
Abstract
Most studies to date of protein quantitative trait loci (pQTLs) have relied on affinity proteomics platforms, which provide only limited information about the targeted protein isoforms and may be affected by genetic variation in their epitope binding. Here we show that mass spectrometry (MS)-based proteomics can complement these studies and provide insights into the role of specific protein isoform and epitope-altering variants. Using the Seer Proteograph nanoparticle enrichment MS platform, we identified and replicated new pQTLs in a genome-wide association study of proteins in blood plasma samples from two cohorts and evaluated previously reported pQTLs from affinity proteomics platforms. We found that >30% of the evaluated pQTLs were confirmed by MS proteomics to be consistent with the hypothesis that genetic variants induce changes in protein abundance, whereas another 30% could not be replicated and are possibly due to epitope effects, although alternative explanations for nonreplication need to be considered on a case-by-case basis.
Similar content being viewed by others


Main
Protein quantitative trait loci (pQTLs) are important tools in drug target discovery and for generating new hypotheses regarding protein function1,2,3,4,5. Most pQTL studies to date have relied on affinity proteomics platforms6,7,8,9,10,11,12,13,14,15, which provide only limited information about the targeted protein isoforms and may be affected by genetic variation in their epitope binding. Mass spectrometry (MS)-based proteomics can complement these studies and provide insights into the role of specific protein isoform and epitope-altering variants. Here we report a genome-wide association study (GWAS) to detect pQTLs using an MS-based proteomics platform with blood plasma samples from a discovery cohort of 1,260 individuals from the USA and a replication cohort of 325 individuals from Asia. We analyzed 1,980 proteins that were quantified in at least 80% of the samples and identified 364 pQTLs, of which 102 were replicated; among these, 35 have not been reported previously. We further investigated cis-pQTLs identified by previous affinity proteomics GWASs for possible epitope effects. In our dataset, 30% of the evaluated pQTLs were confirmed by MS proteomics to be consistent with the hypothesis that genetic variants induce changes in protein abundance, whereas another 30% could not be replicated and are possibly due to epitope effects, although alternative explanations for nonreplication need to be considered on a case-by-case basis. Our study demonstrates the complementarity of the different proteomics approaches and reports pQTLs of biomedical relevance that are not accessible through affinity proteomics, suggesting that many more pQTLs remain to be discovered using MS-based technologies.
The development of high-throughput affinity proteomics platforms has spurred an increasing number of GWASs with protein traits (see ref. 16 for a comprehensive list). The largest published pQTL studies to date come from deCODE, utilizing the SOMAscan platform with 4,907 aptamers in 35,559 samples from Icelanders17, and from the UK Biobank Pharma Proteomics Project (UKB-PPP) consortium using the Olink platform with dual antibodies targeting 2,923 proteins in 54,219 samples from UK Biobank participants18. A few MS-based GWASs have also been reported but were limited to a smaller number of proteins19,20,21. Collectively, these GWASs have reported thousands of pQTLs that are now available for further exploration such as through Mendelian randomization experiments15 to identify new drug targets and to further the development of protein-based biomarkers16.
However, it is important to note that pQTLs discovered using affinity proteomics methods represent genetic associations with protein-binding affinity rather than direct protein abundance, presuming a reproducible link between the number of reagents binding to their targets and the target’s abundance. This link may break down when a protein-altering variant (PAV) is located at the aptamer-binding or antibody-binding site, leading to a genotype-dependent readout that does not correspond to a real change in protein abundance9.
For example, a study on blood pressure identified a strong pQTL for circulating natriuretic peptide precursor A associated with a protein-coding variant (rs5063), but this finding failed to replicate in a sixfold larger study22. The authors concluded that the association was artefactual, because the discovery study used an antibody against an epitope in the midregion of the molecule, in contrast to the amino-terminal epitopes used in the replication study.
Such epitope effects can invalidate conclusions drawn from Mendelian randomization experiments, because their basic hypothesis requires that changes in the exposure, that is, the protein abundance, are causal for changes in the disease outcome. Epitope effects can also skew the prediction of protein levels using polygenic scores and confound correlations with other -omics modalities. Therefore, it is important to validate key pQTLs on an independent platform that is not susceptible to the same epitope-binding effects.
However, it should be noted that MS methods do not measure protein abundance directly either, but infer it indirectly from peptide abundances, which are proportional to the amount of digested and ionized protein fragments and their mapping to the protein isoforms that are assumed to be found in the sample.
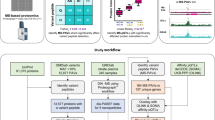
We previously demonstrated that MS proteomics readouts can reliably distinguish between epitope QTLs and protein abundance QTLs by applying a specific data analysis protocol that excludes PAV-containing peptides from the protein quantification23. In the present study, we extended this approach to a larger study cohort and conducted a full GWAS using the MS-based Proteograph proteomics platform (Seer)24,25.
The Seer technology enhances proteome coverage by using nanoparticle enrichment, followed by a data-independent acquisition protocol implemented on a Bruker timsTOF Pro 2 mass spectrometer (Bruker Daltonics). The following analysis focused on 1,980 proteins that were quantified in at least 80% of the samples (Supplementary Table 1 and Supplementary Fig. 1), out of 5,753 proteins quantified across a discovery cohort (Tarkin) of 1,260 US study participants of diverse backgrounds and a replication phase (Qatar Metabolomics study of Diabetes (QMDiab)) comprising 325 samples from participants of mainly Arab, Indian and Filipino backgrounds (Supplementary Fig. 2).
Results
Identification of protein QTLs
In the discovery stage (Tarkin), 364 independent protein associations reached Bonferroni’s level of significance (P < 5 × 10−8), involving 295 genetic loci and 274 different proteins, with 177 of these associations located in cis. Replication was attempted using 325 samples from the QMDiab study, which included matching genotype and proteomics data. To account for differences in genetic structure between the cohorts, a pQTL was considered replicated if it colocalized between the discovery and replication studies and reached a genome-wide and proteome-wide significance level ofP < 2.53 × 10−11(5 × 10−8 ∕1,980) in the joint analysis. A total of 102 pQTLs (28.0%) met these criteria. All replicated pQTLs exhibited concordant effect direction (Figs. 1 and 2, Supplementary Table 2, Supplementary Figs. 3 and 4 and Supplementary Data 1 and 2).

Grid plot of the genomic position of the variant (SNP position) versus the position of the gene coding for the pQTL protein (protein position). The cis-pQTLs are in red and the trans-pQTLs in blue (plot data in Supplementary Table 2).

a–c, Scatterplots of effect size (β) versus minor allele frequency (MAF) (a), pQTL effect size (β) from the discovery (Tarkin) versus from the replication (QMDiab) study (b) and EAFs for Tarkin versus QMDiab (c). All the protein associations that reached a significance level P < 5 × 10−8 in the discovery study are represented, except for b, where only replicated pQTLs are shown (plot data in Supplementary Table 2).
The primary reason for the nonreplicated pQTLs is the limited size of the replication cohort: of 70 pQTLs that had 80% replication power, 58 (82.9%) replicated and most (36 out of 38) of the nominally significant (P < 0.05) pQTLs had concordant directionality, suggesting that most of the unreplicated pQTLs should be replicable in future larger-scale studies. In addition, 14 of the nonreplicated pQTLs exhibited a significant but different genetic signal in both cohorts, such as intelectin-1 (ITLN1). The ITLN1 pQTL overlaps with a Crohn’s disease risk locus, but the role of ITLN1 in the disease mechanism is not clear26. We detected a strong ITLN1 signal in the QMDiab cohort that was not present in Tarkin, which may be discernible only in that cohort due to differences in lifestyle, environmental factors or population-specific genetic backgrounds (Supplementary Fig. 5). It would therefore be interesting to analyze the genetic architecture of this locus in future Crohn’s disease GWASs in different populations.
Of the 102 replicated pQTLs, 53 and 52 had been identified previously by the deCODE SOMAscan study17 and the UK Biobank Pharma Proteomics Project (UKB-PPP) Olink study18, respectively, and 67 (65.7%) had been reported at least once by these and/or other pQTL studies curated by Open Targets27. A total of 35 replicated pQTLs (34.3%) were new (Table 1), although 11 of the new MS pQTLs were for proteins that had been assayed by the deCODE and/or UKB-PPP studies, but did not reach genome-wide significance at these loci, suggesting that the respective affinity assays may be targeting different isoforms, may not be reaching their detection limits or may be binding off-targets.
Biomedical relevance of new pQTLs
The 35 new pQTLs overlap with several loci of biomedical relevance, including a trans-pQTL for ANGPTL6 at the COLEC11 locus associated with low-density lipoprotein (LDL)-cholesterol levels, a trans-pQTL for BRE (brain and reproductive organ expressed (TNFRSF1A modulator)) at the CFH locus associated with age-related macular degeneration and immunoglobulin (Ig)A nephropathy, a cis-pQTL for galactosylceramidase associated with inflammatory bowel disease and many others that can now be considered as candidate drug targets for these diseases.
We also identified pQTLs that complement findings from affinity-based studies, such as a trans-pQTL on the Olink platform for fucosidase FUCA1 (rs11155297) that we replicated. In addition, we found a cis-pQTL for FUCA2 at the same locus, which was not assayed by the Olink platform, but would be expected to account for the trans-association (Supplementary Table 2). It is interesting that the strongest association by SOMAscan at this locus was with mannosidase MAN2B2. FUCA1, FUCA2 and MAN2B2 are all enzymes involved in the lysosomal degradation of glycoproteins and glycolipids. A look-up using the Open Targets platform additionally revealed a GWAS signal for ‘Total PHF-tau (SNP × SNP interaction) (P = 2 × 10−8)28. These genetic signals obtained from three different proteomics platforms illustrate how pQTLs can be leveraged to generate hypotheses for the drug target discovery process.
Analysis of pQTLs reported by orthogonal platforms
We then asked whether previously reported affinity pQTLs could be confirmed using an orthogonal MS technology and whether any of these pQTLs could be affected by epitope effects. We examined 319 and 392 cis-pQTLs from the deCODE SOMAscan study17 and the UKB-PPP Olink study18, respectively, for which matching genotype and protein data were available in our study (Supplementary Tables 7–10). For these pQTLs, we conducted a genetic analysis of the MS proteomics data at the peptide level and computed summary statistics for the associations of all peptides from a given pQTL protein with the respective pQTL variant. To integrate these summary statistics, we employed two approaches: first, a meta-analysis across all peptides (Supplementary Figs. 6 and 7 and Supplementary Data 3–5) and, second, the derivation of an MS-based peptide association (MSPA) score that represents the support for a true protein abundance pQTL at the peptide level.
The MSPA score is defined as the fraction of peptides that support a pQTL at an α level of 1%, weighted by the number of detections of each peptide (Methods). An MSPA score of 1 indicates that, for all detected peptides, the 99% confidence interval (CI) of the effect (β) does not contain the null, thereby supporting a protein abundance QTL. Conversely, an MSPA score of 0 indicates that none of the peptides from the protein provides statistical support for an association. We argue that the MSPA score is a more intuitive measure for the support of a genuine pQTL at the peptide level than relying on the overall statistical significance at the protein or the peptide meta-analysis level. We therefore focus on the MSPA scores in the following discussion (Fig. 3) and provide additional measures and comparisons of their properties in Supplementary Figs. 8–12. A total of 52 out of the 319 SOMAscan pQTLs (16.3%) and 62 of the 392 Olink pQTLs (15.8%) had an MSPA score ≥0.8.

a–g, Scatterplots of the power to replicate a pQTL from the deCODE SOMAscan (a) and the UKB-PPP Olink (e) studies against the ranks of the affinity proteomics pQTLs. Scatterplots of individual MSPA scores against the ranks of the affinity proteomics pQTLs of the deCODE SOMAscan (b) and the UKB-PPP Olink (f) studies, starting with the lowest P value: 120 out of 319 pQTLs (37.6%) for deCODE and 167 out of 392 pQTLs (42.6%) for UKB-PPP had >99% power at a significance level of P < 0.05/319 and P < 0.05/319 for SOMAscan and Olink, respectively, colored to indicate likely protein abundance QTLs (MSPA score >0.8; green) and likely epitope effect-driven pQTLs (MSPA score <0.2; red). MSPA scores were limited to 46 pQTLs that were reported on the same variant in deCODE (c) and UKB-PPP (g). Scatterplots of the effect size (β) of the 46 pQTLs reported deCODE and UKB-PPP (d) (plot data in Supplementary Tables 9, 10 and 12).
Identification of an epitope effect, however, is more challenging, because lack of support for an affinity proteomics pQTL using MS methods may also result from other factors, including insufficient statistical power, limited sensitivity of the MS method, differences between targeted isoforms between platforms or the possibility that genetic variants represent different genetic signals between study populations.
To determine which affinity pQTLs could be expected to be replicated using our MS proteomics data, we conducted a power analysis (Methods). A total of 120 (for deCODE) and 167 (for UKB-PPP) pQTLs had >99% probability of reaching a multiple-testing corrected P value of P < 0.05/319 (for deCODE) and P < 0.05/392 (for UKB-PPP) in Tarkin. Of these sufficiently powered pQTLs, 39 (32.5% for deCODE) and 49 (29.3% for UKB-PPP) replicated and had an MSPA score >0.8, whereas 36 (30.0% for deCODE) and 55 (32.9% for UKB-PPP) did not replicate and had an MSPA score <0.2 (Supplementary Tables 9 and 10). Our analysis also confirms a previously reported29 epitope effect for the pQTL of GDF15, which was reported by deCODE (rank 71 in Supplementary Table 9) and UKB-PPP (rank 133 in Supplementary Table 10) and exhibited an MSPA score of 0 in our study.
Genetic association with protein isoforms
Conflicting directionality of QTLs at the peptide level can indicate the concurrent presence of different isoforms. We identified ten such cases in deCODE and eight in UKB-PPP (Supplementary Tables 9 and 10). One example is variant rs2052534, which is a pQTL for the Serine Peptidase Inhibitor Kazal Type 5 (SPINK5) reported by both UKB-PPP and deCODE, as well as in our study on variants in linkage disequilibrium (LD) (r2 = 1). The pQTL had an MSPA score of 0.5, indicating support by some, but not all, analyzed peptide associations. SPINK5, also known as Lymphoepithelial Kazal-Type-Related Inhibitor or LEKTI, plays a role in skin and hair morphogenesis and protection of the mucous epithelia. Mutations in SPINK5 have been linked to skin disorders characterized by ichthyosis30, such as Netherton’s syndrome, as well as to hair abnormalities. GWASs have further associated SPINK5 variants with lung phenotypes (for example, chronic obstructive pulmonary disease, forced expiratory volume) and pancreatitis. It has been previously shown that that the SPINK5 gene generates three classes of transcripts encoding three different LEKTI isoforms, which vary in their carboxy-terminal portion31, corresponding to three UniProt entries: the 1,064 amino acid canonical form Q9NQ38, isoform Q9NQ38-3 that contains a 30-amino acid insert at position 915 and isoform Q9NQ38-2, which is truncated at position 913, followed by an additional three amino acids (Fig. 4). A fourth isoform (E5RFU9) is truncated at position 158 with an additional 43 amino acids and is likely to be proteolytically degraded. The additional amino acids of Q9NQ38-2 and E5RFU9 were not detected in the MS analysis. Consequently, four protein groups were generated (color coded black, green, red and blue in the Forest plot for the meta-analysis in Fig. 4). These different protein groups associated with the genetic variant rs2052534 at different strengths (for a multiple alignment of the detected peptides and the SPINK5 isoforms, see Supplementary Table 11). These observations can be explained parsimoniously by a genetic variant that influences splicing near residue Lys913, thereby increasing the generation of Q9NQ38-3 isoforms while decreasing that of Q9NQ38. Supporting this hypothesis, the Genotype-Tissue Expression (GTEx) project reports a strong splice QTL for this variant in the relevant tissues (skin and esophagus). This example underscores both the complexity and the level of detail that can be obtained by combining MS proteomics approaches with genetic analyses.

a–c, Violin plots of a splice QTL from GTEx (a), SPINK5 at the protein level (b) and SPINK5 at the peptide level for representative peptides of the four protein groups (c). d, Forest plot of the peptide level meta-analysis. e, UniProt isoforms and protein groups. f, Gene structure indicating the alternative splice forms (black, green, red and blue) from Ensembl with the position of rs2052534 indicated. The C-allele of rs2052534 leads to a higher exon excision ratio and results in lower levels of the peptide QVQNEAEDAK, which is specific for the intron-retaining Q9NQ38-3 isoform (note the reverse order of the alleles in a versus b and c). This SPINK5 pQTL illustrates how peptide level information can be used to identify different isoform QTLs. However, it also reveals current limitations in the automated quantification of such isoforms, because this information did not translate to the protein level QTL, which in this case reflects the average of the isoform signals. A multiple alignment of all detected peptides is shown in Supplementary Table 11, violin plots for all peptide associations are provided as Supplementary Data 5 and regional association plots are in Supplementary Figure 16.
Validity of the MSPA score
To further support the validity of the MSPA score as a proxy for the detection or nondetection of a genetic association and its potential to identify true positive protein abundance pQTLs, we selected all pQTLs that were reported on the same genetic variant by deCODE and UKB-PPP, and for which matching protein and genetic data were also available in Tarkin and QMDiab. This set of 46 pQTLs served as a reference set for comparing effect sizes and directionality without having to rely on proxy SNPs (Supplementary Table 12). Several key observations emerged from this set of pQTLs. First, in contrast to the overall distribution of the MSPA scores (Fig. 3b,f)—where there were many high ranking pQTLs that had low or zero MSPA scores—the top ranking pQTLs in this reference set almost all had high MSPA scores (Fig. 3c,g). This finding suggests that these pQTLs were likely unaffected by epitope effects. This inference is further supported by the near-perfect correlation of the effect sizes between the Olink and the SOMAscan platforms (Fig. 3d). Given that these pQTLs were preselected based on their detection across both affinity platforms, it appears that a pQTL being detected by both affinity platforms is a strong indicator of a true protein abundance QTL and the absence of an epitope effect. This is a reasonable assumption because it is unlikely that two different affinity binders target the same surface area of a protein and produce a similar epitope readout.
We further investigated whether a candidate epitope-changing variant could be identified for the pQTLs colored in red in Fig. 3b,f. We queried the Ensembl database32 for coding variants that could potentially alter epitopes (Supplementary Table 13) and found that, for 22 out of 36 SOMAscan pQTLs and 29 out of 55 Olink pQTLs, such a variant had been reported (requiring LD r2 > 0.8). The lead pQTL SNP or a SNP in perfect LD (r2 = 1) was apparently epitope changing in all but 5 of the 51 cases. In addition, in 15 out of these 51 cases, a PAV-containing peptide was also detected on the Proteograph platform, with heterozygotes exhibiting approximately half the protein level, further confirming the quantification of the protein variant in blood (Supplementary Fig. 13). In eight cases (SERPING1, CPN2, SERPINA1, ENO3, HDGF, APOBR, IGFBP3 and APOL1), both alleles—the reference (REF) and alternate (ALT)—were detected and showed significant associations with the coding variant, whereas all non-PAV-containing peptides did not associate with the variant. Thus, the absence of a protein abundance pQTL in these cases is not attributable to limitations in peptide quantification.
To rule out that differences in genetic architecture contributed to a low MSPA score for some of the pQTLs, we computed a coloc-MSPA score based on genetic colocalization, defined as the weighted fraction of those peptides for which coloc favors a shared (H4) or a different (H3) signal between the UKB-PPP and Tarkin studies (Methods and Supplementary Table 10). Out of 224 pQTLs with MSPA < 0.2, only one pQTL (ITGA2) had a coloc-MSPA > 0.8 for H3, none for H4 and 205 had a coloc-MSPA < 0.2 for the combination of H3 or H4.
Discussion
In summary, our analysis suggests that >30% of the affinity proteomics pQTLs are reproducible by MS proteomics in a study of our size, whereas another 30% cannot be replicated and may thus be attributable to epitope effects. The remainder might be cases where a genetic variant interferes with the affinity binding, but at the same time might affect protein abundance via some other biological pathway. Among the 76 pQTLs with MS support (Supplementary Table 17) were 24 that had a disease-relevant association in the GWAS catalog (Table 2), whereas, among the 91 pQTLs without MS support (Supplementary Table 13), there were 17 (Table 3).
This study presents a comprehensive GWAS using the MS-based Seer Proteograph platform, accompanied by a full replication, and employs a proteomics data analysis protocol that accounts for genetic variants within the analyzed peptide23. Our methodology not only identified new pQTLs on proteins previously unassessed by affinity proteomics platforms but also re-examined previously reported affinity pQTLs for potential confounding due to epitope-altering variants. We estimate that 30% of the sufficiently powered pQTLs that we evaluated may be influenced by such effects, regardless of the affinity platform. However, given the limitations in statistical power that restricted evaluation to only the strongest affinity pQTLs—which are the ones most likely to be enriched for epitope effects due to ascertainment bias—this estimate should be considered as an upper bound and affinity binders targeting different isoforms may also explain some cases.
We reported a total of 364 pQTLs, of which 102 were successfully replicated in an independent population, at a replication rate that matches expectations based on post-hoc power calculations. We also identified instances where nonreplication may be explained by differences in genetic architecture and possibly also differences in environmental and lifestyle factors between the discovery and replication cohorts, which are drawn from very different populations. Nevertheless, this diversity enhances the robustness and translatability of the replicated pQTLs across populations.
The use of a proteome FASTA library that accounts for PAVs was central to our study. Without using this approach, a very high number of false-positive pQTLs would have been detected, as we previously discussed23. Traditional bottom-up proteomics data analysis pipelines often rely on a limited peptide library for protein quantification, where the presence of a single peptide with a large effect can skew the overall quantification. A PAV-containing peptide would not be detected in homozygotes of the alternative allele and heterozygotes would have half the peptide level. Inclusion of such PAV-containing peptides in protein quantification would thus lead to the equivalent of an epitope effect—that is, a pQTL signal that does not fully reflect genotype-dependent protein abundance. Moreover, PAV peptides corresponding to the alternative allele are absent from an in-silico digest library of the standard UniProt database. Indeed, using data from a standard proteomics data-processing run, we observed cases where fragment spectra of these peptides mismatched to peptides from other proteins and, in extreme cases, led to false protein identifications.
Our study also has some caveats. Mapping proteins across platforms using UniProt identifiers can be challenging, because affinity proteomics platforms sometimes report multiple protein identifiers when they are targeting protein complexes or use the UniProt identifier specific to the proteoform employed to generate the affinity binder, whereas alternative proteoforms may be included in the MS library, occasionally leading to annotations by protein groups rather than specific proteins. Furthermore, mapping UniProt IDs to gene names can also be challenging, especially when the underlying database versions do not match between studies. The level of matching between the MS and affinity protein identifiers has therefore been reported in the respective tables.
It should also be acknowledged that nanoparticle-enriched MS proteomics methods are not entirely free of potential ‘epitope-like’ effects. Although less likely due to their less specific electrostatic and hydrophobic interactions, protein–nanoparticle binding can, in principle, be modified by genetic variation. Genotype-associated missingness could potentially indicate such effects, though an association observed with more than one nanoparticle run makes them less likely. We found that 15.3% of the proteins involved in our pQTLs had a missingness pQTL, whereas 51.4% of our pQTL associations are supported by more than one nanoparticle run (Supplementary Note 1).
Although MS proteomics is not biased toward any particular set of preselected proteins, it is biased toward protein isoforms that are present in the utilized database, proteins that are enriched using one of the five nanoparticles and proteins that can be cleaved into peptides detected by the applied MS proteomics method, such as highly abundant proteins. There are also some differences in the protein panels covered by the different technologies. Relative to their respective panel size, the Seer platform covers the largest fraction of cytoplasmic proteins, whereas Olink leads in membrane proteins. SOMAscan has the lowest fraction of extracellular proteins but most proteins originate from the nucleus. As expected, low abundance proteins, such as cytokines, are less frequently detected by the Seer platform (Supplementary Table 6). As an additional level of quality control, we also analyzed associations with age and sex and found that these were generally concordant between the affinity and MS-based proteomics platforms (Supplementary Note 2).
Taken together, we demonstrate the complementarity of MS proteomics with affinity approaches by validating associations that may be driven by epitope effects and by substantially extending the panel of proteins accessible to pQTL studies. We report new pQTLs of biomedical relevance and provide important insights at the peptide level regarding the genetic architecture of previously reported pQTLs. We also show that MS-based and affinity-based methods are complementary when it comes to interpreting pQTLs in the presence of multiple isoforms, because affinity methods discriminate proteins at the level of the folded protein, whereas MS methods work on the peptide level at a higher ‘resolution’ by mapping multiple peptides to different parts of the protein.
Methods
Ethics
The original Tarkin study was reviewed and approved by the Institute of Regenerative and Cellular Medicine (IRCM) Institutional Review Board (IRB) and the Massachusetts General Brigham (MGB) IRB. Participants provided written informed consent to take part in the study. The Weill Cornell Medicine—Qatar (WCMQ) IRB determined that use of the Tarkin data for the present project did not meet the definition of human research for this study (IRB document no. HRP-532). The QMDiab study was approved by the institutional research boards of WCMQ under protocol no. 2011-0012 and Hamad Medical Corporation under protocol no. 11131/11, and complies with all relevant ethical regulations. For forthgoing work with the study nonhuman participant research, determination was obtained. The study design and conduct complied with all relevant regulations regarding the use of human study participants and was conducted in accordance with the criteria set by the Declaration of Helsinki.
Cohorts
The samples used in the Tarkin study were obtained from the MGB Biobank. Joint phenotype and genotype data were available for 1,260 samples, comprising 662 women and 598 men with an age range of 23–99 years (median 70 years, mean 67.2 years). Of the participants 1,057 self-reported as white. This subset of MGB samples together with its deep omics characterization is referred to as the ‘Tarkin study’ in this paper33. For replication, a total of 345 previously unthawed, citrate blood plasma samples from participants of the Qatar Metabolomics study of Diabetes (QMDiab), including women and men of predominantly Arab, Indian and Filipino ancestries, with and without diabetes in an age range of 18–80 years were assayed using the Proteograph platform (Seer)23,34,35.
Genotyping
Imputed genotype data for 1,980 samples of the Tarkin study was received on a per-chromosome basis in vcf format (build 37, imputed using Minimac3, no insertions and/or deletions (indels)). The genotype data were filtered for biallelic variants and variant names were standardized using bcftools (v.1.16), converted to the plink format and filtered using PLINK2 (v.2.00a5LM) with the options --geno 0.1--mac 10 --maf 0.05 --hwe 1E-15. For 1,260 samples, proteomics data were available. These samples were merged into a single genotype file and further filtered using PLINK2 with the options --maf 0.05 --hwe 1E-6 --geno 0.02, leaving data for 5,461,287 genetic variants. The first ten genetic principal components were then computed using PLINK2 with the --pca option. QMDiab samples were genotyped using the Illumina Omni 2.5 array (v.8) and imputed using the SHAPEIT software with 1000 Genomes (phase 3) haplotypes. Genotyping data were available for 325 of the 345 samples with proteomics data.
Proteomics
The workflow used for the proteomics analyses of the Tarkin and the QMDiab samples were essentially identical and have been previously described in detail23. Briefly, plasma samples were prepared using the Proteograph workflow24,25 (Seer) to generate purified peptides that were then analyzed using a dia-PASEF method36 on a timsTOF Pro 2 mass spectrometer (Bruker Daltonics). Each study was conducted at independent times using two mass spectrometers. One mass spectrometer coincidentally was reused in both studies. DIA-NN (1.8.1)37 was used to derive peptide and protein intensities. A library-free search based on UniProt UP000005640_9606 was used, processing the data a second time using the match between runs (MBR) option. Two additional libraries were created, one excluding common (minor allele frequency (MAF) >10%) PAV-containing peptides and one injecting the alternative alleles into the reference protein sequences. These libraries were referred to as the PAV-exclusive library and the PAV-inclusive library, respectively. Details of this library generation process have been described elsewhere23. DIA-NN’s normalized intensities (PG.Normalised) were used as protein readout. Some 5,753 unique protein groups were quantified, 4,109 were detected in at least 20% of the samples and 1,980 had <20% of missing values. Wherever a protein was detected at this level in more than one nanoparticle run, the one with the largest sum of protein intensities was retained. Protein levels were then log(scaled), residualized using age, sex and the ten first genotype principal components and finally inverse-normal scaled.
Genome-wide association
Genetic associations of 1,980 residualized and inverse-normal scaled protein levels with 5,461,287 genetic variants in 1,260 samples were evaluated using linear models (PLINK v.1.90b7.1, option --linear). Missing data points were excluded. For six proteins, the residualization did not entirely remove association with these confounders. Three of these proteins presented with inflated GWAS statistics and the corresponding pQTLs were removed from the analysis (UniProt V9GYE7, B1AKG0 and O15230). For every protein, the strongest association reaching genome-wide significance (P < 5 × 10−8) within a ±10-Mb window was retained. Lead pQTLs were then clumped into loci using an LD cutoff of r2 = 0.9 and r2 = 0.6, which led to the identification of 308 and 295 loci, respectively (Supplementary Table 2). Conditionally independent variants were identified by iteratively conditioning on the previously identified genetic variants until no further variant reaching an ad-hoc significance level of P < 10−6 was found.
Replication
Replication was attempted using data from 325 samples of the QMDiab study that had joint genotype and proteomics information. Power to replicate 80% of the pQTLs was determined as the 80% quantile of the P values obtained from 1,000 random samples from the Tarkin dataset using the number of samples available in the QMDiab for the respective genotype–protein pair. Colocalization analysis using the coloc software38 was conducted using summary statistics for all variants shared between Tarkin and QMDiab within a ±1-Mb window around the respective lead variant. A pQTL was considered replicated if (1) coloc suggested H4 (presence of a shared genetic signal) as the most likely hypothesis and (2) the joint P value computed for the strongest association on a shared variant between Tarkin and QMDiab reached a genome-wide and proteome-wide significance level of P < 5 × 10−8/1,980. Concordance of directionality was verified. We also implemented an alternative replication strategy as follows: for every pQTL with matching protein data in QMDiab (n = 328), we identified all proxies of the lead SNP (r2 ≥ 0.1) using LD information from Tarkin. We then identified all proxies of these variants in QMDiab (r2 ≥ 0.1) using LD information from QMDiab. We considered a pQTL replicated when the strongest association of these proxies had a P < 0.05/328 (Supplementary Table 2).
Overlap with previous Olink and SOMAscan pQTLs
Summary statistics from the UKB-PPP Olink study18 and deCODE SOMAscan study17 were downloaded from the respective sites. Associations were reported for 4,660 proteins by deCODE and for 2,908 proteins by UKB-PPP. All protein associations on variants that matched one of the Proteograph pQTLs were retrieved. Matching at the protein level was conducted in three steps: (1) identical UniProt ID between the MS and affinity study; (2) match to one of the UniProt IDs in a protein group, regardless of the isoform version (ignoring the dash-number in the UniProt ID); and (3) match at the level of the protein-coding gene. An association reported by deCODE and Olink was considered as significant at levels of P < 5 × 10−8 and as marginally significant P < 0.05 and P > 5 × 10−8. The respective information regarding the level of the match is recorded in the respective Supplementary Tables.
Evaluation of age and sex associations
Summary statistics for age and sex associations were retrieved from the supplementary Excel files of ref. 17 for deCODE SOMAscan (sheet ST01) and of ref. 18 for UKB-PPP Olink (sheet ST5). Associations with age and sex for the Tarkin study were computed with PLINK using identical datasets and models as for the genome-wide association (option --linear no-snp). Proteins were matched using UniProt identifiers. In rare cases when there were matches to multiple protein groups, the strongest association was retained.
Evaluation of Olink and SOMAscan cis-pQTLs
The cis-pQTLs were obtained from the supplementary tables of the respective studies (ST02 from ref. 17 for deCODE SOMAscan and ST10 from ref. 18 for UKB-PPP Olink). The pQTLs were limited to variants that were located on autologous chromosomes, had a suitable replication SNP available in both Tarkin and QMDiab (LD r2 > 0.8) and an MAF > 5% in the Tarkin study. Matching of proteins between platforms was done using UniProt IDs, allowing for matches to protein groups that contained the UniProt ID and matches at the gene level, as described above for the overlap with previous Olink and SOMAscan pQTLs. Ambiguous cases where more than one matching protein group was found were omitted. In cases where protein readouts for multiple nanoparticle runs were available, the one with the highest single number of peptide detections was used. Cases where the number of quantified proteins in Tarkin was <80% were excluded. The SOMAscan platform sometimes uses multiple aptamers. In these cases, the strongest association was retained.
Annotation of pQTLs
The Open Targets27 platform (v.22.10) was used via the API to annotate pQTL variants with most likely causal genes, variant effect, overlapping disease GWAS hits and gene expression, splice variant and proteomics QTLs, ordered by increasing P value and reported separately for cis-QTLs and trans-QTLs. The Open Targets look-up comprised both, same variant across stored datasets and reference to GWAS regional lead signals using LD, limited to LD r2 > 0.7 between the pQTL and GWAS lead signal. Hyperlinks to individual Open Targets pages with more detailed information, including references for the disease associations, are provided in Supplementary Table 2. A comprehensive list of all queried studies can be found in the Open Targets release notes (https://genetics-docs.opentargets.org/release-notes). Protein epitope changing variants were identified using Ensembl32.
Definition of the MSPA score
We define the MSPA score of a pQTL as follows:
where \({k}_{\text{pep}}\) is the number of different peptides that have been detected for a given pQTL protein, \({n}_{i}\) is the number of samples in which a peptide i from the given protein has been detected, \({n}_{\text{tot}}={\sum }_{i=1}^{{k}_{\text{pep}}}\,{n}_{i}\) is the total number of individual peptide detections, \({c}_{i}^{\text{upper}}\) and \({c}_{i}^{\text{lower}}\) are the upper and lower 99% bound of the CI for the effect size of the genetic association of the pQTL variant with peptide i, and \(\delta \left(\text{condition}\right)\) is a function that takes a value of 1 if the condition in its argument is true and 0 otherwise. We further computed MSPA scores based on genetic colocalization (coloc-MSPA) between Tarkin and UKB-PPP, where the score is defined as the weighted fraction of a given hypothesis being considered most likely by coloc38. The coloc-MSPA score for a shared genetic signal (hypothesis H4) is thus computed using the above formula with \(\delta \left(\text{H}4\text{most likely}\right)\) and similar for a different genetic signal (hypothesis H3).
Power analysis
We determined power to replicate pQTLs from the deCODE and UKB-PPP studies using an F test. For the computation of the noncentrality parameter (NCP), we used the effect size (βaff) from the UKB-PPP or deCODE pQTLs, whereas the s.e. (s.e.MS) was taken from Tarkin protein associations at the respective loci to account for the variability of the MS measurements, that is, NCP = (βaff/s.e.MS)2. Power was then computed as 1 − P(F < Fcrit, d.f.1, d.f.2, NCP) where P represents the cumulative probability of the F distribution with the NCP. The degree of freedom d.f.1 was set to the number of samples in Tarkin with valid data for the respective pQTL − 2 and d.f.2 was set to 2 (slope and offset). Fcrit was determined as the F value corresponding to a significance level of P < 0.05/319 and P < 0.05/392 to account for the number of tested pQTLs for deCODE and UKB-PPP, respectively (Fig. 3 and Supplementary Tables 9 and 10).
Peptide mapping
Protein quantification was performed using algorithms implemented in DIA-NN37. Although DIA-NN applies calibrated methods and in particular accounts for false discovery rates (FDRs) in the mapping of peptides to proteins, the number of peptides mapped to a given protein and their coverage of the protein can vary and are a measure for the robustness of the protein identification and its quantification. We therefore aligned all peptides to their respective protein amino acid sequences (R Biostrings, v.2.60.2) and plotted their coverage against their sample-wise detection rates (Supplementary Fig. 14 and Supplementary Data 6). The average (median) number of peptides detected per sample was 9.6 (5.6) for Tarkin and 6.4 (2.9) for QMDiab (Supplementary Fig. 15g). To examine the overall between-study variation of the protein and peptide quantifications, we compared the mean intensities and their s.d. values between Tarkin and QMDiab. The average intensities of the 1,980 analyzed proteins correlated with r2 = 0.78 (slope = 0.84) and their s.d. values correlated with r2 = 0.69 (slope = 0.71). The average intensities of the individual peptides that map to these proteins correlate with r2 = 0.61 (slope = 0.76) (Supplementary Fig. 15a–f and Supplementary Table 14).
Protein- and peptide-level associations for all nanoparticle runs
To reduce the multiple-testing burden, we analyzed only a single nanoparticle run for every protein in the main GWAS, selecting the one with the highest intensity at <20% missingness. However, in many cases, a same protein or peptide can be detected during multiple nanoparticle runs. Their associations can provide additional insights and support for the validity of a given pQTL. We therefore computed the summary statistics of all pQTL variants with all proteins and peptides in all nanoparticle runs using identical methods as for the main GWAS (Supplementary Table 15).
Missingness analysis
We conducted a GWAS on missingness using PLINK (v.1.90b7.1, option --model fisher) including all proteins in all nanoparticle runs that had a missingness <95%. For each variant association, we retained the lower of the two P values obtained from Fisher’s exact test with a dominant and a recessive genetic model. To identify the lead variants across multiple nanoparticle runs, we clumped the associations into loci, that is, for every protein, the strongest association reaching an ad-hoc significance level of P < 5 × 10−12 within a ±10-Mb window was retained. Lead pQTLs were then clumped into loci using an LD cutoff of r2 = 0.9.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The MS-proteomics data for the Tarkin study have been deposited with ProteomeXchange under project accession no. PXD048709. The MS-proteomics data of QMDiab are available on ProteomeXchange with accession no. PXD042852. Summary statistics of this GWAS were deposited in the GWAS catalog with identifiers GCST90570713 to GCST90572692. Consent obtained from the study participants does not allow deposition of genetic information in public databases. Researchers affiliated with a research institution may request access to genetic data on an individual basis from the corresponding authors (K.S., WCMQ, Doha, Qatar for QMDiab and J.A.L.-S. for Tarkin). Access is subject to approval by the respective institutional research boards of WCMQ and the MGB Biobank.
References
Plenge, R. M., Scolnick, E. M. & Altshuler, D. Validating therapeutic targets through human genetics. Nat. Rev. Drug Discov. 12, 581–594 (2013).
Mountjoy, E. et al. An open approach to systematically prioritize causal variants and genes at all published human GWAS trait-associated loci. Nat. Genet. 53, 1527–1533 (2021).
Minikel, E. V., Painter, J. L., Dong, C. C. & Nelson, M. R. Refining the impact of genetic evidence on clinical success. Nature 629, 624–629 (2024).
Carss, K. J. et al. Using human genetics to improve safety assessment of therapeutics. Nat. Rev. Drug Discov. 22, 145–162 (2023).
Suhre, K., McCarthy, M. I. & Schwenk, J. M. Genetics meets proteomics: perspectives for large population-based studies. Nat. Rev. Genet. 22, 19–37 (2021).
Suhre, K. et al. Connecting genetic risk to disease end points through the human blood plasma proteome. Nat. Commun. 8, 14357 (2017).
Sun, B. B. et al. Genomic atlas of the human plasma proteome. Nature 558, 73–79 (2018).
Lourdusamy, A. et al. Identification of cis-regulatory variation influencing protein abundance levels in human plasma. Hum. Mol. Genet. 21, 3719–3726 (2012).
Enroth, S., Johansson, A., Enroth, S. B. & Gyllensten, U. Strong effects of genetic and lifestyle factors on biomarker variation and use of personalized cutoffs. Nat. Commun. 5, 4684 (2014).
Emilsson, V. et al. Co-regulatory networks of human serum proteins link genetics to disease. Science 361, 769–773 (2018).
Pietzner, M. et al. Mapping the proteo-genomic convergence of human diseases. Science 374, eabj1541 (2021).
Gudjonsson, A. et al. A genome-wide association study of serum proteins reveals shared loci with common diseases. Nat. Commun. 13, 480 (2022).
Zhao, J. H. et al. Genetics of circulating inflammatory proteins identifies drivers of immune-mediated disease risk and therapeutic targets. Nat. Immunol. 24, 1540–1551 (2023).
Carland, C. et al. Proteomic analysis of 92 circulating proteins and their effects in cardiometabolic diseases. Clin. Proteom. 20, 31 (2023).
Folkersen, L. et al. Genomic and drug target evaluation of 90 cardiovascular proteins in 30,931 individuals. Nat. Metab. 2, 1135–1148 (2020).
Sun, B. B., Suhre, K. & Gibson, B. W. Promises and challenges of populational proteomics in health and disease. Mol. Cell Proteom. 23, 100786 (2024).
Ferkingstad, E. et al. Large-scale integration of the plasma proteome with genetics and disease. Nat. Genet. 53, 1712–1721 (2021).
Sun, B. B. et al. Plasma proteomic associations with genetics and health in the UK Biobank. Nature 622, 329–338 (2023).
Johansson, Å et al. Identification of genetic variants influencing the human plasma proteome. Proc. Natl Acad. Sci. USA 110, 4673–4678 (2013).
Xu, F. et al. Genome-wide genotype-serum proteome mapping provides insights into the cross-ancestry differences in cardiometabolic disease susceptibility. Nat. Commun. 14, 896 (2023).
Niu, L. et al. Plasma proteome variation and its genetic determinants in children and adolescents. Nat. Genet. 57, 635–646 (2025).
Newton-Cheh, C. et al. Association of common variants in NPPA and NPPB with circulating natriuretic peptides and blood pressure. Nat. Genet. 41, 348–353 (2009).
Suhre, K. et al. Nanoparticle enrichment mass-spectrometry proteomics identifies protein-altering variants for precise pQTL mapping. Nat. Commun. 15, 989 (2024).
Liu, Y. et al. Nano-bio interactions in cancer: from therapeutics delivery to early detection. Acc. Chem. Res. 54, 291–301 (2021).
Ferdosi, S. et al. Enhanced competition at the nano-bio interface enables comprehensive characterization of protein corona dynamics and deep coverage of proteomes. Adv. Mater. 34, e2206008 (2022).
Nonnecke, E. B. et al. Human intelectin-1 (ITLN1) genetic variation and intestinal expression. Sci. Rep. 11, 12889 (2021).
Ochoa, D. et al. Open Targets Platform: supporting systematic drug-target identification and prioritisation. Nucleic Acids Res. 49, D1302–D1310 (2021).
Wang, H. et al. Genome-wide interaction analysis of pathological hallmarks in Alzheimer’s disease. Neurobiol. Aging 93, 61–68 (2020).
Pietzner, M. et al. Synergistic insights into human health from aptamer- and antibody-based proteomic profiling. Nat. Commun. 12, 6822 (2021).
Sun, Q. et al. The genomic and phenotypic landscape of ichthyosis: an analysis of 1000 kindreds. JAMA Dermatol. 158, 16–25 (2022).
Tartaglia-Polcini, A. et al. SPINK5, the defective gene in Netherton syndrome, encodes multiple LEKTI isoforms derived from alternative pre-mRNA processing. J. Invest. Dermatol. 126, 315–324 (2006).
Martin, F. J. et al. Ensembl 2023. Nucleic Acids Res. 51, D933–D941 (2022).
Chen, Q. et al. OMICmAge: an integrative multi-omics approach to quantify biological age with electronic medical records. Preprint at bioRxiv https://doi.org/10.1101/2023.10.16.562114 (2023).
Mook-Kanamori, D. O. et al. 1,5-Anhydroglucitol in saliva is a noninvasive marker of short-term glycemic control. J. Clin. Endocrinol. Metab. 99, E479–E483 (2014).
Yousri, N. A. et al. A systems view of type 2 diabetes-associated metabolic perturbations in saliva, blood and urine at different timescales of glycaemic control. Diabetologia 58, 1855–1867 (2015).
Meier, F. et al. diaPASEF: parallel accumulation—serial fragmentation combined with data-independent acquisition. Nat. Methods 17, 1229–1236 (2020).
Demichev, V. et al. dia-PASEF data analysis using FragPipe and DIA-NN for deep proteomics of low sample amounts. Nat. Commun. 13, 3944 (2022).
Giambartolomei, C. et al. Bayesian test for colocalisation between pairs of genetic association studies using summary statistics. PLoS Genet. 10, e1004383 (2014).
Acknowledgements
We are grateful to all study participants for providing their time and blood. K.S. and F.S. are supported by the Biomedical Research Program at WCMQ, a program funded by the Qatar Foundation. K.S. is also supported by Qatar National Research Fund (grant nos. NPRP11C-0115-180010 and ARG01-0420-230007). J.A.L.-S. is supported by the National Institutes of Health (grant nos. R01HL123915, R01HL155742 and U19AI168643). The statements made herein are solely the responsibility of the authors. We thank G. Venkataraman, A. Alavi and J. Wang for developing software workflows that were used in the proteogenomic analyses in this work.
Author information
Authors and Affiliations
Contributions
Financial support: K.S. and J.A.L.-S. Study design: K.S. and J.A.L.-S. Data analysis: K.S., H.G. and S.B. Provided materials and conducted experiments: Q.C., A.H., K.M., A.D., N.S., G.T., H.S., V.B.D., R.S. and F.S. Manuscript writing: K.S., S.B. and J.A.L.-S. All authors contributed to the interpretation of the results and critically reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
H.G. and S.B. are employees and/or stockholders of Seer R.S. and J.A.L.-S. are scientific advisors to Precion and TruDiagnostic. J.A.L.-S. has a sponsored research agreement with TruDiagnostic and has previously consulted for Cambrian and Ahara. The remaining authors declare no competing interests.
Peer review
Peer review information
Nature Genetics thanks Anders Mälarstig and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–16 and Notes 1–2.
Supplementary Data 1 (download PDF )
Violin plots of protein and up to 4 peptide levels by genotype for 364 pQTLs identified in this study.
Supplementary Data 2 (download PDF )
Regional association plots for 364 pQTLs identified in this study.
Supplementary Data 3 (download PDF )
Meta-analysis Forest plots and violin plots of protein and up to 19 peptide levels by genotype for 364 pQTLs identified in this study.
Supplementary Data 4 (download PDF )
Meta-analysis Forest plots and violin plots of protein and up to 19 peptide levels by genotype for 319 cis-pQTLs identified by the deCODE SOMAscan study.
Supplementary Data 5 (download PDF )
Meta-analysis Forest plots and violin plots of protein and up to 19 peptide levels by genotype for 392 cis-pQTLs identified by the UKB-PPP Olink study.
Supplementary Data 6 (download PDF )
Plots visualizing the peptide mapping to all 1,980 analyzed proteins.
Supplementary Tables (download XLSX )
Supplementary Tables 1–17.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Suhre, K., Chen, Q., Halama, A. et al. A genome-wide association study of mass spectrometry proteomics using a nanoparticle enrichment platform. Nat Genet 57, 2987–2996 (2025). https://doi.org/10.1038/s41588-025-02413-w
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41588-025-02413-w
This article is cited by
-
Cross-ancestry comparison of aptamer and antibody protein measures
Nature Communications (2026)
