
Abstract
Cutaneous melanoma (CM), with a continuously rising incidence worldwide, represents the most aggressive type of skin cancer, and it leads to the majority of skin cancer-related deaths. Approximately 50% of CM carry the activating BRAFV600 mutation and, although BRAF inhibitors have demonstrated clinical efficacy, most patients often develop early resistance to treatment. Aberrant expression of non-coding RNAs (ncRNAs), which represent less than 2% of the entire transcriptome, has been implicated in CM development and progression. By using BRAFV600-mutant CM in vitro and in vivo models, we have recently demonstrated that the loss of Spry1 expression impairs BRAFV600-mutant CM progression. Therefore, the extensive long and small ncRNA datasets generated in this study might represent a valuable resource for the characterization of their roles in BRAFV600-mutant CM initiation and progression upon Spry1 loss, thus providing a comprehensive resource to support future studies on BRAFV600-mutant CM.
Similar content being viewed by others
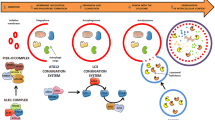
Background & Summary
Cutaneous melanoma (CM) is a growing public health challenge due to its aggressive nature, rising incidence, and high mortality rates. Despite making up only 4% of all skin cancer cases, CM is responsible for nearly 75% of skin cancer-related deaths worldwide, pointing out its severity and the crucial need for improved detection and treatment strategies1. Early-stage localized CM is usually, successfully, treated with surgery2, significantly increasing five-year survival rate3. However, a significant number of cases are diagnosed during a metastatic-advanced stage, when CM has already spread to nearby lymph nodes or other anatomic regions, resulting in frequent unresponsiveness to conventional chemotherapy and leading to a decrease of overall five-year survival to less than 10%4.
Approximately 50% of CM patients harbor activating mutation in codon 600 of the B-Raf Proto-Oncogene Serine/Threonine kinase gene (BRAF)5. Of these, the substitution of valine with glutamic acid variant (V600E) accounts for 70–80% of the BRAFV600 mutations, resulting in the constitutive activation of the mitogen-activated protein kinase (MAPK) pathway, a key driver of CM pathogenesis6. As a result, FDA-approved small molecule inhibitors targeting BRAF (BRAFi) or its downstream effector MEK (MEKi), either alone or in combination, have significantly improved progression-free and overall survival of patients affected by BRAFV600-mutant CM7,8,9. However, intrinsic or acquired resistance to BRAFi/MEKi poses a significant therapeutic challenge, since CM patients frequently do not respond or rapidly develop resistance to this pharmacological regimen10,11, highlighting the urgent need of improved diagnostic and therapeutic strategies.
In the attempt to provide novel insight into the molecular mechanisms involved in CM responsiveness to targeted therapies, we have recently identified the protein Sprouty RTK Signaling Antagonist 1 (Spry1) as a potential oncogene in BRAFV600E-mutant metastatic CM. In fact, by using in vitro and in vivo models, we have demonstrated that Spry1 knock-out (Spry1KO) reduced cell proliferation, causing cell cycle arrest and apoptosis, and enhanced the response to the BRAFi vemurafenib or the MEKi trametininb12. More recently, we have unveiled that Spry1 was mainly localized in mitochondria and that its depletion induced a metabolic rewiring by disrupting mitochondrial homeostasis and impairing glycolytic metabolism in BRAF-mutant CM cells, observing a substantial impairment of angiogenesis following Spry1KO 13, thus suggesting that Spry1 might be exploited as a potential target to improve BRAFV600-mutant CM treatment.
Non-coding RNAs (ncRNAs), which account for 98% of the human genome, represent a class of RNAs without predominant protein coding function. In recent years, ncRNAs have been identified to play a pivotal role in cancer biology and treatment response, emerging as promising diagnostic and prognostic biomarkers in various types of cancer14,15, including CM16,17. These molecules include a broad class of endogenous RNA transcripts such as long non-coding RNAs, linear (in this context, conventionally referred to as lncRNAs) and circular RNAs (circRNAs), as well as small non-coding RNA (sncRNAs), as microRNAs (miRNAs), piwi-interacting RNAs (piRNAs), transfer RNAs (tRNAs), small interfering RNAs (siRNAs), small nuclear RNAs (snRNAs) and small nucleolar RNAs (snoRNAs)18.
LncRNAs are characterized by a linear structure, with transcript lengths equal or greater than 200 nucleotides, while circRNAs have a closed-loop structure, lacking both 3′ and 5′ ends. Both molecules play crucial roles as competitive endogenous RNAs (ceRNAs), accounting for an additional layer of post-transcriptional regulation19. LncRNAs regulate gene expression at epigenetic, transcriptional, and post-transcriptional levels. They can act as “molecular sponges” by competing with miRNAs, DNA, or transcription factors, modulating the deregulation of target proteins. In the nuclear compartment, lncRNAs can recruit or block the binding of transcription factors, directly affecting transcriptional programs. Additionally, they interact with splicing machinery or nascent RNAs, guiding specific splicing patterns20. In the cytoplasm, lncRNAs have proven to impact on gene expression mainly at post-transcriptional level by regulating the polysome loading on messenger RNAs (mRNAs), by controlling internal ribosomal entry sites, or by facilitating mRNA decay21. On the other hand, circRNAs can also act as miRNA sponges to regulate gene expression by controlling alternative splicing, transcription, and protein translation, as well as by influencing the ceRNA mechanism22.
Both lncRNAs and circRNAs have raised increasing interest in CM biology due to their roles in influencing tumor growth, invasion, and resistance to targeted therapies17. Given their aberrant expression in CM, they might serve as promising biomarkers and therapeutic vulnerabilities to be targeted by RNA interference (RNAi), antisense oligonucleotides (ASOs) and CRISPR/Cas9-based genome editing 17. On the other hand, miRNAs represent the most studied sncRNAs in CM as these molecules of approximately 21–23 nt in length play a crucial role in gene regulation by controlling the expression of complementary target mRNAs at the post-transcriptional level, either by repressing translation or promoting their degradation23. Considering their tumor-enhancing or -inhibiting properties, miRNAs are classified as oncomiRs or tumor-suppressor miRNAs24, and an imbalance of their expression levels has been shown to influence the major molecular events that occur during CM development and progression, including cell proliferation, invasion, metastasis, and resistance to MAPK inhibitors25,26,27. As a result, an increasing amount of studies have investigated the predictive and prognostic potential of specific miRNA signatures in CM28,29. Additionally, miRNAs have also emerged as potential non-invasive diagnostic molecules due to their ability to be released into biological fluids30, where they exhibit notable stability and are easily detectable through liquid biopsy31,32.
Considering these premises, the present study aims to provide a useful resource for the characterization of lnc-, circ-, and snc-RNAs in BRAFV600E-mutant metastatic CM. A schematic representation of the study design and workflow is presented in Fig. 1a.

Characterization of lncRNAs, circRNAs, and sncRNAsexpression profiles in BRAFV600E-mutant CM cells and their SpryKO clones. (a) Schematic representation of the experimental workflow followed to generate and validate the sncRNA datasets, including cross-analysis with lncRNAs and circRNAs from both in vitro and in vivo total RNA-seq experiments. (b) Bar plot showing the number of expressed lncRNAs and circRNAs in the indicated cell lines. Log2-transformed values represent the average of replicates. (c) Bar plot showing the number of expressed miRNAs, novel miRNAs (nov-miRNAs), piRNAs, snRNAs, snoRNAs, and tRNAs in the indicated cell lines. Values represent an average of three replicates. (d) Circos plot showing genomic distribution of the first 1000 circRNAs, lncRNAs, and miRNAs detected (reads ≥ 10 for lncRNAs and circRNAs, reads ≥ 3 for miRNAs) in parental cell lines (left) and Spry1KO clones (right).
It is worth noting that despite the availability of studies highlighting the impact of single ncRNA in CM has already been published, our work is the first to broadly encompasses the wide spectrum of expressed lnc- and sncRNAs in BRAFV600E-mutant CM, proposing newly generated dataset from in vitro Mel 593 BRAFV600E-mutant cells. LncRNAs and circRNAs datasets were extracted and annotated and validated by using our already available RNA sequencing (RNA-seq) data12,13. Then, for sncRNA expression profiles, we newly sequenced the small RNA fractions isolated from Mel 593, Mel 599, and Mel 611 BRAFV600E-mutant cells, to characterize all major known sncRNA classes, including miRNAs, piRNAs, snoRNAs, snRNAs, tRNAs and rRNAs.
The analysis of the above-mentioned parental cell lines identified lncRNAs, circRNAs, and sncRNAs commonly expressed (Supplementary Table 1), along with patient-specific ncRNAs.
Evidence suggests that Spry1 may influence the expression of non-coding RNAs indirectly through its role in modulating key signaling pathways, such as MAPK, in BRAFV600-mutant CM cells12,33,34, exerting a regulatory control over gene expression beyond coding transcripts.
To corroborate the specificity of our data, the profile of lncRNAs and circRNAs expressed in xenografts tumors established by the subcutaneous inoculation of the BRAF-mutant Mel 272 parental and Spry1KO cells into athymic nude mice was analyzed according to the analytical steps detailed in the methods section. This additional new dataset may contribute to the identification of ncRNA signatures useful for the stratification of BRAFV600E -mutant metastatic CM patients, supporting their reuse in investigations aimed at elucidating ncRNAs involved in the CM pathogenesis and response to BRAFi.
As shown in Table 1, a total number of 5172 and 5064 distinct sncRNAs, and 29224 and 29115 lncRNAs, were detected in BRAFV600E -mutant parental and Spry1KO CM cells, respectively. The abundance of detected lncRNAs, circRNAs (Fig. 1b), and sncRNA subtypes (Fig. 1c) for each sample was detailed in Table 1. The expression patterns of previously unannotated sequences (novel-miRNAs) were also predicted as a resource for future investigations (Fig. 1c, Supplementary Table 2). Moreover, the chromosomal position of lncRNAs, circRNAs, and miRNAs were investigated in parental and Spry1KOcells (Fig. 1d, Table 2). ncRNA transcripts were found to be expressed on almost all chromosomes in both conditions, except for chromosomes Y. Globally, regions of high miRNA expression were identified on chromosomes X and 1, whereas lncRNA and circRNA genomic positions were found enriched on chromosomes 1,2, and 3 (Table 2).
Methods
Cell lines and generation of Spry1KO BRAFV600E-mutant CM clones
Mel 593, Mel 599, and Mel 611 cell lines were established from metastatic lesions of BRAFV600E-mutant CM patients referred to the National Cancer Institute of Aviano (Italy)35. Spry1KO was performed using the CRISPR/Cas9 gene-editing system, as previously described by Montico and colleagues12. Parental and Spry1KO cell lines were routinely grown in RPMI-1640 medium, supplemented with 10% heat-inactivated fetal calf serum (FCS, Lonza), 100 μg/mL streptomycin, and 100 IU/mL penicillin. Cells were maintained in a humidified incubator at 37 °C in 5% CO2 and routinely tested for mycoplasma contamination.
In vivo models
Xenograft models were established by subcutaneously injecting Mel 272 parental and Spry1KO cells into six-week-old female athymic nude/nude mice, as detailed by Montico and colleagues12. All the preclinical studies were performed in accordance with the Internal Review Board of the Centro di Riferimento Oncologico, IRCCS-National Cancer Institute, Aviano, Italy (IRB number 07-2017) and the Italian Ministry of Health (no. 788/2015/PR).
Western blot analysis for Spry1KO validation
Total protein lysate extraction and western blot were performed as previously reported12,14. The membranes were immunoblotted with the following primary antibodies according to their manufacturer’s instructions: rabbit monoclonal anti-Spry (#13013, Cell Signaling Technologies) and mouse monoclonal anti-β Tubulin (#86298, Cell Signaling Technologies). Images were captured and analyzed using the Chemidoc XRS + system (Bio-Rad). Densitometry was performed by ImageJ software analysis36.
RNA isolation and quality controls
Total RNA for small RNA-seq experiment was extracted from Mel 593, Mel 599, and Mel 611 parental and Spry1KOcells, using Trizol reagent (Invitrogen). Three biological replicates were processed for each sample, resulting in 18 samples. RNA concentration was determined with a NanoDrop ND-2000 spectrophotometer (Thermo Fisher Scientific) by evaluating the absorbance ratios A260/A280 and A230/A280. RNA integrity was assessed using the TapeStation System (Agilent Technologies, Milan, Italy).
Small RNA-seq library preparation and sequencing
For small RNA-seq, 1 μg of total RNA from each cell line was used for library preparation with the Illumina TruSeq Small RNA Sample Preparation Kit (Illumina Inc., San Diego, CA, USA), according to the manufacturer’s protocol. In summary, 3′ and 5′ adapters were ligated to small RNAs in a sequential manner, followed by reverse transcription to synthesize cDNA. The samples were then amplified and indexed through 15 cycles of PCR. The resulting PCR products were purified using a 6% polyacrylamide gel, selecting fragments shorter than 200 nucleotides, and subsequently precipitated with ethanol. The experiment was performed in three independent replicates for each condition. Final library concentrations and sizes were assessed using the Quant-IT DNA Assay Kit and TapeStation System. All libraries were then equimolarly pooled, diluted to a final concentration of 1,3 pM, and sequenced as single reads (75 cycles) on the Illumina NextSeq. 500 platform (Illumina, San Diego, CA).
RNA-seq analysis
Quality control analysis of lncRNAs and circRNAs raw sequences files (fastq files), obtained from previously published RNA-seq datasets12,13 (Mel 599 and Mel 611 clones) as well as the newly available mel 593 RNA-seq data, was performed using FastQc (v0.11.8) (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). Adapter sequences were trimmed using Cutadapt (v3.7)37 with default parameters, setting a minimum read length of 20 bp. Filtered reads were aligned on the hg38 human genome assembly using STAR (v2.7.11b)38 with default parameters and gene quantification was obtained with featureCount (v2.0.3)39 to assess the expression of lncRNAs and circRNAs. The counts were then imported in R (v4.5.0) and DEseq. 2 (v1.48.1)40 was used to identify differentially expressed transcripts, applying a threshold of fold change (|FC|) ≥ 1.5 and an adjusted p-value ≤ 0.05, to evaluate the impact of Spry1KO on lncRNA and circRNA expression profiles. The chromosomal distribution of lnRNAs and circRNAs (reads ≥ 10) was mapped onto the hg38 human genome assembly, whereas Circos41 was used to generate Circos plots. The function EnhancedVolcano (https://github.com/kevinblighe/EnhancedVolcano) was used to generate volcano plots. Heatmaps and hierarchical clustering were generated using countMatrix.
Small RNA-seq analysis
After sequencing, sncRNAfastq files were processed using the miRMaster tool (v2.0)42 selecting the standard parameters for quality filtering, alignment, and annotation of sncRNAs. The miRMaster pipeline was configured to reference the following annotation databases: miRBase (v22.1)43, Ensembl ncRNA (v100) (https://www.ensembl.org/), RNACentral piRNA (v15)44, GtRNAdb (v18.1)45, circBase (accessed on 25.10.20)46, NCBI RefSeq for bacteria and viruses (v74)47, and NONCODE (v5)48. Adapter trimming was performed with Cutadapt (v3.7) using the following small RNA adapter sequences: 3′ adapter TGGAATTCTCGGGTGCCAAGG and 5′ adapter GTTCAGAGTTCTACAGTCCGACGATC. DEseq. 2 (v1.48.1)40 was used to perform a differential expression analysis, applying a threshold of |FC| ≥ 1.5 and an adjusted p-value ≤ 0.05, to detect miRNAs whose expression was significantly modulated by Spry1 depletion. miRNA target prediction was performed using the miRMaster tool. Novel miRNA candidates were predicted by miRMaster through genome-wide mapping of unannotated small RNA reads, followed by secondary structure prediction and evaluation of typical miRNA features such as stem-loop formation, read distribution bias, and minimum free energy criteria. The chromosomal distribution of miRNAs (reads ≥ 3) was mapped onto the hg38 human genome assembly, whereas Circos41 was used to generate Circos plots. The function EnhancedVolcano (https://github.com/kevinblighe/EnhancedVolcano) was used to generate volcano plots. Heatmaps and hierarchical clustering were generated using countMatrix.
Data Records
Raw data generated in the study were deposited in ArrayExpress repository with E-MTAB-1518649 (Mel 593 RNA-seq parental and Spry1KO clones), E-MTAB-1518550 for small RNA-seq (Mel 593, Mel 599, and Mel 611 parental and Spry1KO cells) and E-MTAB-1536951 (Mel 272 RNA-seq parental and Spry1KO in vivo clones). All relevant information concerning samples, useful for their reuse, are available in Table 3.
Technical Validation
Spry1KO validation
To validate the reliability and consistency of our experimental model, we have reported the western blot analysis of Spry1 depletion. As shown in Fig. 2a, Spry1 was not expressed in Spry1KOcells, thus confirming the complete loss of Spry1 protein expression, and ensuring the robustness of subsequent experimental analyses.

Validations of the experimental procedure. (a) Representative western blot and relative densitometry analysis of Spry1 protein expression in parental BRAFV600E-mutant CM cell lines and their respective Spry1KO clones. β-tubulin was used as a loading control. Images were processed with ImageJ software (https://imagej.Net) for densitometry readings. (b) Gel-like images of tape station analysis of total RNA samples from parental and Spry1KO cell lines, showing RIN values. One representative replicate of three is shown for each cell line. EL indicates the electronic ladder. PCA of sncRNAs (c) and ncRNAs (d) in parental and Spry1KO cells. Different shapes (circles, triangles, and squares) indicate biological/technical replicates, color-coded according to cell lines. The first two components (PC1 and PC2) are shown. (e) Correlation heatmaps of miRNAs (left), lncRNAs (middle), and circRNAs (right) expression across the samples.
Assessment of RNA quality
RNA purity and integrity are essential when preparing samples for RNA-seq experiments to ensure the reliability of subsequent experimental analyses. Hence, stringent quality control of RNA samples was performed before library preparation, as described in the methods section. All samples used for the small RNA-seq experiments had an RNA integrity number (RIN) ≥ 9.5, with 260/280 and 260/230 ratios between 1.9 and 2.2, indicating that the RNAs were intact and free of phenolic and protein contaminants (Fig. 2b, Table 4).
Total and Small RNA-seq summarization and quality controls
To assess the sequencing data’s quality and reliability, we evaluated key sequencing parameters from both small RNA-seq (Table 5) and total RNA-seq (Table 6) experiments, including read count and base-call accuracy. The average number of raw reads per sample was 21.08 ± 4.94 million for small RNA-seq and 72.04 ± 20.8 million for total RNA-seq. Notably, the evaluation of alignment efficiency of small RNA-seq experiment revealed a mean alignment percentage to human genome of 92.58 ± 1.86%. The mean GC content per sample was 48.47 ± 0.96% for small RNA-seq and 45.13 ± 0.81% for total RNA-seq, and the average read length was consistent with the expected fragment size for each sequencing approach. The mean Phred score was 35 for both small RNA-seq and total RNA-seq, indicating high base calling accuracy. Additionally, we assessed the same key sequencing parameters for in vivo total RNA-seq experiment (Supplementary Table 3).
Reproducibility validation
Principal component analysis (PCA) was performed to assess the biological significance of Spry1KOon sncRNA and ncRNA expression profiles and to evaluate the accuracy of library preparation and sequencing procedures. The variation along the principal component (PC) 1 axis was above 60% for sncRNAs and above 95% for ncRNAs across all three cell lines, indicating that Spry1KO significantly altered both expression profiles. The variation along the PC2 was below 20% for sncRNAs and 2% for ncRNAs, validating the reproducibility of biological replicates and repeatability of technical assessments (Fig. 2c,d). Furthermore, we performed a correlation analysis on the normalized read counts of miRNAs, lncRNAs, and circRNAs across all samples. The resulting correlation heatmap showed a high intra-group correlation among biological and technical replicates for all ncRNA classes, both before and after Spry1 depletion (Fig. 2e).
ncRNAs differential expression analyses
To further validate our new datasets, lncRNAs and circRNAs derived from Mel 593 were compared to those obtained from Mel 599, and Mel 611 cell lines following Spry1KO 12,13, to enable comprehensive overview of ncRNAs that might be related to Spry1 in BRAFV600E-mutant CM. This revealed 3869, 9397, and 781 differentially expressed lncRNAs (Fig. 3a, Supplementary Table 4) and 1804, 3909, and 656 differentially expressed circRNAs (Fig. 4a, Supplementary Table 5) in Mel 593, Mel 599, and Mel 611, respectively. Moreover, 79 lncRNAs and 90 circRNAs were commonly deregulated across all three cell lines, validating our new datasets, as showed by their expression levels in both parental cell lines and Spry1KO clones (Figs. 3b, 4b, Supplementary Tables 6, 7). Then, we newly assessed their expression in xenograft tumors derived from parental and Mel272 Spry1KO Cl4 cells and observed similar dysregulation for most of them (Figs. 3c, 4c, Supplementary Tables 6, 7). The same approach was applied for miRNAs. The analysis led to the identification of 256, 130, and 96 differentially expressed miRNAs in Mel 593, Mel 599, and Mel 611, respectively (Fig. 5a,b, Supplementary Table 8). This confirmed the reliability of our RNA-seq data sustaining that the experiments were conducted appropriately and match among different methods and settings.

Analysis of lncRNA expression profiles. (a) Volcano plot summarizing significantly up-regulated (red) and down-regulated (green) lncRNAs in SpryKO clones; lncRNAs with insignificant expression values are reported in grey. Significance was determined based on log2 FC cutoff of ± 1.5 and adjusted p-values threshold of 0.05. (b) Heatmaps showing expression (left) with respect to FC (right) of common, concordantly deregulated lncRNAs (|FC| ≥ 1.5, adjusted p-value ≤ 0.05) across Spry1KO clones compared to the parental cell lines. (c) Heatmap showing expression of the same commonly deregulated lncRNAs in xenograft tumors derived from Mel272 parental and Spry1KO cells. RNA-seq data supporting Fig. 3 are available in ArrayExpress with accession number E-MTAB-1518649 (Mel 593 in vitro clones) and E-MTAB-1536951 (Mel 272 in vivo clones).

Analysis of circRNA expression profiles. (a) Volcano plot summarizing significantly up-regulated (red) and down-regulated (green) circRNAs in SpryKO clones;circRNAs with insignificant expression values are reported in grey. Significance was determined based on log2FC cutoff of ± 1.5 and adjusted p-values threshold of 0.05. (b) Heatmaps showing expression (left) with respect to FC (right) of common, concordantly deregulated circRNAs (|FC| ≥ 1.5, adjusted p-value ≤ 0.05) across Spry1KO clones compared to the parental cell lines. (c) Heatmap showing expression of the same commonly deregulated circRNAs in xenograft tumors derived from Mel272 parental and Spry1KO cells. RNA-seq data supporting Fig. 4 are available in ArrayExpress with accession number E-MTAB-1518649 (Mel 593 in vitro clones) and E-MTAB-1536951 (Mel 272 in vivo clones).

Analysis of miRNA expression profiles. (a) Heatmaps showing up-regulated (red) and down-regulated expressed (green) miRNAs upon Spry1 depletion (|FC|) ≥ 1.5, adjusted p-value ≤ 0.05) in the indicated cell lines. (b) Volcano plot summarizing significantly up-regulated (red) and down-regulated (green) miRNAs in SpryKO clones; miRNAs with insignificant expression values are reported in grey. Significance was determined based on log2 FC cutoff of ± 1.5 and adjusted p-values threshold of 0.05. Commonly deregulated miRNAs across the clones are highlighted in bold. Small RNA-seq data supporting Fig. 5 are available in ArrayExpress with accession number E-MTAB-1518550 (Mel 593, Mel 599, and Mel 611 parental and Spry1KO cells).
References
Gosman, L. M., Țăpoi, D.-A. & Costache, M. Cutaneous melanoma: A review of multifactorial pathogenesis, immunohistochemistry, and emerging biomarkers for early detection and management. Int. J. Mol. Sci. 24 (2023).
Orzan, O. A., Șandru, A. & Jecan, C. R. Controversies in the diagnosis and treatment of early cutaneous melanoma. J. Med. Life 8, 132–141 (2015).
Savage, D. J. et al. Patterns in progression from early-stage melanoma to late-stage melanoma: implications for survivorship follow-up. Melanoma Manag. 11, 2424708 (2024).
Kudchadkar, R. R., Lowe, M. C., Khan, M. K. & McBrien, S. M. Metastatic melanoma. CA Cancer J. Clin. 70, 78–85 (2020).
Castellani, G. et al. BRAF mutations in melanoma: biological aspects, therapeutic implications, and circulating biomarkers. Cancers (Basel) 15 (2023).
Guo, W., Wang, H. & Li, C. Signal pathways of melanoma and targeted therapy. Signal Transduct. Target. Ther. 6, 424 (2021).
Giugliano, F. et al. First line treatment of BRAF mutated advanced melanoma: Does one size fit all? Cancer Treat. Rev. 99, 102253 (2021).
Tanda, E. T. et al. Current state of target treatment in BRAF mutated melanoma. Front. Mol. Biosci. 7, 154 (2020).
Grimaldi, A. M. et al. MEK inhibitors in the treatment of metastatic melanoma and solid tumors. Am. J. Clin. Dermatol. 18, 745–754 (2017).
Dulgar, O., Kutuk, T. & Eroglu, Z. Mechanisms of Resistance to BRAF-Targeted Melanoma Therapies. Am. J. Clin. Dermatol. 22, 1–10 (2021).
Luebker, S. A. & Koepsell, S. A. Diverse mechanisms of BRAF inhibitor resistance in melanoma identified in clinical and preclinical studies. Front. Oncol. 9, 268 (2019).
Montico, B. et al. Loss of Spry1 reduces growth of BRAFV600-mutant cutaneous melanoma and improves response to targeted therapy. Cell Death Dis. 11, 392 (2020).
Montico, B. et al. Suppression of Spry1 reduces HIF1α-dependent glycolysis and impairs angiogenesis in BRAF-mutant cutaneous melanoma. J. Exp. Clin. Cancer Res. 44, 53 (2025).
Zhang, X. et al. Non-coding RNAs’ function in cancer development, diagnosis and therapy. Biomed. Pharmacother. 167, 115527 (2023).
Eldakhakhny, B. et al. Exploring the role of noncoding RNAs in cancer diagnosis, prognosis, and precision medicine. Noncoding RNA Res 9, 1315–1323 (2024).
Peng, Q. & Wang, J. Non-coding RNAs in melanoma: Biological functions and potential clinical applications. Mol. Ther. Oncolytics 22, 219–231 (2021).
Montico, B. et al. The pleiotropic roles of circular and long noncoding RNAs in cutaneous melanoma. Mol. Oncol. 16, 565–593 (2022).
Russell, S. J. et al. An atlas of small non-coding RNAs in human preimplantation development. Nat. Commun. 15, 8634 (2024).
Ala, U. Competing Endogenous RNAs, Non-Coding RNAs and Diseases: An Intertwined Story. Cells 9 (2020).
Fernandes, J. C. R., Acuña, S. M., Aoki, J. I., Floeter-Winter, L. M. & Muxel, S. M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Noncoding RNA 5 (2019).
Statello, L., Guo, C.-J., Chen, L.-L. & Huarte, M. Author Correction: Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 22, 159 (2021).
Conn, V. M., Chinnaiyan, A. M. & Conn, S. J. Circular RNA in cancer. Nat. Rev. Cancer 24, 597–613 (2024).
O’Brien, J., Hayder, H., Zayed, Y. & Peng, C. Overview of microrna biogenesis, mechanisms of actions, and circulation. Front Endocrinol (Lausanne) 9, 402 (2018).
Otmani, K., Rouas, R., Berehab, M. & Lewalle, P. The regulatory mechanisms of oncomiRs in cancer. Biomed. Pharmacother. 171, 116165 (2024).
Poniewierska-Baran, A. et al. Role of miRNA in Melanoma Development and Progression. Int. J. Mol. Sci. 24 (2022).
Gajos-Michniewicz, A. & Czyz, M. Role of miRNAs in Melanoma Metastasis. Cancers (Basel) 11 (2019).
Motti, M. L., Minopoli, M., Di Carluccio, G., Ascierto, P. A. & Carriero, M. V. Micrornas as key players in melanoma cell resistance to MAPK and immune checkpoint inhibitors. Int. J. Mol. Sci. 21 (2020).
Lu, T. et al. Identification of a five-miRNA signature predicting survival in cutaneous melanoma cancer patients. PeerJ 7, e7831 (2019).
Dika, E. et al. Defining the prognostic role of micrornas in cutaneous melanoma. J. Invest. Dermatol. 140, 2260–2267 (2020).
Pozniak, T., Shcharbin, D. & Bryszewska, M. Circulating microRNAs in Medicine. Int. J. Mol. Sci. 23 (2022).
Sabato, C. et al. A novel microRNA signature for the detection of melanoma by liquid biopsy. J. Transl. Med. 20, 469 (2022).
Van Laar, R. et al. Validation of a microRNA liquid biopsy assay for diagnosis and risk stratification of invasive cutaneous melanoma. Br. J. Dermatol. 189, 292–301 (2023).
Saj, A. & Lai, E. C. Control of microRNA biogenesis and transcription by cell signaling pathways. Curr. Opin. Genet. Dev. 21, 504–510 (2011).
Ishikawa, T. et al. Long Non-Coding RNAs Associated with Mitogen-Activated Protein Kinase in Human Pancreatic Cancer. Cancers (Basel) 15 (2023).
Altomonte, M. et al. Differential expression of cell adhesion molecules CD54/CD11a and CD58/CD2 by human melanoma cells and functional role in their interaction with cytotoxic cells. Cancer Res. 53, 3343–3348 (1993).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675 (2012).
Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet j. 17, 10 (2011).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Liao, Y., Smyth, G. K. & Shi, W. The R package Rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 47, e47 (2019).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. 2. Genome Biol. 15, 550 (2014).
Krzywinski, M. et al. Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–1645 (2009).
Fehlmann, T. et al. miRMaster 2.0: multi-species non-coding RNA sequencing analyses at scale. Nucleic Acids Res. 49, W397–W408 (2021).
Kozomara, A., Birgaoanu, M. & Griffiths-Jones, S. miRBase: from microRNA sequences to function. Nucleic Acids Res. 47, D155–D162 (2019).
The RNAcentral Consortium. RNAcentral: a hub of information for non-coding RNA sequences. Nucleic Acids Res. 47, D221–D229 (2019).
Chan, P. P. & Lowe, T. M. GtRNAdb 2.0: an expanded database of transfer RNA genes identified in complete and draft genomes. Nucleic Acids Res. 44, D184–9 (2016).
Glažar, P., Papavasileiou, P. & Rajewsky, N. circBase: a database for circular RNAs. RNA 20, 1666–1670 (2014).
Pruitt, K. D., Tatusova, T. & Maglott, D. R. NCBI reference sequences (RefSeq): a curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 35, D61–5 (2007).
Fang, S. et al. NONCODEV5: a comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 46, D308–D314 (2018).
Memoli, D. https://identifiers.org/arrayexpress:E-MTAB-15186 (2025).
Memoli, D. https://identifiers.org/arrayexpress:E-MTAB-15185 (2025).
Memoli, D. https://identifiers.org/arrayexpress:E-MTAB-15369 (2025).
Acknowledgements
This work was supported by AIRC Foundation for Cancer Research, Grant No. IG-23068 (to A.W.), Italian Ministry of University and Research PNRR-MUR NextGenerationEU PRIN 2022, cod. 202282CMEA - CUP: D53D23007790001 (to G.N.) and cod. 2022Y79PT4 – CUP: D53D23008040006 (to A.W.); PNRR-MUR NextGenerationEU PRIN-PNRR 2022 cod. P2022N28FJ - CUP: D53D23016530001 (to G.N.); Italian Ministry of Health (Ricerca Corrente), Young Researcher Grant GR-2021-12373937 (to G.N.) and Grant GR-2018-12366312 (to E.F. and A.W); 5 × 1000 Institutional Grant from CRO Aviano, National Cancer Institute, Istituto di Ricovero e Cura a Carattere Scientifico (IRCCS) seed grant to B.M. and E.F., no grant number provided.
Author information
Authors and Affiliations
Contributions
Conceptualization: E.F., G.N., A.S.; investigation: J.L., B.M., D.M., R.G., F.C., A.S., E.F.; bioinformatic analyses: D.M., F.S.; writing and original draft preparation, J.L., G.N., A.S.; writing, review, and editing: J.L., E.F., B.M., G.N., A.S., A.W.; funding acquisition: E.F., B.M., G.N., A.W. All the authors approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lamberti, J., Memoli, D., Montico, B. et al. Non-coding RNA profiling in BRAFV600E-mutant cutaneous melanoma before and after Spry1 depletion. Sci Data 12, 1538 (2025). https://doi.org/10.1038/s41597-025-05807-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41597-025-05807-x
