
Abstract
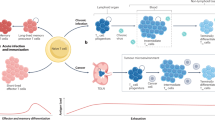
The durability of an antitumor immune response is mediated in part by the persistence of progenitor exhausted CD8+ T cells (Tpex). Tpex serve as a resource for replenishing effector T cells and preserve their quantity through self-renewal. However, it is unknown how T cell receptor (TCR) engagement affects the self-renewal capacity of Tpex in settings of continued antigen exposure. Here we use a Lewis lung carcinoma model that elicits either optimal or attenuated TCR signaling in CD8+ T cells to show that formation of Tpex in tumor-draining lymph nodes and their intratumoral persistence is dependent on optimal TCR engagement. Notably, attenuated TCR stimulation accelerates the terminal differentiation of optimally primed Tpex. This TCR-reinforced Tpex development and self-renewal is coupled to proximal positioning to dendritic cells and epigenetic imprinting involving increased chromatin accessibility at Egr2 and Tcf1 target loci. Collectively, this study highlights the critical function of TCR engagement in sustaining Tpex during tumor progression.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others


Data availability
Raw scRNA-seq, ATAC-seq, WGBS and CUT&Tag data have been deposited in the NCBI Gene Expression Omnibus database under the SuperSeries accession code GSE262845. All other data are available in the article, source data and Supplementary Information, or from the corresponding author upon request. Source data are provided with this paper.
Code availability
No new algorithms were developed for this paper. All analysis code is available upon request.
References
Gubin, M. M. et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 515, 577–581 (2014).
Philip, M. & Schietinger, A. CD8+ T cell differentiation and dysfunction in cancer. Nat. Rev. Immunol. 22, 209–223 (2022).
Rosenberg, S. A. et al. Durable complete responses in heavily pretreated patients with metastatic melanoma using T-cell transfer immunotherapy. Clin. Cancer Res. 17, 4550–4557 (2011).
Rosenberg, S. A. & Restifo, N. P. Adoptive cell transfer as personalized immunotherapy for human cancer. Science 348, 62–68 (2015).
Moskophidis, D., Lechner, F., Pircher, H. & Zinkernagel, R. M. Virus persistence in acutely infected immunocompetent mice by exhaustion of antiviral cytotoxic effector T cells. Nature 362, 758–761 (1993).
Zajac, A. J. et al. Viral immune evasion due to persistence of activated T cells without effector function. J. Exp. Med. 188, 2205–2213 (1998).
Blank, C. U. et al. Defining ‘T cell exhaustion’. Nat. Rev. Immunol. 19, 665–674 (2019).
Hirano, F. et al. Blockade of B7-H1 and PD-1 by monoclonal antibodies potentiates cancer therapeutic immunity. Cancer Res. 65, 1089–1096 (2005).
Barber, D. L. et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 439, 682–687 (2006).
Day, C. L. et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 443, 350–354 (2006).
Korman, A. J., Garrett-Thomson, S. C. & Lonberg, N. The foundations of immune checkpoint blockade and the ipilimumab approval decennial. Nat. Rev. Drug Discov. 21, 509–528 (2022).
Utzschneider, D. T. et al. T cells maintain an exhausted phenotype after antigen withdrawal and population reexpansion. Nat. Immunol. 14, 603–610 (2013).
Im, S. J. et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature 537, 417–421 (2016).
He, R. et al. Follicular CXCR5-expressing CD8+ T cells curtail chronic viral infection. Nature 537, 412–428 (2016).
Utzschneider, D. T. et al. T cell factor 1-expressing memory-like CD8+ T cells sustain the immune response to chronic viral infections. Immunity 45, 415–427 (2016).
Miller, B. C. et al. Subsets of exhausted CD8+ T cells differentially mediate tumor control and respond to checkpoint blockade. Nat. Immunol. 20, 326–336 (2019).
Siddiqui, I. et al. Intratumoral Tcf1+PD-1+CD8+ T cells with stem-like properties promote tumor control in response to vaccination and checkpoint blockade immunotherapy. Immunity 50, 195–211.e10 (2019).
Zehn, D., Thimme, R., Lugli, E., de Almeida, G. P. & Oxenius, A. ‘Stem-like’ precursors are the fount to sustain persistent CD8+ T cell responses. Nat. Immunol. 23, 836–847 (2022).
Blackburn, S. D., Shin, H., Freeman, G. J. & Wherry, E. J. Selective expansion of a subset of exhausted CD8 T cells by αPD-L1 blockade. Proc. Natl Acad. Sci. USA 105, 15016–15021 (2008).
Huang, A. C. et al. T-cell invigoration to tumour burden ratio associated with anti-PD-1 response. Nature 545, 60–65 (2017).
Sade-Feldman, M. et al. Defining T cell states associated with response to checkpoint immunotherapy in melanoma. Cell 175, 998–1013.e20 (2018).
Thommen, D. S. et al. A transcriptionally and functionally distinct PD-1+ CD8+ T cell pool with predictive potential in non-small-cell lung cancer treated with PD-1 blockade. Nat. Med. 24, 994–1004 (2018).
Krishna, S. et al. Stem-like CD8 T cells mediate response of adoptive cell immunotherapy against human cancer. Science 370, 1328–1334 (2020).
Wu, T. et al. The TCF1–Bcl6 axis counteracts type I interferon to repress exhaustion and maintain T cell stemness. Sci. Immunol. 1, eaai8593 (2016).
Wieland, D. et al. TCF1+ hepatitis C virus-specific CD8+ T cells are maintained after cessation of chronic antigen stimulation. Nat. Commun. 8, 15050 (2017).
Burger, M. L. et al. Antigen dominance hierarchies shape TCF1+ progenitor CD8 T cell phenotypes in tumors. Cell 184, 4996–5014.e26 (2021).
Shakiba, M. et al. TCR signal strength defines distinct mechanisms of T cell dysfunction and cancer evasion. J. Exp. Med. 219, e20201966 (2022).
Cheever, M. A. et al. The prioritization of cancer antigens: a national cancer institute pilot project for the acceleration of translational research. Clin. Cancer Res. 15, 5323–5337 (2009).
Aleksic, M. et al. Different affinity windows for virus and cancer-specific T-cell receptors: implications for therapeutic strategies. Eur. J. Immunol. 42, 3174–3179 (2012).
van der Most, R. G. et al. Identification of Db- and Kb-restricted subdominant cytotoxic T-cell responses in lymphocytic choriomeningitis virus-infected mice. Virology 240, 158–167 (1998).
Moran, A. E. et al. T cell receptor signal strength in Treg and iNKT cell development demonstrated by a novel fluorescent reporter mouse. J. Exp. Med. 208, 1279–1289 (2011).
Martinez-Usatorre, A., Donda, A., Zehn, D. & Romero, P. PD-1 blockade unleashes effector potential of both high- and low-affinity tumor-infiltrating T cells. J. Immunol. 201, 792–803 (2018).
Tsui, C. et al. MYB orchestrates T cell exhaustion and response to checkpoint inhibition. Nature 609, 354–360 (2022).
Daniel, B. et al. Divergent clonal differentiation trajectories of T cell exhaustion. Nat. Immunol. 23, 1614–1627 (2022).
Giles, J. R. et al. Shared and distinct biological circuits in effector, memory and exhausted CD8+ T cells revealed by temporal single-cell transcriptomics and epigenetics. Nat. Immunol. 23, 1600–1613 (2022).
Hudson, W. H. et al. Proliferating transitory T cells with an effector-like transcriptional signature emerge from PD-1+ stem- like CD8+ T cells during chronic infection. Immunity 51, 1043–1058.e4 (2019).
Zander, R. et al. CD4+ T cell help is required for the formation of a cytolytic CD8+ T cell subset that protects against chronic infection and cancer. Immunity 51, 1028–1042.e4 (2019).
Dammeijer, F. et al. The PD-1/PD-L1-checkpoint restrains T cell immunity in tumor-draining lymph nodes. Cancer Cell 38, 685–700.e8 (2020).
Connolly, K. A. et al. A reservoir of stem-like CD8+ T cells in the tumor-draining lymph node preserves the ongoing antitumor immune response. Sci. Immunol. 6, eabg7836 (2021).
Huang, Q. et al. The primordial differentiation of tumor-specific memory CD8+ T cells as bona fide responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell 185, 4049–4066.e25 (2022).
Jansen, C. S. et al. An intra-tumoral niche maintains and differentiates stem-like CD8 T cells. Nature 576, 465–470 (2019).
Schenkel, J. M. et al. Conventional type I dendritic cells maintain a reservoir of proliferative tumor-antigen specific TCF-1+ CD8+ T cells in tumor-draining lymph nodes. Immunity 54, 2338–2353.e6 (2021).
Dahling, S. et al. Type 1 conventional dendritic cells maintain and guide the differentiation of precursors of exhausted T cells in distinct cellular niches. Immunity 55, 656–670.e8 (2022).
Jadhav, R. R. et al. Epigenetic signature of PD-1+ TCF1+ CD8 T cells that act as resource cells during chronic viral infection and respond to PD-1 blockade. Proc. Natl Acad. Sci. USA 116, 14113–14118 (2019).
Beltra, J. C. et al. Developmental relationships of four exhausted CD8+ T cell subsets reveals underlying transcriptional and epigenetic landscape control mechanisms. Immunity 52, 825–841.e8 (2020).
Chen, Z. et al. In vivo CD8+ T cell CRISPR screening reveals control by Fli1 in infection and cancer. Cell 184, 1262–1280.e22 (2021).
Hwang, S. S. et al. mRNA destabilization by BTG1 and BTG2 maintains T cell quiescence. Science 367, 1255–1260 (2020).
Wagle, M. V. et al. Antigen-driven EGR2 expression is required for exhausted CD8+ T cell stability and maintenance. Nat. Commun. 12, 2782 (2021).
Shankar, D. B. et al. The role of CREB as a proto-oncogene in hematopoiesis and in acute myeloid leukemia. Cancer Cell 7, 351–362 (2005).
Grenningloh, R. et al. Ets-1 maintains IL-7 receptor expression in peripheral T cells. J. Immunol. 186, 969–976 (2011).
Kim, C. J. et al. The transcription factor Ets1 suppresses T follicular helper type 2 cell differentiation to halt the onset of systemic lupus erythematosus. Immunity 49, 1034–1048.e8 (2018).
Ghoneim, H. E. et al. De novo epigenetic programs inhibit PD-1 blockade-mediated T cell rejuvenation. Cell 170, 142–157.e19 (2017).
Prinzing, B. et al. Deleting DNMT3A in CAR T cells prevents exhaustion and enhances antitumor activity. Sci. Transl. Med. 13, eabh0272 (2021).
Shin, H. et al. A role for the transcriptional repressor Blimp-1 in CD8+ T cell exhaustion during chronic viral infection. Immunity 31, 309–320 (2009).
Nah, J. & Seong, R. H. Kruppel-like factor 4 regulates the cytolytic effector function of exhausted CD8 T cells. Sci. Adv. 8, eadc9346 (2022).
Yoshikawa, T. et al. Genetic ablation of PRDM1 in antitumor T cells enhances therapeutic efficacy of adoptive immunotherapy. Blood 139, 2156–2172 (2022).
Martens, A. W. J. et al. Chronic lymphocytic leukemia presence impairs antigen-specific CD8+ T-cell responses through epigenetic reprogramming towards short-lived effectors. Leukemia 37, 606–616 (2023).
Olesin, E., Nayar, R., Saikumar-Lakshmi, P. & Berg, L. J. The transcription factor Runx2 is required for long-term persistence of antiviral CD8+ memory T cells. ImmunoHorizons 2, 251–261 (2018).
Seo, H. et al. BATF and IRF4 cooperate to counter exhaustion in tumor-infiltrating CAR T cells. Nat. Immunol. 22, 983–995 (2021).
Pritykin, Y. et al. A unified atlas of CD8 T cell dysfunctional states in cancer and infection. Mol. Cell 81, 2477–2493.e10 (2021).
Zhong, Y. et al. Hierarchical regulation of the resting and activated T cell epigenome by major transcription factor families. Nat. Immunol. 23, 122–134 (2022).
Kaya-Okur, H. S. et al. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 10, 1930 (2019).
Hajaj, E. et al. SLAMF6 deficiency augments tumor killing and skews toward an effector phenotype revealing it as a novel T cell checkpoint. eLife 9, e52539 (2020).
Kasmani, M. Y. et al. Clonal lineage tracing reveals mechanisms skewing CD8+ T cell fate decisions in chronic infection. J. Exp. Med. 220, e20220679 (2023).
Utzschneider, D. T. et al. Early precursor T cells establish and propagate T cell exhaustion in chronic infection. Nat. Immunol. 21, 1256–1266 (2020).
Westcott, P. M. K. et al. Low neoantigen expression and poor T-cell priming underlie early immune escape in colorectal cancer. Nat. Cancer 2, 1071–1085 (2021).
Alfei, F. et al. TOX reinforces the phenotype and longevity of exhausted T cells in chronic viral infection. Nature 571, 265–269 (2019).
Khan, O. et al. TOX transcriptionally and epigenetically programs CD8+ T cell exhaustion. Nature 571, 211–218 (2019).
Scott, A. C. et al. TOX is a critical regulator of tumour-specific T cell differentiation. Nature 571, 270–274 (2019).
McGill, J., Van Rooijen, N. & Legge, K. L. Protective influenza-specific CD8 T cell responses require interactions with dendritic cells in the lungs. J. Exp. Med. 205, 1635–1646 (2008).
Chen, Z. et al. TCF-1-centered transcriptional network drives an effector versus exhausted CD8 T cell-fate decision. Immunity 51, 840–855.e5 (2019).
Moustaki, A. et al. Antigen cross-presentation in young tumor-bearing hosts promotes CD8+ T cell terminal differentiation. Sci. Immunol. 7, eabf6136 (2022).
Hanawa, H. et al. Efficient gene transfer into rhesus repopulating hematopoietic stem cells using a simian immunodeficiency virus-based lentiviral vector system. Blood 103, 4062–4069 (2004).
Grandi, F. C., Modi, H., Kampman, L. & Corces, M. R. Chromatin accessibility profiling by ATAC-seq. Nat. Protoc. 17, 1518–1552 (2022).
Utzschneider, D. T. et al. High antigen levels induce an exhausted phenotype in a chronic infection without impairing T cell expansion and survival. J. Exp. Med. 213, 1819–1834 (2016).
Acknowledgements
We thank G. Lennon, R. Cross, K. Hays, S. Fatima and the Flow Cytometry and Cell Sorting Shared Resource at St. Jude Children’s Research Hospital for cell sorting assistance and flow instrument maintenance; S. Konduru, S. Trivedi and P. Kottapalli (St. Jude Children’s Research Hospital) for WGBS library preparation and Illumina sequencing assistance; T. Mori and Y. Wang (St. Jude Children’s Research Hospital) for statistical consulting; the Animal Resource Center at St. Jude Children’s Research Hospital for mouse husbandry; and A. Moustaki (Yale University) for providing the lentiviral vector backbone. This work was supported by the National Institutes of Health (R01AI114442 and R01CA237311 to B.Y. and F32CA250155 to S.B.) and the American Lebanese Syrian Associated Charities (ALSAC to B.Y.). The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author information
Authors and Affiliations
Contributions
X. Lan and B.Y. designed the experiments and conceptualized the study. X. Lan performed the experiments, and analyzed and visualized the data with help from the co-authors. T.M. and X. Lan analyzed the scRNA-seq, ATAC-seq, CUT&Tag and WGBS data. C.G. performed and analyzed imaging experiments with assistance from X. Liu. M.N.D. performed peak calling and comparison analysis for CUT&Tag. S.A., S.B. and P.C. helped with mouse harvests. M.H. provided technical guidance for CUT&Tag assay. D.Z. and Y.F. provided constructive feedback. X. Lan and B.Y. wrote the paper.
Corresponding author
Ethics declarations
Competing interests
B.Y. declares patents related to epigenetic biomarkers and methods for enhancing T cell function (US11020430B2). The other authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: N. Bernard, in collaboration with the Nature Immunology team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 TCR stimulation strength controls the magnitude of T cell activation during priming.
a, Amino acid sequences of indicated Gp33 variants with alterations highlighted75. b,c, Percentages of GFP (Nr4a)+ P14s in vitro stimulated by indicated concentrations of Gp33 peptides (b) or MC38 tumor cells stably expressing indicated Gp33 variants (c). d, LLC-Gp33 tumor growth in C57BL/6 mice, n = 7–12. e, Percentages of mCherry+ LLC-Gp33 at D18 post tumor inoculation, n = 5 per group. f, LLC-Gp33 tumor growth in Rag-KO mice, n(M9) = 5, n(C6M9) = 6. g, Representative flow plot of P14 percentages in tdLN, related to Fig. 1b. h,i, Percentages of CD44+(h, n = 6–8) and PD1+ (i, n = 3–8) P14s in tdLN. j,m, GMFI of GFP, n = 5. Gated on total (j) or CD44+ (m) P14-Nur77-GFP CD8+ T cells. P value 0.0022 (j). k, GMFI of Tox (left) and Tpex percentages (right) in Rag-KO, n = 4–6. Gated on CD44+ tdLN-P14s. P value 0.0029 (left). l, Percentages of CD62L+ P14s in tdLN, n = 4–6. Gated on CD44+ P14s. n, Percentages of PD1+Tim3+ P14s in tdLN, n = 5–9. d–f,j,k,m, Representative data of two independent experiments, two-tailed unpaired Student’s t test (mean ± s.e.m.), P value ** < 0.01, **** < 0.0001, ns represents not significant. h,i,l,n, Representative data of more than three independent experiments, n represents numbers of mice per group, Multiple two-tailed unpaired t test (mean ± s.e.m.), FDR = 0.05, q value indicated in each graph.
Extended Data Fig. 2 Tpex-associated programs are minimal in suboptimally primed T cells in tdLNs.
a–f Related to scRNA-seq in Fig. 1f, naïve T cell cluster 1 and cluster 9 (low cell count) excluded from analysis. a, Cell number in each cell cluster. b, UMAP colored by cell cycle phases (left) and percentage of cells in each phase (right). c, Barplot displaying percentage of cells in each cell cluster. d, Pathway enrichment analysis of differentially expressed genes (DEGs) in each cluster. Bubble graph displays the top enriched pathways determined by log(q-value). e, Heatmap of the top 10 DEGs for each cluster. f, Relative mRNA expression of representative genes.
Extended Data Fig. 3 Suboptimal TCR stimulation results in a heightened decline in intratumoral Tpex.
a–g Related to Fig. 1a. a, Representative flow plot of P14 percentages in tumors, related to Fig. 2a. b,c, Percentages of Ki67+ (b, n = 4–7) and Gzmb+ (c, n = 3–7) P14s in tumors. d, GMFI of PD1 in tumors, n = 3–5. b–d Gated on P14s, representative data of more than two independent experiments. Multiple two-tailed unpaired t test (mean ± s.e.m.), FDR = 0.05, q value indicated in each graph. e,f,h, GMFI and percentages of CX3CR1+ P14s in tumors, n = 5 (e,h), n = 6–8 (f). P value 0.0434 (e, left) and 0.0198 (h, left). g, GMFI of GFP. Gated on P14-Nur77-GFP CD8+ T cells in LLC-M9 tumors, n = 5 per group. e–h, Representative data of two independent experiments. i, Representative flow plot of T cell percentage in peripheral blood with PBS (Ctr) or FTY720 treatment. j,k, Percentages of P14s in tumors (j, n = 8–13) and tdLN (k, n = 11–13), related to Fig. 2d. l, Percentages of P14s in tdLN, related to Fig. 2fn = 4. m, Percentages and absolute cell numbers of P14s in tdLN, related to Fig. 3a, n(M9) = 9, n(C6M9) = 10. P value 0.0254 (left) and 0.0216 (right). n, Percentages of CX3CR1+ P14s in tumors, related to Fig. 3d. Gated on Tim3+ P14s. n(M9) = 8, n(C6M9) = 6. j–n, Representative data of three independent experiments. e–h,j–n, Two-tailed Student’s t test (mean ± s.e.m.). P value * < 0.05, **** < 0.0001, ns represents not significant. n represents numbers of mice per group.
Extended Data Fig. 4 Optimally primed T cells colocalize with DCs and B cells in tdLN.
a, Representative immunofluorescence images of CD90.1+ (red), CD44+ (green) and CD11c+ (pink) cells in M9- and C6M9-tdLN. Scale bars, 100 μm. b, Boxplot displaying relative proportion (Proportion of P14s – Proportion of DCs) of CD44+ and CD44− P14s in DC clusters in M9- and C6M9-tdLN, n(M9) = 22, n(C6M9) = 14. c, Representative immunofluorescence images of CD90.1+ (red), CD44+ (green) and B220+ (blue) cells in M9- and C6M9-tdLN. Scale bars, 100 μm. d, Boxplot displaying correlation of CD44+ and CD44− P14s with B220+ cells, n(M9) = 22, n(C6M9) = 14. b,d, Representative data of two independent experiments, left-tailed Wilcox signed rank test, Bonferroni adjusted significance level, alpha = 0.0015625. n represents total image numbers per group. For all box-and-whisker plots, the center line denotes the median, the box range denotes the 25th and 75th percentiles and whiskers denote the minimum and maximum values.
Extended Data Fig. 5 DNA methylation programming adapts to TCR engagement strength.
a, Cell sorting strategy for ATAC-seq and WGBS. b, PCA plot based on top 3000 significant CpGs, n = 3 for tdLN samples or n = 1 for tumor samples per group, cells pooled from 3 ~ 5 (M9) or 20 ~ 30 mice (C6M9) for each sample. c, Pie chart displaying distributions of cis-element categories of DMRs in M9-tdLN and C6M9-tdLN. d, Volcano plots of differentially methylated genes between M9-tdLN (orange) and C6M9-tdLN (grey) with the top hypomethylated genes displayed. P value ≤ 0.01, two-tailed Wald test. e, Barplot of top pathways enriched in M9-tdLN-DMRs. f, Heatmap of representative DMRs in M9-tdLN and C6M9-tdLN. g, TF motifs enriched in M9-tdLN-DMRs. h,i, Representative genome tracks displaying chromatin accessibility and DNA methylation at Gzmk (h) and Ptpn5 (i) loci. Red: methylated, Blue: unmethylated.
Extended Data Fig. 6 Subset-specific Tcf1 binding patterns in tumor-reactive CD8 T cells.
a, Summary table of Tcf1 differentially binding regions (DBRs) numbers among naïve P14 (Tn), Tpex from tdLN (LN-Tpex) and Tpex from tumors (TU-Tpex). Two-sided empirical Bayes moderated t-test, FC > 2, P < 0.05. b,f, Signal enrichment heatmap of Tcf1 DBRs between LN-Tpex versus Tn (FC > 2, FDR < 0.05) (b) and TU-Tpex versus LN-Tpex (FC > 2, P < 0.05) (f). c,g, Spearman’s correlation of differential chromatin accessibility with differential Tcf1 binding between indicated groups. Gene region counts in each quadrant (n) and correlation coefficient (r) indicated in each graph. d,h, Top TF motifs enriched among the DBRs of indicated subsets. e,i, Heatmap of Tcf1 binding intensity at representative gene loci.
Extended Data Fig. 7 CRISPR-Cas9 editing of TCR-induced Tcf1 binding site at Slamf6.
Sequence of TCR-induced Tcf1 binding site at Slamf6 (171965904–171966678). Blue, Tcf1 motifs; Green and Red, CRSIPR gRNA.
Supplementary information
Supplementary Table 1 (download XLSX )
scRNA-seq DEG list.
Supplementary Table 2 (download XLSX )
ATAC-seq DAR list.
Supplementary Table 3 (download XLSX )
TCF1-CUT&Tag DBR list.
Source data
Source Data Fig. 1 and Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 2 and Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 and Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 4 and Extended Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 and Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Fig. 7 (download XLSX )
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lan, X., Mi, T., Alli, S. et al. Antitumor progenitor exhausted CD8+ T cells are sustained by TCR engagement. Nat Immunol 25, 1046–1058 (2024). https://doi.org/10.1038/s41590-024-01843-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41590-024-01843-8
This article is cited by
-
Inhibitory PD-1 axis maintains high-avidity stem-like CD8+ T cells
Nature (2026)
-
Regulators of CD8+ T cell exhaustion
Nature Reviews Immunology (2026)
-
The ubiquitin ligase KLHL6 drives resistance to CD8+ T cell dysfunction
Nature (2026)
-
Antigen-presenting cells as arbiters of mucosal tolerance and immunity
Nature Immunology (2025)
-
Regulation of T cell exhaustion and stemness: molecular mechanisms and implications for cancer immunotherapy
Cellular & Molecular Immunology (2025)
