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41538-026-00755-1
