
Abstract
Mammals have evolved a more complex brain, exemplified by the transformation of the single-layer dorsal cortex of excitatory projection neurons (ExNs) in ancestors into a multilayered cerebral neocortex1,2,3,4 enriched with diverse intratelencephalic and extratelencephalic ExN subtypes5,6,7, thereby establishing specialized projection systems that enhance brain connectivity and functionality5,6,7,8. This is in contrast to modern reptiles and birds with single-layered or pseudolayered columnar organization of ExNs4,9,10,11,12. However, the mechanisms underlying these mammalian-specific adaptations remain elusive. By comparing the landscape of gene expression and putative cis-regulatory elements (CREs) in mouse ExN subtypes and through cross-species examination, we identified mammalian-specific CREs, including a subset bound by the transcription factor ZBTB18 (also RP58, ZFP238 or ZNF238) and associated with genes defining intratelencephalic and extratelencephalic subtypes and connectivity, which have been implicated in intellectual disability and autism. Deletion of Zbtb18 in mouse ExNs dysregulated target gene expression, reduced molecular diversity, diminished cortico-spinal and callosal projections and increased intrahemispheric cortico-cortical association projections to the prefrontal cortex, thereby resembling non-mammalian brain. ZBTB18 binding motifs are highly enriched in callosally projecting intratelencephalic-biased putative CREs and show higher conservation specifically in mammals. This study uncovers critical components and mammalian-specific evolutionary adaptations within a regulatory node essential for neocortical ExN identity and connectivity.
Similar content being viewed by others

Main
Previous studies identified transcription factors (TFs) that guide subtype specification, laminar positioning and connectivity of mammalian neocortical ExNs5,6,7; however, the evolutionary adaptations and precise molecular mechanisms remain elusive. Here we uncovered gene regulatory subcircuits, particularly involving ZBTB18–CRE interactions that govern key features of neocortical ExNs that have undergone modifications in the mammalian lineage.
Subtype-specific CREs and TFs in neocortical ExNs
To characterize CREs and TFs for neocortical ExNs, we used Arpp21–Gfp or Fezf2–Gfp transgenic mice and enriched GFP-expressing neocortical upper layer (L2–4) intratelencephalic (IT) neurons or deep layer (L5–6) predominantly extratelencephalic (ET) neurons, respectively (Fig. 1a and Supplementary Fig. 1), from neonatal mice (postnatal day (PD) 0), an age at which neocortical ExN identity and connectivity are established. Cells were processed for RNA sequencing (RNA-seq) and chromatin immunoprecipitation (ChIP) assays with DNA sequencing (ChIP–seq) for H3K27ac13,14,15,16,17 (Fig. 1b).

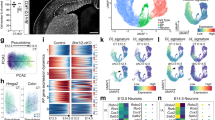
a, Immunolabelling for Arpp21–Gfp+ IT (IT neurons onwards) and Fezf2–Gfp+ ET (ET neurons onwards) neurons and BCL11B in mouse PD 0 neocortex. b, Schematic for isolation and processing of Arpp21–Gfp-positive and Fezf2–Gfp-positive cells from neocortex for RNA-seq and H3K27ac ChIP–seq. c, TFBS for 69 TFs enriched among both IT-biased H3K27ac peaks (red circle) and H3K27ac peaks near IT-biased genes (blue circle) compared with ET-biased peaks and genes, respectively. TFBS enrichments were tested using Fisher’s exact test. Significant TF motifs had a Benjamini–Hochberg-corrected P value < 0.05 and odds ratio > 1. For RNA-seq, n = 3 biological replicates per condition. For ChIP–seq, n = 2 biological replicates per condition. d, Schematic showing two dorsal pallial regions (H and M) microdissected from E17 chicken embryo. e, Venn diagram showing the co-occurrence of peaks derived from ZBTB18-HA ChIP–seq and H3K27ac peaks preferably found in IT and ET neurons and in chicken dorsal pallium. X are peaks in both IT and ET neurons excluded from analysis. f, Heat map showing pairwise alignment distances among vertebrates for six putative CREs overlapping ZBTB18 ChIP–seq peaks linked to IT neurons and axon guidance. The left grid indicates ChIP–seq peak identification method. Columns on the right show if the regions (H3K27ac or ZBTB18 peaks) are orthologous and active in the chicken embryonic dorsal pallium. Grey columns mark non-orthologous regions; red and black columns show the presence or absence of H3K27ac peaks in orthologous regions. Distances are relative to mice. g, Dot plot of Zbtb18-expressing neurons in the dorsal pallium showing cell percentages and co-expression of genes with the ZBTB18 binding motif in IT neurons across mammalian (mouse) and non-mammalian species (chicken, lizard and turtle). h,i, Coronal sections of chicken (h) and mouse (i) brains showing co-localization of BCL11B and SATB2 with ZBTB18. j, Bar plots showing the co-localization percentage for BCL11B and ZBTB18 over total BCL11B+ cells and SATB2 and ZBTB18 over total SATB2+ cells in chicken and mouse. A standard unpaired two-tailed t-test was applied. The graph shows the mean ± s.e.m. **P = 0.0011; ***P = 0.0004 (n = 3 per species). E, entopallium; fL, fetal (immature) layer; H, hyperpallium; HA, apical hyperpallium; Hp, hippocampus; M, mesopallium; N, nidopallium; RPKM, reads per kilobase of transcript per million mapped reads; Str, striatum. Scale bars, 100 µm (a), 1 mm (h,i (mouse brain)), 150 µm (h,i (mouse inset)), 1 mm (h,i (chicken brain)), 300 µm (h,i (chicken inset)), 250 µm (h,i (Cux2)).
Commensurate with the shared developmental lineage of IT and ET neurons, most of the expressed genes (15,120 of 15,902) and the genomic regions enriched for H3K27ac (peaks) (35,173 of 62,448) were common to both cell types (Supplementary Tables 1 and 2). Gene Ontology analysis of the genes and peaks enriched specifically in either IT (304 genes and 54 peaks) or ET (478 genes and 73 peaks) neurons (Extended Data Fig. 1a–c and Supplementary Tables 1–3) revealed terms broadly relevant for IT and ET neurons, respectively (Extended Data Fig. 1d and Supplementary Tables 4–7).
To prioritize TFs for IT and ET neuron development, we identified those highly and differentially expressed and with TF binding sites (TFBSs) enriched among H3K27ac peaks specific to Arpp21–Gfp+ cells (222 TFs; Fig. 1c and Supplementary Table 8). We also highlighted TFBS proximal to genes upregulated in the same cells (154 TFs) compared with Fezf2–Gfp+ cells, and vice versa (Fig. 1c and Supplementary Table 9). Some prioritized TFs are implicated in the development of IT and ET neurons5,6,7, including Meis2, Rara, Rarb and Rxrg, which mediate retinoic acid signalling during the prefrontal cortex (PFC) development18. In Arpp21–Gfp+ cells, 69 TFs had binding motifs enriched in ChIP–seq and showed enhanced expression in RNA-seq datasets. Among these, ZBTB18 was the most highly expressed TF (Fig. 1c and Supplementary Table 8). In Fezf2–Gfp+ cells, 54 TFs had motifs enriched and showed increased expression, with NR2F1 being the most highly expressed (Fig. 1c and Supplementary Table 9). Through integrated analysis, we discovered both shared and cell-type-specific TFs and putative CREs. Notably, our findings highlighted ZBTB18, prompting us to investigate its regulatory network and functions in developing neocortical ExNs.
ZBTB18 targets mammalian-specific ExN enhancers
We sought to validate ZBTB18 binding sites in the developing mouse neocortex using ChIP–seq. Because our trials with commercially available antibodies did not yield high-quality ChIP–seq findings, despite working well for immunostaining (Extended Data Fig. 4a), we conducted ChIP–seq by expressing HA-tagged mouse Zbtb18 plasmid electroporated into PD 0 neocortical ExNs. This revealed that ZBTB18 binds to putative developmental CREs marked by H3K27ac peaks near crucial genes previously implicated in the development and diversification of ExNs, such as Bcl11b, Cux2 and Satb2 (Supplementary Table 11).
We compared mouse ZBTB18 ChIP–seq and H3K27ac peaks with H3K27ac peaks from chicken, a non-mammal (Fig. 1d and Supplementary Table 10). On the basis of multispecies transcriptome datasets4,10,11, we performed this analysis on microdissected chicken hyperpallium and mesopallium regions within the dorsal pallium, considered homologous to the mammalian neocortex, at embryonic day 17 (corresponding to mouse age of PD 0). We identified ten distinct enhancer or promoter peaks bound by ZBTB18 that were also biased to IT neurons in mice but not in chicken (Fig. 1e and Supplementary Table 11). Among these ten peaks, five were in proximity to crucial genes differentially expressed between IT and ET neurons and implicated in their development5,6,7: Cux2 (Cux2 enhancer (E) E1 and Cux2-E4), Satb2 (Satb2-E1), Robo1 (Robo1-E1), and Bcl11b (Bcl11b promoter (P) P1) (Fig. 1e–f and Extended Data Figs. 2a–c and 9a). Within the ZBTB18-bound peak associated with mouse Cux2-E1, no overlapping H3K27ac peaks were detected in the orthologous region in chicken, suggesting that this region serves as an active enhancer in mouse cortex but not in chicken (Fig. 1f and Extended Data Fig. 2a). No orthologous regions or overlapping H3K27ac peaks of mouse Cux2-E4 and Satb2-E1 were seen in chicken (Fig. 1f and Extended Data Fig. 2a,b).
The ZBTB18-bound peak within the mouse Bcl11b-P1 did not have an orthologous region in chicken (Fig. 1f and Extended Data Fig. 2c). We did not identify an orthologous region for mouse Robo1-E1 in chicken or any other non-mammalian species analysed (Fig. 1f and Extended Data Fig. 9a), indicating they lack this putative enhancer. We also identified ZBTB18-bound H3K27ac peaks within corresponding regions in mouse ExNs and chicken dorsal pallium, including promoter region of Zbtb18 (Extended Data Fig. 2d), suggesting a potential conserved autoregulatory role. These findings also indicate that specific ZBTB18-bound neocortical ExN subtype-biased enhancers exhibit distinct mammalian features.
For further review of ZBTB18-interacting regions, we selected Cux2-E1 and Satb2-E1 that contained ZBTB18 binding sites in all analysed mammals. Either lacked H3K27ac peaks, or regions associated with ZBTB18-bound peaks were absent in chicken (Extended Data Fig. 2a,b). In humans, the putative enhancers identified for CUX2 and SATB2 exhibited H3K27ac enrichment in fetal dorsolateral PFC (dlPFC)15, suggesting that these enhancers are conserved in the mammalian developing neocortex (Extended Data Fig. 2a,b). Additionally, we analysed co-expression patterns of Zbtb18 with genes that have ZBTB18-bound peaks in IT neurons (Fig. 1e) using publicly available, single-cell RNA-seq datasets for mouse cortex and dorsal pallium of birds, lizards and turtles12,19,20,21. Among these genes, Zbtb18 exhibited the highest co-expression with Cux2, Satb2 and Robo1 in mice but not in non-mammalian species (Fig. 1g). Conversely, in chickens, lizards and turtles, Zbtb18 showed the highest co-expression with Bcl11b, an ET-specific TF in mammals, suggesting a stronger association with IT neurons in mammals but potentially less ExN class selectivity earlier in evolution (Fig. 1g).
Further, using independently validated anti-ZBTB18 antibodies (Extended Data Fig. 4a), we conducted immunolabelling on ExNs in mouse. ZBTB18 exhibited strong co-localization with IT-enriched TFs SATB2 and Cux2 but not with BCL11B (Fig. 1i,j and Extended Data Fig. 4b). Analysis of embryonic chicken pallium revealed regionally extensive ZBTB18 expression, with a higher density of labelled nuclei in the apical hyperpallium, particularly the interstitial apical hyperpallium (IHA), the main sensory input subregion of the apical hyperpallium, containing neurons molecularly resembling mammalian neocortical thalamorecipient IT ExNs11 (Fig. 1h and Extended Data Fig. 1e). Consistent with previous analyses in embryonic chick pallium4,11, we observed stronger expression of BCL11B and Fezf2 in the apical hyperpallium than in the mesopallium region, whereas SATB2 was expressed predominantly in the mesopallium region (Extended Data Fig. 1e). In contrast to the developing mouse neocortex, ZBTB18 exhibited increased co-localization with BCL11B rather than SATB2 in the apical hyperpallium, IHA and adjacent mesopallium region in chicken pallium (Fig. 1h–j and Extended Data Fig. 1e). We did not detect appreciable Cux2 expression in most regions of the dorsal pallium, except the posterior ventral pallium (Fig. 1h and Extended Data Fig. 1e). Analysis of single-cell RNA-seq datasets12,19,20,21 also revealed little or no appreciable Cux2 expression in Zbtb18-expressing neurons within the dorsal pallium of birds, lizards and turtles (Fig. 1g). Therefore, in mammals, ZBTB18-bound CREs may regulate expression of genes, such as Cux2, Satb2 and Robo1, which are not active or are absent in non-mammalian species, suggesting that the ZBTB18-dependent gene regulatory network contributes to the enhanced diversity and connectivity of mammalian neocortical ExNs.
ZBTB18 regulates mammalian Cux2 neocortex enhancer
Because Cux2 is IT neuron-specific in mice, with minimal expression in embryonic chicken dorsal pallium (Fig. 1h and Extended Data Fig. 1e), and Cux2 H3K27ac peaks in IT neurons were prominent (Extended Data Fig. 2a), we selected Cux2 as a potential target. To validate the enhancer activity, we generated multiple transgenic founders, in which Cux2-E1 was placed 5′ to the human BGN promoter and linked with a Gfp reporter gene. We observed GFP expression in the forebrain by post-conception day (PCD) 14.5, recapitulating the native expression of Cux2 (refs. 22,23) (Extended Data Fig. 3a). By PCD 16.5, forebrain GFP expression was restricted to a subset of neurons in the neocortical plate (Fig. 2a,b) and co-localized with ZBTB18 in the neocortex but not in striatum (Fig. 2b). Consistent with the importance of Cux2-E1 for gene expression broadly in the neocortex and specifically in IT neurons, we found that the Cux2-E1–Gfp expression was predominantly co-localized with IT neuron marker SATB2 but not with ET neuron marker BCL11B at PCD 16.5, 17.5 and PD 0 (Fig. 2c,d and Extended Data Fig. 3b,c). By PD 15, GFP-labelled cells were in L2–L4 and mostly immunopositive for CUX1 but not BCL11B (Extended Data Fig. 3d–f). Further, we observed GFP in the corpus callosum but not in the cortico-spinal tract when surveyed at the pontine region on the ventral surface of the brain at PD 0 (Extended Data Fig. 3c). In contrast to the specificity of Cux2-E1, two putative Cux2 enhancers (Cux2-E2 and Cux2-E3) were active in both IT and ET neurons and lacked ZBTB18 ChIP–seq peak (Extended Data Fig. 3g,h). As expected, neither was able to drive expression selectively in IT neurons but instead expressed mainly in RELN+ L1 neurons and LHX6+ interneurons, respectively (Extended Data Fig. 3g,h).

a, Cux2-E1–Gfp expression in the PCD 16.5 neocortex. b, Cux2-E1–Gfp expression co-localizes with ZBTB18 (closed arrows) in the neocortex. c,d, At PCD 16.5, Cux2- E1–Gfp co-expression with SATB2 (closed arrows) but not with BCL11B (c). Double open arrowheads and triple open arrowheads indicate SATB2+ and BCL11B+ cells, respectively. An unpaired two-tailed t-test was used to detect differences between groups (d). The graph shows the mean ± s.e.m.; n = 349 cells from four biological replicates, *P = 0.0001. e, Line graphs showing the H3K27ac peaks from the IT neurons, ET neurons and ZBTB18-HA ChIP–seq peaks associated with mouse Cux2. f, Luciferase activity of the Cux2-E1 enhancer with ZBTB18. An unpaired two-tailed t-test was applied. The graph represents mean ± s.e.m. *P = 0.000040, 0.00029 and 0.000057 (ZBTB18, POU3F2 and HDAC2+ SIN3A, respectively). g, Luciferase activity of the Cux2-E1 enhancer with ZBTB18 from different species. Ordinary two-way analysis of variance (ANOVA) with Bonferroni’s multiple comparisons test was applied. The graph represents mean ± s.e.m. (n = 6). P values are listed in Supplementary Table 16. h, Luciferase reporter assay comparing the activity of wild-type and ZBTB18-binding site-deleted Cux2-E1 (Δ) enhancer constructs from human and mouse with ZBTB18. The graph represents the mean ± s.e.m. (n = 4 per condition). Two-way ANOVA with Bonferroni’s multiple comparisons correction was applied. P values are listed in Supplementary Table 16. i, Cux2-E1–Gfp expression in Zbtb18fl/+ and Zbtb18fl/fl brains electroporated with pCag–Cre and p-Calsl–Rfp plasmids. Co-localization shown with open (in Zbtb18fl/+) and closed (Zbtb18fl/fl) arrows. j, Activity of the mouse Cux2-E1–Gfp or Cux2-ΔE1–Gfp enhancer in E7 chicken hyperpallial neurons. The graph represents GFP intensity normalized to DAPI; n = 3 (control), 42 (Cux2-E1–Gfp) and 51 (Cux2-DE1–Gfp) cells from three independent experiments. Ordinary two-way ANOVA with Bonferroni’s multiple comparisons test, with single pooled variance, was applied. The graph represents mean ± s.e.m. ***P = 0.0001; **P = 0.0048; *P = 0.0430. k, Reverse transcription–polymerase chain reaction (RT–PCR) for Cux2 and Bcl11b at PCD 14.5 and PD 0. Ordinary two-way ANOVA with Bonferroni’s multiple comparisons test, with single pooled variance, was applied. The graph represents mean ± s.e.m.; n = 3 per condition per time point. *P = 0.0327 and 0.0178 (wild type versus Zbtb18 cKO at E15.5 and PD 0); P = 0.0001 (wild type versus Zbtb18 cKO at E15.5 and PD 0). l,m, Distribution of CUX1 (l) or SATB2 (m) and BCL11B is shown in Neurod6–Cre, Zbtb18 cKO and control brains at PD 1. Quantification of cell-type laminar distribution from l and m, respectively. Standard t-tests were used for cell counts per bin. Mean ± s.e.m. for each bin. For P values, see Supplementary Table 16. For RNA-seq and RT–PCR, n = 3 per time point. For immunofluorescent analyses, we counted neurons from independent sections (n = 3 per condition). CP, cortical plate; h, human; IUE, in utero electroporation; IZ, intermediate zone; lCP, lower cortical plate; m, mouse; SP, subplate; SVZ, subventricular zone; uCP, upper cortical plate; VZ, ventricular zone. Scale bars, 1 mm (a), 100 µm (b,l,m), 50 µm (c,i), 10 µm (j).
We conducted luciferase assays to identify the TFs responsible for neocortical Cux2-E1 activity. Several TFs expressed in the developing mouse and human forebrain, including ZBTB18, exhibit predicted binding motifs. Of the tested TFs, ZBTB18 and POU3F2 increased the Cux2-E1 activity, and a combination of HDAC2 and SIN3A repressed it (Fig. 2f). This is consistent with previous findings that POU3F2 has a conserved function in neocortical progenitors24 and potentially interacts with ZBTB18, which was shown to have dual roles in gene regulation25. Combined with ChIP–seq (Fig. 2e) demonstrating ZBTB18 binding to Cux2-E1, these results indicate that ZBTB18 acts as a direct activator of Cux2. Because Zbtb18 and Cux2 are highly co-expressed in mice and humans (Extended Data Fig. 4), but not in non-mammalian species (Fig. 1g), to assess whether this function is specific to mammals, we generated and tested luciferase constructs containing orthologous Cux2-E1 sequences from human, chimpanzee, macaque, mouse, opossum and chicken. Notably, only the sequences from placental mammals, but not those from opossum or chicken, exhibited a significant increase in luciferase activity upon ZBTB18 co-expression (Fig. 2g), indicating that this activation is an evolutionary adaptation specific to placental mammals. When the ZBTB18 binding site from human and mouse Cux2-E1 was removed (hCux2-ΔE1 and mCux2-ΔE1), their ZBTB18-induced luciferase activity decreased (Fig. 2h), indicating that ZBTB18 motif contributes to this placental mammal-specific activation.
To determine the potential of ZBTB18 for transactivation of Cux2-E1 in vivo, we co-electroporated a Cux2-E1–Gfp plasmid, a CALSL–Rfp reporter plasmid and a CAG–Cre plasmid into mice carrying homozygous (fl/fl) or heterozygous (fl/+) floxed Zbtb18 alleles. Electroporation was performed at PCD 15.5 to predominantly target IT neurons. At PD 0, we observed co-localization of GFP and RFP in upper-layer neurons in the Zbtb18fl/+ brain. By contrast, we observed RFP without commensurate GFP expression in electroporated Zbtb18fl/fl cells (Fig. 2i), indicating ZBTB18 is required for Cux2-E1 transactivation in vivo.
To further investigate whether ZBTB18 activates mammalian Cux2-E1 through its motif, we examined its activity in non-mammalian embryonic chicken hyperpallium neurons expressing ZBTB18 by introducing the Cux2-E1–Gfp plasmid into them. Mouse Cux2-E1 drove robust GFP expression in ZBTB18-immuno-positive chicken hyperpallial neurons, whereas deleting the ZBTB18 binding site (Cux2-ΔE1) resulted in a significant reduction in GFP expression (Fig. 2j). These findings, together with the luciferase data (Fig. 2g,h), demonstrate that Cux2-E1 regulation by ZBTB18 in mammalian cortical ExNs is probably motif-dependent. Consistent with Cux2 being enriched in mammalian IT neurons22 and controls their neurite growth26, we observed that overexpression of mouse CUX2 in chicken ZBTB18 and SATB2 co-immunopositive ExNs increased neurite outgrowth (Extended Data Fig. 3i,j), similar to the reported role of CUX2 in mammalian IT ExNs26.
We identified a consensus ZBTB18 binding site within Satb2-E1 (Extended Data Fig. 2b) and performed luciferase assays by co-expressing Zbtb18 with DNA constructs for either Satb2-E1 or Satb2-ΔE1 (lacking the ZBTB18 site). Zbtb18 markedly increased Satb2-E1 activity, whereas luciferase activity from Satb2-ΔE1 constructs was reduced but not to basal levels (Extended Data Fig. 3k). We further revealed overlaps of putative CREs Cux2-E1, Satb2-E1, Bcl11b-P1, Zbtb18-P1 and Robo1-E1 in both human and developing mouse mammalian neocortex by analysing an independent H3K27ac ChIP–seq database from human midfetal dlPFC15 (Extended Data Figs. 2a–d and 9a), indicating functional conservation of these CREs in mammals and their potential involvement in fundamental neocortical developmental processes. These results highlight the pivotal role of ZBTB18 in regulating these CREs, underscoring the need for a comprehensive investigation.
Enrichment of ZBTB18 in postmigratory IT neurons
We examined ZBTB18 expression at the postmigratory stage, a stage previously shown to be critical for ExN specification5,6,7. It is co-expressed with CUX2 in upper-layer IT neurons in the mouse at PD 0 (Fig. 1i,j). This pattern was replicated in post-conception week (PCW) 20 human neocortex, where strong immunolabelling for ZBTB18 was found in nuclei positive for SATB2 and CUX2 within the upper-layer prospective IT neurons (Extended Data Figs. 4c and 5b,c). However, ZBTB18 immunolabelling was diminished in the BCL11B-immunopositive nuclei of prospective L5B ET neurons (Fig. 1i,j and Extended Data Fig. 5b,c). Analysis of a publicly available mouse single-cell RNA-seq dataset27 revealed Zbtb18 expression was higher in developing IT neurons than in L5 and L6 ET neurons (Extended Data Fig. 5a). The enrichment and sustained ZBTB18 expression in IT neurons suggest its crucial role in their specification.
ZBTB18 regulates postmitotic ExN subspecification
Previously, the role of ZBTB18 in neurogenesis and neuronal migration has been reported28,29,30,31,32, and mutations in the human ZBTB18 gene are linked to intellectual disability and autism33,34,35,36,37. To investigate the effects of Zbtb18 on ExN specification, we tested the expression of genes enriched in IT neurons versus ET neurons following Zbtb18 KO. We performed RNA-seq on Zbtb18−/− (KO) and Zbtb18+/− (control) neocortex at PCD 14.5, a time coinciding with the generation of upper-layer IT neurons (Extended Data Fig. 6a and Supplementary Table 12). Of genes enriched in IT or ET neurons and differentially regulated in Zbtb18 KO at PCD 14.5, disproportionally more genes enriched in IT neurons were downregulated following KO (82.3%), as opposed to the upregulated genes. The opposite trend was observed with ET neuron-enriched genes in Zbtb18 KO (92.4% were upregulated) (Extended Data Fig. 6a and Supplementary Table 12). To test whether ZBTB18 is required for ExN specification postmitotically, we conditionally deleted Zbtb18 in these neurons by crossing Zbtb18fl/fl mice to Neurod6–Cre (also known as Nex1–Cre) transgenic mice and conducted RNA-seq on neocortical tissue derived from Neurod6–Cre, Zbtb18fl/fl (Zbtb18 conditional knockout (cKO)) and Neurod6–Cre; Zbtb18fl/+ (control) mice at PD 0 (Extended Data Fig. 6b and Supplementary Table 13). Similarly, at PD 0, 58.7% of genes enriched in IT neurons and differentially expressed in Zbtb18 cKO were downregulated, whereas 96.7% of genes enriched in ET neurons were upregulated (Extended Data Fig. 6b and Supplementary Table 13). Reduced expression of key IT neuron marker genes within the neocortical plate of Zbtb18 KO and cKO mice, including Cux1, Cux2, Rorb and Satb2, was confirmed by multiple assays (Fig. 2k and Extended Data Fig. 6c,h). Conversely, the ET neuron marker BCL11B expression increased in these same mice (Fig. 2k).
Neurod6–Cre; Zbtb18 cKO mice at PD 0 showed a minor reduction in neocortical size, contrasting with constitutive or Emx1–Cre KO mice28,31 (Extended Data Fig. 6d), but at PD 8, it was substantially smaller than the control (Extended Data Fig. 6d). In Neurod6–Cre; Zbtb18 cKO mice, ZBTB18 protein was detected in neocortical progenitor cells but not in postmitotic cells of the intermediate zone, subplate or cortical plate from PCD 15.5 onwards (Extended Data Fig. 6e), suggesting that the effects observed are not attributable to progenitor-related processes. In Neurod6–Cre; Zbtb18 cKO, CUX2 protein and messenger RNA (mRNA) expression in cortical plate were reduced (Extended Data Fig. 6f–h and Supplementary Fig. 2), similar to Emx1–Cre; Zbtb18 cKO mice and whole-body KO. We also found expansion of the laminar distribution and the number of BCL11B-immunopositive and TBR1-immunopositive neurons. This may occur at the expense of both SATB2-immunopositive and CUX1-immunopositive upper-layer IT neurons at PD 0 (Fig. 2j,k and Extended Data Fig. 7d,e).
We next conducted 5-iodo-2-deoxyuridine (IdU) or 5-chloro-2-deoxyuridine (CIdU) labelling at developmental stages in Neurod6–Cre; Zbtb18 cKO mice and control littermates. At PCD 12.5, we labelled early-born neurons destined for deep layers (L5–6), whereas at PCD 14.5 and 15.5, we labelled later-born neurons primarily destined for L4 or L2–3 (Extended Data Fig. 7a–c). We then analysed the co-localization of IdU or CldU with BCL11B for ET neurons and SATB2 for IT neurons. Following labelling at PCD 12.5, most of IdU-positive nuclei were in the central region of cortical plate in both cKO and control mice, typically where immature L5 neurons resided (Extended Data Fig. 7a). Although there was a significant increase in BCL11B-positive cell numbers and laminar distribution in cKO mice, there was no corresponding increase of cells double positive for IdU and BCL11B (Extended Data Fig. 7a), suggesting that the laminar identity and position of L5 ET neurons are not substantially altered at this age. Following IdU injection at PCD 14.5, in cKO mice, we observed increased proportions of IdU and BCL11B double-labelled nuclei (Extended Data Fig. 7b), although the numbers of IdU-positive and BCL11B-positive cells were greater overall. This suggests that Zbtb18 postmitotic deficiency influences cell fate of late-born nascent neurons. Similarly, we observed a decrease in the percentage of nuclei double positive for CldU and SATB2 following injection at PCD 15.5 in cKO mouse, explained in part by a broader laminar distribution of double-positive neurons (Extended Data Fig. 7c). These results indicate that postmitotic and postmigratory ZBTB18 is required cell autonomously for proper specification of IT neurons, and its deletion leads to misspecification of some IT neurons, including acquisition of certain molecular properties associated with ET neurons.
ZBTB18 regulates mammalian neocortical projections
The above findings suggest that ZBTB18 may also regulate axonal projections, a distinguishing feature of ExN subclasses and crucial for the unique mammalian connectivity. We analysed early postnatal Neurod6–Cre Zbtb18 cKO mice harbouring CAG–CAT–Gfp, a CRE-responsive GFP transgene. The cKO mice exhibited reduced white matter and absent corpus callosum (Fig. 3a). Very few callosal axons reached the midline, and those misrouted ventrally into the septum. Other forebrain commissural projections, including the anterior commissure, were also disrupted, reflecting an overall defect in commissural axon growth (Fig. 3a). Additionally, the internal capsule and different subcerebral projections were significantly reduced (Fig. 3a).

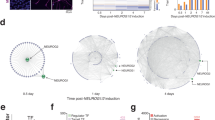
a, Visualization of axonal projections by GFP expression in the Neurod6–Cre; Zbtb18 cKO mouse brain. Full arrows and arrowheads depict wild type, and open arrowheads depict defective tracts in the Zbtb18 cKO brain. b, Whole brain image (left top) and coronal sections of the brain at P7 showing the injection site and extent of the GFP expression after AAvrg–CAG–Gfp injections into the medial PFC at PD 3. c, Left, bar plot showing GFP-positive neurons projecting to PFC in the neocortex of Zbtb18 cKO mice compared with control brains at PD 7, as shown in b. Unpaired two-tailed t-test. The graph represents mean ± s.e.m. *P = 0.03 (n = 3). Right, line graph data showing the percentage distribution of GFP+ neurons projecting to PFC in each bin of the neocortex of Zbtb18 cKO mice compared with wild-type brains at PD 7. The number of labelled neurons in each bin was compared using unpaired t-test. The graph represents mean ± s.e.m. *P = 0.03 (bin 3), 0.004031 (bin 6), 0.03 (bin 7) and 0.012 (bin 14) (n = 3). AC, anterior commissure; AUD, auditory cortex; BLA, basolateral amygdala (excitatory nucleus within amygdala); CC, corpus callosum; CP, cerebral peduncle; cPERI, contralateral perirhinal; CST, cortico-spinal tract; Hip, hippocampus; IC, internal capsule; nLOT, nucleus of lateral olfactory tract; PERI, perirhinal cortex; Str, striatum; Th, thalamus. Scale bars, 1 mm (a), 20 µm (a (CP panel)), 500 μm (b), 125 μm (b (inset)).
To determine the intrahemispheric intracortical connectivity, we injected AAVrg–CAG–Gfp into the PFC of PD 3 mice for retrograde tracing. In control mice, we observed typical intrahemispheric cortico-cortical connectivity originating from transmodal and unimodal association areas, including perirhinal cortex, secondary visual cortex, retrosplenial cortex and parts of the amygdala projecting to PFC (Fig. 3b and Extended Data Fig. 8a,b). However, in Zbtb18 cKOs, this pattern was significantly disrupted: we noted a substantial increase in unilateral afferent connections to the PFC originating from the central region of the neocortex where primary areas are located. Notably, there was a noticeable input reduction from perirhinal cortex, secondary visual cortex and retrosplenial cortex (Fig. 3b and Extended Data Fig. 8a,b). Within neocortex, we also observed an overall increase in GFP-positive cells projecting to PFC (Fig. 3c). Additionally, we observed a reduction in the projections from most of the contralateral inputs and subcortical connections as seen in striatum and thalamus (Fig. 3b and Extended Data Fig. 8b). These results suggest compensatory effects after ZBTB18-mediated loss of callosal and subcortical connections, and collectively demonstrate that ZBTB18 is postmitotically required for the mammalian characteristic neocortical long-range connectivity.
ZBTB18 regulates Robo1 enhancer and callosal wiring
We co-electroporated Neurod1–Cre and CALNL–Gfp reporter vectors into the neocortex of PCD 15 Zbtb18fl/fl mice. This Cre-mediated deletion of Zbtb18 from ExNs resulted in diminished GFP-labelled axons projecting towards the midline at PD 0 (Fig. 4a). Co-electroporation with a Neurod1–Zbtb18 expression vector that restores ZBTB18 in the same cell populations rescued the projection of GFP-positive callosal axons (Fig. 4a), suggesting that ZBTB18 is cell-autonomously required for midline crossing of callosal axons.

a, Coronal sections from PD 0 brain showing ZBTB18 expression and GFP-labelled corpus callosum axons after IUE at PCD 15.5 with Neurod1–Cre and CALNL–Gfp (left) or Neurod1–Zbtb18 (right) plasmids into Zbtb18fl/fl mice. Enlarged panels show GFP images in greyscale. b, Top five Gene Ontology terms for genes that are downregulated (red) or upregulated (blue) in the Zbtb18−/− (KO) and mouse compared with Zbtb18+/− (control) at PCD 14.5 (upper bar plot) and in Neurod6–Cre; Zbtb18 cKO compared with control at PD 0 (lower bar plot). c, Genes encoding axon guidance molecules and upregulated and downregulated in the Zbtb18 KO mouse compared with Zbtb18+/− (control). d, Line graphs showing the H3K27ac peaks from IT neurons, ET neurons and ZBTB18-HA ChIP–seq peaks associated with mouse Robo1. e, Luciferase reporter activity with Robo1-E1 enhancer and ZBTB18. An unpaired two-tailed t-test was performed to detect differences between the control and experimental conditions. The graph represents mean ± s.e.m. **P = 0.00672 (n = 3). f, ISH shows Robo1, Robo2 and Robo3 expressions in the Zbtb18 −/− mouse in the neocortical plate. g–i, Representative images of the IUE with pCag–Robo1 allow Gfp-expressing co-electroporated upper-layer neurons to project GFP-positive axons to and across the corpus callosum (arrows in the inset) analysed at PD 0 (g) and PD 21 (h). Top, number of axons crossing at the midline on the contralateral side of the IUE at PD 21 (i). Bottom, number of cells electroporated on the ipsilateral side (i). An unpaired two-tailed t-test was performed to detect differences between the control and experimental conditions. The graph represents mean ± s.e.m. *P = 0.030. For each IUE experiment, n = 6 (control) and n = 3 (Robo1) animals were analysed at PD 0; n = 3 (control) and n = 3 (Robo1) animals were analysed at PD 21. For RNA-seq, n = 3 per condition. The ISH data shown are representative of the data generated from multiple sections (n = 3 animals). Scale bars, 500 μm (a,g,h), 150 μm (f), 250 μm (g,h (inset)).
We analysed our RNA-seq data from Zbtb18 KO mice to understand the potential downstream effectors. Gene Ontology terms associated with upregulated and downregulated genes at both PCD 14.5 and PD 0 referenced general categories, including nervous system development. Several terms implicating DNA replication and cell cycle were enriched among upregulated genes at PCD 14.5. Axonogenesis, axon guidance and neuron projection development were also enriched among genes downregulated in the Zbtb18 KO at both PCD 14.5 and PD 0 (Fig. 4b). These observations indicate that ZBTB18 not only promotes cell cycle exit and neuronal migration, as previously described28,31,32, but also promotes postmitotic neocortical ExN diversification and axonogenesis. We analysed axon guidance and cell adhesion molecules and found that receptors implicated in callosal development, including members of the Netrin (Dcc and Unc5d38,39,40) and Slit–Robo families, were either downregulated (Robo1 and Robo2; refs. 41,42) or upregulated (Robo3; ref. 43) in Zbtb18 KO neocortex (Fig. 4c). Notably, the Slit receptor Robo1 was the most downregulated. Additionally, ZBTB18 ChIP–seq data revealed an IT neuron-specific putative enhancer Robo1-E1 within the Robo1 locus harbouring a consensus ZBTB18 motif (Fig. 4d and Extended Data Figs. 9a and 11d). Luciferase assays demonstrated significant activation of Robo1-E1 by ZBTB18 (Fig. 4e). In situ hybridization (ISH) at PCD 15.5 confirmed the downregulation of Robo1 and Robo2, as well as the upregulation of Robo3 in the Zbtb18 cKO neocortex (Fig. 4f).
Given these observations and reported callosal defects resulting from disruptions in both mouse and human ROBO1 homologues41,42,44, we electroporated a control (CAG–empty) or Robo1-expression vector (CAG–Robo1) together with Neurod1–Cre and CALNL–Gfp (or CALNL–Rfp) into the PCD 15.5 Zbtb18fl/fl mouse neocortex. We found negligible axons projecting towards or across the midline at PD 0 in any of the six control electroporated Zbtb18fl/fl brains (Fig. 4g). By contrast, we observed recovery of a proportion of GFP+ axons in the corpus callosum following Robo1 overexpression in all electroporated animals (Fig. 4g). Further, we observed similar restoration of a subset of GFP+ axons in the corpus callosum upon overexpression of Robo1 at PD 21, a stage when its development is complete (Fig. 4g,i). The fact that the recovery was not extensive suggests that midline crossing is a complex process, and other factors, such as cell-intrinsic effectors (including DCC and ROBO2), the actions of midline glia8 or a combination of these, may be required, especially in humans and in relation to pathology.
ZBTB18 binding sites conserved in mammalian IT CREs
ExN subtype-specific CREs controlled by ZBTB18 showing either mammal-specific characteristics or unique mammalian ZBTB18 binding sites led us to explore whether this regulatory system might mark an evolutionary milestone. We analysed the protein sequences and dorsal pallial expression of Zbtb18 and found that they are highly conserved across vertebrates (Fig. 1h,i and Extended Data Figs. 1e and 10a). Next, we considered the conservation of broad populations of ZBTB18 binding motifs across placental mammals, marsupials, monotremes and non-mammalian species. We then assessed evolutionary footprints of specific CREs and/or ZBTB18 binding motifs plausibly regulating key genes associated with major subtypes of ExNs, including Cux2, Satb2, Bcl11b and Robo1. However, because variations in TFBSs may lead to interspecies differences13,14,16,17, we sought to determine whether variations are conserved within specific clades.
We assessed the relative conservation of putative ZBTB18 binding sites among putative CREs near genes enriched in IT neurons (Extended Data Fig. 10a) and compared this to putative CREs unrelated to IT or ET neuron subspecifications or corpus callosum formation (Methods). We also considered putative CREs near genes enriched in ET neurons or was upregulated or downregulated in Zbtb18 KO mice. We devised a bootstrap-based strategy to evaluate robustness (Extended Data Fig. 10d and Methods). When precisely matching the core motif as in the JASPAR database (Methods), ZBTB18 binding sites were significantly conserved in IT neuron enhancers in placental mammals and marsupials but not in monotremes or non-mammals (Fig. 5b), an enrichment not observed in ET neuron enhancers. Moreover, we found significant conservation of ZBTB18 binding sites in CREs of genes upregulated in Zbtb18 KO mice specifically throughout Eutheria but no other mammals (Fig. 5a). We also assessed the conservation of ZBTB18 binding sites with one, two or three mutations in the core sequence, reasoning that heavily mutated sites should not exhibit similar enrichment. Expectedly, we found no enrichment among putative IT neuron enhancers or those upregulated in Zbtb18 KO mice when assessed with either two or three mutations, suggesting that the ZBTB18 binding site conservation is specific among IT neurons (Fig. 5a).

a, Conserved ZBTB18 binding motifs are identified in putative CREs associated with genes exhibiting enriched expression in Arpp21–Gfp+ IT neurons. b, Between marsupials and placental mammals, enhancers associated with Arpp21–Gfp+ IT neurons have a significantly higher percentage of conserved ZBTB18 motifs than those associated with Fezf2–Gfp+ ET neurons or background sequences. No differences in ZBTB18 motif conservation in non-mammals or monotremes were detected. Asterisks indicate where Fisher’s exact test revealed a significant enrichment of conservation, with a false discovery rate (FDR)-corrected P value < 0.05. c, Among the TFBS catalogued in the JASPAR database and expressed in either IT neurons or ET neurons, the ZBTB18 motif stands out as one of three motifs with consensus sequences uniquely conserved within enhancers linked to genes enriched in IT neurons across placental mammals and marsupials. Fisher’s exact test was used to assess enrichment of conservation. Red dots represent motifs with FDR-corrected P value < 0.05 (see Supplementary Table 17 for detailed statistics). Mut., mutation.
Considering the possibility that this conservation may be nonspecific, we performed a similar analysis using 267 motifs from the JASPAR database, corresponding to TFs expressed in either IT or ET neurons, or both, at PD 0 according to our RNA-seq dataset. We again assessed enrichment of binding sites with perfect core motif conservation in putative CREs associated with IT or ET neuron-enriched genes or those differentially regulated in the Zbtb18 KO (Fig. 5c). We grouped the significant motifs according to independently tested enrichment in consecutive phylogenetic groups (Placentalia + Marsupalia, Placentalia + Marsupalia + Monotrema, and so on). Of the 267 motifs, only two other motifs (GMEB2 and SMAD2, SMAD3 and SMAD4, involved in transforming growth factor-β signalling) were specifically conserved in IT putative CREs in Theria, demonstrating that ZBTB18 is a member of a rare group of TFs with binding sites that are singularly conserved among placental mammals and marsupials in IT neuron-related CREs (Fig. 5c). This further supports a critical role for ZBTB18 in the evolution of the mammalian diversification of ExNs. Among genes significantly upregulated in the neocortex of Zbtb18 KO mice, the enriched conservation of ZBTB18 motifs was only significant across placental mammals (Extended Data Fig. 10b).
We next examined the motif evolutionary footprints within ZBTB18-bound CREs of critical IT and ET genes, including those we functionally validated to regulate Cux2, Satb2 and Robo1, and the promoter region of Bcl11b and Zbtb18, by performing alignment across 60 vertebrates (Extended Data Figs. 2a–d and 9c). Although motifs within Zbtb18-P1 demonstrate high conservation across vertebrates (Extended Data Fig. 11a), motifs in key IT and ET neuron CREs are eutherian-specific or mammalian-specific (Extended Data Fig. 11b–f). We next inferred the emergence time of these motifs (Extended Data Fig. 10c). Notably, all consensus ZBTB18 motifs in Zbtb18-P1 evolved before the divergence of monotremes from non-mammalian vertebrates (Fig. 1h,i and Extended Data Fig. 1e), suggesting a possible conserved self-regulation mechanism of ZBTB18. By contrast, most motifs in key IT and ET CREs appeared after the divergence of placental mammals from marsupials, suggesting ZBTB18 may have used eutherian-evolved IT and ET CREs to finely tune gene expression.
Finally, we estimated the evolutionary age of ZBTB18 genes by protein sequence similarity and found that, although they are ancient, their protein sequences do not change among chordates (Extended Data Fig. 10a and Methods). These results further suggest that the evolutionary-derived CREs are essential to drive the expression pattern of key genes involved in the subspecification and connectivity of mammalian neocortical ExNs.
Discussion
We addressed a fundamental question at the intersection of neuroscience and evolutionary biology: the emergence of the mammalian neocortex, particularly its diverse ExN subtypes and intricate long-range projections. We revealed mammalian-specific adaptations at the level of CREs linked to TFs associated with ET and IT neuron specification, contributing to the hallmarks of mammalian neocortex evolution. Consistent with these findings, the loss of ZBTB18 function in mice leads to simplified neocortical laminar organization and long-range axonal projection patterns that resemble ancestral forms. This research also expands upon the study of ExNs in mouse piriform cortex (palaeocortex) that revealed diminished ExN subspecification compared with neocortex, with piriform ExNs retaining molecular signatures reminiscent of those shared with reptiles and amphibians45.
Human genetic studies associated ZBTB18 variants with corpus callosum agenesis, microcephaly, autism and intellectual disability35,36,37,46,47. Mutations in the identified target genes, including CUX2, SATB2, BCL11B and ROBO1, are associated with similar disorders36,48,49,50. We revealed that CREs and the expression patterns of these genes in neocortex are conserved across mammalian species and differ from their non-mammalian counterparts. ZBTB18 binding sites are highly conserved in eutherian IT-biased CREs. This heightened evolutionary constraint in mammalian neocortex, although advantageous, may render them more susceptible to various neurodevelopmental and neuropsychiatric disorders. Our research has demonstrated potential predictive value for autism pathogenesis, as individuals with ZBTB18 loss-of-function mutations may show increased cortico-cortical connectivity between PFC and temporal areas, a pattern also seen in functional magnetic resonance imaging studies of autism51,52,53. Although we did not directly examine connectivity within sensory-motor and visual networks, owing to an apparent expansion of temporal heteromodal projections in the Zbtb18 cKO mice, we predict that these areas are hypoconnected in our model. Further, similar to ZBTB18, we identified that its direct target Satb2 also maintains spatially organized connectivity patterns across primary sensorimotor and association neocortical areas in mammals54. These findings highlight how evolutionary and developmental insights relate to neurodevelopmental and psychiatric disorders.
Methods
Mice
All experiments involving animals were approved by the Yale University Institutional Animal Care and Use Committee and conducted in compliance with all relevant university, state and federal guidelines. The day of vaginal plug detection was designated as PCD 0.5. The day of birth was designated as PD 0. Fezf2–Gfp (000293-UNC) and Arpp21–Gfp (011848-UCD) transgenic mice were obtained from the Gene Expression Nervous System Atlas55. Cux2-E1–Gfp, Cux2-E2–Gfp and Cux2-E3–Gfp transgenic mice were generated by delivering linearized DNA constructs by pronuclear injection. Three to seven founders from each line were examined for reproducible GFP expression. Zbtb18 KO mice were generated by the Masai laboratory28, and Zbtb18fl/fl mice were generated by the Heng laboratory. Emx1–Cre (The Jackson Laboratory; 005628) mice, Cux2–Cre mice, Neurod6–Cre (also known as Nex1–Cre) mice and CAG–Cat–Gfp (The Jackson Laboratory; 024636) mice were previously generated and described56,57,58,59 (Cre Driver Network at the National Institutes of Health (NIH) Blueprint for Neuroscience Research). Genotyping primers are shown in Supplementary Table 15.
The mice were provided with food and water ad libitum, maintained on a 12-h light/12-h dark cycle, provided veterinary care by the Yale Animal Resources Center and housed in clean groups. All mice used or bred for these experiments were in good health, as approved by the Yale Animal Resources Center and confirmed through regular veterinary monitoring. To maintain genetic diversity, multiple concurrent breeding pairs were maintained and siblings were never mated. Both males and females were used randomly throughout this study.
Although blinding was not relevant for the primary mutant versus control comparison, other aspects ofthe study required careful design to minimize bias. Randomization was implemented during data acquisition. Littermates (WT, HET and KO) were housed together to avoid confounding housing effects on statistical analyses. The experimental cohort comprised age-matched male and female littermates. Including samples from multiple litters further enhanced reproducibility.
Tissue preparation and fluorescence-activated cell sorting
Neocortices from PD 0.5 Fezf2–Gfp, with GFP-expressing neurons enriched in the deep layer (L5–6) predominantly ET neurons (7), and Arpp21–Gfp, with GFP-expressing neurons enriched in predominantly IT neurons55, were dissected under a dissection microscope and minced with a sterile blade. Tails were collected for determining sex and genotyping. Single-cell suspensions from neocortical tissue were prepared by dissociation with a papain-based solution60 and incubated at 37 °C for 15 min with intermittent trituration using autoclaved, fire-polished glass Pasteur pipettes. Cells were then pelleted by centrifugation at 4 °C for 5 min, washed with sterile 1× phosphate-buffered saline (PBS) and filtered through a 40-μm strainer. Cells were then sorted to collect GFP+ cells by fluorescence-activated cell sorting (FACS) using FACSAria II (BD Biosciences) sorter or Beckman Coulter MoFlo sorter. Hibernate solution (Gibco) supplemented with 2% fetal bovine serum (Gibco) was used to collect FACS-sorted cells. Cell preparations were maintained at 4 °C during the entire process. For RNA-seq, immediately after FACS, cells were pelleted by centrifugation at 350g at 4 °C for 10 min, washed with PBS, pelleted and flash-frozen in liquid nitrogen and stored at −80 °C. For ChIP–seq, FACS-sorted GFP+ cells were pelleted by centrifugation at 350g for 10 min at 4 °C, washed with PBS and immediately crosslinked with formaldehyde solution at a final concentration of 1% for 10 min at room temperature. Glycine (AmericanBio) was added at a final concentration of 125 mM, and samples were incubated for 5 min at room temperature to quench crosslinking. Cells were washed again with PBS, pelleted, flash-frozen in liquid nitrogen and stored at −80 °C.
RNA-seq and initial analysis
Total RNA was extracted from FACS-purified cells or neocortical tissue using TRIzol reagent, according to the manufacturer’s instructions. DNase I (Invitrogen) was added to the extracted total RNA, incubated for 15 min at 37 °C to eliminate DNA contaminants and then inactivated according to the manufacturer’s instructions. RNA concentration and integrity were measured using a NanoDrop spectrophotometer (Thermo Fisher Scientific) and TapeStation 2200 (Agilent). Samples with RNA integrity number of 8 or higher were used for subsequent experiments. Libraries were prepared using the TruSeq Stranded Total RNA with Ribo-Zero preparation kit (Illumina), according to the manufacturer’s instructions. Libraries were quality controlled using the TapeStation 2200 (Agilent) and sequenced on the HiSeq 2000 platform (Illumina) at the Yale Center for Genome Analysis (YCGA) to generate 75-bp single-end reads. Sequencing data were quality controlled using FastQC and aligned to the mouse genome (NCBI38/mm10) using TopHat (v.1.0.13) with up to two mismatches61. An average of 40 million uniquely mapped reads were obtained for each sample. Differential expression analysis was performed using the R package DESeq, and principal component analysis was performed using the R package prompt. Differential expression of transcripts was detected using FDR < 0.01.
ChIP–seq and initial analysis
Pooled GFP+ cells from FACS were used for ChIP–seq. A total of 2.5 × 107 cells per condition were crosslinked with a formaldehyde solution (Sigma-Aldrich) at a final concentration of 1% for 10 min at room temperature. L-Glycine (AmericanBio) was added at a final concentration of 125 mM and incubated for 5 min at room temperature to quench the crosslinking. Cells were washed with PBS and then disrupted using lysis buffer I (50 mM HEPES–KOH (pH 7.5), 140 mM NaCl, 1 M EDTA (pH 8.0), 10% glycerol, 0.5% Nonidet P-40 (NP-40), 0.25% Triton X-100 and 1× protease inhibitor) for 20 min at 4 °C and lysis buffer II (200 mM NaCl, 1 M EDTA (pH 8.0), 0.5 mM EGTA (pH 8.0), 10 mM Tris–HCl (pH 8.0) and 1× protease inhibitor) for 10 min at room temperature. Cells were centrifuged at 300g for 15 min at 4 °C, and pellets were dissolved in 400–600 μl of lysis buffer III (1 mM EDTA (pH 8.0), 0.5 mM EGTA (pH 8.0), 10 mM Tris–HCl (pH 8.0), 0.5% sarkosyl and 1× protease inhibitor) before being sheared into 200–500 bp fragments with a sonicator (Bioruptor; Diagenode). Dynabeads Protein G (Invitrogen) was pre-blocked with 5 mg ml−1 of ice-cold bovine serum albumin (BSA) and incubated with 5 μg anti-H3K27ac antibody (Abcam) at 4 °C with constant rotation for 12 h. Chromatin (25 μg) was added to the bead–antibody complex mixture per reaction and incubated with constant rotation for 16 h at 4 °C. Beads were washed with ice-cold radioimmunoprecipitation assay (RIPA) buffer (Thermo Fisher Scientific) eight times, rinsed with 1× Tris–EDTA (1× Tris–EDTA) solution, eluted by adding 200 μl of ChIP elution buffer (1% SDS; 1× Tris–EDTA) and incubated in a shaker for 20 min at 65 °C. ChIP DNA was incubated for 12 h at 65 °C for reverse crosslinking, treated with RNAse A (Thermo Fisher Scientific; 1 h; 37 °C) and Proteinase K (Sigma-Aldrich; 2 h; 55 °C) and then purified on PCR purification columns. For input control, 5 μg of chromatin from whole-cell extract of each sample was subjected to reverse crosslinking, RNase A treatment (Thermo Fisher Scientific; EN0531) and Proteinase K treatment (Sigma-Aldrich; 3115887001), together with immunoprecipitated samples, and purified using PCR purification columns. DNA amounts were quantified using the PicoGreen assay (Thermo Fisher Scientific; P7589). Immunoprecipitated DNA (5 ng) and input from each sample were used to prepare ChIP libraries with TruSeq ChIP Library Preparation Kit (Illumina; IP-202-1012), according to the manufacturer’s instructions. Libraries were size selected to enrich 300–400 bp size fragments, quality controlled and sequenced on Hiseq 2000 platform (Illumina) (YCGA). Approximately 20–25 million reads were obtained from each sample. FASTA files were mapped to the mouse genome (NCBI37/mm10) using TopHat v.1.0.13 (http://tophat.cbcb.umd.edu/) and Bowtie 2 (http://bowtie-bio.sourceforge.net/bowtie2/index.shtml)62. Peaks were identified using MACS2 (SCR_013291)63.
H3K27ac ChIP–seq in the chicken
Embryonic day 15 or Hamilton–Hamburger stage 41 (ref. 64) chicken eggs were obtained from Charles River Laboratories and incubated in the laboratory for 2 days at 37 °C in a humidified chamber. The dorsal pallial regions (hyperpallium apicale and mesopallium) were microdissected from the embryonic day 17 (Hamilton–Hamburger stage 43) chicken embryos. Ten pallia were pooled and crosslinked with a formaldehyde solution (Sigma-Aldrich) at a final concentration of 1% for 10 min at room temperature per sample. L-Glycine (AmericanBio; 56-40-6) at a final concentration of 125 mM was added and incubated for 5 min at room temperature to quench the crosslinking. Cells were washed with PBS thrice and lysed in a hypotonic solution (50 mM Tris–HCl (pH 7.5), 0.5% NP-40, 0.25% sodium deoxycholate, 0.1% SDS and 150 mM NaCl) on ice for 10 min to obtain the nuclei. The nuclei were centrifuged at 600g for 5 min at 4 °C, and pellets were resuspended in the SDS lysis buffer (1% SDS, 10 mM EDTA and 50 mM Tris–HCl (pH 8.1)) before being sheared into 200–500 bp size fragments using a sonicator (M220 Focused-ultrasonicator; Covaris). The sheared DNA was diluted with the ChIP dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris–HCl (pH 8.1) and 167 mM NaCl) and pre-cleared with Magnetic Protein A/G Beads (MilliporeSigma) for 1 h at 4 °C. The beads were discarded, and anti-H3K27ac antibody (Abcam; ab4729) was added. The samples were incubated on constant rotation for overnight at 4 °C. Magnetic Protein A/G Beads (MilliporeSigma) blocked with 1 mg ml−1 BSA (Sigma-Aldrich) and transfer RNA were added to the chromatin–antibody complexes for 4 h at 4 °C. The beads were washed with low salt (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris–HCl (pH 8.1) and 150 mM NaCl), high salt (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris–HCl (pH 8.1) and 500 mM NaCl), LiCl (0.25 M LiCl, 1% IGEPAL CA-630, 1% deoxycholic acid (sodium salt), 1 mM EDTA and 10 mM Tris–HCl (pH 8.1)) and 1× Tris–EDTA (AmericanBio; AB14033-01000), sequentially for 3 min each. ChIP DNA was incubated overnight at 65 °C for reverse crosslinking and subjected to RNase A (Thermo Fisher Scientific; EN0531) treatment (37 °C; 1 h) and Proteinase K (Sigma-Aldrich; 3115887001) treatment (55 °C; 2 h) and then purified on PCR purification columns. For input control, 5 μg of crosslinked chromatin from each sample was also treated for reverse crosslinking, RNase A (Thermo Fisher Scientific; EN0531) and Proteinase K (Sigma-Aldrich; 3115887001), together with immunoprecipitated samples and purified by PCR purification columns. DNA amounts were quantified using PicoGreen assay (Thermo Fisher Scientific; P7589). Immunoprecipitated DNA (10 ng) and input from each sample were used to prepare libraries using the KAPA HyperPrep Kit (Roche; KK8500), according to the manufacturer’s instructions. Sample multiplexing was done using combinatorial dual indexes (Illumina) (YCGA). Libraries were size selected to enrich 300–400 bp size fragments and quality controlled and sequenced on a HiSeq 2000 platform (Illumina). Approximately 60 million reads were obtained per sample. Reads were mapped to the chicken genome (Galgal6) using Bowtie 2 v.2.4.2 (Research Resource Identifier (RRID): SCR_016368; http://bowtie-bio.sourceforge.net/bowtie2/index.shtml). Duplicates were removed, and unique reads with mapping quality > 20 were selected using SAMtools v.1.16 (RRID: SCR_003030; https://github.com/samtools/samtools). Peaks were called using MACS2 v.2.2.7.1 (RRID: SCR_008036).
ZBTB18 ChIP–seq in the mouse
Epitope-tagged ChIP was performed to identify ZBTB18 binding sites. Cortices from PD 0 were isolated and subjected to enzymatic dissociation. Cells from 15 cortices were pooled and treated as one sample. Cells were washed and transfected with HA-tagged Zbtb18 using Amaxa Basic Nucleofector Kit (Lonza Bioscience; VPI-1003) for primary mammalian neurons following the manufacturer’s instructions. After 48 h of culture, the medium was removed, and the cells were crosslinked with a formaldehyde solution at a final concentration of 1% for 10 min at room temperature. L-Glycine (AmericanBio; 56-40-6) was added at a final concentration of 125 mM and incubated for 5 min at room temperature to quench the crosslinking. The cells were scraped and collected in a 50-ml conical tube and lysed in the hypotonic solution (50 mM Tris–HCl (pH 7.5), 0.5% NP-40, 0.25% sodium deoxycholate, 0.1% SDS and 150 mM NaCl) on ice for 10 min to obtain the nuclei. The nuclei were centrifuged at 600g for 5 min at 4 °C, and pellets were resuspended in the SDS lysis buffer (1% SDS, 10 mM EDTA and 50 mM Tris–HCl (pH 8.1)) before being sheared into 200–500 bp size fragments using a sonicator (M220 Focused-ultrasonicator; Covaris). The sheared DNA was diluted with the ChIP dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM Tris–HCl (pH 8.1) and 167 mM NaCl) and pre-cleared with Protein A/G Magnetic BEads (Thermo Fisher Scientific) for 1 h at 4 °C. For epitope-tagged ChIP, 5 μg of HA antibody (MilliporeSigma; 11867423001) was used. Samples were incubated on constant rotation for overnight at 4 °C. Magnetic Protein A/G Beads (MilliporeSigma; 88803) were blocked with 1 mg ml−1 BSA (Sigma-Aldrich; A2153), and transfer RNAs were added to the chromatin–antibody complexes for 4 h at 4 °C. The beads were washed with low salt (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris–HCl (pH 8.1) and 150 mM NaCl), high salt (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris–HCl (pH 8.1) and 500 mM NaCl), LiCl (0.25 M LiCl, 1% IGEPAL CA-630, 1% deoxycholic acid (sodium salt), 1 mM EDTA and 10 mM Tris–HCl (pH 8.1)) and 1× Tris–EDTA (AmericanBio) sequentially for 3 min each. ChIP DNA was incubated overnight at 65 °C for reverse crosslinking, treated with RNase A (37 °C for 1 h) and Proteinase K (55 °C for 2 h) and then purified using PCR purification columns. For input control, 5 μg of crosslinked chromatin from each sample was also treated by reverse crosslinking, RNase A (Thermo Fisher Scientific) and Proteinase K (MilliporeSigma), together with immunoprecipitated samples, and purified using PCR purification columns. DNA amounts were quantified by PicoGreen assay (Thermo Fisher Scientific; P7589). Immunoprecipitated DNA (10 ng) and input from each sample were used to prepare libraries using KAPA HyperPrep Kit (Roche; KK8500) following the manufacturer’s instructions and multiplexing using combinatorial dual indexes (Illumina). The libraries were size-selected to enrich 300–400 bp size fragments, quality controlled and sequenced on the HiSeq 2000 platform (Illumina) (YCGA). Approximately 60 million reads were obtained per sample. The reads were mapped to the mouse genome (mm10) using Bowtie 2 v.2.4.2. Duplicates were removed, and unique reads with mapping quality > 20 were selected using SAMtools (v.1.16). We explored different options to determine putative ZBTB18 peaks in terms of replicates and parameters, including using TRANSFAC65, PROMO66, MatInspector67 and JASPAR databases68. Hence, peaks were called using four different approaches: (1) MACS2 (ref. 63) with default parameters, requiring the peak to be present in at least two biological replicates; (2) MACS2 with the ‘--nomodel’ and ‘--nolambda’ parameters, requiring consistency across replicates; (3) MACS2 with default parameters but only requiring the peak to be present in replicate 2; and (4) MACS3 with the peaks of replicate 2.
TF binding site prediction of candidate CREs
We obtained the DNA sequence for all 62,448 H3K27ac peaks using the twoBitToFa69 tool (https://genome.ucsc.edu/goldenPath/help/twoBit.html). We then ran the tool Find Individual Motif Occurrences (https://meme-suite.org/meme/doc/fimo.html) with default parameters to predict TFBSs in those sequences70. We used the JASPAR 2016 CORE dataset71 in MEME format containing 635 motifs. For the ET neuron-specific TFBS enrichment analysis, we calculated the number of bases present in the binding sites for a particular TF but not in the binding sites within IT neuron-associated peaks and compared them with the same numbers in ET neuron-associated peaks through Fisher’s exact test. Motifs with an FDR-corrected P value < 0.05 were considered significantly enriched.
For the six key CREs (Cux2-E1, Cux2-E4, Satb2-E1, Robo1-E1, Bcl11b-P1 and Zbtb18-P1), we further used the regular expression (Regex) method with the ZBTB18 consensus motif sequence 5′-CAGATGT-3′, allowing 1-nt mismatch to scan for any non-canonical or low-affinity motifs. Next, we used PROMO66 to test whether there are more sites in all six CREs. Finally, we reanalysed public ZBTB18 ChIP–seq data from HEK293 cells72 to derive an empirical motif using MEME–ChIP, obtaining the top motif significantly enriched in centrality through CentriMo, and searched for the top motif (MEME–TOP) in all six CREs.
Generation of enhancer reporter transgenic mice
Putative enhancers (Cux2-E1, Cux2-E2 and Cux2-E3) were amplified from mouse genomic DNA, cloned into pBgn–Gfp vector (Sestan laboratory) and sequence verified by Sanger sequencing (YCGA). The enhancers were placed upstream of the human BGN minimal promoter73 to drive GFP expression. The primers and oligonucleotides used for cloning DNA constructs are shown in Supplementary Table 15. Enhancer-inserted plasmids were linearized using appropriate restriction enzymes, size-selected by gel electrophoresis and then purified by phenol/chloroform extraction. A final concentration of 2.5 ng μl−1 of DNA was used for pronuclear injection. At PD 0, pups were examined for GFP expression under a fluorescence microscope, and tail samples were collected for genotyping to confirm the presence of Gfp loci. For GFP+ founders, brains were collected and proceeded with immunohistological analysis to examine the GFP expression pattern. Three to seven founders from each transgenic line with stable GFP expression patterns were acquired and analysed.
Quantitative RT–PCR
Total RNA was extracted using TRIzol reagent (Invitrogen; 15596018) from freshly isolated neocortical tissue from PCD 15.5 and PD 0 wild-type, Neurod6–Cre; Zbtb18 fl/fl cKO and Neurod6–Cre; Zbtb18fl/+ (control) mice and subjected to DNase I (Invitrogen; AM1907) treatment as previously described. Complementary DNA (cDNA) was synthesized using reverse transcriptase (Invitrogen; 18080093) following the manufacturer’s instructions, and quantitative PCR was performed in triplicate for each sample using an RT–PCR machine (iQ5 system; Bio-Rad) with primer sets spanning exon junctions of the targeted transcripts. Identical or near-identical-sized transcript fragments from the mouse Tbp gene were used as an internal control, and the expression level of each gene was normalized to wild type for relative fold changes. Sequences of primers used are provided in the Supplementary Table 15. Unpaired two-tailed t-test was used to detect differences between samples.
Single-cell RNA-seq analysis across species
To assess the expression patterns of Zbtb18 across different species, we reanalysed public single-nucleus transcriptome datasets for amygdala in turtle (Trachemys scripta elegans), lizard (Pogona vitticeps), chicken (Gallus gallus) and mouse (Mus musculus). We checked the expression level of Cux2, Satb2, Robo1, Bcl11b, Ccdc1167, Rpl38-ps1, Arhgef16, n-R5s136, 3830408C21Rik, Gm24089 and Gm42489 across major cell types, as identified by the original studies12,19,20,21. For cross-species comparison, we included only the excitatory neurons.
Postmortem human tissue
De-identified postmortem human brain tissue was acquired from the Sestan laboratory collection or the NIH NeuroBioBank (https://neurobiobank.nih.gov), as previously published30. All tissue was collected with informed consent from parents or next of kin and with approval of all relevant review boards or committees of the Yale University School of Medicine and the NIH. Similarly, tissue was handled in accordance with the appropriate constraints, regulations and ethical guidelines for the research use of human brain tissue set forth by the NIH (http://bioethics.od.nih.gov/humantissue.html) and the World Medical Association Declaration of Helsinki (https://www.wma.net/policies-post/wma-declaration-of-helsinki-ethical-principles-for-medical-research-involving-human-subjects/). The samples used in this study were analysed without regard to gender, and the influence of sex-specific characteristics of male versus female samples was not considered in this study.
The samples were fixed in 4% paraformaldehyde (PFA) (Electron Microscopy Sciences) for 2 days at 4 °C. Tissue sections were mounted and dried overnight. Antigen retrieval was performed using R-Buffer A pH 6.0 (Electron Microscopy Sciences) and the bench-top Antigen Retriever device (Electron Microscopy Sciences) according to the manufacturer’s instructions. Sections were washed three times in PBS, each for 15 min, and incubated in blocking solution containing 5% (v/v) normal donkey serum (Jackson ImmunoResearch Laboratories), 1% (w/v) BSA and 0.4% (v/v) Triton X-100 (Sigma-Aldrich) in PBS for 1 h at room temperature. Primary antibodies were diluted in blocking solution as follows: ZBTB18 (Proteintech; 12714-1-AP), 1:1,000; BCL11B (Abcam; ab18465), 1:500; and SATB2 (Genway Biotech; GWB-9F2D9F), 1:200. Tissue sections were incubated with these antibodies for two nights at 4 °C. Sections were washed three times with PBS, each for 15 min, before being incubated with the appropriate fluorescent secondary antibodies (Jackson ImmunoResearch Laboratories) for 1.5 h at room temperature. All secondary antibodies were raised in donkey and diluted at 1:250 in blocking solution. Finally, sections were washed with PBS with 0.3% Triton X-100, treated with the Autofluorescence Eliminator Reagent (MilliporeSigma; 2160) according to the manufacturer’s instructions and coverslipped with aqueous mounting medium (VECTASHIELD; Vector Laboratories). These sections were digitized using confocal microscope (ZEISS; LSM 510 Meta), and images were assembled in ZEISS ZEN, ImageJ, Adobe Photoshop and Adobe Illustrator.
Cell line and transfection
Neuro-2a cells (American Type Culture Collection; CCL-131) were maintained in Dulbecco’s modified Eagle’s medium (Gibco) supplied with 10% fetal bovine serum (Gibco), L-Glutamine (Thermo Fisher Scientific), penicillin (Gibco) and streptomycin (Gibco). The cell line was authenticated by morphology or genotyping, and no commonly misidentified lines were used. All lines tested negative for mycoplasma contamination, checked monthly using the MycoAlert Mycoplasma Detection Kit (Lonza). The cells were passaged every 2–3 days upon reaching 80% confluency. For transfection experiments, the Neuro-2a cells were seeded at an appropriate density the day before transfection; when the cells reached 60–70% confluency, transfection was performed 20–24 h later, with Lipofectamine 2000 (Thermo Fisher Scientific; 11668027) mixed with appropriate vectors at a DNA:lipid ratio of 1:3. Either 24 h or 48 h after transfection, the cells were dissociated with 0.25% trypsin (Gibco), washed with PBS and processed for corresponding assays.
In utero electroporation
IUE was performed on PCD 14.5 and PCD 15.5 timed-pregnancy embryos (n = 3–6 for each condition), and littermates were used as controls. A 0.5-μl DNA preparation (4 μg μl−1 of DNA mixed with 0.05% Fast Green FCF Dye (Sigma-Aldrich) was injected into the lateral ventricle of the embryos and electroporated using a square-wave pulse electroporator (Harvard Apparatus; BTX) at 35–38 V, five pulses, 50 ms ON and 950 ms OFF to deliver DNA constructs to the ventricular zone. At PD 0, pups were screened for GFP or RFP expression under a fluorescence-attached dissection microscope. Pups with the fluorescence signal were euthanized, and brains were analysed as previously described.
Neuronal tracing and imaging
Neonatal mice injections were performed with a motorized microinjector (RWD Life Science; MM-500 and R480). In brief, mice at PD 3 were anaesthetized through hypothermia, and approximately 50 nl of retrograde AAV viral particles carrying pCAG–Gfp cassette (Addgene; 37825-AAVrg; titre ≥ 7 × 1012 vg ml−1) was injected into the medial prefrontal cortex visually with the guide of a stereo binocular microscope. At PD 7, the injected mice that were properly targeted were perfused with ice-cold PBS, followed by 4% PFA (Electron Microscopy Sciences). The collected brains underwent postfixation at 4 °C overnight and washed with PBS before cutting coronally at 60-μm thickness. The sections were stained with anti-GFP antibody (Abcam; ab13970) before imaging. Images were taken using an automated slide scanner (Olympus; VS200) with ×20 objective. For analysis, the images from wild-type mice were aligned with the Allen Brain Atlas coordinates at PD 6, and the Neurod6–Cre Zbtb18 cKO was approximated with the wild-type counterpart. The cell numbers from each bin were counted using the ImageJ tool (RRID: SCR_003070). The total cell number from the neocortex, as shown in Fig. 3c from three mice of each genotype, was compared. For the percentage of distribution, we divided the cell number from each bin by the total number of cells in the neocortex, and comparisons between each bin were made with an unpaired t-test.
BrdU/CIdU/IdU birth-date labelling
Either 5′-bromo-2′-deoxyuridine (BrdU) (Sigma-Aldrich; B5002) or CIdU (Sigma-Aldrich; C6891) was dissolved in distilled water at 100 mg ml−1, and IdU (Sigma-Aldrich; I7125) was dissolved in distilled water at 50 mg ml−1 and stored at −20 °C in the dark. At the proscribed ages, a single dose of IdU, or CIdU was redissolved by intermittent vortexing and given to the timed-pregnant mice by intraperitoneal injection at a dosage of 1 mg (20 g)−1 body weight (10 μl of CIdU or 20 μl of IdU). Pups were euthanized within the PD 1, and brains were fixed in 4% PFA (Electron Microscopy Sciences) for 12 h at 4 °C. Heterozygous littermates were used as controls. The brains were embedded with 4% agarose and sectioned at 50-μm thickness on a vibratome (Leica; VT1000S). Sections were treated with 2 M HCl at room temperature for 30 min and co-immunostained with BrdU (Sigma-Aldrich) and other markers for immunofluorescence.
Recombinant DNA
DNA constructs used for making transgenic mice were subcloned from the pBgn–GFP (Sestan laboratory) backbone with Cux2-E1, Cux2-E2 and Cux2-E3 inserted. Vectors for overexpression experiments were subcloned from the pCAGIG backbone (Addgene; 11159), with inserts of the full coding sequences of the following genes: mouse Zbtb18 (also known as Rp58, Znf238 and Zfp238), BC054529; mouse Pou3f2, NM_008899; mouse Trim28 (also known as Rnf96), BC058391; mouse Lhx2, BC055741; mouse Satb2, BC138626; mouse Hdac2, BC138517; mouse E2F1, BC052160; mouse Sin3A, BC052716; mouse Foxn2, NM_001355743; mouse Coup-tf1 (also known as Nr2f1), BC108408; and mouse Robo1, NM_019413.2. Coding sequences in DNA constructs used for IUE experiments were cloned into an expression construct with a Neurod1 promoter74 and verified by Sanger sequencing (YCGA). Other DNA constructs used in this study included pCAG–Cre, pCALNL–GFP, pCAGIG, pCALSL–RFP, pNeurod1–GFP and subcloned plasmids. The plasmids pCAG–Cre, pCALNL–GFP and pCAGIG were gifts from C. Cepko (Addgene; 11159, 13770 and 13775, respectively), whereas pCALSL–RFP and pNeurod1–GFP have been previously described74,75. Vectors carrying enhancers for luciferase assay were subcloned from pGL4.24 backbone (Promega). For enhancer mutagenesis, desired sites within specific enhancers were mutated using a site-directed mutagenesis kit (New England Biolabs; Q5 Site-Directed Mutagenesis Kit) following the manufacturer’s instructions. The mutated products were validated by Sanger sequencing. Oligos and primers used for cloning are listed in Supplementary Table 15. For neurite outgrowth analysis in chicken primary neurons, pCagen (empty vector; a gift from C. Cepko; Addgene; 11160), pCag–mCherry (a gift from Phil Sharp laboratory, through Addgene; 41583) and pCag–Cux2 (mouse Cux2 expression construct, cloned in pCAG backbone using NsiI and NotI restriction enzymes, with Cux2 open reading frame subcloned from MGC clone ID 30532644) were used.
Cux2-E1 enhancer activity assay and Cux2 overexpression in chicken hyperpallial neurons
Primary hyperpallial cells were isolated from chick embryos at embryonic days 7 and 17. After carefully opening the eggshells, the embryos were collected and immediately transferred into ice-cold artificial cerebrospinal fluid supplemented with the following components: 92 mM N-methyl-D-glucamine (Sigma-Aldrich; M2004), 20 mM HEPES (Sigma-Aldrich; H3375), 5.5 mM glucose (Sigma-Aldrich; G7021), 30 mM sodium bicarbonate (Sigma-Aldrich; S5761), 5 mM sodium L-ascorbate (Sigma-Aldrich; A4034), 2.5 mM potassium chloride (Sigma-Aldrich; P9541), 1.25 mM sodium phosphate (Sigma-Aldrich; S0751), 2 mM thiourea (Sigma-Aldrich; T7875), 3 mM 196 sodium pyruvate (Sigma-Aldrich; P2256), 5.5 mM urea (Sigma-Aldrich; U5128), 10 mM magnesium sulfate (Sigma-Aldrich; M7506) and 0.5 mM calcium chloride (Sigma-Aldrich; 21115). Following dissection, tissues were enzymatically dissociated by incubation in 2 mg ml−1 of papain (Transnetyx; PAP) for 20–30 min at 37 °C. After enzymatic digestion, 0.1 mg ml−1 of DNase I (STEMCELL Technologies; 07900) was added, and the tissue was gently triturated using a 1-ml glass pipette to obtain a single-cell suspension. Dissociated cells were transfected with various plasmids using the Chicken Neuron Nucleofector Kit (Lonza; VPG-1002) according to the manufacturer’s instructions. A total of 4 µg of each plasmid was used in all experiments. Post-transfection cells were seeded at a density of 4 × 105 cells per well in 24-well plates (ibidi; 82406) precoated with poly-D-lysine (Gibco; A3890401) and laminin (Gibco; 23017015). Cultures were maintained in neurobasal medium (Gibco; 21103049) supplemented with 1× B27 (Gibco; 17504044), 1% GlutaMAX (Gibco; 35050061), 30 nM sodium selenite (Sigma-Aldrich; S5261), 5 µg ml−1 of human insulin (Sigma-Aldrich; I9278), 1% penicillin–streptomycin (Gibco; 15140122), 10 ng ml−1 of brain-derived neurotrophic factor (PeproTech; 450-02) and 10 ng ml−1 of NT-3 (PeproTech; 450-03). Media were changed every 2 days. After 48 h, primary chicken neuronal cultures were fixed with 4% PFA for 10 min, followed by three washes with PBS. Cells were permeabilized with 0.3% Triton X-100 (Sigma-Aldrich; T8787) in PBS for 10 min at room temperature and blocked with 10% (v/v) normal donkey serum (Jackson ImmunoResearch Laboratoires; 017-000-121) in PBS for 1 h at room temperature. The primary antibodies were diluted in the same blocking solution and incubated with the cells overnight at 4 °C. The primary antibodies used were anti-SATB2 (1:100; Abcam), anti-ZBTB18 (1:250; Proteintech), anti-GFP (1:2,000; Abcam) and anti-mCherry (1:1,000; Aves Labs). After three washes with PBS, cells were incubated with the respective secondary antibodies prepared in blocking solution for 2 h at room temperature. Nuclei were stained with DAPI (1 μg ml−1; Sigma-Aldrich; D9542) for 5 min at room temperature. Fluorescent images were acquired using a confocal microscope (LSM880; Zeiss). Post hoc, the GFP intensity was quantified using the ImageJ tool (RRID: SCR_003070), and the neurite length was traced following mCherry signal using the NeuronJ plugin of the ImageJ tool. Because deletion constructs used as controls may alter the local DNA context and possibly disrupt cryptic overlapping motifs, we further performed in silico analysis of the 30-bp DNA sequencing flanking the deleted region and found no high-affinity binding motifs for other relevant TFs.
Luciferase assay
Neuro-2a (American Type Culture Collection; CCL-131) cells were plated into 96-well or 24-well plates at a density of 10,000 or 50,000 cells per well, respectively. Sixteen hours after plating, cells were transfected with a DNA mixture consisting of 100 ng and 500 ng overexpression plasmids, 30 ng and 500 ng of firefly luciferase (pGL4.24; Promega) plasmids containing candidate enhancers, together with 20 ng and 25 ng pRL-SV40 plasmid (Promega) as a control for 96-well and 24-well plates, respectively, using Lipofectamine 2000 (Thermo Fisher Scientific). Transfected cells were lysed and assayed 24–48 h after transfection using the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer’s instructions. Relative luciferase activity was calculated by normalizing the firefly luciferase luminescence to the Renilla luciferase luminescence value. Luciferase activity was measured and quantified by GloMax-Multi Detection System (Promega). Primers used for amplifying the candidate enhancer and promoter regions are shown in Supplementary Table 15. Experiments were performed in triplicate or quadruplicate, and the data were normalized to the empty vector control (pCAGEN versus the pZBTB18). P values were calculated using unpaired two-tailed t-tests to compare the overexpression and control. To perform the luciferase assay on multiple mutated enhancers, a one-way ANOVA test was used to detect differences among more than two enhancers; alpha value = 0.05.
In situ hybridization for the human, mouse and chicken brain sections
RNA probes for mouse and human were generated using the respective neocortical tissue cDNA as template (mouse Zbtb18, ENSMUSG00000063659; mouse Cux2, ENSMUSG00000042589; mouse Satb2, ENSMUSG00000038331; mouse Robo1, ENSMUSG00000022883; mouse Robo2, ENSMUSG00000052516; mouse Robo3, ENSMUSG00000032128; mouse Rorb, ENSMUSG00000036192; mouse Dcc, ENSMUSG00000060534; human ZBTB18, ENSG00000179456; and human CUX2, ENSG00000111249) by TA Cloning Kit (Invitrogen) and followed by in vitro transcription (Roche) according to the manufacturer’s instructions. For chicken probes, the cDNA fragments coding for SATB2 (NM_001199110) and CUX2 (XM_415167), were amplified by RT–PCR and subcloned into the plasmid vector pTA2 (Toyobo). Templates purified by phenol/chloroform extraction and digoxigenin (DIG)-labelled probes were synthetized using T3 (Roche) and T7 RNA polymerases (Roche), respectively. RNA labelling was performed mix (Roche) according to the manufacturer’s instructions. Probes were purified by phenol/chloroform extraction, quantified, quality controlled and stored at −80 °C until hybridization. Primers used for generating probes are provided in Supplementary Table 15.
For single-colour ISH, free-floating or slide-mounted cryosections at 20–30 μm thickness were processed according to a previously described protocol76. In brief, brains were fixed overnight at 4 °C in 4% PFA (Electron Microscopy Sciences) diluted in Dulbecco’s phosphate-buffered saline (DPBS) (Thermo Fisher Scientific), equilibrated for 12 h at 4 °C in 10% sucrose and another 12 h at 4 °C in 30% sucrose (AmericanBio) in DPBS (Thermo Fisher Scientific). Fixed brains were then embedded in O.C.T. (Scigen) and sliced on a cryostat (Leica Biosystems; CM1800). Slides were stored at −80 °C until processed for ISH. Sections were first postfixed in 4% PFA (Electron Microscopy Sciences) in PBS for 15 min at room temperature, washed with PBS and submerged in hybridization buffer (5× saline–sodium citrate, 50% formamide, 5× Denhardt’s solution, 500 µg ml−1 of salmon sperm DNA and 250 µg ml−1 of torula yeast RNA) supplemented with 1,000 ng ml−1 of the appropriate DIG-labelled probe at 70 °C overnight. Sections were washed twice for 60 min at 70 °C in 2× saline–sodium citrate, 50% formamide and 0.1% Tween, followed by washing with 100 mM Tris–HCl (pH 7.5), 150 mM NaCl and 0.1% Tween. These sections were then blocked with 10% sheep inactivated serum (Sigma-Aldrich) and incubated overnight at 4 °C with an anti-DIG antibody conjugated to alkaline phosphatase (1:2,000; Roche). The sections were then rinsed in the substrate buffer (100 mM Tris–HCl (pH 9.5), 100 mM NaCl, 50 mM MgCl2 and 0.1% Tween-20) before being overlaid with NBT–BCIP substrate (Roche). Revelation was done at room temperature in the dark until the desired signal was reached. Finally, the sections were rinsed in DPBS (Thermo Fisher Scientific), postfixed with 4% PFA (Electron Microscopy Sciences) in DPBS (Thermo Fisher Scientific), washed with water and mounted with Permount mounting medium (Electron Microscopy Sciences).
For two-colour ISH, probes were synthesized by either DIG-labelled (Roche) or fluorescein-labelled (Roche) RNA labelling mixes in vitro transcription (Roche). Free-floating or slide-mounted cryosections (20–30 μm thickness) were postfixed in 4% PFA (Electron Microscopy Sciences) for 15 min, washed and hybridized overnight at 70 °C in 14-ml hybridization solution with 500 ng ml−1 of DIG-labelled (Roche) human ZBTB18 probe and 500 ng ml−1 of fluorescein-labelled (Roche) human CUX2 probe for human tissue. For mouse tissue, 500 ng ml−1 of DIG-labelled mouse Zbtb18 and 500 ng μl−1 of fluorescein-labelled mouse Cux2 probes were used. The signals were sequentially detected with an alkaline phosphatase-conjugated anti-DIG antibody (MilliporeSigma), NBT–BCIP substrate (Roche) and horseradish peroxidase-conjugated anti-fluorescein antibody. Horseradish peroxidase activity was initially amplified using TSA (Akoya) and subsequently strengthened with 3,3′-diaminobenzidine tetrahydrochloride hydrate (DAB) (Sigma-Aldrich).
Immunostaining and immunoblotting
Brains dissected from embryonic and neonatal mice were fixed by immersion in 4% PFA (Electron Microscopy Sciences) overnight at 4 °C. Adult brains were perfused with 10 ml PBS followed by 10 ml of 4% PFA (Electron Microscopy Sciences), isolated and postfixed by immersion in 4% PFA (Electron Microscopy Sciences) overnight at 4 °C. The brains were then sectioned at a thickness of 50 μm (postnatal) and 80 μm (embryonic) on a vibratome (Leica; VT1000S). The brain sections were blocked using blocking solution (5% BSA, 10% donkey serum and 0.3% Triton X-100 in 1× PBS solution) for 1 h at room temperature and incubated with appropriate primary antibodies for 12–24 h at 4 °C, washed three times with 1× PBS and incubated with appropriate secondary antibodies for 1 h at room temperature. DAPI was used to stain nuclei. The antibody dilutions used for immunostaining were as follows: anti-ZBTB18 (rabbit; 1:1,000; Proteintech), anti-GFP (chicken; 1:3,000; Abcam), anti-RFP (rabbit; 1:1,000; Abcam), anti-CUX1 (rabbit; 1:250; Santa Cruz Biotechnology), anti-CUX2 (rabbit; 1:250; Abcam), anti-SATB2 (mouse; 1:200; Genway), anti-BCL11B (rat; 1:500; Abcam), anti-TBR1 (rabbit; 1:250; Santa Cruz Biotechnology), anti-LHX6 (mouse; 1:300; Santa Cruz Biotechnology), anti-LHX6 (goat; 1:250; Santa Cruz Biotechnology), anti-RELN (mouse; 1:300; Millipore), anti-BrDU (mouse; 1:250; BD Biosciences) and anti-BrDU (rat; 1:250; Accurate Chemical). For the quantitative analysis of total number of BCL11B+ and SATB2+ cells, z-stack images from three per species were used to segment DAPI signals from individual nuclei, and fluorescence signals from the nuclei were obtained using Volocity (v.6.3.1). The positive signals for each marker protein were defined on the basis of the median nuclear intensity from all cells analysed using the Spotfire software (v.11.2.0).
Immunoblot analyses were performed using a previously described protocol77. In brief, neocortical tissue from PD 0 Zbtb18fl/fl; Emx1–Cre mice (n = 3), Zbtb18fl/+; Emx1–Cre mice (n = 3) and wild-type mice (n = 3) were isolated and snap-frozen in liquid nitrogen. The samples were then minced and mixed with lysis buffer (150 mM NaCl, 1.0% NP-40 and 50 mM Tris–HCl (pH 8.0)), including protease inhibitors (Roche). Equal amounts of protein from each sample were loaded for SDS–PAGE, followed by western blot analysis with signals detected using the enhanced chemiluminescence detection reagent. Antibody dilutions used for immunoblot were as follows: anti-GAPDH (rabbit; 1:5,000; Abcam; ab9485), anti-CUX2 (rabbit; 1:1,000; Abcam), anti-ZBTB18 (rabbit; 1:2,000; Proteintech) and anti-ZBTB18 (goat; 1:2,000; Santa Cruz Biotechnology). For detailed antibody information, refer to Supplementary Table 14.
Obtaining multispecies alignments of TF binding sites
Beginning with the lists of putative TFBSs identified by Find Individual Motif Occurrences, as described earlier70, we selected only those motifs that are recognized by TFs expressed (reads per kilobase of transcript per million mapped reads ≥ 1) in our RNA-seq samples derived from either IT or ET neurons for further analysis. This produced a list of 267 motifs. The total number of binding sites for those motifs ranged from 0 to 244,332.
For each of the 267 motifs, we obtained the multiple-species sequence alignment of 60 vertebrate species from the MULTIZ60 (ref. 78) track in the University of California, Santa Cruz (UCSC) browser using mafsInRegion69 from the UCSC tools. We stitched maf alignments and converted them to fasta using the script maf_to_concat_fasta.py from bx-python (https://github.com/bxlab/bx-python/blob/main/scripts/maf_to_concat_fasta.py).
Conservation of TF binding sites across species
We used an in-house script to analyse the multiple sequence alignment of each binding site. First, we calculated the information content for each position in each MEME motif using the R package TFBSTools79. For each motif, we identified positions with high information content (0.75 or higher) and considered them ‘core’ positions. Second, we divided the species in the 60-way multiple sequence alignment into four groups: ‘placental mammals’, ‘marsupials’, ‘monotremes’ and ‘non-mammals’. Finally, for each binding site, we counted the number of species in each group in which the motif was present without any single nucleotide substitutions in any of the core positions compared with the mouse reference sequence mm10. We repeated the analysis with one, two and three mutations. A motif was not considered conserved if it contained an indel spanning the core positions.
For each group of species, we produced a binary vector in which, for each binding site, a value of 1 was assigned if the binding site was conserved and 0 if it was non-conserved. For placental mammals, we required at least six species fulfilling the conservation criteria stated above. For marsupials, monotremes and non-mammalian species, we required at least one species.
Enrichment in TF binding sites in sets of H3K27ac peaks
We selected seven sets of ChIP–seq peaks: peaks annotated to differentially expressed genes between IT and ET neurons (DESeq2 PAdj < 0.05; one set for upregulated genes and another for downregulated genes), peaks differentially enriched between IT and ET neurons (as determined by DESeq2 with FC > 2 and PAdj < 0.1; one set for IT neuron-biased peaks and another for ET neuron-biased peaks), peaks annotated to genes differentially expressed between the Zbtb18 knockout (KO) or Zbtb18 heterozygous mice (DESeq2 PAdj < 0.05; one set for KO and another for heterozygous-biased genes) and a final set composed by a manually curated list of genes important for corpus callosum formation extracted from the literature. In addition, we produced a list of background peaks composed of all H3K27ac peaks, which did not fall in any of the previous categories. We then grouped these CREs into four categories to evaluate the enrichment of TFBSs across species: (1) H3K27ac peaks associated with ET neuron-biased genes (n = 1,433), which include peaks annotated to ET neuron-enriched genes and peaks enriched in ET neurons; (2) peaks associated with IT neuron-biased genes (n = 1,409), which include peaks annotated to IT neuron-enriched genes and peaks enriched in IT neurons; (3) peaks associated with CREs near genes with expression that was downregulated in Zbtb18 KO (n = 8,187); and (4) peaks associated with CREs near genes with expression that was upregulated in Zbtb18 KO (n = 3,705). For associating peaks with genes in the identification of putative CREs, we associated peaks with the nearest genes.
For each group of species, we counted the number of conserved and non-conserved binding sites observed in each set of peaks. We compared the proportion of conserved versus non-conserved binding sites to the same ratio in the background peaks using Fisher’s exact test. Motifs with FDR-adjusted P values < 0.05 were considered significant.
The number of motifs tested in each category of enhancers, including those selected as background supplementary, is shown in Supplementary Table 17. To evaluate the robustness of these enrichments, we devised a bootstrap-based strategy (Extended Data Fig. 10d). For each comparison, we counted ZBTB18 motifs within each category and performed 1,000 random samplings, with replacement, of an equal number of ZBTB18 motifs from background H3K27ac regions (considered invariant putative regulatory elements). We then compared the observed ratios of conserved and non-conserved motifs at each phylogenetic depth to the distribution of 1,000 such ratios obtained from the background peaks. The percentile occupied by the observed ratio represents an empirical P value, indicating how likely it is to obtain this ratio by chance among the invariant peaks.
Conservation of H3K27ac and ZBTB18 peaks
We used mafsInRegion69 from the UCSC tools to obtain MULTIZ60 alignments for each selected enhancer. We stitched maf alignments and converted them to fasta using the script maf_to_concat_fasta.py from bx-python. The pairwise alignment distance between species of each of the selected H3K27ac peaks was obtained using the function dist.alignment from the seqinr package in R.
The intersection of chicken and mouse H3K27ac and ZBTB18 ChIP–seq peaks was conducted using the ‘IntersectBed’ function within bedtools. To facilitate this intersection, chicken peaks were initially converted into mouse coordinates (from gg6 to mm10) using LiftOver with -minMatch = 0.1. Orthologous regions in chicken of all ZBTB18 ChIP–seq peaks identified in mice were determined using LiftOver from mm10 to gg6 using the same minMatch value.
Evolutionary age of the ZBTB18 gene
To evaluate the evolutionary age of the ZBTB18 gene, we used protein sequence similarity (BLASTP80 and HMMER81) to search our reference database, which is adapted from UniProt82 and trimmed with a taxonomically informed procedure that optimizes database size while ensuring that species with well-resolved genomes are kept in all major branches of the phylogenetic tree of life83. We partitioned the similarity hits according to their evolutionary distance from humans and the species in which they appear. Here the evolutionary age of a gene is defined as the taxonomic restriction of the most ancient fragment of the protein encoded by the gene83,84,85,86. If an ancestral gene duplicated and expanded, all descendant genes are considered of the same evolutionary gene as the ancestral gene86. For any protein of at least 40 amino acids, the taxonomic restriction is indicated as the index of the NCBI taxonomic node87 at which the query species and subject species diverged. The evolutionary age of each taxonomic (phylogenetic) node is provided by the TimeTree database88. Thus, a human protein also present in bacteria is ancient; a human protein also present in sea squirts is restricted to the Chordata phylum; a human protein also present in platypus is restricted to the Mammalia class; a human protein present also in the Coquerel’s sifaka lemur is restricted to the Primates order; a human protein present only in humans is restricted to the human species. First, we found that few bacterial species in our database harbour proteins similar to the human ZBTB18 protein (four species with hits in 1,600 species of our database; 0.025%). Of the four bacterial proteins similar to ZBTB18, three of them have two or four zinc finger domains, whereas the remaining protein has a BTB domain. Most eukaryotic species harbour proteins similar to the human ZBTB18 (67 of 100 eukaryotic species; 67%). The three eukaryotic proteins most similar to ZBTB18 have two or four zinc finger domains. Second, we found that in all subsequent evolutionary nodes of the human lineage, all outgroup species for each node harbour proteins similar to ZBTB18. Thus, ZBTB18 is an ancient gene that probably arose at the origin of eukaryotes. The few bacterial hits may indicate an even more ancient origin in all cellular organisms. The presence of genes similar to ZBTB18 in only 0.025% of bacterial species is consistent either with an extremely ancient origin in all cellular organisms followed by many events of gene loss in most bacterial species, or with two to four events of horizontal gene transfer from eukaryotes to a few bacterial species. Although both explanations are possible, the second requires fewer events. In any case, ZBTB18 is an ancient gene that arose in eukaryotes approximately two billion years ago or even earlier at the origin of cellular organisms approximately 3.6 billion years ago.
Cell counting and data analysis
For each cell counting analysis, neocortical somatosensory regions from three different brains of each condition were used. For PD 0 mouse and late to midfetal human brain tissues, 200-μm-wide neocortical columns covering from layer 1 to subplate were used as standard fields to conduct counting. For PCD 14.5 and PCD 16.5 mouse tissues, 200-μm-wide neocortical columns covering from layer 1 to ventricular zone were used as standard fields. Unpaired two-tailed t-tests were used to detect differences between samples. P < 0.05 (α = 0.05) was set as the cutoff for significance.
Replicates
For ChIP–seq, two replicates were used for each condition. For RNA-seq, a minimum of three replicates were used for each condition. For ChIP–quantitative polymerase chain reaction analysis, a minimum of four replicates were used for each condition. For RT–PCR analysis, three replicates from each time point were used for each condition. For Cux2-E1, Cux2-E2 and Cux2-E3 transgenic mice, three to seven founders from each line were analysed. For IUE experiments, three to six successfully electroporated animals were examined for each analysis. For IDU/CIDU in vivo labelling, a minimum of two litters were used for each experiment, and three to five brains from each litter were examined. For ISH and immunofluorescence experiments using mouse tissue, a minimum of three animals were used for each experiment, and between 3 and 20 sections from each animal were examined. For ISH and immunofluorescence experiments using non-mouse mammals, two to five sections from two individuals were used. For cell culture experiments, cells were isolated from three different mouse brains per condition, and two replicates from each brain were used for subsequent observations. For luciferase assays, a minimum of three replicates were used for each transfection and analysis. Statistical methods were not used to determine sample size. For all experiments, n = 3 or >3 was used.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Data were deposited into the NCBI Sequence Read Archive under BioProject accession number PRJNA1391524.
References
Striedter, G. & Northcutt, R. Brains Through Time: A Natural History of Vertebrates (Oxford Univ. Press, 2019).
Krubitzer, L. & Kaas, J. The evolution of the neocortex in mammals: how is phenotypic diversity generated?. Curr. Opin. Neurobiol. 15, 444–453 (2005).
Rowe, T. B., Macrini, T. E. & Luo, Z. X. Fossil evidence on origin of the mammalian brain. Science 332, 955–957 (2011).
Belgard, T. G. et al. Adult pallium transcriptomes surprise in not reflecting predicted homologies across diverse chicken and mouse pallial sectors. Proc. Natl Acad. Sci. USA 110, 13150–13155 (2013).
Molyneaux, B. J., Arlotta, P., Menezes, J. R. & Macklis, J. D. Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci. 8, 427–437 (2007).
Leone, D. P., Srinivasan, K., Chen, B., Alcamo, E. & McConnell, S. K. The determination of projection neuron identity in the developing cerebral cortex. Curr. Opin. Neurobiol. 18, 28–35 (2008).
Kwan, K. Y., Sestan, N. & Anton, E. S. Transcriptional co-regulation of neuronal migration and laminar identity in the neocortex. Development 139, 1535–1546 (2012).
Suarez, R. et al. A pan-mammalian map of interhemispheric brain connections predates the evolution of the corpus callosum. Proc. Natl Acad. Sci. USA 115, 9622–9627 (2018).
Jarvis, E. D. et al. Avian brains and a new understanding of vertebrate brain evolution. Nat. Rev. Neurosci. 6, 151–159 (2005).
Suzuki, I. K., Kawasaki, T., Gojobori, T. & Hirata, T. The temporal sequence of the mammalian neocortical neurogenetic program drives mediolateral pattern in the chick pallium. Dev. Cell 22, 863–870 (2012).
Briscoe, S. D. & Ragsdale, C. W. Homology, neocortex, and the evolution of developmental mechanisms. Science 362, 190–193 (2018).
Tosches, M. A. et al. Evolution of pallium, hippocampus, and cortical cell types revealed by single-cell transcriptomics in reptiles. Science 360, 881–888 (2018).
Nord, A. S., Pattabiraman, K., Visel, A. & Rubenstein, J. L. R. Genomic perspectives of transcriptional regulation in forebrain development. Neuron 85, 27–47 (2015).
Villar, D. et al. Enhancer evolution across 20 mammalian species. Cell 160, 554–566 (2015).
Li, M. et al. Integrative functional genomic analysis of human brain development and neuropsychiatric risks. Science 362, eaat7615 (2018).
Andrews, G. et al. Mammalian evolution of human cis-regulatory elements and transcription factor binding sites. Science 380, eabn7930 (2023).
Zemke, N. R. et al. Conserved and divergent gene regulatory programs of the mammalian neocortex. Nature 624, 390–402 (2023).
Shibata, M. et al. Regulation of prefrontal patterning and connectivity by retinoic acid. Nature 598, 483–488 (2021).
Hain, D. et al. Molecular diversity and evolution of neuron types in the amniote brain. Science 377, eabp8202 (2022).
Colquitt, B. M., Merullo, D. P., Konopka, G., Roberts, T. F. & Brainard, M. S. Cellular transcriptomics reveals evolutionary identities of songbird vocal circuits. Science 371, eabd9704 (2021).
Yao, Z. et al. A taxonomy of transcriptomic cell types across the isocortex and hippocampal formation. Cell 184, 3222–3241 (2021).
Nieto, M. et al. Expression of Cux-1 and Cux-2 in the subventricular zone and upper layers II-IV of the cerebral cortex. J. Comp. Neurol. 479, 168–180 (2004).
Zimmer, C., Tiveron, M. C., Bodmer, R. & Cremer, H. Dynamics of Cux2 expression suggests that an early pool of SVZ precursors is fated to become upper cortical layer neurons. Cereb. Cortex 14, 1408–1420 (2004).
Barao, S. et al. Conserved transcriptional regulation by BRN1 and BRN2 in neocortical progenitors drives mammalian neural specification and neocortical expansion. Nat. Commun. 15, 8043 (2024).
Lu, C. et al. Essential transcription factors for induced neuron differentiation. Nat. Commun. 14, 8362 (2023).
Cubelos, B. et al. Cux1 and Cux2 regulate dendritic branching, spine morphology, and synapses of the upper layer neurons of the cortex. Neuron 66, 523–535 (2010).
Di Bella, D. J. et al. Molecular logic of cellular diversification in the mouse cerebral cortex. Nature 595, 554–559 (2021).
Okado, H. et al. The transcriptional repressor RP58 is crucial for cell-division patterning and neuronal survival in the developing cortex. Dev. Biol. 331, 140–151 (2009).
Johnson, M. B. et al. Functional and evolutionary insights into human brain development through global transcriptome analysis. Neuron 62, 494–509 (2009).
Kang, H. J. et al. Spatio-temporal transcriptome of the human brain. Nature 478, 483–489 (2011).
Xiang, C. et al. RP58/ZNF238 directly modulates proneurogenic gene levels and is required for neuronal differentiation and brain expansion. Cell Death Differ. 19, 692–702 (2012).
Clement, O. et al. Rp58 and p27(kip1) coordinate cell cycle exit and neuronal migration within the embryonic mouse cerebral cortex. Neural Dev. 12, 8 (2017).
Orellana, C. et al. Corpus callosum abnormalities and the controversy about the candidate genes located in 1q44. Cytogenet. Genome Res. 127, 5–8 (2009).
Ballif, B. C. et al. High-resolution array CGH defines critical regions and candidate genes for microcephaly, abnormalities of the corpus callosum, and seizure phenotypes in patients with microdeletions of 1q43q44. Hum. Genet. 131, 145–156 (2012).
de Munnik, S. A. et al. A de novo non-sense mutation in ZBTB18 in a patient with features of the 1q43q44 microdeletion syndrome. Eur. J. Hum. Genet. 22, 844–846 (2014).
De Rubeis, S. et al. Synaptic, transcriptional and chromatin genes disrupted in autism. Nature 515, 209–215 (2014).
Wang, T. et al. Large-scale targeted sequencing identifies risk genes for neurodevelopmental disorders. Nat. Commun. 11, 4932 (2020).
Srivatsa, S. et al. Unc5C and DCC act downstream of Ctip2 and Satb2 and contribute to corpus callosum formation. Nat. Commun. 5, 3708 (2014).
Marsh, A. P. et al. Mutations in DCC cause isolated agenesis of the corpus callosum with incomplete penetrance. Nat. Genet. 49, 511–514 (2017).
Jamuar, S. S. et al. Biallelic mutations in human DCC cause developmental split-brain syndrome. Nat. Genet. 49, 606–612 (2017).
Andrews, W. et al. Robo1 regulates the development of major axon tracts and interneuron migration in the forebrain. Development 133, 2243–2252 (2006).
Lopez-Bendito, G. et al. Robo1 and Robo2 cooperate to control the guidance of major axonal tracts in the mammalian forebrain. J. Neurosci. 27, 3395–3407 (2007).
Zelina, P. et al. Signaling switch of the axon guidance receptor Robo3 during vertebrate evolution. Neuron 84, 1258–1272 (2014).
Darki, F. et al. Human ROBO1 regulates white matter structure in corpus callosum. Brain Struct. Funct. 222, 707–716 (2017).
Zeppilli, S. et al. Single-cell genomics of the mouse olfactory cortex reveals contrasts with neocortex and ancestral signatures of cell type evolution. Nat. Neurosci. 28, 937–948 (2025).
Trost, B. et al. Genomic architecture of autism from comprehensive whole-genome sequence annotation. Cell 185, 4409–4427 (2022).
Cargnin, F. et al. FOXG1 orchestrates neocortical organization and cortico-cortical connections. Neuron 100, 1083–1096 (2018).
Talkowski, M. E. et al. Sequencing chromosomal abnormalities reveals neurodevelopmental loci that confer risk across diagnostic boundaries. Cell 149, 525–537 (2012).
Satterstrom, F. K. et al. Large-scale exome sequencing study implicates both developmental and functional changes in the neurobiology of autism. Cell 180, 568–584 (2020).
Trubetskoy, V. et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 604, 502–508 (2022).
Uddin, L. Q. et al. Salience network-based classification and prediction of symptom severity in children with autism. JAMA Psychiatry 70, 869–879 (2013).
Hahamy, A., Behrmann, M. & Malach, R. The idiosyncratic brain: distortion of spontaneous connectivity patterns in autism spectrum disorder. Nat. Neurosci. 18, 302–309 (2015).
Ilioska, I. et al. Connectome-wide mega-analysis reveals robust patterns of atypical functional connectivity in autism. Biol. Psychiatry 94, 29–39 (2023).
Tsyporin, J. et al. Competing programs shape cortical sensorimotor-association axis development. Preprint at bioRxiv https://doi.org/10.1101/2025.06.26.660775 (2025).
Gong, S. et al. A gene expression atlas of the central nervous system based on bacterial artificial chromosomes. Nature 425, 917–925 (2003).
Gorski, J. A. et al. Cortical excitatory neurons and glia, but not GABAergic neurons, are produced in the Emx1-expressing lineage. J. Neurosci. 22, 6309–6314 (2002).
Nakamura, T., Colbert, M. C. & Robbins, J. Neural crest cells retain multipotential characteristics in the developing valves and label the cardiac conduction system. Circ. Res. 98, 1547–1554 (2006).
Schwab, M. H. et al. Neuronal basic helix-loop-helix proteins (NEX and BETA2/Neuro D) regulate terminal granule cell differentiation in the hippocampus. J. Neurosci. 20, 3714–3724 (2000).
Goebbels, S. et al. Genetic targeting of principal neurons in neocortex and hippocampus of NEX-Cre mice. Genesis 44, 611–621 (2006).
Sestan, N., Artavanis-Tsakonas, S. & Rakic, P. Contact-dependent inhibition of cortical neurite growth mediated by notch signaling. Science 286, 741–746 (1999).
Trapnell, C., Pachter, L. & Salzberg, S. L. TopHat: discovering splice junctions with RNA-seq. Bioinformatics 25, 1105–1111 (2009).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Zhang, Y. et al. Model-based analysis of ChIP-seq (MACS). Genome Biol. 9, R137 (2008).
Hamburger, V. & Hamilton, H. L. A series of normal stages in the development of the chick embryo. 1951. Dev. Dyn. 195, 231–272 (1992).
Matys, V. et al. TRANSFAC: transcriptional regulation, from patterns to profiles. Nucleic Acids Res. 31, 374–378 (2003).
Messeguer, X. et al. PROMO: detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 18, 333–334 (2002).
Cartharius, K. et al. MatInspector and beyond: promoter analysis based on transcription factor binding sites. Bioinformatics 21, 2933–2942 (2005).
Khan, A. et al. JASPAR 2018: update of the open-access database of transcription factor binding profiles and its web framework. Nucleic Acids Res. 46, D1284 (2018).
Kent, W. J. et al. The human genome browser at UCSC. Genome Res 12, 996–1006 (2002).
Grant, C. E., Bailey, T. L. & Noble, W. S. FIMO: scanning for occurrences of a given motif. Bioinformatics 27, 1017–1018 (2011).
Mathelier, A. et al. JASPAR 2016: a major expansion and update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 44, D110–115 (2016).
Schmitges, F. W. et al. Multiparameter functional diversity of human C2H2 zinc finger proteins. Genome Res. 26, 1742–1752 (2016).
Ghiasvand, N. M. et al. Deletion of a remote enhancer near ATOH7 disrupts retinal neurogenesis, causing NCRNA disease. Nat. Neurosci. 14, 578–586 (2011).
Guerrier, S. et al. The F-BAR domain of srGAP2 induces membrane protrusions required for neuronal migration and morphogenesis. Cell 138, 990–1004 (2009).
Hashimoto-Torii, K. et al. Interaction between Reelin and Notch signaling regulates neuronal migration in the cerebral cortex. Neuron 60, 273–284 (2008).
Sousa, A. M. M. et al. Molecular and cellular reorganization of neural circuits in the human lineage. Science 358, 1027–1032 (2017).
Kwan, K. Y. et al. Species-dependent posttranscriptional regulation of NOS1 by FMRP in the developing cerebral cortex. Cell 149, 899–911 (2012).
Blanchette, M. et al. Aligning multiple genomic sequences with the threaded blockset aligner. Genome Res. 14, 708–715 (2004).
Tan, G. & Lenhard, B. TFBSTools: an R/bioconductor package for transcription factor binding site analysis. Bioinformatics 32, 1555–1556 (2016).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
El-Gebali, S. et al. The Pfam protein families database in 2019. Nucleic Acids Res. 47, D427–D432 (2019).
UniProt Consortium UniProt: the Universal Protein Knowledgebase in 2025. Nucleic Acids Res. 53, D609–D617 (2025).
Arendsee, Z. et al. phylostratr: a framework for phylostratigraphy. Bioinformatics 35, 3617–3627 (2019).
Tautz, D. & Domazet-Loso, T. The evolutionary origin of orphan genes. Nat. Rev. Genet. 12, 692–702 (2011).
Domazet-Loso, T. et al. No evidence for phylostratigraphic bias impacting inferences on patterns of gene emergence and evolution. Mol. Biol. Evol. 34, 843–856 (2017).
Weber, J. A. et al. The whale shark genome reveals how genomic and physiological properties scale with body size. Proc. Natl Acad. Sci. USA 117, 20662–20671 (2020).
Schoch, C. L. et al. NCBI taxonomy: a comprehensive update on curation, resources and tools. Database https://doi.org/10.1093/database/baaa062 (2020).
Kumar, S. et al. TimeTree 5: an expanded resource for species divergence times. Mol. Biol. Evol. https://doi.org/10.1093/molbev/msac174 (2022).
Acknowledgements
We thank members of the Heng, Richards and Sestan laboratories for their helpful comments. We also thank C. Cepko, E. Grove, F. Polleux and P. Rakic for providing reagents. This study was supported by the Australian Research Council (DP 160103958 to R.S. and L.J.R. and Discovery Early Career Researcher Award Fellowship 160101394 to R.S.), the National Health and Medical Research Council (Principal Research Fellowship to L.J.R. and C.J. Martin Fellowship to P.K.), the Australia Postgraduate Award (to L.R.F.), the UQ-QBI Doctoral Scholarship (to A.P.), the Victorian Government through the Operational Infrastructure Scheme and the National Health and Medical Research Council of Australia (ID 1011505 to J.I.-T.H.), Instituto de Salud Carlos III Spain and European Social Fund grant MS20/00064 (to G.S.), grants PID2019-104700GA-I00 and PID2022-140137NB-100 funded by MICIU/AEI/10.13039/501100011033 and co-funded by FEDER, European Union (to G.S.), Fundació LaMarató de TV3 (to G.S.), NIH grants (MH129981 to K.T.G., NS095654, MH106934, MH116488, MH110926, MH122678, MH124619 and HG010898) and the Simons Foundation 736706 (to N.S.).
Author information
Authors and Affiliations
Contributions
N.S. conceived and designed the study. Z.L., N.K., S.K.S., C.Q. and M.S. performed most of the genetic, molecular and other experiments described in this study. S.K.M., G.S. and X.d.M. performed most of the computational, bioinformatic and related analyses for this study. M.L., X. Xu, P.K., R.S., V.L., A.K., H.C., S.M., S.-K.K. performed further computational analysis. Z.L., O.C., A.S., I.K.S., T.H. and F.O.G. conducted ISH experiments. Z.L., N.K., S.K.S., A.M.M.S., R.D.R., P.Q.D., A.T.N.T., T.S.K., W.H., A.P. and L.R.F. performed analysis of immunohistochemistry or protein content. Z.L., W.H., S.K.S. and Y.B. performed IUE experiments. S.-H.C., H.K. and S.K.S. performed tissue culture experiments for chicken. Z.L., A.T.N.T. and S.P. developed/acquired human tissue resources. Z.L., X. Xing, I.E.G.-N. and H.D.C. developed/acquired mouse model resources. Z.L., S.K.M., G.S., F.O.G. and N.S. prepared and wrote the initial draft of the paper. All authors provided edits and/or comments on subsequent drafts. F.O.G., N.K., A.M.M.S., K.T.G., L.J.R., J.I.-T.H. and N.S. provided supervision and guidance for this study. L.J.R., J.I.-T.H., G.S. and N.S. obtained and provided funding for this study.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Tomomi Shimogori and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Integration of RNA-Seq and ChIP-Seq to Identify Gene Enrichment and CREs in IT and ET Neurons.
a-b) Venn diagrams illustrating the overlap between genes differentially expressed and genes regulated by differentially enriched H3K27ac peaks between Arpp21- and Fezf2- labeled cells. b) As in A but representing overlap of H3K27ac peaks rather than genes. c) Most genes and H3K27ac peaks were not differentially expressed or enriched, respectively, between IT neurons and ET neurons (left panel). However, certain genes were enriched in either IT neurons or ET neurons and associated with putative CREs enriched or present in that same cell subtype (middle and right panels). Data shown represent ChIP-Seq data collected from 2 independent biological replicates per condition. d) Bar graph showing the top 15 gene ontology terms in the H3K27ac peaks either present in both (grey) or in Arpp21-Gfp + (green) or in Fezf2-Gfp + (red) cells but are absent in the H3K27ac peaks from E 17 chicken. The X-axis depicts the log2 P-value. e) Schematics showing different areas of the chicken brain across the anterior-posterior axis in the coronal plane. Coronal sections of chicken brain showing ZBTB18, Cux2, Satb2, BCL11B, and Fezf2 expression domain ZBTB18 and BCL11B images showing ZBTB18 and BCL11b immunostaining are inverted to enhance visualization. In inset Cux2 in situ expression signal is shown at posterior plane. This demonstrates the Cux2 probe’s sensitivity to chicken Cux2. Scale bar: ZBTB18, Cux2, (500 µm); Satb2, BCL11B, Fezf2 (250 µm). Hp, hippocampus; HA, hyperpallium apicale; M, mesopallium; N, nidopallium; E, entopallium; Str, striatum; D, dorsal; L, lateral.
Extended Data Fig. 2 CREs bound by ZBTB18 at the Genomic Loci of Cux2, Satb2, Bcl11b and Zbtb18 and Luciferase Reporter Assay.
a-d) Line graphs showing the H3K27ac peaks from, Arpp21-Gfp+ cells (line 1 and 2), Fezf2-Gfp+ cells (line 3 and 4), from PCD 14.5, and PD 0 forebrain DNaseI Hypersensitivity tracks (DHS) by Digital DNaseI from ENCODE, (line 5 and 6, respectively) and mammal conservation as reference (line 7), ZBTB18-HA ChIP-seq peaks from mouse (line 8), H3K27ac peaks from fetal (dlPFC (line 9), gene locus in mice (line 10) H3K27ac peaks collapsed together in the Arpp21-Gfp + IT neurons and Fezf2-Gfp + ET neurons (line 11) and H3K27ac peaks from E17 chicken dorsal pallium (12) at the genomic locus of Cux2, Satb2, Bcl11b and Zbtb18. The H3K27ac and ZBTB18- ChIP peaks in mice are shown in light beige. In the chicken genome, the regions orthologous to the ZBTB18-ChIP peaks are shown in a dark beige color, with the dotted blue boxes indicating the exact orthologous regions obtained after liftover.
Extended Data Fig. 3 Assessment of Cux2 Enhancer Activities Through Mouse Transgenic Reporter Assays.
a) Transgenic mouse Cux2-E1-Gfp shows no GFP expression at PCD 12.5, but strong GFP expression at PCD 14.5 GFP in the embryonic mouse forebrain. Scale bar: 1 mm (left), 2 mm (right). b) At PCD 17.5, GFP is expressed in the BCL11B negative cells in upper CP and CC in Cux2-E1-Gfp mice. Scale bar: 1 mm (whole brain), 50 µm (right). c) At PD 0, GFP is mostly expressed in BCL11B negative upper layer neurons and absent from CST as seen at pons in Cux2-E1-Gfp mice. Scale bar: 1 mm (whole brain), 50 µm (right). (d-f). At PD 15, the majority of Cux2-E1-Gfp positive cells reside in upper layers and co-label with CUX1 (closed arrows). Double open arrowheads (CUX1) and triple open arrowheads (BCL11B) again indicate cells immunolabeled for specific molecular markers but not by Cux2-E1-Gfp. CUX1 and BCL11B positive cell populations were compared using an unpaired, two-tailed t-test. The graph represents mean ± s.e.m. n = 1,116 cells from 3 biological replicates * P = 0.000352. Scale bars: 100 µm (e); inset, 50 µm (f) ;100 µm. (g) In the PCD 17.5, Cux2-E3-Gfp transgenic mice brain, GFP is mostly expressed in L1 cell and co-localize with RELN, but not with BCL11B, GFP labeled axons can be seen passing through CC. Scale bar: 100 µm (left, right), 20 µm (middle). (h) In the PCD 17.5 Cux2-E2-Gfp transgenic mice brain, GFP-positive cells are scattered throughout CP and largely co-localize with LHX6; GFP is not expressed in CC. Scale bar: 100 µm (left, right), 20 µm (middle). i) Cux2 overexpression increases neurite length of the SATB2 and ZBTB18 immunopositive developing chicken neurons. The image shows chicken neurons from E7 chicken embryonic forebrain overexpressing Cux2 using pCag-Cux2 and pCaggs-mCherry to label electroporated neurons. Scale bar: 100 µm. n = 43 neurons for control & n = 24 for Cux2 OE. j) The graph represents the length of neurites of the pCag-Cux2 + pCaggs-mCherry neurons that co-express SATB2 and ZBTB18 over controls pCagen + pCaggs-mCherry. Ordinary two-way ANOVA with Bonferroni’s multiple comparisons test, with single pooled variance, was applied. The graph represents mean ± s.e.m. n = 210 from control and 139 from Cux2 overexpressing cells from 3 experimental replicates. **** P = 0.0001. k) Luciferase reporter activity driven by the Satb2 E1 enhancer is significantly increased by ZBTB18 and reduces when the ZBTB18 binding site is mutated (Satb2 ΔE1). The graph represents mean ± s.e.m. Ordinary two-way ANOVA with Bonferroni’s multiple comparisons test, with single pooled variance, was applied. **** P = 0.0001 (Satb2 E1 (Control vs Zbtb18)), (Satb2 ΔE1 (Control vs Zbtb18)), and P = 0.003 for Satb2 E1 vs Satb2 ΔE1 (Zbtb18), n = 3.
Extended Data Fig. 4 ZBTB18 Expression in the Developing Cerebral Cortex of Humans and Mice.
a) Coronal sections showing the immunolabeling for three anti- ZBTB18 antibodies on Neurod6-Cre; Zbtb18 cKO as compared to the control. Arrow points to the expression of ZBTB18 in the ventral region of the cKO where Neurod6-Cre appears not be active. Scale bar: 150 µm. b) Zbtb18 mRNA are co-expressed with Cux2 mRNA in PD 0 mouse neocortex in L2-3 (arrows) as well as in L5 (arrowheads). Scale bar: 1 mm, 20 µm (right). c) ZBTB18 mRNA is co-expressed with CUX2 mRNA (arrow) in 20 PCW human frontal cortex. Scale bar: 2 mm, 40 µm (right) CP, cortical-plate; IZ, intermediate zone; SVZ, subventricular zone; VZ, ventricular zone. All images of mouse neocortex are representative of at least 3 independent biological replicates. In situ hybridization of a postmortem postconception weeks (PCW 20) human neocortex reflects multiple sections from a single brain but is consistent with an additional PCW 18 postmortem human brain sample (not shown).
Extended Data Fig. 5 Enrichment of ZBTB18 in Postmigratory IT Neurons.
a) Dot-plot showing expression of Zbtb18 and other canonical layer markers across ExN types along developmental ages of mouse cortex27. Zbtb18 co-expresses Satb2 and Cux2 in IT neurons but not with ET neurons’ TFs Bcl11b, Fezf2, Sox5, Foxp2, and Zfpm2. b-c) ZBTB18 co-localizes with SATB2, but not BCL11B, in the 20 PCW human neocortex. Scale bar: 100 µm. Immunofluorescent analysis of the mouse brain utilized independent sections from at least 3 animals. Images and quantification shown in A are representative of analysis conducted on at least 2 sections from the postmortem human brain. The graph represents mean ± s.e.m. An unpaired two tailed t-test was applied. * P = 0.0001, n = 3.
Extended Data Fig. 6 Downregulation of Essential IT Neuron Marker Genes in Mice Lacking ZBTB18.
a-b) Bulk RNA-Seq of PCD 14.5 and PD 0 neocortex was performed in whole body Zbtb18 -/- knockout (KO) and Zbtb18 +/- (Control) mice and in Neurod6-Cre; Zbtb18fl/fl (Neurod6-Cre; Zbtb18 cKO) and Neurod6-Cre; Zbtb18fl/+ (Neurod6-Cre; Zbtb18 cHET) (Control) respectively. 82.3% of genes upregulated in Arpp21-Gfp+ cells were downregulated in the Neurod6-Cre; Zbtb18 cKO [93 of 113], while 92.4% of genes enriched in Fezf2-Gfp+ cells were upregulated in the cKO [219 of 237]. c) In situ hybridization shows mRNA expression of upper layer marker genes Cux1, Satb2, and Rorb. Scale bar: 150 µm. d) The size of the dorsal brain of Neurod6-Cre; Zbtb18fl/fl (cKO) mice don’t change at PD 0 but appears to be smaller at PD 8 as compared with Neurod6-Cre; Zbtb18fl/+; mice. (e) Immunofluorescence staining of ZBTB18 is shown in the PCD 15.5 Neurod6-Cre; Zbtb18 cKO brain, and ZBTB18 expression is lost in postmitotic neurons but not SVZ progenitors. f) Western blot shows ZBTB18 protein expression is abolished, and CUX2 expression is significantly diminished in PD 0 Emx-Cre; Zbtb18 cKO mouse brain. g) Quantified signal intensity shows CUX2 expression is reduced by 70% in Emx-Cre; Zbtb18 cKO mouse brain compared with control. The graph represents mean ± s.e.m. n = 3 biological replicates. ZBTB18, CUX2 and GAPDH were run on individual gels with equal amount of lysate from same samples to avoid overlapping of smaller isoforms. Raw image of uncropped gel is in Supplementary Fig. 1. An unpaired t-test was used to compare groups. * P = 0.0465. h) Cux2 mRNA expression is significantly reduced in the neocortical plate but not the striatum of Zbtb18 knockout mice at multiple developmental periods. 150 µm. i) CUX2 expression in control and ZBTB18 cKO brains detected by CUX2 antibody. Representative image of coronal sections of the P0 brains stained with anti-Cux2 antibody are shown. Upper panel: Neurod6-Cre + ; Zbtb18fl/+; Lower panel: Neurod6-Cre + ; Zbtb18fl/fl. Scale bar: 500 µm. CP, cortical plate; IZ, intermediate zone; SP, subplate; SVZ, subventricular zone; VZ, ventricular zone. All immunofluorescent analyses and in situ hybridization represent images selected from multiple sections from at least 3 independent biological replicates. Western blotting is representative of 3 independent biological replicates.
Extended Data Fig. 7 Impact of Postmitotic Depletion of ZBTB18 on IT Neuron Development and Upper Layer Formation.
a) Top: IdU labeling (red) expected deep layer, BCL11B-immunopositive neurons at PCD 12.5 and immunofluorescence analysis with BCL11B (green) at PD 0 shows a greater number and broader distribution of BCL11B- positive neurons without a concomitant change in BCL11B and, IdU double-positive neurons. Bottom: Quantification of cells per laminar position (bins). b) Similar to A, with IdU labeling putative upper layer neurons at PCD 14.5, we observed a significant increase in the number of BCL11B and IdU double-positive neurons at PD 0. c) Similar to a and b, with CIdU labeling (green) expected upper layer neurons at E15.5 and immunofluorescence analysis with SATB2 (red) indicating upper layer neurons at P1. Zbtb18 deficient cells exhibited an alteration in the distribution of SATB2-immunopositive neurons that are also CIdU- positive relative to control mice. For each experiment, 145 to 356 cells from independent sections from at least three different mice were analyzed. Two-tailed T-tests were used to compare control and knockout cell counts per bin. * Bonferroni corrected P-value < 0.05. Please refer to the Supplementary Table 18 for detailed p values. Scale bar: 100 µm. d-e) In early postnatal Emx1-Cre; Zbtb18fl/fl mice brain, layers 2-4 are absent, BCL11B- positive cells populate the top layers of the neocortex, CUX1 expression is absent from upper layers, but present in SP. Scale bars: 50 µm. All analyses depicted in this figure were conducted on at least 3 independent biological replicates.
Extended Data Fig. 8 ZBTB18 Depletion Leads to Increased Intrahemispheric Cortico-Cortical Projections into the mPFC.
a) Coordinates (33) of bins used to analyze tracing experiments depicted in b. b) Coronal sections of traced PD 7 brains showing more intensive GFP labeling in GI, S2, S1Ul in Neurod6-Cre/+; Zbtb18flfl cKO mice than control. GI, granular insular cortex; S2, secondary somatosensory cortex; S1UL, primary somatosensory cortex, upper lip. Scale bar: 500 µm.
Extended Data Fig. 9 ZBTB18 Enhances Robo1 E1 Activity through Direct Binding.
a) Line graphs showing the H3K27ac peaks from, Arpp21-Gfp+ cells (line 1 and 2), Fezf2-Gfp+ cells (line 3 and 4), from PCD 14.5, and PD 0 forebrain DNaseI Hypersensitivity tracks (DHS) by Digital DNaseI from ENCODE, (line 5 and 6, respectively) and mammal conservation as reference (line 7), ZBTB18-HA ChIP-seq peaks from mouse (line 8), H3K27ac peaks from fetal dlPFC (line 9), gene locus in mice (line 10) H3K27ac peaks collapsed together in the Arpp21-Gfp + IT neurons and Fezf2-Gfp + ET neurons (line 11) and H3K27ac peaks from E17 chicken dorsal pallium (12) at the genomic locus of Robo1. The regions detected by H3K27ac and ZBTB18- ChIP seq are in light beige color.
Extended Data Fig. 10 ZBTB18 Binding Motifs Exhibit High Conservation in Putative CREs of Eutherian IT Neurons.
a) Taxonomic tree of the 23 Chordate species ZBTB18 SST protein sequence features of ZBTB18 were examined. For every species, the Pfam architecture of protein domains is indicated to the right. b) The ZBTB18 binding motif, as well as 16 other motifs from the JASPAR database, have consensus sequences enriched in regulatory regions related to genes downregulated in the Zbtb18 KO that are specifically conserved in placental mammals. Fisher’s exact test was used to test for an enrichment of conservation. Red dots represent motifs with a False Discovery Rate corrected P-value < 0.05. For detailed statistics, please refer to Supplementary Table 17. c) Evolutionary divergence of the key enhancers and/or ZBTB18 binding motifs. The evolutionary divergence of ZBTB18 motifs in key enhancers (Zbtb18 P1, Cux2 E1, Cux2 E4, Robo1 E1, Satb2 E1, and Bcl11b P1) are plotted onto a phylogenetic tree that shows the divergence of placental mammals (black line), marsupials (red line), monotremes (green line) and non-mammal vertebrates (purple line). All three motifs in Zbtb18 P1(Zbtb18 P1_motif1, Zbtb18 P1_motif2, Zbtb18 P1_motif3) and Cux2 E1_motif2 (purple) had evolved before the divergence of monotremes from non-mammal vertebrates, and Satb2 E1_motif2 appeared between the divergence of monotremes to the rest of mammals. In contrast, Cux2 E1_motif1, Satb2 E1_motif1, and all motifs in Cux2 E4, Robo1 E1, and Bcl11b P1 appeared after the divergence of placental mammals from marsupials. MYA: million years ago. d) Bootstrap-based strategy to evaluate the robustness of motif enrichments. The percentile occupied by the observed ratio represents an empirical P-value (Red dash line), indicating how likely it is to obtain this ratio by chance among the invariant peaks. The categories IT Placentalia, IT Marsupialia, and KO up Placentalia present significantly extreme observed ratios (P = 0.005, P = 0.003, and P < 0.001, respectively).
Extended Data Fig. 11 Evolutionary footprints analysis of ZBTB18 motifs in key CREs.
DNA sequence from mouse Zbtb18 P1, Cux2 E1, Cux2 E4, Robo1 E1, Satb2 E1, and Bcl11b P1 are analyzed for ZBTB18 motif discovery, including non-canonical motifs. For Regex motif, identifying the canonical “CAGATGT” pattern allowing one mismatch, only the core sequence is shown. Multi-species alignment was performed by comparing ZBTB18 motifs across 60 vertebrate species, which were sub-grouped in placental mammal (black), marsupial (red), monotreme (green), and non-mammal vertebrate (purple). Gaps are represented by black vertical bars. a) Three Jaspar motifs are detected in Zbtb18 P1, all of which show a relatively high level of conservation in a majority of the 60 species examined, but they are less conserved in species belonging to the fish clades. b) Two Regex motifs are detected in Cux2 E1, and Cux2 E1_ motif1 shows universal conservation in placental mammals, and 1 bp mutation is observed in marsupials, but the motif is lost in non-mammal vertebrates because of gaps (black vertical bar, panel b). Cux2 E1_ motif2 is less conserved with 1 bp mutation across mammalian species. c) Two motifs are detected in Cux2 E4, one is Jaspar motif (red) and the other is Regex motif (blue). d) Three motifs are detected in Robo1 E1, one is Jaspar motif (red), one is Regex motif (blue), and the other is detected by both Regex(blue) and MEME-TOP (#). All three motifs are only present in placental mammals. e) Two Jaspar motifs(red) and one PROMO (*) /Regex(blue) motif are detected in Satb2 E1, showing high level of conservation in mammals. All motifs are not present in monotremes or non-mammal vertebrates. f) Four motifs are detected in Bcl11b P1, one being Jaspar motif(red), two being Regex motifs(blue), and the last one being MEME-TOP (#). While motif1 and motif2 show a higher level of conservation, motif3 and motif4 have more sequence variation across mammals. Jaspar consensus motif (Red). Regular expression motif (regex motif, Blue). * PROMO motif. # MEME-TOP motif. The E-box core sequences are highlighted in rectangles.
Supplementary information
Supplementary Information (download PDF )
Supplementary Discussion and Figs. 1 and 2.
Supplementary Tables (download XLSX )
Supplementary Tables 1–18.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, Z., Kaur, N., Santpere, G. et al. Adaptive evolution of gene regulatory networks in mammalian neocortex. Nature 653, 156–166 (2026). https://doi.org/10.1038/s41586-026-10226-y
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-026-10226-y
