
Abstract
Tropical storms and hurricanes pose a significant impediment to southbound Nearctic-Neotropical migratory songbirds. The effects of severe weather on transient songbirds may be significant but have been difficult to assess. We used two meteorological indices of inclement weather, Accumulated Cyclone Energy (ACE) values from the western Atlantic Basin and the Southern Oscillation Index (SOI), to investigate whether the effects of inclement weather during southward migration are manifest in the population morphometrics of a migratory songbird at a North American breeding site in Delaware and Pennsylvania, USA. Over a 23-year period (2000–2023), both ACE and SOI were predictors of wing chord and body condition of returning Veeries (Catharus fuscescens). Elevated September ACE (more severe en route weather in September) was a predictor of shorter wing chord and lower body condition indices in returning adult females. The effects on SY birds differed. Years with higher September ACE were a predictor of higher body condition indices in returning SY females and elevated September and October ACE values were a predictor of longer wing chord in returning SY birds. Positive SOI values (La Niña type years) were a predictor of shorter wing chord and higher body condition indices in adult Veeries. In general, en route severe weather was a stronger predictor for females than males. Sex- and age-specific differences are likely due to differing spatiotemporal migratory patterns. Our results show that inclement tropical weather is a source of morphological variation in a Nearctic-breeding migratory songbird and are consistent with the premise that severe tropical weather constrains Nearctic-breeding migratory songbird populations. These data contribute to the mounting evidence that the projected increase in the severity of en route tropical storms and hurricanes due to climate change is a significant emerging threat to Nearctic-Neotropical migratory songbirds.
Similar content being viewed by others
Introduction
Over the past several decades, the decline of Nearctic-Neotropical migratory songbird populations has been a primary focus of ornithological study1,2,3. Understanding the breadth of seasonal constraints on populations is of paramount importance toward an understanding of population regulation. Barriers along migratory routes, whereby birds must circumnavigate obstacles, may have significant effects on avian fitness4,5. Severe tropical weather, including tropical storms and hurricanes, pose significant and re-occurring impediments to southward passage migratory birds, particularly the species that traverse large expanses of open water including the Atlantic Ocean, the Gulf of Mexico and the Caribbean Sea6,7,8. Since 1950, > 80% of all hurricanes and > 90% of major hurricanes (Saffir-Simpson Hurricane Wind Scale Categories ≥ 3) in the Atlantic Basin have developed in September and October during peak songbird migration9,10. Because of their small size, songbirds may be especially susceptible to mortality associated with severe weather. The expanse of hurricanes may be massive, and unlike permanent geographic barriers, the spatiotemporal properties of hurricanes are unpredictable and chaotic, likely to exacerbate the physical burden on passage birds8,11. The radius of hurricanes, typically measured by the outer closed isobar, can reach > 1000 km creating a barrier to migration nearly the size of the Gulf of Mexico12. Although migratory birds can maximize their survival probability by departing over open water under favorable conditions13 as centuries of human history have shown, tropical storms and hurricanes may be difficult to detect until shortly before they are encountered – there is no reason to think this could not be the case for songbirds. Elsewhere, mass mortality due to severe weather events encountered post-departure, has been well documented14.
Unfortunately, it has been nearly impossible to quantify the effects of severe storms on transient migratory birds in part because some effects may be indirect. For example, indirect mortality may occur when birds encountering severe storms are blown off-course and lost at sea, are forced to make landfall in inhospitable regions, succumb to exhaustion while circumnavigating large storms over open water or succumb to exposure during strong winds and torrential downpours when faced with inadequate cover8,15,16. Mortality of Nearctic-Neotropical migrant songbirds is now thought to be greatest during migratory periods, accounting for 44–85% of annual mortality17,18,19,20,21. Recent studies suggest that mortality related to weather events and their global climate cycles, including tropical storms and hurricanes, may be an underappreciated source of this mortality potentially limiting songbird populations6,16,17. There are few direct observations of migratory songbird mortality events during severe tropical storms; however, indirect evidence that mass mortality occurs is mounting, and there is at least one historic eye-witness account of birds “falling from the sky” en masse during an autumn hurricane at sea22. Butler6 found that some North American bird populations fluctuate in relation to the severity of tropical weather during southbound migration providing evidence that mass mortality events likely occur. In the Gulf of Mexico, juvenile tiger sharks (Galeocerdo cuvier) congregate annually apparently to feed on significant numbers of birds killed in relation to inclement weather16. Thus, this avian mortality is so predictable that it provides a pulsed resource upon which these sharks may have become somewhat dependent16. Indirect effects of related weather events in the western Atlantic Basin have also been documented; for example, following strong westerly winds over the Caribbean Sea in relation to inclement weather, a sharp increase in the abundance of southbound Swainson’s Thrush (Catharus ustulatus) was recorded in northern Colombia well east of their normal migratory route23. Whimbrels (Numenius phaeopus) that continued migration after encountering storms in flight, terminated flights prematurely presumably because of depleted energy stores8. It is widely believed that hurricanes are becoming more severe due to recent climate change, especially with regard to rapid intensification, sustained winds, prolonged torrential downpours, and decreasing translation speed24,25,26,27,28. Therefore, en route inclement weather, including more severe and more frequent tropical depressions, storms, and hurricanes, may pose a rapidly emerging threat to Nearctic-Neotropical migratory songbirds that cross the Gulf of Mexico, Caribbean Sea, and portions of the Atlantic Ocean.
Over the last few decades, multiple studies have reported a shift in the morphometrics of many migratory songbird species including changes in both wing chord and mass29,30,31. Body size is generally assumed to be heritable32,33. Therefore, variability in morphometrics over time may have consequences for individual fitness, population biology, and species conservation32,33. Long-term changes in the body size of songbirds have become a topic of particular interest given the potential effects of a rapidly changing climate34,35. For example, Bergmann’s Rule predicts that body size in endotherms will decrease in warm climates because a small surface-to-body ratio is more efficient in regulating heat transfer; therefore, it follows that a warming climate may result in decreased body size as found in some studies30,34,36,37. However, long-term changes in body size have been equivocal as some studies have reported changes in morphology opposite that expected if selection on body size was simply the result of a warming environment31,35,37. Thus, empirical support for a direct causal effect of a warming climate on body size in wild birds has remained elusive38. This ambiguity may in part be due to the inability to parse the interannual effect of fluctuating temperature on body size from simply associating a trend in morphological change over a period of warming37. In addition to warming ambient temperatures, alternate proximate causes have been hypothesized to explain long-term changes such as episodic environmental conditions (e.g., intensifying drought, heat waves), variable food availability and associated changes in primary productivity, the loss or degradation of breeding and wintering habitat, or a combination of these factors33,35,39,40. Effects other than those associated directly with increasing temperatures, environmental conditions, or habitat also seem plausible. Anderson et al.41 and Lank et al.42 hypothesized that changes in morphology, including smaller body and wing size, has favored increased aerial maneuverability in shorebirds – possibly the result of increased selective pressure from aerial predators from growing falcon populations during the post-DDT era. Identifying the drivers of morphological change, particularly those that may be caused or exacerbated by climate change, is considered a research priority43.
Many studies that have used long-term data to quantify body size changes in bird species have relied on data from migratory stopover sites30,31,41. This may be problematic for multiple reasons. Migratory habitat use may differ for some species depending on the age or sex of individuals which is often impossible or difficult to determine at stopover sites41. Moreover, sexual size dimorphism is prevalent in most species, potentially compromising the ability to detect directional changes in body size when males and females cannot be parsed. Perhaps most importantly, if Bergman’s Rule is the foundation of hypotheses used to explain decreasing body size, it could be problematic to collect data from a site where breeding individuals may originate from a broad latitudinal range. For these reasons, we believe that data collected from long-term studies at breeding sites is more appropriate to investigate morphological changes. Collection of long-term data from a breeding population will control for the effect of latitude and will accurately incorporate age and sex effects while in many cases enabling the ability to parse the effect of interannual environmental fluctuation40,44,45.
Given the potential effect of inclement weather on migrating songbirds, and the ambiguity in data associating phenotypic shifts with warming ambient temperatures, we investigated whether the effects of severe en route tropical weather – that ultimately may be exacerbated by global warming–might be manifest in the morphometrics of a migratory songbird population. Our study subject is the Veery (Catharus fuscescens), a single-brooded (typically one successful brood per year) forest-dependent thrush (Passeriformes: Turdidae) that migrates annually between temperate and boreal forests in northern North America and mature stunted nutrient-poor forests in the Cerrado-Amazonia transition zone of the southern Amazon Basin, a roundtrip distance of up to 10,000 km46. Research using light level geolocators, GPS loggers, and at stopover sites, have revealed that individuals use a variety of migratory routes, either westward along the Caribbean coast of eastern Central America or eastward through the Greater and sometimes Lesser Antilles, or otherwise directly across both the Gulf of Mexico and Caribbean Sea, or combinations thereof46,47,48,49,50,51,52. During southward migration, Veeries from western North America converge with Veeries from eastern North America (4,000 km separation of breeding regions) at stopover sites along the north shores of the Gulf Coast. The Yucatán Peninsula and Darien Gap regions of Central America, and the Greater Antilles (mostly Cuba), are primary stopover locations, as are parts of northern South America especially Sierra Nevada de Santa Marta in northern Colombia47,48,49,50,51,52,53. Veeries undertake three long-distance migrations annually including an intratropical migration in South America between their south and northbound migrations54. Specifically, we investigated whether the effect of inclement tropical weather (e.g., tropical depressions, tropical storms, hurricanes) during southward migration are apparent in Veery morphometrics including returning body condition which may have lateral effects on recruitment and reproductive success. Because incidents of severe tropical weather may accompany a period of climate warming, such an effect might be one causal explanation of body size changes in Nearctic-Neotropical migrants that have shown relationships with periods of warming. Considering the random interannual fluctuations in the severity of hurricane seasons, such a relationship might explain some of the ambiguity observed in the morphometric data. Secondarily, establishing a link between interannual phenotypic change and severe tropical weather, we can assume that severe weather events actively result in selection pressure likely via direct and indirect mortality, providing further justification for concern regarding the projected future effect of increasing en route severe tropical weather on Nearctic-Neotropical migratory bird populations. An additional impetus for our investigation was the discovery that the Veery breeding season is largely constrained by the propensity for severe tropical weather on southward migration with early cessation of the breeding season coinciding with years of greater subsequent storm activity presumably resulting in an earlier departure date from breeding grounds or better physical condition of individuals prior to the peak of hurricane season55. Importantly, the observed constraint of global climate cycles on breeding performance suggest that severe tropical weather likely plays a role in exerting selective pressure on passage birds which we hypothesized may be reflected in Veery morphometrics including a decrease or increase in body size (i.e., wing chord) or compromised body condition. Thus, our objective is somewhat exploratory in that we are attempting to detect a signal in our dataset and assessing the magnitude of that signal: our goal is not to identify all drivers of Veery morphometrics but rather to determine whether the effects of severe tropical weather have been an influential source of phenotypic variability in our study population.
Methods
For twenty-five years (1998–2023), a resident population of Veeries has been studied from May through July at White Clay Creek State Park, New Castle County, Delaware, and the contiguous White Clay Creek Preserve, Chester County, Pennsylvania, USA (centered at 39.739569°, −75.763802°). Our study area consists of broadleaf forests of mixed seral stages typical of the North American Mid-Atlantic Appalachian Piedmont physiographic province56. Elevation of forests range from 50 to 150 masl. From May through mid-July 1998 to 1999, territorial male Veeries were target-netted (i.e., locating a territorial adult and erecting nets to catch that individual), while females were captured only as by-catch. Nonetheless, all captured Veeries were color-marked but few morphometric data were recorded. From 2000 on, un-flattened wing chord and mass were measured at each capture and represent the only morphometrics recorded consistently throughout the remaining 23-year period. Target netting males, and occasionally females at nest sites, was the dominant type of field capture method until 2008 when constant-effort netting (large array of open nets at various locations to catch all bird species) was commenced. Therefore, females were rarely captured, weighed, and measured, prior to 2008, whereas nearly all active males at our study site were captured and processed each year prior. This resulted in a greater number of males in the 23-year dataset than females. Once on the breeding grounds after completing migration, Veery do not show fat reserves; therefore, all measurements of mass were of fat-free mass.
Statistical analysis
We separated birds by sex and age using Pyle57. We aged and grouped birds as second-year (SY) and after second year (ASY). Birds categorized as SY birds were hatch-year (HY) birds the preceding southward migration–hereafter, our primary period of interest. As is common practice, below we sometimes refer to ASY birds as “adult” to distinguish between SY and ASY cohorts.
We used Accumulated Cyclone Energy (ACE) as a primary predictor of returning wing chord and body condition. ACE is the most advanced seasonal metric used by meteorologists to convey the strength and duration of named tropical storms and cyclones in each Atlantic hurricane season (1 June to 30 November)10. ACE is calculated for each storm separately. The value of ACE for each storm in a season is combined to convey the overall seasonal severity of tropical weather. Importantly, despite the fact ACE is only calculated for named storms (i.e., tropical storms and hurricanes), total ACE values reflect the relative degree of tropical storm activity within a season, including inclement weather events caused by un-named tropical disturbances; therefore, higher ACE values are reflective of a greater number of tropical disturbances and inclement weather events relative to periods of lower ACE (P. Klotzbach, pers. comm., Colorado State University). Thus, although ACE is calculated from individual named storms, seasonal ACE values act as the best index to the frequency and severity of tropical disturbances in the respective region during our period of interest. Yearly ACE values are independent as there is no interannual relationship among hurricane seasons.
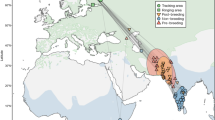
For each hurricane season, we used ACE values from a targeted area of the Atlantic Basin that directly affected regions known to be traversed by southward migrating Veeries during late August, September, October, and early November – the peak of Atlantic hurricane season. These dates are consistent with the timing of migration from previously tracked individuals from our study population47,50,52. Our target area included the region within an area from 100° West to 58° West, and 32° North to 7° North (Fig. 1). This area encompasses part of the western Atlantic Ocean, the entire Gulf of Mexico, the Caribbean Sea, east coastal Central America, and both the Lesser and Greater Antilles – the regions traversed by southward migrating Veeries. It also includes the western third of the Main Development Region (Caribbean Sea, Greater Antilles, Lesser Antilles) – an area of the Atlantic Basin where the majority of hurricanes develop10. ACE values for individual storms were obtained from the Department of Atmospheric Science, Colorado State University, Fort Collins, Colorado (Supplementary Table S1). Because we were interested in the temporal effects of inclement weather on passage birds, we divided ACE values by month into September ACE, October ACE (October and the first week of November, combined), September/October ACE (September and October ACE combined), and total ACE (the total ACE for the entire Atlantic Basin hurricane season including outside our target area). We included early November storms in October ACE because, although most Veeries will have entered South America by that time48,50 migration continues within our focal area in northern South America and Veeries often make prolonged use of stopover sites there before continuing southward movement58. Storms that straddled the end of September and the beginning of October were included within the time frame in which the majority of the named storm days in our target area occurred. Similarly, we included storms that straddled August and September as September ACE, and those that straddled October and November as October ACE values. Because the El Niño- Southern Oscillation (ENSO) cycles have been shown to be predictors of tropical climate in both our target area and the Amazon River basin during our period of interest10,59 we included Southern Oscillation Index (SOI) values as a meteorological metric in our modeling. SOI is a measure of the difference in sea-level air pressure across the Pacific Ocean from Tahita in the southeast, to Darwin in northern Australia60. SOI tracks the strength of the Walker Circulation–a prominent atmospheric wind circulation that is strongly linked to ENSO cycles60. ENSO cycles drive annual precipitation patterns, including those in our target area during our period of interest. El Niño years produce less frequent Atlantic tropical storms and hurricanes while La Niña years usually produce more frequent Atlantic tropical storms and hurricanes. SOI is usually negative during El Niño, and positive during La Niña years. We used SOI instead of other climate metrices (e.g., North Atlantic Oscillation index, Atlantic Multidecadal Oscillation) because the SOI is a predictor of seasonal Atlantic hurricane activity10 and has previously been linked to the annual survival and population recruitment of North American Nearctic-Neotropical migratory bird species including Veery17,55. We used the mean SOI values over a 12-month period prior to each breeding season (June to May) which includes the prior hurricane season. We obtained SOI values from the Australian Government Bureau of Meteorology (http://www.bom.gov.au/; Supplementary Table S2). It is important to realize that even if a weather event did not directly affect an individual, tropical climate patterns could have lateral indirect effects including subsequent seasonal carry-over effects17,61.

Veery migratory corridor (rectangle) used to compile Accumulated Cyclone Energy (ACE) predictor values. Annual ACE values were generated from Atlantic Basin hurricanes and tropical storms that formed within or entered the target area from the last week of August through the first week of November 2000 to 2023. Rectangular area is bounded by 100° West, 58° West, 32° North, 7° North (imagery: ESRI, United States Geological Survey).
We chose a model selection framework using Akaike Information Criterion (AIC)62,63. Initially, our chosen predictor variables were (1) September ACE, (2) October ACE, (3) September/October ACE, (4) Total ACE, (5) SOI values. Our response variables were wing chord and a body condition index, all obtained from returning birds in the breeding season following our predictor values, grouped by all combinations of age (SY and ASY) and sex (male and female). Only the first capture of each individual in a season was used for analyses. Throughout the course of our study, after arrival at our study site, we found that a weak negative relationship existed between adult female body mass and ordinal date; however, there was no consistent relationship between ordinal date and adult male mass (females: R2 = 0.05, P = 0.04; males: R2 = 0.006, P = 0.22). Given the power of our 23-year dataset and considering our predictor values lack a temporal component, we consider this weak relationship among females inconsequential in relation to our predicted hypotheses. Descriptive statistics for all response variable categories are included in Supplementary Table S3. For body condition, we converted mass to size-corrected body mass. We scaled body mass to adjust for size following methods described by Peig and Green64which considers the average wing chord of the study population. The index is calculated as:
where \(\:{\widehat{M}}_{l}\) is the predicted mass of individual i with wing length \(\:{L}_{i}\) in mm, Mi is the body mass in grams, \(\:{L}_{0}\) is the population average of wing chord length for the species, and bSMA is the scaling exponent estimated from the standardized major axis (SMA) regression of body mass on wing chord length. To determine if body size needed to be scaled, we conducted a simple linear regression to confirm a significant relationship (P < 0.05) between body mass and wing size and proceeded to calculate scaled body mass64. Because females were not targeted until 2008, they were caught incidentally in only some of the years prior to 2008. As such, data were missing for females in the years 2002 through 2006, with the exception of wing chord in 2004. A total of 346 individual Veeries were used in analysis including 232 males, 100 females, and 14 of unknown sex. Of these, 155 were SY birds including 106 males, 41 females, and 8 of unknown sex. All birds of unknown sex were eliminated from sex-specific analyses.
Initially, all combinations of predictor variables were tested for co-linearity. September ACE and September/October ACE were correlated (Pearson’s Correlation: 0.85, P < 0.0001) as were September ACE and Total ACE (Pearson’s Correlation: 0.65, P < 0.0001); thus, September/October ACE and Total ACE were dropped from further modeling. None of the remaining predictor variables were correlated (P > 0.05). There was no correlation between year (the season from which response variables were measured) and ACE or SOI (P > 0.05).
For all data analysis, we used the statistical program R (version 4.3.1). We first used general linear models fit by maximum likelihood using the R package “lmne”. For ASY birds only, we used a mixed-effect model with individuals as a random effect to account for repeat individuals represented in multiple years. We then applied Akaike’s Information Criterion for small sample sizes (AICc) to assess the best fitting models for each of our response variables using the R package “AICcmodavg”. We used all seven possible combinations of predictors as candidates and included the null model in the candidate set. To enable comparison with analyses at stopover sites where the effect of sex would have been impossible to consider, we analyzed Veeries first by age (ASY, SY) and then separately by male and female. For each response variable, we considered models with a ΔAICc < 2 to be the most parsimonious models62. We chose an 85% CI because it is the most parsimonious CI associated with the AIC paradigm65. We examined each candidate model and eliminated those with probable uninformative parameters65,66. We considered nested parameters likely uninformative if the log likelihood was similar to models with fewer parameters and if the suspected predictor had an 85% CI that crossed zero. We applied the same for top candidate models that were ranked within a ΔAICc < 2 of the null model. In all cases, the single remaining model with the highest rank and the least number of informative parameters was used to draw inference67.
All research was conducted with appropriate state and federal permits. The Delaware State University Institutional Animal Care and Use Committee approved all research methods. This manuscript conforms to the ARRIVE guidelines.
Results
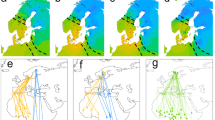
SOI and ACE, both indices to tropical weather in South America, Central America, the Gulf of Mexico, and the Caribbean Sea regions during migratory periods, were represented in various candidate models that showed weak to moderate directional effects on Veery wing morphology and returning body condition (Table 1). SOI was the most parsimonious model explaining adult (male and female) wing chord and showed an inverse relationship with ASY wing chord resulting in a 2 mm decrease across the predictor gradient (−12.5%, range: 90–106 mm, ꞵSOI = −0.54, 85% CI: −0.8 to −0.3; Fig. 2a). When parsed by sex, SOI was the most parsimonious model explaining ASY male wing chord (Table 1). SOI showed an inverse relationship with ASY male wing chord, resulting in a decline of 1.0 mm across the gradient (−7.6%, range: 93–106 mm, ꞵSOI = −0.36, 85% CI: −0.61 to −0.11; Fig. 2b). Although SOI also showed an inverse relationship with ASY female wing chord, consistent with the results of SOI on adult wing chord (−2.0 mm, −18.2%, range: 90–101 mm, ꞵSOI = −0.56, 85% CI: −1.04 to −0.09), September ACE was the more parsimonious model for ASY female wing chord resulting in a 1.8 mm decline in wing chord (−11.2%, range: 90–106 mm, ꞵSept = −0.02, 85% CI: −0.04 to −0.01; Fig. 2c). September ACE + October ACE was the best performing model explaining SY wing chord (Table 1). September ACE showed a positive relationship with SY wing chord resulting in a 2.2 mm increase (+ 17%, range: 89–102 mm, ꞵSept = 0.02, 85% CI: 0.005 to 0.03; Fig. 2d). October ACE also showed a positive relationship with SY wing chord resulting in a 1.2 mm increase (+ 9.0%, range: 89–102 mm, ꞵOct = 0.02, 85% CI: 0.005 to 0.04; Fig. 2e). There was no support for the candidate models in relation to male (n = 106) and female (n = 41) SY wing chord.

Relationship between three independent predictor variables and the wing chord of Veeries returning to a Delaware and Pennsylvania, USA, breeding site over a 23-year period (2000–2023): (a) relationship between the Southern Oscillation Index (SOI) and adult (after second-year; ASY) wing chord (n = 359); (b) relationship between SOI and adult male wing chord (n = 252); (c) relationship between September ACE values and adult female wing chord (n = 99); (d) relationship between September ACE and second-year (SY) wing chord (n = 155); (e) relationship between October ACE and second-year wing chord (n = 155). Shaded area represents 85% confidence interval boundaries. SOI values are the mean value from the 12-month period prior to capture, and ACE values are from the western Atlantic Basin during the period of southward passage preceding capture (August to November; see text).
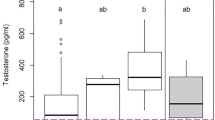
Body condition of returning Veeries showed a relationship with both SOI and ACE (Table 1). Both were included in the top model for ASY body condition (Table 1). However, the second ranked model (SOI) was considered the best model to draw primary inference because of fewer parameters and because September ACE was considered uninformative (ꞵSept = −0.006, 85% CI: −0.14 to 0.002). SOI showed a positive relationship with ASY body condition resulting in a 3 g increase in scaled mass across the gradient (+ 14.0%, range: 20.5–41.8 g, ꞵSOI = 0.32, 85% CI: 0.01 to 0.63; Fig. 3a). However, when parsed by sex, September ACE showed a meaningful inverse relationship with ASY female body condition resulting in a 1.2 g decline in scaled mass (−6.8%, range: 23.6–41.2 g, ꞵSept = −0.03, 85% CI: −0.05 to −0.004; Fig. 3b). September ACE showed a positive effect on SY female body condition resulting in a 2.5 g increase in scaled mass (+ 18.0%, range: 27.4–41.3 g, ꞵSept = 0.04, 85% CI: 0.004 to 0.07; Fig. 3c). There was no support for an effect of the candidate models on male (ASY and SY; n = 348) or female (ASY and SY; n = 138) body condition, ASY male body condition (n = 246), SY body condition (male and female combined; n = 151), or SY male body condition (n = 102).

Relationship between two independent predictor variables and the body condition indices (scaled body mass) of Veeries returning to a Delaware and Pennsylvania, USA, breeding site over a 23-year period (2000–2023): (a) relationship between the Southern Oscillation Index (SOI) and adult (after-second year; ASY) body condition (n = 359); (b) relationship between September Accumulated Cyclone Energy (ACE) and adult female body condition (n = 99); (c) relationship between September ACE and second-year (SY) female body condition (n = 41). Shaded area represents 85% confidence interval boundaries. SOI values are the mean value from the 12-month period prior to capture, and ACE values are from the western Atlantic Basin during the period of southward passage preceding capture (August to November; see text).
Discussion
We revealed a relationship between tropical climate and the phenotypic profile of a Nearctic-breeding songbird over a 23-year period and provided support that direct selective pressure on morphology resulted from inclement tropical weather events (e.g., tropical storms and hurricanes) encountered on southward migration. We believe these data are significant because they are the first to show that severe en route tropical weather is (1) likely a driver of inter-annual morphological variation in a Nearctic-breeding migratory songbird which presumably has heritable effects33 (but see Adams et al.43), and (2) is likely a source of significant mortality and carry-over effects that may include fitness consequences. These results complement those from other studies that suggest tropical climate and inclement weather may be an important contributor to annual population fluctuation and regulation of some Nearctic-breeding songbird populations6,16,17,20,55.
Wing chord
Wing morphology, including shape and size, affects the energetic efficiency of migratory flight68,69. Therefore, phenotypes that optimize flight performance during migration are under strong selection pressure68,69. Turbulence and atmospheric disturbances, more frequent in years with positive SOI and greater ACE, increase energy expenditure by migrants resulting in further selective pressure to optimize efficient wing morphology11. The effect of September ACE on adult female wing chord showed a negative relationship (i.e., smaller wing chord in years with more en route storm activity). However, the effect of ACE on SY (males and females combined) wing chord was opposite that of adult females – wing chord of SY birds was greater in years following more storm activity. During southward migration, HY birds migrate through the tropics later than adults48likely explaining the additive effect of October ACE on SY wing chord. The differences among males, females, and SY birds, suggest that vulnerability to inclement en route weather depends on the temporal and spatial aspects of migratory behavior which are known to be sex and age dependent48,50,52. Severe tropical weather during September correlated with shorter wing chord in female Veeries possibly because short wing chord favors increased maneuverability in dense forest. Veeries that arrive in northern South America after non-stop flights over open water are in poor physical condition in comparison to those that arrive via an overland route through Central America and the Darien region, suggesting that those arriving via Central America frequently stop to refuel48,53. We believe that females may be more likely to travel over land through Central America during southward migration than males, rather than take long flights over open water50. If so, females likely make more frequent and lengthier stopovers than males50. Overland routes with frequent stops to refuel will favor birds that possess shorter wings and a lower wing aspect ratio than those that may travel long distances over open water68,69. The positive effect of ACE on SY wing chord (i.e., greater wing chord in seasons with higher ACE) suggests that SY birds that can maximize efficiency during long sustained flights (i.e., fewer stopovers later in the season), perhaps while circumnavigating storms, are more likely to return to our study site.
Increased SOI was a predictor of smaller wing chord in adults (males and females combined) and for adult males alone. September ACE was the most parsimonious predictor of adult female wing chord, showing an inverse effect. SOI is a predictor of tropical meteorological disturbances during our period of interest, but ENSO cycles are also a predictor of rainfall anomalies in the southern Amazon Basin when Veeries are occupying their non-breeding grounds. It is impossible to know which might be more influential, bearing in mind the effects might not be mutually exclusive. Notably, the effect of SOI on adult wing chord (i.e., smaller wing chord in relation to increased inclement weather) is consistent with the effects of September ACE on adult females (i.e., smaller wing chord and lower body condition in years with higher ACE).
Body condition
Returning body condition is of interest because a challenging autumn migration could result in compromised physical condition, which could prompt detrimental carry-over effects that transcend multiple seasons, similar to those previously detected in our study population and others50,70. For example, arriving in South America in a compromised state could affect an individual’s ability to secure high quality foraging resources which in turn could affect their ability to properly prepare for northward migration70. Compromised body condition of returning birds could affect subsequent reproductive success and therefore population recruitment over time69,71.
Our analyses showed support, albeit of limited strength, for seasonal carry-over effects from inclement weather events. SOI was a predictor of returning ASY body condition. As a group, male and female adult birds returned in better condition following years with positive SOI values. Birds in better physical condition may be more likely to survive harsh environmental conditions72. This suggests that birds in good physical condition were more likely to return than those in less optimal condition after years with more en route severe weather. Alternatively, because positive SOI values are a predictor of rainfall anomalies in the southern Amazon Basin, these conditions could have a positive effect on returning adult body condition if dietary resources (e.g., fruit) are more plentiful or higher quality in years with positive SOI values (i.e., greater moisture). Paxton et al.61 found that vegetation greenness, as measured by the Normalized Difference Vegetation Index, is greater in South America during La Niña-type conditions (i.e., increased precipitation) and likely affects Nearctic-Neotropical migratory songbird energetics prior to northward migration. A combination of both (i.e., selection against compromised body condition and increased quality of dietary resources) could result in our observed effects on returning body condition in positive ENSO (La Niña) type years. When adults were parsed by sex, SOI was not an influential predictor of adult body condition suggesting that the ability to detect the effect of SOI was limited in the respective smaller sample sizes.
September severe weather was a predictor of female body condition for both adult and SY birds. Consistent with the results found for wing chord, adult female body condition indices were lower in years following high September ACE values. The mean date of adult female entry into South America occurs in the first week of October revealing that most individuals are en route across our region of interest in mid to late September during such time they would be susceptible to severe September weather events. However, SY birds on average returned in better body condition during years of increased severe September weather. Although the effect of severe weather in September may have, in some unrecognized way, positively affected female SY body condition, it seems more likely that individuals in good physical condition were more likely to have survived a challenging migratory period than those in a more compromised state. Male body condition did not show a meaningful response to our predictors. In comparison to females, males will prematurely terminate parental care at late season nests presumably due to the seasonal constraints of oncoming fall migration (e.g., need for timely molt)50. Thus, in comparison to females, males may be more readily able to avoid the consequences of severe weather encountered on migration if they are in better physical condition than females that may prolong parental care (see Heckscher et al.55) prior to departure.
Despite our results, the models for body condition narrowly outperformed our null intercept. Resources at our study site could have affected our ability to detect the effects of tropical climate on returning birds. For example, our study site sits in a fragmented and somewhat urban landscape and much of the forests at our study site are infested with alien invasive plants56. Both landscape fragmentation and alien invasive plants can affect arthropod biomass and species richness73,74 the effects of which could mask other drivers of body condition indices such as tropical climate. In addition, we considered the annual breeding-season decrease in female Veery body mass inconsequential to our objectives, yet it remains possible the weak relationship could have affected our ability to detect body condition changes that might have persisted over the intervening life history stages. Regardless, we were able to detect a “storm signal” reflected in our body condition indices across our 23-year dataset, albeit of limited strength. Future studies should examine the relationship between tropical climate and the local breeding-season on the body condition of returning Nearctic-Neotropical migrant songbirds.
Summary
The interplay among our predictors and response variables, supports our hypotheses that en route tropical weather is a driver of morphology and body condition of Veeries returning to a North American breeding site. Annual ACE and SOI values are both related to incidents of inclement weather in the Veery migratory corridor. Frequent encounters with atmospheric disturbances – most likely to occur in years with higher ACE and positive SOI values – significantly increase the energy demands required to complete migration and should result in directional selection on wing morphology and body condition as our data suggest11. For example, for Catharus thrushes, a correlation between wind speed encountered in flight and energy expenditure (both for headwinds and tailwinds) has been documented11. In our study, we detected a negative relationship of SOI with adult Veery wing chord (smaller adult wing chord in La Niña type years), a negative effect of September ACE on female wing chord (i.e., smaller wing chord after severe September storms), and a positive relationship of ACE with SY wing chord (i.e., larger returning wing chord in La Niña type years). In La Niña years, adult Veeries returned in better physical condition, while females returned in a compromised body condition in breeding seasons following severe September en route weather in our target corridor (e.g., tropical storms and hurricanes). On average, SY females returned in better physical condition following greater September ACE. Rainfall anomalies in South America which may affect food resources during the Veery’s non-transient periods in relation to positive SOI (ENSO) values, may also play a role in our observed effects of SOI on Veery morphometrics and body condition (cf. Paxton et al.)61. Ideally, a meta-analysis from breeding sites at a regional or continental scale should be conducted to assess whether these effects hold for other single-brooded species that migrate from temperate or boreal breeding grounds through the tropics. The differing effects of ACE among males, females, and age groups, are expected based on the differences in the spatiotemporal variability of southward migration previously determined by individual tracking devices using both light-level geolocators and GPS loggers, and from prior studies at stopover sites. For example, October severe weather was influential for SY birds likely because HY birds migrate later than adults. Adult females returned to the breeding site in a compromised body condition following years of more severe September storm activity. This was not the case for males possibly because males may be, on average, in better physical condition prior to southward migration than females as females are often providing parental care later in the season than males50. Finally, it is impossible to know the extent of direct versus indirect effects of ACE and SOI on our study subjects, including seasonal carry over effects which could have fitness consequences.
It is important to emphasize that our results do not preclude the possibility of other sources of selection on the wing morphology and body condition of Nearctic-Neotropical migrants returning to our study site. Our focus was inclement weather, and our period of interest was post-breeding southward migration. Paxton et al.61 showed effects of ENSO cycles experienced by songbirds in South America during non-transient periods and Buler et al.75 revealed patterns in songbird wing morphology in relation to near-shore stopover locations along an ecological barrier suggesting sources of en route selection during northward migration. It is possible the environmental effects encountered after southward migration may have masked our ability to detect some directional changes in our study population (i.e., during subsequent non-transient, transient, or breeding periods). Regardless, to that end, our contribution advances the over-arching goal of ornithologists to “disentangle the sources of selection”43 on songbird morphology which is crucial toward understanding how Nearctic-breeding birds are likely to respond to a changing climate37.
It has been nearly impossible to assess the effect of severe tropical storms and hurricanes on migratory songbirds. In lieu of the ability to detect direct mortality, we identified a corresponding signal in our 23-year dataset indicating that inclement weather events during southward migration, and possibly during stationary periods in South America, have been influential in our population. We believe that this “storm signal” should be present in other long-term datasets that have yet to be explored with the potential to further elucidate the effects of storms and hurricanes on migratory songbirds.
Taken cumulatively, our results and those of others6,16,55 show that direct and indirect effects of en route tropical storms and hurricanes, and associated atmospheric disturbances, are likely drivers of both Nearctic-breeding songbird morphometrics and population regulation. Importantly, we believe that sex- and age-specific spatiotemporal differences in migratory patterns may be an adaptation, or perhaps an important exaptation, to unpredictable severe weather events resulting in differential annual survival among cohorts that reinforces population persistence over time in the face of mass mortality events. Although we have not identified fitness consequences, the apparent compromising effect of severe tropical weather on the body condition of returning adult females is an important result of our analyses. We agree with others8,11,55,61 that the projected increase in the severity and frequency of ENSO events associated with climate change, including more severe drought and precipitation events, more hurricanes, rapid storm intensification, larger storms, greater sustained high wind speed, excessive downpours, and slower hurricane translation speed within migratory corridors24,25,26,27,28 are likely an emerging threat to migratory bird populations, the effects of which warrant increased attention from ornithologists and conservationists concerned with the future of Nearctic-Neotropical migratory songbirds.
Data availability
Data used for predictor variables in all models are available in Supplementary Material. Data used for response variables are archived at Delaware State University and are available upon reasonable request from the corresponding author.
References
Robbins, C. S., Sauer, J. R., Greenberg, R. S. & Droege, S. Population declines in North American birds that migrate to the neotropics. Proc. of the Nat. Acad. of Science 86, 7658–7662 (1989).
Sauer, J. R., Pendleton, G. W. & Peterjohn, B. G. Evaluating causes of population change in North American insectivorous songbirds. Cons Bio. 10, 465–478 (1996).
Ballard, G., Geupel, G. R., Nur, N. & Gardi, T. Long-term declines and decadal patterns in population trends of songbirds in Western North america, 1979–1999. Condor 105, 737–755 (2003).
Moore, F. & Kerlinger, P. Stopover and fat deposition by North American wood-warblers (Parulinae) following spring migration over the Gulf of Mexico. Oecologia 74, 47–54 (1987).
Smolinsky, J. A., Diehl, R. H., Radzio, T. A. & Delaney, D. K. Moore. Factors influencing the movement biology of migrant songbirds confronted with an ecological barrier. Behav. Ecol. Socio. 67, 2041–2051 (2013).
Butler, R. W. Stormy seas for some North American songbirds: are declines related to severe storms during migration? Auk 117, 518–522 (2000).
Barrow, W. Jr. et al. US Department of the Interior, U.S. Geological Survey. Circular 1306,. Broad-scale response of landbird migration to the immediate effects of Hurricane Katrina. in Science and the storms: the USGS response to the hurricanes of 2005 (eds., Farris, G. S., Smith G. J., Crane M. P., Robbins, L. L., Lavoie, D. L.) 131–136 (2006).
Watts, B. D. et al. Whimbrel populations differ in trans-Atlantic pathways and cyclone encounters. Sci. Rep. 11, 12919. https://doi.org/10.1038/s41598-021-92429-z (2021) (PMID: 34155292).
Klotzbach, P. J. & Gray, W. M. Updated 6–11-month prediction of Atlantic basin seasonal hurricane activity. Weath Forecast. 19, 917–934 (2004).
Klotzbach, P. J., Saunders, M. A., Bell, G. D. & Blake, E. S. North Atlantic seasonal hurricane prediction: underlying science and an evaluation of statistical models in Climate Extremes: Patterns and Mechanisms (eds Wang, S., Yoon, S. Y., Funk, J. H. & Gillies, R. R.) C. C., 315–328 (Geophysical Monograph 226, American Geophysical Union. John Wiley & Sons, Inc, (2017).
Bowlin, M. S. Wikelski. Pointed wings, low wingloading and calm air reduce migratory flight costs in songbirds. PLOS ONE. 3 (5), e2154. https://doi.org/10.1371/journal.pone.0002154 (2008).
Kimball, S. K. & Mulekar, M. S. A 15-year climatology of North Atlantic tropical cyclones. Part I: size parameters. J. Clim. 17, 3555–3575 (2004).
Ward, M. et al. (ed, P.) Estimating apparent survival of songbirds crossing the Gulf of Mexico during autumn migration. Proc. R Soc. B 285 20181747 https://doi.org/10.1098/rspb.2018.1747 (2018).
Diehl, R. H., Bates, J. M., Willard, D. E. & Gnoske, T. P. Bird mortality during nocturnal migration over lake michigan: A case study. Wilson J. Ornithol. 126, 19–29 (2014).
Wiley, J. W. & Wunderle, J. M. Jr. The effects of hurricanes on birds, with special reference to Caribbean Islands. Bird. Cons Int. 3, 319–349 (1993).
Drymon, J. M. et al. Tiger sharks eat songbirds: scavenging a windfall of nutrients from the Sky. Ecology 100 (9), e02728. https://doi.org/10.1002/ecy.2728 (2019).
Sillett, T. S., Holmes, R. T. & Sherry, T. Impacts of global climate cycle on population dynamics of a migratory Songbird. Science 288, 2040–2042 (2000).
Sillett, T. S. & Holmes, R. T. Variation in survivorship of a migratory Songbird throughout its annual cycle. J. Anim. Ecol. 71, 296–308 (2002).
Rushing, C. S. et al. Spatial and Temporal drivers of avian population dynamics across the annual cycle. Ecology 98, 2837–2850 (2017).
Rockwell, S. et al. Seasonal survival Estimation for a long-distance migratory bird and the influence of winter precipitation. Oecologia 183, 715–726 (2017).
Newton, I. Migration mortality in birds. Ibis https://doi.org/10.1111/ibi.13316 (2024).
Archer, B. Loss of His Majesty’s Ship Phoenix, off the Island of Cuba, in the Year 1780. In, The Mariner’s Chronicle, (ed. Treadway, R. M.) 418–431 (New Haven, 1834).
Gómez, C. & Bayly, N. J. Rosenberg. Seasonal variation in stopover site use: Catharus thrushes and vireos in Northern Colombia. J. Ornithol. https://doi.org/10.1007/s10336-012-0876-5 (2013).
Knutson, T. R. et al. Tropical cyclones and climate change. Nat. Geosci. 3, 157–163 (2010).
Walsh, K. J. E. et al. Tropical cyclones and climate change. WIRE Clim. Change 7, 65–89 (2015).
Kossin, J. P. A global slowdown of tropical-cyclone translation speed. Nature 558, 104–107 (2018).
Bhatia, K. et al. A potential explanation for the global increase in tropical cyclone rapid intensification. Nat. Commun. 13, 6626. https://doi.org/10.1038/s41467-022-34321-6 (2022).
Mendez-Téjeda, R. & Hernández-Ayala, J. J. Links between climate change and hurricanes in the North Atlantic. PLOS Clim. 2 (4), e0000186 (2023).
Salewaski, V., Siebenrock, K., Hochachka, W. M. & Woog, F. Fiedler. Morphological change to birds over 120 years is not explained by thermal adaptation to climate change. PLOS ONE. 9 (7), e101927 (2014). Doi:101371/journal.pone.0101927.
Van Buskirk, J., Mulvihill, R. S. & Leberman, R. C. Declining body sizes in North American birds associated with climate change. Oikos 119, 1047–1055 (2010).
Collins, M. D., Relyea, G. E., Blustein, E. C. & Badami, S. M. Neotropical migrants exhibit variable body-size changes over time and space. Northeastern Naturalist. 24, 82–96 (2017).
Gardner, J. L. et al. Dynamic size responses to climate change: prevailing effects of rising temperature drive long-term body size increases in a semi-arid passerine. Global Ch Biology. https://doi.org/10.1111/gcb.12507 (2014).
Gardner, J. L. et al. Australian songbird body size tracks climate variation: 82 species over 50 years. Proc. R. Soc. B 286, 20192258. https://doi.org/10.1098/rspb.2019.2258 (2019).
Sheridan, J. A. & Bickford, D. Shrinking body size as an ecological response to climate change. Nat. Clim. Change. 1, 401–406 (2011).
Goodman, R. E., Lebuhn, G., Seavy, N. E. & Gardali, T. Bluso-Demers. Avian body size changes and climate change: warming or increasing variability? Glob Change Biol. 18, 63–73 (2012).
Millien, V., Lyons, S. K., Olson, L., Smith, F. A. & Wilson, A. B. Yom-Tov, Y. Ecotypic variation in the context of global climate change: revisiting the rules. Ecol. Lett. 9, 853–869 (2006).
Weeks, B. C. et al. Shared morphological consequences of global warming in North American migratory birds. Ecol. Lett. 23, 316–325. https://doi.org/10.1111/ele.13434 (2020).
Teplitsky, C. & Millien, V. Climate warming and bergmann’s rule through time: is there any evidence? Evol. Appl. 7, 156–168. 10.111/eva (2013).
Yom-Tov, Y. & Geffen, E. Recent Spatial and Temporal changes in body size of terrestrial vertebrates: probable causes and pitfalls. Biol. Rev. 86, 531–541. https://doi.org/10.1111/j.1469-185X.2010.00168.x (2010).
Remacha, C., Rodríguez, C., de la Puente, J. & Pérez-Tris, J. Climate change and maladaptive wing shortening in a long-distance migratory bird. Auk 137, 1–15 (2020).
Anderson, A. M. et al. Consistent declines in wing lengths of Calidridine sandpipers suggest a rapid morphometric response to environmental change. PLOS ONE. 14 (4), e0213930 (2019).
Lank, D. B. et al. Long-term continental changes in wing length, but not bill length, of a long-distance migratory shorebird. Eol & Evol. Doi. https://doi.org/10.1002/ece3.2898 (2017).
Adams, N. et al. Genetic and morphological shifts associated with climate change in a migratory bird. BMC Biol. https://doi.org/10.1186/s12915-024-02107-5 (2025).
Karell, P., Ahola, K., Karstinen, T., Valkama, J. & Brommer, J. E. Climate change drives microevolution in a wild bird. Nat. Comm. https://doi.org/10.1038/ncomms1213 (2011).
Telleria, J. L., de la Hera, I. & Pérez-tris, J. Morphological variation as a tool for monitoring bird populations: A review. Ardeola 60, 191–224. 10.13.157/arla.60.2.2013.191 (2013).
Heckscher, C. M. et al. Miniature GPS dataloggers reveal habitat affiliations and movement of veeries (Catharus fuscescens) during their first non-transient period in South America. Orn Neotrop. 32, 1–9 (2021).
Heckscher, C. M., Taylor, S. M., Fox, J. W. & Afanasyev, V. Veery (Catharus fuscescens) wintering locations, migratory connectivity, and a revision of its winter range using geolocator technology. Auk 128, 531–542 (2011).
González-Prieto, A. M., Hobson, K. A., Bayly, N. J. & Gómez, C. Geographic origins and timing of fall migration of the veery in Northern Colombia. Condor 113, 860–868 (2011).
Hobson, K. A. & Kardynal, K. J. The pull of the central flyway? Veeries breeding in Western Canada migrate using an ancestral Eastern route. J. Field Ornithol. 88, 262–273 (2015).
Heckscher, C. M., Gutierrez, M., Kneidel, A. H. & Ramirez & Reproductive outcomes determine the timing of arrival and settlement of a single-brooded Nearctic-Neotropical migrant Songbird (Catharus fuscescens) in South America. Auk 134, 842–856 (2017).
Hobson, K. A. Kardynal. Western veeries use an Eastern shortest-distance pathway: new insights to migration routes and phenology using light-level geolocators. Auk 132, 540–550 (2017).
Mendez, D. C. Migratory routes and stopover regions of eastern breeding Veeries (Turdidae: Catharus fuscescens) as revealed by archival light-level geolocators. Master’s Thesis, Delaware State University, Dover. 93 pp. (2018).
Gómez, C., Bayly, N. J. & Rosenberg, K. V. Fall stopover strategies of three species of thrush (Catharus) in Northern South America. Auk 131, 702–117 (2014).
Heckscher, C. M., Halley, M. R. & Stampul, P. M. Intratropical migration of a Nearctic-Neotropical migratory Songbird (Catharus fuscescens) in South America with implications for migration theory. J. Trop. Ecol. 31, 285–289 (2015).
Heckscher, C. M. A Nearctic-Neotropical songbird’s nesting phenology and clutch size are predictors of accumulated cyclone energy. Sci. Rep. 8, 9899. https://doi.org/10.1038/s41598-018-28302-3 (2018).
Heckscher, C. M. Veery nest sites in a Mid-Atlantic Piedmont forest: vegetative physiognomy and use of alien shrubs. Am. Midl. Nat. 151, 326–337 (2004).
Pyle, P. Identification Guide to North American Birds, part 1: Columbidae to Ploceidae (Slate Creek, 1997).
Bayly, N. J., Gomez, C., Hobson, K. A., González, A. M. & Rosenberg, K. V. Fall migration of the veery (Catharus fuscescens) in Northern colombia: determining the energetic importance of a stopover site. Auk 129, 449–459 (2012).
Bookhagen, B. & Strecker, M. R. Modern Andean rainfall variation during ENSO cycles and its impact on the Amazon drainage basin. In Amazonia: Landscape and Species Evolution (eds Hoorn, C. & Wesselingh, F.) 223–241 (Blackwell Publishing, 2010).
Power, S. B. & Kociuba, G. The impact of global warming on the Southern Oscillation index. Clim. Dyn. 37, 1745–1754 (2010).
Paxton, K. L., Cohen, E. B., Paxton, E. H., Németh, Z. & Moore, F. R. El Niño-Southern Oscillation is linked to decreased energetic condition in long-distance migrants. PLOS ONE. 9 (5), e95383. https://doi.org/10.1371/journal.pone.0095383 (2014).
Burnham, K. P. & Anderson, D. R. Springer,. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, second edition. (2002).
Tredennick, A. T., Hooker, G., Ellner, S. P. & Adler, P. B. A practical guide to selecting models for exploration, inference, and prediction in ecology. Ecology 102, e03336. https://doi.org/10.1002/ecy.3336 (2021).
Peig, J. & Green, A. J. New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method. Oikos 118, 1883–1891 (2009).
Sutherland, C. et al. Practical advice on variable selection and reporting using Akaike information criterion. Proc. R Soc. B. 290, 20231261. https://doi.org/10.1098/rspb.2023.1261 (2023).
Leroux, S. J. On the prevalence of uninformative parameters in statistical models applying model selection in applied ecology. PLOS ONE. 14 (2), e0206711 (2019).
Cade, B. S. Model averaging and muddled multimodel inferences. Ecology 96, 2370–2382 (2015).
Lockwood, R., Swaddle, J. P. & Rayner, J. M. C. Avian wingtip shape reconsidered: wingtip shape indices and morphological adaptations to migration. J. Avian Biol. 29, 273–292 (1998).
Stojanovic, D. Altered wing phenotypes of captive-bred migratory birds lower post-release fitness. Ecol. Lett. 26, 789–796 (2023).
Marra, P. P., Hobson, K. A. & Holmes, R. T. Linking winter and summer events in a migratory bird by using stable-Carbon isotopes. Science 282, 1884–1886 (1998).
Milenkaya, O., Catlin, D. H., Legge, S. & Walters, J. R. Body condition indices predict reproductive success but not survival in a sedentary, tropical bird. PLOS ONE. 10 (8), e0136582. https://doi.org/10.1371/journal.pone.0136582 (2015).
Evans, P. R. Winter fat deposition and overnight survival of yellow buntings (Emberiza Citronella L). J. Anim. Ecol. 38, 415–423 (1969).
Zanette, P., Doyle, P. & Tremont, S. M. Food shortage in small fragments: evidence from an area-sensitive passerine. Ecology 81, 1654–1666 (2000).
Heleno, R. H., Ceia, R. S., Ramos, J. A. & Memmott, J. Effects of alien plants on insect abundance and biomass: a food-web approach. Conserv. Biol. 23, 410–419 (2009).
Buler, J. J., Lyon, R. J., Smolinsky, J. A., Zenzal, T. J. & Moore, F. R. Body mass and wing shape explain variability in broad-scale bird species distribution of migratory passerines along an ecological barrier during stopover. Physiol. Ecol. 185, 205–212 (2017).
Acknowledgements
We thank the many students and volunteers that assisted with data collection during the 23-year study period especially: S. M. Taylor, P. Stampul, B. Bruce, M. R. Halley, M. Gutierrez Ramirez, A. H. Kneidel, A. A. Aispuro, D. C. Mendez, M. Mancuso, N. Kleponis, M. Gerveni, A. Pickett. D. Baker created Fig. 1. The Delaware Division of Parks and Recreation (DDPR), Delaware Division of Fish and Wildlife (DDFW), and Pennsylvania Game Commission approved research permits for work in Delaware and Pennsylvania, respectively. Thanks to R. Line, C. Bennett and L. Lee of DDPR for continued support of the research at White Clay Creek State Park. Federal bird banding permits were issued by the Bird Banding Laboratory, United States Geological Survey. The University of Delaware and the DDFW provided logistical support prior to 2008. Special thanks to Roland R. Roth for advice and assistance from 1998 to 2008.
Author information
Authors and Affiliations
Contributions
C. M. H. conceived the study, designed the research, conducted and supervised the fieldwork, analyzed the data and wrote the manuscript. T. M. assisted with the creation of figures and data analysis and provided important comments to improve the manuscript. L. A. L. organized data for analysis, assisted with statistical analyses, assisted with the supervision of field assistants, and provided valuable comments to improve the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have declared no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Heckscher, C.M., Mohyuddin, T. & Lester, L.A. Severe en route tropical weather is a predictor of morphological variation and body condition of a Nearctic-Neotropical migratory songbird. Sci Rep 15, 25864 (2025). https://doi.org/10.1038/s41598-025-11395-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-11395-y
