
Abstract
Artificial light pollution is increasing worldwide with pervasive effects on ecosystem structure and function, yet its influence on ecosystem metabolism remains largely unknown. Here we combine artificial light at night (ALAN) intensity metrics with eddy covariance observations across 86 sites in North America and Europe to show that ALAN indirectly decreases annual net ecosystem exchange by enhancing ecosystem respiration (Re). At half-hourly and daily scales, we detect consistent nonlinear interactions between ALAN and night duration, with Re increasing under higher ALAN and partially decoupling from gross primary production. At the annual scale, gross primary production shows no direct ALAN response and is instead influenced by the growing season length and urban proximity, whereas Re responds more strongly and consistently across timescales. Our findings show that ALAN disrupts the fundamental energetic constraints on ecosystem metabolism, warranting the inclusion of light pollution in global change and carbon–climate feedback assessments.
Similar content being viewed by others


Main
Artificial light pollution is accelerating across the globe1,2 and has widespread consequences for people3,4 and the planet5,6,7. Shifts in the luminance and spectral composition of the nocturnal environment modify the physiology, behaviour and ecological interactions of organisms7,8,9,10,11, which together play a fundamental role in ecosystem metabolism12,13. Ecosystem metabolism, comprising gross primary production (GPP) and ecosystem respiration (Re), directs the magnitude and direction of carbon–climate feedbacks via net ecosystem exchange (NEE)14. Around one quarter of global terrestrial ecosystems are exposed to artificial light at night (ALAN)15, but the effects on ecosystem metabolism are currently unknown.
Changing daily and seasonal cycles of light and dark10 could decouple the timing of biological processes across trophic networks16. Trophic groups are also exposed to ALAN at different intensities and have varying sensitivities to luminance and spectral composition17. Plant responses to photoperiod are influenced even at low ALAN intensities18,19, and longer-term exposure influences seasonal phenology, growth form, resource allocation and, thus, potentially carbon fixation20. High ALAN intensity exposure in urban areas disrupts the behavioural patterns of nocturnally migrating birds21 and plant diversity22 and restructures soil microbial communities, reducing the functional genes involved in nutrient regulation and plant health23. Together, the observed effects of ALAN across levels of biological organization and diverse taxa suggest a potential cascading impact on ecosystem structure and function. Previous studies of ALAN effects, however, have focused on local or experimental manipulations, leaving uncertainty about whether ALAN effects persist at the ecosystem level and longer timescales.
GPP and Re are fundamentally constrained by shortwave (solar) radiation (SW) and temperature (T), respectively24,25,26. That is, SW determines the direction and duration of energy flow between the atmosphere and ecosystems, and T determines the rate of reactions12. Although ALAN is not expected to influence SW or T directly, artificial light could disrupt the processing of energy according to these fundamental constraints via acclimation, compensation and adaptation strategies27,28. A better understanding of the magnitude and direction of ALAN effects on ecosystem metabolism could help constrain carbon–climate processes in Earth system models (ESMs)29. Specifically, largely uncertain ESM processes and their response to climatic factors could be compounded by the chronic effects of pervasive anthropogenic stressors, such as ALAN.
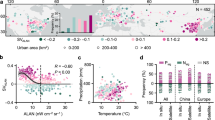
Global efforts to measure carbon exchange across diverse ecosystems30 combined with satellite observations of ALAN distribution and intensity across the land surface2,31 enable the exploration of artificial light’s influence on terrestrial ecosystem metabolism. Here, we leverage the harmonized nighttime light dataset of Li et al.32 and eddy covariance observations from FLUXNET201530 to investigate the instantaneous and aggregated influence of ALAN on ecosystem-scale NEE, GPP and Re fluxes. Although both datasets have global coverage, the location of eddy covariance flux towers are biased towards dark sky regions (Extended Data Fig. 1). Following definitions by Li et al.32 and others33, we use three digital number (DN, higher values represent greater luminance of light at night; Methods) groups representative of low (DN <10), medium (DN ≥ 10 < 30) and high (≥30, representative of urban boundaries) ALAN intensity to identify regions with FLUXNET2015 sites across a range of ALAN intensities. North America and Europe were the only regions, globally, with more than one high ALAN intensity FLUXNET2015 site (Methods; Fig. 1a,d). Within both North America and Europe, sites were selected on the basis of latitudinal ranges at which medium or high ALAN intensity sites were present (Fig. 1b,e) to minimize climatic factors in higher or lower latitude sites being ascribed to low ALAN intensities. In total, 86 FLUXNET2015 sites were selected, 34 sites in North America (4, 5 and 25 sites at high, medium and low ALAN intensities, respectively) and 52 sites in Europe (13, 17 and 22 sites at high, medium and low ALAN intensities, respectively) (Methods; Fig. 1 and Supplementary Table 1). Despite regional imbalances in FLUXNET2015 site distribution across ALAN intensity levels, the dataset captures a diverse range of ALAN intensities across temperate regions experiencing similar seasonal fluctuations in T and SW.

a,d, The location of 86 eddy covariance flux tower sites from FLUXNET2015 (symbols, colours indicate ALAN intensity according to DN (higher values represent greater luminance of light at night) (as in d) displayed over a harmonized global nighttime light map for 2012 (for visualization only) in North America (n = 34) (a) and Europe (n = 52) (d). b,e, The latitudinal distribution of sites with different ALAN intensities for North America (b) and for Europe (e), in 2° N intervals. c,f, The ALAN intensities of selected FLUXNET2015 sites, averaged across site years (the number of years with observational data per site), for North America (c) and Europe (f) according to DN with symbol size indicating number of site years (range: 1–20 years per site between 1992 and 2014, total site years in c is 211 and in f is 412). Basemaps in a and d were generated with QGIS using the harmonized global nighttime light dataset32 under a Creative Commons license CC BY 4.0.
To detect the potential influence of ALAN on ecosystem metabolism, we investigate half-hourly and mean daily ecosystem carbon fluxes (Fc; Fc: NEE, GPP and Re) measurements against their fundamental constraints, T and SW, according to the modified Arrhenius equation of Weyhenmeyer12:
where Fc is ecosystem C flux (NEE, GPP, Re) (in µmol CO2 m−2 s−1), T is temperature in Kelvin, σ is the Stefan–Boltzmann constant (in J m−2 s−1 K−4) (5.67 × 10−8), SW is incoming SW (in J m−2 s−1,) kFc is the slope of the linear relationship and b is the intercept. The function establishes a biophysically grounded baseline for different Fc by capturing their shared fundamental constraints (T and SW). The use of the modified Arrhenius function in this study primarily serves as a comparative baseline rather than a mechanistic model, enabling deviations attributable to chronic ALAN effects to be identified relative to fundamental energetic constraints.
The null models for NEE, GPP and Re were linear mixed effect models (LMMs) or generalized additive mixed models (GAMMs) fitted to equation (1) (Methods) with FLUXNET2015 site and latitude as random effects and fundamental constraint (\({T}^{4}\sigma \,{{\rm{e}}}^{\frac{-\mathrm{SW}}{\sigma {T}^{4}}}\), J m−2 s−1) as a fixed effect (Fig. 2). The null models were tested against models with additional explanatory variables, including continent, climate, International Geosphere–Biosphere Programme land use classifications, growing season (GS), night duration (ND, hours), vapour pressure deficit (VPD, hectopascals), precipitation (P, millimetres), ALAN intensity (DN), distance to nearest urban polygon (DtNUP, kilometres) and proportion of urban land cover in 3- and 10-km buffers around each site (pULC_3km, pULC_10km). The LMM selection criteria for explanatory variables followed a trade-off between explanatory power and parsimony, with the condition that additional degrees of freedom (df) were accompanied by lower Akaike information criteria (AIC) and higher marginal R2 (R2m) goodness-of-fit measures (ΔAICdf <−5 and ΔR2mdf >0.01 compared with the null model (Methods; Fig. 2a,c,e). All Fc LMMs selected GS; GPP and Re LMMs selected ND; the GPP LMM selected DtNUP; and the Re LMM selected VPD and ALAN (Supplementary Tables 2 and 3).

a,c,e, The symbols are half-hourly FLUXNET2015 measurements for NEE (gold symbols) (a), daytime GPP (blue symbols) (c) and nighttime Re (magenta symbols) (e) for 86 sites across North America and Europe. The linear regression lines in a, c and e indicate fixed-effect relationships of fundamental constraints on ecosystem carbon fluxes according to the modified Arrhenius function (null models as in equation (1)). b,d,f, The box plots display the distribution of measured fluxes across bins of the modified Arrhenius function (axes labels and units in b, d and f are the same as in a, c and e, respectively) grouped by ALAN intensity to illustrate variation in carbon fluxes relative to energetic constraints as a function of ALAN. The boxes represent interquartile ranges (IQR), the horizontal lines denote medians, the whiskers extend to 1.5 × IQR and the points indicate outliers.
Backward selection and variance-weighting were applied to GAMMs fitted to half-hourly NEE, GPP and Re observations (Methods; Supplementary Table 4) with the explanatory variable identified in the LMMs. In the final GAMMs, all Fc retained a significant interaction between ALAN and ND (Fig. 3 and Supplementary Table 5). Notably, the NEE GAMM did not retain GS as a significant predictor, suggesting that the seasonality in instantaneous NEE responses was captured by GPP and Re, which both retained GS effects (Supplementary Table 5). During GAMM selection, VPD (selected in the Re LMM) exhibited consistently high concurvity (>0.8) with other smooth terms, including models in which ALAN was removed, indicating substantial collinearity with the modified Arrhenius function. DtNUP, selected in the GPP LMM, contributed no additional explanatory power in all GAMMs.

a–i, GAMMs fitted to half-hourly carbon flux measurements consistently selected smooth tensor product interactions between ALAN and ND for NEE (a–c), GPP (d–f) and Re (g–i). a (R2adj = 0.430; fREML = 1.35 × 107), d and g show observed versus predicted fluxes for the final variance-weighted GAMMs (Supplementary Table 5), which account for heteroscedasticity across the range of observed fluxes (Supplementary Table 4 and Extended Data Fig. 2). b, e and h illustrate smooth estimates of the tensor product interaction across the gradient of ALAN intensity and ND (coloured lines, with shaded ribbons representing mean predictions ± 95% confidence intervals). For GPP, e, and Re, h, predictions are shown separately for the GS and non-GS, whereas for NEE, b, the GS was not selected as a significant predictor. c, f and i depict GAMM-derived estimates of the relative change in each flux across gradients of ALAN intensity and ND, expressed relative to ALAN = 0, with shaded areas denoting 95% confidence intervals around the mean.
The variance weighting substantially reduced residual heteroscedasticity across all Fc, with the scale estimate reduced by ~95% and adjusted R² (R2adj) reduced by 0.06–0.11 in weighted compared with unweighted final GAMMs, indicating improved model stability through decreased overfitting to high-variance observations (Supplementary Table 4 and Extended Data Fig. 2). Figure 3, right panels, shows weighted GAMM estimates of relative changes in each Fc across gradients of ALAN intensity and ND. Partial effect surfaces illustrating nonlinear ALAN × ND interactions at half-hourly timescales are presented in Extended Data Fig. 3, along with residual diagnostics indicating no substantial autocorrelation after model fitting.
LMMs and GAMMs fitted to mean daily NEE, GPP and Re yielded more consistent trends compared with models fitted to half-hourly observations. All daily LMMs and GAMMs identified GS and ND as significant predictors, with the GPP LMM selecting DtNUP and the Re LMM selecting ALAN as explanatory variables (Supplementary Tables 6 and 7). All of the daily GAMMs selected the smooth tensor product between ALAN and ND (Supplementary Tables 8 and 9). Compared with the half-hourly models, the daily GAMMs exhibited smoother and more monotonic relationships between ALAN and Fc, reflecting the reduction in diel and short-term variability through temporal aggregation (Extended Data Fig. 4). Temporal aggregation led to clearer trends in predicted relative changes in Fc across gradients of ALAN intensity (Fig. 4), in contrast to more variable patterns in the half-hourly GAMM predictions (Fig. 3c,f,i). Notably, whereas ALAN consistently increased Re in half-hourly GAMMs and particularly during short nights (Fig. 3h,i), the daily GAMMs showed a contrasting pattern, with Re increasing most with ALAN intensity during longer nights (Fig. 4c,f). This divergence demonstrates how the aggregation of diel variability can modify the apparent direction and magnitude of ALAN effects on Fc. The partial effect surfaces from daily models showed more regular gradients and reduced nonlinear complexity, whereas residual autocorrelation was minimal, supporting the suitability of daily models for capturing net ALAN effects on ecosystem metabolism (Extended Data Fig. 5).

a–c, The box plots display the distribution of measured mean daily NEE (a), GPP (b) and Re (c) across ND groups (short ND: <9.5, mid ND: ≥9.5 < 13.9, long ND: ≥13.9 h, defined by 25% and 75% quartiles) coloured by ALAN intensity (n = 197,247). The boxes represent the interquartile ranges (IQR), the horizontal lines denote medians, the whiskers extend to 1.5× the IQR and the points indicate outliers. d–f, The variance-weighted GAMM predictions (Supplementary Table 9) for relative changes in daily mean NEE (d), GPP (e) and Re (f), expressed relative to ALAN = 0 for ND groups as in Fig. 3, with shaded ribbons representing mean predictions ± 95% confidence intervals.
The role of ALAN along with longer-term drivers of ecosystem metabolism was evaluated by constructing a piecewise structural equation model (SEM) integrating multiple exogenous predictors and hypothesized mediation pathways. The final SEM incorporated GS length, ALAN intensity and climatic variables including SW, VPD and T, along with the urban metric DtNUP (Fig. 5a). The modified Arrhenius function was not selected, reflecting how annual temporal aggregation reduces positive and negative deviations in fundamental constraints compared with short-term flux variability. The aggregated measures of ND were also not selected, with phenological drivers such as GS length more important at annual timescales (Supplementary Table 10). The mediation analysis, using nonparametric bootstrap resampling to quantify both direct and indirect effects of GS length and indirect effects of ALAN on NEE, supports the inference that the influence of ALAN on ecosystem metabolism is primarily mediated through increased Re (Fig. 5b). The influence of GS length on NEE was significantly mediated through increased GPP (Fig. 5b). The leave-one-out sensitivity analysis of the SEM indicated that no alternative model performed better than the full model (Supplementary Table 11). Notably, the exclusion of ALAN, DtNUP, VPD or GS length led to significant declines in model performance, reflecting the importance of these predictors in explaining the interannual variation in ecosystem metabolism (Fig. 5c).

a, The hypothesized pathways linking explanatory variables to ecosystem carbon fluxes show the standardized path coefficients of the final SEM with significance levels (****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05, exact P values in Supplementary Table 10). The tests are two-sided, with no adjustments for multiple comparisons. The arrow thickness indicates the magnitude of standardized effect sizes, the black arrows and text indicate direct pathways, the double-headed grey arrows and text indicate the residual correlations and the blue arrows and text indicate the mediation pathways supported by the bootstrap analysis. The SEM was fitted to complete cases for all variables (605 site years, 84 sites). b, The bootstrap-derived estimates shown are means and 95% percentiles from 1,000 replicates for the direct and indirect effects of GS length and ALAN on NEE. c, The outputs from a leave-one-out analysis show the change in marginal (lighter-coloured symbols and lines) and conditional (darker-coloured symbols and lines) R² (ΔR²) for NEE (gold), GPP (blue) and Re (magenta) relative to the full model after systematically removing each exogenous predictor (Supplementary Table 11).
To ensure our data analysis was robust to site bias across ALAN intensities (17, 22 and 47 sites at high, medium and low ALAN intensities, respectively, and 34 sites in North America and 52 sites in Europe), we repeated all GAMM and SEM analyses using a balanced dataset with an equal representation of low, medium and high ALAN intensity sites per continent (Extended Data Figs. 6–9). The models fitted to the balanced dataset showed consistently significant nonlinear interactions between ALAN and ND across temporal scales (Extended Data Fig. 6), and the annual SEM retained the core structure of ALAN indirectly influencing NEE through increased Re (Extended Data Fig. 9). Whereas several weaker interactions (for example Re ~ T and NEE ~ GS length) were no longer significant owing to reduced sample size, the SEM retained dominant pathways, and the overall explanatory power was comparable (R2m = 0.64, R2c = 0.70 for NEE). Notably, the standardized coefficients strengthened between ALAN ~ Re and Re ~ NEE when the SEM was fitted to the balanced dataset. Our observed ALAN effects on ecosystem metabolism are therefore robust to spatial imbalances in ALAN intensity across FLUXNET2015 site distribution.
Our study provides cross-continental evidence of ALAN’s influence on ecosystem metabolism across timescales. We demonstrate that ALAN consistently modifies the relationship between Fc and their fundamental energetic constraints (Figs. 2–4). The Re response to fundamental constraints was particularly sensitive to ALAN intensity at short (half-hourly and daily) timescales (Fig. 2 and Supplementary Tables 2 and 6). Alongside Re, GPP and NEE exhibited significant nonlinear interactions between ND and ALAN intensity, revealing the importance of ALAN magnitude and timing in modulating ecosystem metabolism across scales (Figs. 3 and 4). At annual timescales, the influence of ALAN on NEE was primarily mediated through increased Re rather than the direct suppression of GPP (Fig. 5). Taken together, our findings demonstrate the role of ALAN as a pervasive stressor capable of disrupting carbon balance across spatial and temporal scales.
The nonlinear influence of ALAN on ecosystem metabolism was strongly modulated by diel cycles and seasonality, demonstrating the importance of phenological dynamics34 and biogeochemical feedbacks in shaping long-term carbon balance35. The temporal aggregation led to notable shifts in the strength and direction of ALAN effects on Re, whereas GPP and NEE displayed more consistent nonlinear responses to ND across timescales (Figs. 3 and 4). At the half-hourly resolution, short nights showed the strongest ALAN-induced increases in Re (Fig. 3h,i), reflecting immediate physiological and microbial responses such as prolonged stomatal opening36, sustained leaf dark respiration37 and elevated microbial decomposition under disrupted circadian regulation38. By contrast, daily mean nighttime Re estimates indicated larger ALAN-related increases during longer nights (Fig. 4f), demonstrating how aggregation dampens short-term variability while revealing broader shifts in Re across longer nights. GPP exhibited consistent positive or nonlinear ALAN effects across timescales (Figs. 3 and 4), probably driven by nocturnal illumination extending photosynthetic activity at medium ALAN intensities39,40. Temporal scale and ND thus collectively shape ALAN’s ecological impact, whereas diel averaging can obscure short-lived physiological responses while reflecting cumulative nighttime effects.
Re exhibited greater sensitivity to ALAN than GPP across timescales (Figs. 2 and 3 and Supplementary Tables 2–11), and the SEM confirmed that ALAN primarily influences NEE indirectly via increased Re at annual timescales (Fig. 5). The destabilizing effect of ALAN on production-respiration coupling will arise from shifts in multiple autotrophic and heterotrophic processes controlling carbon allocation and use efficiency17,38. The greater Re sensitivity may reflect a higher capacity of autotrophs to acclimate to ALAN through conservative growth strategies such as increased shoot-to-root ratios20,41. In ecosystems dominated by C3 plants, for instance, prolonged ALAN exposure can disrupt circadian regulation and prolong stomatal opening, reducing carbon uptake efficiency, increasing mortality and senescence, and leading to reduced GPP over time42,43. Such trophic mismatches and shifts in carbon allocation are likely to accumulate across levels of biological organization, space and time44,45, leading to progressive declines in NEE in illuminated ecosystems.
The ecological impacts of ALAN have primarily been examined at local scales6,17, but landscape-scale factors will confound or amplify these localized effects2. Urban proximity influenced GPP in our analysis, whereas ALAN directly influenced Re (Fig. 5), suggesting distinct pathways through which nighttime lighting and urban characteristics modify ecosystem metabolism. Balancing sites across low, medium and high ALAN intensities further indicates potentially stronger mediating effects of ALAN on NEE via Re (Extended Data Fig. 9). Despite the pervasive nature of light pollution, ALAN remains overlooked in ESM carbon–climate projections that otherwise account for climate and land use changes. Current observational data, however, do not enable the disentangling of the contribution of ALAN relative to sunlight in shaping Fc, and future targeted experimental studies will be needed to resolve these relationships.
Global eddy covariance networks such as FLUXNET are vital for monitoring ecosystem metabolism across diverse climates and land use types, but they are typically biased towards temperate regions, seminatural landscapes and dark skies (Extended Data Fig. 1). Urban flux towers are particularly scarce, and although networks such as Urban PLUMBER have been established, they do not measure Fc (ref. 46). Similarly, available nighttime light satellite products used here (Visible Infrared Imaging Radiometer Suite (VIIRS) and Defence Meteorological Satellite Programme (DMSP)) are coarse in spatial resolution, are insensitive to blue light emitted by white light-emitting diode (LED) lighting47 and cannot fully capture local heterogeneity in ALAN exposure at flux tower sites.
Enhanced satellite sensors with improved spectral and spatial resolution would advance ALAN monitoring2, but ground-based measurements are also needed to capture how cloud cover exacerbates or reduces skyglow (brightening of the night sky) in high or low ALAN intensity areas, respectively48. The coordinated expansion of eddy covariance flux tower networks along with complementary measurements, such as chamber-based respiration estimates and isotopic tracers, will be critical to disentangle the mechanisms by which ALAN alters ecosystem metabolism. Expanding ecosystem-level Fc measurements into urbanized, tropical, arid and high-latitude regions is vital to evaluate the global relevance of ALAN impacts on carbon cycling. While monitoring is essential, mitigation is also readily achievable.
Artificial light is ubiquitous and often beneficial, but the negative ecological effects of light pollution can be reduced while balancing societal benefits. Retrofitting LED lighting can reduce light pollution5, but it often results in over-illumination due to their higher efficiency49. Given that lighting accounts for 20% of global electricity consumption and 6% of CO2 emissions50 and can exacerbate degraded air quality51,52, mitigation interventions such as directional, dimmable and adaptive lighting designs2 offer wider cobenefits. Unlike climate and land use change, the effects of light pollution could be mitigated overnight53. Our study demonstrates the pervasive influence of light pollution on ecosystem metabolism across scales and highlights the urgent need to integrate ALAN into global change research, assessments of carbon–climate feedbacks and mitigation strategies. Developing a higher resolution understanding of species, community and ecosystem sensitivity to ALAN will be central to designing interventions that both safeguard biodiversity and preserve the land carbon sink.
Methods
We combined satellite-derived ALAN intensity metrics with eddy covariance flux measurements from 86 FLUXNET2015 sites in North America and Europe. The analyses were conducted at half-hourly, daily and annual timescales to capture short-term physiological responses, aggregated diel patterns and long-term ecosystem dynamics. All Fc (NEE, GPP and Re) were first evaluated against their fundamental energetic constraints of T and SW according to the modified Arrhenius function (equation (1)) using LMMs. To explore nonlinear interactions, we then applied GAMMs which allow the flexible estimation of smooth terms. At annual timescales, we used piecewise SEM to account for collinearity among multiple drivers and to partition direct and indirect effects of ALAN, GS length and climatic variables on Fc. This hierarchical modelling framework enabled the consistent evaluation of ALAN effects across temporal scales.
Global harmonized nighttime light dataset
The Day/Night Band of the VIIRS is the only satellite radiometer currently acquiring imagery of the Earth at night, providing single-band nightscapes at a resolution of 750 m since 2012 across the globe. Prior global nightscapes were monitored by DMSP–Operational Linescan System (OLS) at a resolution of 1 km between 1992 and 2013. A harmonized global nighttime light dataset, developed by Li et al.32, provides a consistent annual time-series of overall luminance between 1992 and 2018 at ~1 km resolution through the intercalibration of DMSP-like DN values. The DNs for all FLUXNET2015 sites (see below) and site years were derived from the harmonized dataset of Li et al.32. The low DN sites (DN <10) were cross-checked in Google Earth for each site year to verify low DN values reflected remote locations by noting distances to the nearest built-up area in QGIS (version 3.30.3). The satellite-derived DN values from DMSP–Operational Linescan System and VIIRS represent relative radiance indices and cannot be directly converted into absolute illuminance units such as lux, as they do not capture spectral composition or ground-level variability54. The ALAN categories used here should thus be interpreted as relative exposure gradients rather than specific ecological thresholds.
FLUXNET dataset and site selection
FLUXNET is a global network of micrometeorological sites providing eddy covariance CO2 exchange observations between terrestrial ecosystems and the atmosphere30. The FLUXNET2015 dataset used in this study includes measurements from 210 eddy covariance flux towers across the globe30. A total of five tier-2 sites and two arctic sites outside the latitudinal range of the global nighttime light dataset (latitude >75° N) were excluded. Originally, DNs from the harmonized nighttime light dataset were extracted for 203 FLUXNET2015 sites from 1992 to 2014 (1,474 site years) (Extended Data Fig. 1). The 203 sites were composed of 1,116 low (DN <10), 243 medium (DN ≥ 10 < 30) and 115 high (DN >30) ALAN intensity site years. Only one high ALAN intensity FLUXNET2015 site (JP-SMF) was located outside of North America or Europe, with no replication of low ALAN intensity sites in a 2° latitudinal or longitudinal range. The site selection was therefore restricted to North America and Europe to reduce noise from additional climatic and ecosystem properties at low ALAN intensity FLUXNET2015 sites globally. Within both North America and Europe, the latitudinal and longitudinal ranges of selected FLUXNET2015 sites were based on the presence of medium or high ALAN intensity sites at 2° intervals (Fig. 1 and Supplementary Table 1).
Disentangling respiration and photosynthesis fluxes during the day is complex and relies on modelling techniques with high uncertainty, particularly under low turbulence or during transitional periods around dawn and dusk. The FLUXNET2015 dataset undergoes processing to check data quality, filter low turbulence periods and CO2 flux partitioning into respiration and photosynthesis using established methods30. The measurements were compiled from the FLUXNET2015 dataset55, which in this study includes non-gap-filled half-hourly and annual air temperature (TA_F), incoming shortwave (SW_IN_F), NEE (NEE_VUT), nighttime Re (RECO_NT) and daytime GPP (GPP_DT) measurements for 86 sites across 623 site years. Along with the use of nighttime Re and daytime GPP, the half-hourly data were filtered for Re by selecting timepoints with GPP <0.001 µmol CO2 m−2 s−1 and outgoing SW greater than incoming SW and vice versa for daytime GPP.
Additional environmental and urban variables were derived to check for confounding effects, including half-hourly and annual VPD (VPD_F) and P (P_F) from FLUXNET2015. Urban metrics pULC_3km, pULC_10km and DtNUP were calculated by quantifying the proportion of land cover classified as urban within 3- and 10-km buffers around each site, using the ESA CCI Land Cover dataset, and computing the Euclidean distance (km) from the site centroid to the nearest urban polygon in the Copernicus Urban Centre Database56. To account for latitudinal and climatic variation in phenology across the 86 sites, GS was classified using site-specific 25th percentiles of daily GPP per site year. GPP measurements above the 25th percentile threshold was classified as occurring within the GS, and other observations were classified as non-GS. To avoid classifying transient periods of activity as part of the GS, we implemented a hybrid phenological rule requiring ≥5 consecutive candidate days for GS initiation, and ≥5 consecutive non-GS days to mark the GS finish. To estimate the daily duration of night at each study site, we calculated the time between astronomical sunset and sunrise (UTC) using site-specific latitude, longitude and observation dates using the suncalc package. The ND was calculated as the time in hours elapsed between sunset on a given day and sunrise on the following day.
Model analysis
All model analyses were conducted in R statistical software (version 4.2.2)57. The null models (equation (1)) describe the relationship between Fc (NEE, GPP and Re) and their fundamental constraints (SW and T) according to a modified Arrhenius function12. Unlike alternative functions such as those used in metabolic ecology26, the modified Arrhenius function enables the exploration of NEE, GPP and Re according to a single measure of shared fundamental constraints and analysis of untransformed Fc measurements. The null models for NEE, GPP and Re according to equation (1) were fitted to half-hourly, daily and annual FLUXNET2015 measurements, with LMMs and GAMMs applied to half-hourly and daily measurements and an SEM developed for annual timescales.
LMMs
First, LMMs were incrementally tested for each carbon flux and explanatory variable: continent (North America and Europe), climate (boreal, temperate and Mediterranean), International Geosphere–Biosphere Programme land use classifications (CRO, CSH, DBF, EBF, ENF, GRA, MF and WET), GS (Y and N), ND (hours), month, hour of the day, VPD, P, pULC_3km, pULC_10km, DtNUP and ALAN. FLUXNET2015 site (n = 86) and latitude (n = 80) were included as random effects to account for spatial clustering. The model selection thresholds (ΔAICdf <−5 and ΔR2mdf >0.01) ensured that any increase in explanatory power was proportionate to model complexity and prevented LMM overfitting by only relying on ΔAIC selection criteria. The bootstrapped 95% confidence intervals for LMM marginal and conditional R2 fits were computed by resampling model residuals using 500 semiparametric bootstrap replicates. For the half-hourly datasets (4–10 million observations) we used random 10% subsamples to provide reliable estimates while avoiding computational limitations inherent in very large datasets.
GAMMs
GAMMs were used to explore nonlinear relationships and interactions between variables, with initial GAMMs including variables identified as potentially important in the LMMs for the temporal resolution (half-hourly and daily) under consideration. The smooth terms were specified for continuous explanatory variables, and the categorical variables were treated as parametric effects. The tensor product interactions and stratification of categorical variables were also tested where ecologically feasible, and FLUXNET2015 site and latitude were included as random smooth terms. All final GAMMs identified latitude as a redundant random effect, defined statistically as a lack of improvement in model fit and a concurvity value of 1, indicating complete collinearity with other smooth terms. We use a backward selection approach to sequentially simplify the initial models, with GAMM selection based on a combination of penalized likelihood (fREML), R2adj, deviance explained (%) and approximate significance of smooth terms. We did not apply R2adj per df thresholds as with LMMs, as GAMMs inherently penalize smooth term complexity during estimation to optimize effective df.
To address the risk of overfitting and concurvity (collinearity between smooth terms), we evaluated GAMM diagnostics and smooth terms exhibiting high concurvity values (>0.8) were identified as potentially redundant. Preference during model selection was given to simpler models that retained comparable, although usually lower, explanatory power while reducing concurvity. Additional explanatory variables were then reintroduced to the final backward selected GAMMs to compare model performance. Finally, heteroscedasticity (non-constant residual variance) in the final GAMMs was accounted for by comparing the final selected GAMMs to variance-weighted GAMMs, which provide lower weight to observations associated with higher residual variance. Model performance and smooth term significance were compared between the unweighted and weighted GAMMs to ensure robust evidence for the selected explanatory variables. For all GAMMs, residual autocorrelation was evaluated using partial autocorrelation functions.
SEM
To investigate the relationships between environmental drivers and annual NEE, GPP and Re, we developed a piecewise SEM comprising three linked LMMs, including NEE ~ GPP + Re, GPP ~ SW and Re ~ GPP + T and testing additional exogenous predictors (GS length, mean ND, VPD, P, ALAN, pULC_3km, pULC_10km and DtNUP). The annual dataset included 605 site years across 84 sites after excluding site years with missing variables. Both LMMs and GAMMs were evaluated for component models. Given the relatively small sample size, GAMMs presented a higher risk of overfitting and unstable smooth functions at the annual timescale. The annual aggregation of Fc measurements also inherently smoothed diel and seasonal nonlinearities observed in half-hourly and daily measurements. Exploratory diagnostics further indicated that annual relationships were approximately linear, supporting the use of LMMs as a parsimonious framework capable of accounting for site-level random intercepts while estimating fixed effects on annual Fc. The SEMs included residual covariance terms among exogenous predictors to account for collinearity, and model fit between SEM’s was evaluated through Fisher’s C and P, AIC and df and marginal and conditional R2 for NEE, GPP and Re.
The mediation pathways in the final SEM quantified the indirect effect of GS length on NEE via GPP and the indirect effect of ALAN on NEE via Re. The uncertainty in direct and indirect effects was estimated through nonparametric bootstrap resampling (1,000 iterations). In each iteration, the three component LMMs were refitted on a bootstrap-resampled dataset with replacement, and indirect effects were calculated as the product of relevant path coefficients. Percentile bootstrap confidence intervals (95%) were derived for each estimated effect and considered significant if they did not overlap zero. The relative importance of each exogenous predictor in the final SEM was inferred through a leave-one-out sensitivity analysis, in which each variable was removed in turn and the reduced SEM fit compared, rather than absolute effect sizes. The model fit for each alternative SEM specification fitted to the same dataset was compared with the full model, with higher model sensitivity indicated by a significantly poorer fit (P < 0.05), higher AIC, or reduced explanatory power (R2m and R2c for NEE, GPP and Re) relative to the final selected SEM.
Sensitivity analysis
The sensitivity of models to the composition of FLUXNET2015 sites was evaluated by generating a balanced, stratified subset of the full dataset with equal representation of low, medium and high ALAN intensity sites across both continents at half-hourly, daily and annual timescales. A random sample of sites equal to the stratum with the fewest available sites (high ALAN sites in North America, n = 4) was selected without replacement from each stratum (4 × 2 continents × 3 ALAN intensity groups = 24 sites in the balanced dataset). The half-hourly and daily GAMMs and the annual SEM were refitted to the balanced dataset using the same model specification in the main analysis.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The FLUXNET2015 data analysed in this study are available at https://fluxnet.fluxdata.org/data/fluxnet2015-dataset/ (ref. 55) and are subject to the FLUXNET data policy (https://fluxnet.org/data/data-policy). As the redistribution of raw half-hourly flux data is not permitted, we provide only derived products, including daily and annual summaries, processed variables and model outputs, which are available under a CC-BY 4.0 license via Figshare at https://doi.org/10.6084/m9.figshare.29958455 (ref. 58). The ALAN metrics used here are available at https://doi.org/10.3390/rs9060637 (ref. 32). Summaries for each FLUXNET site are also provided in Supplementary Table 1.
Code availability
The R code used for data processing and analysis in this study is available via Figshare at https://doi.org/10.6084/m9.figshare.29958455 (ref. 58).
References
Kyba, C. C. M. et al. Artificially lit surface of Earth at night increasing in radiance and extent. Sci. Adv. 3, e1701528 (2017).
Linares Arroyo, H. et al. Monitoring, trends and impacts of light pollution. Nat. Rev. Earth Environ. 5, 417–430 (2024).
Xue, P., Nôga, D. A. & Benedict, C. The dark side of light: light at night may raise the risk of type 2 diabetes. Lancet Reg. Health Eur. 42, 100955 (2024).
Jones, R. R. Exposure to artificial light at night and risk of cancer: where do we go from here?. Br. J. Cancer 124, 1467–1468 (2021).
Jägerbrand, A. K. & Spoelstra, K. Effects of anthropogenic light on species and ecosystems. Science 380, 1125–1130 (2023).
Sanders, D., Frago, E., Kehoe, R., Patterson, C. & Gaston, K. J. A meta-analysis of biological impacts of artificial light at night. Nat. Ecol. Evol. 5, 74–81 (2021).
Cieraad, E., Strange, E., Flink, M., Schrama, M. & Spoelstra, K. Artificial light at night affects plant–herbivore interactions. J. Appl. Ecol. 60, 400–410 (2023).
Liu, J. A., Meléndez-Fernández, O. H., Bumgarner, J. R. & Nelson, R. J. Effects of light pollution on photoperiod-driven seasonality. Horm. Behav. 141, 105150 (2022).
Owens, A. C. S. et al. Light pollution is a driver of insect declines. Biol. Conserv. 241, 108259 (2020).
Gaston, K. J., Davies, T. W., Nedelec, S. L. & Holt, L. A. Impacts of artificial light at night on biological timings. Annu. Rev. Ecol. Evol. Syst. 48, 49–68 (2017).
Knop, E. et al. Artificial light at night as a new threat to pollination. Nature 548, 206–209 (2017).
Weyhenmeyer, G. A. Toward a fundamental understanding of ecosystem metabolism responses to global warming. One Earth 7, 1886–1898 (2024).
Schramski, J. R., Dell, A. I., Grady, J. M., Sibly, R. M. & Brown, J. H. Metabolic theory predicts whole-ecosystem properties. Proc. Natl Acad. Sci. USA 112, 2617–2622 (2015).
Weiskopf, S. R. et al. Biodiversity loss reduces global terrestrial carbon storage. Nat. Commun. 15, 4354 (2024).
Falchi, F. et al. The new world atlas of artificial night sky brightness. Sci. Adv. 2, e1600377 (2016).
Maggi, E. et al. Artificial light at night erases positive interactions across trophic levels. Funct. Ecol. 34, 694–706 (2020).
Gaston, K. J., Visser, M. E. & Hölker, F. The biological impacts of artificial light at night: the research challenge. Philos. Trans. R. Soc. B 370, 20140133 (2015).
Runkle, E., Heins, R., Cameron, A. & Carlson, W. Flowering of herbaceous perennials under various night interruption and cyclic lighting treatments. HortScience 33, 672–677 (1998).
Blanchard, M. G. & Runkle, E. S. Intermittent light from a rotating high-pressure sodium lamp promotes flowering of long-day plants. HortScience 45, 236–241 (2010).
Bennie, J., Davies, T. W., Cruse, D. & Gaston, K. J. Ecological effects of artificial light at night on wild plants. J. Ecol. 104, 611–620 (2016).
Van Doren, B. M. et al. High-intensity urban light installation dramatically alters nocturnal bird migration. Proc. Natl Acad. Sci. USA 114, 11175–11180 (2017).
Stanhope, J., Liddicoat, C. & Weinstein, P. Outdoor artificial light at night: a forgotten factor in green space and health research. Environ. Res. 197, 111012 (2021).
Li, X.-M. et al. Artificial light at night triggers negative impacts on nutrients cycling and plant health regulated by soil microbiome in urban ecosystems. Geoderma 436, 116547 (2023).
Nemani, R. R. et al. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300, 1560–1563 (2003).
Enquist, B. J. et al. Scaling metabolism from organisms to ecosystems. Nature 423, 639–642 (2003).
Johnston, A. S. A. et al. Temperature thresholds of ecosystem respiration at a global scale. Nat. Ecol. Evol. 5, 487–494 (2021).
Kehoe, R., Sanders, D. & van Veen, F. J. Towards a mechanistic understanding of the effects of artificial light at night on insect populations and communities. Curr. Opin. Insect Sci. 53, 100950 (2022).
Altermatt, F. & Ebert, D. Reduced flight-to-light behaviour of moth populations exposed to long-term urban light pollution. Biol. Lett. 12, 20160111 (2016).
O’Sullivan, M. et al. Process-oriented analysis of dominant sources of uncertainty in the land carbon sink. Nat. Commun. 13, 4781 (2022).
Pastorello, G. et al. The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data. Sci. Data 7, 225 (2020).
Sánchez de Miguel, A., Bennie, J., Rosenfeld, E., Dzurjak, S. & Gaston, K. J. First estimation of global trends in nocturnal power emissions reveals acceleration of light pollution. Remote Sens. 13, 3311 (2021).
Li, X., Zhou, Y., Zhao, M. & Zhao, X. A harmonized global nighttime light dataset 1992–2018. Sci. Data 7, 168 (2020).
Zhou, Y., Li, X., Asrar, G. R., Smith, S. J. & Imhoff, M. A global record of annual urban dynamics (1992–2013) from nighttime lights. Remote Sens. Environ. 219, 206–220 (2018).
Wang, L. et al. Artificial light at night outweighs temperature in lengthening urban growing seasons. Nat. Cities 2, 506–517 (2025).
Davidson, E. A. & Janssens, I. A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165–173 (2006).
Matzke, E. B. The effect of street lights in delaying leaf-fall in certain trees. Am. J. Bot. 23, 446–452 (1936).
de Dios, V. R. et al. Processes driving nocturnal transpiration and implications for estimating land evapotranspiration. Sci. Rep. 5, 10975 (2015).
Hölker, F., Wolter, C., Perkin, E. K. & Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 25, 681–682 (2010).
Meng, L. et al. Artificial light at night: an underappreciated effect on phenology of deciduous woody plants. PNAS Nexus 1, pgac046 (2022).
Lo Piccolo, E. et al. Shedding light on the effects of LED streetlamps on trees in urban areas: Friends or foes?. Sci. Total Environ. 865, 161200 (2023).
Qi, Y., Wei, W., Chen, C. & Chen, L. Plant root-shoot biomass allocation over diverse biomes: a global synthesis. Glob. Ecol. Conserv. 18, e00606 (2019).
Didaran, F. et al. The mechanisms of photoinhibition and repair in plants under high light conditions and interplay with abiotic stressors. J. Photochem. Photobiol. B 259, 113004 (2024).
Anic, V., Gaston, K. J., Davies, T. W. & Bennie, J. Long-term effects of artificial nighttime lighting and trophic complexity on plant biomass and foliar carbon and nitrogen in a grassland community. Ecol. Evol. 12, e9157 (2022).
Johnston, A. S. A. Predicting emergent animal biodiversity patterns across multiple scales. Glob. Change Biol. 30, e17397 (2024).
Sanders, D. et al. How artificial light at night may rewire ecological networks: concepts and models. Philos. Trans. R. Soc. B 378, 20220368 (2023).
Lipson, M. et al. Harmonized gap-filled datasets from 20 urban flux tower sites. Earth Syst. Sci. Data 14, 5157–5178 (2022).
Miller, S. D. et al. Upper atmospheric gravity wave details revealed in nightglow satellite imagery. Proc. Natl Acad. Sci. USA 112, E6728–E6735 (2015).
Kyba, C. C. M., Ruhtz, T., Fischer, J. & Hölker, F. Cloud coverage acts as an amplifier for ecological light pollution in urban ecosystems. PLoS ONE 6, e17307 (2011).
Ściężor, T. Effect of street lighting on the urban and rural night-time radiance and the brightness of the night sky. Remote Sens. 13, 1654 (2021).
UN Environment Programme. The Rapid Transition to Energy Efficient Lighting: An Integrated Policy Approach (United Nations Environment Programme, 2013); https://www.unep.org/resources/report/rapid-transition-energy-efficient-lighting-integrated-policy-approach
Kocifaj, M. & Barentine, J. C. Air pollution mitigation can reduce the brightness of the night sky in and near cities. Sci. Rep. 11, 14622 (2021).
Stark, H. et al. City lights and urban air. Nat. Geosci. 4, 730–731 (2011).
Smith, M. Time to turn off the lights. Nature 457, 27–27 (2009).
Pandey, B., Zhang, Q. & Seto, K. C. Comparative evaluation of relative calibration methods for DMSP/OLS nighttime lights. Remote Sens. Environ. 195, 67–78 (2017).
FLUXNET2015 dataset. FLUXNET (accessed 12 January 2024); https://fluxnet.fluxdata.org/data/fluxnet2015-dataset/
Pesaresi, M. et al. Advances on the global human settlement layer by joint assessment of earth observation and population survey data. Int. J. Digit. Earth 17, 2390454 (2024).
R Core Team. R: A language and environment for statistical computing. (R Foundation for Statistical Computing, 2024); https://www.r-project.org/ (2024).
Johnston, A. S. A., Kim, J. & Harris, J. A. Widespread influence of artificial light at night on ecosystem metabolism: data and R script. Figshare https://doi.org/10.6084/m9.figshare.29958455 (2025)
Acknowledgements
We thank Z. Nasar, D. Simms and W. Rust for helpful discussions on the original idea for this manuscript. This work was supported by a Natural Environment Research Council grant (grant no. NE/W003031/1 to A.S.A.J. and J.A.H.).
Author information
Authors and Affiliations
Contributions
A.S.A.J. conceived the idea and designed the methodology, acquired and curated data, developed the code, performed all analyses and data visualization and wrote the original draft and revisions. J.A.H. contributed to writing and editing of the manuscript, and J.K. contributed to data acquisition. All authors, including A.S.A.J., J.K. and J.A.H., discussed the methods and results and approved the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Climate Change thanks Gesa Weyhenmeyer, Xuanzhou Zhang and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Global distribution of FLUXNET2015 sites across artificial light intensity gradients.
White symbols indicate FLUXNET2015 sites within the latitudinal range of the harmonised nighttime light map (n = 203, see Methods). Panel a) shows number of sites and b) shows artificial light at night (ALAN) digital number (DN) averaged over site years, plotted against latitude. Bar and symbol colours in a) and b) indicate low (DN < 10), medium (DN ≥ 10 ≤ 30) and high (DN > 30) ALAN intensities. Sites for analysis in this study were selected based on replication of FLUXNET2015 sites across high, medium and low ALAN intensities (Fig. 1 in the main text, see Methods). Basemap generated with QGIS using the harmonized global nighttime light dataset32 under a Creative Commons license CC BY 4.0.
Extended Data Fig. 2 Residual diagnostics for unweighted and weighted GAMMs of half-hourly carbon fluxes.
Final unweighted GAMMs (red symbols and lines) and weighted GAMMs (black symbols and lines) were fitted to half-hourly flux observations (see Supplementary Table 4) for a, b) NEE, c, d) GPP, e, f) Re. The left-hand panel (a, c, e) show GAMM residuals plotted against mean fitted values, with fitted loess lines. The right-hand panel (b, d, f) show residual variance by fitted value bins, indicating a reduction in heteroscedasticity across the range of predicted values in the weighted GAMMs (black symbols and lines), presented in Fig. 3 and Supplementary Table 5.
Extended Data Fig. 3 Partial effect surfaces and residual autocorrelation for half-hourly GAMMs.
Final weighted half-hourly GAMMs for a, b) NEE, c, d) GPP and e, f) Re are shown. Partial effect surfaces (a, c, e) display the estimated tensor-product smooth interaction between ALAN and night duration, as derived from the final GAMMs in Supplementary Table 5. Panels b, d and e show partial autocorrelation functions of the model residuals (at hourly lags), indicating the degree of remaining temporal autocorrelation after model fitting. Dashed blue lines represent approximate 95% confidence intervals for the null hypothesis of white noise.
Extended Data Fig. 4 Predicted conditional smooths of daily GAMMs for carbon fluxes during growing and non-growing seasons.
Panels show: a, b) NEE, c, d) GPP and e, f) Re in the growing season (a, c, e) and non-growing season (b, d, f) (Supplementary Table 9). Smooth curves illustrate the tensor-product interaction between ALAN and night duration (ND, coloured lines: 9, 12 and 15 h, with shaded ribbons representing mean predictions ±95% confidence intervals).
Extended Data Fig. 5 Partial effect surfaces and residual autocorrelation for daily GAMMs of carbon fluxes.
Final variance-weighted daily GAMMs for a, b) NEE, c, d) GPP and e, f) Re are shown. Partial effect surfaces (a, c, e) display the estimated tensor-product smooth interaction between ALAN and night duration, as derived from the final GAMMs in Supplementary Table 9. Panels b, d and e show partial autocorrelation functions of the model residuals (at day of the month lags), indicating the degree of remaining temporal autocorrelation after model fitting. Dashed blue lines represent approximate 95% confidence intervals for the null hypothesis of white noise.
Extended Data Fig. 6 Balanced dataset GAMM predictions of half-hourly and daily carbon flux responses to ALAN and night duration.
Variance-weighted GAMMs were fitted to the balanced ALAN intensity dataset, comprising an equal number of sites (n = 4 each in low, medium, and high ALAN intensity categories per continent). Panels (a–c) show predicted relative changes in half-hourly (a-c) and mean daily (d-f) carbon fluxes for NEE (a, d), GPP (b, e), Re (c, f) as a function of ALAN (DN) intensity. Coloured lines represent night duration (ND) categories of 9, 12, and 15 h, with shaded ribbons representing mean predictions ±95% confidence intervals. The horizontal dashed line denotes zero relative change.
Extended Data Fig. 7 Distributions of mean daily carbon fluxes in the balanced versus full datasets.
The balanced dataset represents equal distribution of sites in ALAN intensity groups (n = 34,351 across 24 sites) compared to the full dataset (n = 231,598 across 86 sites). Boxplots for (a, b) NEE, (c, d) GPP, and (e, f) Re are stratified by growing season (N: non-growing season, Y: growing season) and grouped according to (left: a, c, e) ALAN intensities or (right: b, d, f) Night Duration (ND). Boxes represent interquartile ranges (IQR), horizontal lines denote medians, whiskers extend to 1.5 × IQR, and points indicate outliers.
Extended Data Fig. 8 Comparison of annual carbon fluxes and correlations with environmental variables in full and balanced datasets.
Panels show annual: a) NEE, b) GPP and c) Re grouped by ALAN intensity. Boxes (a-c) represent interquartile ranges (IQR), horizontal lines indicate medians, whiskers extend to 1.5 × IQR, and points denote outliers for the balanced dataset (with equal representation of sites across ALAN intensity groups, n = 120) and the full dataset (n = 605). Correlation matrices between annual environmental and urban variables with carbon fluxes are shown for (d) the balanced dataset and (e) the full dataset. Pearson correlation coefficients are shown, with colour intensity indicating the strength (lighter: weaker, darker: stronger correlation) and direction (red: positive, blue: negative, white: none) of correlations.
Extended Data Fig. 9 Sensitivity analysis of annual structural equation models fitted to the balanced dataset.
Panel (a) illustrates the original structural equation model (SEM) structure (Fig. 5, fitted to the full dataset). The balanced dataset yielded an acceptable SEM fit to the original model in (a) Fisher’s C = 25.05, P = 0.199, df = 20, n = 120, AIC = 4,939, R2m = 0.64, R2c = 0.71 (NEE) but interactions between GPP ~ SW + DtNUP, Re ~ T + VPD, and NEE ~ GS Length were no longer significant (p > 0.05, red lines in a). Retained pathways (blue lines in a) were fitted to the balanced dataset alongside Re ~ GS Length, which was identified as a missing pathway in the balanced SEM. Standardized path coefficients are shown for the balanced SEM in (b), adjacent to arrows with line thickness indicating the relative strength of relationships. Significance levels for path coefficients (solid black arrows in b), p < 0.0001: ****, p < 0.001 ***, p < 0.01 **, p < 0.05 *) are based on two-sided tests, with no multiple comparison adjustments. The refitted SEM to the balanced dataset (b) had a better global fit (SEM diagnostics shown in plot) than (a) but yielded a higher AIC due to the original SEM in (a) having better explanatory power for Re (a: R2m = 0.51; b: R2m = 0.38).
Supplementary information
Supplementary Information (download PDF )
Supplementary Tables 1–11.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Johnston, A.S.A., Kim, J. & Harris, J.A. Widespread influence of artificial light at night on ecosystem metabolism. Nat. Clim. Chang. 15, 1371–1377 (2025). https://doi.org/10.1038/s41558-025-02481-0
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41558-025-02481-0
This article is cited by
-
Artificial light reduces ecosystem carbon sinks
Nature Climate Change (2025)
