
Abstract
Given the absence of approved treatments for pathogenic variants in Peripherin-2 (PRPH2), it is imperative to identify a universally effective therapeutic target for PRPH2 pathogenic variants. To test the hypothesis that formation of the elongated discs in presence of PRPH2 pathogenic variants is due to the presence of the full complement of rhodopsin in absence of the required amounts of functional PRPH2. Here we demonstrate the therapeutic potential of reducing rhodopsin levels in ameliorating disease phenotype in knockin models for p.Lys154del (c.458-460del) and p.Tyr141Cys (c.422 A > G) in PRPH2. Reducing rhodopsin levels improves physiological function, mitigates the severity of disc abnormalities, and decreases retinal gliosis. Additionally, intravitreal injections of a rhodopsin-specific antisense oligonucleotide successfully enhance the physiological function of photoreceptors and improves the ultrastructure of discs in mutant mice. Presented findings shows that reducing rhodopsin levels is an effective therapeutic strategy for the treatment of inherited retinal degeneration associated with PRPH2 pathogenic variants.
Similar content being viewed by others
Introduction
Inherited retinal diseases (IRDs) encompass a wide range of heterogeneous disorders (https://web.sph.uth.edu/RetNet/disease.htm) resulting in the degeneration of photoreceptors and progressive visual impairment worldwide1,2,3. Advancements in genetic engineering have shed light on their etiology, leading to the development of the first FDA-approved IRD gene therapy and various clinical trials1,4.
Of the many genes associated with IRDs, mutations in Peripherin-2 (PRPH2) make up some of the most prevalent disease-causing pathogenic variants with over 200 already identified5,6. PRPH2 is a photoreceptor-specific tetraspanin located in the photoreceptor outer segment (OS) disc rim region and forms homo-tetramers as well as hetero-tetramers with its homolog, rod outer segment membrane protein 1 (ROM1)7,8. These tetramers subsequently assemble into octamers and higher order oligomers, which play a crucial role in OS disc rim formation9. Minimum threshold PRPH2 protein levels ( ~ 80% of wild type) are known to be essential for OS disc morphogenesis and maintenance10. Therefore, mice heterozygous for the Prph2 null allele (Prph2+/-) exhibit highly abnormal OSs, while homozygotes (Prph2−/−) fail to develop OSs11,12.
To date, there are no approved treatments or ongoing clinical trials specifically for PRPH2 associated diseases6. While many studies have provided insights into the correlation between PRPH2 genotypes and the resulting phenotypes in animal models13,14,15,16,17,18, establishing these associations in human patients is challenging. This difficulty arises from the substantial degree of interfamilial and intrafamilial phenotypic variability observed among individuals carrying identical PRPH2 pathogenic variants and, the complexity is further compounded by the occurrence of digenic inheritance involving its binding partner ROM119,20,21,22,23,24. Prph2 mutations can exert their effects through loss-of-function, dominant-negative, and/or gain-of-function mechanisms13,14,16, making traditional gene supplementation therapies inapplicable for all pathogenic variants. Supplementation alone cannot effectively treat dominant-negative and gain-of-function pathogenic variants13, while combining gene knockdown with supplementation could be a potential solution. However, the large number of unique pathogenic variants and their low occurrence renders this approach economically unfeasible25. These complex pathogenic mechanisms along with the vast array of different disease-causing pathogenic variants makes the identification of a ubiquitous target for the treatment of PRPH2-associated diseases highly desirable.
The ratio of PRPH2 to the rod-specific protein rhodopsin (RHO) plays a crucial role in morphogenesis of rod OS discs26,27. RHO is a light-sensitive protein located in the disc lamellae and is the most abundant retinal protein28,29. It is highly regulated due to its essential role in phototransduction and visual function27,30,31,32,33. Murine models with an increased RHO/PRPH2 ratio exhibited formation of discs with significantly larger diameters than wild type, accelerated retinal degeneration, and decreased physiological function26,27,32. In contrast, models with a decreased ratio display properly oriented rod outer segments (ROSs) with decreased diameters27,34,35 but possess reduced sensitivity and faster flash-response kinetics before degenerating over a slow time course34,35,36. Through the utilization of a mouse model that overexpresses PRPH2, we successfully alleviated the stress associated with abnormally high RHO:PRPH2 ratio, which emphasized the significance of maintaining this ratio for proper photoreceptor structure and function27. It is imperative to acknowledge that any therapeutic interventions targeting PRPH2-affected photoreceptors must focus on generating sufficient protein levels to maintain the RHO:PRPH2 ratio as close as it is in wild type retinas. These findings also shed light on the delicate nature of rod photoreceptors, indicating that their heightened sensitivity to excessive opsin levels. Pathogenic variants in PRPH2 are known to decrease the levels of functional protein contributing to the formation of the elongated discs37. Therefore, we hypothesized that an increased RHO to functional PRPH2 ratio contributes to disc elongation and disease progression of PRPH2-associated disorders.
In this study, we demonstrate that modifying the RHO/PRPH2 ratio in favor of PRPH2 positively impacts the disease phenotype in previously characterized knockin mouse models expressing the patient pathogenic variants c.461_463del, p.Lys154del19 (herein called K153∆ for the knockin mouse model13) and c.422 A > G, p.Tyr141Cys22,23 (herein called Y141C for the knockin mouse model14) in PRPH2. Proof-of-concept studies presented here revealed that reducing one allele of Rho in the heterozygous models improves photoreceptor ultrastructure and physiological function. Furthermore, in order to demonstrate the translational applicability of this strategy, we employed a previously characterized antisense oligonucleotide (ASO) mRho ASO1 shown to effectively reduce Rho transcript levels following intravitreal administration38. ASO-mediated reduction of RHO levels in mice heterozygous for the Prph2 Y141C knockin mutation resulted in improved retinal function, OS ultrastructure, and delayed photoreceptor degeneration. These findings suggest that reducing RHO levels is a potential therapeutic strategy to ameliorate the disease phenotype in patients with PRPH2-associated inherited retinal disorders.
Results
Reducing RHO levels in Prph2 knockin mouse models expressing mutant PRPH2 leads to functional improvements
In Rho+/- mice, retinal development proceeds normally with proper lamination but gradually undergoes degeneration over time34. The reduction of RHO manifests specific effects on the ultrastructure of ROS, including an elliptical disk shape and decreased surface area35. To determine the impact of reducing RHO levels in ROS discs on functionality of the retinas from mice expressing mutant forms of PRPH2, comprehensive electroretinography measurements (ERG) were conducted on Prph2K153∆/+ and Prph2Y141C/+ mice that were also hemizygous for Rho (Rho+/-). These Prph2 heterozygous mice were chosen for their clinical relevance, as PRPH2-associated retinitis pigmentosa typically exhibits autosomal dominant inheritance16,39. ERGs were performed at various postnatal (P) stages (P17, P30, and P90) to evaluate differences in scotopic and photopic maximum amplitudes during photoreceptor maturation and disease progression (Fig. 1A–F).

Representative scotopic (A) and photopic (B) ERG waveforms at P30 from all genotypes. C–F Mean maximum amplitudes of scotopic a-waves and b-waves, as well as photopic b-waves, plotted as mean ± SD at P17, P30, and P90. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, ∗∗∗∗P < 0.0001 by one-way ANOVA with Tukey’s post-hoc comparison (see Table S3 for corresponding p values). N = 9-15 animals/genotype/age. *denotes comparisons between (C, D) Prph2Y141C/+ and Prph2Y141C/+/Rho+/- or (E, F) Prph2K153∆/+ and Prph2K153∆/+/Rho+/-.
Eliminating one allele of Rho in Prph2Y141C/+ mice led to a significant improvement in mean maximum amplitudes of scotopic a- and b-waves as early as P17 compared to Prph2Y141C/+controls ( ~ 53% and ~43% increases respectively, Fig. 1C). Additionally, there was an increase in the photopic ( ~ 29%) b-wave mean maximum amplitude, although this increase did not reach statistical significance (Fig. 1D). These improvements persisted throughout the study, as Prph2Y141C/+/Rho+/- retinas exhibited significantly better mean maximum amplitudes of scotopic a-waves ( ~ 37%) and photopic b-wave ( ~ 25%) at P30, as well as scotopic a-waves ( ~ 60%) and b-waves ( ~ 40%) at P90 (Fig. 1C, D).
For P17 Prph2K153∆/+/Rho+/- mice, the assessment revealed modest non-significant functional improvements in the scotopic a- and b- as well as photopic b-wave mean maximum amplitudes compared to Prph2K153∆/+ ( ~ 14%, ~10%, and ~45%, respectively) (Fig. 1E, F). As the mice matured and their retinas fully developed, the partial ablation of Rho significantly enhanced scotopic a- and photopic b-wave amplitudes at P30 ( ~ 58% and ~51%, respectively). However, this statistically significant restorative effect was only apparent in the scotopic a-wave mean maximum amplitude ( ~ 42%) at P90 (Fig. 1E, F).
Reduced RHO levels result in histologic improvements
To determine if the observed functional improvements were attributed to increased photoreceptor survival, we conducted outer nuclear layer (ONL) counts for the different genotypes at P30 and P90. Despite the functional improvements, histologic and morphometric analyses at the light level revealed only minor changes between the Prph2 mutant models and their compound heterozygote/hemizygote counterparts (Fig. 2). At P30, ONL counts were similar among WT, Rho+/-, Prph2Y141C/+, and Prph2Y141C/+/Rho+/- (Fig. 2B, left). As the mice aged, significant loss of ONL nuclei was observed in Rho+/-, Prph2Y141C/+, and Prph2Y141C/+/Rho+/- compared to WT at P90 (Fig. 2B, right and see Table S4). Modest ONL loss was noted in Prph2K153∆/+ and Prph2K153∆/+/Rho+/- at P30, progressing to significant photoreceptor loss in both models by P90 (Fig. 2C and see Table S5). However, a reduction in RHO levels was observed to improve average nuclear counts in the central regions of the Prph2K153∆/+/Rho+/- retina at both time points (Fig. 2C).

A Representative images of H&E stained retinal sections at P30 and P90. B, C Nuclei counts were performed in 100 µm width windows in retinal sections taken from the indicated genotypes. Images were captured every 500 µm starting at the optic nerve head sections, and nuclei counts from multiple regions were plotted as mean ± SD for the listed genotypes (n = 3 for each genotype). ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001 by two-way ANOVA (P30 Prph2Y141C/+: Interaction P = 0.8626, Row Factor P < 0.0001, and Column Factor P < 0.0001. P90 Prph2Y141C/+: Interaction P = 0.0001, Row Factor P < 0.0001, and Column Factor P < 0.0001. P30 Prph2K153∆/+: Interaction P = 0.0026, Row Factor P < 0.0001, and Column Factor P < 0.0001. P90 Prph2K153∆/+: Interaction P < 0.0001, Row Factor P < 0.0001, and Column Factor P < 0.0001.) with Tukey’s post-hoc comparison. *denotes comparisons between (B) Prph2Y141C/+ or (C) Prph2K153∆/+and (B) Prph2Y141C/+/Rho+/- or (C) Prph2K153∆/+/Rho+/-. Scale bar corresponds to 100 µm.
Reducing RHO levels leads to enhanced structural integrity of the outer segments
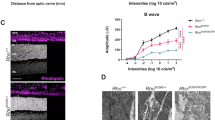
To explore additional factors contributing to the observed functional improvements, retinas underwent ultrastructural analyses. Low magnification ultrastructure images revealed that the elimination of one allele of Rho resulted in an overall enhancement of OS structure in both Prph2 models (Fig. 3A, top panels). OSs display improved disc stacking and a reduction in the number of membranous whorls, particularly in Prph2Y141C/+/Rho+/- (arrowheads in Fig. 3A top panel).

A Representative TEM images at low and high-magnification of tannic acid/uranyl acetate-stained retinas from WT, Rho+/-, Prph2Y141C/+, Prph2Y141C/+/Rho+/-, Prph2K153∆/+, and Prph2K153∆/+/Rho+/- at P16. B Quantification of open discs at the base of the OS in the listed genotypes at P16, utilizing data from a previous publication for WT and Prph2Y141C/+ open discs37. C Quantification of OS diameters measured in the listed genotypes at P16, with each data point representing a single outer segment. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, ∗∗∗∗P < 0.0001 by one-way ANOVA (P < 0.0001 for both open disc and OS diameter) with Tukey’s post-hoc comparison. Arrowheads highlight misaligned, overgrown, and sagittally aligned discs, as well as whorl-like structures commonly observed in Prph2Y141C/+ and Prph2K153∆/+ ROSs. N (number of independent eyes used for analyses) values for (B): WT: 111, Rho+/-: 112; Prph2Y141C/+: 152; Prph2Y141C/+/Rho+/-: 105; Prph2K153∆/+: 98, Prph2K153∆/+/Rho+/-: 43 and for C: WT: 86, Rho+/-: 138, Prph2Y141C/+: 80, Prph2Y141C/+/Rho+/-: 139, Prph2K153∆/+: 63, Prph2K153∆/+/Rho+/-: 90. Scale bar, 1 µm. Error bars represent mean ± SD.
Mutant PRPH2-associated defects in protein oligomerization have been linked to impaired disc closure, resulting in enlarged OS diameters and increased numbers of open, nascent discs37. Tannic acid staining allows differentiation between nascent and mature discs, as the exposed nascent discs exhibit a darker staining pattern due to their exposure to the extracellular space40. Higher magnification images were utilized to measure OS diameters and count open discs, providing a quantitative assessment of the impact of RHO reduction on these morphological abnormalities (Fig. 3A, lower panels). In the presence of WT PRPH2 levels, removing one allele of Rho decreased the average number of open nascent discs from ~9 in WT to ~5 in Rho+/- (Fig. 3B). Similarly, the reduction in RHO levels resulted in decreased numbers of open discs, with averages of ~18 for Prph2Y141C/+ and ~12 for Prph2K153∆/+ reduced to ~6 and ~5 in Prph2Y141C/+/Rho+/- and Prph2 K153∆/+/Rho+/-, respectively (Fig. 3B). The rescue effect was also evident in OS diameter measurements (µm), with mean values of 1.2, 1.5, and 1.4 in WT, Prph2Y141C/+, and Prph2K153∆/+, respectively, reduced to 1.0, 0.9, and 1.1 in Rho+/-, Prph2Y141C/+/Rho+/-, and Prph2K153∆/+/Rho+/- (Fig. 3C). These findings demonstrate that improved ultrastructure of OSs underlies the observed functional improvements in Prph2Y141C/+/Rho+/- and Prph2K153∆/+/Rho+/- mice.
Partial ablation of one allele of Rho leads to reduced levels of RHO relative to PRPH2
To establish the correlation between ablation of one Rho allele and the observed functional and structural improvements, immunodot blots were performed on retinal extracts from P30 mice. This aimed to determine the degree of decrease in RHO levels and the resulting RHO/PRPH2 ratio across all models (Fig. S1A). The partial genetic ablation of Rho resulted in reduction in protein levels in Rho+/- ( ~ 56%), Prph2Y141C/+/Rho+/- ( ~ 55%), and Prph2K153Δ/+/Rho+/- ( ~ 65%) compared to WT, Prph2Y141C/+, and Prph2K153Δ/+, respectively (Fig. S1B). Notably, this reduction did not impact the levels of PRPH2 (Fig. S1C). These findings highlight that the partial genetic ablation of Rho effectively decreases the ratio of RHO to PRPH2 in Rho+/-, Prph2Y141C/+/Rho+/-, and Prph2K153Δ/+/Rho+/- retinas.
Partial ablation of RHO does not impact the formation of higher-order complexes of PRPH2
Since proper PRPH2 oligomerization is essential for disc formation and OS structure, we examined the oligomerization status of PRPH2 in these mice following partial ablation of RHO37,41. Retinal extracts were subjected to immunoblotting after separation by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) under non-reducing conditions (Fig. S2A). Under these conditions, WT PRPH2 typically separates into monomers ( ~ 37 kDa) and disulfide-linked dimers ( ~ 75 kDa), while Prph2Y141C/+ and Prph2K153∆/+ mutants have been shown to exhibit additional abnormal high-molecular weight protein complexes13,14. Our experimental results confirmed the presence of this abnormal complexes in Prph2Y141C/+/Rho+/- and Prph2 K153∆/+/Rho+/- retinas (Fig. S2A, red arrows). Quantification of the different complex forms was performed by measuring the signal intensity of each complex for PRPH2 and ROM1 and dividing it by the total signal intensity for the respective lane (Fig. S2B and C). No changes in the distribution of PRPH2 complexes were observed in either Prph2Y141C/+/Rho+/- (Fig. S2B, left) or Prph2K153∆/+/Rho+/- (Fig. S2B, right) compared to their respective heterozygous mutant counterparts. These findings indicate that the reduction of RHO does not affect the oligomerization of PRPH2.
Reducing RHO level attenuates retinal gliosis in a mutation-independent manner
Müller cell gliosis, implicated in numerous retinal disorders42, is characterized by the upregulation of glial fibrillary acidic protein (GFAP) as a non-specific marker of active gliosis and stress during retinal degeneration43,44,45. Since reducing RHO levels in both Prph2 models led to functional and structural improvements, we next addressed the impact of that reduction on GFAP upregulation and its cellular distribution. Immunofluorescence and immunoblotting were conducted on P30 Prph2Y141C/+ and Prph2K153∆/+ retinas in the presence of WT levels of RHO or after ablation of one allele (Fig. S3). In WT and Rho+/- retinal sections, GFAP was predominantly localized in the nerve fiber layer consistent with previous literature (Fig. S3A, upper panels), whereas in Prph2Y141C/+ and Prph2K153∆/+, GFAP extended to other retinal layers, including the ONL (Fig. S3A, arrows in middle left and lower left panels). Notably, the ablation of one Rho allele led to a reduction in retinal stress, evidenced by GFAP restriction to the nerve fiber, ganglion, and inner plexiform layers (Fig. S3A, arrows in middle right and lower right panels). However, quantification by immunodot blotting of P30 retinal extracts from the listed genotypes (Fig. S3B) revealed a non-statistically significant trend in GFAP reduction upon elimination of one Rho complement (Fig. S3C). These findings collectively suggest that reducing RHO expression diminishes retinal stress associated with mutant Prph2 expression, although further research is warranted to confirm this observation and its significance.
Intravitreal treatment with Rho-specific ASO slows functional decline in Prph2 Y141C/+ mice
We next investigated the potential of modulating RHO levels in a clinically relevant manner by the introduction of a rhodopsin specific ASO called mRho ASO138. This ASO had already been tested and demonstrated efficacy in reducing endogenous mouse RHO38. Murray et al. showed “a dose-dependent reduction in rhodopsin mRNA was observed in eyes treated with mRho ASO1”38. Titrations of a single intravitreal injection of mRho ASO1 were performed at ages P15 and P45 to explore the dose-dependent effects of early and late-stage therapeutic intervention in Prph2Y141C/+ mice. Therapeutic efficacy was assessed by recording ERGs at ages P60 and P90 to identify changes in visual function. Results were expressed as percentages normalized to the vehicle control values for each dosage (Figs. S4 and S5). Due to procedure related complications with the smaller eyes of P15 mice, we report the initial titration results as the maximum amplitude (µV) measured for independent mRho ASO1 treated samples divided by the mean maximum amplitude of independent vehicle injected controls (Fig. S4B–E). The results for P45 injected mice are reported as the maximum amplitude measured for the ASO treated eye divided by that of the contralateral vehicle injected control eye (Fig. S5B–E).
The titration experiments revealed that optimal dosages were age at intervention dependent. Following P15 intervention, a dose of 3.125 µg showed the most improvement at P60 as determined by maximum scotopic a- ( ~ 146%), scotopic b- ( ~ 137%), and photopic b- ( ~ 124%) amplitudes compared to vehicle control eyes (Fig. S4B and C). As the mice aged and the disease progressed, similar improvements were observed at P90 for both 1.56 µg (scotopic a-wave: ~167%, scotopic b-wave: 153%, and photopic b-wave: 120%) and 3.125 µg (scotopic a-wave: ~167%, scotopic b-wave: ~157%, and photopic b-wave: ~121%) treated eyes compared to their vehicle control eyes (Fig. S4D and E). Based on the observed increases in maximum amplitudes at P60 and P90 with the administration of 3.125 µg of mRho ASO1, we chose this dosage for the P15 intervention in the remainder of the study. As to the intervention at P45, improvements in maximum amplitudes were observed at P60 following the administration of 6.25 µg (scotopic a-wave: ~135%, scotopic b-wave: ~117%, and photopic b-wave: ~105%) and 12.5 µg (scotopic a-wave: ~117%, scotopic b-wave: ~97%, and photopic b-wave: ~117%) dosages (Fig. S5B and C). The effects of the 6.25 µg dosage persisted at P90 (scotopic a-wave: ~133%, scotopic b-wave: ~112%, and photopic b-wave: ~108%), while the beneficial effects of the 12.5 µg dosage appeared to diminish (scotopic a-wave: ~92%, scotopic b-wave: ~80%, and photopic b-wave: ~105%) (Fig. S5D and E).
To demonstrate the specificity of this ASO, a control ASO (5’-CCTATAGGACTATCCAGGAA-3’) was generated and used. We assessed the impact of intravitreal injecting the control ASO to ensure there were no adverse effects on retinal function. For this study, the most effective dose of 6.25 µg in 1 µl was used to evaluate the efficacy of the control ASO in P30 Prph2Y141C/+ mice. At 15 days post injection (PI), mice were evaluated functionally, structurally and biochemically (Figs. S6, S7 and S8, respectively). Results were compared to P45 old untreated and PI-15 vehicle injected animals. Functional assessments revealed scotopic a- and b-waves, as well as photopic a- and b-waves, were indistinguishable from the untreated or vehicle treated controls (Fig. S6). Quantification of the ERG responses similarly showed no changes compared to the controls, both untreated and vehicle-injected (Fig. S6C). These results were further confirmed through light microscopy, and the number of photoreceptor nuclear counts showed no significant changes compared the control eyes (Fig. S7B and C). To verify that the injection of the control ASO did not affect the levels of endogenous RHO or PRPH2, immunodot blot analyses were conducted. Once again, no changes in the levels of RHO or PRPH2 were observed following the injection of the control ASO compared to untreated and treated samples (Fig. S8).
To further investigate our findings related to mRho ASO1 intervention at P15, we recorded ERGs from Prph2Y141C/+ mice at PI-15, PI-45 and P75, respectively (Fig. 4A). Comparing ERG values for ASO-treated eyes with their untreated contralateral eyes revealed significant increases in mean treated maximal scotopic a-wave amplitudes at each evaluated time point (PI-15: ~107%, PI-45: ~112%, and PI-75: ~113%), while b-wave maximum amplitudes remained unchanged following therapeutic intervention (Fig. 4B, C). This pattern of specific improvement was also observed in photopic responses, where photopic a-wave maximum amplitudes were significantly improved at each time point of assessment compared to their corresponding control eye (PI-15: ~120%, PI-45: ~124%, and PI-75: ~130%), and no significant differences were observed when evaluating photopic b-wave maximum amplitudes (Fig. 4D).

A Schematic representation of the design for early-stage preclinical intervention studies. Created with BioRender.com released under a Creative Commons Attribution-Non-commercial-NoDervis 4.0 International license. B Representative waveforms of scotopic and photopic responses recorded 15 days after injection at P30. Mean ± SD maximum amplitudes of scotopic a-waves and scotopic b-wave (C) and photopic a-wave and b-wave amplitudes (D) of treated (3.125 µg mRho ASO1) and untreated contralateral control eyes recorded in preclinical early injection studies at ages P30, P60, and P90. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001 by Mann-Whitney U test (PI-15: N = 51, PI-45: N = 34, and PI-75: N = 18).
Functional changes were evaluated at PI-15 and PI-45 following P45 mRho ASO1 intervention (Fig. 5A). Comparing ASO-treated eyes with their vehicle control contralateral counterparts at PI-15 revealed higher scotopic ( ~ 120%) and photopic ( ~ 144%) a-wave maximum amplitudes, while no observable differences were noted in scotopic and photopic b-wave maximum amplitudes (Fig. 5B, D). This improvement in a-wave amplitudes persisted at PI-45 (scotopic a-wave: ~124% and photopic a-wave: ~118%), with no enhancement observed in b-wave maximum amplitudes (Fig. 5C and D).

A Design of late-stage preclinical intervention studies. Created with BioRender.com released under a Creative Commons Attribution-Non-commercial-NoDervis 4.0 International license. B Representative scotopic and photopic waveforms recorded at 15- and 45-days post injections at P45. C, D Mean maximum amplitudes of scotopic a-waves and b-waves, as well as photopic a-waves and b-waves were plotted as mean ± SD for treated eyes (6.25 µg mRho ASO1) and vehicle control contralateral eyes in the late-stage intervention studies conducted at P60 and P90. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, ∗∗∗∗P < 0.0001 by Mann-Whitney U test (PI-45: N = 38 and PI-75: N = 18).
ASO treatment lowers the ratio of RHO to PRPH2
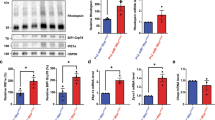
Next, we explored the effects of ASO intervention on RHO and PRPH2 levels by quantifying protein amounts through immunodot blots and employing a two-tailed t-test for statistical analysis (Fig. 6A–F). The evaluation time points following intravitreal injection at P15 included ages P30, P60, and P90 corresponding to PI-15, PI-45, and PI-75. Treated eyes exhibited a reduction in mean RHO levels by ~32% at P30, and ~49% at P60, and P90, albeit this reduction was statistically insignificant at P30 (Fig. 6A). ASO administration at P15 did not significantly affect PRPH2 protein levels at any of the assessed time points (Fig. 6B). A statistically significant reduction in mean ratio of RHO to PRPH2 was observed at all three time points (P30: ~43%, P60: ~49%, and P90: 52%) (Fig. 6C). These improved ratios indicate the successful achievement of the desired therapeutic effect by efficiently reducing RHO levels. It is important to avoid excessive reduction of RHO, as it may exacerbate the degeneration and subsequently reduce PRPH2 levels.

Quantification of (A) RHO and (B) PRPH2 in retinal extracts at P30, P60, and P90 after injections with mRho ASO1 at P15 (3.125 µg mRho ASO1). C Graphs depicting the ratio of RHO to PRPH2 following P15 injection, determined by dividing the RHO signal intensity value by that of PRPH2. Quantification of (D) RHO and (E) PRPH2 in retinal extracts at P60 and P90 after injections with mRho ASO1 at P45 (6.25 µg mRho ASO1). F Graphs illustrating the ratio of RHO to PRPH2 following P45 injections. G–I Transcript levels assessed by qRT-PCR 15 days post injection at either P15 or P45. Quantification of (G) Rho and (H) Prph2 mRNA transcript levels relative to Gapdh for P15 and P45 injected samples. I Quantification of Rho mRNA transcript levels relative to Prph2 for P15 and P45 injected samples. Data are presented as mean ± SD. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, ∗∗∗∗P < 0.0001 as determined by student t-test analysis. N = 3-5 retinas per treatment condition.
Protein quantification was performed at PI-15 and PI-45 for intervention at P45 (P60 and P90, respectively). Average RHO protein levels showed a reduction of ~53% at P60 and ~71% at P90 compared to contralateral control eyes (Fig. 6D). Similar to the early intervention group, ASO treatment did not induce significant changes in average PRPH2 levels at both P60 and P90, although a slight decrease in the mean was observed at P90 (Fig. 6E). However, assessing the ration of RHO to PRPH2 revealed that the therapeutic effect of ASO treatment was successfully achieved, resulting in a decreased mean ratio of RHO to PRPH2 (P60: ~60% and P90: ~64%). Furthermore, this reduction was statistically significant at both time points (Fig. 6F).
To verify the mechanism of action for mRho ASO1 intervention, qRT-PCR was employed (primers used are in Table S1) to quantify Rho and Prph2 transcript levels post injection (Fig. 6G–I). Quantification of Rho transcript levels normalized to that of Gapdh revealed significant decreases in mRNA levels following injections at P15 ( ~ 35%) and P45 ( ~ 24%) (Fig. 6G). In contrast, evaluation of Prph2 transcript levels normalized to Gapdh revealed no significant changes between uninjected and treated eyes following P15 injections. However, we observed a significant increase ( ~ 11%) in Prph2 transcript levels following P45 injection of mRho ASO1 compared to vehicle control (Fig. 6H). We also assessed the relative abundance of the Rho transcript compared to Prph2 transcript to gain insight into how the intervention affects this ratio. This revealed similar levels of significant reduction in the ratio of Rho to Prph2 mRNA following P15 ( ~ 24%) and P45 ( ~ 31%) injections (Fig. 6I). These findings further support the mechanism of action of mRho ASO1 intervention in effectively reducing Rho transcript levels while exhibiting no significant impact on Prph2 transcript levels.
Age-dependent impact of mRho ASO1 intervention in mitigating histopathologic defects
To assess the histologic changes in Prph2Y141C/+ following ASO treatment, morphometric analyses were conducted on retinal sections from P90 mice injected with either 3.125 µg mRho ASO1 at P15 or 6.25 µg mRho ASO1 at P45. Animals injected at P15 exhibited significant improvement in the number of photoreceptor nuclei compared to vehicle and untreated controls (Fig. 7A, B). However, injections at P45 did not show a significant improvement in number of photoreceptor nuclei, except for one area in the far superior periphery (Fig. 7C). When directly comparing the two intervention time points, it is evident that P15 intervention leads to improved ONL nuclear count throughout most of the retina, with a statistically significant improvement in the superior portion of the retina (Fig. 7D). Taken together, these results demonstrate the efficacy of early intervention using mRho ASO1 as a therapeutic strategy to delay photoreceptor death caused by the Prph2Y141C/+ mutation.

A Representative images of retinal sections stained with H&E at P90. B–D Nuclear counts from 100 µm-windows at every 500 µm distance from the optic nerve and across the superior-inferior plane of retinal sections collected from P90 un-injected, 1X PBS injected as a control and mRho ASO1 injected animals following (B) early (P15) or (C) late-stage (P45) therapeutic intervention. B, C WT and Prph2Y141C/+ controls were added for comparison. B, C ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001 by two-way ANOVA (P15 Injected: Interaction P < 0.0001, Row Factor P < 0.0001, and Column Factor P < 0.0001. P45 Injected: Interaction P < 0.0001, Row Factor P < 0.0001, and Column Factor P < 0.0001. P15 vs. P45 Injected: Interaction P < 0.0001, Row Factor P < 0.0001, and Column Factor P < 0.0001.) with Tukey’s post-hoc comparison. *denotes comparisons between Prph2Y141C/+ and (B) Prph2Y141C/+ 3.125 µg ASO or (C) Prph2Y141C/+ 6.25 µg ASO. #denotes comparisons between (B) Prph2Y141C/+ 3.125 µg ASO and P15 injected Prph2Y141C/+ 1X PBS. D Plotted are mean ± SD values from (B) and (C) for WT, Prph2Y141C/+ P15 Injected 3.125 µg ASO, and P45 injected 6.25 µg ASO Prph2Y141C/+ for ease of comparison. + denotes comparisons P15 injected and P45 injected. N = 3 animals for all genotypes and experimental conditions. Scale bar corresponds to 50 µm.
mRho ASO1 intervention improves OS ultrastructure
Ultrastructural examination of ROSs was performed 45 days post injection at both P15 and P45. ASO treated photoreceptors showed notable improvements in their upright appearance, organization and a reduction in whorl formations (Figs. S9, S10 and S11), which are characteristic features of mutant PRPH2 murine retinas7,12,13,16,27,37. Evaluating the percentage these whorls represent of the total ROSs throughout the retina revealed ~50% reduction after early ASO injections and ~62% reduction associated with late therapeutic intervention compared to contralateral controls (Fig. S11B, right panel). Additionally, numerous nucleated cells were frequently observed in the subretinal space of control P90 Prph2Y141C/+ eyes, containing a large amount of phagocytosed material (Fig. S11A, arrow in right panel). While these cells were also present in ASO-injected animals, their presence was significantly reduced (compare Fig. S10A (arrowheads) to B). Evaluation of PBS-treated control Prph2Y141C/+ eyes at P45 and collected at P90 revealed the presence of likely activated microglia. These cells were characterized by their nucleated appearance (Fig. S12A, arrowhead), localization next to the RPE (Fig. S12), extended processes (Fig. S12B, arrowhead), and the presence of phagocytized material and numerous lysosomes (Fig. S12C, arrowhead).
To further investigate the observed improvements in ROS ultrastructural, measurements of OS diameters and open disc counts were performed at PI-45 following both early and late treatment to quantitatively assess the known morphological defects (Fig. 8A). Treatment with mRho ASO1 resulted in a reduction in the number of open nascent discs compared to contralateral controls following both time points of intervention (P15 intervention: control ~12.4 and treated 7.1) (P45 intervention: control ~11.8 and treated ~7.6) (Fig. 8B and Table 1). This decrease was also evident in OS diameter measurements (P15 intervention: control ~1.8 µm and treated ~1.3 µm) (P45 intervention: control ~2 µm and treated ~1.3 µm) (Fig. 8C and Table 1). Similar to what was observed with genetic ablation, these results indicate that mitigation of these morphological defects plays a crucial role in driving the observed improvements in ROS ultrastructural following mRho ASO1 administration.

A Representative high-magnification TEM images of tannic acid/uranyl acetate-stained retinas taken from P90 mRho ASO1 treated and contralateral control eyes that were injected at P15 (3.125 µg mRho ASO1) and P45 (6.25 µg mRho ASO1). Quantification of open discs at the base of the OS (B) and OS diameters (C) in mRho ASO1 injected and contralateral control eyes 45 days after treatment. Each data point represents a single OS. ∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001, ∗∗∗∗P < 0.0001 by one-way ANOVA (P < 0.0001 for both open discs and OS diameter) with Tukey’s post-hoc comparison. N = 50 OSs per each sample for (B) and (C). Scale bar, 1 µm. Error bars represent mean ± SD.
Discussion
Photoreceptors exhibit high metabolic demands and are prone to the accumulation of toxic photo-oxidative products associated with the visual cycle. To ensure their proper function and overall health, photoreceptors undergo a daily physiological renewal process that is crucial46,47. This process involves diurnal shedding of the distal portion of the outer segment, where discs are phagocytosed by the RPE, followed by the formation of new disc at the proximal end to replace them46,47. The presence of PRPH2 is crucial for proper disc morphogenesis, as its absence leads to failure in outer segment formation and the release of membrane evaginations from the connecting cilium in the form of ectosomes27,48. PRPH2 plays a dual role, not only inhibiting ectosome release but also participating in the proper enclosure of mature photoreceptor discs37. Defects in discs enclosure lead to misaligned overgrown discs and aberrations in the form of whorls, which are characteristics features observed in photoreceptor outer segments of animal models expressing Prph2 pathogenic variants13,14,16,17,18,49,50. These structural abnormalities in the outer segments significantly impact photoreceptor function, leading to progressive loss of visual function and a reduction in the number of viable photoreceptor cells13,14. Despite the identification of numerous pathogenic variants in the PRPH2 gene in patients5,6, finding an effective therapeutic approach remains challenging.
In pursuit of finding approaches to delay disease progression in a comprehensive manner, we explored a novel therapeutic strategy targeting the ratio between RHO and PRPH2 to address pathological structural phenotype. Mutations in Prph2 often lead to reduced levels and/or functionality of PRPH2, disrupting the essential photoreceptor protein balance needed for cellular homeostasis. Given that the abundance of RHO influences the size and structure of rod outer segment discs26,28, we hypothesized that reducing RHO levels in Prph2 mutant models could mitigate the phenotypic overgrowth abnormalities.
In proof of concept studies, we utilized partial genetic ablation of Rho to modulate protein levels in two well-established mouse models expressing mutant PRPH2, Prph2Y141C/+ and Prph2K153∆/+13,14. This reduction in RHO levels resulted in improved maximum physiological responses driven by rods in both models due to enhanced rod outer segment structure. Interestingly, despite being a rod-specific protein, the decrease in RHO reduction also improved cone-driven responses in both models. This finding is in line with the well-documented symbiotic relationship between rods and cones, where cone degeneration is often secondary to rod degeneration in retinitis pigmentosa51,52. The proper alignment of rod outer segment may contribute to the upright positioning of cone outer segments, resulting in enhanced responses to light stimulation. Previous studies have demonstrated that rods produce factors supportive of cones53,54. It is conceivable that mRho ASO1 treatment improved rod outer segment structures, thereby reducing cellular stress and promoting the production of these supportive factors, consequently leading to improved cone responses.
Although we observed only transient improvements in physiological function for Prph2K153∆/+, the potential of RHO reduction as a therapeutic strategy for this mutation is promising. Previous efforts to supplement the Prph2 gene in Prph2K153∆/+ mice by crossing them with a PRPH2-overexpresser line demonstrated limited efficacy, resulting in minimal improvements in scotopic a-wave ERGs at P3013. Therefore, the transient rescue of scotopic a-, scotopic b-, and photopic b-wave maximum amplitudes associated with RHO reduction is particularly exciting. One possible explanation for this transient rescue is based on our observations from ASO titration studies, suggesting that levels of RHO reduction may offer therapeutic benefits in the early stages but become detrimental as the disease progresses. In future studies, it will be crucial to explore whether controlled modulation of RHO levels in Prph2K153∆/+ mice can result in sustained therapeutic benefits.
To demonstrate the clinical relevance of this strategy and achieve controlled reduction of RHO levels, we opted to use a previously characterized ASO, mRho ASO138. This ASO was employed as a control by Murray et al. and demonstrated specificity for wild type RHO38. ASOs are single-stranded nucleic acids with chemically modified backbones designed to bind to their target mRNA and regulate protein expression55. Traditionally, this heteroduplex formation aims to decrease aberrant protein expression through various mechanisms, including RNase H mediated cleavage of the heteroduplex, splicing modulation, and steric hindrance of ribosomal binding55,56. ASOs have shown a good safety profile in numerous clinical trials, resulting in 10 FDA approved therapeutics56. The specific oligonucleotide, mRho ASO1, has the added benefit of previously demonstrating in vivo efficacy in reducing Rho mRNA in a dose-dependent manner following intravitreal injection38. Intravitreal administrations have become a standard procedure for the treatment of retinal diseases since the first FDA approved intravitreal injection therapeutic in 199857. With millions of intravitreal treatments performed annually, significant advancements have been made in injection-related procedures and tools to minimize patient discomfort and injection complications, making intravitreal injection the current preferred method of delivery for retinal disease therapy58.
To explore the therapeutic window of our approach, we evaluated two intervention time points, considering the wide variability in age of onset and severity for PRPH2 associated phenotypes6. For these experiments, we selected Prph2Y141C/+ mice, known for their slower rate of degeneration, allowing for a longer window of intervention and assessment7,14,59. Early intervention was initiated shortly after mice open their eyes at P15, while late intervention took place at P45 after full development of the retina and the initiation of functional decline in rods and cones7,14,59. Both early and late therapeutic interventions resulted in improved scotopic and photopic maximum a-wave responses. While it was expected that earlier intervention would yield superior functional rescue, the maximum a-wave values were similar between early and late intervention when evaluated at P60 and P90. This could possibly be due to a waning effect of the therapeutic benefit over time, as P15 intervention was administered 30 days earlier. Future studies involving repeated dosage administration may provide more insight into this aspect. However, no significant changes in maximum b-wave amplitudes were observed following mRho ASO1 treatment, despite observing improvements in b-wave responses following partial genetic ablation of RHO in Prph2Y141C/+ mice.
In addition to the observed improvements in ERG responses, early mRho ASO1 intervention led to significant increases in the number of photoreceptor nuclei. By administering the mRho ASO1 to modulate protein levels, we were able to optimize RHO reduction to achieve the proper RHO:PRPH2 ratios, resulting in maximum physiological response improvements. This dosage preserved the photoreceptors and slowed down the rate of degeneration. An inadequate reduction of RHO would lead to insignificant changes and minimal preservation, while excessive reduction of RHO would worsen photoreceptor death and degeneration. Late stage mRho ASO1 intervention failed to delay photoreceptor cell death. It is possible that, despite optimization for maximum functional improvement, P45 might be beyond the threshold of disease progression for mRho ASO1 to effectively improve photoreceptor survival. These findings further underscore the importance of maintaining proper RHO:PRPH2 ratios and choosing the appropriate time point for intervention to effectively prevent photoreceptor death using the reduction of RHO as a strategy.
In line with our observations in the Prph2 double mutants, ultrastructural analyses revealed that mRho ASO1 therapy improved rod outer segment ultrastructure throughout the retina, as evidenced by a reduction in large membranous whorls and sagittally oriented discs. Additionally, reduction of RHO following mRho ASO1 treatment reduced the structural abnormalities of increased open nascent discs and outer segment diameters associated with Prph2 mutations. Furthermore, we observed a potential decrease in microglia infiltration following late stage mRho ASO1 intervention at P90. As microglia infiltration is often observed in photoreceptor degenerative conditions60,61,62, this decrease further illustrates the overall benefit of mRho ASO1 on overall photoreceptor health and structure.
Taken together, these results signify the therapeutic efficacy of reducing levels of RHO as a strategy in impeding retinal degeneration caused by PRPH2 pathogenic variants. However, while these findings are exciting, there are limitations to consider when translating this approach into clinical relevance. The Prph2Y141C/+ knockin model has been shown to recapitulate many known patient phenotypes, including photoreceptor structural defects, decreased scotopic and photopic ERG values, and fundus flecking14. However, differences between human and mouse retinas hinder complete replication of the underlying disease mechanisms63,64,65. Additionally, genetic variations, variable penetrance, and diverse age-of-onset observed in humans19,20,21,22,23,24,66 present challenges in determining which patient would benefit from therapeutic intervention. Nonetheless, the strategy of reducing RHO levels holds promise as a preventative therapeutic approach for individuals with PRPH2 mutations, although long-term investigations in healthy control animals would be necessary before implementation.
Despite these limitations, these studies present significant findings that we aim to explore further in the future. We have introduced a distinctive strategy with the potential to yield therapeutic benefits across various PRPH2 pathogenic variants by targeting critical protein ratios essential for outer segment morphogenesis and structural stability. Future investigations will focus on expanding the application of this strategy to include additional Prph2 mutations, exploring the long-term effects of Rho knockdown in healthy controls, and assessing the feasibility of repeated dosing in long-term Prph2 mutants.
Methods
Study approvals and generation/acquisition of mutant lines
All animal experiments were approved by the University of Houston Institutional Animal Care and Use Committee (IACUC) and adhered to NIH guidelines and the recommendations from the Association for Research in Vision and Ophthalmology (Rockville, MD).
The knockin mouse models, Prph2K153∆/K153Δ and Prph2Y141C/Y141C, were generated and characterized previously13,14. Additionally, an in-house WT mouse strain was generated as described previously67. The homozygous Rho knockout mouse line (Rho−/−) was originally obtained as a generous gift from Dr. Janis Lem (Tufts University, Boston, MA, USA) and has been previously described34. Knockin heterozygotes (Prph2K153∆/+ and Prph2Y141C/+) mice were generated by crossing the WT mice with either Prph2K153∆/K153∆ or Prph2Y141C/Y141C mice, respectively. Double heterozygotes (Prph2K153∆/+/Rho+/- and Prph2Y141C/+/Rho+/-) were produced through crossbreeding of Prph2K153∆/K153∆ or Prph2Y141C/Y141C mice with Rho−/− mice. All mice were raised under cyclic light conditions, with a 12h-light/12-dark cycle (12 L/12D), and an illuminance of 30 lux. Mice were fed ad libitum mouse diet 9 F (5020) from LabDiet (Quakertown, PA, USA)). Mice were euthanized by carbon dioxide asphyxiation followed by cervical dislocation. Both mouse sexes were used in this study and no differences were observed between them.
The p.Tyr141Cys variant, identified as (c.422 A > G), has been previously characterized in patients22,23. Our Y141C knockin mouse model precisely carries the same mutation, Y141C (c.422 A > G), and has been well characterized7,14,68. The p.Lys154del variant, identified as (c.461-463del), was initially reported as 3-bp deletion at position 153/15419. The deletion affects one of the two conserved lysines in both human and mouse PRPH2 at positions 153 and 154, corresponding to c.458-460 and c.461-463 in human (see below). The human cDNA sequence at this site is AAG-AAG, and patients have a deletion of one of the AAG. The mouse cDNA at this site is AAA-AAG. To mirror the human condition, the K153∆ knockin model was generated by deleting AAA at position 153 site, resulting in a mutant protein lacking on of the two lysines. As a result, this mouse line aligns with both the initial patient report referring to the deletion of 153/15419, and the updated information in the Leiden Open Variation Database (LOVD) and in recent literature, where the mutation is identified as c.461_463del, p.Lys154del (db-ID PRPH2_000084)69. This mouse line, K153del, has been previously characterized13.
Human PRPH2
cDNA: 428-478 458 461
GAC ACA GAC ACC CCT GGC AGG TGT TTC ATG AAG AAG ACC ATC GAC ATG CTG
D T D T P G R C F M K K T I D M L
AA: 143-159
Mouse Prph2
cDNA:428-478 458 461
GAT ACG GAC ACC CCC GGC CGG TGC TTC ATG AAA AAG ACC ATC GAC ATG CTC
D T D T P G R C F M K K T I D M L
AA: 143-159
Electroretinography
Full field scotopic and photopic electroretinograms (ERGs) were recorded as described before70, with minor modifications using a UTAS system (LKC; Gaithersburg, MD). Eyes of overnight dark-adapted mice were dilated with 1% cyclopentolate solution (Bausch+Lomb; Bridgewater, NJ) prior to inducing anesthesia via intramuscular injection of 85 mg/kg ketamine (Covetrus; Portland, ME) and 14 mg/kg xylazine (Akorn, Inc.; Lake Forest, IL). The reference needle electrode was placed subdermally between the ears instead of the cheek as previously described70. Values obtained for ERG experiments using the double heterozygote animals were averaged and plotted. For injected animals, following the recording of the initial values, the probes were replaced, and a second measurement was taken to ensure observed trends were accurate. Only the initially recorded values were used for analysis.
Histology and morphometry
Histology and morphometric analyses were performed as previously described67. Briefly, eyes were enucleated from euthanized mice and fixed in modified Davidson’s fixative71 (12% formaldehyde, 15% ethanol, and 5% glacial acetic acid) overnight at 4 °C. Fixed eyes were then dehydrated and embedded in paraffin before being sectioned at 10 µm thickness. Sections containing the optic nerve head were stained with hematoxylin and eosin (H&E) (Sigma Aldrich, Burlington, MA, MHS16 and HT110116) and mounted with Permount (Fisher Scientific, Waltham, MA, P15-100) mounting medium. For morphometric analysis, 100 µm width window images were captured every 500 µm starting at the optic nerve head using a Zeiss Axioskop and a 20X objective lens. Image J was used for morphometric analysis by manually counting the number of nuclei in the 100 µm window images captured at the indicated distances. Representative images were taken using a 40X objective.
Protein extraction and quantification
Retinas from the indicated genotypes and treatment groups were extracted from euthanized mice and immediately flash frozen using liquid nitrogen before being stored at -80 °C until processing. Retinas were homogenized and then sonicated in 50 µL 1X RIPA Buffer (ABCAM, Boston, MA, ab156034) per retina. Samples were then incubated in the same extraction buffer over night at 4 °C. Following extraction, samples were centrifuged at 18,200 × g for 15 minutes at 4 °C to remove insoluble debris. Extracted protein concentrations were measured using the colorimetric Bradford Assay (Bio-Rad, Hercules, CA, #5000006).
Immunodot blots were used to quantify protein levels. Extract titrations were performed to determine the optimal range of total protein level needed from each genotype prior to quantification experiments. A MINIFOLD I microsample filtration manifold (Whatman® Schleicher & Schuell®, Keene, NH #27510) was used to load the protein extracts onto a nitrocellulose membrane (Bio-Rad, Hercules, CA #1620112). The membrane was allowed to dry before blocking with 5% non-fat milk in 1X TBST (0.1% Tween®). Following blocking, membranes were incubated with either unconjugated primary antibody (anti-RHO, -PRPH2, or -GFAP) or anti-actin primary antibody conjugated with Horseradish Peroxidase (HRP; Table S2) at room temperature. Membranes were washed prior to incubation with the appropriate HRP-conjugated secondary antibodies at room temperature. Following a final wash, the membrane was incubated with ECL reagent (SuperSignal™ West PICO PLUC Chemiluminescent Substrate #34577) for 1 minute, chemiluminescence imaging was performed using ChemiDocTM MP imaging system (Bio-Rad, Hercules, CA), and quantification was performed using Image Lab software v6.0.1 (Bio-Rad).
For protein quantification using immunodot blot, three to five replicates were used for each genotype and for each treatment. Each sample was independently measured in triplicate on each blot, averaged to be considered one value and presented as a mean + SD. For all immunoblotting quantification involving double heterozygote mice, chemiluminescent signal intensity values obtained for PRPH2, RHO, and GFAP were normalized by β-actin signal intensity before further normalization to the internal WT control. Quantification for the ASO treatment experiments involved normalization of measured RHO and PRPH2 chemiluminescent signal intensity values by β-actin signal intensity to obtain RHO/β-actin and PRPH2/β-actin values. Additionally, RHO was normalized by PRPH2 in order to obtain RHO:PRPH2 ratios.
Protein oligomerization was analyzed by performing non-reducing sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by immunoblotting using previously described methods14. Chemiluminescent signal intensity values obtained for each band were divided by the total lane signal intensity to obtain the percent distribution.
Transmission electron microscopy (TEM)
Fixation and processing of mouse eyes for TEM was performed as described previously40. In short, anesthetized mice were transcardially perfused with 2% paraformaldehyde, 2% glutaraldehyde and 0.05% calcium chloride in 50 mM MOPS (pH 7.4). Eyes were post-fixed for an additional 2 hour. Eyecups were embedded in 2.5% low-melt agarose (Precisionary Instruments, Greenville, NC) and sectioned on a Vibratome (VT1200S; Leica, Buffalo Grove, IL). 200 µm agarose sections were stained with 1% tannic acid (Electron Microscopy Sciences, Hatfield, PA) and 1% uranyl acetate (Electron Microscopy Sciences). Stained sections were gradually dehydrated with ethanol and infiltrated and embedded in Spurr’s resin (Electron Microscopy Sciences). 70 nm plastic sections were cut, placed on copper grids and stained with 2% uranyl acetate and 3.5% lead citrate (19314; Ted Pella, Redding, CA). Samples were imaged on a JEM-1400 electron microscope (JEOL, Peabody, MA) at 60 kV with a digital camera (BioSprint; AMT, Woburn, MA). Image analysis and processing was performed with ImageJ.
mRho ASO1 synthesis
A previously characterized Rho-specific ASO mRho ASO1 targeting exon 5 (5′-AGCTACTATGTGTTCCATTC−3′) are chimeric 20-mers with phosphorothioate backbone containing 20-O-methoxyethyl (MOE) modification at positions 1-5 and 15-20 was used in this study38. The control (5’ CCTATAGGACTATCCAGGAA- 3’) and mRho ASO1, were prepared by Integrated DNA Technologies, Inc. (Coralville, Iowa) using identical chemistry38. Synthesis was performed according to company’s standard operating procedures using proprietary processes. The control ASO has been previously employed as a control in various contexts: to suppress polypyrimidine tract binding protein 1 in neurons72, to increase microglial phagocytosis73, to deliver to the cornea74, to alleviate hepatic fibrosis75, in finding a protective factor in Dravet Syndrome76, in knockdown of GADD34 to ameliorate ALS77, to inhibit mTORC278 and to activate human frataxin expression79.
Intravitreal Injections
Intravitreal injections were performed as previously described80. Briefly, Prph2Y141C/+ mice were injected intravitreally at P30 for control ASO, P15 for early-stage intervention and P45 for late-stage intervention. Eyes were dilated with 1% cyclopentolate solution (Bausch + Lomb; Bridgewater, NJ) for 5 minutes prior to intramuscular injection of 85 mg/kg ketamine (Covetrus; Portland, ME) and 14 mg/kg xylazine (Akorn, Inc.; Lake Forest, IL). Adequate anesthesia was confirmed prior to injection by pedal withdrawal reflex assessment. A 30 gauge sterile needle was used to puncture the cornea and provide a guide hole. A Hamilton syringe was inserted through the guide hole to manually dispense material [mRho ASO1 or Endotoxin-Free Dulbecco’s PBS (1X) (EMD Millipore; Billerica, MA)] into the vitreal chamber. Volumes injected for all studies were 0.5 µL in P15 animals and 1.0 µL in P45 animals. Dosages used for P15 titration studies included 1.5625 µg, 3.125 µg, 6.25 µg, 12.5 µg, and 25 µg, while dosages used for P45 titrations included 3.125 µg, 6.25 µg, 12.5 µg, 25 µg, and 50 µg. Following dosage titration studies, a concentration of 6.25 µg/µL was used for all injections so that P15 mice received 3.125 µg mRho ASO per injection while P45 mice received 6.25 µg mRho ASO per injection. Following injection, Triple antibiotic ointment (Taro pharmaceuticals Inc., Hawthorne, NY, USA) was applied to each eye. All animals were closely monitored during recovery from anesthesia until fully ambulatory and provided adequate access to food and water. Mice experiencing injection related complications, such as significant accumulation of blood in the vitreous chamber, retinal detachment, damage to the iris or lens, intraocular infection, or cataract development were excluded from the study. All injections were performed in a sterile surgical suite. In the titration studies, both eyes were injected for all mice with either mRho ASO1 or 1X PBS vehicle control. Due to procedure related difficulties experienced during the titration studies when attempting to inject both eyes at P15, all characterization studies were performed injecting one eye with 3.125 µg ASO and the other serving as an uninjected contralateral control. All P45 characterization studies were performed using the same procedure as titration studies with 6.25 µg ASO.
Immunofluorescence
Immunofluorescence was performed as previously described59. Briefly, mice were euthanized by carbon dioxide asphyxiation followed by cervical dislocation, eyes were enucleated, and then fixed in modified Davidson’s fixative overnight at 4 °C71. Fixed eyes were then dehydrated and embedded in paraffin before being sectioned at 10 µm thickness. Before use, samples were incubated in xylene to remove paraffin and tissue was rehydrated in sequential ethanol dilutions. Following rehydration, antigen retrieval was performed by incubating samples in sodium citrate at 100 °C for 30 minutes before being placed in blocking buffer (5% bovine serum albumin, 1% donkey serum, and 0.5% Triton X-100 in PBS) for 1 hour. Following blocking, antigen labeling was performed by incubating sections with primary antibodies overnight, washing with 1X PBS, incubating with secondary antibodies for 2 hours, and then washing with 1X PBS. DAPI staining was then performed for 15 minutes at a concentration of 0.1 µg/mL (Thermo Fisher Scientific, Waltham, MA). Slides were mounted using anti-fade mounting media and sealed with clear nail polish. Imaging was performed using a 20x objective on a Zeiss LSM800 confocal microscope (Zeiss, White Plains, NY). Image processing was performed in Zen 3 lite software.
Antibodies
All antibodies used for immunoblotting and immunofluorescence are listed in Supplementary Table 2.
Statistical analysis
Statistical Analysis was performed using GraphPad Prism version 8 (GraphPad Software, La Jolla, CA). One-way ANOVA with Tukey’s post-hoc comparisons was employed for quantitative analysis between groups for all double mutant ERG data and comparison of protein levels in double mutant figures. Two-way ANOVA with Tukey’s post-hoc comparisons was employed for quantitative analysis between groups for all morphometry analysis. ASO treated ERGs were compared using two-tailed Mann–Whitney U test. Student’s two-tailed t-test was used for statistical analysis of protein quantification studies. Significance was set at P < 0.05. Pairwise comparison significances are indicated as *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001 throughout the text.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data associated with this study are presented in the paper or the Supplementary Materials. Source data are provided with this paper Source data are provided with this paper.
Change history
19 July 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41467-024-50140-3
References
Nuzbrokh, Y., Ragi, S. D. & Tsang, S. H. Gene therapy for inherited retinal diseases. Ann. Transl. Med. 9, 1278 (2021).
Georgiou, M., Fujinami, K. & Michaelides, M. Inherited retinal diseases: Therapeutics, clinical trials and end points-A review. Clin. Exp. Ophthalmol. 49, 270–288 (2021).
Sanjurjo-Soriano, C. & Kalatzis, V. Guiding lights in genome editing for inherited retinal disorders: implications for gene and cell therapy. Neural plasticity 2018, 5056279 (2018).
Nuzbrokh, Y., Kassotis, A. S., Ragi, S. D., Jauregui, R. & Tsang, S. H. Treatment-emergent adverse events in gene therapy trials for inherited retinal diseases: a narrative review. Ophthalmol. Ther. 9, 709–724 (2020).
Conley, S. M. et al. Delineating the clinical phenotype of patients with the c.629C>G, p.Pro210Arg mutation in peripherin-2. Investig. Ophthalmol. Vis. Sci. 63, 19 (2022).
Coco-Martin R. M., Sanchez-Tocino H. T., Desco C., Usategui-Martin R., Telleria J. J. PRPH2-related retinal diseases: broadening the clinical spectrum and describing a new mutation. Genes 11, 773 (2020).
Conley, S. M., Stuck, M. W., Watson, J. N. & Naash, M. I. Rom1 converts Y141C-Prph2-associated pattern dystrophy to retinitis pigmentosa. Hum. Mol. Genet. 26, 509–518 (2017).
Loewen, C. J., Moritz, O. L. & Molday, R. S. Molecular characterization of peripherin-2 and rom-1 mutants responsible for digenic retinitis pigmentosa. J. Biol. Chem. 276, 22388–22396 (2001).
Loewen, C. J., Moritz, O. L., Tam, B. M., Papermaster, D. S. & Molday, R. S. The role of subunit assembly in peripherin-2 targeting to rod photoreceptor disk membranes and retinitis pigmentosa. Mol. Biol. cell 14, 3400–3413 (2003).
Nour, M., Ding, X. Q., Stricker, H., Fliesler, S. J. & Naash, M. I. Modulating expression of peripherin/rds in transgenic mice: critical levels and the effect of overexpression. Investig. Ophthalmol. Vis. Sci. 45, 2514–2521 (2004).
Hawkins, R. K., Jansen, H. G. & Sanyal, S. Development and degeneration of retina in rds mutant mice: photoreceptor abnormalities in the heterozygotes. Exp. eye Res. 41, 701–720 (1985).
Cheng, T. et al. The effect of peripherin/rds haploinsufficiency on rod and cone photoreceptors. J. Neurosci. Off. J. Soc. Neurosci. 17, 8118–8128 (1997).
Chakraborty, D., Conley, S. M., Zulliger, R. & Naash, M. I. The K153Del PRPH2 mutation differentially impacts photoreceptor structure and function. Hum. Mol. Genet. 25, 3500–3514 (2016).
Stuck, M. W., Conley, S. M. & Naash, M. I. The Y141C knockin mutation in RDS leads to complex phenotypes in the mouse. Hum. Mol. Genet. 23, 6260–6274 (2014).
Ding, X. Q. et al. The R172W mutation in peripherin/rds causes a cone-rod dystrophy in transgenic mice. Hum. Mol. Genet. 13, 2075–2087 (2004).
Chakraborty, D. et al. Novel molecular mechanisms for Prph2-associated pattern dystrophy. FASEB J.: Off. Publ. Federation Am. Societies Exp. Biol. 34, 1211–1230 (2020).
Stricker, H. M., Ding, X. Q., Quiambao, A., Fliesler, S. J. & Naash, M. I. The Cys214->Ser mutation in peripherin/rds causes a loss-of-function phenotype in transgenic mice. Biochem. J. 388, 605–613 (2005).
Kedzierski, W. et al. Deficiency of rds/peripherin causes photoreceptor death in mouse models of digenic and dominant retinitis pigmentosa. Proc. Natl Acad. Sci. USA 98, 7718–7723 (2001).
Weleber, R. G., Carr, R. E., Murphey, W. H., Sheffield, V. C. & Stone, E. M. Phenotypic variation including retinitis pigmentosa, pattern dystrophy, and fundus flavimaculatus in a single family with a deletion of codon 153 or 154 of the peripherin/RDS gene. Arch. Ophthalmol. 111, 1531–1542 (1993).
Boon, C. J. et al. Mutations in the peripherin/RDS gene are an important cause of multifocal pattern dystrophy simulating STGD1/fundus flavimaculatus. Br. J. Ophthalmol. 91, 1504–1511 (2007).
Francis, P. J. et al. Genetic and phenotypic heterogeneity in pattern dystrophy. Br. J. Ophthalmol. 89, 1115–1119 (2005).
Yang, Z. et al. A novel RDS/peripherin gene mutation associated with diverse macular phenotypes. Ophthalmic Genet. 25, 133–145 (2004).
Vaclavik, V., Tran, H. V., Gaillard, M. C., Schorderet, D. F. & Munier, F. L. Pattern dystrophy with high intrafamilial variability associated with Y141C mutation in the peripherin/RDS gene and successful treatment of subfoveal CNV related to multifocal pattern type with anti-VEGF (ranibizumab) intravitreal injections. Retina 32, 1942–1949 (2012).
Dryja, T. P., Hahn, L. B., Kajiwara, K. & Berson, E. L. Dominant and digenic mutations in the peripherin/RDS and ROM1 genes in retinitis pigmentosa. Investigative Ophthalmol. Vis. Sci. 38, 1972–1982 (1997).
Wouters, O. J., McKee, M. & Luyten, J. Estimated research and development investment needed to bring a new medicine to market, 2009-2018. Jama 323, 844–853 (2020).
Wen, X. H. et al. Overexpression of rhodopsin alters the structure and photoresponse of rod photoreceptors. Biophys. J. 96, 939–950 (2009).
Chakraborty, D., Conley, S. M., Al-Ubaidi, M. R. & Naash, M. I. Initiation of rod outer segment disc formation requires RDS. PloS one 9, e98939 (2014).
Rakshit, T. & Park, P. S. Impact of reduced rhodopsin expression on the structure of rod outer segment disc membranes. Biochemistry 54, 2885–2894 (2015).
Jastrzebska, B. et al. Rhodopsin-transducin heteropentamer: three-dimensional structure and biochemical characterization. J. Struct. Biol. 176, 387–394 (2011).
Toda, K., Bush, R. A., Humphries, P. & Sieving, P. A. The electroretinogram of the rhodopsin knockout mouse. Vis. Neurosci. 16, 391–398 (1999).
Sung, C. H., Makino, C., Baylor, D. & Nathans, J. A rhodopsin gene mutation responsible for autosomal dominant retinitis pigmentosa results in a protein that is defective in localization to the photoreceptor outer segment. J. Neurosci. Off. J. Soc. Neurosci. 14, 5818–5833 (1994).
Tan, E. et al. The relationship between opsin overexpression and photoreceptor degeneration. Investigative Ophthalmol. Vis. Sci. 42, 589–600 (2001).
Hollingsworth, T. J. & Gross, A. K. Defective trafficking of rhodopsin and its role in retinal degenerations. Int. Rev. cell Mol. Biol. 293, 1–44 (2012).
Lem, J. et al. Morphological, physiological, and biochemical changes in rhodopsin knockout mice. Proc. Natl Acad. Sci. USA 96, 736–741 (1999).
Makino, C. L. et al. Rhodopsin expression level affects rod outer segment morphology and photoresponse kinetics. PloS one 7, e37832 (2012).
Humphries, M. M. et al. Retinopathy induced in mice by targeted disruption of the rhodopsin gene. Nat. Genet. 15, 216–219 (1997).
Lewis, T. R. et al. Photoreceptor Disc Enclosure Is Tightly Controlled by Peripherin-2 Oligomerization. J. Neurosci. Off. J. Soc. Neurosci. 41, 3588–3596 (2021).
Murray, S. F. et al. Allele-Specific Inhibition of Rhodopsin With an Antisense Oligonucleotide Slows Photoreceptor Cell Degeneration. Inv. Ophthalmol. Vis. Sci. 56, 6362–6375 (2015).
Keen, T. J. & Inglehearn, C. F. Mutations and polymorphisms in the human peripherin-RDS gene and their involvement in inherited retinal degeneration. Hum. Mutat. 8, 297–303 (1996).
Ding, J. D., Salinas, R. Y. & Arshavsky, V. Y. Discs of mammalian rod photoreceptors form through the membrane evagination mechanism. J. Cell Biol. 211, 495–502 (2015).
Conley, S. M. et al. Prph2 initiates outer segment morphogenesis but maturation requires Prph2/Rom1 oligomerization. Hum. Mol. Genet. 28, 459–475 (2019).
Bringmann, A. & Reichenbach, A. Role of Muller cells in retinal degenerations. Front. Biosci. J. virtual Libr. 6, E72–E92 (2001).
Bringmann, A. et al. Muller cells in the healthy and diseased retina. Prog. Retinal Eye Res. 25, 397–424 (2006).
de Hoz, R. et al. Retinal macroglial responses in health and disease. BioMed. Res. Int. 2016, 2954721 (2016).
Fernandez-Sanchez, L., Lax, P., Campello, L., Pinilla, I. & Cuenca, N. Astrocytes and muller cell alterations during retinal degeneration in a transgenic rat model of retinitis pigmentosa. Front. Cell. Neurosci. 9, 484 (2015).
Wensel, T. G. et al. Structural and molecular bases of rod photoreceptor morphogenesis and disease. Prog. Retinal Eye Res. 55, 32–51 (2016).
Kocaoglu, O. P. et al. Photoreceptor disc shedding in the living human eye. Biomed. Opt. Express 7, 4554–4568 (2016).
Spencer, W. J., Lewis, T. R., Pearring, J. N. & Arshavsky, V. Y. Photoreceptor discs: built like ectosomes. Trends Cell Biol. 30, 904–915 (2020).
Kedzierski, W., Lloyd, M., Birch, D. G., Bok, D. & Travis, G. H. Generation and analysis of transgenic mice expressing P216L-substituted rds/peripherin in rod photoreceptors. Investigative Ophthalmol. Vis. Sci. 38, 498–509 (1997).
McNally, N. et al. Murine model of autosomal dominant retinitis pigmentosa generated by targeted deletion at codon 307 of the rds-peripherin gene. Hum. Mol. Genet. 11, 1005–1016 (2002).
Sahel, J. A., Marazova, K. & Audo, I. Clinical characteristics and current therapies for inherited retinal degenerations. Cold Spring Harb. Perspect. Med. 5, a017111 (2014).
Newton F., Megaw R. Mechanisms of photoreceptor death in retinitis pigmentosa. Genes 11, 1120 (2020).
Kelley, R. A. et al. Ablation of the riboflavin-binding protein retbindin reduces flavin levels and leads to progressive and dose-dependent degeneration of rods and cones. J. Biol. Chem. 292, 21023–21034 (2017).
Leveillard, T. et al. Identification and characterization of rod-derived cone viability factor. Nat. Genet. 36, 755–759 (2004).
Rinaldi, C. & Wood, M. J. A. Antisense oligonucleotides: the next frontier for treatment of neurological disorders. Nat. Rev. Neurol. 14, 9–21 (2018).
Xiong H., Veedu R. N.& Diermeier S. D. Recent advances in oligonucleotide therapeutics in oncology. Int. Mol. Sciences 22, 3295 (2021).
Hartman, R. R. & Kompella, U. B. Intravitreal, subretinal, and suprachoroidal injections: evolution of microneedles for drug delivery. J. Ocul. Pharmacol. Therapeutics: Off. J. Assoc. Ocul. Pharmacol. Therapeutics 34, 141–153 (2018).
Petri, A. S. et al. Intravitreal injections with vascular endothelial growth factor inhibitors: a practical approach. Ophthalmol. Ther. 9, 191–203 (2020).
Genc A. M. et al. Retbindin: A riboflavin Binding Protein, Is Critical for Photoreceptor Homeostasis and Survival in Models of Retinal Degeneration. Int. J. Mol. Sci. 21, 8083 (2020).
Yu, C., Roubeix, C., Sennlaub, F. & Saban, D. R. Microglia versus monocytes: distinct roles in degenerative diseases of the retina. Trends Neurosci. 43, 433–449 (2020).
Makabe, K., Sugita, S., Mandai, M., Futatsugi, Y. & Takahashi, M. Microglia dynamics in retinitis pigmentosa model: formation of fundus whitening and autofluorescence as an indicator of activity of retinal degeneration. Sci. Rep. 10, 14700 (2020).
Huang, H., Parlier, R., Shen, J. K., Lutty, G. A. & Vinores, S. A. VEGF receptor blockade markedly reduces retinal microglia/macrophage infiltration into laser-induced CNV. PloS one 8, e71808 (2013).
Peichl, L. Diversity of mammalian photoreceptor properties: adaptations to habitat and lifestyle? Anat. Rec. Part A, Discoveries Mol., Cell., Evolut. Biol. 287, 1001–1012 (2005).
Huberman, A. D. & Niell, C. M. What can mice tell us about how vision works? Trends Neurosci. 34, 464–473 (2011).
Volland, S., Esteve-Rudd, J., Hoo, J., Yee, C. & Williams, D. S. A comparison of some organizational characteristics of the mouse central retina and the human macula. PloS one 10, e0125631 (2015).
Boon, C. J. et al. The spectrum of retinal dystrophies caused by mutations in the peripherin/RDS gene. Prog. retinal eye Res. 27, 213–235 (2008).
Strayve, D. et al. ROM1 contributes to phenotypic heterogeneity in PRPH2-associated retinal disease. Hum. Mol. Genet. 29, 2708–2722 (2020).
Ikelle, L. et al. Correction: Comparative study of PRPH2 D2 loop mutants reveals divergent disease mechanism in rods and cones. Cell Mol. Life Sci. 80, 290 (2023).
Reeves, M. J. et al. Genotype-phenotype associations in a large PRPH2-related retinopathy cohort. Hum. Mutat. 41, 1528–1539 (2020).
Nour, M., Quiambao, A. B., Peterson, W. M., Al-Ubaidi, M. R. & Naash, M. I. P2Y(2) receptor agonist INS37217 enhances functional recovery after detachment caused by subretinal injection in normal and rds mice. Investigative Ophthalmol. Vis. Sci. 44, 4505–4514 (2003).
Latendresse, J. R., Warbrittion, A. R., Jonassen, H. & Creasy, D. M. Fixation of testes and eyes using a modified Davidson’s fluid: comparison with Bouin’s fluid and conventional Davidson’s fluid. Toxicol. Pathol. 30, 524–533 (2002).
Maimon, R. et al. Therapeutically viable generation of neurons with antisense oligonucleotide suppression of PTB. Nat. Neurosci. 24, 1089–1099 (2021).
Schoch K. M., et al. Acute Trem2 reduction triggers increased microglial phagocytosis, slowing amyloid deposition in mice. Proc. Nat. Acad. Sci. USA 118, e2100356118 (2021).
Chau, V. Q. et al. Delivery of antisense oligonucleotides to the cornea. Nucleic Acid Ther. 30, 207–214 (2020).
Win, S. et al. Hepatic mitochondrial SAB deletion or knockdown alleviates diet-induced metabolic syndrome, steatohepatitis, and hepatic fibrosis. Hepatology 74, 3127–3145 (2021).
Lenk, G. M. et al. Scn8a Antisense Oligonucleotide Is Protective in Mouse Models of SCN8A Encephalopathy and Dravet Syndrome. Ann. Neurol. 87, 339–346 (2020).
Ghadge, G. D. et al. Knockdown of GADD34 in neonatal mutant SOD1 mice ameliorates ALS. Neurobiol. Dis. 136, 104702 (2020).
Chen, C. J. et al. Therapeutic inhibition of mTORC2 rescues the behavioral and neurophysiological abnormalities associated with Pten-deficiency. Nat. Med. 25, 1684–1690 (2019).
Shen, X. et al. Efficient electroporation of neuronal cells using synthetic oligonucleotides: identifying duplex RNA and antisense oligonucleotide activators of human frataxin expression. RNA 25, 1118–1129 (2019).
Eblimit A. et al. Co-injection of sulfotyrosine facilitates retinal uptake of hyaluronic acid nanospheres following intravitreal injection. Pharmaceutics 13, 1510 (2021).
Acknowledgements
All authors acknowledge Larissa Ikelle and Drs. Lars Tebbe and Shannon Conley for their thoughtful review and valuable insights on the manuscript. This work was supported by grants from the NIH/NEI [EY010609 (MIN and MRA), EY034671 (MIN), EY030451 (VYA), EY005722 (VYA), and EY033763 (TRL). NIH had no role in the collection, analyses, or interpretation of the data; in the writing of the manuscript or in the decision to submit the manuscript for publication.
Author information
Authors and Affiliations
Contributions
C.R.W., M.I.N and M.R.A designed the study. C.R.W. gathered and executed most of the experiments, analyzed the data and wrote the first draft of the manuscript. M.S.M performed and analyzed the P17 and P30 double heterozygote ERG studies, nuclear counts, intravitreal injections of the control ASO, and the blots in Fig. S2. R.C. performed intravitreal injections of the Rho-ASO and participated in the ASO titration experiments. SZ was responsible for obtaining and analyzing the control ASO data presented in Figs. S6, S7 and S8. T.R.L., C.M.C. and V.Y.A were responsible for all of the EM experiments and the data presented in Figs. 3, 8, S9, S10, S11, and S12 and Table 1, M.K. performed GFAP IHC presented in Fig. S3A, MIN and MRA edited the manuscript to its final form.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rutan Woods, C.T., Makia, M.S., Lewis, T.R. et al. Downregulation of rhodopsin is an effective therapeutic strategy in ameliorating peripherin-2-associated inherited retinal disorders. Nat Commun 15, 4756 (2024). https://doi.org/10.1038/s41467-024-48846-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-024-48846-5
This article is cited by
-
Anesthetic effects on electrophysiological responses across the visual pathway
Scientific Reports (2024)
