
Abstract
Several dog skeletons were excavated at the Roman town of Augusta Raurica and at the military camp of Vindonissa, located in the northern Alpine region of Switzerland (Germania Superior). The relationships between them and the people, the nature of their lives, and the circumstances of their deaths are unclear. In order to gain insight into this dog population, we collected 31 dogs deposited almost simultaneously in two wells (second half of the third century CE), three dogs from burial contexts (70–200 CE and third to fifth century CE) at Augusta Raurica, and two dogs from burial contexts at Vindonissa (ca. first century CE). We detected a mixed population of young and adult dogs including small, medium and large sized individuals. Three small dogs had conspicuous phenotypes: abnormally short legs, and one with a brachycephalic skull. Stable isotope analysis of a subset of the dogs showed that their diets were omnivorous with a substantial input of animal proteins and little variation, except one with a particularly low δ15N value, indicating a diet low in animal proteins. Partial mitochondrial DNA sequences from 25 dogs revealed eight haplotypes within canine haplogroup A (11 dogs; 44%; 5 haplotypes), C (8 dogs; 32%; 1 haplotype), D (4 dogs, 16%; 1 haplotype) and B (2 dogs, 8%; 1 haplotype). Based on shotgun sequencing, four Roman mitogenomes were assembled, representing sub-haplogroups A1b3, A1b2 and C2. No canine pathogens were identified, weakening the assumption of infectious disease as a cause for dog disposal. The genetic and morphological diversity observed in dogs of Augusta Raurica and Vindonissa is similar to modern dog diversity.
Similar content being viewed by others
Introduction
Since their domestication before the emergence of Neolithic farming, dogs diversified in appearance, behaviour and function, a process still ongoing1,2,3,4,5,6,7,8. While accompanying people across the world a close relationship was established and as consequence of their various exploitations by humans, dogs gained the ability to digest starchy food and underwent bodily changes, e.g. coat colour and floppy ears9.
Although various body sizes and coat colours are already attested for in the Magdalenian time period (18′000–12′000 BCE) and throughout prehistory10, in Europe dogs increasingly diversified during the Roman period, a process which likely began around the 3rd century BCE in Italy. This is well documented in texts, depicted in mosaics, and supported by objects (see Fig. 1) and archaeology11,12,13,14,15,16,17,18,19. During the Roman period dogs were particularly esteemed as pets, with a number of small dogs appearing in the Roman homeland and provinces16,20. For example, small-sized dogs with a brachycephalic skull (similar to Pekingese) and/or with abnormally short limbs (similar to Dachshund, Basset Hound), have been described in the archaeozoological record from a variety of Roman sites in Britannia17,21,22, Hispania16,23, Italia24 and Pannonia25. Together, the data suggest dog breeding for the purpose of selection for a specific phenotype, such as size, shortened limbs or slenderness.

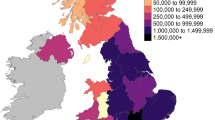
Map with approximate location of sites in Switzerland. Free available map downloaded from justfreeslide.com (https://justfreeslide.com/download/editable-europe-map-for-powerpoint/) and edited using Microsoft Office 2016. Dog figurine: Bronze key handle of a dozing dog from Augusta Raurica. Courtesy of Augusta Raurica.
Normally dogs were not part of the human diet in the Roman period and were not slaughtered26. In some cases, their dead bodies were even carefully buried, for example, at the Roman pet cemetery in Berenice, Egypt27. However, more often their bodies were deposited in rubbish pits or in wells out of use28,29. However, dog health, feeding and welfare issues and their complex relationship to people have rarely been addressed or brought into focus30,31.
In addition to archaeozoology, stable isotopes and ancient DNA offer further perspectives for addressing social relationships between humans and dogs. Stable isotope ratios in skeletal remains reflect dietary compositions32 and have been widely used to study faunal dietary diversity33. Therefore, stable isotope analysis of dog bones may shed light on their diet and living conditions. Roman authors actually recommended specific diets depending on dogs’ tasks and life stage and presumably the composition of dog food in Roman times may have varied due to the individual feeding preferences of dog owners31.
Genetically dogs are highly diverse. Based on mitochondrial DNA (mtDNA), European dogs belong to one of the four major haplogroups A-D34,35,36 with a dominance of A lineages in modern times, in contrast to a dominance of haplogroup C lineages during the Neolithic, suggesting a prehistoric turnover (introductions/replacements) of mtDNA lineages in Europe1,4,37. However, haplogroup A lineages have continuously dominated in Iberia since the Mesolithic and other lineages in central Europe have also likely survived since the Neolithic3,19,38. For the Roman era genetic information about dogs is scarce. Only two genetic studies addressing Roman dogs mtDNA diversity in Iberia/Morocco20 and Bulgaria5 are currently available. Their results show high mtDNA diversity with uneven geographic distribution of haplotypes belonging to the major dog haplogroups A and D (Iberia) and A, B and D (Bulgaria), denoting different histories of maternal dog lineages between the different regions of the western and eastern part of the Roman Empire, and possibly suggesting diverse centers of dog “breeding” activities.
With the northward expansion of the Roman Empire beyond the Alps and the establishment of the imperial province Germania Superior, economic, cultural and military centers such as the colonial town of Augusta Raurica and the military camp of Vindonissa in the north of Switzerland were founded39,40 (see also Supplementary Information). Large socio-economical changes occurred such as the introduction of horticulture, new crops and spices, new domestic animals and new technologies to Switzerland. The region was prospering and trading intensified41. People were very mobile in the Roman Empire42,43 and they transported not only objects related to daily life but they were also accompanied by their animals such as dogs. Therefore, the region considered here probably became a melting pot not only for people but also for animals. In the 2nd half of the third century CE, while the once prosperous city Augusta Raurica gradually deteriorated, several dogs were deposited in different wells, a common praxis of animal disposal during the Roman period in Europe28,29,44. In the case of the wells of Augusta Raurica, however, the archaeological evidence points to quick depositions within a few years or even within a few months suggesting that the animals were living at the same time and deposited almost simultaneously. Potential circumstances accompanying this event are still under debate39,44,45,46,47,48. The dogs from Vindonissa were from burial contexts and date to the 1st century CE.
This research combined osteometry, mitochondrial DNA D-loop typing, mitogenome analysis, shotgun sequencing to screen for pathogens and stable isotope analyses of dogs from Augusta Raurica and Vindonissa. The aim was to gain insight into the size, age and skeletal particularities, into the genetic variation in terms of mtDNA haplogroups and haplotypes and potential diseases as well as the diet of dogs from a local and near contemporaneous context of two wells at Augusta Raurica. The study was extended to include dogs from different time periods of Augusta Raurica and Vindonissa (Fig. 1).
Material and methods
Study sites and archaeological material
Samples from 36 almost complete individual Roman dog skeletons were used in this study. They originate from the town of Augusta Raurica (n = 34) and from the legionary camp of Vindonissa (n = 2) and were made available for examination by the archaeological services of Augusta Raurica and the canton of Aargau. Both sites are located in the northern Alpine margins of the Roman Imperium (today Switzerland) (Fig. 1; Table 1, Supplementary Information; Table S1). Most of the specimens from Augusta Raurica (n = 31) were discovered in two wells, namely well MR 12 ("Sodbrunnen MR 12") located in the Lower Town and well house in Insula 8 ("Brunnenhaus") in the Upper Town, filled with animal remains of different wild and domestic species (e.g., horses, pigs, bears) a custom only observed in the 2nd half of the third century CE at this site and likely coinciding with the urban decline of the city44,47. Numerous artefacts were also found in the wells, such as short term dated coin casting molds and pottery. Matching ceramic fragments were found scattered throughout the wells, proving that the filling is to be considered as a unit and that it took place in a short time or even in one event44 and for both wells at the same time span. This is also supported by the nearly completeness of the animal skeletons (Fig. S1).
Additionally, two individuals were from pits and one from a hypocaust room in the residential area of Augusta Raurica49,50,51 (Table 1, Table S1). One of the dogs from Vindonissa was found in a cemetery52, the other (headless) dog within an officer’s house close to two neonate burials40 (Table 1, Table S1). Only samples from distinguishable dog individuals were used in this study.
Metrics and morphology
Osteological analyses were carried out at IPAS, Basel, Switzerland. Withers height calculations (cm) were attempted for all individuals according to Clark53 and Koudelka54 and particular morphological features of limb bones and, in few cases skulls were also recorded (see Supplementary Information).
Stable isotope analysis
Stable isotope analyses were carried out at IPAS, Basel, Switzerland. Stable carbon and nitrogen isotope analysis was performed on 18 dog individuals from Augusta Raurica, of which 9 were also analysed for aDNA. The same bone specimens were used for both analyses (Table S1). Analytical protocols followed Longin55 with modifications as described in56 (for more details see Supplementary Material).
Ancient DNA methods
All aDNA data of the bones were produced independently in Basel, Switzerland and Kiel, Germany.
Authentication of aDNA and mtDNA analysis at Basel University
To prevent contaminations, handling and processing of ancient samples followed strict standards in aDNA research established at Integrative Prehistory and Archaeological Science (IPAS)57,58,59. For details see Supplementary Information.
A 97-bp diagnostic fragment of the dog mitochondrial D-loop1 was PCR-amplified and sequenced to determine mtDNA haplotypes in 28 dog individuals (Table 1, Supplementary Methods, Table S1). To study relatedness in a historical context, obtained sequences were compared to published Eurasian modern, Roman and pre-Roman dog sequences with known haplogroup affiliations by using Median-Joining Network (MJN) analysis (Supplementary Methods, Table S2). Genetic diversity for 23 dogs from Augusta Raurica was calculated with Arlequin 10.060 and haplogroup frequencies for Roman and pre-Roman periods and for modern dog breeds were estimated from published data (Table S2, Supplementary Information).
Sample processing and aDNA analysis at Kiel University
Sample processing and DNA analysis were performed in clean room facilities dedicated to aDNA research at Kiel University and following the guidelines on contamination control in aDNA studies61,62,63 as described in Krause-Kyora et al.64. Dog samples from Augusta Raurica were pre-screened for pathogens using a shotgun sequencing approach (Table S3). Four complete dog mitochondrial genomes were assembled. A maximum likelihood tree was generated and visualized by using FigTree (Supplementary Information). Haplogroup/type assignment was according to Duleba et al.36.
Results
Morphometrics and morphology
Dog withers heights were calculated from different complete elements: femur, metapodia, humerus, tibia, ulna, calcaneus and radius. For 33 out of 36 dogs analyzed, representing 27 full-grown adult and 9 juvenile dogs highly variable sizes were obtained (Table 1, Table S1, Fig. S2). Three dogs were only represented by fragments; therefore, no withers heights could be calculated. The smallest dog had a withers height of 19 cm and the tallest reached a size of 76 cm. Adult dogs were either small (< 39 cm, n = 7), medium-sized (40–59 cm, n = 13) or large (> 60 cm, n = 6). The data show features of a morphologically varied and age-mixed population with slight differences in size distribution between wells: more medium-sized adult dogs were found in well MR 12 (“Sodbrunnen MR 12”) (n = 9, out of 12 adults) whereas smaller and larger adult dogs (n = 5 and n = 4, respectively, out of 11 adults) stem from the well house in Insula 8 (“Brunnenhaus”). The two dogs from Vindonissa (HAV1, HAV2) found in a burial context were small sized although HAV1 was a juvenile individual which likely had not attained its full size yet (see Fig. S2).
Four small adult dogs stand out morphologically. Dog HAL16/HAL10 from well MR 12 (“Sodbrunnen MR 12”) was abnormally short-legged (i.e. brachymelic, as confirmed by two bone elements). Further two dogs HAU12 and HAU14 from well house in Insula 8 (“Brunnenhaus”) were brachymelic too, but additionally the skull of subject HAU14 showed the characteristic traits of a brachycephalic (“short-headed”) dog (Table S1). To our best knowledge this is the first report of brachymelia and brachycephaly in dogs from this part of the Roman Empire (e.g. Roman Switzerland). Additionally, one small dog (individual HAU2) was slender. Altogether, four small adult dogs from a total of ten small dogs (seven adults) showed particular morphologies.
Carbon and nitrogen isotope analysis
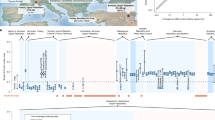
Collagen of all 18 samples fulfilled the quality criteria for ancient collagen as suggested by Ambrose65 and van Klinken66. They yielded between 2.9 and 17.0% of collagen, 28.6 to 44.2% of C, 10.5 to 16.0% of N, and had atomic C/N ratios of between 3.1 and 3.4. The analytical data are listed in Table S1. The δ13C and δ15N values of the dog samples from Augusta Raurica varied between − 20.7 and − 18.2‰ and 7.4 and 9.8‰ respectively, and are in the range of published stable isotope data for dogs (see Fig. 2) indicative of an omnivorous diet56,67,68,69,70,71,72. The difference in δ13C and δ15N between dogs and herbivores from Augusta Raurica73 accounts for almost one trophic level74 indicating also substantial animal protein intake. δ15N variation was limited to 2.4‰, and δ13C showed a range of 2.5‰. Most samples clustered within a range of 1.7‰ in respect to both δ13C and δ15N, however, with the exception of outlier value HAU5 (Grubbs’s test (2-sided): δ13C: Z = 2.708, p = 0.038; δ15N: Z = -2.260, but p = 0.265). This large adult individual displayed a combination of low δ15N and high δ13C (Fig. 2) indicating a different dietary signature. Three dogs derive from features other than wells. Two of these are of earlier date (HAU9, HAU18) and displayed moderate but statistically significant differences to the dogs from wells (dating to the 2nd half of the third century CE) in respect to δ13C (Student’s t-test: t(15) = 2.698; p = 0.017) suggesting potential differences between these time periods.

δ13C and δ15N values measured in bone collagen sampled from dog skeletal remains from Augusta Raurica (AR). Symbol shades denote haplogroups (Hg) (A–D) and unknown haplogroups. Isotopic values are plotted against herbivore and herbivorous omnivore stable isotope data from the same site (mean ± 1SD) and selected dog stable isotope data from Roman contexts across Europe and from the Late Iron Age site of Basel-Gasfabrik (2nd-1st century BCE), located < 15 km from Augusta Raurica.
Mitochondrial DNA D-loop diversity
PCR amplification of the short mtDNA D-loop segment (97 bp, without primer) was successful in 24 dogs (out of 26) from Augusta Raurica and in one dog (out of two) from Vindonissa. Attempts to obtain a longer D-loop fragment (255 bp, without primer) failed in all samples positive for the shorter fragment. In total, 8 haplotypes could be assigned to one of the four major haplogroups A-D (Table 1, Tables S1, S2, Fig. 3, 4). Five haplotypes (RS-a1 – RS-a5) clustered within haplogroup A (11 dogs, 44%) which constitutes the most diverse dog haplogroup known today36. Haplotype RS-a5 (HAU7) is a private variant matching no identical sequence in public databases. Further, haplotype RS-a3 found in three dogs from Augusta Raurica (HAL2, HAL8, HAU3) is not shared with any other Roman dog (Fig. 3, Table S2). Three haplotypes clustered within haplogroup C (RS-c1, 8 dogs, 32%), D (RS-d1, 4 dogs, 16%,) and B (RS-b1, 2 dogs, 8%) (Fig. 3). The relatively high haplotype (0.86 ± 0.04) diversity is comparable to genetic diversity indices reported from modern native breeds from Portugal and Iran75,76.

Median-Joining Network of Roman (n = 45) and modern (n = 34) mtDNA sequences (82 bp). Haplogroups (A–D) follow nomenclature of Duleba et al. (2015) based on whole mitochondrial genomes. Eight different haplotypes were found in Roman Switzerland. Median vector (filled rhombous) denotes hypothetical haplotype. Further details as listed in Table S2.

Visualization of haplogroup (Hg) (A–D) frequencies (0–100%, y-axis) in pre-Roman (a) and Roman (b) times in Eurasia: Iberia (I), Iberia/Morocco (I/MA), Switzerland (CH), Bulgaria (BG), Italy (IT), Central Western Europe (CWE), Southeast Europe (SEE), Western Northern Europe/Russia and Southwest and Central Asia (SWCA) as listed in Table S2. Free available map downloaded from slidelizard.com (https://slidelizard.com/en/blog/powerpoint-world-map) and edited using Microsoft Office 2016.
Haplotype distribution between Roman dogs and relatedness to pre-Roman dogs
To date, only two genetic studies of Roman dogs from Iberia/Morocco20 and Bulgaria5 are available. Consequently, dog mtDNA sequences from Augusta Raurica and Vindonissa were compared to mtDNA sequences from these two studies (Table S2, Fig. 3, Fig. S4). Two haplogroups were found in dogs of Roman Iberia/Morocco20 and, except for one (LYEP60), all their haplotypes are shared with dogs from Augusta Raurica and Vindonissa (e.g., RS-a1, RS-a2, RS-a4; and RS-d1) (Fig. 3). Four dog samples from Roman Bulgaria clustered within haplogroups A, B and D5 but haplotypes were neither shared with Augusta Raurica and Vindonissa nor with Iberia/Morocco.
By comparing Roman with pre-Roman sequences, shifts in haplotype frequencies were observed. For example, while most haplogroup A sequences affiliate with cluster “RS-a4” in pre-Roman Iberia/Morocco, a second cluster “RS-a2” appears with increased numbers of sequences and haplogroup D appears for the first time in Roman Iberia/Morocco (Table S2, Fig. S4). Similarly, haplogroup A and B haplotypes from Roman Bulgaria differ from pre-Roman haplotypes, and haplogroup D emerges for the first time in Roman Bulgaria (Fig. S4, Table S2). In Roman Switzerland new A and D haplotypes replace haplotypes from previous periods and haplogroup B is newly introduced (Fig. S4, Table S2). Interestingly, cluster “RS-d1” with shared sequences between Roman Iberia and Switzerland, is the most frequent D cluster in pre-Roman Southeast Europe (Table S2). While some haplogroup C lineages may have persisted through times in Switzerland (Fig. S4) others are new (see mitogenome HAL11 1700–1650 (CH), Fig. 5).

Maximum-likelihood tree of mitochondrial dog genome sequences. The tree contains four mitogenomes from Augusta Raurica (this work, in red text) along with published mitogenomes of modern (n = 512) and ancient (n = 38) dogs as listed in Table S6. An asterix (*) indicates no bootstrap values provided as the outgroup was used for tree rooting after inference. Bootstrap values (in percentage, from 500 replicates) are provided for internal branches of the tree.
Shifts in the proportion of haplogroups are also observed (see Fig. 4): an increase in haplogroup A and D frequencies, the disappearance of C lineages in Roman Iberia/Morocco20,38; a decrease in haplogroup A and an increase in haplogroup D frequencies in Roman Bulgaria5. Notably, Bulgaria preserves the highest ratio of haplogroup B (ca. 30%) over time (Fig. 4). An increase in haplogroup A and B ratios, and a reduction of haplogroup D proportions is noted in Roman Switzerland. Pre-Roman frequencies of haplogroups A, B, C and D vary considerably between regions in Eurasia and contrast to frequencies reported from modern dogs (see Fig. S5).
Relationships between phenotypes, genetics and diet
No obvious relationships between haplotypes, sizes, age or isotopic signatures was detected. Haplotype and accordingly haplogroup distribution did not show any particular association with size groups (Figs. S2 and S6). Stable isotope ratios of small, medium and large dogs overlapped with no apparent trend, that is, the dogs fed on similar, slightly varying omnivorous diets with small differences irrespective of their size (Fig. S3). Interestingly, stable isotope values of the three smallest dogs with abnormally short limbs HAL16/HAL10 and HAU12 (brachymelic) and HAU14 (brachymelic/brachycephalic) were within the same range and the only dog (HAU5) with outlier stable isotope values (Fig. 2, Table S1) showed no particular exceptional morphologies besides being large.
Shotgun sequencing, pathogen screening and recovery of four mitogenomes
Shotgun sequences from 14 dog individuals from Augusta Raurica were generated. In general, the content of endogenous dog DNA was low (Table S5). The DNA displayed the damage pattern expected for aDNA (see Supplementary Methods). The genome-wide data were screened for pathogens (see Supplementary Information)77 but no bacterial or viral pathogens were detected. High-coverage mitochondrial genomes could be assembled for samples HAL2, HAL8, HAL11, HAL16. A maximum likelihood tree was constructed to view their position within other dog mitogenomes (Fig. 5). The tree contained published whole mitochondrial genome sequences from modern and ancient dogs representing the four well-supported major clades A, B, C and D78 and the new proposed clade X found in ancient Siberian dogs79 (see Table S6). One Roman mitogenome HAL11 located within clade C (close to C2), and three within clade A1: mitogenome HAL2 and HAL8 were slightly different from each other (otherwise identical based on the D-loop fragment) and clustered within sub-haplogroup A1b3 whereas mitogenome HAL16 clustered within sub-haplogroup A1b2 (see Fig. 5). PCR-based haplogroup classification based on the short D-loop fragment for individuals HAL2, HAL8, HAL11 and HAL16 was confirmed by their respective mitogenomes. Interestingly, the tree position of the four Roman mitogenomes differed from the two available Palaeolithic mitogenomes from Kesslerloch cave in Switzerland that cluster within C and A (TU11 and TH11, respectively)78,80.
Discussion
The present study used an interdisciplinary research approach to analyze a large number of Roman dog individuals including dogs deposited in two wells that likely lived contemporaneously around the mid-third century CE in the town of Augusta Raurica, Germania Superior, a unique assemblage of bone deposits in Northern Switzerland. Osteometric analysis of all individuals (n = 33), revealed a broad range of sizes providing the first morphometric information from a large population of dogs living on the edge of the Roman Empire. The dogs were classified into three size categories, e.g. small (n = 10), medium (n = 17) and large (n = 6) and included young and adult individuals. Overall, the dogs show a similar size variation to dogs across other Roman regions and sites, e.g. Pompeii (Italia)24, Vindolanda (Britannia)17,81, and Hispania Tarraconensis16,17,24. The detection of three small-sized adult and abnormally short-limbed dogs (HAL16/HAL10, HAU12, HAU14) one of which with a brachycephalic skull (HAU14), and a slender dog (HAU2), denotes distinctive morphotypes of small dogs co-occurred at Augusta Raurica. A similar short-limbed small brachycephalic dog was reported from the Roman cemetery of Llanos del Pretorio (Córdoba, Spain)23, implying that such combined traits were not unique and restricted to one particular region. This raises the question of where this dog type was first bred and whether they were indeed lapdogs, as previously suggested for these kinds of small individuals23,24,82. Two dogs, one juvenile (HAV1) and one adult (HAV2) from the military camp of Vindonissa were small and shared no particular phenotype but were found in burial contexts, potentially being esteemed household dogs of legionary people.
In contrast, it is rather surprising to find a brachycephalic dog with such a particular trait disposed in a well. Was this dog not considered as something special (e.g. prestige pet?) and worth a proper burial? Was it subject to a ritual act, or was it a precipitous disposal with another background? The same questions may also apply to the slender dog. The remains of several other wild and domestic animals (including horses, pigs and bears) were also disposed of in the wells along with the dogs44,47, raising questions around the way that these deposits should be interpreted. Various theories are being discussed for this phenomenon such as “only” waste disposal, victims of war or epidemic, political crisis, the contamination of water or ritual deposits44,48. The short period of time during which the animals were deposited and the relatively large number of dogs and other animals disposed in the wells suggest that an epizootic disease was a potential cause. Shotgun DNA sequence analysis of dog viral and DNA based pathogens was, however, negative. The general low quality and quantity of endogenous nuclear DNA of the samples might explain the negative screening results for viral and bacterial pathogens. Or, pathogens attacking primarily soft tissues remained unperceived. Therefore, we cannot completely rule out infectious disease or other pathologies as cause of dog death and disposal.
The capacity in dogs to digest starches has been demonstrated since the transition to the Neolithic6,83 suggesting that dogs followed similar diets as humans with substantial proportions of vegetal food components besides animal proteins. It is known that the Romans were feeding their dogs differently according to a number of factors, including function and age, but most likely the majority of Roman dogs were relying primarily on table scraps31. Stable carbon and nitrogen isotope ratios obtained from dogs of Augusta Raurica of mixed size, age and haplotype composition (Fig. 2, Table S1) show that they fed on similar, only slightly varying diets. The data are consistent with a mainly mixed omnivorous diet including variable meat and vegetable proportions, similar to other Roman sites across Europe23,67,68,69,70,71,72,84 and the Celtic dogs from Basel Gasfabrik (150– 80 BCE), a site located close by Augusta Raurica (Fig. 2)56. This matches numerous archaeological, epigraphic and literary evidence for dog food in the Roman Empire, that show a large proportion of vegetal components, e.g. cereals, bread (made of barley, wheat or spelt), and a minor component of meat, bones, milk and whey (Harvey31). The substantial difference in δ13C and δ15N values between herbivores from Augusta Raurica, i.e. horses73 suggests that the proportion of animal proteins in the dogs’ diet should not be underestimated. The low isotopic variability may support the hypothesis that size, age or function, may have only moderately impacted stable isotope compositions at Augusta Raurica. However, stable isotope compositions as found in the dogs from Augusta Raurica do not allow for reliable assignments of functions to individual dogs. The isotopic signals show no significant differences between the small dogs with particular morphologies (brachymely and brachycephaly) and the others (Fig. 2). This could indicate that these particular dogs did not receive any special food. Two dogs (HAU9, HAU18) with earlier dates (ca. 200 CE and 70–100 CE, respectively) and different archaeological contexts displayed more negative δ13C values compared to the dogs disposed of in wells. Sample sizes are very small, one tentative explanation could be that the two dogs fed on specific (plant) components low in δ13C that eventually became scarce by the 2nd half of the third century CE when the town of Augusta Raurica supposedly was in decline, another explanation could be diachronically varying climatic, e.g. more humid, conditions. The only distinct outlier, HAU5, fed on an exceptionally low-protein and more plant based diet (Fig. 2). One possible explanation would be a diet including C4 plants such as millet, reported to be grown at Augusta Raurica39. The combination of a low meat and high plant intake could be seen as an indication for unbalanced nutrition in the individual HAU5, but the reasons behind this remain unclear since there are no unusual findings in archaeology, morphology or aDNA.
Mitochondrial DNA variation in a 97 bp fragment of the control region in 25 individuals points to a genetically diverse dog population at Augusta Raurica and Vindonissa. Eight haplotypes representing all four widely recognized major haplogroups A-D34 were detected but no association between haplotype and size/morphology was apparent. The concomitant presence of all the four major haplogroups in this dog population is remarkable as modern dog breeds generally do not harbor all four haplogroups at once36,75,76,85. The observed haplotype diversity corresponds to the increasing morphotype diversity in the Roman world23 and provides insights into a diversifying dog population in situ, with a sharp increase in effective population size starting around 2,500 years ago through the Roman era into modern times36,78. Therefore, we may assume Roman people likely propagated and dispersed dog lineages across the Empire into the Colonies (and back) as reflected in dog lineage diversity in Augusta Raurica and Vindonissa.
Some of the lineages from Iberia, Bulgaria and from Augusta Raurica were unique to different Roman sites and either went extinct, were subject to genetic drift or were not yet detected due to relatively small datasets especially for archaeological dogs. Because of the lack of genetic data from Roman Italy and also because of the rarity of mtDNA data from archaeological dogs elsewhere and from other time periods to compare with, interpretations are limited. However, some scenarios might be considered taking into account the known trade/contact routes in Roman Europe: the dogs from Augusta Raurica share many haplotypes with dogs from Roman Iberia/Morocco (haplogroups A and D), but not with Roman Bulgaria suggesting a connection between dog lineages from both regions. From the Median-Joining Network analysis of pre-Roman dog lineages (Fig. S4, Table S2) other links are conceivable such as A haplotype cluster “RS-a2” or cluster “RS-a4, might have reached Augusta Raurica via Italy. Interestingly, Chalcolithic to Bronze Age Italy shows frequencies of haplogroup A (50%) and C (42%) comparable to Roman Switzerland86. Pre-Roman and Roman Bulgaria is a known centre of haplogroup B5 which is also reflected in the haplogroup frequency distribution in Europe (Fig. 4). Comparison to some pre-Roman data from Southeast Europe in particular haplogroup B and D haplotypes are found at Augusta Raurica, hints to a connection also to that region. (Fig. S4, Table S2). The situation for haplogroup C lineages suggests potential continuity for some C haplotypes in Switzerland from the Neolithic (ca. 5500 – 2200 BCE) into Roman times (ca. 1st century BCE–4th century CE). Future research is needed to test these hypotheses.
We assembled four new mitogenomes from dogs of Augusta Raurica with three diverse A haplotypes and one C haplotype. This allows the subdivision of individuals in haplogroup A into sub-haplogroups A1b2 and A1b3 and in haplogroup C into sub-haplogroup C2 (assignment after Duleba et al.36). The four Roman mitogenomes differed from dog mitogenomes from Palaeolithic Switzerland (Fig. 5)78,80. The past history of dogs in Switzerland is marked by continuity and replacements of lineages but remains largely unknown/unresolved due to a lack of studies. The dogs of Augusta Raurica and Vindonissa fill, therefore, a small part of this knowledge gap.
The mtDNA diversity documented here in Roman dogs of northern Switzerland is only known from modern dogs. The high diversity of haplogroups and haplotypes concentrated at Augusta Raurica suggests that diverse maternal lineages were kept at the same time and place simultaneously. This is surprising as much of the diversity of today’s dogs is generally seen as the result of breeding activities over the last 200 years and is not expected to originate from earlier, e.g. Roman times, ca. 2000 years ago. Some dogs likely accompanied their owners to Augusta Raurica and Vindonissa (e.g. traders, legionary families, i.e. people known to come in from any part of the Roman Empire), underpinning the region as a trade and transport hub north of the Alps under the rule of Rome.
Conclusion
This study has provided the first archaeozoological and biomolecular insights into a hitherto unstudied dog population from Augusta Raurica and Vindonissa, Germania Superior, emphasizing the utility of wells and burials as sources for the reconstruction of dog diversity in Roman times in general. Based on our data no function was assigned to the dogs. However, it is a reasonable assumption that dogs from wells were a mixture of free-roaming individuals and animals cared for by people. The dogs were highly variable in size and include dogs with short-limbs, slenderness and one extreme case of brachycephaly.
Overall, the dogs of Augusta Raurica were similarly omnivorous with a tendency towards animal proteins. Whatever the living conditions were, the dogs showed no sign of undernourishment (except maybe for outlier HAU5 with a lower animal protein ratio in its diet) in line with general observations from other Roman regions30.
Mitochondrial D-loop and mitogenome diversity suggests that diverse maternal lines were kept in Augusta Raurica and Vindonissa and probably had different geographical origins and haplogroup frequencies in the Roman period, showing a specific pattern compared to other European regions. Some of the dogs shared the same maternal lineage and, since they were living almost contemporaneously and all very likely in Augusta Raurica, they might have been closely related, raising questions of kinship and potential local breeding without excluding the possibility of mixing by chance.
Data availability
Ancient partial mitochondrial DNA and whole mitochondrial genome dog sequences generated in the current study are publicly available from GenBank accession numbers OP082250-OP082275 and from the European Nucleotide Archive (ENA) with project number PRJEB55253, respectively. All raw data are available upon request.
References
Frantz, L. A. et al. Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science 352, 1228–1231 (2016).
Vonholdt, B. M. & Driscoll, C. A. in The Domestic Dog: Its Evolution, Behavior and Interactions with People (ed James Serpell) 22–41 (Cambridge University Press, 2016).
Botigué, L. R. et al. Ancient European dog genomes reveal continuity since the Early Neolithic. Nat. Commun. 8, 16082 (2017).
Ollivier, M. et al. Dogs accompanied humans during the Neolithic expansion into Europe. Biol. Lett. 14, 52 (2018).
Yankova, I. et al. Evidence for early European neolithic dog dispersal: New data on Southeastern European subfossil dogs from the prehistoric and antiquity ages. Genes 10, 757 (2019).
Bergström, A. et al. Origins and genetic legacy of prehistoric dogs. Science 370, 557–564 (2020).
Feuerborn, T. R. et al. Modern Siberian dog ancestry was shaped by several thousand years of Eurasian-wide trade and human dispersal. Proc. Natl. Acad. Sci. 118, 52 (2021).
Perri, A. R. et al. Dog domestication and the dual dispersal of people and dogs into the Americas. Proc. Natl. Acad. Sci. U S A 118, 69 (2021).
Losey, R. J. Domestication is not an ancient moment of selection for prosociality: Insights from dogs and modern humans. J. Soc. Archaeol. 22, 131–148 (2022).
Horard-Herbin, M.-P., Tresset, A. & Vigne, J.-D. Domestication and uses of the dog in western Europe from the Paleolithic to the Iron Age. Anim. Front. 4, 23–31 (2014).
Bisig, V. Osteometrische Untersuchungen an zwei fast vollständig erhaltenen Hundeskeletten der Römischen Colonia Augusta Raurica (Universität Zürich, 1971).
Lepetz, S. L’animal dans la société gallo-romaine de la France du nord. Revue Archéologique de Picardie 5, 96 (1996).
Peters, J. D. Hund in der Antike aus archäozoologischer Sicht. Anthropozoologica 25(26), 511–523 (1997).
Peters, J. Römische Tierhaltung und Tierzucht. Passauer Universitätsschriften zur Archäologie 5. Rahden/Westfalen (1998).
De Grossi, M. J. & Tagliacozzo, A. Dog remains in Italy from the Neolithic to the Roman period. Anthropozoologica 25, 429–440 (1997).
Colominas, L. Morphometric variability of roman dogs in hispania tarraconensis: The case study of the vila de madrid necropolis. Int. J. Osteoarchaeol. 26, 897–905 (2016).
Bennett, D. & Timm, R. M. The dogs of Roman Vindolanda, Part II: Time-stratigraphic occurrence, ethnographic comparisons, and biotype reconstruction. Archaeofauna 25, 107–126 (2016).
De Grossi M. J. & Tagliacozzo, A. in Dogs Through Time: An Archaeological Perspective (ed S. J. Crockford) 141–180 (BAR International Series 889, 2000).
Blaschikoff, L. et al. A multidisciplinary study of Iberian Chalcolithic dogs. J. Archaeol. Sci. Rep. 42, 1–17 (2022).
Pires, A. E. et al. Roman dogs from the Iberian Peninsula and the Maghreb–A glimpse into their morphology and genetics. Quat. Int. 471, 132–146 (2018).
Baxter, I. L. in Dogs and People in Social, Working, Economic or Symbolic Interaction (eds M. S. Lynn & A. M. Elizabeth) 12–23 (Oxbow Books, Proceedings of the 9th ICAZ Conference: Durham, 2006).
Bennett, D. & Timm, R. M. The dogs of Roman Vindolanda, Part III: Quantifying juvenilization and pleiotropic effects of miniaturization. Archaeofauna 27, 57–82 (2018).
Martínez Sánchez, R. M. et al. Who let the dogs in? Lap dogs, canid sacrifices and funerary practices in the Roman cemetery of Llanos del Pretorio (Cordoba, Spain). Archaeol. Anthropol. Sci. 12, 87 (2020).
Zedda, M. et al. Ancient Pompeian dogs-Morphological and morphometric evidence for different canine populations. Anatomia Histol. Embryol. 35, 319–324 (2006).
Bartosiewicz, L. Shuffling Nags, Lame Ducks. The Archaeology of Animal Disease (Oxbow, 2013).
Toynbee, J. M. C. Animals in Roman Life and Art (Thames and Hudson, 1973).
Osypinska, M., Skibniewski, M. & Osypinski, P. Ancient Pets. The health, diet and diversity of cats, dogs and monkeys from the Red Sea port of Berenice (Egypt) in the 1st-2nd centuries AD. World Archaeol. 6, 1–15 (2021).
van Haasteren, M. & Groot, M. The biography of wells: A functional and ritual life history. J. Archaeol. Low Countries 4, 25–51 (2013).
Valenzuela-Lamas, S. & Gardeisen, A. in Onze puits gallo-romains de Lattara (Ier s. av. n. è.-IIe s. de n. è). Fouilles programmées 1986–2000 Vol. Lattara 18 (eds R. Buxó & G. Piqués), 235–270 (2005).
MacKinnon, M. ‘Sick as a dog’: Zooarchaeological evidence for pet dog health and welfare in the Roman world. World Archaeol. 42, 290–309 (2010).
Harvey, S. M. Bread and bones: Feeding roman dogs. Classical World 114, 419–447 (2021).
Layman, C. A. et al. Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol. Rev. 87, 545–562 (2012).
Makarewicz, C. A. & Sealy, J. Dietary reconstruction, mobility, and the analysis of ancient skeletal tissues: Expanding the prospects of stable isotope research in archaeology. J. Archaeol. Sci. 56, 146–158 (2015).
Savolainen, P., Zhang, Y.-P., Luo, J., Lundeberg, J. & Leitner, T. Genetic evidence for an East Asian origin of domestic dogs. Science 298, 1610 (2002).
Pang, J.-F. et al. mtDNA data indicate a single origin for dogs South of Yangtze River, Less Than 16,300 Years ago, from numerous wolves. Mol. Biol. Evol. 26, 2849–2864 (2009).
Duleba, A., Skonieczna, K., Bogdanowicz, W., Malyarchuk, B. & Grzybowski, T. Complete mitochondrial genome database and standardized classification system for Canis lupus familiaris. Forensic Sci. Int. Genet. 19, 123–129 (2015).
Deguilloux, M. F., Moquel, J., Pemonge, M. H. & Colombeau, G. Ancient DNA supports lineage replacement in European dog gene pool: insight into Neolithic southeast France. J. Archaeol. Sci. 36, 513–519 (2009).
Pires, A. E. et al. The curious case of the Mesolithic Iberian dogs: An archaeogenetic study. J. Archaeol. Sci. 105, 116–129 (2019).
Deschler-Erb, S., Stopp, B. & Vandorpe, P. Big Data-65 Jahre archäobiologische Forschungen in Augusta Raurica. Jahresberichte aus Augst und Kaiseraugst 42, 293–368 (2021).
Deschler-Erb, S. & Stopp, B. in Am Südtor von Vindonissa. Die Steinbauten der Grabung Windisch-Spillmannwiese 2003–2006 (V.003.1) im Süden des Legionslagers. Vol. 22 (eds J. Trumm & M. Flück) 463–477 (Veröffentlichungen der Gesellschaft Pro Vindonissa, 2013).
L. Flutsch, U. Niffeler, F. Rossi, Eds., SPM V Römische Zeit (Schweizerische Gesellschaft für Ur- und Frühgeschichte, Basel, 2002), Vol SPM V, p 432.
Schmitz, D. & Sieler, M. Überall zu Hause und doch fremd. Römer unterwegs. Kataloge des Römermuseums im Archäologischen Park Xanten, 5. (Petersberg 2013).
Lo Cascio, E. & Tacoma, L. E. in Proceedings of the Twelfth Workshop of the International Network Impact of Empire (Rome, June 17–19, 2015). Leiden/Boston (2017).
Schmid, D., Peter, M. & Deschler-Erb, S. in L'Empire Romain en Mutation - Répercussions sur les villes dans la deuxième moitié du 3e siècle. Colloque International / Das römische Reich im Umbruch - Auswirkungen auf die Städte in der zweiten Hälfte des 3. Jahrhunderts. Internationales Kolloquium, Bern/August (Schweiz), 3.-5.12.2009. Archéologie et Histoire Romaine Vol. 20 (eds Regula Schatzmann & Stefanie Martin-Kilcher) 125–132 (Éditions Monique Mergoil 2011).
Berger, L. Führer durch Augusta Raurica (Schwabe, 2012).
Schatzmann, R. Die Spätzeit der Oberstadt von Augusta Raurica. Untersuchungen zur Stadtentwicklung im 3. Jahrhundert. (Augusta Raurica, 2013).
Mráz, M. Bärenhatzen und Hundeplagen-Tierknochen aus dem Sodbrunnen MR12 in der Region 17C der Unterstadt von Augusta Raurica. Jahresberichte aus Augst und Kaiseraugst 39, 143–182 (2019).
Lepetz, S. & Bourgois, A. Were sanctuary wells in Roman Gaul intentionally contaminated using animal carcasses (3rd–4th c. AD)?. Gallia 75, 173–188 (2018).
Müller, U., Grolimund, L. & Saner, C. Ausgrabungen in Kaiseraugst im Jahre 2003. Jahresberichte aus Augst und Kaiseraugst 25, 179–196 (2004).
Pfäffli, B. & Rychener, J. in Jahresberichte aus Augst und Kaiseraugst Vol. 26 (ed Römerstadt Augusta Raurica) 71–86 (Römermuseum Augst, 2005).
Breuer, G. Die Tierknochenfunde aus zwei Latrinengruben des 1. Jahrhunderts in Augst (Grabung 1991.65) unter besonderer Berücksichtigung der Hundeskelette. Jahresberichte aus Augst und Kaiseraugst 13, 177–196 (1992).
Trumm, J., Brogli, R. F., Frei-Stolba, R. & Kahlau, T. Bestattet und begraben: ein neu entdecktes römisches Gräberfeld in Vindonissa. Archäologie der Schweiz 36, 26–33 (2013).
Clark, K. M. The later prehistoric and protohistoric dog: The emergence of canine diversity. Archaeozoologia 7, 9–32 (1995).
Koudelka, F. Das Verhältniss der Ossa longa zur Skelethöhe bei den Säugetieren. Verhandlungen des naturforschenden Vereins Brünn 24, 127–153 (1885).
Longin, R. New method of collagen extraction for radiocarbon dating. Nature 230, 241–242 (1971).
Knipper, C. et al. What is on the menu in a Celtic town? Iron Age diet reconstructed at Basel-Gasfabrik, Switzerland. Archaeol. Anthropol. Sci. 9, 1307–1326 (2017).
Elsner, J., Schibler, J., Hofreiter, M. & Schlumbaum, A. Burial condition is the most important factor for mtDNA PCR amplification success in Palaeolithic equid remains from the Alpine foreland. Archaeol. Anthropol. Sci. 7, 505–515 (2015).
Schibler, J., Elsner, J. & Schlumbaum, A. Incorporation of aurochs into a cattle herd in Neolithic Europe: Single event or breeding?. Sci. Rep. 4, 1–6 (2014).
Gilbert, M. T. P., Bandelt, H.-J., Hofreiter, M. & Barnes, I. Assessing ancient DNA studies. Trends Ecol. Evol 20, 541–544 (2005).
Excoffier, L. & Lischer, H. E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 10, 564–567 (2010).
Yang, D. Y. & Watt, K. Contamination controls when preparing archaeological remains for ancient DNA analysis. J. Archaeol. Sci. 32, 331–336 (2005).
Pilli, E. et al. Monitoring DNA contamination in handled vs. directly excavated ancient human skeletal remains. PloS One 8, e52524 (2013).
Knapp, M., Clarke, A. C., Horsburgh, K. A. & Matisoo-Smith, E. A. Setting the stage–Building and working in an ancient DNA laboratory. Annal. Anatom. Anatomischer Anzeiger 194, 3–6 (2012).
Krause-Kyora, B. et al. Ancient DNA study reveals HLA susceptibility locus for leprosy in medieval Europeans. Nat. Commun. 9, 1569 (2018).
Ambrose, S. H. Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17, 431–451 (1990).
van Klinken, G. J. Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26, 687–695 (1999).
Prowse, T., Schwarcz, H. P., Saunders, S., Macchiarelli, R. & Bondioli, L. Isotopic paleodiet studies of skeletons from the Imperial Roman-age cemetery of Isola Sacra, Rome, Italy. J. Archaeol. Sci. 31, 259–272 (2004).
Jørkov, M. L. S., Jørgensen, L. & Lynnerup, N. Uniform diet in a diverse society. Revealing new dietary evidence of the Danish Roman Iron Age based on stable isotope analysis. Am. J. Phys. Anthropol. 143, 523–533 (2010).
Redfern, R. C., Hamlin, C. & Athfield, N. B. Temporal changes in diet: A stable isotope analysis of late Iron Age and Roman Dorset, Britain. J. Archaeol. Sci. 37, 1149–1160 (2010).
Pate, F. D., Henneberg, R. J. & Henneberg, M. Stable carbon and nitrogen isotope evidence for dietary variability at ancient Pompeii, Italy. Mediterranean Archaeol. Arch. 16, 127–133 (2016).
Bourbou, C., Arenz, G., Dasen, V. & Lösch, S. Babes, bones, and isotopes: A stable isotope investigation on nonadults from Aventicum, Roman Switzerland (first–third century CE). Int. J. Osteoarchaeol. 29, 974–985 (2019).
Varano, S. et al. The edge of the Empire: Diet characterization of medieval Rome through stable isotope analysis. Archaeol. Anthropol. Sci. 12, 1–16 (2020).
Gerling, C. Neither fish nor fowl Isotopic evidence of a plant-based diet in (captive?) brown bears from Roman Augusta Raurica, Switzerland. Anthropozoologica 58, 59–72 (2023).
Bocherens, H. & Drucker, D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: Case studies from recent and ancient terrestrial ecosystems. Int. J. Osteoarchaeol. 13, 46–53 (2003).
Pires, A. E. et al. Mitochondrial DNA sequence variation in Portuguese native dog breeds: Diversity and phylogenetic affinities. J. Hered. 97, 318–330 (2006).
Ghanatsaman, A. Z. et al. Mitochondrial DNA sequence variation in Iranian native dogs. Mitochondrial. DNA Part A 29, 394–402 (2018).
Susat, J. et al. Yersinia pestis strains from Latvia show depletion of the pla virulence gene at the end of the second plague pandemic. Sci. Rep. 10, 14628 (2020).
Thalmann, O. et al. Complete mitochondrial genomes of ancient canids suggest a European Origin of domestic dogs. Science 342, 871 (2013).
Ameen, C. et al. Specialized sledge dogs accompanied Inuit dispersal across the North American Arctic. Proc. Biol. Sci. 286, 20191929 (2019).
Loog, L. et al. Ancient DNA suggests modern wolves trace their origin to a Late Pleistocene expansion from Beringia. Mol. Ecol. 2, 1–15 (2019).
Bennett, D., Campbell, G. & Timm, R. M. The dogs of Roman Vindolanda, Part I: Morphometric techniques useful in differentiating domestic and wild canids. Archaeofauna 25, 79–105 (2016).
Colominas, L. Animals i ideologia en l’àmbit funerari: estudi arqueozoològic de la Necròpolis de la plaça de la vila de Madrid. Quarhis Quaderns d’Arqueologia i Història de la Ciutat de Barcelona 2, 82–101 (2007).
Ollivier, M. et al. Amy2B copy number variation reveals starch diet adaptations in ancient European dogs. R. Soc. Open Sci. 3, 160449 (2016).
Craig, O. E. et al. Stable isotopic evidence for diet at the Imperial Roman coastal site of Velia (1st and 2nd Centuries AD) in Southern Italy. Am. J. Phys. Anthropol. 139, 572–583 (2009).
Marinov, M., Teofanova, D., Gadjev, D., Radoslavov, G. & Hristov, P. Mitochondrial diversity of Bulgarian native dogs suggests dual phylogenetic origin. PeerJ 6, e5060 (2018).
K. Koupadi et al., Population Dynamics in Italian Canids between the Late Pleistocene and Bronze Age. Genes 11, 1409 (2020).
Acknowledgements
The present work was funded by the Swiss National Science Foundation (SNSF; HumAnimAl Project, Grant number 178834). We thank Sandra Ammann from Augusta Raurica for providing us with dog samples from Augusta Raurica. We also thank Susanne Schenker from Augusta Raurica for providing us with the picture of dog figurine. We further thank Regine Fellmann from Vindonissa who provided us with the two dog samples from Vindonissa. Moritz Lehmann kindly provided access to mass spectrometry facilities and Thomas Kuhn kindly performed stable isotope analysis. We express our gratitude to Ben Jennings for English language editing.
Author information
Authors and Affiliations
Contributions
M.S.-M. and S.D.-E. conceived the idea. M.S.-M., C.G., A.S., S.D.-E and B.K.-K. designed the study. J.G., J.S., C.G. and M.S.-M. performed laboratory work and contributed to data analysis. J.G. and J.S. led the writing of the manuscript. All authors provided critical input on drafts and agreed with the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Granado, J., Susat, J., Gerling, C. et al. A melting pot of Roman dogs north of the Alps with high phenotypic and genetic diversity and similar diets. Sci Rep 13, 17389 (2023). https://doi.org/10.1038/s41598-023-44060-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-023-44060-3
