
Abstract
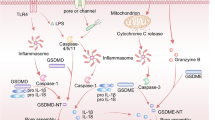
Whether pyroptosis is controllable and reversible remains an enigma. Here we revealed that autophagy could eliminate the pore-formed N terminus of GSDME (GSDME-NT) located on membranes at different locations, suppressing pyroptosis. Crucially, GSDME-NT on the plasma membrane was eliminated through endocytic internalization, where GSDME-NT-laden vesicles were targeted and degraded as intact units. Specifically, GSDME-NT pores on the plasma membrane induced endocytosis, generating endocytosed but leaky vesicles carrying GSDME-NT. Leakage prevented acidification, necessitating further degradation through autophagy. Upon endocytosis, GSDME-NT on the vesicle membrane was labelled with ubiquitin by calcium-activated E3 ligase NEDD4L. These labelled vesicles were recognized by TOLLIP, guiding subsequent autophagosome formation, and enabling further acidification, fusion with lysosomes and eventual GSDME-NT degradation. Furthermore, in several tumour models, either disturbing autophagy or interfering with the recognition of GSDME-NT vesicles by targeting TOLLIP increased tumour cell pyroptosis, activating antitumour immunity and promoting chemotherapeutic efficacy.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
All data generated and analysed in this study are included in the article, Supplementary Information or source data files. The single-cell RNA sequencing datasets from the NPC cohort were obtained from a published study (https://doi.org/10.1038/s41591-023-02369-6)56. The mass spectrometry proteomics data have been deposited to the iProX partner repository under dataset identifier PXD073218. Other key raw data have been deposited on the Research Data Deposit public platform (https://www.researchdata.org.cn/) under approval no. RDDB2026269282. All other data supporting the findings of this study are available from the corresponding author on reasonable request. Source data are provided with this paper.
Code availability
No custom code was developed or used in this study. All analyses were performed using publicly available tools, including R (v.4.4.1), Seurat (v.4.4.0), ggplot2 (v.3.5.0) and ggpubr (v.0.6.0).
References
Ding, J. et al. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 535, 111–116 (2016).
Broz, P., Pelegrín, P. & Shao, F. The gasdermins, a protein family executing cell death and inflammation. Nat. Rev. Immunol. 20, 143–157 (2020).
Shi, J. et al. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 526, 660–665 (2015).
Kayagaki, N. et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 526, 666–671 (2015).
Xia, S. et al. Gasdermin D pore structure reveals preferential release of mature interleukin-1. Nature 593, 607–611 (2021).
Liu, X. et al. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 535, 153–158 (2016).
Deng, W. et al. Streptococcal pyrogenic exotoxin B cleaves GSDMA and triggers pyroptosis. Nature 602, 496–502 (2022).
Zhong, X. et al. Structural mechanisms for regulation of GSDMB pore-forming activity. Nature 616, 598–605 (2023).
Hou, J. et al. PD-L1-mediated gasdermin C expression switches apoptosis to pyroptosis in cancer cells and facilitates tumour necrosis. Nat. Cell Biol. 22, 1264–1275 (2020).
Wang, Y. et al. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 547, 99–103 (2017).
Zhou, B. et al. Full-length GSDME mediates pyroptosis independent from cleavage. Nat. Cell Biol. 26, 1545–1557 (2024).
De Schutter, E. et al. Punching holes in cellular membranes: biology and evolution of gasdermins. Trends Cell Biol. 31, 500–513 (2021).
Zhang, Z. et al. Gasdermin E suppresses tumour growth by activating anti-tumour immunity. Nature 579, 415–420 (2020).
Erkes, D. A. et al. Mutant BRAF and MEK inhibitors regulate the tumor immune microenvironment via pyroptosis. Cancer Discov. 10, 254–269 (2020).
Wu, J. et al. Methionine metabolite spermidine inhibits tumor pyroptosis by enhancing MYO6-mediated endocytosis. Nat. Commun. 16, 2184 (2025).
Miao, R. et al. Gasdermin D permeabilization of mitochondrial inner and outer membranes accelerates and enhances pyroptosis. Immunity 56, 2523–2541.e2528 (2023).
Rogers, C. et al. Gasdermin pores permeabilize mitochondria to augment caspase-3 activation during apoptosis and inflammasome activation. Nat. Commun. 10, 1689 (2019).
Qu, Y. et al. Gasdermin D mediates endoplasmic reticulum stress via FAM134B to regulate cardiomyocyte autophagy and apoptosis in doxorubicin-induced cardiotoxicity. Cell Death Dis. 13, 901 (2022).
Karmakar, M. et al. N-GSDMD trafficking to neutrophil organelles facilitates IL-1β release independently of plasma membrane pores and pyroptosis. Nat. Commun. 11, 2212 (2020).
Gatica, D., Lahiri, V. & Klionsky, D. J. Cargo recognition and degradation by selective autophagy. Nat. Cell Biol. 20, 233–242 (2018).
Mizushima, N. A brief history of autophagy from cell biology to physiology and disease. Nat. Cell Biol. 20, 521–527 (2018).
Chen, Y. et al. Metabolic regulation of homologous recombination repair by MRE11 lactylation. Cell 187, 294–311.e221 (2024).
Ai, Y. L. et al. Mannose antagonizes GSDME-mediated pyroptosis through AMPK activated by metabolite GlcNAc-6P. Cell Res. 33, 904–922 (2023).
Du, G. et al. ROS-dependent S-palmitoylation activates cleaved and intact gasdermin D. Nature 630, 437–446 (2024).
Rogers, C. et al. Cleavage of DFNA5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat. Commun. 8, 14128 (2017).
Suda, K. et al. Plasma membrane damage limits replicative lifespan in yeast and induces premature senescence in human fibroblasts. Nat. Aging 4, 319–335 (2024).
Thurston, T. L., Wandel, M. P., von Muhlinen, N., Foeglein, A. & Randow, F. Galectin 8 targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature 482, 414–418 (2012).
Lazarou, M. et al. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 524, 309–314 (2015).
Li, C. M. et al. Cytosolic FKBPL and ER-resident CKAP4 co-regulates ER-phagy and protein secretion. Nat. Commun. 15, 7886 (2024).
De Belly, H. et al. Membrane tension gates ERK-mediated regulation of pluripotent cell fate. Cell Stem Cell 28, 273–284.e276 (2021).
Idone, V. et al. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. J. Cell Biol. 180, 905–914 (2008).
Thiery, J. et al. Perforin activates clathrin- and dynamin-dependent endocytosis, which is required for plasma membrane repair and delivery of granzyme B for granzyme-mediated apoptosis. Blood 115, 1582–1593 (2010).
Cullen, P. J. & Steinberg, F. To degrade or not to degrade: mechanisms and significance of endocytic recycling. Nat. Rev. Mol. Cell Biol. 19, 679–696 (2018).
Uemura, T., Suzuki, T., Dohmae, N. & Waguri, S. Clathrin adapters AP-1 and GGA2 support expression of epidermal growth factor receptor for cell growth. Oncogenesis 10, 80 (2021).
Schindler, C., Chen, Y., Pu, J., Guo, X. & Bonifacino, J. S. EARP is a multisubunit tethering complex involved in endocytic recycling. Nat. Cell Biol. 17, 639–650 (2015).
Chauhan, S. et al. TRIMs and galectins globally cooperate and TRIM16 and galectin-3 co-direct autophagy in endomembrane damage homeostasis. Dev. Cell 39, 13–27 (2016).
Novellasdemunt, L. et al. NEDD4 and NEDD4L regulate Wnt signalling and intestinal stem cell priming by degrading LGR5 receptor. EMBO J. 39, e102771 (2020).
Wiesner, S. et al. Autoinhibition of the HECT-type ubiquitin ligase Smurf2 through its C2 domain. Cell 130, 651–662 (2007).
Escobedo, A. et al. Structural basis of the activation and degradation mechanisms of the E3 ubiquitin ligase Nedd4L. Structure 22, 1446–1457 (2014).
Li, Z. L. et al. Autophagy deficiency promotes triple-negative breast cancer resistance to T cell-mediated cytotoxicity by blocking tenascin-C degradation. Nat. Commun. 11, 3806 (2020).
Jin, S. et al. Suppression of ACE2 SUMOylation protects against SARS-CoV-2 infection through TOLLIP-mediated selective autophagy. Nat. Commun. 13, 5204 (2022).
Hayashi, Y. et al. TOLLIP acts as a cargo adaptor to promote lysosomal degradation of aberrant ER membrane proteins. EMBO J. 42, e114272 (2023).
Zhou, Z. et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells. Science 368, eaaz7548 (2020).
Liu, Y. et al. Gasdermin E-mediated target cell pyroptosis by CAR T cells triggers cytokine release syndrome. Sci. Immunol. 5, eaax7969 (2020).
Tian, W. et al. An antibody for analysis of autophagy induction. Nat. Methods 17, 232–239 (2020).
Yu, T. et al. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell 170, 548–563.e516 (2017).
Wang, X. et al. Oncogenic RAS induces a distinctive form of non-canonical autophagy mediated by the P38-ULK1-PI4KB axis. Cell Res. https://doi.org/10.1038/s41422-025-01085-9 (2025).
Yim, W. W., Yamamoto, H. & Mizushima, N. A pulse-chasable reporter processing assay for mammalian autophagic flux with HaloTag. eLife 11, e78923 (2022).
Karabiyik, C., Vicinanza, M., Son, S. M. & Rubinsztein, D. C. Glucose starvation induces autophagy via ULK1-mediated activation of PIKfyve in an AMPK-dependent manner. Dev. Cell 56, 1961–1975.e1965 (2021).
Karlstaedt, A., Khanna, R., Thangam, M. & Taegtmeyer, H. Glucose 6-phosphate accumulates via phosphoglucose isomerase inhibition in heart muscle. Circ. Res. 126, 60–74 (2020).
Zhang, C. S. et al. Fructose-1,6-bisphosphate and aldolase mediate glucose sensing by AMPK. Nature 548, 112–116 (2017).
González, A., Hall, M. N., Lin, S. C. & Hardie, D. G. AMPK and TOR: The yin and yang of cellular nutrient sensing and growth control. Cell Metab. 31, 472–492 (2020).
Kim, J., Kundu, M., Viollet, B. & Guan, K. L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 13, 132–141 (2011).
Egan, D. F. et al. Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science 331, 456–461 (2011).
Yamamoto, K. et al. Autophagy promotes immune evasion of pancreatic cancer by degrading MHC-I. Nature 581, 100–105 (2020).
Lv, J. et al. The tumor immune microenvironment of nasopharyngeal carcinoma after gemcitabine plus cisplatin treatment. Nat. Med. 29, 1424–1436 (2023).
Charoentong, P. et al. Pan-cancer Immunogenomic analyses reveal genotype-immunophenotype relationships and predictors of response to checkpoint blockade. Cell Rep. 18, 248–262 (2017).
Shi, X. et al. Identification and validation of an autophagy-related gene signature for predicting prognosis in patients with esophageal squamous cell carcinoma. Sci Rep. 12, 1960 (2022).
Luo, M. et al. Multi-omics characterization of autophagy-related molecular features for therapeutic targeting of autophagy. Nat. Commun. 13, 6345 (2022).
Hansen, J. M. et al. Pathogenic ubiquitination of GSDMB inhibits NK cell bactericidal functions. Cell 184, 3178–3191.e3118 (2021).
Ren, Y. et al. USP48 stabilizes gasdermin E to promote pyroptosis in cancer. Cancer Res. 83, 1074–1093 (2023).
Thiery, J. et al. Perforin pores in the endosomal membrane trigger the release of endocytosed granzyme B into the cytosol of target cells. Nat. Immunol. 12, 770–777 (2011).
Wu, X. et al. MLKL-dependent signaling regulates autophagic flux in a murine model of non-alcohol-associated fatty liver and steatohepatitis. J. Hepatol. 73, 616–627 (2020).
Liu, S. et al. MLKL polymerization-induced lysosomal membrane permeabilization promotes necroptosis. Cell Death Differ. 31, 40–52 (2024).
Poillet-Perez, L. et al. Autophagy promotes growth of tumors with high mutational burden by inhibiting a T-cell immune response. Nat. Cancer 1, 923–934 (2020).
Zhang, G., Chen, Y., Huang, X. & Liang, T. Cancer immunotherapeutic challenges from autophagy-immune checkpoint reciprocal regulation. Trends Cancer 11, 169–184 (2025).
Vodnala, S. K. et al. T cell stemness and dysfunction in tumors are triggered by a common mechanism. Science 363, eaau0135 (2019).
Ma, Z. et al. Interferon-dependent SLC14A1(+) cancer-associated fibroblasts promote cancer stemness via WNT5A in bladder cancer. Cancer Cell 40, 1550–1565.e1557 (2022).
Li, Z. et al. Tumor-repopulating cells evade ferroptosis via PCK2-dependent phospholipid remodeling. Nat. Chem. Biol. https://doi.org/10.1038/s41589-024-01612-6 (2024).
Zhang, C. et al. HO-1 impairs the efficacy of radiotherapy by redistributing cGAS and STING in tumors. J. Clin. Invest. 134, e181044 (2024).
Ma, Z. et al. AhR diminishes the efficacy of chemotherapy via suppressing STING dependent type-I interferon in bladder cancer. Nat. Commun. 14, 5415 (2023).
Acknowledgements
We thank W.-Y. Sun (Guangdong Engineering Research Center of Chinese Medicine & Disease Susceptibility, State Key Laboratory of Bioactive Molecules and Druggability Assessment, Jinan University), L. Ge (Tsinghua University), L. Wang and T.-M. Li (Institute of Biophysics, Chinese Academy of Sciences) for technical assistance. This research was supported by the National Natural Science Foundation of China (82272883) to L.T.; the National Natural Science Foundation of China (82472822), Young Talents Program of SYSUCC (YTP-SYSUCC-0072) to X.L.; Noncommunicable Chronic Diseases-National Science and Technology Major Project (2024ZD0520700), National Natural Science Foundation of China (82430085), Cancer Innovative Research Program of SYSUCC (CIRP-SYSUCC-0005), Overseas Expertise Introduction Project for Discipline Innovation (111 Project, B14035) to J.M.; and the National Natural Science Foundation of China (82504101) to Z.L. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
X.L., Z.X., J.M. and L.T. conceived the experiments. Z.X., Z.L. and Z.D. carried out and analysed the data for most of the in vitro experiments. Z.X., Z.L., C.D. and L.L. collected NPC samples and performed the IHC experiments. Z.X., Z.D., J.W. and C.Z. designed and performed the animal experiments. Z.X., Z.L., H.W. and T.H. designed and performed LC–MS/MS analysis. X.L. and Z.X. wrote and revised the paper. J.M., X.L. and L.T. supervised the study. All authors reviewed and discussed the final version of the paper.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Cell Biology thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Autophagy regulation of GSDME-NT and pyroptosis.
(a) GSDME-knockout cells were transfected with GSDME-NT-HATet-On viruses to create cells that express GSDME-NT-HA upon Dox treatment, triggering pyroptosis. (b) Immunoblot analysis of GSDME-knockout GSDME-NT-HATet-On cells treated with Dox, using GSDME-FL-HATet-On as a control. (c) Phase-contrast images of pyroptotic cell death morphology. Scale bar: 50 μm. (d) Time-course analysis of 7-AAD staining, LDH activity and secreted HMGB1 levels in cells treated with Dox (0.5 μg/mL). (e-f) Secreted HMGB1 and of IL-6 levels in HK1 GSDME-NT-HATet-On cells pretreated with indicated inhibitors (e) or inducers (f) and treated with Dox. (g-i) Secreted HMGB1 and of IL-6 levels in HK1 GSDME-NT-HATet-On cells with ATG5 (g), BECN1 (h), or ATG4B (i) knockout and Dox treatment. (j) Immunoblot analysis of GSDME-knockout HK1 cells overexpressing GSDME-FL treated with raptinal for 3 h. (k)Immunoblots of GSDME expression in decitabine-treated HK1 cells, and of LC3 and p62 in HK1 cells pretreated with SAR405 or MRT68921. (l-u) Decitabine-treated HK1 cells were exposed to raptinal (l-o) or cisplatin (p-u) with pharmacological or genetic modulation of autophagy. Cells pretreated with SAR405 or MRT68921 (l, m, p, s), Torin-1 or rapamycin (r, u), or subjected to ATG5 knockout (n, o, q, t) were analysed by immunoblotting, with HMGB1, 7-AAD, LDH, and IL-6 levels assessed, as indicated. Data are shown as Mean ± SD. Data are representative of at least three independent experiments (b-u). One-way ANOVA (e-i, m-u) and two-way ANOVA (d) were used for analysis.
Extended Data Fig. 2 Autophagy regulation of N-terminal fragments of GSDME and other gasdermins.
(a-d) SH-SY5Y (a, b) and NCI-H226 (c, d) cells with pharmacological (SAR405 or MRT68921) (a, c) or genetic (ATG5 knockout) inhibition (b, d) of autophagy were treated with raptinal and analysed by immunoblotting and HMGB1, 7-AAD, LDH, IL-6 and HMGB1 assays. Inhibition of autophagy were also verified by immunoblotting. (e) Immunoblot analysis of GSDME expression in HK1 GSDME-NT-HATet-On cells treated with increasing Dox doses. (f) Immunoblot analysis of decitabine-treated HK1 cells pretreated with SAR405/MRT68921 under raptinal treatment. (g) Immunoblot analysis of GSDME-knockout HK1 cells pretreated with SAR405/MRT68921 under raptinal treatment. (h) Immunoblot analysis of GSDME-knockout HK1 cells pretreated with SAR405/MRT68921 and under cisplatin treatment. (i) Interaction analysis of GSDME with CASP3C/Ap17-HA/p12 in cells treated with SAR405/MRT68921. (j-l) Immunoblots, secreted HMGB1, 7-AAD, and LDH analysis in HK1 GSDMB-NT-HATet-On (j), GSDMD-NT-HATet-On (k), GSDMD-FL-V41A-HATet-On (l) cells treated with Dox (0.5 μg/mL), pretreated with SAR405 or MRT68921. (m) Immunoblots, cell death via LDH and IL-1β release of THP-1 cells pretreated with SAR405/MRT68921 under LPS + Nigericin. (n) Immunoblots, cell death via LDH and IL-1β release of THP-1 cells pre-transfected with ATG5 siRNA under LPS + Nigericin. (o) Inhibition of autophagy and knockdown efficiency were verified by immunoblotting. Data are shown as Mean ± SD. Data are representative of at least three independent experiments (a-o). One-way ANOVA (a-d, j-n) was used for analysis.
Extended Data Fig. 3 Endocytosis-autophagy axis mediates regulation of GSDME-NT upon membrane damage.
(a) GSDME-knockout cells were transduced with Tet-On viruses expressing GSDME-NT-HA mutants and induced with Dox. (b) Immunoblots of GSDME-knockout HK1 cells and their corresponding GSDME-NT-T6E/F2A + K40A-HATet-On derivatives after Dox treatment. (c-d) Representative immunofluorescence images (c) and 7-AAD/LDH analysis (d) of Dox-induced cells. Scale bar: 10 μm; Dox, 0.5 μg/mL. (e) Immunoblots of cells overexpressing GSDME-NT-T6E/F2A + K40A-HA transfected with GSDME-NT-FLAGTet-On and treated with Dox. (f-h) MFI of pHrodo dextran in DNM1 knockdown cells to assess endocytosis inhibition (f), immunoblots validating ER-phagy inhibition (FAM134B knockdown, g), and FACS analysis validating mitophagy inhibition (PINK1 knockdown, h). (i-j) MFI of pHrodo dextran showing inhibition of endocytosis by Dyngo-4a (i). Immunoblots and 7-AAD/LDH analysis of cells treated with Dox with or without Dyngo-4a (j). (k) Knockdown efficiency. (l) Knockout efficiency. (m-p) Immunofluorescence images and colocalization of GSDME-NT with EEA1 in DNM1 knockdown (m, o) or Dyngo-4a-treated (n, p) HK1 cells. Scale bar: 10 μm; n = 30 cells. Plasma membrane-to-total cellular fluorescence ratio was calculated (m, n). Z-stack images show GSDME-NT and EEA1 distribution (o, p). Z-interval: 2 μm. (q-r) Z-stack images of GSDME-NT with EEA1 (q) or LAMP1 (r) in SAR405-treated HK1 cells. Z-interval: 2 μm. (s) 7-AAD/LDH analyses of cells with RAB4A or RAB11A knockdown, treated with Dox. (t) 7-AAD staining of ATG5 knockout cells transfected with RAB4A siRNA and treated with Dox. (u) Immunofluorescence images and colocalization (n = 30 cells) of GSDME-NT with RAB4A. Scale bar: 10 μm. Data are shown as Mean ± SD. Data are representative of at least three independent experiments (b-u). One-way ANOVA (d, f, h, k, s, t) or unpaired two-tailed t-test (i, j, m, n, u) were used for analysis.
Extended Data Fig. 4 Ubiquitination of endocytosed GSDME-NT at K161 by NEDD4L regulates pyroptosis.
(a-d) qRT–PCR analysis of indicated genes (a). Immunoblots (b) and 7-AAD staining (c) of siRNA-transfected cells treated with Dox. qRT–PCR analysis of knockdown efficiency (d). (e) Mass spectrometry analysis of ubiquitination sites on GSDME-NT in HK1, and shared sites in HK1 and HONE1 cells. (f) Sequence alignment of GSDME-NT across species using the ClustalW2 algorithm and presented with ESPript 3.0 (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi), highlighting conserved lysines and the caspase-3 cleavage motif. (g) Three-dimensional structure of GSDME-NT, highlighting K83 and K161 positions. (h) Immunoprecipitation of GSDME-FL-HA from Dox-treated GSDME-FL-HA WT/K161RTet–On cells, followed by ubiquitin immunoblot. (i) Immunoblots of GSDME-FL-HA and grayscale analysis in Dox-treated GSDME-FL-K161R-HATet-On cells treated with CHX. (j-m) Dox-treated cells transfected with the indicated siRNAs were subjected to 7-AAD staining (j), immunoblot analysis (l), and HA immunoprecipitation followed by ubiquitin immunoblotting (m). Corresponding knockdown efficiencies are presented in (k). (n) Knockout efficiency of NEDD4L as assessed by immunoblotting. (o) Ubiquitination of immunoprecipitated GSDME-NT-HA from Dox-treated GSDME-NT-HA WT/K161RTet-On cells transfected with Flag–NEDD4L. (p) Immunoblots of cells overexpressing NEDD4L. Data are shown as Mean ± SD. Data are representative of at least three independent experiments (a-e, h-p). One-way ANOVA (c, d, j, k) or two-way ANOVA (i) were used for analysis.
Extended Data Fig. 5 TOLLIP mediated autophagic degradation of GSDME-NT.
(a-c) 7-AAD staining (a), LDH analysis (b) and immunoblots (c) of cells transfected with indicated siRNAs and treated with Dox. (d) Knockdown efficiency of siRNAs. (e-f) Z-stack images of GSDME-NT with LC3 (e) or LAMP1 (f) in TOLLIP-knockout HK1 cells (Z interval: 2 μm). (g) Immunoblots of ATG5/BECN1-knockout HK1 cells transfected with GSDME-NT-HA and Flag–TOLLIP. (h) Immunoblots of ATG5/BECN1-knockout HK1 cells. (i) Knockout efficiency of TOLLIP. (j) GSDME-FL protein stability in Dox-treated TOLLIP-knockout GSDME-FL-HATet-On cells treated with CHX. (k-l) Immunoblots (k), 7-AAD staining, LDH activity, secreted IL-6 and HMGB1 levels (l) in TOLLIP-knockout HK1 cells overexpressing GSDME-FL treated with raptinal. Knockout efficiency of TOLLIP was also shown (k). (m) Secreted IL-6 and HMGB1 levels in TOLLIP knockout decitabine-treated HK1 cells treated with raptinal. And knockout efficiency of TOLLIP. (n-o) Immunoblots of TOLLIP-overexpressing, decitabine-treated HK1 cells with or without ATG5 knockout (n), and corresponding secreted IL-6 and HMGB1 levels following raptinal treatment (o). (p) Secreted IL-6 and HMGB1 levels in TOLLIP knockout HK1 cells pretreated with decitabine and treated with cisplatin. Data are shown as Mean ± SD. Data are representative of at least three independent experiments (a-p). One-way ANOVA (a, b, d, l, m, o, p) or two-way ANOVA (j) were used for analysis.
Extended Data Fig. 6 TOLLIP regulate N-terminal fragments of GSDME and other gasdermins.
(a-d) Immunoblots and analysis of 7-AAD, LDH, HMGB1, and IL-6 in TOLLIP-knockout or TOLLIP-overexpressing cells with/without ATG5 knockout, under raptinal treatment. Knockout efficiency was shown for TOLLIP (a, c), and immunoblots of cells with ATG5 knockout and TOLLIP overexpression (b, d). Experiments were performed in SH-SY5Y (a, b) and NCI-H226 (c, d) cells. (e) NEDD4L-knockout cells transfected with Flag–TOLLIP, analysed for GSDME-NT-TOLLIP interaction by immunoprecipitation. (f) HK1 GSDME-NT-HA WT/K161RTet-On cells were transfected with Flag–TOLLIP and analysed for GSDME-NT-TOLLIP interaction. (g-h) HEK 293 T cells were transfected with Flag–TOLLIP and GSDMB-NT-HA (g) or GSDMD-NT-HA (h), and interactions were analysed by immunoprecipitation. (i-j) Immunoblots, 7-AAD staining and LDH activity of HK1 GSDMB-NT-HATet-On (i) or GSDMD-NT-HATet-On (j) cells with TOLLIP knockdown and treated with Dox. Knockdown efficiency of TOLLIP were also shown. (k-m) Immunoblots of TOLLIP knockdown efficiency in THP-1 cells (k), and analysis of cell death and IL-1β release in THP-1 cells under LPS + Nigericin exposure (l, m). Data are shown as Mean ± SD. Data are representative of at least three independent experiments (a-m). One-way ANOVA (a-d, i, j, m) was used for analysis.
Extended Data Fig. 7 Peptide CUE3 enhanced pyroptosis.
(a) Immunofluorescence images and the localization ratio (n = 30 cells) of LC3 with TOLLIP in cells treated with Dox and Peptide CUE3. Scale bar: 10 μm. (b) Secreted IL-6 and HMGB1 levels in HK1 GSDME-FL-HA cells treated with raptinal and Peptide CUE3. (c-d) Immunoblots (c), HMGB1 release, 7-AAD staining, LDH activity and IL-6 secretion (d) analysis of HK1 cells pretreated with decitabine, under cisplatin with or without Peptide CUE3 exposure. (e) Flow cytometry analysis of HER2 surface expression in HONE1, HK1 and MDA-MB-361 cells. (f-h) Immunoblots (f), HMGB1 release (g), 7-AAD staining, LDH activity and IL-6 secretion (h) in HK1 cells pretreated with decitabine, exposed to CAR-T cells, with or without Peptide CUE3. Data are shown as Mean ± SD. Data are representative of at least three independent experiments (a-h). Two-tailed unpaired Student’s t test (a, b, d, h) was used for analysis.
Extended Data Fig. 8 Crosstalk of autophagy and GSDME-NT mediated pyroptosis.
(a) Representative immunofluorescence images of mRFP-GFP-LC3B-transfected cells treated with Dox. Scale bar: 10 μm. (b) Representative TEM images showing the diaminobenzidine staining pattern in APEX2-LC3B-transfected cells. (c) Schematic of the processing assay using Halo-LC3 as an autophagy reporter. Halo-LC3 is completely degraded in autolysosomes. Upon labelling with a TMR-conjugated ligand, LC3 is degraded in autolysosomes, releasing ligand-bound Halo fragments (Halo-ligand) that are resistant to further degradation. (d) Immunoblots of HK1 GSDME-NT-T6E-HATet-On and GSDME-NT-F2A + K40A-HATet-On cells treated with Dox for 6 h, with or without CQ. (e) Immunoblots showing the expression of AMPKα in AMPK DKO cells. (f) Immunoblots of HK1 GSDME-NT-HATet-On cells treated with Dox for 6 h, with or without MK-8722. (g) Immunoblots of lysates of HONE1 and HK1 cells overexpressing GSDME-NT-HATet-On treated with Dox (left) or cells overexpressing GSDME-FL treated with raptinal (right). Data are representative of at least three independent experiments (a, b, d-g).
Extended Data Fig. 9 Inhibition of GSDME-NT autophagic degradation improves chemotherapeutic response in vivo.
(a) Immunoblots of CT26 and B16 cells overexpressing mGSDME. (b) B16 mGSDME ATG4B(C74A)Tet-On cells were implanted into female C57BL/6 mice, with Dox and/or cisplatin (n = 6). (c) Immunoblots and analysis of mouse GSDME in tumours. (d) Immunoblots showing the subcellular localization of mouse GSDME-NT in tumours. (e) Representative immunofluorescence images and the percentage of negative nuclear HMGB1 cells in tumour sections treated as in (b). Scale bar: 100 µm. (f) Gating strategy of CD45 + , F4/80-, CD11B-, CD11C + DC cells. (g-h) Flow cytometry analysis of CD80 and MHC II expression in DCs (g), CD8 + T cells and IFN-γ, TNF production (h) in tumours treated as in (b). (i) Knockout efficiency of mouse GSDME. (j-k) Gsdme knockout CT26 mGSDME Flag–ATG4B(C74A)Tet-On cells were implanted in BALB/c mice and treated with Dox and/or cisplatin (n = 6). Tumour growth curves (j). Flow cytometric analysis of CD8 + T cells and IFN-γ, TNF-α production (k). (l) Growth of CT26 mGSDME Flag–ATG4B(C74A)Tet-On cells in BALB/c mice treated with anti-CD8 antibody. Mice received Dox and/or cisplatin (n = 6). (m) Knockdown efficiency of mouse TOLLIP. (n-q) B16 mGSDMETet-Tollip shRNA cells were implanted into C57BL/6 mice, with Dox and/or cisplatin (n = 6). Tumour growth curves (n). Immunoblots and analysis of mouse GSDME in tumours (o). Flow cytometry analysis of CD80 and MHC II of DCs (p), CD8 + T cells and IFN-γ, TNF production (q). Data are shown as Mean ± SD. n = 6 biologically independent mice. Two-way ANOVA with Tukey’s multiple-comparison test (b, j, l, n), two-tailed unpaired Student’s t test (c, d, o) or one-way ANOVA (e, g, h, k, p, q) were used for analysis.
Extended Data Fig. 10 GSDME and TOLLIP antibody validation and HONE1 cell line authentication.
(a) Immunohistochemical staining for GSDME was performed on tumour sections from HK1 GSDME-knockout xenografts in NSG mice (negative control) and human colon tissue sections (positive control). Scale bar, 40 μm. (b) Immunohistochemical staining for TOLLIP was performed on tumour sections from HK1 TOLLIP-knockout xenografts in NSG mice and human pancreas tissue sections (positive control). Scale bar: 40 μm. (c) STR profiles of HONE1 provided by Shanghai QiDa Biotechnology. (d) STR profiles of HONE1 provided by Pricella Biotechnology. (e) Comparison of STR profiles between samples and the STR information of the HONE1 cell line.
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–11.
Supplementary Video 1 (download MP4 )
Live-cell imaging of cells expressing GSDME-NT–EGFP and EEA1–mCherry.
Supplementary Video 2 (download MP4 )
Live-cell imaging of cells transfected with GSDME-NT–EGFP and treated with LysoTracker stain.
Supplementary Video 3 (download MP4 )
Live-cell imaging of cells transfected with GSDME-NT–EGFP and mCherry–TOLLIP.
Supplementary Tables 1–7 (download XLSX )
Supplementary Table 1: Proteins identified from immunoprecipitants pulled down with GSDME-NT-HA. Supplementary Table 2: HPLC–MS analysis of the supernatants of cells with GSDME-NT pores. Supplementary Table 3: Used signature in this paper. Supplementary Table 4: The clinicopathological characteristics of patients in SYSUCC NPC cohort 1. Supplementary Table 5: The clinicopathological characteristics of patients in SYSUCC NPC cohort 2. Supplementary Table 6: Primer sequences for qRT–PCR, sgRNA, siRNA and shRNA assays. Supplementary Table 7: List of antibodies used in this study.
Supplementary Data 1 (download PDF )
Unprocessed western blots for Supplementary Figs. 1–11.
Supplementary Data 2 (download XLSX )
Statistical source data for Supplementary Figs. 1–11.
Source data
Source Data Figs. 1–8 (download XLSX )
Statistical source data.
Source Data Figs. 1–8 (download PDF )
Unprocessed western blots.
Source Data Extended Data Figs. 1–7 and 9 (download XLSX )
Statistical source data.
Source Data Extended Data Figs. 1–9 (download PDF )
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Xu, Z., Li, Z., Deng, Z. et al. TOLLIP targets GSDME-NT-carrying endocytic vesicles for autophagy to regulate pyroptosis and chemotherapy efficacy. Nat Cell Biol 28, 812–827 (2026). https://doi.org/10.1038/s41556-026-01902-2
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41556-026-01902-2
