
Abstract
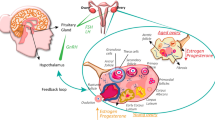
Female reproductive aging has systemic health implications, yet tissue-level dynamics remain poorly understood. Here we integrate deep learning analysis of 1,112 histology images with RNA sequencing from 659 samples across seven female reproductive organs in donors aged 20–70 years. We uncover asynchronous trajectories: the ovary ages gradually, whereas the uterus shows an abrupt molecular and morphological shift around menopause. This uterine transition is independently supported by plasma proteomics data from a large population cohort, indicating that organ-linked aging signatures are detectable in circulation. Tissue segmentation highlights the myometrium as strongly age affected, with extracellular matrix remodeling and immune activation. Epithelial tissues also show coordinated age-related remodeling, with a sharp menopausal transition in the vaginal epithelium. Multi-omics factor analysis links these histological changes to nonlinear gene-expression shifts enriched for reproductive traits, including pelvic organ prolapse and age at menarche. Together, these findings establish menopause as a key inflection point in female aging and provide a tissue-resolved, multi-dataset framework for late-life health.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others


Data availability
The data used for the analyses described in this manuscript were obtained from the GTEx Portal v10 (counts and TPMs) on 27 November 2024 and dbGaP accession number phs000424.v8 (metadata). The fallopian tube single-cell dataset was downloaded from https://cellxgene.cziscience.com/collections/380ade76-e561-49a8-afb2-0f10b39c2c72, and the myometrium single-cell and spatial datasets were downloaded from https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE236660. UKB data are available upon request to qualified researchers through a standard protocol (https://www.ukbiobank.ac.uk/use-our-data/apply-for-access/).
Code availability
Analysis scripts and pipeline are available via GitHub at https://github.com/Mele-Lab/2025_GTEx_Menopause.
References
Monteleone, P., Mascagni, G., Giannini, A., Genazzani, A. R. & Simoncini, T. Symptoms of menopause - global prevalence, physiology and implications. Nat. Rev. Endocrinol. 14, 199–215 (2018).
El Khoudary, S. R. et al. The menopause transition and women’s health at midlife: a progress report from the Study of Women’s Health Across the Nation (SWAN). Menopause 26, 1213 (2019).
Davis, S. R., Pinkerton, J., Santoro, N. & Simoncini, T. Menopause-biology, consequences, supportive care, and therapeutic options. Cell 186, 4038–4058 (2023).
Lambrinoudaki, I., Paschou, S. A., Armeni, E. & Goulis, D. G. The interplay between diabetes mellitus and menopause: clinical implications. Nat. Rev. Endocrinol. 18, 608–622 (2022).
Uddenberg, E. R. et al. Menopause transition and cardiovascular disease risk. Maturitas 185, 107974 (2024).
Liu, J. et al. Development and validation of a predictive model for carotid atherosclerosis in postmenopausal women. Sci. Rep. 15, 5079 (2025).
Mou, H., Zhang, J., Guo, Y., Xu, L. & Luo, X. Effects of key physiological parameters on cardiovascular disease and osteoporosis risk in perimenopausal and postmenopausal women. Sci. Rep. 15, 2814 (2025).
Mosconi, L. et al. Menopause impacts human brain structure, connectivity, energy metabolism, and amyloid-beta deposition. Sci. Rep. 11, 10867 (2021).
Gilmer, G. et al. Female aging: when translational models don’t translate. Nat. Aging 3, 1500–1508 (2023).
Benonisdottir, S., Straub, V. J., Kong, A. & Mills, M. C. Genetics of female and male reproductive traits and their relationship with health, longevity and consequences for offspring. Nat. Aging 4, 1745–1759 (2024).
te Velde, E. R. & Pearson, P. L. The variability of female reproductive ageing. Hum. Reprod. Update 8, 141–154 (2002).
López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M. & Kroemer, G. Hallmarks of aging: an expanding universe. Cell 186, 243–278 (2023).
Murphy, C. T. Aging research: a field grows up. PLoS Biol. 21, e3002132 (2023).
van der Laak, J., Litjens, G. & Ciompi, F. Deep learning in histopathology: the path to the clinic. Nat. Med. 27, 775–784 (2021).
Foroughi pour, A., White, B. S., Park, J., Sheridan, T. B. & Chuang, J. H. Deep learning features encode interpretable morphologies within histological images. Sci. Rep. 12, 1–12 (2022).
Ramirez, J. M. et al. The molecular impact of cigarette smoking resembles aging across tissues. Genome Med. 17, 66 (2025).
Abila, E. et al. Tissue clocks derived from histological signatures of biological aging enable tissue-specific aging predictions from blood. Preprint at bioRxiv https://doi.org/10.1101/2024.11.14.618081 (2024).
Sheehan, S. et al. A machine learning approach for quantifying age-related histological changes in the mouse kidney. Geroscience 46, 2571–2581 (2023).
Chakradeo, K. et al. Contrastive deep learning reveals age biomarkers in histopathological skin biopsies. Preprint at arXiv https://doi.org/10.48550/arXiv.2411.16956 (2024).
Marx, G. A. et al. Histopathologic brain age estimation via multiple instance learning. Acta Neuropathol. 146, 785–802 (2023).
Ribeiro, R., Moraes, A., Moreno, M. & Ferreira, P. G. Integration of multi-modal datasets to estimate human aging. Mach. Learn. 113, 7293–7317 (2024).
Vaswani, A. et al. Attention is all you need. In Adv. Neural Inf. Process. Syst. 30 (eds Guyon, I. et al.) 5998–6008 (2017).
Cisternino, F. et al. Self-supervised learning for characterising histomorphological diversity and spatial RNA expression prediction across 23 human tissue types. Nat. Commun. 15, 5906 (2024).
Fanizzi, A. et al. Comparison between vision transformers and convolutional neural networks to predict non-small lung cancer recurrence. Sci. Rep. 13, 1–10 (2023).
Lian, J. et al. Early stage NSCLS patients’ prognostic prediction with multi-information using transformer and graph neural network model. Elife 11, e80547 (2022).
Park, S.-Y. et al. Vision transformers for low-quality histopathological images: a case study on squamous cell carcinoma margin classification. Diagnostics 15, 1042 (2025).
Jin, C. et al. Molecular and genetic insights into human ovarian aging from single-nuclei multi-omics analyses. Nat. Aging 5, 275–290 (2025).
Wu, M. et al. Spatiotemporal transcriptomic changes of human ovarian aging and the regulatory role of FOXP1. Nat. Aging 4, 527–545 (2024).
Weigert, M. et al. A cell atlas of the human fallopian tube throughout the menstrual cycle and menopause. Nat. Commun. 16, 372 (2025).
Punzon-Jimenez, P. et al. Effect of aging on the human myometrium at single-cell resolution. Nat. Commun. 15, 945 (2024).
Winkler, I. et al. The cycling and aging mouse female reproductive tract at single-cell resolution. Cell 187, 981–998.e25 (2024).
Loid, M. et al. Aging promotes accumulation of senescent and multiciliated cells in human endometrial epithelium. Hum. Reprod. Open 2024, hoae048 (2024).
Aguet, F. et al. The GTEx Consortium atlas of genetic regulatory effects across human tissues. Science https://doi.org/10.1126/science.aaz1776 (2020).
Winstanley, Y. E. et al. Emerging therapeutic strategies to mitigate female and male reproductive aging. Nat. Aging 4, 1682–1696 (2024).
Holzinger, A., Goebel, R., Mengel, M. & Müller, H. Artificial Intelligence and Machine Learning for Digital Pathology: State-of-the-Art and Future Challenges (Springer Nature, 2020).
Komura, D., Ochi, M. & Ishikawa, S. Machine learning methods for histopathological image analysis: updates in 2024. Comput. Struct. Biotechnol. J. 27, 383–400 (2025).
Dos Santos, C. O., Dolzhenko, E., Hodges, E., Smith, A. D. & Hannon, G. J. An epigenetic memory of pregnancy in the mouse mammary gland. Cell Rep. 11, 1102–1109 (2015).
Gray, G. K. et al. A human breast atlas integrating single-cell proteomics and transcriptomics. Dev. Cell 57, 1336–1352 (2022).
Alex, A., Bhandary, E. & McGuire, K. P. Anatomy and physiology of the breast during pregnancy and lactation. In Diseases of the Breast during Pregnancy and Lactation (eds Poli, E., Schattner, A. & Sklair-Levy, M.) 3–7 (Springer, 2020).
Twigger, A.-J. et al. Transcriptional changes in the mammary gland during lactation revealed by single cell sequencing of cells from human milk. Nat. Commun. 13, 1–12 (2022).
InterLACE Study Team Variations in reproductive events across life: a pooled analysis of data from 505 147 women across 10 countries. Hum. Reprod. 34, 881–893 (2019).
Selvaraju, R. R. et al. Grad-CAM: visual explanations from deep networks via gradient-based localization. Int. J. Comput. Vis. 128, 336–359 (2020).
Wang, X., Liu, J., Zou, J., Luo, C. & Wei, D. The study of vaginal wall thickness in adults based on histopathological measurements. Sci. Rep. 14, 18644 (2024).
Binder, R. L. et al. Histological and gene expression analysis of the effects of menopause status and hormone therapy on the vaginal introitus and labia majora. J. Clin. Med. Res. 11, 745–759 (2019).
Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 14, 1–20 (2013).
Goeminne, L. J. E. et al. Plasma protein-based organ-specific aging and mortality models unveil diseases as accelerated aging of organismal systems. Cell Metab. 37, 205–222.e6 (2025).
Oh, H. S.-H. et al. Organ aging signatures in the plasma proteome track health and disease. Nature 624, 164–172 (2023).
Oh, H. S.-H. et al. Plasma proteomics links brain and immune system aging with healthspan and longevity. Nat. Med. 31, 2703–2711 (2025).
Sun, B. B. et al. Plasma proteomic associations with genetics and health in the UK Biobank. Nature 622, 329–338 (2023).
Caron, M. et al. Emerging properties in self-supervised vision transformers. In Proc. IEEE/CVF International Conference on Computer Vision 9650–9660 (IEEE, 2021).
Perry, J. R. B., Murray, A., Day, F. R. & Ong, K. K. Molecular insights into the aetiology of female reproductive ageing. Nat. Rev. Endocrinol. 11, 725–734 (2015).
Gorodeski, G. I. Vaginal-cervical epithelial permeability decreases after menopause. Fertil. Steril. 76, 753–761 (2001).
Stirling, D. R. et al. CellProfiler 4: improvements in speed, utility and usability. BMC Bioinformatics 22, 433 (2021).
Argelaguet, R. et al. Multi-omics factor analysis-a framework for unsupervised integration of multi-omics data sets. Mol. Syst. Biol. 14, e8124 (2018).
Levine, M. E. et al. Menopause accelerates biological aging. Proc. Natl Acad. Sci. USA 113, 9327–9332 (2016).
Sklias, A. et al. Epigenetic remodelling of enhancers in response to estrogen deprivation and re-stimulation. Nucleic Acids Res. 49, 9738–9754 (2021).
Mu, L. et al. Physiological premature aging of ovarian blood vessels leads to decline in fertility in middle-aged mice. Nat. Commun. 16, 72 (2025).
Franceschi, C., Garagnani, P., Parini, P., Giuliani, C. & Santoro, A. Inflammaging: a new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 14, 576–590 (2018).
Kriseman, M. et al. Uterine double-conditional inactivation of and in mice causes endometrial dysregulation, infertility, and uterine cancer. Proc. Natl Acad. Sci. USA 116, 3873–3882 (2019).
Kriseman, M. L. et al. SMAD2/3 signaling in the uterine epithelium controls endometrial cell homeostasis and regeneration. Commun. Biol. 6, 261 (2023).
Marečková, M. et al. An integrated single-cell reference atlas of the human endometrium. Nat. Genet. 56, 1925–1937 (2024).
Wu, Y., Li, M., Zhang, J. & Wang, S. Unveiling uterine aging: much more to learn. Ageing Res. Rev. 86, 101879 (2023).
Wang, B., Han, J., Elisseeff, J. H. & Demaria, M. The senescence-associated secretory phenotype and its physiological and pathological implications. Nat. Rev. Mol. Cell Biol. 25, 958–978 (2024).
Rodrigues, J. et al. Oestrogen enforces the integrity of blood vessels in the bone during pregnancy and menopause. Nat. Cardiovasc. Res. 1, 918–932 (2022).
Broer, S. L., Broekmans, F. J. M., Laven, J. S. E. & Fauser, B. C. J. M. Anti-Müllerian hormone: ovarian reserve testing and its potential clinical implications. Hum. Reprod. Update 20, 688–701 (2014).
Kentistou, K. A. et al. Publisher correction: Understanding the genetic complexity of puberty timing across the allele frequency spectrum. Nat. Genet. 56, 1763–1764 (2024).
Ruth, K. S. et al. Genetic insights into biological mechanisms governing human ovarian ageing. Nature 596, 393–397 (2021).
Stankovic, S. et al. Genetic links between ovarian ageing, cancer risk and de novo mutation rates. Nature 633, 608–614 (2024).
Lehallier, B. et al. Undulating changes in human plasma proteome profiles across the lifespan. Nat. Med. 25, 1843–1850 (2019).
Shen, X. et al. Nonlinear dynamics of multi-omics profiles during human aging. Nat. Aging 4, 1619–1634 (2024).
Márquez, E. J. et al. Sexual-dimorphism in human immune system aging. Nat. Commun. 11, 751 (2020).
Gartoulla, P., Worsley, R., Bell, R. J. & Davis, S. R. Moderate to severe vasomotor and sexual symptoms remain problematic for women aged 60 to 65 years. Menopause 22, 694–701 (2015).
Pujol-Gualdo, N. et al. Advancing our understanding of genetic risk factors and potential personalized strategies for pelvic organ prolapse. Nat. Commun. 13, 3584 (2022).
Hendrix, S. L. et al. Pelvic organ prolapse in the Women’s Health Initiative: gravity and gravidity. Am. J. Obstet. Gynecol. 186, 1160–1166 (2002).
Smith, F. J., Holman, C. D. J., Moorin, R. E. & Tsokos, N. Lifetime risk of undergoing surgery for pelvic organ prolapse. Obstet. Gynecol. 116, 1096–1100 (2010).
Genovese, F. et al. Plasma levels of PRO-C3, a type III collagen synthesis marker, are associated with arterial stiffness and increased risk of cardiovascular death. Atherosclerosis 388, 117420 (2024).
Loomba, R. & Adams, L. A. Advances in non-invasive assessment of hepatic fibrosis. Gut 69, 1343–1352 (2020).
Ravel, J., Moreno, I. & Simón, C. Bacterial vaginosis and its association with infertility, endometritis, and pelvic inflammatory disease. Am. J. Obstet. Gynecol. 224, 251–257 (2021).
Amjadi, F., Salehi, E., Mehdizadeh, M. & Aflatoonian, R. Role of the innate immunity in female reproductive tract. Adv. Biomed. Res. 3, 1 (2014).
Davis, S. R. et al. Menopause. Nat. Rev. Dis. Primers 1, 15004 (2015).
Faddy, M. J., Gosden, R. G., Gougeon, A., Richardson, S. J. & Nelson, J. F. Accelerated disappearance of ovarian follicles in mid-life: implications for forecasting menopause. Hum. Reprod. 7, 1342–1346 (1992).
Alexander, D. H., Novembre, J. & Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 19, 1655 (2009).
GENCODE Consortium. GENCODE - Human Release 39. https://www.gencodegenes.org/human/ (2021).
Xuan, H. et al. Exploring the impact of temperature scaling in Softmax for classification and adversarial robustness. Preprint at https://arxiv.org/html/2502.20604v1 (2025).
Muñoz-Aguirre, M., Ntasis, V. F., Rojas, S. & Guigó, R. PyHIST: a histological image segmentation tool. PLOS Comput. Biol. 16, e1008349 (2020).
Otsu, N. A threshold selection method from gray-level histograms. IEEE Trans. Syst. Man Cybern. 9, 62–66 (1979).
Simonyan, K. & Zisserman, A. Very deep convolutional networks for large-scale image recognition. In 3rd Asian Conference on Pattern Recognition (ACPR) 730–773 (2015).
Deng, J. et al. ImageNet: a large-scale hierarchical image database. in 2009 IEEE Conference on Computer Vision and Pattern Recognition (IEEE, 2009); https://doi.org/10.1109/cvpr.2009.5206848.
Guo, C., Pleiss, G., Sun, Y. & Weinberger, K. Q. In Proc. 34th International Conference on Machine Learning, PMLR Vol. 70, 1321–1330 (2017).
Yao, Y., Rosasco, L. & Caponnetto, A. On Early Stopping in Gradient Descent Learning. Constr. Approx. 26, 289–315 (2007).
Haghighat, M. et al. Automated quality assessment of large digitised histology cohorts by artificial intelligence. Sci. Rep. 12, 1–16 (2022).
Kerr, J. B., Myers, M. & Anderson, R. A. The dynamics of the primordial follicle reserve. Reproduction 146, R205–R215 (2013).
Hoffman, G. E. & Schadt, E. E. variancePartition: interpreting drivers of variation in complex gene expression studies. BMC Bioinformatics 17, 483 (2016).
Davies, R. B. Hypothesis testing when a nuisance parameter is present only under the alternative: linear model case. Biometrika 89, 484–489 (2002).
Swapna, L. S., Huang, M. & Li, Y. GTM-decon: guided-topic modeling of single-cell transcriptomes enables sub-cell-type and disease-subtype deconvolution of bulk transcriptomes. Genome Biol. 24, 190 (2023).
Tabula Sapiens Consortium* et al. The Tabula Sapiens: a multiple-organ, single-cell transcriptomic atlas of humans. Science 376, eabl4896 (2022).
Jin, C. et al. Molecular and genetic insights into human ovarian aging from single-nuclei multi-omics analyses. Nat. Aging. 5, 275–290 (2025).
Aitchison, J. The statistical analysis of compositional data. J. R. Stat. Soc. B 44, 139–160 (2018).
Breiman, L. Random forests. Mach. Learn. 45, 5–32 (2001).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 43, e47 (2015).
Law, C. W., Chen, Y., Shi, W. & Smyth, G. K. voom: precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 15, R29 (2014).
Ferreira, P. G. et al. The effects of death and post-mortem cold ischemia on human tissue transcriptomes. Nat. Commun. 9, 490 (2018).
Tukiainen, T. et al. Landscape of X chromosome inactivation across human tissues. Nature 550, 244–248 (2017).
Aguet, F. et al. Cell type–specific genetic regulation of gene expression across human tissues. Science https://doi.org/10.1126/science.aaz8528 (2020).
The GTEx Consortium et al. The Genotype-Tissue Expression (GTEx) pilot analysis: multitissue gene regulation in humans. Science https://doi.org/10.1126/science.1262110 (2015).
Hron, K., Filzmoser, P., de Caritat, P., Fišerová, E. & Gardlo, A. Weighted pivot coordinates for compositional data and their application to geochemical mapping. Math. Geosci. 49, 797–814 (2017).
Wang, W. et al. Single-cell transcriptomic atlas of the human endometrium during the menstrual cycle. Nat. Med. 26, 1644–1653 (2020).
Swain, M. & Kulkarni, A. D. Endometrium at menopause: the pathologist’s view. J. Midlife Health 12, 310–315 (2021).
Wu, T. et al. clusterProfiler 4.0: a universal enrichment tool for interpreting omics data. Innovation 2, 100141 (2021).
Talukdar, N. et al. Effect of long-term combined oral contraceptive pill use on endometrial thickness. Obstet. Gynecol. 120, 343–349 (2012).
Gallagher, C. S. et al. Author correction: Genome-wide association and epidemiological analyses reveal common genetic origins between uterine leiomyomata and endometriosis. Nat. Commun. 13, 5543 (2022).
Moolhuijsen, L. M. E. et al. Genomic and proteomic evidence for hormonal and metabolic foundations of polycystic ovary syndrome. Preprint at medRxiv 2024.04.18.24306020 https://doi.org/10.1101/2024.04.18.24306020 (2024).
Rahmioglu, N. et al. The genetic basis of endometriosis and comorbidity with other pain and inflammatory conditions. Nat. Genet. 55, 423–436 (2023).
Pathare, A. D. S. et al. A large-scale genome-wide association study on female genital tract polyps highlights role of DNA repair, cell proliferation, and cell growth. Hum. Reprod. https://doi.org/10.1093/humrep/deaf025 (2025).
Pujol Gualdo, N. et al. Atlas of genetic and phenotypic associations across 42 female reproductive health diagnoses. Nat. Med. https://doi.org/10.1038/s41591-025-03543-8 (2025).
Pujol-Gualdo, N. et al. Circulating anti-Müllerian hormone levels in pre-menopausal women: novel genetic insights from a genome-wide association meta-analysis. Hum. Reprod. 39, 1102–1113 (2024).
Ruth, K. S. et al. Using human genetics to understand the disease impacts of testosterone in men and women. Nat. Med. 26, 252–258 (2020).
Sollis, E. et al. The NHGRI-EBI GWAS Catalog: knowledgebase and deposition resource. Nucleic Acids Res. 51, D977–D985 (2023).
The 1000 Genomes Project A global reference for human genetic variation. Nature 526, 68–74 (2015).
Privé, F. Smart Principal Component Analysis. https://search.r-project.org/CRAN/refmans/smartsnp/html/smart_pca.html (2023).
Meyer Lab. Identification of related individuals. https://meyer-lab-cshl.github.io/plinkQC/reference/check_relatedness.html (2020).
Ribeiro Reis, A. P. et al. Retinal morphology across the menstrual cycle: insights from the UK Biobank. npj Womens Health 2, 38 (2024).
Acknowledgements
N.P.-G. acknowledges her AI4S fellowship within the “Generación D” initiative by Red.es, Ministerio para la Transformación Digital y de la Función Pública, for talent attraction (C005/24-ED CV1). This study was funded by the NextGenerationEU funds through PRTR. M.M. was supported by a grant PID2019-107937GA-I00 funded by MCIN/AEI/10.13039/501100011033 and a grant RYC-2017-22249 funded by MCIN/AEI/10.13039/501100011033 and by “ESF Investing in your future.” D. Torrents acknowledges Instituto de Salud Carlos III (ISCIII) and “Unión Europea NextGenerationEU/Mecanismo para la Recuperación y la Resilencia (MRR)/PRTR” under project PMP21/00015, the Departament de Recerca i Universitats de la Generalitat de Catalunya (code: 2021 SGR 01626), the Science and Innovation Spanish Ministry under project PREVDIS (PID2023-152867NB-I00) and European Commission (EU-HORIZON NEARDATA GA.101092644). M.S.-R. was supported by a predoctoral AGAUR-FI Joan Oró fellowship from the Secretaria d’Universitats i Recerca, Departament de Recerca i Universitats, Generalitat de Catalunya and the European Social Fund Plus (FI-3 2024-0065). J.M.R. was supported by a predoctoral fellowship from “la Caixa” Foundation (ID 100010434) with code LCF/BQ/DR22/11950022. A.R.-C. was supported by a predoctoral fellowship “Formación Personal Investigador (FPI)” from “el Ministerio de Ciencia, Innovación y Universidades (MCIN)” and “la Agencia Estatal de Investigación (AEI)” (MCIN/AEI FPI with code PRE2019-090193). We are deeply grateful to W. Oliveros for her invaluable input and timely advice, providing the genetic ancestry annotation. We thank C. Castelo-Branco for his insightful gynecological perspective on reproductive aging. Last, we are grateful to the Melé laboratory at the Barcelona Supercomputing Center (BSC) for their support, discussions and valuable feedback throughout this project.
Author information
Authors and Affiliations
Contributions
M.M. conceived the study; M.M., O.S. and L.V.-S.P. designed and supervised all analyses; O.S., L.V.-S.P., N.P.-G., A.E.H., D. Tabares and M.S.-R. analyzed the data; M.M., O.S., L.V.-S.P. and N.P.-G. wrote the manuscript with input from all co-authors; J.O. provided histopathological annotation and interpretation; M.A.P.-E. and D. Torrents contributed to proteomics analyses; A.R.-C. and J.M.R. advised in data analysis, provided helpful insight and contributed to manuscript editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Aging thanks Qingling Yang and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Age classification and visual interpretation from histology and gene expression across female reproductive organs.
A Image classification-based aging trajectories for the uterus, ovary, and vagina, only for the donors belonging to validation and middle-age sets; each dot corresponds to a sample; solid lines represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions). B CNN-predicted probabilities of being classified as old based on ovary or uterus images from the same donor. C,D Grad-CAM heatmaps for ovary (C) and vagina (D). Intensity indicates the level of importance of the region to age classification. E Continuous age trajectories fitted to a linear model; predictions obtained with a regression-based model on image tile to predict continuous age. F Number of differentially expressed genes with age in the uterus, ovary, and vagina across 20-year sliding windows (1-year offset); each dot corresponds to a number of age-DEGs between donors in age windows (Age - 10 years) and (Age + 10 years). G Proteomics-inferred organ-specific trajectories across time since menopause, solid lines represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions) H-M Proteomics-inferred organ-specific trajectories adjusting for cycle phase (H-I), hormone therapy use (J), contraceptives use (K) and reproductive history (L,M), solid lines represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions).
Extended Data Fig. 2 Vision transformer histology organ segmentation and tile examples.
A Clustering accuracies per donor and tile across organs. B-H Per-organ UMAP projections of a subset of identified tissues and tissue structures. A-G Organ segmentation examples showing tissue and structure classifications. H Age-related histological changes in myometrium. I Follicle count per age group and sample size per group, boxes of the violin plot show median and quartiles, and the whiskers extend to the minimum and maximum values within 1.5×IQR. Sample sizes per age bin are in Extended Data Table 26 J Z-scored proportions of peg cells, ciliated epithelial cells, secretory epithelial cells of fallopian tube samples; dots represent individual samples; solid lines represent locally estimated scatterplot smoothing (LOESS). Significant associations (FDR < 0.05) are marked with asterisks. K The left panel displays the number of cells analyzed per cell type (Supplementary Table 6). The right panel displays estimates from a compositional analysis modeling changes in cell type abundance with age adjusted by ethnicity group. Bar lengths correspond to the estimated age effects (β coefficients) from linear regression models of CLR-transformed cell-type proportions; error bars denote the associated 95% confidence intervals. Significant associations (FDR < 0.05) are marked with filled circles. L-O Age-related histological changes in vagina (higher nuclear-to-citoplasm ratio in epithelia)(L), ectocervix (M), accompanied with the flattening of columnar cells endocervical glandular (N), and fallopian tube epithelium (O) in old donors.
Extended Data Fig. 3 Histological segmentation and changes with age.
A-G Organ segmentation examples showing tissue and structure classifications. H Age-related histological changes in myometrium. I Follicle count per age group and sample size per group, boxes of the violin plot show median and quartiles, and the whiskers extend to the minimum and maximum values within 1.5×IQR. Sample sizes per age bin are in Extended Data Table 26 J Z-scored proportions of peg cells, ciliated epithelial cells, secretory epithelial cells of fallopian tube samples; dots represent individual samples; solid lines represent locally estimated scatterplot smoothing (LOESS). Significant associations (FDR < 0.05) are marked with asterisks. K The left panel displays the number of cells analyzed per cell type (Supplementary Table 6). The right panel displays estimates from a compositional analysis modeling changes in cell type abundance with age adjusted by ethnicity group. Bar lengths correspond to the estimated age effects (β coefficients) from linear regression models of CLR-transformed cell-type proportions; error bars denote the associated 95% confidence intervals. Significant associations (FDR < 0.05) are marked with filled circles. L-O Age-related histological changes in vagina (higher nuclear-to-citoplasm ratio in epithelia)(L), ectocervix (M), accompanied with the flattening of columnar cells endocervical glandular (N), and fallopian tube epithelium (O) in old donors.
Extended Data Fig. 4 Image features trajectories and demographic variability across donors and organs.
A-D Image feature trajectories for each tissue structure in uterus (A), ovary (B), vagina (C), breast (D); each dot corresponds to a sample; curves represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions). E BMI and genetic ancestry distributions of image and RNA-Seq samples (biological replicates, sample sizes in Extended Data Table 1). Box plots show median, quartiles, and 1.5× IQR whiskers. F Mean variance explained by demographic traits across tissues including the residuals (upper plot) and excluding the residuals (middle plot), indicating sample sizes (bottom plot). G BMI-related histological differences in breast adipose tissue, with larger adipocytes and reduced intercellular space in high-BMI donors. H Genetic ancestry-associated differences in vaginal epithelium, appearing denser in donors of African ancestry compared to European ancestry.
Extended Data Fig. 5 Tissue-specific and cell type-specific aging dynamics.
A Balanced accuracies for tissue-specific Elastic-Net classifiers. B Tissue and tissue structure proportions across organs. Asterisk (*) denotes the tissues that change significantly with age (FDR < 0.05) in the compositional data analysis; each dot corresponds to a sample; curves represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions). C Number of images and RNAseq samples with and without endometrium. D Proportion of endometrium within uterus samples across age groups. Each dot corresponds to a sample. E-G Cell-type proportion estimate changes with age for samples of uterus (E), myometrium (F) and ovary (G), bar lengths correspond to the estimated age effects (β coefficients) on mean donor-level cell-type proportions; error bars denote 95% confidence intervals around the coefficient estimates. Sample sizes are indicated in the title (uterus n = 130, myometrium n = 61, ovary n = 169).
Extended Data Fig. 6 Multimodal analysis for ovary and breast and up and downregulated functions with age.
A-B Multi-Omics Factor Analysis (MOFA) results for ovary (A) and breast (B): heatmaps show variance explained by 10 latent factors across modalities (bottom) and their correlation with donor age (top). Asterisks denote significance (*FDR < 0.05 & >0.005, **FDR < 0.005 & >0.0005, ***FDR < 0.0005). C Gene set enrichment analysis (GSEA) using Gene Ontology for ranked genes from pseudobulked menopausal-status-differential expression analyses per each myometrial cell type (FDR < 0.05). Bold terms are collagen or extracellular-matrix related terms. D Trajectory of the most weighted feature for Factor 4 in myometrium, showing the difference between low values in young donors (left) and high values in old donors (right); each dot corresponds to a sample; solid line represents locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions). E Spatial gene expression counts of the top-ranked gene MFAP5 in Factor 4 of the uterus, comparing the expression in one peri (left) and one post-menopausal (right) samples. F Gene Set Enrichment Analysis (GSEA) of the top 7 pathways with positive (NES > 0) and negative (NES < 0) normalized enrichment scores using Gene Ontology for the genes contributing to Factor 4 in the uterus. Enrichment significance was assessed using a Kolmogorov-Smirnov-like running-sum statistic (one-sided test), FDR < 0.05.
Extended Data Fig. 7 Gene Set Enrichment Analysis (GSEA) on MOFA factors.
GSEA of the top 7 pathways with positive (NES > 0) and negative (NES < 0) normalized enrichment scores using Gene Ontology: Factor 5 in the ovary (A), Factor 5 in the vagina (B), and Factor 6 in the uterus (C) For the ovary, only 2 top pathways with negative normalized enrichment scores appeared, so 12 top pathways with positive scores are shown. Enrichment significance was assessed using a Kolmogorov-Smirnov-like running-sum statistic (one-sided test), FDR < 0.05.
Extended Data Fig. 8 Gene expression trajectory clustering across organs.
A Gene count aging trajectories in the uterus, with Gene Ontology enrichments shown below clusters with significant enrichment. Clusters are grouped by the similarity of LOESS-smoothed trajectories (see Methods), with the thicker lines representing the average trajectory for each cluster. B Gene count aging trajectories in the vagina, with Gene Ontology enrichment displayed next to the significantly enriched cluster. Clusters are grouped by the similarity of LOESS-smoothed trajectories (see Methods), with the thicker lines representing the average trajectory for each cluster. In both A and B, Gene Ontology enrichment analysis significance was assessed using a one-sided hypergeometric test; FDR < 0.05.
Extended Data Fig. 9 Gene expression trajectory clustering across organs and GWAS overlap.
A Gene count aging trajectories in the ovary, with Gene Ontology enrichments shown below significantly enriched clusters. Gene Ontology enrichment analysis significance was assessed using a one-sided hypergeometric test; FDR < 0.05. B Heatmap for 65 age-DEGs in the uterus, ovary, and/or the myometrium overlapping with menopause-associated GWAS genes. Asterisks indicate significant age-DEGs (FDR < 0.05). Grey squares indicate missing data.
Extended Data Fig. 10 Methods section analyses.
A Regression models (clocks) trajectories for uterus, ovary and vagina, each dot corresponds to a sample, curves represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions). B Curve of training loss across epochs. C Correlation values between PEER factors and tissue proportions. D Variance explained by age in uterine tissues in the image samples that have both endometrium and myometrium. Each violin shows the distribution of per-donor means (n = 97 donors) over 1000 tiles of the tissue randomly selected once. Box plots show median, quartiles, and 1.5×IQR whiskers. E Variance explained by age by all tissues with overall tile numbers per tissue (analyses were conducted on randomly sampled 1000 tiles per tissue). Box plots show median, quartiles, and 1.5×IQR whiskers. F Variance explained by age in uterine tissues in downsampling analysis. Each violin shows the distribution of per-donor means (n = 97 donors) over 1000 tiles of the tissue randomly selected once. Box plots show median, quartiles, and 1.5×IQR whiskers. G-H Uterine tissue classification trajectories in downsampling analysis (F) and classification accuracies (G). In G, each dot corresponds to a sample and curves represent locally estimated scatterplot smoothing (LOESS) fitted mean trajectories with 95% confidence intervals around the LOESS estimate (shaded regions).
Supplementary information
Supplementary Tables (download XLSX )
Supplementary Tables 1–28.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Soldatkina, O., Ventura-San Pedro, L., Pujol-Gualdo, N. et al. Multimodal data analysis reveals asynchronous aging dynamics across female reproductive organs. Nat Aging 6, 1177–1192 (2026). https://doi.org/10.1038/s43587-026-01098-y
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s43587-026-01098-y
