
Abstract
High Mobility Group Box 1 (HMGB1) is a critical regulator of cardiac injury and repair. However, there is conflicting evidence regarding whether HMGB1 is beneficial or deleterious. We evaluated and synthesised available evidence regarding the molecular functions as well as the clinical and therapeutic utility of HMGB1 in heart disease. We found that overall, the effects depend on the redox state and subcellular location, although most studies failed to identify these parameters clearly. Nuclear upregulation or exogenous administration of fully reduced HMGB1 was beneficial. The partially oxidised form of HMGB1 (dsHMGB1) in the cytoplasm depleted intranuclear HMGB1, and extracellular dsHMGB1 was proinflammatory. Clinically, elevated circulating HMGB1 levels correlate with disease severity and may be a useful prognostic biomarker. Future research should specify the subcellular location and redox state of HMGB1. The development of methods to identify the different redox isoforms would help uncover the therapeutic potential of this multifaceted protein.
Similar content being viewed by others
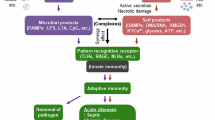

Introduction
Cardiovascular disease (CVD) is the leading cause of mortality worldwide, with ischaemic heart disease affecting ~200 million people annually1. A recent meta-analysis estimated that 3.8% of individuals aged under 60 and 9.5% of those over 60 years suffered myocardial infarction (MI) globally2. Despite advances in revascularisation strategies following MI, ~40% of patients develop heart failure (HF)3,4. An estimated 65 million adults worldwide suffer from HF1,5, and the prevalence continues to rise with an ageing population and increasing rates of obesity, diabetes, and hypertension. By 2030, the number of HF cases in the USA is projected to increase by 45%, reaching 8 million, with total direct medical costs surging to $53 billion6,7. Despite guideline-directed therapies, the 5-year survival rate for HF remains around 50%, underscoring the need for new treatments5,8.
A potential novel therapeutic avenue for improving outcomes in cardiac disease involves targeting alarmins, endogenous danger signals capable of modulating both pathological inflammation and physiological repair9. Among these, High Mobility Group Box 1 (HMGB1) is the most extensively characterised9.
HMGB1 is a highly conserved non-histone chromatin-binding protein whose function is dictated by its subcellular location and redox state10. In the nucleus, HMGB1 maintains chromatin structure and regulates DNA repair, replication, and transcription10. Under cellular or redox stress, nuclear HMGB1 translocates to the cytoplasm, primarily through modifications of nuclear localisation signals (NLS1, NLS2) via acetylation or deacetylation11,12. Other post-translational modifications, including ADP-ribosylation, phosphorylation, and methylation, also facilitate shuttling and intracellular signalling12.
HMGB1 can be actively secreted by immune cells, fibroblasts, cardiomyocytes, epithelial or endothelial cells11,12,13,14, or passively released from necrotic or damaged cells, when it functions as an alarmin9,15. The extracellular activity is determined by its redox state and corresponding receptor interactions16,17.
HMGB1 consists of two structurally homologous DNA-binding domains (Box A and Box B) and a C-terminal acidic tail (Fig. 1). The three cysteine residues (C23, C45, C106) are critical to modulating it's function16,18. The fully reduced form (frHMGB1) promotes tissue repair and chemotaxis via CXCL12-CXCR4 signalling19,20,21,22,23. However, frHMGB1 can be partially oxidised into dsHMGB1, with a disulfide bridge between C23 and C45 in the Box A domain24. The resulting conformation change modulates receptor binding. dsHMGB1 interacts with Toll-like receptor-2 (TLR-2), TLR-4, and Receptor for Advanced Glycation End-products (RAGE) to promote inflammation25 and platelet activation26.

HMGB1 consists of two DNA-binding domains, Box A and Box B, followed by a C-terminal acidic tail. In the fully reduced form (frHMGB1), thiol groups are present at C23 and C45 in Box A, and at C106 in Box-B. Oxidation results in the formation of a disulfide bridge between the cysteines at C23 and C45, yielding disulfide HMGB1 (dsHMGB1). Created in BioRender. Mao, S-H. (2026) https://BioRender.com/ltw7v97.
frHMGB1 has a half-life of ~17 min in human tissue fluids in vitro, converting entirely to dsHMGB1 within 50 min, whilst dsHMGB1 has a half-life 642 min24. In vivo, dsHMGB1 binds to haptoglobin, and following binding to CD163 on macrophages, the complex is cleared by endocytosis27. A study investigating HMGB1 dynamics in trauma patients identified two distinct peaks in the circulation28. The initial release occurred immediately following injury and declined rapidly, with a half-life of 26 min. Levels correlated with the extent of injury. This initial wave was likely due to the passive release of frHMGB1 from damaged tissue. The second peak emerged around 3 h and declined by around 6 h post-trauma, and was strongly predictive of deleterious outcomes and systemic inflammation. Although the redox states were not assessed, this secondary peak likely corresponded to the pro-inflammatory disulfide isoform (dsHMGB1)28. Fully oxidised HMGB1, when all cysteines are oxidised, lacks chemotactic or inflammatory properties16.
HMGB1 is elevated in serum and myocardial tissue in response to cardiac diseases29,30,31. Inhibition of HMGB1 has been shown to reduce inflammation and myocardial injury29,32,33,34,35,36,37,38,39 and exogenous administration can lead to worse outcomes29,32,33,34,40,41,42,43. However, HMGB1 inhibition can also be detrimental30,44, and exogenous administration or cardiac-specific overexpression may be beneficial in MI31,45,46,47 and HF48,49,50. These apparently contradictory effects have made it challenging to understand the role of HMGB1 in cardiac disorders.
The apparently contradictory effects of HMGB1 mean that in order to understand its potential as a therapeutic target, it is crucial to appreciate how its function in cardiac disease is modulated by its location (intracellular or extracellular), redox state, and specific disease context. Whilst previous reviews have highlighted conflicting data regarding these roles16,17,51,52, this scoping review with a systematic search provides a detailed update on the current understanding of HMGB1 in cardiac disorders, aiming to resolve these discrepancies by analysing and synthesising the available evidence.
Results
Overview
Our search identified 3818 articles, with 352 meeting the eligibility criteria (Fig. 2a). These studies were categorised by heart disease type, including ischaemic heart disease (IHD), HF, myocarditis, chemotherapy-induced, sepsis-induced, trauma or stress-induced, and other cardiomyopathies and cardiac disorders (Fig. 2b). Each category was subdivided into three sub-categories: HMGB1 as a mediator (Supplementary Data 1), a prognostic biomarker, and HMGB1 perturbation (Fig. 2c).

a PRISMA flowchart showing the study selection process*. b Distribution of heart diseases covered in the included studies. c Categorisation of the literature based on study type and focus. *Created using an adaptation of a web-based tool (Haddaway et al., 2022 https://doi.org/10.1002/cl2.1230).
More than half of the included studies characterised HMGB1 as a critical mediator of disease pathology (Fig. 2c). Consistently, HMGB1 levels were found to be elevated in cardiac disorders, whilst therapeutic interventions that improved clinical outcomes were associated with the downregulation of HMGB1 (Supplementary Data 1). However, most studies reported associations rather than causality. Studies reporting on HMGB1 perturbation provided mechanistic insights and others highlighted the clinical prognostic value of HMGB1 in cardiac diseases.
Ischaemic heart disease (IHD)
IHD remains the most prevalent cardiovascular disorder globally1. Coronary artery occlusion, typically driven by atherosclerotic thrombosis or embolism, precipitates myocardial ischaemia, necrosis, and inflammation53. The consequent loss of cardiomyocytes and formation of fibrotic scar tissue drive adverse ventricular remodelling and functional decline, ultimately leading to HF53. During ischaemia, HMGB1 is released passively from necrotic cells or actively translocated from the nucleus to the cytoplasm and extracellular space29,30,31. Whilst there is evidence that levels of HMGB1 in the circulation may have prognostic value54,55,56,57,58, its specific pathophysiological effects in IHD remain unclear, particularly regarding the impact of subcellular location and redox state.
HMGB1 levels in the myocardium or circulation of animals were reported as elevated following ischaemic myocardial injury, with or without reperfusion, as well as intracellularly and extracellularly in cardiomyocytes subjected to hypoxia in vitro29,34,59. Circulating levels of HMGB1 peaked at 12 h in patients and on day 1 in rats, whilst HMGB1 mRNA expression in the infarcted myocardium peaked at day 7 post-MI in rodents29,30,31.
Human clinical studies highlighted the correlation between circulating HMGB1 levels and coronary artery disease (CAD) severity and progression, particularly in acute coronary syndromes (ACS), including ST-elevation myocardial infarction (STEMI), non-STEMI (NSTEMI), and unstable angina (UA). Elevated HMGB1 levels were associated with infarct size and left ventricular ejection fraction (LVEF)54,55. HMGB1 levels also correlated with key biomarkers such as high-sensitivity C-reactive protein (hs-CRP) and cardiac troponin I (cTnI)54,56.
HMGB1 may also serve as a prognostic marker for major cardiovascular events (MACE) and mortality post-ACS. High Gensini scores and elevated IL-6, together with higher HMGB1 levels in patients with extensive coronary lesions, were predictive of adverse outcomes, including heart-related death, infarction recurrence, HF, and stroke57. Higher HMGB1 levels were observed in non-survivors of STEMI58. Multivariate analysis confirmed the prognostic value of HMGB1, and combination with hs-CRP, cTnI, and BNP enhanced early risk stratification in UA and NSTEMI patients56. Additionally, higher HMGB1 levels correlated with worse cardiopulmonary function (poor exercise tolerance and low LVEF) and angiographic evidence of the extent of atherosclerotic plaques in CAD patients. Elevated levels of HMGB1 were associated with CAD development, reinforcing its potential as a biomarker for disease progression and prognosis (Supplementary Note 3).
Taken together, the evidence indicates that HMGB1 is released into the circulation and levels positively correlate with infarct size, the extent of coronary occlusion, and clinical outcomes. However, it is important to note that the ELISA-based assays used to measure circulating levels are unable to distinguish between the two redox states. The rapid oxidation together with the longer half-life of dsHMGB1 suggests that the disulfide form likely predominates in the circulation, especially at later timepoints24,28.
Consistent with higher circulating levels being associated with worse outcomes is evidence that exogenous administration of recombinant HMGB1 (rHMGB1) increased infarct size, and was associated with elevated proinflammatory cytokine levels (e.g., TNF, IL-6, IL-18, and IL-1β) and increased cell death in rodent models of permanent ligation (PL) and ischaemia-reperfusion (IR)29,32,33,34,40,41 (Table 1). Conversely, inhibition of HMGB1 by Box A29,32,33,34, anti-HMGB1 antibodies35,36,37, immunoprecipitation41,60, HMGB1 silencing using siRNA or shRNA60,61,62,63, or glycyrrhizin34,40,59,64,65 ameliorated myocardial injury (Table 1).
Mechanistically, these pro-inflammatory effects result from the engagement of TLRs and RAGE, suggesting that the detrimental effects of HMGB1 are due to the disulfide form. Several studies reported that rHMGB1 exacerbated myocardial damage following IR via JNK/ERK/NF-κB signalling pathways32,34,40. The deleterious effects of rHMGB1 were significantly reduced in transgenic mice lacking TLR-2, TLR-4, TLR-9, or RAGE29,32,33,34,41,66. Consistent with these findings, anti-HMGB1 downregulated inflammation by limiting dendritic cell activation via the TLR-4/MyD88/NF-κB axis in an IR model in rats37. Furthermore, glycyrrhizin-mediated HMGB1 inhibition protected the myocardium from cell death (apoptosis and ferroptosis) in IR models in rats by suppressing TLR-4 signalling59,64 and inhibiting the ERK/p38 MAPK/JNK pathways65. HMGB1 has been shown to transcriptionally regulate FOXP1, a tumour suppressor gene which can modify the inflammatory response in macrophages following MI. Hypoxia resulted in increased proinflammatory cytokine expression by RAW264.7 macrophages, with concomitant increase in HMGB1 and decrease in Foxp1 expression61. This indicates that intracellular HMGB1 in macrophages and extracellular dsHMGB1 are both deleterious in IHD.
In contrast, other studies have reported that HMGB1 plays a beneficial role in MI (Table 2). HMGB1 inhibition was reported to disrupt reparative processes following MI, and animals treated with anti-HMGB1 antibodies developed larger infarcts and impaired cardiac function30,44. Furthermore, several studies have reported that exogenous administration can also be beneficial. Exogenous administration of rHMGB1 reduced infarct size, oxidative stress, and expression of proinflammatory cytokines in rodent MI models45,46,47,67,68,69,70,71,72,73. Intramyocardial injection of rHMGB1 four hours after left anterior descending (LAD) ligation significantly improved cardiac function and reduced both infarct size and fibrosis in murine and ovine models of MI45,46,47.
Several mechanisms were proposed to explain these findings, including cardioprotection, angiogenesis, and cardiac regeneration. The cardioprotective effects of HMGB1 were mediated through upregulation of HIF-1α, p38 MAPK inhibition, and activation of the PI3K/AKT pathway, leading to increased VEGF protein levels, an effect that was abolished by PI3K inhibitors69,70,71. Additionally, HMGB1 promoted autophagy through AMPK-dependent inhibition of mTOR and p-AKT, reduced apoptosis73 and improved cardiac remodelling by activating Wnt/β-catenin signalling and reducing collagen deposition68. Angiogenesis is essential for the repair of damaged myocardium and maintaining endothelial integrity. rHMGB1 enhanced angiogenesis in the border zone around the infarcted area45,46,47,72, with a concomitant upregulation of VEGF expression69. These effects were absent in TLR-9−/− mice72, suggesting angiogenesis is promoted, at least in part, through TLR-9 signalling. rHMGB1 treatment was reported to enhance cardiac regeneration by increasing the number of proliferative c-kit⁺/Ki67⁺ cells and upregulating early cardiomyogenic transcription factors, including MEF2C and NKX2-5, thereby promoting the renewal and replenishment of cardiomyocytes and cardiac progenitors45,46,47. Transcriptomic profiling at day 3 post-treatment revealed activation of regenerative processes via the Notch signalling pathway, with a 35% and 58% increase in Notch intracellular cytoplasmic domain expression in cardiomyocytes and cardiac progenitor cells, respectively74. However, cardiac regeneration in adult mammals through c-kit+ cells has now been largely discredited75.
Although the improved repair following exogenous administration is likely to be attributed to the fully reduced form, only one study clearly defined the redox state of the exogenous rHMGB1 used as frHMGB1. The authors showed that intramyocardial injection of frHMGB1 4 h after PL improved ejection fraction and other parameters of left ventricular function at one week, reduced fibrosis and left ventricle dilatation at 4 weeks via CXCL12/CXCR4 signalling21. Interestingly, a non-oxidisable HMGB1 variant (3SHMGB1), engineered by substituting the three cysteine residues (C23, C45, and C106) in frHMGB1 with serine, exacerbated adverse remodelling and impaired cardiac function following MI, partly by abolishing angiogenic effects and promoting chemotaxis of cardiac fibroblasts by signalling via CXCR4 independently of CXCL1221. These findings underscore the critical role of defining the redox state, structure, and ligand–receptor interaction of HMGB1 when considering its effects in MI.
Interestingly, the overexpression of HMGB1 yielded beneficial effects comparable to those observed with exogenous administration. Mice with cardiomyocyte-specific HMGB1 overexpression had a 30% reduction in infarct size and 90% improvement in survival rate compared to wild-type controls following MI, with enhanced angiogenesis in the border zone31. Plasma HMGB1 levels were also elevated in these mice31. This finding stands in contrast to prior reports where elevated circulating HMGB1 was consistently correlated with worse clinical outcomes. GFP-labelled bone marrow cells injected into irradiated recipient mice before MI surgery showed increased chemotaxis and differentiation of bone marrow-derived endothelial progenitor cells in the border zone of mice overexpressing HMGB148. It is unclear whether the beneficial effects of transgenic upregulation were due to higher intracellular levels of HMGB1, increased release of frHMGB1 following cell injury, or both.
In summary, the role of HMGB1 in IHD remains controversial. However, current evidence suggests that the redox state and subcellular location may act as critical determinants of its function. Effects of exogenous administration of HMGB1 are explained by downstream signalling. Only one study identified the redox form of the HMGB1 used as frHMGB1 and found that it signalled via CXCL12 and CXCR421. 3SHMGB1 signals directly via CXCR4 independently of CXCL12 and resulted in increased fibrosis21. HMGB1 constructs that signalled via TLRs or RAGE were deleterious29,32,33,34,41,66.
Distinct from MI, which is characterised by direct ischaemic necrosis, conditions such as sepsis-induced cardiomyopathy, myocarditis, and trauma- or stress-induced cardiomyopathy do not primarily involve acute ischaemic cell death. Instead, HMGB1 upregulation in these pathologies appears to be a response to stress-related stimuli, such as excessive inflammation or catecholamine surges. In these contexts, the exogenous administration of HMGB1 consistently results in the exacerbation of myocardial dysfunction. It is likely that the effects were due to dsHMGB1, as they are abrogated by the knockout of RAGE76. Interestingly, cardiomyocyte-specific conditional knockout of HMGB1 was detrimental77, suggesting that intracellular HMGB1 in these cells is protective.
Sepsis-induced cardiomyopathy
Sepsis-induced cardiomyopathy is a reversible form of cardiac dysfunction driven by the systemic cytokine storm associated with severe infection78. HMGB1 has been identified as a potential prognostic biomarker for evaluating therapeutic efficacy and monitoring disease progression79,80,81. Circulating levels inversely correlated with outcomes, and exogenous administration of rHMGB1 resulted in acute myocardial depression and impaired contractility82.
Clinical trials of Sivelestat and vitamin C reported improved cardiac outcomes and survival rates associated with lower systemic HMGB1 levels in patients with septic cardiomyopathy79,80. HMGB1 may also be useful as part of a biomarker panel to assess the risk of left ventricular diastolic dysfunction (LVDD) in septic patients when combined with cTnI and BNP, with a sensitivity of 90% and specificity of 94%81.
rHMGB1 administration to ex vivo lipopolysaccharide (LPS)-treated rat hearts induced a transient negative inotropic effect, reducing LVEF, LV +dP/dtmax, and LV −dP/dtmin, whilst increasing left ventricular end-diastolic pressure. This occurred within 1–10 min of perfusion, and the effects were reversed upon rHMGB1 withdrawal, suggesting that acute HMGB1 exposure transiently impaired cardiac contractility during endotoxemia83. Inhibition of HMGB1/TLR-4 signalling with glycyrrhizin reduced oxidative stress and cytokine release in LPS-stimulated cardiomyocytes and improved cardiac function in mice exposed to LPS82. Additionally, LPS-induced HMGB1 expression in cardiomyocytes via TLR-4-mediated PI3Kγ phosphorylation. Genetic deletion or pharmacological inhibition of PI3Kγ significantly reduced LPS-induced HMGB1 expression and secretion, thereby ameliorating depression of myocardial contractility84.
Cardiomyocyte-specific conditional deletion of HMGB1 (Hmgb1f/f TgCre/+) exacerbated myocardial injury in a model of Coxsackievirus B3 virus (CVB3)-induced myocarditis. This exacerbation was associated with increased susceptibility of HMGB1-deficient cardiomyocytes to early apoptosis via the p53-mediated Bax mitochondrial pathway, suggesting that intracellular HMGB1 in cardiomyocytes protects against the development of myocarditis77. Collectively, these data suggest that intracellular HMGB1 in cardiomyocytes is protective and that the detrimental effects were likely due to dsHMGB1 in sepsis-induced cardiomyopathies.
Autoimmune and stress-induced cardiomyopathies
Autoimmune myocarditis is associated with inflammation and stress, with concomitant upregulation of HMGB1 levels in preclinical models85. Administration of exogenous rHMGB1 resulted in increased myocardial fibrosis in murine experimental autoimmune myocarditis (EAM) through upregulation of MMP1/2 and TIMP1 in cardiac fibroblasts and myofibroblasts via the PKCβ/Erk1/2 signalling pathway86. Inhibition of HMGB1 with a neutralising antibody reduced cardiac fibrosis in EAM, accompanied by a reduction in immune infiltrates, including the autoimmunity-contributing Th17+ cells85. Inhibition of HMGB1 with an antibody or glycyrrhizin attenuated EAM, and RAGE−/− mice were protected from the development of EAM76. Given that dsHMGB1 signals via RAGE, these findings suggest that targeting the dsHMGB1–RAGE axis may be of benefit in autoimmune myocarditis.
Stress-induced cardiomyopathy is an acute cardiac dysfunction precipitated by a massive surge of catecholamines following severe physical or emotional trauma87. It is characterised by transient apical ballooning of the left ventricle, mimicking MI, but is typically reversible87. Haemorrhagic shock and resuscitation resulted in myocardial injury in animals and were associated with upregulation of HMGB188,89. Administration of anti-HMGB1 reduced cardiomyocyte apoptosis, downregulated TLR-4 and HMGB1 in the myocardium, and decreased pro-inflammatory cytokine (IL-1β, IL-6, TNF) levels88,89.
Unlike MI, which is characterised by direct cellular necrosis and the passive release of frHMGB1, acute cardiomyopathies result from cytokine or catecholamine storms. These conditions generate a pro-oxidative environment that facilitates the secretion of HMGB1 by immune and stromal cells. In this context, HMGB1 is likely to be present predominantly in the pro-inflammatory dsHMGB1, consistent with inflammatory pathology mediated through TLRs82,84,88,89 and RAGE76 signalling. Passive release of frHMGB1 from injured cardiomyocytes acts as a chemokine to recruit more immune cells, which in turn actively secrete dsHMGB1, thereby amplifying local inflammation in a feed-forward manner. Therefore, inhibition of HMGB1 is likely to be beneficial in acute cardiomyopathies.
Heart failure/adverse remodelling/hypertrophy
The prevalence of HF and chronic heart disease is rising, driven by an ageing population, increased comorbidities90, and improved survival rates following the implementation of early revascularisation strategies for MI5. In this chronically stressed myocardial environment, HMGB1 may play a role distinct from its function in acute cardiac disorders. However, as in the acute disease setting, there are conflicting accounts on its role in chronic disease, again likely explained by the subcellular location and redox state.
HMGB1 has been proposed as a prognostic biomarker for HF, aiding in monitoring disease progression and treatment outcomes. Levels correlated with established markers of HF, including NT-proBNP, and functional parameters such as left ventricular end diastolic volume (LVEDV), left ventricular end systolic volume (LVESV), LVEF, left ventricular mass index, and myocardial wall stress91,92,93,94. Circulating HMGB1 levels were elevated in HF patients and correlated with disease severity92,93. Patients with advanced HF (New York Heart Association [NYHA] stages III/IV) had significantly higher HMGB1 levels compared to those with less severe disease (NYHA I/II) or healthy controls92. Multivariate analysis confirmed HMGB1 as an independent predictor of disease severity and adverse outcomes, including necessity for heart transplantation or death92.
HMGB1 levels were also significantly increased in patients with pulmonary arterial hypertension (PAH) due to congenital heart disease and correlated with pulmonary arterial pressure and pulmonary vascular resistance. After 6 months of sildenafil therapy, HMGB1 levels in PAH patients decreased, with concomitant reduction in right atrial and ventricular diameters and pulmonary artery pressure95. However, HMGB1 levels can also be influenced by comorbidities. Patients with concurrent conditions had higher HMGB1 levels compared to those with HF alone93, highlighting the need to consider all underlying conditions when interpreting its prognostic value.
Most studies suggested that HMGB1 overexpression, particularly within the nuclei of cardiomyocytes or immune cells, was protective49,50,96,97. Exogenous HMGB1 administration demonstrated inconsistent results42,43,98,99, presumably depending on the redox state, which was not defined in any of the articles retrieved.
In a mouse model of heart failure with preserved ejection fraction (HFpEF), administration of an HMGB1 inhibitor, glycyrrhizin, resulted in attenuation of fibrosis and diastolic dysfunction, although there was no improvement in systolic function38. Mechanistically, there was decreased neutrophil infiltration and neutrophil extracellular traps, suggesting that the functional improvement was due to reduced inflammation38 (Table 3).
HMGB1 injected into the myocardium at the time of transverse aortic constriction (TAC) surgery exacerbated cardiac remodelling and reduced cardiac function over four weeks42,43. This was associated with upregulation of expression of collagen type I, TGF-β, and MMP2 in the myocardium43. Mechanistically, rHMGB1-induced stress protein expression and nuclear translocation of NFAT3, promoting hypertrophy via the PI3K/AKT/NFAT signalling pathway, which was reversed by PI3K inhibition100. The deleterious effects of intramyocardial injection of HMGB1 in a TAC model were partially antagonised by Box A42. Whilst the redox state of the rHMGB1 used was not specified, we can surmise that it was dsHMGB1.
However, the role of RAGE in mediating these processes remains unclear. HMGB1 exacerbated mechanical stress-induced cardiomyocyte hypertrophy in vitro by activating ERK1/2 through RAGE101. Nevertheless, despite elevated HMGB1 levels in the myocardium, RAGE−/− mice did not exhibit improved cardiac function compared to wild-type controls 4 weeks post-TAC. Furthermore, administration of neutralising RAGE antibody did not inhibit the effects of exogenous HMGB1-induced cardiomyocyte hypertrophy102. Whilst these deleterious effects in TAC-induced heart failure can likely be attributed to dsHMGB1, targeting RAGE alone may be insufficient, as dsHMGB1 can also activate downstream inflammatory pathways via TLR-2 and TLR-4. TLR-4−/− mice subjected to TAC showed reduced cardiac hypertrophy compared to wild-type controls103.
Other studies have reported beneficial effects of exogenous administration of HMGB1 in heart failure models (Table 4). Intramyocardial injection of HMGB1 administered 2–3 weeks following PL of the LAD in rodent models of HF significantly improved cardiac function. A 6–10% increase in LVEF was observed four weeks post-injection98,99,104. There was a concomitant increase in left ventricular wall thickness and reduced collagen deposition98,99. Cardiac regeneration and neovascularisation were suggested as potential mechanisms, evidenced by increases in new cardiomyocytes derived from c-kit+ cells and higher vascular density98. Others suggested HMGB1 modulated inflammation by reducing infiltration of inflammatory cells in the border zone, particularly OX62+ dendritic cells, via activation of ERK1/2104.
Most studies have utilised Box A as a competitive antagonist to HMGB1. However, one study reported that systemic administration of Box A fragment in MI-induced HF resulted in 15% improvement of LVEF as a result of enhanced angiogenesis and reduced fibrosis, mediated by the recruitment of PDGFRα+ mesenchymal cells from the bone marrow to the border zone105.
Subcellular location is critical to the function of HMGB1 in HF, particularly within the nucleus. Preserving nuclear HMGB1 by limiting its export to the cytoplasm has been shown to protect against the development of HF. In vitro stressing of rat neonatal cardiomyocytes resulted in acetylation of nuclear HMGB1 and export to the cytoplasm49. In the human failing heart, levels of nuclear HMGB1 negatively correlated with DNA damage. In these samples, nuclear HMGB1 expression was downregulated as the protein translocated from the nucleus to the cytoplasm50.
Overexpression of poly(ADP-ribose) polymerase 1 (PARP1) promoted nuclear export and extracellular secretion of HMGB1, contributing to pathological cardiac hypertrophy, whereas PARP1 inhibitors mitigated these effects96. Cardiomyocyte-specific HMGB1 overexpression, under the control of an α-MHC promoter, attenuated hypertrophy and improved cardiac function in TAC-induced heart failure and angiotensin II-induced hypertrophy mouse models, protecting the heart from pathological hypertrophy by reducing DNA damage49,50.
Furthermore, mice with myeloid-specific HMGB1 deletion had worse cardiac function in a TAC-induced heart failure model compared to wild-type controls97. Loss of HMGB1 in macrophages was associated with an increased ratio of M1/M2 macrophages and elevated inflammatory cytokines, indicating a shift towards proinflammatory macrophage differentiation in the myocardium. Collectively, these data suggest that increased intracellular levels of HMGB1, particularly intranuclear, are protective.
In the context of HF and adverse remodelling, the role of HMGB1 remains unclear and less well-characterised compared to MI. The impact of extracellular HMGB1 administration is particularly difficult to interpret, as none of the studies have defined the redox state of HMGB1 used42,43,98,99,104. However, based on the downstream receptors, it can be deduced that deleterious effects in models of heart failure can likely be attributed to dsHMGB1. From the available evidence, it is not possible to use a similar approach to deduce that the beneficial effects were due to frHMGB1. In contrast, the upregulation of nuclear HMGB1 expression appears consistently protective49,96, underscoring the critical effect of subcellular location. Mechanistically, whilst HMGB1-induced angiogenesis is well-supported, other proposed reparative pathways, such as the differentiation of c-kit+98 or bone marrow-derived cells105 to replace dead myocardium, have been largely discounted75,106, with evidence suggesting that the injection of killed cells can yield similar non-specific effects. Consequently, resolving these discrepancies requires a rigorous definition of the redox state and reappraisal of the mechanism of action.
Diabetic cardiomyopathy
Diabetes is associated with the development of cardiomyopathy through exacerbation of inflammation, endoplasmic reticulum stress and cellular death, leading to increased fibrosis and cardiac dysfunction39,107,108,109. There was concurrent upregulation of cytoplasmic translocation of nuclear HMGB1 in cardiomyocytes39,107,108,109.
Metabolic stress induced by a high-fat, high-glucose diet in mice triggered the expression of CCAAT/enhancer-binding protein homologous protein (CHOP), a transcriptional regulator of cardiac HMGB1. Mice treated with a high-fat and high-glucose diet showed increased expression of acetylated HMGB1 in cardiomyocytes. Exogenous administration of rHMGB1 promoted M1 macrophage polarisation in vitro, especially in media with high fat and high glucose. Treatment with glycyrrhizin improved cardiac function and reduced immune infiltration in the myocardium of mice with diabetic cardiomyopathy107. Inhibition of HMGB1 by shRNA or neutralising antibody downregulated the expression of MMP-2 and MMP-9 in cardiac fibroblasts under high-glucose conditions. This inhibition was associated with a reduction in hyperglycaemia-induced activation of E26 transformation-specific-1 (Ets-1) through ERK1/2 signalling. Inhibiting Ets-1 also reduced apoptosis in the myocardium of mice with diabetic cardiomyopathy. Collectively, these findings indicate that HMGB1 inhibition reduces apoptosis and MMP expression in cardiac fibroblasts via the ERK-dependent Ets-1 pathway39.
Exogenous rHMGB1 promoted cardiac fibroblast proliferation and migration under conditions of normal glucose, comparable to cells incubated in high glucose109. High glucose-induced upregulation of HMGB1 in cardiomyocytes was associated with a downregulation of IL-33 and an increase in collagen production by cardiac fibroblasts110. Administration of Box A or exogenous IL-33 reduced collagen deposition and attenuated cardiac dysfunction in diabetic mice. Additionally, TLR-4−/− diabetic mice exhibited higher myocardial HMGB1 expression without a corresponding increase in fibrosis and downregulation of IL-33 in the fibroblasts, suggesting that HMGB1 regulates fibrosis through the TLR-4/IL-33 axis110.
Taken together, the data show that diabetic cardiomyopathy is associated with elevated levels of cytoplasmic HMGB1, leading to ER stress and apoptosis. rHMGB1 enhanced M1 polarisation, inflammation, and fibrosis, whilst inhibition of HMGB1 improved function. Based on the downstream signalling pathways (TLR-4), it is likely that the deleterious effects of HMGB1 in diabetic cardiomyopathy can be attributed to the disulfide form.
Chemotherapy-induced cardiomyopathy
Chemotherapy agents induced cellular stress and damage, with concomitant upregulation of HMGB1, leading to activation of programmed cell death111,112,113,114,115,116. The majority of studies showed that antagonising HMGB1 reduced cell death.
HMGB1 has been reported to play a pivotal role in cell death associated with chemotherapy-induced cardiotoxicity and cardiomyopathy, whilst antagonising HMGB1 has been shown to be beneficial. Inhibition, including administration of Box A, silencing of HMGB1 with shRNA, or TLR-4 knock-out, resulted in reduced apoptosis of cardiomyocytes exposed to doxorubicin111. Silencing HMGB1 or inhibition with glycyrrhizin downregulated autophagy markers LC3II and p62, leading to improved cardiomyocyte viability112,113. Inhibition of HMGB1 attenuated maladaptive autophagy through downregulation of the Akt/mTOR pathway112. Moreover, overexpression of Yes-Associated Protein (YAP)/transcriptional coactivator downregulated HMGB1 expression, suggesting a regulatory role for YAP113.
Ferroptosis has also been implicated in doxorubicin-induced cardiomyopathy. Administration of the iron chelator, dexrazoxane, attenuated doxorubicin-induced cardiotoxicity115,116. Overexpression of HMGB1 reduced the efficacy of dexrazoxane by promoting ferroptosis and oxidative stress in doxorubicin-induced cardiotoxicity, whereas silencing HMGB1 had the opposite effect115. However, the authors induced HMGB1 expression via intravenous lentiviral delivery rather than using a cardiac-specific approach. This likely resulted in systemic overexpression across multiple cell types, thereby complicating the interpretation of the data. HMGB1 promoted inflammation during ferroptosis via immune cell activation. Depletion of neutrophils reduced the formation of neutrophil extracellular traps (NETs), downregulated HMGB1 expression, and attenuated ferroptosis, thereby improving cardiac function in doxorubicin-induced cardiomyopathy. The administration of rHMGB1 partially reversed the effect of neutrophil depletion through the YAP/TLR-4 axis116, suggesting that the effects were due to the disulfide form.
Dasatinib, a tyrosine kinase inhibitor used for treating chronic myeloid leukaemia, has been associated with significant cardiovascular toxicity, mainly through the induction of necroptosis in cardiomyocytes via intracellular activation of HMGB1. Genetic deletion of HMGB1 through CRISPR-Cas9 knockout increased the survival of cardiomyocytes exposed to dasatinib114.
Mice with cardiac-specific (α-MHC promoter) overexpression of HMGB1 exhibited reduced doxorubicin-induced cardiotoxicity compared to wild-type controls. Silencing HMGB1 in cardiomyocytes decreased mitochondrial membrane potential, which was partially restored with HMGB1 transfection. HMGB1 promoted cardiomyocyte survival by upregulating heat shock protein beta-1 (HSPB1) through activation of the heat shock element promoter. Silencing HSPB1 and heat shock factor 2 abrogated the protective effects of HMGB1. The results show that overexpression of HMGB1 in cardiomyocytes protected against doxorubicin-induced cardiotoxicity by restoring mitochondrial function through modulation of HSPB1117.
Other cardiomyopathies/cardiac disorders
HMGB1 has been shown to exert detrimental effects in congenital dilated cardiomyopathy, metabolism-, ageing-, cancer-associated cardiomyopathy and arrhythmias. This literature is summarised in Supplementary Note 4.
The majority of studies suggest that HMGB1 exerts a detrimental role in the progression of chronic heart disease, as its inhibition attenuates cellular stress and damage, largely via TLR-4 signalling, suggesting that these effects are mediated by dsHMGB1110,111,116. Although cardiac-specific overexpression of HMGB1 has generally demonstrated protective effects in doxorubicin-induced cardiotoxicity117, other studies report that HMGB1 overexpression that is not restricted to cardiomyocytes can induce cell death via ferroptosis and oxidative stress115. Furthermore, the knockout of HMGB1 has been shown to improve the survival of cardiomyocytes treated with dasatinib114. Taken together, the studies on cardiomyopathies suggest that the role of HMGB1 in cardiomyopathies depends on the redox state, cellular location, as well as disease context.
Discussion
The multifaceted role of HMGB1 is determined by subcellular location and redox state. The redox state dictates the downstream signalling—frHMGB1 forms a heterocomplex with CXCL12 and signals via CXCR419,20,21,22,23, whilst dsHMGB1 signals via RAGE, TLR-2, and TLR-425. A summary of the mechanisms of action of HMGB1 in cardiac diseases, depending on subcellular location and redox state, is illustrated in Fig. 3.

HMGB1 is a non-histone chromatin-binding protein in the nucleus. Under stress conditions or following injury, HMGB1 can be passively released into the extracellular space or transported to the cytoplasm and then actively secreted into the extracellular space by packing into intracellular vesicles (lysosomes or autophagosomes) and fusing with the cytoplasmic membrane12. HMGB1 can undergo post-translational modifications (PTM) affecting nuclear localisation signals (NLS) and nuclear export signal (NES) to facilitate translocation of nuclear HMGB1 to the cytoplasm11,12. These PTMs include acetylation, phosphorylation, methylation, N-glycosylation, and oxidation. The redox state of HMGB1 modulates function both intra- and extracellularly25,118,119. Conformational changes as a result of the disulfide bridge between C23 and C45 alter the affinity of HMGB1 for intracellular shuttling proteins and extracellular receptors. Intracellularly, upregulation of nuclear HMBG1 has been shown to be beneficial. The function of cytoplasmic HMGB1 depends on post-translational modifications and is not fully understood. Extracellular HMGB1 acts as an alarmin. frHMGB1 forms a heterocomplex with CXCL12 and signals via CXCR4 to promote chemotaxis and tissue repair. dsHMGB1 signals via TLR-2, TLR-4, and RAGE to promote the release of pro-inflammatory cytokines. The dsHMGB1–RAGE axis also activates platelets. Various strategies have been explored to inhibit HMGB1, including Box A, glycyrrhizin, anti-HMGB1 antibodies, or small interfering RNA. Box A acts as a competitive inhibitor of HMGB1, whereas glycyrrhizin or neutralising antibodies bind to the HMGB1. However, none of these approaches can target a specific redox form of HMGB1. Created in BioRender. Mao, S-H. (2026) https://BioRender.com/scr20ip frHMGB1 fully reduced HMGB1, dsHMGB1disulfide HMGB1.
The subcellular location of HMGB1 is a critical determinant of its function. Whilst the protective role of nuclear HMGB1 is increasingly recognised42,49,50,96,117, the specific intracellular functions of HMGB1 and the impact of post-translational modifications in cardiac diseases remain poorly understood. The available evidence suggests that maintaining nuclear HMGB1 is beneficial, with the inhibition of nuclear-to-cytoplasmic translocation reducing myocardial damage in heart failure96. Furthermore, reduced nuclear HMGB1 expression has been observed in murine pressure overload models and in human heart failure, particularly following the onset of decompensation, indicating that the loss of nuclear HMGB1 may contribute to disease progression49.
Physiologically, newly synthesised HMGB1 is predominantly transported into the nucleus, reinforcing the concept that upregulation of the nuclear pool is cardioprotective. However, under stress conditions, HMGB1 undergoes post-translational modifications, such as acetylation or oxidation, which facilitate its translocation to the cytoplasm12,42. Intracellular oxidation of frHMGB1 to dsHMGB1 promotes translocation from the nucleus to the cytoplasm118 and sustains autophagy through interaction with Beclin1119. Yet only a few studies in the context of cardiac disease have characterised the differential expression of HMGB1 in the nucleus versus the cytoplasm49,96. Given the importance of cytosolic HMGB1 in determining cell survival119, further research is required to elucidate the specific cytoplasmic functions and post-translational modifications of HMGB1 in cardiac pathology.
Genetic deletion of HMGB1 has also been shown to be harmful in both physiological and pathological states, whereas its overexpression is mostly protective. Global HMGB1 deletion (Hmgb1−/−)-induced lethal hypoglycemia in mice120, whilst mice with cardiomyocyte-specific conditional deletion (cTnICre/+−Hmgb1fl/fl) exhibited downregulated glucocorticoid receptor and PGC1-α signalling, leading to upregulation of glycolytic metabolism and impairment of heart development121. Conditional deletion of HMGB1 exacerbated CVB3-induced viral myocarditis77.
Conversely, overexpression of endogenous HMGB1 reduced DNA and myocardial damage in several disease models48,49,50,97,117. Whilst some studies reported that HMGB1 overexpression via viral vector transfection was deleterious61,62,76,115, this may have resulted from the use of non-native promoters that drive excessive protein expression, potentially leading to disrupted molecular interactions, mis-localisation, and cellular toxicity122. However, gene knock-in at the native locus ensures physiologically regulated expression, maintaining appropriate feedback mechanisms, post-translational modifications, and physiological subcellular location122, resulting in improved outcomes48,49,50,97,117.
A crucial aspect of HMGB1 biology is its redox state25. Although the majority of studies did not specify the redox state, it can be inferred based on receptor interactions and downstream signalling pathways. Extracellular frHMGB1 exclusively signals via CXCL12/CXCR418,22,23, whereas dsHMGB1 binds TLR-2, TLR-4 and RAGE18,25. There is no overlap between frHMGB1 and dsHMGB1 signalling25. dsHMGB1 triggers a proinflammatory cascade, leading to the upregulation of proinflammatory cytokines25 and platelet activation26, which can exacerbate myocardial damage. Conversely, frHMGB1 promotes chemotaxis and repair19,22 (Fig. 3). frHMGB1 has also been reported to accelerate tissue repair of skeletal muscle, bone, and blood by transitioning tissue-resident stem cells from G0 to GAlert19.
Investigations into the effect of exogenous administration of rHMGB1 in IHD and HF yielded mixed results, likely dependent on the redox state of the preparation used. The only publication that clearly defined the redox state of the rHMGB1 reported that frHMGB1 signalling via CXCL12/CXCR4 was beneficial following MI21. The detrimental effects of rHMGB1 were largely associated with the activation of TLR-2, TLR-4, and RAGE signalling37,59,64,82,88,89,101,116, implicating dsHMGB1 as the primary mediator. Deletion of these receptors abolished the adverse effects of HMGB1, further supporting the notion that dsHMGB1 was used in these experiments29,32,33,34,41,66,110,111.
We attempted to clarify the redox state of the rHMGB1 when not stated in the publication by contacting the suppliers described in the materials (Tables 1–4). The majority responded that the redox status of their product was unknown. Only one supplier confirmed that one of their four preparations of the HMGB1 molecule was dsHMGB1 (1690-HMB, R&D), but the redox state of the remaining preparations could not be identified. Interestingly, four studies using HMGB1 from R&D Systems all reported beneficial effects in animals with acute MI or MI-induced HF71,72,99,104; however, none of these studies reported the product code or the redox state. The downstream signalling pathways reported included TLR-972, TGF-β-Smad, ERK1/2, and HIF-1α/PI3K/AKT71,99,104, but none of them reported signalling via CXCL12-CXCR4, TLR-2, TLR-4 or RAGE71,72,99,104. Thus, we could not infer the redox state based on the information provided.
Other studies, despite not specifying the redox state, reported favourable outcomes following HMGB1 administration through upregulation of recruitment of bone marrow cells48,105, angiogenesis45,46,47,69,72,98, myocyte regeneration via c-kit+ stem cells45,46,47,74,98, and improvement in remodelling98,99,104,105 in MI and HF models. The contribution of c-kit+ cardiac stem cells to the repair of the adult mammalian heart has subsequently been largely discounted75. Interestingly, a non-oxidisable HMGB1 variant (3SHMGB1), engineered by substituting three conserved cysteine residues (C23, C45, and C106) with serine, exacerbated adverse remodelling and impaired cardiac function following MI, partly by abolishing angiogenic effects and promoting chemotaxis of cardiac fibroblasts via CXCR4, independent of CXCL1221. This contrasts with other tissues, including bone, blood, skeletal muscle, and liver, where the administration of 3SHMGB1 was beneficial19,20. These findings underscore the critical role of defining the ligand-receptor interaction as well as the redox state of HMGB1 when considering its effects on different tissues and cells.
None of the HMGB1 inhibition approaches selectively targeted either frHMGB1 or dsHMGB1. Glycyrrhizin binds to Box A or Box B, and dsHMGB1 had a higher binding affinity to glycyrrhizin compared to frHMGB1, resulting in a more stable glycyrrhizin–dsHMGB1 complex123,124. This could explain the beneficial effect of glycyrrhizin38,40,59,64,65. Monoclonal or polyclonal antibodies bind circulating HMGB1 without differentiating between the two redox forms25. Box A competitively inhibits HMGB1 by binding to TLR-4 and RAGE125,126, and also inhibits signalling via CXCL12/CXCR425. Although the specific redox form of HMGB1 neutralised by antagonists remains unclear, dsHMGB1 may be predominantly affected in conditions characterised by chronic inflammation. Furthermore, the beneficial effect of inhibiting HMGB1 were largely associated with TLR-2/TLR-4/RAGE and their downstream signalling pathways29,32,33,34,37,41,59,64,66,84,89,101,102,108,110,127,128,129. However, non-selective inhibition of HMGB1 may also counteract the beneficial effects of frHMGB1, potentially worsening adverse remodelling30,44.
Modulation of HMGB1 in IHD29,32,35,45,46,47,67,73,130, HF42,43,49,98,99,102,104, myocarditis76,77,86, and chemotherapy-induced cardiomyopathy111,112,113,114,115,116,117 yielded conflicting results. However, in cardiomyopathies induced by diabetes39,107,108,109,110, trauma, stress88,89, or sepsis82,84, HMGB1 was consistently detrimental. In diabetic cardiomyopathy, chronic metabolic stress upregulates HMGB1, promoting inflammation and myocardial fibrosis. Although mice with cardiomyocyte-specific HMGB1 overexpression exhibited a reduced infarct size following MI31, mice with cardiomyocyte-specific HMGB1 overexpression with a streptozotocin-induced diabetic phenotype had larger infarcts and lower plasma HMGB1 levels compared to wild-type controls131. This suggests that the diabetic environment and trauma-, stress- or sepsis-induced cardiomyopathies may alter the functions of HMGB1, potentially by favouring the disulfide form.
The different effects of HMGB1 between disease states highlight the importance of distinct disease contexts and progression patterns. The available evidence supports the pivotal effect of the redox state of HMGB1. However, direct evidence is lacking due to the inability to differentiate frHMGB1 and dsHMGB1 in vivo and the failure to report the redox state of HMGB1. It is possible that in IHD, acute ischaemia results in cell necrosis, leading to passive release of frHMGB1 from the nucleus. However, prolonged ischaemia or excessive redox stress creates a microenvironment that is acidic due to anaerobic metabolism and lactate accumulation, which is likely to promote conversion of extracellular or intracellular frHMGB1 to dsHMGB1, leading to persistent inflammation, fibrosis, and adverse remodelling25.
HMGB1 is released actively under stress conditions and passively from damaged or necrotic cells. Circulating HMGB1 levels correlated positively with disease progression and severity in IHD, HF, and myocarditis54,55,56,57,58,92,93. Therapeutic interventions that improved cardiac health in both clinical and preclinical studies were associated with lower HMGB1 levels79,80 (Supplementary Data 1). However, HMGB1 is expressed in all nucleated cells, and thus, release is not limited to cardiac pathology, making it unsuitable for diagnostic purposes due to poor specificity as a result of its high sensitivity to comorbidities and injuries93 (Supplementary Note 3). Combining HMGB1 with other biomarkers (e.g. hsCRP, cTnI, NT-proBNP) may enhance its prognostic value56,81,91,92,93,94.
Despite the extensive literature, our understanding of the role of HMGB1 in cardiac pathologies remains incomplete. Firstly, some of the mechanisms of action described were not robustly verified. For example, HMGB1 was reported as promoting regeneration via resident c-kit+ cardiac stem cells45,98,130. However, the role of this mechanism has been questioned given the extremely limited contribution of these cells to cardiomyocyte renewal in adult mammals75. More sophisticated models, such as clonal expansion models using confetti mice132 or FUCCI (fluorescent ubiquitination-based cell cycle indicator) reporter mice133, are required to investigate this hypothesis further.
Second, distinguishing dsHMGB1 from frHMGB1, especially in vivo18,25, remains a significant challenge, limiting our understanding of the intracellular and extracellular oxidation dynamics. Redox-sensitive mass spectrometry approaches using differential cysteine alkylation could enable direct discrimination of HMGB1 redox isoforms in cardiac tissue and plasma. The development and validation of redox-state-specific HMGB1 antibodies or aptamers may provide an additional approach, although sensitivity and specificity could be limiting factors.
Finally, the timing and duration of HMGB1 release during cardiac events are still poorly defined. The temporal dynamics of HMGB1 release can determine whether it exerts protective or detrimental effects42, emphasising the importance of characterising how and when HMGB1 is released, and the redox state during different phases of cardiac injury. High-resolution time-course analyses of circulating and myocardial HMGB1 levels following injury may help define phase-specific functions. In parallel, cell-specific HMGB1 reporter or conditional knockout models could clarify the relative contribution of cardiomyocyte versus immune-derived HMGB1 over time, whilst single-cell and spatial transcriptomic approaches may provide complementary insight into the temporal and spatial context of HMGB1 signalling.
In conclusion, this review highlights the importance of HMGB1 in cardiac disorders. Its role is determined by its location and redox state. Intracellular HMGB1, particularly the nuclear fraction, is largely protective. However, its role following translocation to the cytoplasm and the effect of post-translational modification remain unclear. Cytoplasmic dsHMGB1 appears detrimental by facilitating nuclear export, thereby depleting the protective nuclear pool. Extracellularly, HMGB1 effects are dependent on its redox state: frHMGB1 exerts beneficial effects via the CXCL12–CXCR4 axis, whereas dsHMGB1 acts as a proinflammatory mediator through TLR-2, TLR-4, and RAGE signalling. To reconcile these apparently conflicting findings, more rigorous reporting of HMGB1 redox state and subcellular location is required. In addition, there is a pressing need to develop reliable methods for distinguishing between fully reduced HMGB1 (frHMGB1) and disulfide HMGB1 (dsHMGB1) in both tissue samples and circulation. These advances will be essential to realise the translational potential of targeting HMGB1-associated pathways for therapeutic benefit in cardiac disease.
Methods
Protocol and registration
This scoping review followed the Joanna Briggs Institute (JBI) methodology134 and was registered with the Open Science Framework (OSF) under their Scoping Review Protocol Guidance (https://osf.io/ajg9d/files/osfstorage/675575878fb4f95587563d2b). Results were reported according to the PRISMA extension for scoping reviews, ensuring structured reporting, transparency, and reproducibility (Supplementary Note 1)135.
Eligibility criteria
Primary research studies investigating the role of HMGB1 in heart diseases were included. The exclusion criteria were as follows:
-
Non-primary research, including correspondence, letters to the editor, editorials, book chapters, reviews, and conference abstracts.
-
Studies that do not address HMGB1 as a primary or secondary focus.
-
Studies without full-text availability or retracted.
-
Studies unrelated to heart diseases, including those focused on cerebrovascular conditions, vascular diseases, or cardiac surgery.
The inclusion and exclusion criteria were confirmed by both primary reviewers before the formal screening process. Any disagreements regarding eligibility were resolved through discussion or consultation with a senior reviewer.
Information sources
The search strategy encompassed three databases: Embase (via Ovid), Medline (via Ovid), and Web of Science Core Collection, searched separately without restrictions on date or language. The search terms included “HMGB1” and “heart diseases.” The strategy was developed collaboratively by the lead author (S.H.M.) and an academic librarian (K.S.). A full electronic search strategy was provided in Supplementary Note 2. The search was conducted on 8 October 2024. Citation management and deduplication were handled using EndNote and Covidence, and screening was performed in Rayyan.
Selection of sources of evidence
Two independent reviewers evaluated all retrieved citations based on the pre-specified eligibility criteria using Rayyan. Full-text reviews were conducted for citations that meet the inclusion criteria or for undecidable abstracts. Reasons for exclusion at the full-text stage were documented. Conflicts were resolved through discussion or consultation with a senior reviewer. The outcomes of the search and study selection process were detailed in the scoping review and presented in a PRISMA flow diagram (Fig. 2).
Data charting process
Data were extracted independently by two reviewers, focusing on study characteristics, outcomes, key findings, and proposed mechanisms relevant to the scoping review questions related to HMGB1 and heart diseases. A pilot extraction of ~10 articles was conducted by the lead author to refine the extraction tool. Extracted data were stored in Rayyan or Excel. Discrepancies between reviewers were documented and resolved through discussion or consultation with a senior reviewer. Adjustments were recorded in the final report to ensure transparency and consistency in the data extraction process.
Synthesis of results
The results were presented using a PRISMA flow diagram. Data were summarised in graphical, diagrammatic, or tabulated formats, depending on the characteristics of the data. A narrative summary accompanied these visualisations, describing how the results align with the study objectives.
Data availability
All relevant data extracted and analysed during this study are included in this published article (and its supplementary information and data files). Additional raw extraction sheets used during the scoping process are available from the corresponding author upon reasonable request.
References
Organization, W. H. The Top 10 Causes of Death. World Health Organization.
Salari, N. et al. The global prevalence of myocardial infarction: a systematic review and meta-analysis. BMC Cardiovasc. Disord. 23, 206 (2023).
Hung, J. et al. Trends from 1996 to 2007 in incidence and mortality outcomes of heart failure after acute myocardial infarction: a population-based study of 20,812 patients with first acute myocardial infarction in Western Australia. J. Am. Heart Assoc. 2, e000172 (2013).
Sulo, G. et al. Heart failure complicating acute myocardial infarction; burden and timing of occurrence: a Nation-wide Analysis Including 86 771 patients from the Cardiovascular Disease in Norway (CVDNOR) Project. J. Am. Heart Assoc. 5, https://doi.org/10.1161/JAHA.115.002667 (2016).
Gerber, Y. et al. A contemporary appraisal of the heart failure epidemic in Olmsted County, Minnesota, 2000 to 2010. JAMA Intern. Med. 175, 996–1004 (2015).
Heidenreich, P. A. et al. Forecasting the impact of heart failure in the United States: a policy statement from the American Heart Association. Circ. Heart Fail. 6, 606–619 (2013).
Ziaeian, B. & Fonarow, G. C. Epidemiology and aetiology of heart failure. Nat. Rev. Cardiol. 13, 368–378 (2016).
Bozkurt, B. Contemporary pharmacological treatment and management of heart failure. Nat. Rev. Cardiol. 21, 545–555 (2024).
Chan, J. K. et al. Alarmins: awaiting a clinical response. J. Clin. Investig. 122, 2711–2719 (2012).
Kang, R. et al. HMGB1 in health and disease. Mol. Asp. Med. 40, 1–116 (2014).
Bonaldi, T. et al. Bonaldi-et-al-2003-monocytic-cells-hyperacetylate-chromatin-protein-hmgb1-to-redirect-it-towards-secretion.pdf. EMBO J. 22, 5551–5560 (2003).
Chen, R., Kang, R. & Tang, D. The mechanism of HMGB1 secretion and release. Exp. Mol. Med. 54, 91–102 (2022).
Lee, W. J. et al. Profibrogenic effect of high-mobility group box protein-1 in human dermal fibroblasts and its excess in keloid tissues. Sci. Rep. 8, 8434 (2018).
Pisetsky, D. S. The expression of HMGB1 on microparticles released during cell activation and cell death in vitro and in vivo. Mol. Med. 20, 158–163 (2014).
Scaffidi, P., Misteli, T. & Bianchi, M. E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 418, 191–195 (2002).
Raucci, A. et al. The Janus face of HMGB1 in heart disease: a necessary update. Cell. Mol. Life Sci. 76, 211–229 (2019).
Wasim, R. et al. High Mobility Group Box 1 and cardiovascular diseases: study of act and connect. Cardiovasc. Toxicol. 24, 1268–1286 (2024).
Viñals Guitart, Á. et al. https://doi.org/10.1101/2024.11.06.622383 (2024).
Lee, G. et al. Fully reduced HMGB1 accelerates the regeneration of multiple tissues by transitioning stem cells to GAlert. Proc. Natl. Acad. Sci. USA 115, E4463–E4472 (2018).
Tirone, M. et al. High Mobility Group Box 1 orchestrates tissue regeneration via CXCR4. J. Exp. Med. 215, 303–318 (2018).
Di Maggio, S. et al. Non-oxidizable HMGB1 induces cardiac fibroblasts migration via CXCR4 in a CXCL12-independent manner and worsens tissue remodeling after myocardial infarction. Biochim. Biophys. Acta 1863, 2693–2704 (2017).
Schiraldi, M. et al. HMGB1 promotes recruitment of inflammatory cells to damaged tissues by forming a complex with CXCL12 and signaling via CXCR4. J. Exp. Med. 209, 551–563 (2012).
Mantonico, M. V. et al. The acidic intrinsically disordered region of the inflammatory mediator HMGB1 mediates fuzzy interactions with CXCL12. Nat. Commun. 15, 1201 (2024).
Zandarashvili, L. et al. Real-time kinetics of high-mobility group box 1 (HMGB1) oxidation in extracellular fluids studied by in situ protein NMR spectroscopy. J. Biol. Chem. 288, 11621–11627 (2013).
Venereau, E. et al. Mutually exclusive redox forms of HMGB1 promote cell recruitment or proinflammatory cytokine release. J. Exp. Med. 209, 1519–1528 (2012).
Stark, K. et al. Disulfide HMGB1 derived from platelets coordinates venous thrombosis in mice. Blood 128, 2435–2449 (2016).
Yang, H. et al. Identification of CD163 as an antiinflammatory receptor for HMGB1–haptoglobin complexes. JCI Insight 3, https://doi.org/10.1172/jci.insight.126617 (2018).
Ottestad, W. et al. Biphasic release of the alarmin High Mobility Group Box 1 protein early after trauma predicts poor clinical outcome. Crit. Care Med. 47, e614–e622 (2019).
Volz, H. C., Kaya, Z., Katus, H. A. & Andrassy, M. The role of HMGB1/RAGE in inflammatory cardiomyopathy. Semin. Thromb. Hemost. 36, 185–194 (2010).
Kohno, T. et al. Role of high-mobility group box 1 protein in post-infarction healing process and left ventricular remodelling. Cardiovasc. Res. 81, 565–573 (2009).
Kitahara, T. et al. High-mobility group box 1 restores cardiac function after myocardial infarction in transgenic mice. Cardiovasc. Res. 80, 40–46 (2008).
Andrassy, M. et al. High-mobility group box-1 in ischemia-reperfusion injury of the heart. Circulation 117, 3216–3226 (2008).
Mersmann, J. et al. Attenuation of myocardial Injury by HMGB1 blockade during ischemia/reperfusion is toll-like receptor 2-dependent. Mediators Inflamm. 2013, https://doi.org/10.1155/2013/174168 (2013).
Xu, H. et al. Endogenous HMGB1 contributes to ischemia-reperfusion-induced myocardial apoptosis by potentiating the effect of TNF-a/JNK. Am. J. Physiol. Heart Circ. Physiol. 300, H913–H921 (2011).
Hu, X. R., Zhang, K., Chen, Z. Q., Jiang, H. & Xu, W. P. The HMGB1-IL-17A axis contributes to hypoxia/reoxygenation injury via regulation of cardiomyocyte apoptosis and autophagy. Mol. Med. Rep. 17, 336–341 (2018).
Xia, J. et al. [The role of high-mobility group box protein 1 in the signaling pathways of myocardial ischemia-reperfusion injury in rats]. Zhonghua yi xue za zhi 98, 3268–3273 (2018).
Xue, J. Y. et al. The role of dendritic cells regulated by HMGB1/TLR4 signalling pathway in myocardial ischaemia reperfusion injury. J. Cell. Mol. Med. 23, 2849–2862 (2019).
Zhang, X. L. et al. HMGB1-promoted neutrophil extracellular traps contribute to cardiac diastolic dysfunction in mice. J. Am. Heart Assoc. 11, https://doi.org/10.1161/JAHA.121.023800 (2022).
Wang, W.-K. et al. HMGB1 mediates hyperglycaemia-induced cardiomyocyte apoptosis via ERK/Ets-1 signalling pathway. J. Cell. Mol. Med. 18, 2311–2320 (2014).
Zhai, C. L. et al. Glycyrrhizin protects rat heart against ischemia-reperfusion injury through blockade of HMGB1-dependent phospho-JNK/Bax pathway. Acta Pharmacol. Sin. 33, 1477–1487 (2012).
Tian, Y. K. et al. The myocardial infarct-exacerbating effect of cell-free DNA is mediated by the high-mobility group box 1-receptor for advanced glycation end products-Toll-like receptor 9 pathway. J. Thorac. Cardiovasc. Surg. 157, 2256 (2019).
Zhang, L. et al. Extracellular high-mobility group box 1 mediates pressure overload-induced cardiac hypertrophy and heart failure. J. Cell. Mol. Med. 20, 459–470 (2016).
Zhang, L. et al. HMGB1 aggravates pressure overload-induced left ventricular dysfunction by promoting myocardial fibrosis. Int. J. Hypertens. 2020, https://doi.org/10.1155/2020/7270351 (2020).
Oozawa, S. et al. Effects of HMGB1 on ischemia-reperfusion injury in the rat heart. Circ. J. 72, 1178–1184 (2008).
Limana, F. et al. Exogenous high-mobility group box 1 protein induces myocardial regeneration after infarction via enhanced cardiac c-kit+ cell proliferation and differentiation. Circ. Res. 97, E73–E83 (2005).
He, Y. Y., Wen, Y., Zheng, X. X. & Jiang, X. J. Intramyocardial delivery of HMGB1 by a novel thermosensitive hydrogel attenuates cardiac remodeling and improves cardiac function after myocardial infarction. J. Cardiovasc. Pharmacol. 61, 283–290 (2013).
Bauzá, M. D. et al. High-dose intramyocardial HMGB1 induces long-term cardioprotection in sheep with myocardial infarction. Drug Deliv. Transl. Res 9, 935–944 (2019).
Nakamura, Y. et al. High Mobility Group Box 1 promotes angiogenesis from bone marrow-derived endothelial progenitor cells after myocardial infarction. J. Atheroscler. Thromb. 22, 570–581 (2015).
Funayama, A. et al. Cardiac nuclear High Mobility Group Box 1 prevents the development of cardiac hypertrophy and heart failure. Cardiovasc. Res. 99, 657–664 (2013).
Takahashi, T. et al. Cardiac nuclear High-Mobility Group Box 1 ameliorates pathological cardiac hypertrophy by inhibiting DNA damage response. JACC-Basic Transl. Sci. 4, 234–247 (2019).
Foglio, E., Pellegrini, L., Russo, M. A. & Limana, F. HMGB1-mediated activation of the inflammatory-reparative response following myocardial infarction. Cells 11, https://doi.org/10.3390/cells11020216 (2022).
Pellegrini, L. et al. HMGB1 and repair: focus on the heart. Pharm. Ther. 196, 160–182 (2019).
Frangogiannis, N. G. Pathophysiology of myocardial infarction. Compr. Physiol. 5, 1841–1875 (2015).
Yao, H. C. et al. Correlation between serum high-mobility group box-1 levels and high-sensitivity C-reactive protein and troponin I in patients with coronary artery disease. Exp. Ther. Med. 6, 121–124 (2013).
Andrassy, M. et al. HMGB1 as a predictor of infarct transmurality and functional recovery in patients with myocardial infarction. J. Intern. Med. 270, 245–253 (2011).
Hashimoto, T. et al. Circulating high-mobility group box 1 and cardiovascular mortality in unstable angina and non-ST-segment elevation myocardial infarction. Atherosclerosis 221, 490–495 (2012).
He, W. et al. IL-6 and HMGB1 levels for predicting major adverse vascular events after percutaneous coronary intervention in patients with acute coronary syndrome. Heart Surg. Forum 27, E960–E967 (2024).
Sorensen, M. V., Pedersen, S., Mogelvang, R., Skov-Jensen, J. & Flyvbjerg, A. Plasma high-mobility group box 1 levels predict mortality after ST-segment elevation myocardial infarction. JACC-Cardiovasc. Interv. 4, 281–286 (2011).
Yuan, Y. G., Li, B., Peng, W. Z. & Xu, Z. S. Protective effect of glycyrrhizin on coronary microembolization-induced myocardial dysfunction in rats. Pharmacol. Res. Perspect. 9, https://doi.org/10.1002/prp2.714 (2021).
Lusha, E. & Nan, J. [Effects of HMGB1 inhibition on endoplasmic reticulum stress after myocardial ischemia/reperfusion injury in rats]. Zhonghua wei zhong bing. ji jiu yi xue 29, 916–920 (2017).
Hu, J., Liu, X. J. & Tang, Y. HMGB1/Foxp1 regulates hypoxia-induced inflammatory response in macrophages. Cell Biol. Int. 46, 265–277 (2022).
Ouyang, F. et al. HMGB1 induces apoptosis and EMT in association with increased autophagy following H/R injury in cardiomyocytes. Int. J. Mol. Med. 37, 679–689 (2016).
Chen, Q. F. et al. Role of high-mobility group B1 in myocardial injury induced by coronary microembolization in rats. J. Cell. Biochem.120, 4238–4247 (2019).
Zhu, K. Y. et al. Glycyrrhizin attenuates myocardial ischemia reperfusion injury by suppressing Inflammation, oxidative stress, and ferroptosis via the HMGB1-TLR4-GPX4 pathway. Exp. Cell Res.435, https://doi.org/10.1016/j.yexcr.2024.113912 (2024).
Cai, X., Wang, X., Li, J. & Chen, S. Protective effect of glycyrrhizin on myocardial ischemia/reperfusion injury-induced oxidative stress, inducible nitric oxide synthase and inflammatory reactions through high-mobility group box 1 and mitogen-activated protein kinase expression. Exp. Ther. Med 14, 1219–1226 (2017).
Ren, Y. et al. Regulatory effects of toll-like receptor 4 knockout on CD4+ and CD8+ T lymphocytes and interleukin-17 during myocardial ischemia. Ann. Clin. Lab. Sci. 50, 761–768 (2020).
Abarbanell, A. M. et al. Exogenous high-mobility group box 1 improves myocardial recovery after acute global ischemia/reperfusion injury. surgery 149, 329–335 (2011).
Zhou, X. Y. et al. Exogenous high-mobility group box 1 protein injection improves cardiac function after myocardial infarction: involvement of Wnt signaling activation. J. Biomed. Biotechnol. https://doi.org/10.1155/2012/743879 (2012).
Zhou, Y. H. et al. HMGB1 protects the heart against ischemia-reperfusion injury via PI3K/AkT pathway-mediated upregulation of VEGF expression. Front. Physiol. 10, https://doi.org/10.3389/fphys.2019.01595 (2020).
Zhou, Y. H. et al. High Mobility Group Box 1 protein attenuates myocardial ischemia reperfusion injury via inhibition of the p38 mitogen-activated protein kinase signaling pathway. Exp. Ther. Med. 14, 1582–1588 (2017).
Yao, H.-C. et al. Intravenous High Mobility Group Box 1 upregulates the expression of HIF-1alpha in the myocardium via a protein kinase B-dependent pathway in rats following acute myocardial ischemia. Mol. Med. Rep. 13, 1211–1219 (2016).
Liu, F. Y. et al. TLR9 is essential for HMGB1-mediated post-myocardial infarction tissue repair through affecting apoptosis, cardiac healing, and angiogenesis. Cell Death Disease 10, https://doi.org/10.1038/s41419-019-1718-7 (2019).
Foglio, E., Puddighinu, G., Germani, A., Russo, M. A. & Limana, F. HMGB1 inhibits apoptosis following MI and induces autophagy via mTORC1 inhibition. J. Cell. Physiol. 232, 1135–1143 (2017).
Limana, F. et al. Transcriptional profiling of HMGB1-induced myocardial repair identifies a key role for notch signaling. Mol. Ther. 21, 1841–1851 (2013).
Chien, K. R. et al. Regenerating the field of cardiovascular cell therapy. Nat. Biotechnol. 37, 232–237 (2019).
Bangert, A. et al. Critical role of RAGE and HMGB1 in inflammatory heart disease. Proc. Natl. Acad. Sci. USA 113, E155–E164 (2016).
Sun, T., Dong, C. & Xiong, S. Cardiomyocyte-derived HMGB1 takes a protective role in CVB3-induced viral myocarditis via inhibiting cardiac apoptosis. Immunol. Cell Biol. 101, 735–745 (2023).
Hollenberg, S. M. & Singer, M. Pathophysiology of sepsis-induced cardiomyopathy. Nat. Rev. Cardiol. 18, 424–434 (2021).
Jiang, N. et al. Impact of Vitamin C on inflammatory response and myocardial injury in sepsis patients. Altern. Ther. Health Med. 30, 427–431 (2024).
Lv, H. et al. The clinical effectiveness of sivelestat in treating sepsis patients with both acute respiratory distress syndrome and septic cardiomyopathy. J. Cardiothorac. Surg. 19, https://doi.org/10.1186/s13019-024-02835-3 (2024).
Lv, X. W., Shi, X. H., Maihemuti, M., Yang, D. P. & Xiao, D. Correlation of HMGB1, TLR2 and TLR4 with left ventricular diastolic dysfunction in sepsis patients. Scand. J. Immunol. 97, https://doi.org/10.1111/sji.13260 (2023).
Li, W. et al. Discovery and synthesis of novel glyrrhizin-analogs containing furanoylpiperazine and the activity against myocardial injury in sepsis. Bioorg. Chem. 153, 107846 (2024).
Hagiwara, S., Iwasaka, H., Uchino, T. & Noguchi, T. High mobility group box 1 induces a negative inotropic effect on the left ventricle in an isolated rat heart model of septic shock—a pilot study. Circ. J. 72, 1012–1017 (2008).
Xu, H. et al. The alarmin cytokine, high mobility group box 1, is produced by viable cardiomyocytes and mediates the lipopolysaccharide-induced myocardial dysfunction via a TLR4/phosphatidylinositol 3-kinase gamma pathway. J. Immunol. (Baltimore, MD: 1950) 184, 1492–1498 (2010).
Su, Z. L. et al. HMGB1 blockade attenuates experimental autoimmune myocarditis and suppresses Th17-cell expansion. Eur. J. Immunol. 41, 3586–3595 (2011).
Su, Z. et al. Up-regulated HMGB1 in EAM directly led to collagen deposition by a PKCbeta/Erk1/2-dependent pathway: cardiac fibroblast/myofibroblast might be another source of HMGB1. J. Cell. Mol. Med. 18, 1740–1751 (2014).
Bybee, K. A. & Prasad, A. Stress-related cardiomyopathy syndromes. Circulation 118, 397–409 (2008).
Cui, H., Li, Y., Sun, M., Zhao, X. & Wu, M. HMGB1 neutralizing antibody attenuates hemorrhagic shock-induced cardiac injury and modulates apoptotic gene expression in murine heart. Natl. Med. J. China 94, 704–708 (2014).
Zhou, Y., Li, Y. & Mu, T. HMGB1 neutralizing antibody attenuates cardiac injury and apoptosis induced by hemorrhagic shock/resuscitation in rats. Biol. Pharm. Bull. 38, 1150–1160 (2015).
Conrad, N. et al. Temporal trends and patterns in heart failure incidence: a population-based study of 4 million individuals. Lancet 391, 572–580 (2018).
Xi, A. & Liao, B. Study on change in levels of HMGB1, TNF-alpha, chemerin in patients with congestive heart failure and its relationship with ventricular remodeling. J. Am. Coll. Cardiol. 66, C210 (2015).
Volz, H. C. et al. HMGB1 is an independent predictor of death and heart transplantation in heart failure. Clin. Res. Cardiol. 101, 427–435 (2012).
Wahid, A. et al. Serum HMGB1 is a biomarker for acute myocardial infarction with or without heart failure. Clin. Transl. Sci. 16, 2299–2309 (2023).
Wang, L. J. et al. Increased serum high-mobility group box-1 and cleaved receptor for advanced glycation endproducts levels and decreased endogenous secretory receptor for advanced glycation endproducts levels in diabetic and non-diabetic patients with heart failure. Eur. J. Heart Fail. 13, 440–449 (2011).
Huang, Y. Y. et al. Elevated serum HMGB1 in pulmonary arterial hypertension secondary to congenital heart disease. Vasc. Pharmacol. 85, 66–72 (2016).
Li, Q. et al. PARP1 interacts with HMGB1 and promotes its nuclear export in pathological myocardial hypertrophy. Acta Pharmacol. Sin. 40, 589–598 (2019).
Yang, X. et al. HMGB1 in macrophage nucleus protects against pressure overload induced cardiac remodeling via regulation of macrophage differentiation and inflammatory response. Biochem. Biophys. Res. Commun. 611, 91–98 (2022).
Limana, F. et al. HMGB1 attenuates cardiac remodelling in the failing heart via enhanced cardiac regeneration and miR-206-mediated inhibition of TIMP-3. PLoS ONE 6, https://doi.org/10.1371/journal.pone.0019845 (2011).
He, Y., Zhou, X., Zheng, X. & Jiang, X. Exogenous high-mobility group box 1 protein prevents postinfarction adverse myocardial remodeling through TGF-beta/Smad signaling pathway. J. Cell. Biochem. 114, 1634–1641 (2013).
Su, F., Shi, M., Zhang, J., Li, Y. & Tian, J. Recombinant high-mobility group box 1 induces cardiomyocyte hypertrophy by regulating the 14-3-3eta, PI3K and nuclear factor of activated T cells signaling pathways. Mol. Med. Rep. 23, https://doi.org/10.3892/mmr.2021.11853 (2021).
Zhang, L. et al. HMGB1 enhances mechanical stress-induced cardiomyocyte hypertrophy in vitro via the RAGE/ERK1/2 signaling pathway. Int. J. Mol. Med. 44, 885–892 (2019).
Lin, H. R. et al. HMGB1–RAGE axis makes no contribution to cardiac remodeling induced by pressure-overload. PLoS ONE 11, https://doi.org/10.1371/journal.pone.0158514 (2016).
Ha, T. et al. Reduced cardiac hypertrophy in toll-like receptor 4-deficient mice following pressure overload. Cardiovasc. Res. 68, 224–234 (2005).
Takahashi, K. et al. Modulated inflammation by injection of high-mobility group box 1 recovers post-infarction chronically failing heart. Circulation 118, S106–S114 (2008).
Goto, T. et al. High-mobility group box 1 fragment suppresses adverse post-infarction remodeling by recruiting PDGFRα-positive bone marrow cells. PLoS ONE 15, https://doi.org/10.1371/journal.pone.0230392 (2020).
Vagnozzi, R. J. et al. An acute immune response underlies the benefit of cardiac stem cell therapy. Nature 577, 405–409 (2020).
Kaur, N. et al. Paracrine signal emanating from stressed cardiomyocytes aggravates inflammatory microenvironment in diabetic cardiomyopathy. iScience 25, https://doi.org/10.1016/j.isci.2022.103973 (2022).
Volz, H. C. et al. HMGB1: the missing link between diabetes mellitus and heart failure. Basic Res. Cardiol. 105, 805–820 (2010).
Wang, W. K. et al. Inhibition of high-mobility group box 1 improves myocardial fibrosis and dysfunction in diabetic cardiomyopathy. Int. J. Cardiol. 172, 202–212 (2014).
Tao, A. B. et al. Cardiomyocyte–fibroblast interaction contributes to diabetic cardiomyopathy in mice: role of HMGB1/TLR4/IL-33 axis. Biochim. Biophys. Acta-Mol. Basis Disease 1852, 2075–2085 (2015).
Yao, Y. W. et al. Role of HMGB1 in doxorubicin-induced myocardial apoptosis and its regulation pathway. Basic Res. Cardiol. 107, https://doi.org/10.1007/s00395-012-0267-3 (2012).
Lv, X. L. et al. Glycyrrhizin improved autophagy flux via HMGB1-dependent Akt/mTOR signaling pathway to prevent Doxorubicin-induced cardiotoxicity. Toxicology 441, https://doi.org/10.1016/j.tox.2020.152508 (2020).
Luo, P. et al. HMGB1 contributes to adriamycin-induced cardiotoxicity via up-regulating autophagy. Toxicol. Lett. 292, 115–122 (2018).
Xu, Z. F. et al. High-mobility group box 1 protein-mediated necroptosis contributes to dasatinib-induced cardiotoxicity. Toxicol. Lett. 296, 39–47 (2018).
Zhang, H. Y., Wang, Z., Liu, Z. X., Du, K. & Lu, X. Protective effects of dexazoxane on rat ferroptosis in doxorubicin-induced cardiomyopathy through regulating HMGB1. Front. Cardiovasc. Med. 8, https://doi.org/10.3389/fcvm.2021.685434 (2021).
Zhao, P. et al. Neutrophil extracellular traps mediate cardiomyocyte ferroptosis via the Hippo-YAP pathway to exacerbate doxorubicin-induced cardiotoxicity. Cell. Mol. Life Sci.81, 122 (2024).
Narumi, T. et al. High-mobility group box 1-mediated heat shock protein beta 1 expression attenuates mitochondrial dysfunction and apoptosis. J. Mol. Cell. Cardiol. 82, 1–12 (2015).
Kwak, M. S. et al. Peroxiredoxin-mediated disulfide bond formation is required for nucleocytoplasmic translocation and secretion of HMGB1 in response to inflammatory stimuli. Redox Biol. 24, 101203 (2019).
Tang, D. et al. Endogenous HMGB1 regulates autophagy. J. Cell Biol. 190, 881–892 (2010).
Calogero, S. et al. The lack of chromosomal protein Hmg1 does not disrupt cell growth but causes lethal hypoglycaemia in newborn mice. Nat. Genet. 22, 276–280 (1999).
Yu, P. et al. Cardiomyocyte-restricted high-mobility group box 1 (HMGB1) deletion leads to small heart and glycolipid metabolic disorder through GR/PGC-1alpha signalling. Cell Death Discov. 6, 106 (2020).
Prelich, G. Gene overexpression: uses, mechanisms, and interpretation. Genetics 190, 841–854 (2012).
Mollica, L. et al. Glycyrrhizin binds to high-mobility group box 1 protein and inhibits its cytokine activities. Chem. Biol. 14, 431–441 (2007).
Vergoten, G. & Bailly, C. Analysis of glycyrrhizin binding to protein HMGB1. Med. Drug Discov. 7, https://doi.org/10.1016/j.medidd.2020.100058 (2020).
He, M., Bianchi, M. E., Coleman, T. R., Tracey, K. J. & Al-Abed, Y. Exploring the biological functional mechanism of the HMGB1/TLR4/MD-2 complex by surface plasmon resonance. Mol. Med. 24, 21 (2018).
Yang, H. et al. Inhibition of HMGB1/RAGE-mediated endocytosis by HMGB1 antagonist box A, anti-HMGB1 antibodies, and cholinergic agonists suppresses inflammation. Mol. Med. 25, 13 (2019).
Zhao, X. Y. et al. High mobility group box 1 derived mainly from platelet microparticles exacerbates microvascular obstruction in no reflow. Thromb. Res. 222, 49–62 (2023).
He, D. W. et al. HMGB1–RAGE axis contributes to myocardial ischemia/reperfusion injury via regulation of cardiomyocyte autophagy and apoptosis in diabetic mice. Biol. Chem. 405, 167–176 (2024).
Du, D. et al. Synthesis, biological evaluation, and molecular modeling of glycyrrhizin derivatives as potent high-mobility group box-1 inhibitors with anti-heart-failure activity in vivo. J. Med. Chem. 56, 97–108 (2013).
Limana, F. et al. Myocardial infarction induces embryonic reprogramming of epicardial c-kit+ cells: role of the pericardial fluid. J. Mol. Cell. Cardiol. 48, 609–618 (2010).
Sato, A. et al. DPP4 inhibition ameliorates cardiac function by blocking the cleavage of HMGB1 in diabetic mice after myocardial infarction. Int. Heart J. 58, 778–786 (2017).
Li, Z. et al. Single-cell transcriptome analyses reveal novel targets modulating cardiac neovascularization by resident endothelial cells following myocardial infarction. Eur. Heart J. 40, 2507–2520 (2019).
Alvarez, R. Jr. et al. Cardiomyocyte cell cycle dynamics and proliferation revealed through cardiac-specific transgenesis of fluorescent ubiquitinated cell cycle indicator (FUCCI). J. Mol. Cell. Cardiol. 127, 154–164 (2019).
Peters, M. D. J. et al. Updated methodological guidance for the conduct of scoping reviews. JBI Evid. Synth. 18, 2119–2126 (2020).
Tricco, A. C. et al. PRISMA extension for Scoping Reviews (PRISMA-ScR): checklist and explanation. Ann. Intern. Med. 169, 467–473 (2018).
Han, D. et al. Enhancing cardioprotection through neutrophil-mediated delivery of 18beta-glycyrrhetinic acid in myocardial ischemia/reperfusion injury. Adv. Sci. (Weinh.) 11, e2406124 (2024).
Liu, W. W. et al. HMGB1 may modulates proliferation and migration of endothelial cells by YAP1 under hypoxia. Biol. Bull. 51, 233–242 (2024).
Lozano, J. et al. Scalable generation of nanovesicles from human-induced pluripotent stem cells for cardiac repair. Int. J. Mol. Sci. 23, https://doi.org/10.3390/ijms232214334 (2022).
Rossini, A. et al. HMGB1-stimulated human primary cardiac fibroblasts exert a paracrine action on human and murine cardiac stem cells. J. Mol. Cell. Cardiol. 44, 683–693 (2008).
Author information
Authors and Affiliations
Contributions
J.N. and S.-H.M. conceived the study. S.-H.M. and R.G.-J.L. screened the articles. J.N. resolved any disputes. All authors provided critical feedback and contributed to writing the manuscript, and approved the contents.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mao, SH., Luo, R.GJ., Lee, C. et al. Scoping review of preclinical and clinical studies on the role of HMGB1 in heart disease. npj Cardiovasc Health 3, 20 (2026). https://doi.org/10.1038/s44325-026-00119-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44325-026-00119-4
