
Abstract
Metastasis is the leading cause of cancer-related deaths, and obesity is associated with increased breast cancer (BC) metastasis. Preclinical studies have shown that obese adipose tissue induces lung neutrophilia associated with enhanced BC metastasis to this site. Here we show that obesity leads to neutrophil-dependent impairment of vascular integrity through loss of endothelial adhesions, enabling cancer cell extravasation into the lung. Mechanistically, neutrophil-produced reactive oxygen species in obese mice increase neutrophil extracellular DNA traps (NETs) and weaken endothelial junctions, facilitating the influx of tumor cells from the peripheral circulation. In vivo treatment with catalase, NET inhibitors or genetic deletion of Nos2 reversed this effect in preclinical models of obesity. Imaging mass cytometry of lung metastasis samples from patients with cancer revealed an enrichment in neutrophils with low catalase levels correlating with elevated body mass index. Our data provide insights into potentially targetable mechanisms that underlie the progression of BC in the obese population.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others


Data availability
Numerical and western blot source data for Figs. 1–8 and Extended Data Figs. 1–8 are provided with this paper. Any additional data that support the findings of this study are available from the corresponding author upon reasonable request. Further information on research design is available in the Nature Research Reporting Summary linked to this article. All RNA-seq data were deposited in the Gene Expression Omnibus under the accession codes GSE165442 (lung neutrophils, Figs. 4a–d and 6c and Extended Data Fig. 6f) and GSE165441 (lung endothelial cells, Fig. 2a–c). Databases used for RNA-seq analysis included MSigDB v.7.2 http://www.gsea-msigdb.org/gsea/msigdb/index.jsp, DoRothEA v.1.3.0 https://saezlab.github.io/dorothea/ and IPA v.01-13 https://digitalinsights.qiagen.com.
Code availability
MATLAB code used for IMC cell segmentation is available upon request.
References
Lauby-Secretan, B. et al. Body fatness and cancer-viewpoint of the IARC working group. N. Engl. J. Med. 375, 794–798 (2016).
Calle, E. E., Rodriguez, C., Walker-Thurmond, K. & Thun, M. J. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N. Engl. J. Med. 348, 1625–1638 (2003).
Risk, N. C. D. Factor collaboration. Trends in adult body-mass index in 200 countries from 1975 to 2014: a pooled analysis of 1698 population-based measurement studies with 19.2 million participants. Lancet 387, 1377–1396 (2016).
Calle, E. E. & Kaaks, R. Overweight, obesity and cancer: epidemiological evidence and proposed mechanisms. Nat. Rev. Cancer 4, 579–591 (2004).
Ligibel, J. A. et al. American Society of Clinical Oncology position statement on obesity and cancer. J. Clin. Oncol. 32, 3568–3574 (2014).
Weigelt, B., Peterse, J. L. & van ‘t Veer, L. J. Breast cancer metastasis: markers and models. Nat. Rev. Cancer 5, 591–602 (2005).
Ewertz, M. et al. Effect of obesity on prognosis after early-stage breast cancer. J. Clin. Oncol. 29, 25–31 (2011).
Osman, M. A. & Hennessy, B. T. Obesity correlation with metastases development and response to first-line metastatic chemotherapy in breast cancer. Clin. Med. Insights Oncol. 9, 105–112 (2015).
Olson, O. C., Quail, D. F. & Joyce, J. A. Obesity and the tumor microenvironment. Science 358, 1130–1131 (2017).
Quail, D. F. et al. Obesity alters the lung myeloid cell landscape to enhance breast cancer metastasis through IL5 and GM-CSF. Nat. Cell Biol. 19, 974–987 (2017).
Adams, T. D. et al. Long-term mortality after gastric bypass surgery. N. Engl. J. Med. 357, 753–761 (2007).
Garcia-Alonso, L., Holland, C. H., Ibrahim, M. M., Turei, D. & Saez-Rodriguez, J. Benchmark and integration of resources for the estimation of human transcription factor activities. Genome Res. 29, 1363–1375 (2019).
Czymai, T. et al. FOXO3 modulates endothelial gene expression and function by classical and alternative mechanisms. J. Biol. Chem. 285, 10163–10178 (2010).
Potente, M. et al. Involvement of Foxo transcription factors in angiogenesis and postnatal neovascularization. J. Clin. Invest. 115, 2382–2392 (2005).
Sang, T., Cao, Q., Wang, Y., Liu, F. & Chen, S. Overexpression or silencing of FOXO3a affects proliferation of endothelial progenitor cells and expression of cell cycle regulatory proteins. PLoS ONE 9, e101703 (2014).
Weber, C., Fraemohs, L. & Dejana, E. The role of junctional adhesion molecules in vascular inflammation. Nat. Rev. Immunol. 7, 467–477 (2007).
Phillipson, M. & Kubes, P. The neutrophil in vascular inflammation. Nat. Med. 17, 1381–1390 (2011).
Boivin, G. et al. Durable and controlled depletion of neutrophils in mice. Nat. Commun. 11, 2762 (2020).
Gomez-Moreno, D., Adrover, J. M. & Hidalgo, A. Neutrophils as effectors of vascular inflammation. Eur. J. Clin. Invest. 48, e12940 (2018).
Rohn, T. T. et al. Priming of human neutrophils by peroxynitrite: potential role in enhancement of the local inflammatory response. J. Leukoc. Biol. 65, 59–70 (1999).
Manda-Handzlik, A. et al. Nitric oxide and peroxynitrite trigger and enhance release of neutrophil extracellular traps. Cell. Mol. Life Sci. 77, 3059–3075 (2020).
Yabe, Y., Nishikawa, M., Tamada, A., Takakura, Y. & Hashida, M. Targeted delivery and improved therapeutic potential of catalase by chemical modification: combination with superoxide dismutase derivatives. J. Pharmacol. Exp. Ther. 289, 1176–1184 (1999).
Chen, A. Y., Lu, J. M., Yao, Q. & Chen, C. Entacapone is an antioxidant more potent than vitamin C and vitamin E for scavenging of hypochlorous acid and peroxynitrite, and the inhibition of oxidative stress-induced cell death. Med. Sci. Monit. 22, 687–696 (2016).
Song, E. et al. Deamidated lipocalin-2 induces endothelial dysfunction and hypertension in dietary obese mice. J. Am. Heart Assoc. 3, e000837 (2014).
Florence, J. M., Krupa, A., Booshehri, L. M., Allen, T. C. & Kurdowska, A. K. Metalloproteinase-9 contributes to endothelial dysfunction in atherosclerosis via protease activated receptor-1. PLoS ONE 12, e0171427 (2017).
Saha, P. et al. Bacterial siderophores hijack neutrophil functions. J. Immunol. 198, 4293–4303 (2017).
Ward, P. P., Mendoza-Meneses, M., Park, P. W. & Conneely, O. M. Stimulus-dependent impairment of the neutrophil oxidative burst response in lactoferrin-deficient mice. Am. J. Pathol. 172, 1019–1029 (2008).
Metzler, K. D. et al. Myeloperoxidase is required for neutrophil extracellular trap formation: implications for innate immunity. Blood 117, 953–959 (2011).
Clark, S. R. et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 13, 463–469 (2007).
Albrengues, J. et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 361, eaao4227 (2018).
Szczerba, B. M. et al. Neutrophils escort circulating tumour cells to enable cell cycle progression. Nature 566, 553–557 (2019).
Perreault, M. & Marette, A. Targeted disruption of inducible nitric oxide synthase protects against obesity-linked insulin resistance in muscle. Nat. Med. 7, 1138–1143 (2001).
Amano, S. U. et al. Local proliferation of macrophages contributes to obesity-associated adipose tissue inflammation. Cell Metab. 19, 162–171 (2014).
Kichenadasse, G. et al. Association between body mass index and overall survival with immune checkpoint inhibitor therapy for advanced non-small cell lung cancer. JAMA Oncol. 6, 512–518 (2019).
McQuade, J. L. et al. Association of body-mass index and outcomes in patients with metastatic melanoma treated with targeted therapy, immunotherapy, or chemotherapy: a retrospective, multicohort analysis. Lancet Oncol. 19, 310–322 (2018).
Domigan, N. M., Charlton, T. S., Duncan, M. W., Winterbourn, C. C. & Kettle, A. J. Chlorination of tyrosyl residues in peptides by myeloperoxidase and human neutrophils. J. Biol. Chem. 270, 16542–16548 (1995).
Harris, I. S. & DeNicola, G. M. The complex interplay between antioxidants and ROS in cancer. Trends Cell Biol. 30, 440–451 (2020).
Blot, W. J. et al. Nutrition intervention trials in Linxian, China: supplementation with specific vitamin/mineral combinations, cancer incidence, and disease-specific mortality in the general population. J. Natl Cancer Inst. 85, 1483–1492 (1993).
Keaney, J. F. Jr. et al. Obesity and systemic oxidative stress: clinical correlates of oxidative stress in the Framingham Study. Arterioscler. Thromb. Vasc. Biol. 23, 434–439 (2003).
Aleman, J. O. et al. Effects of rapid weight loss on systemic and adipose tissue inflammation and metabolism in obese postmenopausal women. J. Endocr. Soc. 1, 625–637 (2017).
Aruoma, O. I., Halliwell, B., Hoey, B. M. & Butler, J. The antioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radic. Biol. Med. 6, 593–597 (1989).
Nagareddy, P. R. et al. Adipose tissue macrophages promote myelopoiesis and monocytosis in obesity. Cell Metab. 19, 821–835 (2014).
Herishanu, Y., Rogowski, O., Polliack, A. & Marilus, R. Leukocytosis in obese individuals: possible link in patients with unexplained persistent neutrophilia. Eur. J. Haematol. 76, 516–520 (2006).
Gentles, A. J. et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 21, 938–945 (2015).
Coffelt, S. B., Wellenstein, M. D. & de Visser, K. E. Neutrophils in cancer: neutral no more. Nat. Rev. Cancer 16, 431–446 (2016).
Kwak, H. J. et al. Myeloid cell-derived reactive oxygen species externally regulate the proliferation of myeloid progenitors in emergency granulopoiesis. Immunity 42, 159–171 (2015).
Adrover, J. M. et al. A neutrophil timer coordinates immune defense and vascular protection. Immunity 50, 390–402 e310 (2019).
Ode, Y., Aziz, M. & Wang, P. CIRP increases ICAM-1(+) phenotype of neutrophils exhibiting elevated iNOS and NETs in sepsis. J. Leukoc. Biol. 103, 693–707 (2018).
Bronte, V. et al. Recommendations for myeloid-derived suppressor cell nomenclature and characterization standards. Nat. Commun. 7, 12150 (2016).
Xia, S. et al. Gr-1+ CD11b+ myeloid-derived suppressor cells suppress inflammation and promote insulin sensitivity in obesity. J. Biol. Chem. 286, 23591–23599 (2011).
Clements, V. K. et al. Frontline science: high fat diet and leptin promote tumor progression by inducing myeloid-derived suppressor cells. J. Leukoc. Biol. 103, 395–407 (2018).
Ng, L. G., Ostuni, R. & Hidalgo, A. Heterogeneity of neutrophils. Nat. Rev. Immunol. 19, 255–265 (2019).
Eiserich, J. P. et al. Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science 296, 2391–2394 (2002).
Wang, Q. et al. Myeloperoxidase deletion prevents high-fat diet-induced obesity and insulin resistance. Diabetes 63, 4172–4185 (2014).
Zhou, J., Wang, Q., Ding, Y. & Zou, M. H. Hypochlorous acid via peroxynitrite activates protein kinase Ctheta and insulin resistance in adipocytes. J. Mol. Endocrinol. 54, 25–37 (2015).
Kessenbrock, K. et al. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 15, 623–625 (2009).
Moorthy, A. N., Tan, K. B., Wang, S., Narasaraju, T. & Chow, V. T. Effect of high-fat diet on the formation of pulmonary neutrophil extracellular traps during influenza pneumonia in BALB/c Mice. Front. Immunol. 7, 289 (2016).
Tohme, S. et al. Neutrophil extracellular traps promote the development and progression of liver metastases after surgical stress. Cancer Res. 76, 1367–1380 (2016).
Cools-Lartigue, J. et al. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Invest. 123, 3446–3458 (2013).
Brotfain, E. et al. Neutrophil functions in morbidly obese subjects. Clin. Exp. Immunol. 181, 156–163 (2015).
Shah, D. et al. Obesity-induced adipokine imbalance impairs mouse pulmonary vascular endothelial function and primes the lung for injury. Sci. Rep. 5, 11362 (2015).
Chedid, P. et al. Adiponectin and its globular fragment differentially modulate the oxidative burst of primary human phagocytes. Am. J. Pathol. 180, 682–692 (2012).
Tilg, H. & Moschen, A. R. Adipocytokines: mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 6, 772–783 (2006).
Miyazaki, Y. et al. Effect of pioglitazone on circulating adipocytokine levels and insulin sensitivity in type 2 diabetic patients. J. Clin. Endocrinol. Metab. 89, 4312–4319 (2004).
Quail, D. F. & Dannenberg, A. J. The obese adipose tissue microenvironment in cancer development and progression. Nat. Rev. Endocrinol. 15, 139–154 (2019).
Guy, C. T., Cardiff, R. D. & Muller, W. J. Induction of mammary tumors by expression of polyomavirus middle T oncogene: a transgenic mouse model for metastatic disease. Mol. Cell. Biol. 12, 954–961 (1992).
Gibby, K. et al. Early vascular deficits are correlated with delayed mammary tumorigenesis in the MMTV-PyMT transgenic mouse following genetic ablation of the NG2 proteoglycan. Breast Cancer Res. 14, R67 (2012).
Ewens, A., Mihich, E. & Ehrke, M. J. Distant metastasis from subcutaneously grown E0771 medullary breast adenocarcinoma. Anticancer Res. 25, 3905–3915 (2005).
Nishikawa, M. et al. Inhibition of metastatic tumor growth by targeted delivery of antioxidant enzymes. J. Control. Release 109, 101–107 (2005).
Kaushik, N. et al. Responses of solid tumor cells in DMEM to reactive oxygen species generated by non-thermal plasma and chemically induced ROS systems. Sci. Rep. 5, 8587 (2015).
Avci, B., Bahadir, A., Tuncel, O. K. & Bilgici, B. Influence of alpha-tocopherol and alpha-lipoic acid on bisphenol-A-induced oxidative damage in liver and ovarian tissue of rats. Toxicol. Ind. Health 32, 1381–1390 (2016).
Davie, S. A. et al. Effects of FVB/NJ and C57Bl/6J strain backgrounds on mammary tumor phenotype in inducible nitric oxide synthase deficient mice. Transgenic Res. 16, 193–201 (2007).
Quail, D. F. et al. The tumor microenvironment underlies acquired resistance to CSF-1R inhibition in gliomas. Science 352, aad3018 (2016).
Huang, Y. et al. Pulmonary vascular destabilization in the premetastatic phase facilitates lung metastasis. Cancer Res. 69, 7529–7537 (2009).
Najmeh, S., Cools-Lartigue, J., Giannias, B., Spicer, J. & Ferri, L. E. Simplified human neutrophil extracellular traps (NETs) isolation and handling. J. Vis. Exp. 16, 52687 (2015).
Rayes, R. F. et al. Primary tumors induce neutrophil extracellular traps with targetable metastasis promoting effects. JCI Insight 5, e128008 (2019).
Yoo, D. G., Floyd, M., Winn, M., Moskowitz, S. M. & Rada, B. NET formation induced by Pseudomonas aeruginosa cystic fibrosis isolates measured as release of myeloperoxidase-DNA and neutrophil elastase-DNA complexes. Immunol. Lett. 160, 186–194 (2014).
Park, J. et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 8, 361ra138 (2016).
Robinson, B. D., Shaji, C. A., Lomas, A. & Tharakan, B. Measurement of microvascular endothelial barrier dysfunction and hyperpermeability in vitro. Methods Mol. Biol. 1717, 237–242 (2018).
Chan, T. F. & Vese, L. A. Active contours without edges. IEEE Trans. Image Process. 10, 266–277 (2001).
Acknowledgements
We thank the Goodman Cancer Research Centre and Life Sciences Complex at McGill University for core facility support, including the Single Cell and Imaging Mass Cytometry Platform (SCIMAP, Y.W.), Histology facility (Cleber Moraes), the Comparative Medicine and Animal Resource Centre and the Flow Cytometry facility. SCIMAP is supported by a McGill MI4 Platform grant and the Fraser Memorial Trust. This study was funded by the Susan G. Komen Foundation (D.F.Q., grant no. CCR18548032), Canada Foundation for Innovation (D.F.Q., JELF-37488; L.A.W., JELF-39178), Canadian Institutes of Health Research (D.F.Q., PJT-159742; L.A.W., PJT-162137) and the Breast Cancer Research Foundation (A.J.D., BCRF-19-034). D.F.Q. is supported by a Tier II Canada Research Chair in Tumor Microenvironment Research. L.A.W. is supported by a Rosalind and Morris Goodman Chair in Lung Cancer Research. S.A.C.M is supported by a Charlotte and Leo Karassik Foundation Fellowship, a Canada Graduate Scholarship from the Canadian Institutes of Health Research and a Canderel Graduate Studentship Award from the Goodman Cancer Research Centre.
Author information
Authors and Affiliations
Contributions
S.A.C.M., L.A.W. and D.F.Q. designed the study, reviewed the data and wrote the paper. A.J.D. provided essential insight throughout the study and contributed neutrophil RNA-seq data. S.A.C.M. oversaw and performed all experiments and data analysis. R.B.E.L., E.K., M.R., R.R.Y. and B.F. provided bioinformatics support. R.B.E.L., A.A., S.D., N.C.B., V.B., B.S., R.F.R. and L.K. provided experimental support and/or optimized protocols. L.J.M.P., D.M., Y.W. and I.R.W. optimized the IMC antibody panel. K.D.L., M.S.M.I., R.F.R., M-C.G., P.O.F. and J.D.S. provided human tissue samples, built the patient tissue microarray and provided pathology/clinical insight. All authors reviewed and approved the paper.
Corresponding authors
Ethics declarations
Competing interests
P.O.F. has received honoraria and consultancy fees from Amgen Canada, AstraZeneca Canada, Bristol Myers Squibb, EMD Serono, Hoffmann-La Roche, Merck Canada, Pfizer Canada and Roche Canada. J.D.S. has received consultancy fees for BMS, Merck, Amgen, AstraZeneca, Protalix and TransHit Bio, and grant funding from AstraZeneca, CLS Therapeutics, Merck and Hoffman La Roche. All other authors declare no competing interests.
Additional information
Peer review information Nature Cancer thanks Luisa Iruela-Arispe, Elena Piskounova and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Obesity is associated with enhanced lung metastasis in multiple genetic models.
a, Weight curves for C57BL6 MMTV-PyMT mice enrolled on LF or HF diet from 5-25 weeks of age. LF, n = 7 mice; HF, n = 9 mice; mean ± SEM. b, Spider-plot showing primary tumor growth for trial shown in a, measured by digital caliper. c, Quantification of metastatic burden in lung from MMTV-PyMT mice at end point. LF, n = 18 lung sections; HF, n = 31 lung sections; Box = median ± interquartile range, whiskers = min-max, all datapoints shown; two-tailed Mann–Whitney test. d, Control conditions for TEM assays using serum gradients. Upper chamber of the Transwell contains 0% serum in all cases. Bottom conditions include: 2% heat-inactivated (H.I.) FBS (n = 4 Transwells), a dose-response from 0%-10% FBS (2%, n = 8 Transwells, 0,5,10%, n = 4 Transwells), 2% serum isolated from LF or HF mice (n = 4 Transwells), and 2% serum isolated from LF or HF and subject to H.I. (n = 4 Transwells); mean ± SEM; two-tailed Student’s t-test. Transwells represent individual experimental replicates with similar results; mouse serum was obtained from different mice. e, Average weight of the leptin-deficient model of genetic-induced obesity (GIO). Wild type (WT) or B6.Cg-Lepob (ob/ob) mice on a C57BL6 background were raised on a standard rodent diet until 10 weeks of age. WT, n = 10 mice; ob/ob, n = 9 mice; mean ± SEM; two-tailed Student’s t-test. f, Schematic illustration of the in vivo extravasation assay corresponding to g-i. WT or ob/ob mice were injected via tail vein with syngeneic breast cancer cells labelled with a green fluorescent CellTracker, and extravasation was quantified in lung after 48h via fluorescence microscopy. g, Quantification of lung vascular density in the in vivo extravasation assay. Vascular density is quantified as CD31+ cells as a percentage of total DAPI+ cells. WT, n = 10 mice; ob/ob, n = 9 mice; mean ± SEM; two-tailed Student’s t-test. h, Fluorescence quantification of Py230 cancer cell extravasation in lung tissue. Extravasation is quantified as the ratio of cancer cells outside the vessels (CellTracker+ CD31-) versus inside the vessels (CellTracker+ CD31+). WT, n = 10 mice; ob/ob, n = 9 mice; mean ± SEM; two-tailed Student’s t-test. i, Representative immunofluorescence image for data in h.
Extended Data Fig. 2 JAM1 is sufficient to regulate vascular permeability.
a, Schematic illustration of the in vitro TransEndothelial Electrical Resistance (TEER) assay. b, Schematic illustration of the in vitro dextran–FITC permeability assay. c, Representative immunofluorescence image for Fig. 2l, showing JAM1+ CD31+ cells in LF (n = n = 4 mice) and HF (n = 7 mice) lung tissue. d, Representative immunofluorescence images for Fig. 2l showing adhesion protein staining in CD31+ cells in LF (n = 4 mice) and HF lung tissue (N-cadherin, n = 4 mice; all others, n = 5 mice). e, Top: Western blot analysis confirming knockdown of VCAM1 protein in HMEC endothelial cells via infection with shRNA targeting VCAM1. Knockdown was confirmed in 2 independent blots. Bottom: TEM quantification in which monolayers were formed with either shSCR or shVCAM1 HMEC endothelial cells prior to assessing Py230 breast cancer transmigration. n = 4 Transwells per group representing individual experimental replicates; mean ± SEM; two-tailed Student’s t-test. f, Representative western blot confirming knockdown of JAM1 protein in HMEC endothelial cells after infection with 2 shRNA constructs. Knockdown was confirmed in 2 independent blots. g, Quantification of TEER across HMEC monolayers genetically modified to express either a JAM1 shRNA (shJAM11, n = 5 Transwells; shJAM12, n = 4 Transwells), a VCAM1 shRNA (shVCAM1; n = 4 Transwells) or a scramble control shRNA (shSCR). Mean ± SEM; two-tailed Student’s t-test. Transwells represent individual experimental replicates with similar results. h, Schematic illustration of the in vitro TEM assay corresponding to Fig. 2o, in which HMEC monolayers genetically modified to express a JAM1 shRNA (shJAM11) or a scramble control shRNA (shSCR) were formed prior to assessing Py230 breast cancer transmigration.
Extended Data Fig. 3 Neutrophil depletion in vivo.
a, Schematic illustration of a modified in vitro TEM assay, in which endothelial cells are pre-treated with neutrophil conditioned media (nCM) generated from bone marrow-derived neutrophils from LF or HF mice. b, Quantification of control TEM assay where HMEC endothelial cells are pre-treated for 24 hours with serum-free media (SFM), or nCM from either LF or HF mice. PyMT cells are then seeded in the upper chamber, and a 0-2% FBS gradient is used to stimulate TEM. n = 4 Transwells per condition representing experimental replicates using nCM from individual mice; mean ± SEM; two-tailed Student’s t-test. c, Schematic of trial design for in vivo 48h extravasation assay with antibody-based neutrophil depletion using ?Ly6G/?Rat or an IgG control. d, Flow cytometry confirmation that neutrophils are effectively depleted in response to ?Ly6G/?Rat treatment, corresponding to experiments in Fig. 3h,i.
Extended Data Fig. 4 Obesity is associated with enhanced ROS in neutrophils.
a, Flow cytometry gating strategy for neutrophils, monocytes, and macrophages in lung. Gates were first drawn around CD45+ CD11b+ cells (not shown), followed by gating according to Ly6G and Ly6C as shown. CD45+ CD11b+ Ly6G+ Ly6Clo neutrophils were used for RNA-seq, corresponding to Fig. 4a-d. b, Weights of LF (n = 3 mice) and HF (n = 4 mice) mice used for lung neutrophil RNA-sequencing. Mean ± SEM. c, Neutrophil CellROX flow cytometry plot, corresponding to Fig. 4e. d, Quantification of CellROX+ neutrophils (CD45+ CD11b+ Ly6G+ Ly6Clo) in lungs from LF or HF B16F10 primary tumor-bearing mice. n = 8 mice per condition; mean ± SEM; two-tailed Mann-Whitney test. e, Quantification of CellROX+ monocytes (CD45+ CD11b+ Ly6G- Ly6Chi) in lungs from LF or HF Py230 primary tumor-bearing mice. LF, n = 5 mice; HF, n = 7 mice; mean ± SEM; two-tailed Mann-Whitney test. f, Quantification of CellROX+ macrophages (CD45+ CD11b+ Ly6G- Ly6C-) in lungs from LF or HF Py230 primary tumor-bearing mice. LF, n = 5 mice; HF, n = 7 mice; mean ± SEM; two-tailed Mann-Whitney test. g, Quantification of CellROX+ monocytes (CD45+ CD11b+ Ly6G- Ly6Chi) in lungs from LF or HF B16F10 primary tumor-bearing mice. n = 8 mice per condition; mean ± SEM; two-tailed Mann-Whitney test. h, Quantification of CellROX+ macrophages (CD45+ CD11b+ Ly6G- Ly6C-) in lungs from LF or HF B16F10 primary tumor-bearing mice. n = 8 mice per condition; mean ± SEM; two-tailed Student’s t-test. i, Quantification of SOD activity (U/ml) using bone marrow neutrophils isolated from WT or ob/ob mice. LF, n = 3 mice; HF, n = 4 mice; mean ± SEM; two-tailed Mann-Whitney test. j, Quantification of SOD activity (U/ml) using serum isolated from WT or ob/ob mice. n = 5 mice per condition; mean ± SEM; two-tailed Student’s t-test. k, Quantification of MPO activity (mU/ml) using bone-marrow neutrophils isolated from LF or HF mice. n = 4 mice per condition; mean ± SEM; two-tailed Student’s t-test. l, Quantification of hypochlorite concentration in bone marrow neutrophils isolated from LF or HF mice harbouring E0771 mammary tumors (tumor-bearing; TB). LF, n = 6 mice; HF, n = 4 mice; mean ± SEM; two-tailed Student’s t-test.
Extended Data Fig. 5 Catalase reduces breast cancer extravasation to lung in obese hosts.
a, Representative immunofluorescence image for data shown in Fig. 5c. LF + veh, n = 5 mice; LF + cat, n = 4 mice; HF + veh, n = 5 mice; HF + cat, n = 4 mice. b, Schematic illustration of the in vivo extravasation assay corresponding to Fig. 5d. LF or HF mice were injected via tail vein with syngeneic breast cancer cells labelled with a green fluorescent CellTracker, treated with either vitamin E or vehicle and extravasation was quantified in lung after 48h via fluorescence microscopy. c, Representative immunofluorescence image for data shown in Fig. 5f. LF + veh, n = 6 mice; LF + cat, n = 4 mice; HF + veh, n = 8 mice; HF + cat, n = 8 mice. d, Schematic illustration of the in vivo extravasation assay corresponding to Fig. 5g,h. WT or ob/ob mice were injected via tail vein with syngeneic breast cancer cells labelled with a green fluorescent CellTracker, treated with either catalase or vehicle and extravasation was quantified in lung after 48h via fluorescence microscopy. e, Representative immunofluorescence image for data shown in Fig. 5g. WT + veh, n = 9 mice; WT + cat, n = 7 mice; ob/ob + veh, n = 7 mice; ob/ob + cat, n = 4 mice. f, Quantification of TEER across an HMEC monolayer genetically modified to express an shRNA against catalase (shCAT), versus a scramble control (shSCR). n = 4 Transwells per condition representing individual experimental replicates with similar results; mean ± SEM; two-tailed Student’s t-test.
Extended Data Fig. 6 Obesity increases MMP9 from neutrophils.
a, Cytokine array from lung neutrophil conditioned media (nCM) obtained from LF or HF mice. MMP9 and LCN2 were the top two differences observed. b, Outline of 111 cytokine array membrane targets, corresponding to a. c, Immunofluorescence quantification of Py230 cancer cell extravasation in lung tissue from LF or HF mice treated with DNase1 or vehicle. LF + veh, n = 5 mice; LF + DNase1, n = n = 4 mice; HF + veh, n = 7 mice; HF + DNase1, n = 6 mice; mean ± SEM; One-way ANOVA with Bonferroni multiple comparisons test. d, Representative immunofluorescence image for data shown in c. e, Immunofluorescence quantification of MPO and H3cit in lung tissue from obese (HF) mice treated with DNase1 or vehicle. n = 4 mice per group; mean ± SEM; two-tailed Student’s t-test. f, RNA-seq differentially expressed genes in HF (n = 4 mice) relative to LF (n = 3 mice) lung neutrophils, showing representative genes relevant to cell cycle progression during metastasis. Data are displayed as log2fold HF vs LF with standard error.
Extended Data Fig. 7 Diet-induced weight changes between WT and Nos2−/− mice.
a, Weight curves comparing wild type (WT) and Nos2−/− mice enrolled on LF, HF or SRD diet from 5-20 weeks of age (15 weeks total). LF WT, n = 10 mice; HF WT, n = 10 mice; HF Nos2−/− n = 5 mice; SRD WT, n = 10 mice; SRD Nos2−/−, n = 10 mice; mean ± SEM.
Extended Data Fig. 8 Imaging mass cytometry (IMC) allows for single cell and spatial analysis.
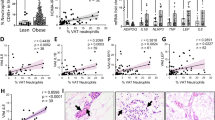
a, Patient information (BMI and primary cancer type) for samples analyzed by IMC. Patients with an asterisk indicate those used for immunofluorescence validation. Patients with BMI<25.9 were assigned to the BMIlow group (ave BMI=24), and patients with a BMI>26 were assigned to the BMIhigh group (ave BMI=30; black line). b, IMC antibody panel information. c, Schematic of IMC staining workflow. Tissue sections were stained with antibody panel (as in b), and subject to CyTOF acquisition using a Hyperion imaging system. d, Immunofluorescence quantification of JAM1+ PyMT tumor cells in lung tissue from WT or ob/ob mice. WT, n = 10 mice; ob/ob, n = 8 mice; mean ± SEM; two-tailed Student’s t-test. e, Quantification of TEM of PyMT breast cancer cells infected with an shRNA against Jam1 (shJAM1) or scramble control (shSCR). n = 4 Transwells per group representing individual experimental replicates; mean ± SEM; two-tailed Student’s T test. f, Quantification of the average number of total CD163- CD68+ M1-like macrophages in BMIlow (n = 8 patients) and BMIhigh (n = 14 patients) lung metastasis samples. Box = median ± interquartile range, whiskers = min-max, all datapoints shown; two-tailed Mann–Whitney test.
Supplementary information
Supplementary Information (download PDF )
Supplementary Fig. 1.
Supplementary Table 1 (download PDF )
Antibody information.
Source data
Source Data Fig. 1 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 2 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 2 (download PDF )
Unprocessed western blots.
Source Data Fig. 3 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 4 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 4 (download PDF )
Unprocessed western blots.
Source Data Fig. 5 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 5 (download PDF )
Unprocessed western blots.
Source Data Fig. 6 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 7 (download XLSX )
Numerical source data for statistics.
Source Data Fig. 7 (download PDF )
Unprocessed western blots.
Source Data Fig. 8 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 1 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 2 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 2 (download PDF )
Unprocessed western blots.
Source Data Extended Data Fig. 3 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 4 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 5 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 6 (download XLSX )
Numerical source data for statistics.
Source Data Extended Data Fig. 8 (download XLSX )
Numerical source data for statistics.
Rights and permissions
About this article

Cite this article
McDowell, S.A.C., Luo, R.B.E., Arabzadeh, A. et al. Neutrophil oxidative stress mediates obesity-associated vascular dysfunction and metastatic transmigration. Nat Cancer 2, 545–562 (2021). https://doi.org/10.1038/s43018-021-00194-9
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s43018-021-00194-9
This article is cited by
-
Neutrophil extracellular traps in cancer
Nature Reviews Cancer (2026)
-
Target neutrophil heterogeneity and plasticity in cancer
Journal of Hematology & Oncology (2025)
-
Neutrophil extracellular traps in cancer: immune modulation, therapy resistance, and the dilemma of targeting
Cell Death & Disease (2025)
-
The NET-DNA-CCDC25 inhibitor di-Pal-MTO suppresses tumor progression and promotes the innate immune response
Cellular & Molecular Immunology (2025)
-
Gemcitabine-induced neutrophil extracellular traps via interleukin-8-CXCR1/2 pathway promote chemoresistance in pancreatic cancer
British Journal of Cancer (2025)
