
Abstract
Cholangiocarcinoma, a malignant tumor originating from the bile duct epithelium, poses significant therapeutic challenges and is often associated with poor prognosis. This study aims to elucidate the relationship between plasmacytoma variant translocation 1 (PVT1), a long non-coding RNA implicated in cancer progression, and suppressor of cytokine signaling 2 (SOCS2), recognized for its tumor-suppressive properties. Utilizing the GEPIA2 database, we investigated the expression levels of PVT1 and SOCS2 in cholangiocarcinoma specimens and cell lines through quantitative real-time PCR (qRT-PCR) and Western blotting. Our results demonstrated that PVT1 expression was markedly upregulated in cholangiocarcinoma tissues and cells, while SOCS2 was significantly downregulated. Functional assays revealed that silencing PVT1 inhibited cholangiocarcinoma cell proliferation, migration, and invasion, suggesting its oncogenic role. Conversely, SOCS2 overexpression diminished these malignancies, highlighting its tumor-suppressive function. Notably, the knockdown of PVT1 correlated with an increase in SOCS2 expression, underscoring the regulatory interplay between these two molecules. Ultimately, our findings indicate that lncRNA PVT1 facilitates cholangiocarcinoma progression by repressing SOCS2 expression, positioning SOCS2 as a potential therapeutic target, and warranting further exploration in therapeutic strategies against cholangiocarcinoma.
Similar content being viewed by others

Introduction
Cholangiocarcinoma (CCA) is a malignant tumor originating from the bile duct system. It has an insidious onset, poses challenges for early diagnosis, and carries an unfavorable prognosis1. Due to the lack of specific early diagnostic markers and effective molecular targets for therapy, CCA is often diagnosed at advanced stages, where treatment options are limited1,2. Therefore, identifying novel diagnostic biomarkers and therapeutic targets is imperative to improve patient outcomes.
Long non-coding RNAs (lncRNAs) are a class of non-coding RNAs longer than 200 nucleotides (nt) that lack protein-coding function. LncRNAs regulate gene expression both in the nucleus and cytoplasm through mechanisms such as chromatin modification, transcriptional regulation, and post-transcriptional regulation, playing crucial roles in tumorigenesis and cancer progression3,4. Research has demonstrated that the development and progression of cholangiocarcinoma are closely associated with abnormal lncRNA expression and dysfunction5. Plasmacytoma variant translocation 1 (PVT1), an lncRNA transcribed from a locus approximately 50 kb downstream of the proto-oncogene Myc, has been implicated in tumorigenesis6. Studies have shown that PVT1 contributes to the development of various malignancies, including colorectal cancer7, ovarian cancer8, and prostate cancer9, and is associated with poor prognosis. Although PVT1 is known to be highly expressed in CCA and to promote the proliferation and migration of cholangiocarcinoma cells10, the molecular mechanisms underlying its oncogenic effects remain poorly understood. Moreover, there are limited studies investigating the clinical significance of PVT1 overexpression in CCA.
Suppressor of cytokine signaling2 (SOCS2) is a member of the SOCS protein family and an essential physiological regulator of cytokine signaling11. SOCS2 functions as a negative feedback regulator in several proliferation-related pathways and is associated with cancer progression and prognosis12. SOCS family proteins are involved in multiple signaling pathways, including the circITFG2/miR-1204/SOCS2 pathway13, the JAK2/STAT6 pathway14, the SOCS2/SLC7A11 pathway15, and the PI3K/AKT pathway16. Among the SOCS family members, research specifically focusing on SOCS2 remains relatively limited. SOCS2 also plays a key role as a negative regulator of growth hormone and prolactin signaling17. In cancer, some studies report that SOCS2 inhibits the development of gastrointestinal stromal tumors and enhances their sensitivity to imatinib through KIT activation18. Conversely, other studies reveal that SOCS2 expression is downregulated in hepatoblastoma and correlates with tumor metastasis; overexpression of SOCS2 suppresses hepatoblastoma cell migration and invasion, whereas SOCS2 knockdown promotes malignant phenotypes19. Although SOCS2 downregulation has been observed in cholangiocarcinoma, its functional role in this cancer type remains unexplored.
In this study, we aimed to investigate the clinical relevance of lncRNA PVT1 and SOCS2 expression levels in CCA, to elucidate the role of lncRNA PVT1 in regulating SOCS2 expression, and to explore the significance of the lncRNA PVT1/SOCS2 axis in CCA progression. Furthermore, we evaluated the potential of SOCS2 as a diagnostic biomarker and therapeutic target for CCA.
Materials and methods
Human tissue specimen collection
Human cholangiocarcinoma tissue specimens were collected from The First Department of General Surgery at the Affiliated Hospital of Chengde Medical University. All participants provided signed informed consent, and the study protocol was approved by the Institutional Ethics Committee of the Affiliated Hospital of Chengde Medical University (No. CYFYLL2021018). Specimens were obtained from patients who underwent radical surgery between January 2021 and July 2024, with no prior treatments administered. Fresh surgical tissues were immediately processed for RNA extraction and subsequent qRT-PCR analysis.
Bioinformatics analysis
The expression level of lncRNA PVT1 in cholangiocarcinoma was analyzed using the GEPIA2 online database (http://gepia2.cancer-pku.cn/), and a t-test was performed to assess statistical significance. SOCS2, which was downregulated in cholangiocarcinoma, was identified through gene chip GSE45001 and further validated using the GEPIA2 database, with results were compared.
Cell culture
Normal bile duct epithelial cells (HIBEC) and the human cholangiocarcinoma cell line RBE were obtained from Wuhan Punosai Life Science Co., Ltd., while the human cholangiocarcinoma cell lines Hccc9810 and HuCCT1 were purchased from Beijing Zhulianbihe Biotechnology Co., Ltd. All cell lines (HIBEC, HuCCT1, Hccc9810, and RBE) were cultured in RPMI 1640 medium (GIBCO, USA) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. Cells were maintained at 37 °C in a humidified incubator with 5% CO₂.
Plasmid extraction
A volume of 100–200 µL of glycerol stock containing plasmid (synthesized by Guangzhou Ruibo Biotechnology Co., Ltd.) was inoculated into 8 mL of LB liquid medium supplemented with the appropriate antibiotic (antibiotic-to-medium ratio = 1:1000). The culture was incubated in a constant-temperature shaker (37 °C, 250 rpm) for 12–14 h. After incubation, the bacterial culture was centrifuged at 4200 rpm for 17 min, the supernatant discarded, and plasmids extracted using a centrifugal column-type endotoxin-free plasmid mini-extraction kit (Tiangen Biochemical Technology Co., Ltd., Beijing) following the manufacturer’s instructions under a clean bench. Plasmid concentrations were measured using a NanoDrop One spectrophotometer.
Cell transfection
Cholangiocarcinoma cells were seeded into six-well plates at a density of (2.5–3.5)×10⁵ cells per well. After 24 h of culture, when the cells reached 60–70% confluency, transfection was performed. The transfection mixture contained Lipofectamine 3000 combined with one of the following: PVT1-targeting siRNA(si-PVT1), negative control siRNA(NC-PVT1), PVT1 overexpression plasmid(oe-PVT1), empty vector control(oe-NC), SOCS2 overexpression plasmid(oe-SOCS2), or untreated control(oe-NC). The mixture was incubated for 15 min before use. Meanwhile, the culture medium in the six-well plates was replaced, after which the transfection mixtures were added to their corresponding treatment groups. Following 24–48 h of incubation, cells were harvested. All siRNAs and plasmids (PVT1-targeting siRNA, PVT1 overexpression plasmid, and SOCS2 overexpression plasmid) were synthesized by Guangzhou Ruibo Biotechnology Co., Ltd. (Table 1).The plasmids used in this study contain a selectable marker gene (e.g., Ampicillin resistance), and selection was applied post-transfection using the appropriate antibiotic (e.g., AMP).
Real-time quantitative PCR (qRT-PCR)
Total RNA was extracted from samples using Trizol reagent (Thermo, USA), followed by purification with chloroform and isopropanol. Real-time quantitative polymerase chain reaction (qRT-PCR) was conducted using a Roche cobas z480 instrument, following the manufacturer’s instructions. GAPDH was used as an internal control. The sequences of the primer used in this study were synthesized by Guangzhou Ruibo Biotechnology Co., Ltd. (Table 2).The primer sequences used were as follows: IncDETECTTM h-PVT1_F (Tm: 60 °C, Amplicon Size: 90 bp); GAPDH Forward Sequence (Tm: 58 °C, Amplicon Size: 226 bp).For calculating relative expression levels, we clearly stated that we calculate relative gene expression using the 2^(-ΔΔCt) method and normalize it to GAPDH.
Western blotting
In the Western blotting experiment, the original culture medium was discarded, and the cells were washed twice with pre-cooled PBS. An appropriate amount of RIPA buffer containing protease and phosphatase inhibitors was added according to the cell density. Cells were lysed using ultrasound (50 W, 10 s pulse / 3 s interval) for 8 min, followed by ice incubation for 15 min. Lysates were centrifuged at 12,000×g at 4 °C for 15 min. Protein concentration was determined using a BCA protein assay kit (Pulile, China). Equal amounts of protein were separated by 15% SDS-PAGE and transferred to a polyvinylidene fluoride (PVDF) membrane. Membranes were blocked with 1× fast blocking solution for 30 min and incubated overnight at 4 °C with primary antibodies against SOCS2 and β-actin (internal control). After washing, membranes were incubated with appropriate secondary antibodies, and signals were detected using the C300 imaging system.
Cell scratch assay
To assess the migratory capacity of cholangiocarcinoma cells, HuCCT1 and Hccc9810 cells were cultured in six-well plates. When the cells reached 70–80% confluency, transfection was performed. Twenty-four hours post-transfection, a 10 µL pipette tip was used to create a scratch wound in the monolayer, and images were captured immediately (0 h). Subsequent images were taken at 12-hour and 24-hour time points to monitor cell migration.We have used ImageJ software (National Institutes of Health, USA) to measure the cell-free area at each time point (0 h and post-scratched time points).The migration index was calculated using the following formula:
[\text{Migration index} = \frac{\text{Area at 0 h} - \text{Area at Tx}}{\text{Area at 0 h}} \times 100\% ]where \( Tx \) represents the time point after scratching (e.g., 12–24 h).
Transwell experiment
The transwell assay was performed to evaluate the migration and invasion abilities of cholangiocarcinoma cells using 24-well transwell plates with 8 μm pores. The procedures for migration and invasion assays were identical, except that migration assays used uncoated transwell chambers, whereas invasion assays used Matrigel-coated chambers.Prior to experimentation, cells were serum-starved for 12–24 h and resuspended in serum-free basal medium. A total of 8 × 10⁴ cholangiocarcinoma cells were seeded into each upper chamber, while the lower chamber contained medium supplemented with 20% FBS. After incubation at 37 °C for 12–24 h, non-migrated cells were removed from the upper surface of the membrane with a cotton swab. Migrated or invaded cells on the lower surface were fixed with methanol for 30 min and stained with 0.1% crystal violet for 15 min. After washing, images were acquired using a digital optical microscope (Leica, Germany).we used ImageJ software (National Institutes of Health, USA) to count the migrated/invaded cells. The cell counting was performed manually using the built-in cell counter tool after image thresholding to ensure accuracy and consistency.
Cell-counting kit 8 (CCK-8)
The CCK-8 was used to assess cell proliferation according to the manufacturer’s instructions. After 24 h of transfection, cells were resuspended in 10% fetal bovine serum (FBS)-supplemented medium and plated in a 96-well plate at 2 × 10³ cells/well. Cells were cultured at 37 °C with 5% CO₂ for 0, 24, 48, 72, and 96 h. At each time point, 100 µl of fresh medium containing 10 µl of CCK-8 reagent was added per well and incubated for 2 h. After incubation, the absorbance was measured at 450 nm with 620 nm as the reference wavelength using the Thermo Scientific microplate reader.
Statistical analysis
Experimental data are expressed as mean ± standard deviation. GraphPad Prism 9.0 software was used for data processing and analysis. Each experiment was independently repeated three times. Graphs were generated based on one-way analysis of variance and independent sample t-tests. P-values < 0.05 were considered statistically significant.
Results
PVT1 is highly expressed in cholangiocarcinoma cells and tissues
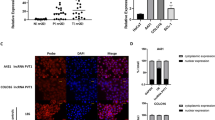
We found that PVT1 was significantly overexpressed in cholangiocarcinoma through the GEPIA2 database analysis(Fig. 1A–C), and it was correlated with the prognosis of patients (Fig. 1D). To further investigate the expression of lncRNA PVT1 in cholangiocarcinoma, PVT1 expression was measured by qRT-PCR in 20 cholangiocarcinoma tissues and paired adjacent normal tissues. Results showed PVT1 was highly expressed in cholangiocarcinoma tissues compared to normal controls (Fig. 1E). Similarly, cholangiocarcinoma cell lines were analyzed, revealing lncRNA PVT1 was highly expressed in HuCCT1, Hccc9810, and RBE relative to normal bile duct epithelial cells HIBEC (Fig. 1F). Based on these findings, HuCCT1 and Hccc9810 were selected for subsequent experiments.

Elevated lncRNA PVT1 expression in cholangiocarcinoma tissues and cells. (A,B) Expression levels of lncRNA PVT1 in tumor tissues compared with paired normal tissues. (C) lncRNA PVT1 expression was significantly elevated in cholangiocarcinoma tissues compared to normal bile duct epithelium. (D) Kaplan-Meier survival analysis of cholangiocarcinoma patients stratified by lncRNA PVT1 expression levels. (E) qRT-PCR analysis of lncRNA PVT1 expression in cholangiocarcinoma tissues versus paired adjacent normal tissues. (F) qRT-PCR detection of lncRNA PVT1 expression in normal bile duct epithelial cells compared with three cholangiocarcinoma cell lines: RBE, Hccc9810, and HuCCT1. *P < 0.05, **P < 0.01,*** P < 0.001.
Knockdown of LncRNA PVT1 can inhibit the proliferation, migration, and invasion of cholangiocarcinoma cells
Since lncRNA PVT1 is upregulated in cholangiocarcinoma, we performed PVT1 knockdown to investigate its detailed role. Functionally, its impact on cholangiocarcinoma cells was assessed through proliferation, migration, and invasion assays.
First, three siRNAs targeting PVT1 (siRNA-1, siRNA-2, and siRNA-3) were designed. siRNAs and a negative control (NC) were transfected into HuCCT1 and Hccc9810 cells. After 48 h, PVT1 expression levels were measured by qRT-PCR. The results showed that all three siRNAs significantly reduced PVT1 expression, with siRNA-3 showing the strongest knockdown efficiency (Fig. 2A). Therefore, siRNA-3 was selected for subsequent experiments. Following siRNA-3 transfection, the effects of PVT1 knockdown on cholangiocarcinoma cell proliferation, migration, and invasion were evaluated using CCK-8, scratch wound healing, and Transwell assays. Compared to the control group, lncRNA PVT1 knockdown significantly impaired the proliferation of HuCCT1 and Hccc9810 cells, as shown by CCK-8 assays(Fig. 2B,C). In addition, migration capacity, assessed by scratch wound healing and Matrigel-free Transwell assays, was markedly reduced (Fig. 2D–J). Similarly, invasion potential, evaluated using Matrigel-coated Transwell assays, was significantly diminished(Fig. 2K–M). These findings establish that lncRNA PVT1 knockdown suppresses the proliferation, migration, and invasion of cholangiocarcinoma cells.

Knockdown of lncRNA PVT1 inhibits the proliferation, migration, and invasion of cholangiocarcinoma cells. (A) Knockdown efficiency of three PVT1-targeting siRNAs in cholangiocarcinoma cells. (B,C) Proliferation capacity of HuCCT1 and Hccc9810 cells following lncRNA PVT1 knockdown, assessed by CCK-8 assay. (D–G) Migratory ability of HuCCT1 and Hccc9810 cells after lncRNA PVT1 knockdown, evaluated by wound healing assay. (H–J) Migration ability of HuCCT1 and Hccc9810 cells following lncRNA PVT1 knockdown, determined by Transwell assay without Matrigel. (K–M) Invasive capacity of HuCCT1 and Hccc9810 cells after lncRNA PVT1 knockdown was examined using matrigel-coated transwell assay. *P < 0.05, **P < 0.01,*** P < 0.001.
Overexpression of SOCS2 can inhibit the proliferation, migration, and invasion of cholangiocarcinoma cells
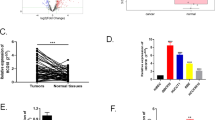
Based on GEPIA2 database analysis, a trend toward downregulation of SOCS2 was observed in cholangiocarcinoma tissues compared to normal adjacent tissues (Fig. 3A–C), and its low expression correlated with poor patient prognosis(Fig. 3D). To investigate the functional role of SOCS2, we overexpressed SOCS2 in cholangiocarcinoma cell lines and confirmed the upregulation of SOCS2 expression by Western blot analysis (Fig. 3G–J).
Functionally, the effects of SOCS2 overexpression on cholangiocarcinoma cells were evaluated through proliferation, migration, and invasion assays.The results showed that in HuCCT1 and Hccc9810 cholangiocarcinoma cell lines, compared with the control group control, after overexpression SOCS2 overexpression of SOCS2, SOCS2 expression was upregulated increased SOCS2 expression in Western blotting experiments analysis(Fig. 3G–J), the proliferation ability proliferation of cholangiocarcinoma cells was reduced decreased in CCK8 experiments assays(Fig. 3E,F), the migration ability of cholangiocarcinoma cells was reduced impaired in scar healing wound healing and Transwell experiments assays without matrix gel Matrigel(Fig. 3K–Q), and the invasive capacity of cholangiocarcinoma cells was reduced in Transwell experiments assays with matrix gel Matrigel (Fig. 3R–T). It was findings demonstrate that overexpression of SOCS2 inhibits the proliferation, migration, and invasion abilities of cholangiocarcinoma cells.

Overexpression of SOCS2 inhibits the proliferation, migration, and invasion of cholangiocarcinoma cells. (A–C) Expression levels of SOCS2 in tumor tissues and their paired adjacent normal tissues. (D) Aberrant SOCS2 expression correlates with the prognosis of cholangiocarcinoma patients. (E,F) Proliferation capacity of HuCCT1 and Hccc9810 cells following SOCS2 overexpression, assessed by CCK-8 assay. (G–J) Western blot analysis showing SOCS2 expression differences in HuCCT1 and Hccc9810 cells after SOCS2 overexpression. (K–N) Migratory ability of HuCCT1 and Hccc9810 cells following SOCS2 overexpression, evaluated by wound healing assay. (O–Q) Migration ability of HuCCT1 and Hccc9810 cells after SOCS2 overexpression, determined by Transwell assay without Matrigel. (R–T) Invasive potential of HuCCT1 and Hccc9810 cells after SOCS2 overexpression, assessed by Matrigel-coated Transwell assay. *P < 0.05, **P < 0.01,*** P < 0.001.
LncRNA PVT1 promotes the proliferation, migration, and invasion of cholangiocarcinoma by inhibiting the expression of SOCS2
To further investigate the regulatory relationship between lncRNA PVT1 and SOCS2, siRNA targeting PVT1 was transfected into HuCCT1 and Hccc9810 cells. After 48 h of culture, SOCS2 expression was assessed by Western blotting. The results demonstrated that PVT1 knockdown upregulated SOCS2 expression compared to the control group, indicating that lncRNA PVT1 negatively regulates SOCS2 expression (Fig. 4A–D).To further verify the regulatory relationship, HuCCT1 and Hccc9810 cells were co-transfected with lncRNA PVT1-overexpressing plasmid and SOCS2-overexpressing plasmid, and divided into three groups: oe-PVT1 + oe-SOCS2, oe-PVT1 + oe-NC and oe-SOCS2 + oe-NC. Western blotting was performed to examine SOCS2 expression across the groups.
Functionally, the CCK-8 assay was conducted to evaluate the effects on cell proliferation. Wound healing and matrix-free Transwell assays were used to assess migration, while Matrigel-coated Transwell assays were used to evaluate invasion capacity.
Western blot analysis showed that compared to the oe-PVT1 + oe-SOCS2 group, SOCS2 expression was downregulated in the oe-PVT1 + oe-NC group, indicating that SOCS2 overexpression partially reversed PVT1-mediated suppression of SOCS2. Additionally, compared to the oe-PVT1 + oe-SOCS2 group, SOCS2 expression was higher in the oe-SOCS2 + oe-NC group, suggesting that PVT1 overexpression counteracted the enhancing effect of SOCS2 overexpression on SOCS2 levels (Fig. 4E,F).
In the CCK-8 assay, cholangiocarcinoma cell proliferation was enhanced in the oe-PVT1 + oe-NC group compared to the oe-PVT1 + oe-SOCS2 group, whereas it was decreased in the oe-SOCS2 + oe-NC group(Fig. 4G,H).In both the scratch wound and matrix-free Transwell migration assays, cholangiocarcinoma cell migration was enhanced in the oe-PVT1 + oe-NC group relative to the oe-PVT1 + oe-SOCS2 group, while it was reduced in the oe-SOCS2 + oe-NC group(Fig. 4I–O). Similarly, in the Matrigel-coated Transwell invasion assay, cholangiocarcinoma cell invasion was increased in the oe-PVT1 + oe-NC group compared with the oe-PVT1 + oe-SOCS2 group, whereas it was decreased in the oe-SOCS2 + oe-NC group(Fig. 4P–R). The results demonstrated that lncRNA PVT1 enhanced the proliferation, migration, and invasion of cholangiocarcinoma cells, whereas SOCS2 suppressed these malignant behaviors. Mechanistically, lncRNA PVT1 facilitates cholangiocarcinoma progression by downregulating SOCS2 expression. PVT1 promotes proliferation, migration, and invasion of cholangiocarcinoma cells, whereas SOCS2 suppresses these malignant behaviors.

lncRNA PVT1 promotes cholangiocarcinoma cell proliferation, migration, and invasion by suppressing SOCS2 expression. (A–D) Western blot analysis showing increased SOCS2 expression in cholangiocarcinoma cell lines following lncRNA PVT1 silencing. (E,F) Western blot analysis of SOCS2 expression following co-transfection. (G,H) CCK-8 assay evaluating the proliferation of HuCCT1 and Hccc9810 cells after co-transfection. (I–L) Wound healing assays assessing the migration ability of HuCCT1 and Hccc9810 cells after co-transfection. (M–O) Matrix-free Transwell assays evaluating the migration capacity of HuCCT1 and Hccc9810 cells after co-transfection. (P–R) Matrigel-coated Transwell assays measuring the invasion ability of HuCCT1 and Hccc9810 cells after co-transfection.
Overexpression of lncRNA PVT1 promotes the proliferation, migration, and invasion of cholangiocarcinoma cells
We overexpressed lncRNA PVT1 and assessed its effects on cholangiocarcinoma cells through proliferation, migration, and invasion experiments. We transfected HuCCT1 and Hccc9810 cells with an overexpression plasmid for PVT1 (oe-PVT1) and the overexpression negative control (OE-NC). We then evaluated the effects of PVT1 overexpression on the proliferation, migration, and invasion of cholangiocarcinoma cells using CCK8, Transwell migration, Transwell invasion, and scratch wound healing assays. The results showed that compared to the control group, cholangiocarcinoma cell proliferation increased after transfection with oe-PVT1 in both HuCCT1 and Hccc9810 cells (Fig. 5H,L). Cholangiocarcinoma cell migration increased in both the scratch wound healing assays and Transwell migration assays without Matrigel (Fig. 5A–G), and invasion ability increased in the Transwell invasion assays with Matrigel (Fig. 5I–K). This suggests that overexpression of lncRNA PVT1 enhances the proliferation, migration, and invasion capabilities of cholangiocarcinoma cells. Furthermore, after overexpressing PVT1, a decrease in SOCS2 expression was observed in Western blot experiments, indicating that lncRNA PVT1 reduces SOCS2 expression(Fig. 5M–P).

Overexpression of lncRNA PVT1 promotes the proliferation, migration, and invasion of cholangiocarcinoma cells. (A–D) The migration ability of cholangiocarcinoma cell lines HuCCT1 and Hccc9810 after overexpression of lncRNA PVT1 was assessed using scratch wound healing assays (**P < 0.01). (H,L) The proliferation ability of cholangiocarcinoma cell lines HuCCT1 and Hccc9810 after overexpression of lncRNA PVT1 was assessed using CCK8 (****P < 0.0001). (E–G) The migration ability of cholangiocarcinoma cell lines HuCCT1 and Hccc9810 after overexpression of lncRNA PVT1 was assessed using Transwell migration assays without Matrigel (*P < 0.05, **P < 0.01). (I–K) The invasion ability of cholangiocarcinoma cell lines HuCCT1 and Hccc9810 after overexpression of lncRNA PVT1 was assessed using Transwell invasion assays with Matrigel (****P < 0.0001). (M–P) The differences in SOCS2 expression in cholangiocarcinoma cell lines after overexpression of lncRNA PVT1 were assessed using Western blot (*P < 0.05, ***P < 0.001).
SOCS2 knockdown promotes the proliferation, migration, and invasion of cholangiocarcinoma cells
To assess its functional impact, we transfected sh-SOCS2 and the negative control sh-NC into HuCCT1 and Hccc9810 cells. We then evaluated the effects of SOCS2 silencing on cell proliferation, migration, and invasion using Cell Counting Kit-8 (CCK8) assays, Transwell assays, and wound healing assays. Compared to the control group, SOCS2 knockdown enhanced the proliferative capacity of both HuCCT1 and Hccc9810 cells in the CCK8 assay (Fig. 6H,L). It also increased their migratory ability in wound healing assays and Transwell migration assays without Matrigel (Fig. 6A–G), as well as their invasive ability in Transwell invasion assays with Matrigel (Fig. 6I–K). These results demonstrate that SOCS2 silencing promotes proliferation, migration, and invasion in cholangiocarcinoma cells. Western blot analysis confirmed reduced SOCS2 expression following knockdown(Fig. 6M–P).

SOCS2 knockdown promotes the proliferation, migration, and invasion of cholangiocarcinoma cells. (A–D) Migratory ability of HuCCT1 and Hccc9810 cells following SOCS2 knockdown was assessed by wound healing assays (** P < 0.01). (H,L) Proliferative ability of HuCCT1 and Hccc9810 cells following SOCS2 knockdown was measured by CCK8 assays (**** P < 0.0001). (E–G) Migratory ability of HuCCT1 and Hccc9810 cells following SOCS2 knockdown was evaluated by Transwell migration assays without Matrigel (* P < 0.05, **** P < 0.0001). (I–K) Invasive ability of HuCCT1 and Hccc9810 cells following SOCS2 knockdown was assessed by Transwell invasion assays with Matrigel (*** P < 0.001). (M–P) Changes in SOCS2 expression levels in cholangiocarcinoma cell lines following SOCS2 knockdown were measured by Western blot assays (* P < 0.05).
Discussion
CCA is a heterogeneous group of malignant tumors originating from the bile ducts. Both the global incidence and mortality rates are increasing20. Its pathogenesis is complex and influenced by multiple factors, including the location of origin and etiology21. Surgical resection remains the only curative option; however, fewer than 20% of cases are resectable22. Although treatment outcomes for CCA patients have improved with advances in medical technology, the 5-year survival rate remains below 5%23. Therefore, there is an urgent need to identify new early diagnostic markers and therapeutic targets to support early diagnosis, improve treatment outcomes, and enhance prognosis in CCA.
PVT1 is a long non-coding RNA implicated in regulating cell survival, proliferation, metabolism, differentiation, and other cellular processes. Located within the cancer risk locus 8q24.21, PVT1 plays a pivotal role in tumorigenesis and progression across multiple cancer types24, consistently exhibiting upregulation in malignancies.For example, in lung cancer, PVT1 acts as an oncogene by modulating key pathways involved in cell proliferation, apoptosis invasion, angiogenesis, and epithelial-mesenchymal transition, thus promoting tumor progression, metastasis, and poor prognosis25. Similarly, in colorectal cancer, PVT1 overexpression enhances cell proliferation, migration, clonogenicity, and invasion7, and its high expression correlates with adverse clinical outcomes, making it a promising diagnostic marker7.In this study, GEPIA2 database analysis revealed significant PVT1 overexpression in cholangiocarcinoma tissues compared to adjacent non-tumorous tissues. Upregulated PVT1 expression was also associated with poor prognosis in cholangiocarcinoma patients. Consistent with database findings, cholangiocarcinoma cell lines exhibited elevated PVT1 levels compared to normal bile duct epithelial cells. Functional experiments showed that PVT1 knockdown significantly attenuated cholangiocarcinoma cell proliferation, migration, and invasion, collectively supporting an oncogenic role for PVT1 in cholangiocarcinoma pathogenesis.
SOCS2 is a member of the SOCS protein family and has been associated with the development of various cancers. SOCS2 plays a crucial role in regulating cellular processes, and its dysregulation is closely linked to malignant tumor progression. Previous studies reported that SOCS2 is upregulated in nasopharyngeal carcinoma patients and cell lines, where high SOCS2 expression correlates with poor prognosis26. Typically, SOCS2 upregulation is associated with unfavorable clinical outcomes. However, emerging research has revealed contradictory roles for SOCS2 in tumorigenesis. For instance, SOCS2 expression is significantly reduced in cervical cancer tissues, and its overexpression inhibits cancer cell proliferation, migration, and invasion27. Additionally, Juan José Cabrera-Galván et al. demonstrated that SOCS2-deficient mice exhibited increased liver tumor burden, higher tumor grade, and enhanced inflammation, fibrosis, and proliferation28, suggesting that SOCS2 functions as a tumor suppressor in hepatocellular carcinoma. Similar tumor-suppressive roles have been reported for SOCS2 in breast cancer29. The mechanisms underlying these context-dependent functions remain unclear, although accumulating evidence indicates that SOCS family genes play critical roles in various human malignancies30. To date, no studies have investigated the role of SOCS2 in cholangiocarcinoma.
In this study, we initially analyzed the GEPIA2 database and observed that SOCS2 was downregulated in cholangiocarcinoma. This downregulation correlated with poor prognosis in cholangiocarcinoma patients. Through Western blotting, we confirmed successful SOCS2 overexpression in cell lines and demonstrated that elevated SOCS2 levels significantly inhibited the proliferation, migration, and invasion of cholangiocarcinoma cells. These findings establish SOCS2 as a tumor suppressor in cholangiocarcinoma. Studies in CCA have revealed that SOX2 downregulates miR-186 through transcriptional activation of lncRNA PVT1, while upregulating SEMA4D expression, thereby promoting CCA progression31. Additionally, lncRNA PVT1 enhances cellular proliferation and migration by modulating ANGPTL4 expression in cholangiocarcinoma10.To explore the regulatory relationship between PVT1 and SOCS2, PVT1 knockdown experiments were performed, followed by Western blotting, which revealed increased SOCS2 expression. This finding suggests that PVT1 negatively regulates SOCS2. Co-transfection experiments with different combinations (oe-PVT1 + oe-SOCS2, oe-PVT1 + oe-NC and oe-SOCS2 + oe-NC) further clarified this relationship: PVT1 overexpression promoted tumorigenic properties in cholangiocarcinoma cells, while SOCS2 overexpression exerted the opposite effect. Rescue experiments confirmed that lncRNA PVT1 promotes cholangiocarcinoma malignancy by suppressing SOCS2 expression. However, the detailed molecular mechanisms underlying this regulatory axis remain to be fully elucidated and warrant further investigation.
In summary, this study revealed a regulatory interaction between lncRNA PVT1 and SOCS2 in cholangiocarcinoma pathogenesis. First, lncRNA PVT1 was significantly overexpressed in cholangiocarcinoma tissues and cell lines. Gene knockdown experiments demonstrated that inhibiting PVT1 effectively reduced cholangiocarcinoma cell proliferation, migration, and invasion, supporting its role as an oncogene. Second, SOCS2 overexpression significantly inhibited the malignant properties of cholangiocarcinoma cells, confirming its tumor-suppressive function. Mechanistic studies further showed that lncRNA PVT1 drives cholangiocarcinoma cell proliferation, migration, and invasion by negatively regulating SOCS2. These findings elucidate the molecular regulatory network involving the PVT1/SOCS2 signaling axis in cholangiocarcinoma and identify a novel therapeutic target.
While this study provides important insights into the PVT1/SOCS2 axis in cholangiocarcinoma, several limitations should be acknowledged. confirming their roles through multiple molecular and phenotypic experiments. Rescue experiments further elucidated their regulatory relationship, demonstrating that lncRNA PVT1 is upregulated in cholangiocarcinoma and promotes tumorigenesis and progression by suppressing SOCS2 expression.
This study has not yet fully explored the underlying regulatory mechanism, such as whether through RNA-protein interactions or chromatin remodeling pathways. And in vivo experiments were not performed, for instance, by constructing conditional gene knockout mouse models to investigate the impact of the PVT1/SOCS2 axis on cholangiocarcinoma proliferation, migration, and invasion. Future studies should focus on more comprehensive mechanistic investigations and incorporate in vivo models to validate the findings and strengthen clinical relevance.
Conclusion
-
(1)
lncRNA PVT1 is significantly overexpressed in cholangiocarcinoma tissues and cells and is strongly associated with poor patient prognosis.
-
(2)
SOCS2 expression is significantly reduced in cholangiocarcinoma tissues and cell lines (p < 0.05) and exhibits an inverse relationship with lncRNA PVT1 expression.
-
(3)
lncRNA PVT1 promotes the proliferation, migration, and invasion of cholangiocarcinoma cells by suppressing SOCS2 expression.
Data availability
The datasets used and analyzed in this study are available from the corresponding author upon reasonable request.
References
Li, Y. et al. Advances in targeted therapy of cholangiocarcinoma. Ann. Med. 56, 2310196 (2024).
Cantallops Vilà, P., Ravichandra, A., Agirre Lizaso, A., Perugorria, M. J. & Affò, S. Heterogeneity, crosstalk, and targeting of cancer-associated fibroblasts in cholangiocarcinoma. Hepatology 79, 941–958 (2024).
Ge, W. J. et al. Long non-coding RNAs in hepatocellular carcinoma. Pathol. Res. Pract. 248, 154604 (2023).
Ghafouri-Fard, S. & Taheri, M. Long non-coding RNA signature in gastric cancer. Exp. Mol. Pathol. 113, 104365 (2020).
Andrade, R., Ribeiro, I. P., Carreira, I. M. & Tralhão, J. G. The diagnostic and prognostic potentials of Non-Coding RNA in cholangiocarcinoma. Int. J. Mol. Sci. 25, 6002 (2024).
Li, C. et al. Long noncoding RNA plasmacytoma variant translocation 1 is overexpressed in cutaneous squamous cell carcinoma and exon 2 is critical for its oncogenicity. Br. J. Dermatol. 190, 415–426 (2024).
Sun, Z., Li, X., Shi, Y. & Yao, Y. LncRNA PVT1 facilitates the growth and metastasis of colorectal cancer by sponging with miR-3619-5p to regulate TRIM29 expression. Cancer Rep. (Hoboken) 7 (2085). (2024).
Li, L., Chen, J., Wang, A. & Yi, K. ALKBH5 regulates ovarian cancer growth via demethylating long noncoding RNA PVT1 in ovarian cancer. J. Cell. Mol. Med. 28, e18066 (2024).
Liang, X. & Liu, B. Exploration of PVT1 as a biomarker in prostate cancer. Med. (Baltim). 103, e39406 (2024).
Yu, Y. et al. Long Non-coding RNA PVT1 promotes cell proliferation and migration by Silencing ANGPTL4 expression in cholangiocarcinoma. Mol. Ther. Nucleic Acids. 13, 503–513 (2018).
Ashfaq, I., Sheikh, N., Fatima, N. & Tayyeb, A. Inhibition of anti-inflammatory pathway through suppressors of cytokine signalling (Socs2/Socs3) in the initiation of hepatocellular carcinoma. Saudi J. Biol. Sci. 29, 103348 (2022).
Jiang, L. et al. Knockdown of m6A methyltransferase METTL3 in gastric cancer cells results in suppression of cell proliferation. Oncol. Lett. 20, 2191–2198 (2020).
Wang, H., Zhang, X., Li, Y., Li, Y. & Pang, T. Lidocaine hampers colorectal cancer process via circITFG2/miR-1204/SOCS2 axis. Anticancer Drugs. 33, 235–244 (2022).
Duan, Y. et al. Lanatoside C inhibits human cervical cancer cell proliferation and induces cell apoptosis by a reduction of the JAK2/STAT6/SOCS2 signaling pathway. Oncol. Lett. 22, 740 (2021).
Wang, J., Jia, Q., Jiang, S., Lu, W. & Ning, H. POU6F1 promotes ferroptosis by increasing lncRNA-CASC2 transcription to regulate SOCS2/SLC7A11 signaling in gastric cancer. Cell. Biol. Toxicol. 40, 3 (2024).
Chi, B. et al. Increased expression of miR-194-5p through the circPVRL3/miR-194-5p/SOCS2 axis promotes proliferation and metastasis in pancreatic ductal adenocarcinoma by activating the PI3K/AKT signaling pathway. Cancer Cell. Int. 22, 415 (2022).
Li, K. et al. SOCS2 regulation of growth hormone signaling requires a canonical interaction with phosphotyrosine. Biosci. Rep. 42, BSR20221683 (2022).
Zhang, L. et al. SOCS2 inhibits the tumorigenesis of gists and increases the sensitivity of gists to Imatinib by suppression of KIT activation. Sci. Rep. 15, 4779 (2025).
Lv, Y. et al. SOCS2 inhibits hepatoblastoma metastasis via downregulation of the JAK2/STAT5 signal pathway. Sci. Rep. 13, 21814 (2023).
Qurashi, M., Vithayathil, M. & Khan, S. A. Epidemiology of cholangiocarcinoma. Eur. J. Surg. Oncol. 51, 107064 (2025).
Khosla, D. et al. Cholangiocarcinoma: recent advances in molecular pathobiology and therapeutic approaches. Cancers (Basel). 16, 801 (2024).
Pawaskar, R. et al. Systematic review of preoperative prognostic biomarkers in Perihilar cholangiocarcinoma. Cancers (Basel). 16, 698 (2024).
Banales, J. M. et al. Cholangiocarcinoma 2020: the next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 17, 557–588 (2020).
Wu, F. et al. Regulation mechanism and pathogenic role of LncRNA plasmacytoma variant translocation 1 (PVT1) in human diseases. Genes Dis. 10, 901–914 (2023).
Hakami, M. A. et al. PVT1 LncRNA in lung cancer: A key player in tumorigenesis and therapeutic opportunities. Pathol. Res. Pract. 253, 155019 (2024).
Dai, B. Q. & Zhang, X. SOCS2 affects the proliferation, migration and invasion of nasopharyngeal carcinoma cells via regulating EphA1. Neoplasma 67, 794–801 (2020).
Li, D., Huang, Y., Wei, M., Chen, B. & Lu, Y. Overexpression of SOCS2 inhibits EMT and M2 macrophage polarization in cervical cancer via IL-6/JAK2/STAT3 pathway. Comb. Chem. High. Throughput Screen. 27, 984–995 (2024).
Cabrera-Galván, J. J. et al. SOCS2 protects against chemical-induced hepatocellular carcinoma progression by modulating inflammation and cell proliferation in the liver. Biomed. Pharmacother. 157, 114060 (2023).
Wang, J. L., Ji, W. W., Huang, A. L., Liu, Z. & Chen, D. F. CEBPA restrains the malignant progression of breast cancer by prompting the transcription of SOCS2. Mol. Biotechnol. 67, 2127–2137 (2025).
Xia, T., Zhang, L., Sun, G., Yang, X. & Zhang, H. Genomic evidence of adaptive evolution in the reptilian SOCS gene family. PeerJ 9, e11677 (2021).
Yu, A. et al. SOX2 knockdown slows cholangiocarcinoma progression through Inhibition of transcriptional activation of LncRNA PVT1. Biochem. J. 477, 3527–3540 (2020).
Acknowledgements
We acknowledge TopEdit LLC for the linguistic editing and proofreading during the preparation of this manuscript.
Funding
Natural Science Foundation of Hebei Province (No. H2021406047).
Author information
Authors and Affiliations
Contributions
Xidong Cao, Liyong Zhang, Kai Chen, Hua Fu and Haizhao Yi performed the in vitro cell experiments and drafted the manuscript. Xidong Cao, Ziyu Bai and Luwen Zhao analyzed and processed the experimental data and images. Xidong Cao and Aijun Yu conceived the study, supervised its progress, and revised the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
I confirm that the research has been approved by the Ethics Committee of the Affiliated Hospital of Chengde Medical University (No. CYFYLL2021018). The research programme complied with the basic elements of the Declaration of Helsinki. We ensure that all procedures strictly adhere to ethical guidelines to maintain the legality and integrity of the study. Participant data is accessible only to authorized researchers.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cao, X., Zhang, L., Chen, K. et al. LncRNA PVT1 promotes proliferation, migration and invasion of cholangiocarcinoma by regulating the expression of SOCS2. Sci Rep 16, 2184 (2026). https://doi.org/10.1038/s41598-025-34019-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-34019-x
