
Abstract
Gammaherpesviruses are DNA tumor viruses that establish lifelong latent infections in lymphocytes. For viruses such as Epstein-Barr virus and murine gammaherpesvirus 68, this is accomplished through a viral gene-expression program that promotes cellular proliferation and differentiation, especially of germinal center B cells. Intrinsic host mechanisms that control virus-driven cellular expansion are incompletely defined. Using a small-animal model of gammaherpesvirus pathogenesis, we demonstrate in vivo that the tumor suppressor p53 is activated specifically in B cells latently infected by murine gammaherpesvirus 68. In the absence of p53, the early expansion of murine gammaherpesvirus 68 latency greatly increases, especially in germinal center B cells, a cell type whose proliferation is conversely restricted by p53. We identify the B cell-specific latency gene M2, a viral promoter of germinal center B cell differentiation, as a viral protein sufficient to elicit a p53-dependent anti-proliferative response caused by Src-family kinase activation. We further demonstrate that Epstein-Barr virus-encoded latent membrane protein 1 similarly triggers a p53 response in primary B cells. Our data highlight a model in which gammaherpesvirus latency gene-expression programs that promote B cell proliferation and differentiation to facilitate viral colonization of the host trigger aberrant cellular proliferation that is controlled by p53.
Similar content being viewed by others
Introduction
Gammaherpesviruses (GHVs) are DNA tumor viruses that include the human pathogens Epstein-Barr virus (EBV) and Kaposi sarcoma-associated herpesvirus (KSHV) and the rodent pathogen murine gammaherpesvirus 68 (MHV68), among others. Following a period of productive viral replication in various host tissues, GHVs establish lifelong chronic infections1,2. This phase of the infectious cycle, referred to as latency, is maintained in B cells. The lifelong infections caused by GHVs place the host at risk for numerous cancers, including Burkitt lymphoma, Hodgkin lymphoma, primary effusion lymphoma, multicentric Castleman disease, and many others3,4. Moreover, GHV-related malignancies represent serious complications for AIDS patients and transplant recipients5. However, the precise mechanisms by which GHVs promote oncogenesis are incompletely defined, as are the host pathways that prevent cellular transformation due to GHV infection.
To establish and maintain latency, GHVs stimulate and usurp normal B cell differentiation processes6. This is accomplished through the actions of viral noncoding RNAs and oncoproteins, which, upon infection of naïve B cells, lead to an activated phenotype reminiscent of a germinal center (GC) reaction7,8. GC B cells are highly proliferative, with an estimated doubling time of every 6 hours9. GC B cells also undergo mutagenic processes known as somatic hypermutation and class-switch recombination, which change the antigenic specificity and effector functions of the B cell receptor and secreted antibodies10. The GC reaction ultimately results in the emergence of long-lived memory B cells or antibody-secreting plasma cells that respectively enable recall responses upon reinfection and facilitate elimination of pathogens11. It is hypothesized that stimulating GC reactions enables GHV-infected B cells to increase in number dramatically as the cells rapidly proliferate while also allowing a mechanism for the establishment of long-term latency in the circulating memory B cell pool12. This model of GHV latency establishment is largely based on EBV infection and immortalization of primary B cells in culture, but it is also supported by kinetic evaluations of cell types that harbor MHV68 in vivo and through analyses of mutant viruses following experimental infection of mice [reviewed in refs. 13,14].
For instance, the MHV68 homolog of the latency-associated nuclear antigen (mLANA) encoded by ORF73 is essential for latency establishment after intranasal inoculation of mice15,16,17. In addition to mediating maintenance of the viral episome during latent cell division, mLANA also promotes c-Myc stabilization, which likely influences latent cell survival18. The ORF72-encoded viral cyclin D ortholog (v-Cyclin) is a bona fide oncogene19, and it is important for viral replication and reactivation from latency20,21. A viral Bcl-2 homolog (v-Bcl-2) encoded by the M11 open-reading frame, prevents activation-induced cell death to support viral latency22,23,24. The M2-derived gene product is a scaffold protein that in and of itself promotes B cell differentiation by activating Src family kinases, phospholipase C-gamma (PLC), and downstream transcription via nuclear factor of activated T cells (NFAT) and interferon regulatory factor 4 (IRF4)25,26,27,28. Interestingly, M2’s role in facilitating MHV68 latency establishment appears to function almost exclusively in the GC B cell compartment29,30. EBV likewise uses a coordinated gene expression program that facilitates latency establishment and promotes cellular proliferation and differentiation1. Of note, latent membrane protein 1 (LMP1) mimics the activating and pro-survival signaling functions of CD4031,32,33, a B cell surface receptor that interacts with CD40L on T follicular helper cells to facilitate GC reactions10. M2 and LMP1 are hypothesized to be functional homologs despite no discernable sequence homology14.
While many of the viral oncogenes that promote cellular proliferation, survival, and differentiation have been studied extensively, less is known regarding the potential for cellular tumor suppressor pathways to be activated in response to manipulation of host-cell physiology by GHV proteins during latency establishment. It is noteworthy that EBV, KSHV, and MHV68 each encode viral proteins capable of inducing cell-cycle entry and transformation19,32,34,35,36, yet none of these viruses efficiently transforms infected cells. This suggests that the viral latency-associated gene expression programs induce host cell responses that limit the potential for cellular transformation during infection. For example, although most, if not all cells are infected and begin to proliferate after EBV infection of primary B cells in culture, only a small percentage of infected cells immortalize to become lymphoblastoid cell lines (LCLs)37,38,39,40. This is because EBV-induced cellular proliferation triggers the activation of ataxia telangiectasia mutated (ATM) and checkpoint kinase 2 (Chk2), DNA damage response (DDR) kinases that inhibit cellular proliferation and subsequent immortalization37. KSHV also triggers ATM activation and proliferative arrest upon infection of primary endothelial cells41. Like EBV, MHV68 can immortalize primary cells in culture42. Although host restriction factors are not yet known, the process is similarly inefficient, requiring weeks of culture prior to the outgrowth of transformed cells. While DDR pathways appear to restrict GHV-mediated cellular transformation in culture, tumor suppressor responses that restrict GHV infection and immortalization of cells in vivo during natural infection of a host are poorly defined and may differ from responses occurring in cell culture. For instance, a role for ATM in restricting MHV68 infection in vivo was tested; however, ATM deficiency did not promote enhanced latent MHV68 infection or cancer43. Rather, ATM facilitates B cell latency44,45, suggesting that ATM does not impose a critical barrier to latent MHV68 infection or related oncogenesis in vivo.
Another candidate host protein for intrinsic latency restriction is tumor suppressor p53. p53 is considered a master regulator of genetic integrity, a postulate emphasized by the presence of inactivating p53 mutations in approximately half of all human cancers46. Though constitutively expressed, p53, under normal physiologic cellular conditions, is constantly targeted for proteolytic degradation by proteins such as ubiquitin ligase mouse double-minute 2 (MDM2)47. In response to potentially genotoxic cellular stresses, including DNA damage, oncogene expression, and viral infection, p53 becomes stabilized and activated through a variety of post-translational modifications47. Active p53 regulates transcription of numerous cellular genes involved in cell-cycle arrest, DNA repair, and apoptosis48. Interestingly, p53 is stabilized, and p53-responsive transcripts are induced during B cell immortalization by EBV49, and the KSHV-encoded cyclin D ortholog is sufficient in and of itself to induce p5334,41,50. Exogenous induction of p53 by MDM2 inhibition reduces LCL formation by EBV51, and conversely, p53 inhibition facilitates endothelial cell proliferation following KSHV infection41. Although both EBV and KSHV encode latency proteins demonstrated to inhibit p53 in various biochemical evaluations52,53,54, established cell lines in which these viral inhibitors of p53 are highly expressed remain responsive to p53 agonists49,55, that these proteins inefficiently suppress p53 functions in latently infected cells. Moreover, it is somewhat counterintuitive that a virus that establishes a life-long chronic infection would universally inhibit a tumor suppressor as critically important as p53. However, p53 mutations do occur frequently in endemic Burkitt lymphoma (eBL), a cancer characterized by stable EBV infection and the presence of chromosomal translocations, which suggests that p53 functions limit GHV-related cancers in vivo2. Whether p53 is activated to limit GHV infection and tumorigenic potential has not been directly tested using in vivo models of pathogenesis.
Here, we describe experiments that test the hypothesis that p53 is activated in response to GHV manipulation of B cell physiology during latency establishment. Using MHV68 infection of WT and p53-deficient mice, we define the functions of p53 in controlling viral latency establishment and maintenance in vivo. We employed a targeted sufficiency screen in primary B cells to identify specific viral gene products from MHV68 and EBV that are latency-related activators of p53, and we confirmed the MHV68 p53 agonist by comparing mutant and WT MHV68 in vivo. Finally, we performed mutational and pharmacologic inhibitor analyses that highlight key signaling events by which the viral factors promote p53 activation. Through this work, we demonstrate that virus-mediated manipulation of B cell activation and differentiation is countered by an intrinsic host tumor suppressor response to limit early proliferative events during latency establishment by a DNA tumor virus.
Results
MHV68 induces p53 during latency establishment
Tumor suppressor p53 is activated in response to aberrant cellular proliferation, differentiation, and genotoxic stress, especially when induced by cellular and viral oncogenes1,34,56. In order to establish latency, GHVs encode proteins that promote cellular proliferation and differentiation3,4,13, which could hypothetically be recognized by the p53 pathway as potentially oncogenic events. While it is known that p53 is induced by GHV infection of many cell types in culture41,57, its activity is putatively inhibited by certain latency gene products1,56. However, whether p53 is activated and functional during GHV latency establishment in a living host is not known.
To begin to define the relationship between p53 and GHV latency, we infected mice with an MHV68 recombinant virus that encodes YFP to enable direct identification of infected cells by flow cytometry58 and evaluated p53 induction in spleens of infected mice on day 16 after infection. At this time point, acute viral replication has waned, latent viral burdens in the spleen are at their peak, and the majority of infected B cells are proliferating and exhibit a germinal center phenotype58,59,60. In this analysis, p53 levels were significantly higher specifically in the MHV68-infected cells (YFP+) than in mock-infected animals or the uninfected cells (YFP-) in spleens of infected mice (Fig. 1a, b). As a downstream transcriptional target of the type I-interferon (IFN-I) signaling pathway during viral infections61, we reasoned it possible that p53 was simply activated as part of the innate antiviral response to MHV68 infection; to determine if p53 activation during MHV68 latency was due to IFN-I signaling, we infected mice lacking the type I-IFN receptor (IFNAR1-/-). Again, on day 16 post-infection p53 induction was evident almost exclusively in YFP+ splenocytes at levels that were equivalent in both IFNAR+/+ and IFNAR1-/- mice (Supplementary Fig. 1). These data indicate that p53 activation during MHV68 latency establishment is independent of IFN-I signaling and occurs specifically within infected cells.

a Flow cytometry analysis of p53 induction in infected (YFP +, red) and uninfected (YFP−, blue) spleen cells on day 16 after mock-infection (gray) or infection of C57BL/6 mice with 104 PFU of H2B-YFP MHV68. b Quantification of data in (a) represented as mean +/− SEM. One-way ANOVA p < 0.0001. Each dot represents an individual mouse. 2-3 independent infections with a minimum of 3 mice per group. c, d Quantitative RT-PCR analyses comparing MHV68 infected to uninfected splenocytes after sorting on day 16 post-infection. The virus-dependent genetic marking strategy to identify infected cells and confirmation of infection is shown in Supplementary Fig. 2. Viral latency transcript abundance is shown in (c). Fold change represented as mean +/− SD. Profiler p53 pathway array data are shown in (d). Relative transcript abundance was determined by comparing virus-positive to virus-negative cells after sorting using the ∆∆CT method. The trp53 transcript is highlighted in red. Transcripts previously evaluated in MHV68 infection are denoted with blue bars.
Since p53 primarily functions as a transcription factor once stabilized, we next sought to evaluate p53-related transcription within latently infected cells. We infected inducible fluorescent reporter mice62 with a recombinant MHV68 virus that expresses Cre recombinase to induce fluorescent protein expression63,64 as a means to mark infected cells. After sorting fluorescent cells and confirming infection by limiting-dilution PCR (LD-PCR)65 and p53 induction by flow cytometry (Supplementary Fig. 2a, c), we performed p53 pathway RT-PCR array analyses comparing infected cells, which were approximately 96% CD19+/B220+ B cells (Supplementary Fig. 2b), to uninfected B cells from infected animals. Further confirming that the cells were indeed infected, viral latency transcripts were highly enriched in the fluorescent cell population, as were numerous p53-associated transcripts (Fig. 1c, d). Of note, the p53 transcript Trp53 was ca. 100-fold higher in infected cells compared to uninfected cells, as were other transcripts with previously demonstrated roles in MHV68 latency, such as Myc, Il6, and Nfkb118,66,67. Together these data support the conclusion that p53 is stabilized and transcriptionally active in vivo during MHV68 latency establishment. The observation that p53 induction occurred almost exclusively in infected cells using two independent marking strategies, as well as IFNAR1-/- mice, demonstrates that p53 activation is not a general consequence of viral infection as might be expected of a typical antiviral response, but rather represents a cell-intrinsic reaction to MHV68 infection.
MHV68 latency establishment is enhanced in p53-deficient mice
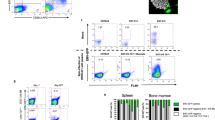
Given that p53 induction was specific to infected cells, we hypothesized that p53 functions in an intrinsic cellular response that controls MHV68 latency establishment. If correct, we predicted that the absence of p53 would correlate with enhanced MHV68 latency establishment. To test this hypothesis, we evaluated MHV68 infection of both p53+/+ and p53-/- mice in side-by-side comparisons. Remarkably, on day 16 post-infection nearly 8% of cells in spleens of p53-/- animals were YFP+ compared to 0.6% of cells in WT mice (Fig. 2a). In agreement with YFP+ percentages, the frequency of splenocytes harboring MHV68 genomes was 14-fold higher in p53-/- animals than WT littermates, with approximately 1 in 20 p53-/- cells infected vs. 1 in ca. 280 WT cells (Fig. 2b, d). Moreover, immunofluorescence analyses of spleen sections revealed that infected cells were more readily detected in B220+ B cell follicles of p53-null mice in comparison to WT mice (Fig. 2e and Supplementary Fig. 4).

a Flow cytometry analyses to determine the percentage of infected splenocytes (YFP +) for each mouse genotype at 16 days after mock infection or infection with 104 PFU of H2B-YFP MHV68. Data represent means +/− SEM for 2-3 independent infections with a minimum of 3 mice per group. Each dot represents an individual mouse, n = 4-5. Welch’s two-sided t test, **p < 0.005. b Quantification of latently infected splenocyte frequencies by limiting-dilution PCR on day 16 post-infection. p53-/- or p53+/+ mice were infected intranasally with 104 PFU of H2B-YFP MHV68. Data represent means +/− SEM for 2 independent infections with a minimum of 3 mice per group. Extra sum-of-squares F test p < 0.0001, F = 40.36 (1, 22). c Quantification of latent MHV68 reactivation efficiency from infected p53-/- or p53+/+ splenocytes on day 16 post-infection. Reactivation was detected by scoring cytopathic effect in a limiting-dilution ex vivo culture. Data represent means +/− SEM for 2-3 independent infections with a minimum of 3 mice per group. Extra sum-of-squares F test p = 0.0024, F = 10.36 (1, 46). Replicate graphs in Supplementary Fig. 3. d table of values from (a, and c). e, Immunohistochemical visualization of B cells (B220+) and MHV68 infected cells (YFP+) in spleen sections from p53-/- or p53+/+ mice on day 16 after infection. Representative images of splenic follicles are shown. Scale bars indicate 100 μm. Additional representative images in Supplementary Fig. 4.
Previous studies suggest that p53 plays roles in innate and adaptive immune responses to certain infections68,69,70, so we reasoned mice lacking p53 might be less able to control MHV68 infection. In evaluating acute viral replication, we found no difference in MHV68 infection of WT and p53 knockout mouse lungs on day 7 post-infection (Supplementary Fig. 3a), the typical peak of acute viral replication in this tissue71. This result suggests that p53 is not critical for controlling the acute phase of MHV68 infection in mice. While the frequency of latently infected splenocytes with the capacity to reactivate upon ex vivo culture was enhanced modestly compared to WT mice (Fig. 2c and Supplementary Fig. 3b), the increase correlated with the higher frequency of cells harboring MHV68 in p53-/- mice (Fig. 2d), and we did not detect evidence of ongoing persistent viral replication72,73. The absence of increased acute replication, enhanced reactivation, or persistent replication, outcomes largely controlled by innate immune responses74,75,76,77,78, suggests that p53 is not directly involved in these host defense mechanisms. Finally, MHV68-specific adaptive immune responses also developed normally in p53-/- mice, as virus-specific IgG in serum, T cell activation, and induction of MHV68 antigen-specific CD8+ T cells were comparable in both WT and p53-null animals infected with MHV68 during latency establishment (day 16) and maintenance (day 42) as immune activation wanes (Supplementary Figs. 5 and 6). Together, these data demonstrate that the absence of p53 correlates with enhanced MHV68 latency establishment and supports the hypothesis that p53 mediates intrinsic cellular resistance to chronic MHV68 infection.
p53 controls cellular proliferation during MHV68 latency establishment
As mentioned above, MHV68 and other GHVs encode latency-associated gene products that promote the expansion of B cells that serve as reservoirs for latent infection. For MHV68 this results in a syndrome akin to infectious mononucleosis caused by EBV79. In addition to enhanced latent MHV68 infection in p53-deficient animals, it was readily apparent that there was also a significantly greater infection-related increase in the number of cells per spleen compared to WT mice (Fig. 3a). This observation suggested that p53 acts to control cellular expansion induced by the virus. In support of this hypothesis, quantification of specific lymphocyte populations by flow cytometry revealed enhanced expansion of B cells in p53-/- mice (Fig. 3b), while T cell and natural killer (NK) cell numbers were similar between WT and knockout animals (Supplementary Fig. 6). For particular B cell subsets, the number of GC B cells was significantly increased (Fig. 3c), but plasmablasts were only modestly affected by the absence of p53 (Fig. 3d). Reflecting MHV68’s latent tropism for B cells and usage of the GC B cell compartment during latency establishment, the number of infected B cells was significantly increased in p53 knockout mice, with the MHV68 infected subset of GC B cells exhibiting a ca. 10-fold relative increase (Fig. 3e–h). Infection of plasmablasts also was significantly enhanced, which may result from the differentiation of infected GC B cells. In contrast to infection of p53-/- mice with WT virus, infection with a latency-deficient mutant virus lacking the MHV68 homolog of the latency-associated nuclear antigen (mLANA-null) did not cause increased expansion of splenocyte populations (Supplementary Fig. 7). Since this mutant virus can still undergo acute replication and stimulate adaptive immunity15,16, these data suggest that the enhanced splenocyte expansion that occurs in p53-/- mice requires the presence of latent MHV68 and is not simply a consequence of the general immune response to viral infection.

a–d Quantification of total splenocytes, B cells, GC B cells, and plasmablasts 16 days after mock infection or infection of p53+/+ or p53-/- mice with 104 PFU of H2B-YFP MHV68. Flow cytometry was performed to identify specific B cell populations. B cells were gated as CD19+/B220+ in (b) GC cells as GL7+/CD38lo sub-population of CD19+/B220+ B cells in (c) and plasmablasts as CD138+/B220lo in (d). Gating strategies are shown in Supplementary Fig. 9. Other lymphocyte populations were also quantified and were equivalent in p53-/- and p53+/+ spleens (Supplementary Fig. 6). e–h Quantification of infected (YFP+ ) total splenocytes, B cells, GC B cells, and plasmablasts as identified in (a–d) on day 16 after infection. a–h Data represent means +/− SEM for 2-3 independent infections with a minimum of 3 mice per group. Each dot represents an individual mouse. Mann-Whitney unpaired two-sided t test, *p < 0.05, **p < 0.005, ***p < 0.0005, ****p < 0.0001.
Given that GHVs drive B cell proliferation in order to establish and maintain latency12,13 and p53 restricts cellular proliferation or promotes apoptosis downstream of oncogene expression56,80,81,82, we hypothesized that p53 functions to limit MHV68-driven cellular proliferation and/or induce death in infected cells. Using in vivo 5-ethynyl-2’-deoxyuridine (EdU)-incorporation to label cells actively replicating their DNA, we detected similar basal levels of EdU incorporation in both p53-/- and WT animals that were mock-infected. Infection with MHV68 led to an increase in the number of EdU+ GC B cells irrespective of genotype; however, spleens of p53-deficient mice contained twice as many EdU+ GC B cells than WT mice (Fig. 4a). The number of apoptotic cells after infection was only modestly reduced in the absence of p53 as determined by flow cytometry-based active caspase 3/7 staining (Fig. 4b). These data suggest that a mechanism by which p53 limits MHV68 latency establishment is through restriction of cellular proliferation, an interpretation supported by the enhanced expansion and infection of the highly proliferative GC B cell compartment in p53-/- mice.

a Quantification of GC B cells undergoing DNA replication on day 16 after mock infection or infection of p53-/- and p53+/+ mice with MHV68. EdU was injected intraperitoneally 2 h prior to sacrifice to label newly replicated DNA. Click chemistry was performed and EdU + GC B cells (gated as CD19+/B220+/GL7+/CD38lo) were detected by flow cytometry. Total (left panel) and percent EdU + GC B cells per spleen (right panel) are shown. Each dot represents an individual mouse, n = 4. b Flow cytometric quantification of GC B cells (gated as CD19+B220+GL7hiCD38lo) undergoing apoptosis on day 16 after infection of p53-/- and p53+/+ mice with MHV68. Apoptotic cells were identified as active caspase 3/7+ after ex vivo staining. Gating strategies are shown in Supplementary Fig. 10. Data represent means +/− SEM. Mann-Whitney unpaired two-sided t test, * p < 0.05, **p < 0.005. Each dot represents an individual mouse, n = 4-5.
Long-term MHV68 latency is not enhanced in p53-deficient mice
After the initial virus-driven B cell activation and expansion associated with the establishment of MHV68 latent infection, both total cell numbers and the frequency of cells harboring viral genomes contract to levels that are maintained at relatively stable levels during long-term infection59,60,83. In testing the impact of p53 on long-term MHV68 latency, we found that viral genome-positive cells in spleens of p53-/- mice were ca. 5-fold lower than WT mice on day 42 post-infection, a phenotype that was maintained up to 100 days post-infection (Supplementary Fig. 8a, b). The total number of cells, including total B cells and GC and plasmablast subsets, were equivalent in spleens of both knockout and WT animals by day 42 after infection (Supplementary Fig. 8c–f). These data suggest that p53 is mainly activated to restrict virus-driven B cell expansion during latency establishment and does not play a critical role in controlling infection as latency is maintained over time. Moreover, it is notable that despite restricting initial latent colonization of the host, the modest reduction in latent viral burdens observed at later time points suggests that p53 induction during latency establishment has a pro-viral role in long-term latent infection by MHV68.
The M2 latency protein promotes p53 activation in primary B cells
Since p53 was stabilized specifically in MHV68 infected cells where it appeared to restrict GC B cell expansion during early latent infection, we hypothesized that viral proteins involved in latency establishment are responsible for activating p53 in B cells. To test this hypothesis, we performed a screen in which primary B cells were transduced with retroviruses that encode the major MHV68 latency genes, M2, M11/v-BCL2, ORF72/v-Cyclin, and ORF73/mLANA15,16,22,84,85 and evaluated p53 induction by flow cytometry. Frameshift-stop mutant versions of each gene were used as negative controls, and high-level transcription of each construct was verified by quantitative RT-PCR (Supplementary Fig. 11a–c). In this screen, only the M2-encoding retrovirus led to p53 induction (Fig. 5a). Similar to MHV68 infected splenocytes, increased p53 detection was only observed in M2 transduced cells and not bystander cells within the culture (Supplementary Fig. 11e), which further supports the notion that p53 induction is an intrinsic cellular response to viral gene expression and does not occur through paracrine mechanisms.

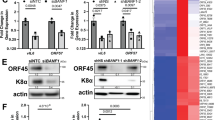
a, b Flow cytometry analysis of p53 induction in primary murine B cells after transduction with retroviruses that encode MHV68 latency proteins. Representative histograms and mean fluorescence intensities of p53 staining in transduced cells are shown. Data in (a) show p53 staining 4 days after transduction of cells with retroviruses that encode v-Cyclin, LANA, M2, M11, or a frame-shift/translation-stop mutant M2 (M2.Stop) as a negative control. Data are biological replicates, n = 2. Data in (b) show p53 staining 2 days after transduction with M2.Stop, M2-WT, or a tyrosine mutant M2-Y120F/Y129F (M2-Tyr). Data represent mean ± SEM. Two-tailed Student’s t test, ns - not significant, *p < 0.05, and ****p < 0.0001. The experiment was performed in technical triplicate. Data in (c) show p53 staining 2 days after transduction with M2.Stop or M2-encoding retroviruses in the presence of DMSO (1:1000), mouse IgG (20 μg/mL), PP2 (10 μM), PP3 (10 μM), CsA (500 ng/mL), or rat anti-IL10R (20 μg/mL). Inhibitors were added 24 h post-transduction. Data represent mean ± SEM. Ordinary one-way ANOVA, ns - not significant and ****p < 0.0001. The experiment was performed in technical triplicate. Gating strategies and additional controls are shown in Supplementary Fig. 11. d, e RNA-seq was performed 4 days after transduction of primary B cells with M2 or M2.Stop retroviruses to evaluate changes in transcription. Data in (e) show gene set enrichment analysis (GSEA) and (d) hallmark gene z-score heatmaps comparing M2 to M2.Stop for the hallmark p53 gene set. Statistical scores are inset into the top right of analysis images. NES normalized enrichment score, q-val FDR-adjusted p-value. The genes presented were derived from GSEA and DEseq2 analysis of all genes with significant expression changes where the expression level increased from M2 to M2.Stop at least 1.5-fold and decrease from M2 to M2.Stop at least 1.5-fold. A table of hallmark gene sets with significant changes caused by M2 is shown in Supplementary Fig. 14.
The M2 latency protein of MHV68 functions similarly to EBV latent membrane proteins to promote B cell proliferation and differentiation1,86. M2 acts as a scaffold protein that activates Src-family kinases28,87, triggering a signaling cascade that results in NFATc1 and IRF4-dependent production of the cytokine IL-1026. This signal transduction is mediated through a structural motif encompassing two tyrosine residues (Y120 and Y129)25,27. M2 function is specifically required in GC B cells to facilitate MHV68 latency establishment30,88, and its expression in naive B cells results in a GC-like activation profile and BCR class switching that is dependent on IL-1086. Given that oncogenes are well known to activate cellular tumor suppressor responses89, we reasoned that the M2-driven signal transduction pathway would be recognized as a potentially oncogenic stimulus by the cell, resulting in p53 activity. To test this hypothesis, we transduced primary B cells with control, WT M2-encoding retroviruses, and an M2 tyrosine mutant (Y120F/Y129F) to disrupt M2-dependent cellular activation. Unlike B cells transduced with retrovirus encoding WT M2, those transduced with M2 mutant (M2Tyr) retrovirus did not exhibit p53 induction (Fig. 5b), suggesting that residues in M2 required for signal transduction are responsible for activating p53 in primary B cells.
To further define the molecular pathways involved in M2-driven activation of p53, we transduced primary B cells with control or M2-expressing retrovirus in the presence or absence of specific pharmacologic inhibitors or an IL-10 inhibitory antibody using concentrations previously described to inhibit M2-related effects on B cells26. Remarkably, Src inhibitor PP2 completely blocked M2-mediated p53 induction while its inactive control analog PP3 had no effect (Fig. 5c). Cyclosporin A (CsA), an inhibitor of the calcineurin/NFAT pathway, partially prevented p53 induction, however M2-mediated p53 induction still occurred in the presence of an antibody that blocks the IL-10 receptor. While we cannot rule out potential off-target effects of the reagents tested, together the M2Tyr mutant and pharmacologic inhibitor data strongly suggest that p53 activation is primarily mediated by M2’s activation of a Src-family kinase signal transduction cascade. This is also consistent with Src kinases acting upstream of NFAT and IL-10 upon expression of M2 in B cells, which agrees with previous studies26.
Detecting p53 induction after expression of M2 in B cells seemed to contradict prior work suggesting that M2 promotes cellular proliferation and survival26,86,90. We, therefore, performed RNA-seq experiments as an unbiased means to evaluate transcription pathways regulated by M2 expression in B cells. In agreement with previous analyses26, M2 transduction led to high-level transcription of Il10 and Irf4 (Fig. 5d). In comparisons of control to M2-transduced cells, gene set enrichment analysis (GSEA) revealed a significant positive enrichment of p53 hallmark transcripts upon M2 expression (Fig. 5d, e and Supplementary Fig. 14). These studies, therefore, demonstrate that M2 causes p53 induction and transcriptional activity in primary murine B cells.
MHV68 M2 induces the p53 pathway in vivo
Having defined M2 as sufficient to promote p53 induction and transcription in primary B cells in culture, we next sought to determine whether M2 promotes p53 activity in vivo. M2 functions primarily in GC B cells to facilitate latency establishment88, which presented a complication since this was also the primary cell type impacted by the presence of p53 during MHV68 infection. We, therefore, utilized two specific reporters to facilitate comparisons between cells infected with either WT or M2-null MHV68. To mark GC B cells, we used AID-Cre Ai14 reporter mice, which express TdTomato upon expression of activation-induced cytidine deaminase (AID), a B cell-specific enzyme required for somatic hypermutation and class-switch recombination during GC reactions62,91. To identify infected cells, we utilized WT and M2.Stop MHV68 reporter viruses in which the enzyme beta-lactamase was fused to the episome-maintenance protein mLANA (73.Bla)59, which is highly expressed in GC B cells92. This reporter was selected based on compatibility with TdTomato in flow cytometry analysis, the increased sensitivity to detect infected cells due to enzymatic amplification of fluorescent signal upon cleavage of the beta-lactamase substrate59, and its compatibility with live-cell sorting for downstream RNA sequencing. The reporters used and the necessity for live-cell sorting, unfortunately, prevented a direct evaluation of p53 protein levels in these experiments. While M2 is essential for efficient viral dissemination and splenic latency establishment93,94,95, M2-deficient viruses are detected in draining lymph nodes at levels comparable to WT MHV68 after IN infection88. We therefore infected AID-Cre Ai14 reporter mice with either WT 73.Bla or M2.Stop 73.Bla MHV68 and sorted infected AID+Bla+ and uninfected AID+Bla- cells from mediastinal lymph nodes for ultra-low-input RNA-seq (Fig. 6a). Principle component analyses revealed that each specific group exhibited unique, non-overlapping transcription profiles that were similar between biological replicates within groups (Fig. 6b). Viral latency transcripts were highly enriched in Bla+ data sets of both M2.Stop and WT MHV68 (Fig. 6c), however, only AID+Bla+ samples from WT MHV68 infection showed increased transcription of Il10 and Irf4 (Fig. 6d), which is consistent with previous work demonstrating that M2 upregulates their expression26,86. Significant negative enrichment in Hallmark p53 Pathway GSEA comparisons between Bla+ M2.Stop and WT MHV68 infected B cells revealed that p53-related transcripts were generally lower in infected AID+ cells when M2 expression was ablated (Fig. 6e, f). The majority of differentially regulated p53-related transcripts in this comparison were also induced by M2 expression alone in primary B cells (highlighted in red text). Together with the demonstration that M2 in and of itself has the capacity to induce p53 and downstream transcription, these results strongly suggest that M2 is both necessary and sufficient to promote p53 pathway activation in infected B cells.

AID-cre Ai14 mice were infected intranasally with 104 PFU of WT or M2.Stop 73.Bla MHV68. At 14 dpi infection, draining lymph nodes were isolated and treated with CCF4-AM. MHV68+ cells are defined by cleavage of CCF4 fluorescent substrate (Bla+). a Sorting strategy for AID+ 73.Bla MHV68 lymphocytes. The AID+ uninfected cell population marked in red and AID+ Bla+ cell population marked in blue. b–f Low input RNA-seq was performed on WT or M2.Stop MHV68 sorted cells to evaluate changes in transcription. b Principle component analysis (PCA) of low input RNA-seq data. c Viral latency transcript abundance compared to uninfected AID+ lymphocytes. d M2-associated transcript expression. Z-score heatmap comparing Il10 and Irf4 expression in WT (UI and I) and M2.Stop (UI and I) populations. Data in (e) show gene set enrichment analysis (GSEA) comparing WT to M2.Stop MHV68 infection for the hallmark p53 gene set. Statistical scores are inset into the bottom left of analysis images. NES, normalized enrichment score. q-val, FDR-adjusted p-value. f Z-score heatmaps show average expression data for WT or M2.Stop MHV68 AID+ lymphocytes. The genes presented were derived from GSEA and DEseq2 analysis of all genes with significant expression changes where the expression level increased from WT to M2.Stop at least 1.5-fold and decrease from WT to M2.Stop at least 1.5-fold. M2-associated transcripts identified in Fig. 5 are denoted in red.
p53 controls M2-driven cellular proliferation
Given p53’s known roles in restricting cell-cycle progression, M2-specific p53 pathway induction both in culture and in vivo, and the apparent p53-related control of GC B cell replication during MHV68 latency establishment in mice, we reasoned that p53 was restricting M2-driven B cell proliferation. To test this hypothesis, we evaluated EdU incorporation after M2-encoding retroviral transduction of WT and p53-/- primary B cells. Confirming our hypothesis, EdU incorporation assays showed a large increase in the percentage of p53-/- cells undergoing DNA replication compared to WT cells upon M2 expression (Fig. 7a). As in WT cells, EdU incorporation was potently reduced by Src kinase inhibitor PP2 in p53-deficient cells, which further highlights the importance of Src family kinases in M2-mediated B cell activation (Fig. 7b). Importantly, RNA-seq analyses revealed that p53 hallmark genes, which include numerous anti-proliferative transcripts, were negatively enriched in p53-/- B cells after M2 transduction (Fig. 7c, d and Supplementary Fig. 14), while target genes of the cell-cycle promoting E2F pathway were induced in the absence of p53 (Fig. 7e, f). M2-responding transcripts Irf4 and Il10 remained highly induced in p53-/- cells, suggesting that p53 does not directly influence transcription via the Src/NFAT/calcineurin pathway activated by M2 (Supplementary Fig. 12). Together, these data provide evidence that M2 drives Src family kinase-related cellular proliferation that is restricted by p53 activation in primary B cells.

a, b Quantification of p53-/- or p53+/+ B cells undergoing DNA replication after transduction with M2 or M2.Stop retroviruses. Cells were pulsed with 10 mM EdU for 4 h prior to harvest at 50 h post-transduction. Cells in (b) were treated with Src-family inhibitor PP2 (10 μM) or controls (DMSO at 1:1000, PP3 at 10 μM) 24 h post-transduction. EdU incorporation in newly synthesized DNA was labeled by Click chemistry. EdU+ transduced B cells (gated on ZsGreen+ cells) were detected by flow cytometry. Data represent means +/− SEM. Two-tailed Student’s t test, ns - not significant, *p < 0.05, ** p < 0.01. The experiment was performed in technical triplicate. Representative flow cytometry plots and additional controls are shown in Supplementary Fig. 12. c–f RNA-seq was performed 4 days after transduction of p53-/- or p53+/+ B cells with M2 or M2.Stop retroviruses to evaluate changes in transcription. Data in (c, d) show gene set enrichment analysis (GSEA) and hallmark gene z-score heatmaps comparing M2 to M2.Stop for the hallmark p53 gene set. Data in (e, f) show the hallmark E2F gene set. Statistical scores are inset into the top right of analysis images. NES, normalized enrichment score. q-val, FDR-adjusted p-value. d, f z-score heatmaps show average expression data for M2 or M2.Stop expressing primary B cells. The genes presented were derived from GSEA and DEseq2 analysis of all genes with significant expression changes where the expression level increased from M2 to M2.Stop at least 1.5-fold and decrease from M2 to M2.Stop at least 1.5-fold. A table of hallmark gene sets with significant changes caused by M2 are shown in Supplementary Fig. 14.
EBV LMP1 causes p53 activation in murine primary B cells
Analogous to MHV68, EBV induces GC-like responses in infected B cells, which the virus usurps to facilitate latency establishment6,8,96. In particular, the EBV “growth program” (also known as Latency III) involves the coordinated expression of several latency factors that lead to B cell activation, proliferation, and eventual immortalization into lymphoblastoid cell lines (LCLs)97,98. Prior studies demonstrated that EBV infection of primary human B cells elicited p53 stabilization49, and p53 agonists limit LCL formation upon EBV infection51. We, therefore, hypothesized that distinct latency factors involved in the EBV growth program may trigger p53 induction, and we focused our analysis on LMP1 due to its functional similarities to M21 and previous reports that LMP1 expression correlates with p53 stabilization99,100. Consistent with our hypothesis, transduction of primary murine B cells with retroviruses encoding LMP1 resulted in increased p53 detection by flow cytometry (Fig. 8a). In contrast, dominant-negative LMP1 (LMP1AAAG) encoding four point-mutations in residues critical for activating NF-κB, STAT, and AP-1 pathways101 did not activate p53 in transduced B cells (Fig. 8b), which suggests that LMP1-dependent signal transduction leads to p53 activation. We again performed RNA-seq analyses on transduced cells and found that p53-related transcripts were significantly enriched in LMP1-transduced cells relative to control transductions (Fig. 8d, e), as were c-Myc signature transcripts (Supplementary Fig. 14). Comparisons of RNA-seq data from p53 competent and knockout cells expressing LMP1 indicated that enrichment of both c-Myc and p53-related transcription in these cells is linked to p53 expression (Supplementary Figs. 13d, e and 14). Interestingly, both LMP1- and M2-transduced p53-/- B cells formed large clusters reminiscent of an activated, blasting phenotype (Fig. 8c) that proliferated and survived in culture for 2-3 weeks (Supplementary Fig. 13b), whereas transduced WT cells could not be maintained. These data demonstrate that EBV-encoded LMP1 elicits p53 activation in mouse B cells in a manner that is consistent with p53 restricting cellular proliferation and survival. This parallel observation for an EBV latency protein suggests that antagonism of the virus-driven GC response by p53 is a common feature of GHV latency establishment.

a, b Flow cytometry analysis of p53 induction in primary murine B cells 4 days after transduction with LMP1, M2, LMP1 dominant-negative cell signaling mutant (LMP1AAAG) or M2.Stop retroviruses. Representative histograms and mean fluorescence intensities of p53 staining in transduced cells are shown. Data represent means +/− SEM. Two-tailed Student’s t test, **p < 0.01. The experiment was performed in technical triplicate. Gating strategies are shown in Supplementary Fig. 12. c Transduced p53-/- B cells were imaged 48 h post-transduction. The scale bar indicates 250 μm. d, e RNA-seq was performed 4 days after transduction of primary B cells with LMP1 or M2.Stop (negative control) retroviruses to evaluate changes in transcription. Data show gene set enrichment analysis (GSEA) and hallmark gene z-score heatmaps comparing LMP1 to M2.Stop for the hallmark p53 gene set. The reference list was derived from Hallmark gene sets and compared with a pre-ranked list (by fold) of global average gene expression. Statistical scores are inset into the top right of analysis images. NES, normalized enrichment score. q-val, FDR-adjusted p-value. e z-score heatmaps show average expression data for LMP1 or M2.Stop expressing primary B cells. The genes presented were derived from GSEA and DEseq2 analysis of all genes with significant expression changes where the expression level increased from LMP1 to M2.Stop at least 1-fold and decrease from LMP1 to M2.Stop at least 1-fold. RNA-seq analysis of LMP1 transduced p53-/- primary murine B cells and a table of hallmark gene sets with significant changes caused by LMP1 are shown in Supplementary Figs. 13 and 14. f Schematic of the model in which viral latency proteins that drive B cell proliferation and differentiation lead to the activation of p53. Lab, F. (2024) https://BioRender.com/b70y129.
Discussion
GHVs take advantage of B cell proliferation and differentiation to facilitate the establishment of life-long latent infection of their hosts. This is accomplished through a coordinated latency gene-expression program that augments and mimics natural developmental processes that guide B cell activation and differentiation as part of the adaptive immune response to infection. We demonstrate here, using an in vivo model of GHV infection that p53 is induced specifically in infected cells during the period in which a murine GHV, MHV68, manipulates the B cell compartment to colonize its host. Although p53 is thought to be repressed by Bcl-6 in GC B cells to facilitate rapid cellular proliferation and mutagenesis of the B cell receptor102, the absence of p53 correlated with large increases in the number of GC B cells, their proliferation, and infection of these cells by MHV68. Further supported by the detection of p53-responsive transcripts within latently infected cells, these data strongly suggest that p53 remains responsive in the GC B cell compartment and is capable of restricting GHV-driven B cell expansion as the virus establishes latency. Since p53 stabilization occurs during EBV infection of primary human B cells49, p53 agonists restrict their immortalization51, and LMP1 induces p53, we propose that p53 is a common intrinsic inhibitor of GHV latency establishment in B cells.
As a master regulator of cell-stress responses, p53 activation during viral infection has been evaluated for numerous viruses, such as influenza virus, herpes simplex virus (HSV), human cytomegalovirus (HCMV), HIV, SV40, adenovirus, and others, including MHV68 and EBV49,81,103,104,105. In fact, p53 was initially identified through its interaction with the SV40 oncoprotein, Large T antigen (Tag)106. It is important to emphasize that these studies primarily focused on p53 function and inhibition during the productive replication phase of the viral infection cycle. For instance, p53 is potently inhibited by Large Tag, adenovirus E1B-55K/E4orf6, and MHV68 mLANA and muSOX during lytic replication104,107. Identical productive viral replication in both WT and p53-/- mice highlights the potent inhibition of p53 during acute MHV68 replication. In contrast, p53 is an interferon-stimulated gene that participates in the antiviral response and host survival during HSV-1 infection of mice108, whereas HCMV integrates p53 activity to promote lytic gene expression and viral replication105. While well-studied in productive replication of numerous and diverse virus infections, surprisingly little is known regarding p53 function in the latent phase of herpesvirus infection. Certainly, prior studies suggest that some GHV latency proteins inhibit p53 (summarized below), but whether interactions between a GHV and p53 impact latency in a living host was previously unexplored. To the best of our knowledge, the data presented here represent the first in vivo evidence of an antagonistic relationship between host p53 and the GHV latency-associated transcription program that influences viral latency outcomes.
Since the absence of p53 correlated with increased GC proliferation and early viral latency establishment, our data strongly suggest that p53 is not simply inhibited by viral latency factors, an idea that has long been proposed in the GHV field, but that has become somewhat controversial in recent years. For instance, EBV nuclear antigen-1 (EBNA-1), a DNA-binding protein that facilitates maintenance of the latent viral episome, is expressed broadly in EBV cancers and hypothesized to facilitate cellular immortalization by promoting p53 proteolysis via an interaction with ubiquitin-specific protease 7 (USP7)54,109. However, despite the expression of EBNA-1 and in agreement with our findings, p53 is activated early during EBV infection of primary B cells, and LCLs remain responsive to p53 agonists49,51. Although the KSHV episome maintenance protein LANA inhibits p53 in overexpression systems52,110,111, p53 remains functional in PEL cells that contain high levels of LANA protein55,112. While p53 can be inhibited by the MHV68 LANA homolog, the inhibitory function appears to be restricted to the lytic replication cycle, where it requires an undefined activating event and is assisted by additional lytic-phase viral proteins57,104.
Given that inactivation of p53 by mutation or suppression is predisposing to cellular transformation, it is perhaps not surprising that viruses that establish lifelong chronic infections as part of their maintenance strategy would evolve to not block p53 function within latently infected cells. Presumably, potent inhibition of p53 would reduce host survival and limit viral transmission. However, it is important to also emphasize that this idea does not rule out the possibility that p53 responses are toned down by viral latency factors either transiently or subtly to enable GHVs to overcome p53-dependent barriers to latency establishment during initial colonization of host lymphoid tissues. It is additionally notable that p53 is frequently mutated in eBL2, suggesting that p53 function is a critical determinant of pathogenesis for this disease. p53 knockout mice are primarily predisposed to thymic lymphoma and sarcomas113, whereas people with p53 mutations exhibit a broad spectrum of cancers114. The differences in tumor spectrum between mice and humans are surely influenced by the types of p53 mutations present, but it is also possible that infectious agents influence the types of cancers that develop when p53 is absent or nonfunctional. Whether GHV infection influences the tumor phenotype of hosts when p53 is non-functional is unknown but could be tested in a p53-deficient setting using the mouse model, as we suspect that enhanced B cell proliferation during MHV68 infection of p53-deficient animals is a hallmark of incipient lymphomagenesis.
With regard to a specific mechanism for p53 induction during GHV latency establishment, increased GC expansion during infection of p53 knockout mice supports a model in which viral latency proteins that drive B cell proliferation and differentiation lead to the activation of p53 (Fig. 8f). In agreement with known responses to viral oncogene expression56,80,81, we suspected that p53 would then serve to counteract the proliferative expansion of latently infected cells promoted by the viral latency program. Indeed, identification of the latency-specific gene product M2, which in and of itself promotes GC B cell expansion30,86, as a potent p53 activator in primary B cells directly supports this hypothesis. M2 serves as a molecular scaffolding protein that binds and activates Src-family kinases27,28,87. This leads to the induction of NFAT/calcineurin signaling pathways that promote IL-10 production and B cell differentiation26. Using M2 mutants and previously published inhibitors, we provide evidence that signal transduction via Src-family kinases is the key event for triggering p53 upon M2 expression, and not IL-10-related effects on the cell. It is well established that aberrant cellular activation and cell-cycle progression via oncogenes such as H-Ras and c-Myc triggers tumor suppressor responses, especially activation of p53-dependent inhibition of cell-cycle progression115,116. Our data, therefore, support the conclusion that M2/Src activity is recognized by an infected B cell as a potential oncogenic threat, leading to p53 activity that restricts infected cell proliferation. Since p53 was equivalently induced in latently infected mice lacking a functional IFN-I receptor, we also conclude that p53 activation in this setting is not part of a traditional interferon-stimulated transcription response.
It will be of interest to determine if MHV68, like EBV37,117,118, elicits ATM and/or ATM and Rad3-related (ATR) DNA damage responses as a mechanism of p53 phosphorylation and stabilization or if other kinases fulfill this role. For EBV, ATM and ATR are activated in response to a period of hyperproliferation and replication stress and limit the immortalization of infected B cells in culture37,119,120. The functions of ATM in MHV68 infection of mice have been evaluated, and MHV68’s relationship with ATM is complex in that ATM expression in B cells does not restrict, but rather facilitates viral latency establishment43,44. A specific role for ATR, which is primarily activated by single-strand DNA breaks that occur as a result of DNA replication stress121, would support the notion that aberrant cell-cycle progression and DNA replication caused by M2/Src is a stimulus for p53 activation. In further support of this notion, a previous RNA-seq analysis of MHV68-infected splenocytes revealed enhanced transcription of genes involved in the ATR signaling pathway, in addition to p53-related transcription122. While the ATR hypothesis is compelling, dissecting this pathway in vivo would be challenging, as ATR-deficient mice exhibit progeria syndrome and immune dysfunction123.
Although M2 is unique to MHV68, it exhibits functional similarities to latency proteins encoded by human GHVs, such as EBV LMP1/LMP2A and KSHV K1/K15, especially integration of signaling events that occur upon B cell receptor and co-stimulatory molecule crosslinking1,96,124,125,126,127. Combined expression of EBV LMP1 and LMP2A in a transgenic mouse model caused GC B cell expansion and B cell differentiation into rapidly growing plasmablasts that have enriched expression of E2F targets and G2M checkpoint genes128, which is analogous to M2 effects on B cells86. Both LMP1 and LMP2A mimic aspects of B cell signaling, but only LMP1 is a bona fide oncogene36,40,99,129, which motivated our testing of LMP1-mediated p53 induction. Depending on the system being tested, LMP1 is reported to both induce p53-stabilization and cell-cycle inhibition or, conversely, to prevent p53 function130,131. Our data support the hypothesis that a p53 anti-proliferative response is activated in response to LMP1, where it prevents the outgrowth of transduced B cells. This interpretation is further supported by the observation that p53 stabilization is enhanced in lymphomas occurring in a transgenic mouse model in which LMP1 is constitutively expressed in B cells99 and also by studies noting that LMP1 enhances p53 stabilization in cells expressing Large T antigen100 and LMP1 expression levels strongly correlate with p53 levels in NPC132. In light of our results, we suspect that LMP1-driven B cell activation triggers p53 in an attempt to control tumor cell growth. While the percentage of cells with stabilized p53 was not evaluated in the transgenic mouse model99, it is interesting to note that tumors were also characterized by NF-κB activation, which is necessary to overcome p53-mediated growth restriction during LCL formation51. Moving forward, it will be of interest to determine if LMP2A, K1, and K15 (and other human GHV latency proteins) also activate p53. Since LMP2A reduces LMP1 hyperactivation of B cells133 and combined LMP1/LMP2A-deficient EBV exhibits diminished transforming capacity in a cord blood model of EBV lymphomagenesis134, it will also be important to investigate whether LMP2A suppresses p53 induction by LMP1.
An important question that remains unresolved is how p53 can restrict early latency expansion, while also supporting long-term latency. This is further complicated by the counterintuitive observation that pro-latency viral gene products serve as triggers for p53. In total, our data suggest that a dynamic exists between p53 and viral latency genes that is analogous to an automobile’s gas pedal and brake – one accelerating the process while the other helps maintain appropriate control (Fig. 8f). The cellular activating functions of the viral latency genes facilitate progression of the latently infected cell from point A (initial infection of a naïve B cell) to point B (progression through the GC reaction to enable long-term latency in memory B cells or reactivation from plasma cells). While beneficial to the virus in the long run, this is inherently a potentially dangerous method for promoting chronic infection as enhanced proliferative and/or differentiation signal transduction are recognized hallmarks of cellular transformation that should activate tumor suppressor responses in non-transformed cells.
In its most basic function, p53 is a cytoprotective molecule, promoting in response to a perceived threat the production of cell-cycle inhibitors that pause DNA replication and cellular proliferation until it is safe (genomically or metabolically) for such cellular processes to continue. If the threat persists or is catastrophic, p53 initiates an apoptotic cascade to eliminate the potential harm altogether135. Without the braking mechanisms provided by p53, we suspect that augmented cellular proliferation and differentiation driven by the viral latent transcription program results in enhanced DNA damage, accumulation of mutations, and, eventually, cellular transformation. Of note, we do find evidence of increased genomic instability in p53-/- mice after MHV68 infection (S.M. Owens and J.C. Forrest, manuscript in preparation), and the capacity to maintain in culture p53-deficient B cells expressing M2 and LMP1 (Supplementary Fig. 13b) suggests the potential for p53-dependent prevention of lymphomagenesis driven by certain GHV gene products.
While our work emphasizes the importance of p53 in limiting the GC expansion phase of MHV68 latency establishment, how the absence of p53 leads to a modest comparative reduction in latency over time, especially if p53 is activated by pro-latency viral genes, is not immediately apparent. It is generally thought that herpesviruses coevolved with their hosts yielding a virus-host relationship that allows the virus to chronically infect the host without causing major disease. This is perhaps reflected in our RNA-seq data, which indicate that p53 transcription is occurring simultaneously with M2-driven transcription related to B cell proliferation, survival, and differentiation. The early increase and late deficit in latently infected cell numbers suggest that p53 is a central player in this finely tuned balance that has evolved over millions of years. It is also interesting to note that M2, which is expressed in GC B cells, is an antigenic target for cytotoxic T lymphocytes (CTLs), as is LMP114,40. While we did not detect any difference in adaptive immune responses with the reagents at our disposal, we have not ruled out the possibility that there is increased CTL clearance over time of M2-expressing B cells in p53-/- mice. As alluded to above, it also is possible that the absence of p53 cytoprotective functions results in an increased mutational load that eventually leads to cellular dysfunction and death. Another possibility is that p53 influences terminal B cell differentiation, and we acknowledge that we have not yet defined the cell types harboring MHV68 during long-term infections of p53-/- mice. These questions, as well as determining whether p53 restricts MHV68-driven B cell lymphomagenesis, highlight some of the important concepts and remaining questions resulting from our studies that could be tested in future experiments.
Methods
Cell culture and viruses
Swiss albino 3T12 fibroblasts were purchased from ATCC. Murine embryonic fibroblasts (MEFs) were harvested from C57BL/6 mice embryos and immortalized as previously described72. All cells were cultured in DMEM supplemented with 10% fetal bovine serum, 2 mM L-glutamine, and 100 µ/ml penicillin/streptomycin. Cells were maintained at 37 °C in 5% CO2. Viruses used in this study include WT MHV68136, H2B-yellow fluorescent protein (YFP)-expressing MHV6858, Cre-recombinase-expressing MHV6864, LANA-beta-lactamase (73.Bla)-expressing MHV6859, and mLANA-null MHV68 (73.STOP)16. We generated an M2.Stop MHV68 on the 73.Bla backbone using en passant mutagenesis137. The MHV68 M2.Stop93 targeting construct was generated by the insertion of a 26-bp linker into the SacII site within the M2 ORF at bp 4314 (oligonucleotides Oligo1 [5“AAGCTTAGGCTAGTTAACTAGCCAGC] and Oligo2 [5“TGGCTAGTTAACTAGCCTAAGCTTGC]). The linker contained a diagnostic HindIII site. Oligo1 and Oligo2 were annealed and ligated into SacII-digested Lit38-M2. The addition of the oligomer resulted in a translational stop codon after residue 108 of the genomic M2 ORF. The γHV68M2.Stop construct was sequenced over the entire ORF with the Big Dye DNA sequencing kit (Applied Biosystems). A silent T-to-C mutation (that did not alter the predicted amino acid sequence but did result in the loss of a PstI site) at bp 4271 in the Lit38-M2.Stop construct was noted. Viral stocks were generated on 3T12 fibroblasts57. Viral titers were determined by MHV68 plaque assay138.
Mice and infections
All mice were housed and cared for according to the guidelines of the UAMS Department of Laboratory Animal Medicine and all state and federal requirements. Mice are housed in IVC cages in a room with a 12:12 light cycle and environmental conditions of 72°F and humidity of 30–70%. Both male and female mice are used for these experiments, and no overt differences in phenotypes due to sex are known to exist. p53-null mice on a C57BL/6 background (B6.129S2-Trp53tm1Tyj/J) were purchased from Jackson Laboratories and bred p53+/- x p53+/- to get a distribution of p53+/+, p53+/-, and p53-/- progeny. Ai14 (B6.Cg-Gt(ROSA)26Sortm14(CAG-tdTomato)Hze/J) and AID-Cre (B6.129P2-Aicdatm1(cre)Mnz/J) mice were purchased from Jackson laboratories and bred Ai14+/+ x AIDCre/Cre to produce hemizygous offspring (Ai14+/-AIDCre/+). IFNAR1-null mice on a C57BL/6 background (B6/129S2-Ifnar1tm1Agt/Mmjax) and Ai6 mice (B6.Cg-Gt(ROSA)26Sortm6(CAG-ZsGreen1)Hze/J) were purchased from Jackson Laboratories. 7–11-week-old mice were infected with 104 PFU of WT MHV68, H2B-YFP MHV68, MHV68-Cre, WT or M2.Stop 73.Bla MHV68, or mLANA-null MHV68 by intranasal inoculation. Mice were sacrificed according to normal endpoint protocols. Blood was collected on days 0, 16, and 42 post-infection via the submandibular vein.
Splenocyte isolation and limiting-dilution analyses
Limiting dilution analyses were performed with at least 2 independent infections with a minimum of 3 mice per group. Spleens were homogenized in a tenBroek tissue disrupter. Red blood cells were lysed by incubating tissue homogenate in 8.3 g/L ammonium chloride for 10 min at room temperature with shaking. Cells were filtered through a 40-micron mesh to reduce clumping. Frequencies of cells harboring MHV68 genomes were determined using a limiting-dilution, nested PCR analysis72. Frequencies of latently-infected cells capable of reactivating were determined using a limiting-dilution analysis for cytopathic effect induced on an indicator MEF monolayer72.
Antibodies, tetramers, and treatments
CD19-BV650 (6D5), IgM-BV421 (RMM-1), CD38-Pacific Blue (90), and purified B220 (RA3-6B2) were purchased from Biolegend (San Diego, CA). CD3e-PCPCy5.5 (145-2c11), CD8α-PCPCy5.5 (53-6.7), CD4-AF700 (RMA-5), B220-AF700 (RA3-6B2), GL7-eF660 (GL-7), CD38-PE-Cy7 (90), CD138-BV711 (281-2), NK1.1-PCPCy5.5 (PK136), and IgD-PE (11-26c) were purchased from eBioscience. Flow antibodies used at 1:300. Other antibodies include mouse anti-p53 Alexa Fluor 647-conjugate (Cell Signaling) used at 1:300, goat anti-GFP to enhance YFP detection (Rockland) used at 1:1000, anti-rat AF647-conjugate used at 1:400 and donkey anti-goat Alexa Fluor 488-conjugate (Jackson ImmunoResearch) used at 1:400. MHV68-specific MHC class I tetramers were generated by the NIH Tetramer Core. As a positive control for p53 induction, bulk splenocytes were treated with 10 grays of gamma radiation by exposure to a cesium-137 source. Following exposure, cells were allowed to recover at 37 °C in a tissue-culture incubator for 1 h prior to staining for flow cytometry.
Flow cytometry
Cells were washed with FACS buffer (0.2% BSA, 1 mM in PBS) before blocking with Fc block (Invitrogen) and incubation with eF780 live/dead viability stain (eBioscience) for 10 min at 4 °C. Surface staining was then performed with antibodies diluted at 1:300 for 30 min incubation time at 4 °C. For intracellular stains, cells were fixed and permeabilized using a FoxP3 staining kit (eBioscience) following the manufacturer’s guidelines. The data were collected using an LSRFortessa (Becton Dickinson) and analyzed using FlowJo (10.4.2) software.
Immunohistology and microscopy
Tissues were fixed in 4% formaldehyde before paraffin embedding and cut into 5 µm sections. Tissue sections were deparaffinized, boiled in citrate buffer to retrieve antigens, blocked in BSA, and incubated with primary antibody overnight. Secondary antibodies were applied for 1 h, and slides were mounted using Vectashield139. All antibodies were utilized at 1:400. To neutralize high autofluorescence in spleen sections, sections were treated with TrueVIEW™ Autofluorescence Quenching Kit (Vector Laboratories) before mounting. Images were captured using a Keyence BZ-9000E.
Real-time PCR and genotyping
Lungs were harvested from mice infected with H2B-YFP MHV68 on day 7 post-infection and homogenized using 0.5 mm silica beads in a Minibeadbeater (BioSpec)72. DNA was extracted from homogenized tissue using a DNeasy kit (Qiagen) according to the manufacturer’s instructions followed by treatment with RNAse I (Qiagen) for 30 minutes at 37 °C. Quantitative PCR was performed on 500 ng of DNA with primers forward corresponding to the MHV68 ORF59 genomic locus (forward: 5’-ATG-CAG-ACC-TTC-CAG-CTT-GAC-3’; reverse: 5’-CTC-TTC-CAA-GGG-AGC-TTG-CG-3’). Cycling parameters were 95 °C for 30 s, 55 °C for 30 s, and then 72 °C for 30 s for 40 cycles on an Applied Biosystems StepOnePlus PCR system. Genotyping of mice was performed using a mouse genotyping kit (Kapa Biosystems). Briefly, DNA was extracted from mouse tail snips and amplified as previously described113.
p53 pathway transcriptional array
Ai6 mice were infected with 104 PFU of MHV68-Cre intranasal inoculation and splenocytes were isolated at 16 days post-infection. One thousand ZsGreen positive and negative splenocytes were sorted in biological triplicate to a 96-well plate using a FACSAria II (BD). Transcriptome amplification was performed using the QIASeq Stranded RNA library kit (Qiagen). RT2 Profiler PCR Array Mouse p53 Signaling Pathway (PAMM-027Z – Qiagen) was used for quantification. Reactions were performed in an Applied Biosystems StepOnePlus PCR system with cycling conditions of 10 min at 95 °C followed by 40 cycles of 15 s at 95 °C and 1 min at 60 °C. Biological triplets were analyzed using GeneGlobe RT2 Profiler PCR Data Analysis software. Latent transcripts were analyzed by RT-PCR using inner primer sets as previously described140. Reactions were performed in an Applied Biosystems StepOnePlus PCR system with cycling conditions of 10 min at 95 °C followed by 40 cycles of 15 s at 95 °C and 1 min at 60 °C. Biological triplicate samples were analyzed in technical triplicate using the ΔΔCT method with β-actin as the cellular housekeeping transcript control. The results shown are technical triplicate for duplicate biological samples.
Plasmids
Retrovirus plasmids were generated by restriction enzyme cloning of latency genes into pRetroX-IRES-ZsGreen (Takara). Latency locus genes were amplified from WT MHV68-BAC DNA136 and the frame-shift stop mutant, M2.Stop, was amplified from M2.Stop MHV68-BAC DNA88 using primers in Table 1. LANA was cloned into the pRetroX vector via SacII and BamHI. vCyclin, M2, M11, and M2.Stop was amplified and cloned via NotI and BamHI. The LMP1 coding region was amplified from MSCV-N LMP1141 (Addgene plasmid #37962) and cloned into pRetroX-IRES-ZsGreen via Gibson Assembly. M2-Tyr was amplified from a gene block (GeneWiz) and cloned into pRetroX-IRES-ZsGreen via NotI and BamHI. LMP1AAAG was generated as previously described. A variant 3’ primer was used to mutate codon 384 from Tyr to Gly. LMP1G was cloned into pRetroX-IRES-ZsGreen via Gibson Assembly. Site-directed mutagenesis was used to mutate the codons 204-208 from PXGXT to AXAXA. All plasmids were confirmed by sequencing (Plasmidsaurus).
B cell cultures and retroviral transduction
Naïve B cells were isolated from spleens by immunomagnetic depletion using MojoSort Pan B cell Isolation Kit (Biolegend) and cultured in RPMI supplemented with L-glutamine, sodium pyruvate, penicillin-streptomycin, 50 μM 2-mercaptoethanol, and 10% fetal bovine serum. B cells were stimulated with 25 μg/ml LPS (Sigma). Retroviral supernatants were obtained from Phoenix cells (ATCC) 72 h after transient transfection (Lipofectamine LTX, ThermoFisher) with the retroviral plasmids and filtered through a 0.45 mm syringe filter. B cells were transduced 24 h after LPS stimulation with retroviral supernatants by centrifugation for 30 mins at 1000 × g at 37 °C in the presence of 5 μg/ml polybrene. Cells were washed, and supernatants were replaced with RPMI. Triplicate wells per condition were analyzed by flow cytometry on day 4 post-transduction.
Drug treatments
Cyclosporin A (500 μg/mL, TCI America), PP2 (10 µM, Selleck Chemical LLC), and PP3 (10 µM, Tocris Bioscience) were reconstituted in DMSO. For antibody treatments, mouse IgG Isotype Control (20 μg/mL, ThermoFisher) and anti-mouse IL-10R (20 μg/mL, BioXCell) were diluted in cRPMI. The drugs were used at a final concentration as indicated in parentheses above.
EdU incorporation assays
Mice were injected with 100 μg of 5-ethynyl-2’deoxyuridine (EdU, Invitrogen) 2 h prior to splenocyte harvest. Cells were fixed with 4% paraformaldehyde in PBS and Click-iT chemistry (BD Pharmagen) was performed according to the manufacturer’s instructions to fluorescently mark EdU + cells.
Caspase 3/7 cleavage assays
Apoptotic cells were labeled using fluorescently labeled inhibitors of caspases (FLICA 660, Immunochemistry Technologies, LLC). Briefly, splenocytes were stained with FLICA 660 for 1 h at 37 °C. Following incubation, cells were washed and surface stained for GC B cells markers as described above.
Enzyme-linked immunosorbent assays
Viral antigen was prepared by infecting 3T12 fibroblasts at an MOI of 0.5 PFU/cell for 96 h. Cells were washed and fixed in 1% PFA. Viral antigen was used to coat high-binding plates (Denville) overnight at 4 °C. After blocking in 5% FBS in PBS, serially diluted serum was incubated on the plate, followed by incubation with HRP-conjugated IgM or IgG Ab (Southern Biotech). SureBlue substrate (KPL) was added to detect the Ag-specific Abs. The wells were read on a FLUOstar Omega plate reader (BMG Labtech).
Ultra-low input RNAseq
Following 73.Bla MHV68 infection, MHV68 + cells are defined by cleavage of CCF4-AM fluorescent substrate59. Briefly, draining lymph nodes from infected animals were collected and homogenized as described above. β-lactamase activity was detected using the LiveBLAzer FRET-BG/Loading Kit with CCF4-AM (Thermo Fischer Scientific #K1095). Cells were incubated for 1 h at room temperature, protected from light, washed 3x with FACS buffer, and immediately subjected to fluorescent-activated cell sorting (FACS) performed on a FACS Aria flow cytometer (BD Biosciences). Sorted cells were frozen in Trizol. The library was constructed by GeneWiz (Azenta) using Illumina RNA prep with enrichment for full-length transcripts. Polyadenylated transcripts were reverse transcribed using SMART-Seq® v4 Ultra® Low Input RNA Kit (Clontech), followed by second-strand synthesis of full-length cDNA and amplification of the product. Illumina®-compatible sequencing libraries were constructed and then validated for sufficient concentration and appropriate fragment sizes. Samples were sequenced on the Illumina HiSeq® 2500 with a 1 × 100 bp single-end configuration. Data processing and analysis are described in RNAseq methods.
RNAseq
Transduced B cells were collected 4 days post-transduction. RNA was harvested using Trizol. The library was constructed by BGI Laboratories using DNBSEQ Eukaryotic transcriptome protocol. Following mRNA isolation, the mRNA was fragmented, and first-strand synthesis performed. Double-stranded cDNA fragments were subjected to end-repair, and a single ‘A’ nucleotide was added to the 3’ end of the blunt fragment. Following library QC, single-strand cDNAs were circularized and replicated via rolling cycle amplification. DNA nanoballs were generated and loaded into patterned nanoarrays using high-intensity DNA nanochip technique and sequenced through combinatorial Probe-Anchor Synthesis (cPAS). Sequencing was performed on a DNBSEQ-G400 (BGI Genomics Co). Raw sequence reads were quality-checked using fastQC and trimmed based on quality (Phred > 30). After trimming, reads < 30 bp in size were discarded. Trimmed sequence reads were mapped to reference genome mm10 using STAR aligner with default parameters142. The gene count tables were extracted from alignment results using bedtool2 software143. Read counts were normalized using the Voom method144. Normalized read counts were analyzed using DEseq2145. Representative gene set enrichment analysis (GSEA)146,147 was performed using the reference list derived from Hallmark gene sets148 and compared with a pre-ranked list (by fold) of global average gene expression.
Statistical analysis
All data was analyzed using GraphPad Prism software (GraphPad Software, http://www.graphpad.com, La Jolla, CA). MFI was analyzed by one-way ANOVA. The total number of immune cells subset per animal were assessed using a Mann-Whitney unpaired two-sided T-test. Based on the Poisson distribution, the frequencies of the viral genome–positive cells and reactivation were obtained from the nonlinear regression fit of the data where the regression line intersected 63.2%. Extrapolations were used for samples that did not intersect 63.2%. Non-linear regressions were assessed by the Extra sum-of-squares F test of the LogEC50. PCR and reactivation assays were performed with at least 2 independent infections with a minimum of 3 mice per group. Edu+ and active caspase cell frequencies were analyzed using two-tailed Student’s t tests. All results are insignificant unless otherwise noted. Dots represent individual animals.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Ex vivo B cell transduction mRNAseq data generated in this study are in the NCBI database under GEO Accession # GSE225579. (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE225579). In vivo lymphocyte mRNAseq data generated in this study are in the NCBI database under GEO Accession # GSE285085 Source data are provided in this paper.
References
Speck, S. H. & Ganem, D. Viral latency and its regulation: lessons from the gamma-herpesviruses. Cell Host Microbe 8, 100–115 (2010).
Bornkamm, G. W. Epstein-Barr virus and its role in the pathogenesis of Burkitt’s lymphoma: an unresolved issue. Semin. Cancer Biol. 19, 351–365 (2009).
Damania, B., Kenney, S. C. & Raab-Traub, N. Epstein-barr virus: Biology and clinical disease. Cell 185, 3652–3670 (2022).
Cesarman, E., Chadburn, A. & Rubinstein, P. G. KSHV/HHV8-mediated hematologic diseases. Blood 139, 1013–1025 (2022).
Yarchoan, R. & Uldrick, T. S. HIV-Associated cancers and related diseases. N. Engl. J. Med. 378, 1029–1041 (2018).
Johnson, K. E. & Tarakanova, V. L. Gammaherpesviruses and B Cells: A relationship that lasts a lifetime. Viral Immunol. 33, 316–326 (2020).
Liang, X., Collins, C. M., Mendel, J. B., Iwakoshi, N. N. & Speck, S. H. Gammaherpesvirus-driven plasma cell differentiation regulates virus reactivation from latently infected B lymphocytes. PLoS Pathog. 5, e1000677 (2009).
Roughan, J. E. & Thorley-Lawson, D. A. The intersection of Epstein-Barr virus with the germinal center. J. Virol. 83, 3968–3976 (2009).
MacLennan, I. C. Germinal centers. Annu. Rev. Immunol. 12, 117–139 (1994).
Victora, G. D. & Nussenzweig, M. C. Germinal centers. Annu. Rev. Immunol. 40, 413–442 (2022).
Suan, D., Sundling, C. & Brink, R. Plasma cell and memory B cell differentiation from the germinal center. Curr. Opin. Immunol. 45, 97–102 (2017).
Thorley-Lawson, D. A. Epstein-barr virus: exploiting the immune system. Nat. Rev. Immunol. 1, 75–82 (2001).
Barton, E., Mandal, P. & Speck, S. H. Pathogenesis and host control of gammaherpesviruses: lessons from the mouse. Annu. Rev. Immunol. 29, 351–397 (2011).
Wang, Y., Tibbetts, S. A. & Krug, L. T. Conquering the host: Determinants of pathogenesis learned from murine gammaherpesvirus 68. Annu. Rev. Virol. 8, 349–371 (2021).
Fowler, P., Marques, S., Simas, J. P. & Efstathiou, S. ORF73 of murine herpesvirus-68 is critical for the establishment and maintenance of latency. J. Gen. Virol. 84, 3405–3416 (2003).
Moorman, N. J., Willer, D. O. & Speck, S. H. The gammaherpesvirus 68 latency-associated nuclear antigen homolog is critical for the establishment of splenic latency. J. Virol. 77, 10295–10303 (2003).
Salinas, E. et al. Conditional mutagenesis in vivo reveals cell type- and infection stage-specific requirements for LANA in chronic MHV68 infection. PLoS Pathog. 14, e1006865 (2018).
Rodrigues, L., Popov, N., Kaye, K. M. & Simas, J. P. Stabilization of Myc through heterotypic poly-ubiquitination by mLANA is critical for γ-herpesvirus lymphoproliferation. PLoS Pathog. 9, e1003554 (2013).
van Dyk, L. F. et al. The murine gammaherpesvirus 68 v-cyclin gene is an oncogene that promotes cell cycle progression in primary lymphocytes. J. Virol. 73, 5110–5122 (1999).
Hoge, A. T., Hendrickson, S. B. & Burns, W. H. Murine gammaherpesvirus 68 cyclin D homologue is required for efficient reactivation from latency. J. Virol. 74, 7016–7023 (2000).
van Dyk, L. F., Virgin, H. W. T. & Speck, S. H. The murine gammaherpesvirus 68 v-cyclin is a critical regulator of reactivation from latency. J. Virol. 74, 7451–7461 (2000).
de Lima, B. D., May, J. S., Marques, S., Simas, J. P. & Stevenson, P. G. Murine gammaherpesvirus 68 bcl-2 homologue contributes to latency establishment in vivo. J. Gen. Virol. 86, 31–40 (2005).
Wang, G. H., Garvey, T. L. & Cohen, J. I. The murine gammaherpesvirus-68 M11 protein inhibits Fas- and TNF-induced apoptosis. J. Gen. Virol. 80, 2737–2740 (1999).
Coleman, C. B. et al. A gammaherpesvirus Bcl-2 ortholog blocks B cell receptor-mediated apoptosis and promotes the survival of developing B cells in vivo. PLoS Pathog. 10, e1003916 (2014).
Rangaswamy, U. S., O’Flaherty, B. M. & Speck, S. H. Tyrosine 129 of the murine gammaherpesvirus M2 protein is critical for M2 function in vivo. PLoS ONE 9, e105197 (2014).
Rangaswamy, U. S. & Speck, S. H. Murine gammaherpesvirus M2 protein induction of IRF4 via the NFAT pathway leads to IL-10 expression in B cells. PLoS Pathog. 10, e1003858 (2014).
Pires de Miranda, M. et al. The Gammaherpesvirus m2 protein manipulates the Fyn/Vav pathway through a multidocking mechanism of assembly. PLoS ONE 3, e1654 (2008).
Pires de Miranda, M., Lopes, F. B., McVey, C. E., Bustelo, X. R. & Simas, J. P. Role of Src homology domain binding in signaling complexes assembled by the murid γ-herpesvirus M2 protein. J. Biol. Chem. 288, 3858–3870 (2013).
Owens, S. M., Oldenburg, D. G., White, D. W. & Forrest, J. C. Deletion of murine gammaherpesvirus gene M2 in activation-induced cytidine deaminase-expressing B cells impairs host colonization and viral reactivation. J. Virol. 95, https://doi.org/10.1128/jvi.01933-20 (2020).
Terrell, S. & Speck, S. H. Murine gammaherpesvirus M2 antigen modulates splenic B cell activation and terminal differentiation in vivo. PLoS Pathog 13, e1006543 (2017).
Mosialos, G. et al. The Epstein-Barr virus transforming protein LMP1 engages signaling proteins for the tumor necrosis factor receptor family. Cell 80, 389–399 (1995).
Kilger, E., Kieser, A., Baumann, M. & Hammerschmidt, W. Epstein-Barr virus-mediated B-cell proliferation is dependent upon latent membrane protein 1, which simulates an activated CD40 receptor. EMBO J. 17, 1700–1709 (1998).
Uchida, J. et al. Mimicry of CD40 signals by Epstein-Barr virus LMP1 in B lymphocyte responses. Science 286, 300–303 (1999).
Verschuren, E. W., Klefstrom, J., Evan, G. I. & Jones, N. The oncogenic potential of Kaposi’s sarcoma-associated herpesvirus cyclin is exposed by p53 loss in vitro and in vivo. Cancer Cell 2, 229–241 (2002).
Curran, J. A., Laverty, F. S., Campbell, D., Macdiarmid, J. & Wilson, J. B. Epstein-Barr virus encoded latent membrane protein-1 induces epithelial cell proliferation and sensitizes transgenic mice to chemical carcinogenesis. Cancer Res. 61, 6730–6738 (2001).
Kulwichit, W. et al. Expression of the Epstein-Barr virus latent membrane protein 1 induces B cell lymphoma in transgenic mice. Proc. Natl. Acad. Sci. USA 95, 11963–11968 (1998).
Nikitin, P. A. et al. An ATM/Chk2-mediated DNA damage-responsive signaling pathway suppresses Epstein-Barr virus transformation of primary human B cells. Cell Host Microbe 8, 510–522 (2010).
Sugden, B. & Mark, W. Clonal transformation of adult human leukocytes by Epstein-Barr virus. J. Virol. 23, 503–508 (1977).
Wang, D., Liebowitz, D. & Kieff, E. An EBV membrane protein expressed in immortalized lymphocytes transforms established rodent cells. Cell 43, 831–840 (1985).
Zhang, B. et al. Immune surveillance and therapy of lymphomas driven by Epstein-Barr virus protein LMP1 in a mouse model. Cell 148, 739–751 (2012).
Koopal, S. et al. Viral oncogene-induced DNA damage response is activated in Kaposi sarcoma tumorigenesis. PLoS Pathog. 3, 1348–1360 (2007).
Liang, X. et al. Murine gamma-herpesvirus immortalization of fetal liver-derived B cells requires both the viral cyclin D homolog and latency-associated nuclear antigen. PLoS Pathog. 7, e1002220 (2011).
Kulinski, J. M. et al. Ataxia telangiectasia mutated kinase controls chronic gammaherpesvirus infection. J. Virol. 86, 12826–12837 (2012).
Darrah, E. J. et al. B Cell-specific expression of Ataxia-Telangiectasia mutated protein kinase promotes chronic gammaherpesvirus infection. J. Virol. 91, https://doi.org/10.1128/jvi.01103-17 (2017).
Darrah, E. J., Stoltz, K. P., Ledwith, M. & Tarakanova, V. L. ATM supports gammaherpesvirus replication by attenuating type I interferon pathway. Virology 510, 137–146 (2017).
Muller, P. A. & Vousden, K. H. Mutant p53 in cancer: new functions and therapeutic opportunities. Cancer Cell 25, 304–317 (2014).
Lavin, M. F. & Gueven, N. The complexity of p53 stabilization and activation. Cell Death Differ. 13, 941–950 (2006).
Lozano, G. The enigma of p53. Cold Spring Harb. Symp. Quant. Biol. 81, 37–40 (2016).
Allday, M. J., Sinclair, A., Parker, G., Crawford, D. H. & Farrell, P. J. Epstein-Barr virus efficiently immortalizes human B cells without neutralizing the function of p53. EMBO J. 14, 1382–1391 (1995).
Chang, P. C. & Li, M. Kaposi’s sarcoma-associated herpesvirus K-cyclin interacts with Cdk9 and stimulates Cdk9-mediated phosphorylation of p53 tumor suppressor. J. Virol. 82, 278–290 (2008).
Forte, E. & Luftig, M. A. MDM2-dependent inhibition of p53 is required for Epstein-Barr virus B-cell growth transformation and infected-cell survival. J. Virol. 83, 2491–2499 (2009).
Friborg, J. Jr., Kong, W., Hottiger, M. O. & Nabel, G. J. p53 inhibition by the LANA protein of KSHV protects against cell death. Nature 402, 889–894 (1999).
Saha, A. et al. Epstein-Barr virus nuclear antigen 3C augments Mdm2-mediated p53 ubiquitination and degradation by deubiquitinating Mdm2. J. Virol. 83, 4652–4669 (2009).
Frappier, L. Contributions of Epstein-Barr nuclear antigen 1 (EBNA1) to cell immortalization and survival. Viruses 4, 1537–1547 (2012).
Petre, C. E., Sin, S. H. & Dittmer, D. P. Functional p53 signaling in Kaposi’s sarcoma-associated herpesvirus lymphomas: implications for therapy. J. Virol. 81, 1912–1922 (2007).
Serrano, M., Lin, A. W., McCurrach, M. E., Beach, D. & Lowe, S. W. Oncogenic <em>ras</em> provokes premature cell senescence associated with accumulation of p53 and p16INK4a. Cell 88, 593–602 (1997).
Forrest, J. C. et al. ORF73-null murine gammaherpesvirus 68 reveals roles for mLANA and p53 in virus replication. J. Virol. 81, 11957–11971 (2007).
Collins, C. M. & Speck, S. H. Tracking murine gammaherpesvirus 68 infection of germinal center B cells in vivo. PLoS ONE 7, e33230 (2012).
Nealy, M. S., Coleman, C. B., Li, H. & Tibbetts, S. A. Use of a virus-encoded enzymatic marker reveals that a stable fraction of memory B cells expresses latency-associated nuclear antigen throughout chronic gammaherpesvirus infection. J. Virol. 84, 7523–7534 (2010).
Willer, D. O. & Speck, S. H. Long-term latent murine Gammaherpesvirus 68 infection is preferentially found within the surface immunoglobulin D-negative subset of splenic B cells in vivo. J. Virol. 77, 8310–8321 (2003).
Rivas, C., Aaronson, S. A. & Munoz-Fontela, C. Dual role of p53 in innate antiviral immunity. Viruses 2, 298–313 (2010).
Madisen, L. et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 13, 133–140 (2010).
Siegel, A. M., Rangaswamy, U. S., Napier, R. J. & Speck, S. H. Blimp-1-dependent plasma cell differentiation is required for efficient maintenance of murine gammaherpesvirus latency and antiviral antibody responses. J. Virol. 84, 674–685 (2010).
Moser, J. M. et al. Role of B-cell proliferation in the establishment of gammaherpesvirus latency. J. Virol. 79, 9480–9491 (2005).
Weck, K. E., Kim, S. S., Virgin, H. W. I. V. & Speck, S. H. B cells regulate murine gammaherpesvirus 68 latency. J. Virol. 73, 4651–4661 (1999).
Krug, L. T., Moser, J. M., Dickerson, S. M. & Speck, S. H. Inhibition of NF-kappaB activation in vivo impairs establishment of gammaherpesvirus latency. PLoS Pathog. 3, e11 (2007).
Sarawar, S. R., Brooks, J. W., Cardin, R. D., Mehrpooya, M. & Doherty, P. C. Pathogenesis of murine gammaherpesvirus-68 infection in interleukin-6-deficient mice. Virology 249, 359–366 (1998).
Miciak, J. & Bunz, F. Long story short: p53 mediates innate immunity. Biochim. Biophys. Acta 1865, 220–227 (2016).
Levine, A. J. P53 and The immune response: 40 Years of exploration-a plan for the future. Int. J. Mol. Sci. 21, https://doi.org/10.3390/ijms21020541 (2020).
Menendez, D., Shatz, M. & Resnick, M. A. Interactions between the tumor suppressor p53 and immune responses. Curr. Opin. Oncol. 25, 85–92 (2013).
Tibbetts, S. A. et al. Establishment and maintenance of gammaherpesvirus latency are independent of infective dose and route of infection. J. Virol. 77, 7696–7701 (2003).
Weck, K. E., Barkon, M. L., Yoo, L. I., Speck, S. H. & Virgin, H. I. Mature B cells are required for acute splenic infection, but not for establishment of latency, by murine gammaherpesvirus 68. J. Virol. 70, 6775–6780 (1996).
Weck, K. E., Kim, S. S., Virgin, H. I. & Speck, S. H. Macrophages are the major reservoir of latent murine gammaherpesvirus 68 in peritoneal cells. J. Virol. 73, 3273–3283 (1999).
Bussey, K. A. et al. Endosomal toll-like receptors 7 and 9 cooperate in detection of murine gammaherpesvirus 68 infection. J. Virol. 93, https://doi.org/10.1128/jvi.01173-18 (2019).
Mandal, P. et al. A gammaherpesvirus cooperates with interferon-alpha/beta-induced IRF2 to halt viral replication, control reactivation, and minimize host lethality. PLoS Pathog. 7, e1002371 (2011).
Weck, K. E. et al. Murine gamma-herpesvirus 68 causes severe large-vessel arteritis in mice lacking interferon-gamma responsiveness: a new model for virus-induced vascular disease. Nat. Med. 3, 1346–1353 (1997).
Barton, E. S., Lutzke, M. L., Rochford, R. & Virgin, H. W. Alpha/beta interferons regulate murine gammaherpesvirus latent gene expression and reactivation from latency. J. Virol. 79, 14149–14160 (2005).
Steed, A. L. et al. Gamma interferon blocks gammaherpesvirus reactivation from latency. J. Virol. 80, 192–200 (2006).
Tripp, R. A. et al. Pathogenesis of an infectious mononucleosis-like disease induced by a murine gamma-herpesvirus: role for a viral superantigen? J. Exp. Med. 185, 1641–1650 (1997).
de Stanchina, E. et al. E1A signaling to p53 involves the p19(ARF) tumor suppressor. Genes Dev. 12, 2434–2442 (1998).
Lowe, S. W. & Ruley, H. E. Stabilization of the p53 tumor suppressor is induced by adenovirus 5 E1A and accompanies apoptosis. Genes Dev. 7, 535–545 (1993).
Hoffman, B. & Liebermann, D. A. Apoptotic signaling by c-MYC. Oncogene 27, 6462–6472 (2008).
Flaño, E., Kim, I. J., Moore, J., Woodland, D. L. & Blackman, M. A. Differential gamma-herpesvirus distribution in distinct anatomical locations and cell subsets during persistent infection in mice. J. Immunol. 170, 3828–3834 (2003).
Husain, S. M. et al. Murine gammaherpesvirus M2 gene is latency-associated and its protein a target for CD8(+) T lymphocytes. Proc. Natl. Acad. Sci. USA 96, 7508–7513 (1999).
Allen, R. D., Dickerson, S. & Speck, S. H. Identification of spliced gammaherpesvirus 68 LANA and v-cyclin transcripts and analysis of their expression in vivo during latent infection. J. Virol. 80, 2055–2062 (2006).
Siegel, A. M., Herskowitz, J. H. & Speck, S. H. The MHV68 M2 protein drives IL-10 dependent B cell proliferation and differentiation. PLoS Pathog. 4, e1000039 (2008).
Rodrigues, L., Pires de Miranda, M., Caloca, M. J., Bustelo, X. R. & Simas, J. P. Activation of Vav by the gammaherpesvirus M2 protein contributes to the establishment of viral latency in B lymphocytes. J. Virol. 80, 6123–6135 (2006).
Owens, S. M., Oldenburg, D. G., White, D. W. & Forrest, J. C. Deletion of murine gammaherpesvirus gene. J. Virol. 95, https://doi.org/10.1128/jvi.01933-20 (2020).
Tornesello, M. L., Annunziata, C., Tornesello, A. L., Buonaguro, L. & Buonaguro, F. M. Human oncoviruses and p53 tumor suppressor pathway deregulation at the origin of human cancers. Cancers 10, https://doi.org/10.3390/cancers10070213 (2018).
Liang, X., Shin, Y. C., Means, R. E. & Jung, J. U. Inhibition of interferon-mediated antiviral activity by murine gammaherpesvirus 68 latency-associated M2 protein. J. Virol. 78, 12416–12427 (2004).
Keim, C., Kazadi, D., Rothschild, G. & Basu, U. Regulation of AID, the B-cell genome mutator. Genes Dev. 27, 1–17 (2013).
Niemeyer, B. F. et al. The gammaherpesvirus 68 viral cyclin facilitates expression of LANA. PLoS Pathog. 17, e1010019 (2021).
Jacoby, M. A., Virgin, H. W. T. & Speck, S. H. Disruption of the M2 gene of murine gammaherpesvirus 68 alters splenic latency following intranasal, but not intraperitoneal, inoculation. J. Virol. 76, 1790–1801 (2002).
Macrae, A. I. et al. Murid herpesvirus 4 strain 68 M2 protein is a B-cell-associated antigen important for latency but not lymphocytosis. J. Virol. 77, 9700–9709 (2003).
Simas, J. P., Marques, S., Bridgeman, A., Efstathiou, S. & Adler, H. The M2 gene product of murine gammaherpesvirus 68 is required for efficient colonization of splenic follicles but is not necessary for expansion of latently infected germinal centre B cells. J. Gen. Virol. 85, 2789–2797 (2004).
Rastelli, J. et al. LMP1 signaling can replace CD40 signaling in B cells in vivo and has unique features of inducing class-switch recombination to IgG1. Blood 111, 1448–1455 (2008).
Kempkes, B. & Robertson, E. S. Epstein-Barr virus latency: current and future perspectives. Curr. Opin. Virol. 14, 138–144 (2015).
Young, L. S. & Murray, P. G. Epstein-Barr virus and oncogenesis: from latent genes to tumours. Oncogene 22, 5108–5121 (2003).
Thornburg, N. J. et al. LMP1 signaling and activation of NF-kappaB in LMP1 transgenic mice. Oncogene 25, 288–297 (2006).
Li, L. et al. The activation of p53 mediated by Epstein-Barr virus latent membrane protein 1 in SV40 large T-antigen transformed cells. FEBS Lett. 582, 755–762 (2008).
Brennan, P., Floettmann, J. E., Mehl, A., Jones, M. & Rowe, M. Mechanism of action of a novel latent membrane protein-1 dominant negative. J. Biol. Chem. 276, 1195–1203 (2001).
Phan, R. T. & Dalla-Favera, R. The BCL6 proto-oncogene suppresses p53 expression in germinal-centre B cells. Nature 432, 635–639 (2004).
Aloni-Grinstein, R., Charni-Natan, M., Solomon, H. & Rotter, V. p53 and the Viral Connection: Back into the Future. Cancers 10, https://doi.org/10.3390/cancers10060178 (2018).
Sifford, J. M., Stahl, J. A., Salinas, E. & Forrest, J. C. Murine gammaherpesvirus 68 LANA and SOX homologs counteract ATM-Driven p53 activity during lytic viral replication. J. Virol. 90, 2571–2585 (2015).
Casavant, N. C. et al. Potential role for p53 in the permissive life cycle of human cytomegalovirus. J. Virol. 80, 8390–8401 (2006).
Lane, D. P. & Crawford, L. V. T antigen is bound to a host protein in SV40-transformed cells. Nature 278, 261–263 (1979).
Sarnow, P., Ho, Y. S., Williams, J. & Levine, A. J. Adenovirus E1b-58kd tumor antigen and SV40 large tumor antigen are physically associated with the same 54 kd cellular protein in transformed cells. Cell 28, 387–394 (1982).
Maruzuru, Y. et al. p53 Is a host cell regulator during Herpes Simplex Encephalitis. J. Virol. 90, 6738–6745 (2016).
Saridakis, V. et al. Structure of the p53 binding domain of HAUSP/USP7 bound to Epstein-Barr nuclear antigen 1 implications for EBV-mediated immortalization. Mol. Cell 18, 25–36 (2005).
Cai, Q. L., Knight, J. S., Verma, S. C., Zald, P. & Robertson, E. S. EC5S ubiquitin complex is recruited by KSHV latent antigen LANA for degradation of the VHL and p53 tumor suppressors. PLoS Pathog. 2, e116 (2006).
Juillard, F. et al. KSHV LANA acetylation-selective acidic domain reader sequence mediates virus persistence. Proc. Natl. Acad. Sci. USA 117, 22443–22451 (2020).
Sarek, G. et al. Reactivation of the p53 pathway as a treatment modality for KSHV-induced lymphomas. J. Clin. Invest. 117, 1019–1028 (2007).
Jacks, T. et al. Tumor spectrum analysis in p53-mutant mice. Curr. Biol. 4, 1–7 (1994).
Valdez, J. M., Nichols, K. E. & Kesserwan, C. Li-Fraumeni syndrome: a paradigm for the understanding of hereditary cancer predisposition. Br J Haematol 176, 539–552 (2017).
Mukhopadhyay, U. K. et al. Doubles game: Src-Stat3 versus p53-PTEN in cellular migration and invasion. Mol. Cell Biol. 30, 4980–4995 (2010).
Thomas, A. F., Kelly, G. L. & Strasser, A. Of the many cellular responses activated by TP53, which ones are critical for tumour suppression? Cell Death Differ. 29, 961–971 (2022).
Hau, P. M. & Tsao, S. W. Epstein-Barr virus Hijacks DNA damage response transducers to orchestrate its life cycle. Viruses 9, (2017). https://doi.org/10.3390/v9110341
Tatfi, M., Hermine, O. & Suarez, F. Epstein-Barr Virus (EBV)-Related Lymphoproliferative Disorders in Ataxia Telangiectasia: Does ATM Regulate EBV Life Cycle? Front Immunol 9, 3060 (2018).
Hafez, A. Y. & Luftig, M. A. Characterization of the EBV-Induced Persistent DNA Damage Response. Viruses 9, https://doi.org/10.3390/v9120366 (2017).
Koganti, S. et al. STAT3 interrupts ATR-Chk1 signaling to allow oncovirus-mediated cell proliferation. Proc. Natl. Acad. Sci. USA 111, 4946–4951 (2014).
Blackford, A. N. & Jackson, S. P. ATM, ATR, and DNA-PK: The Trinity at the Heart of the DNA Damage Response. Mol Cell 66, 801–817 (2017).
Canny, S. P. et al. Latent gammaherpesvirus 68 infection induces distinct transcriptional changes in different organs. J. Virol. 88, 730–738 (2014).
Ruzankina, Y. et al. Deletion of the developmentally essential gene ATR in adult mice leads to age-related phenotypes and stem cell loss. Cell Stem Cell 1, 113–126 (2007).
Lee, H. et al. Identification of an immunoreceptor tyrosine-based activation motif of K1 transforming protein of Kaposi’s sarcoma-associated herpesvirus. Mol. Cell Biol. 18, 5219–5228 (1998).
Lagunoff, M., Majeti, R., Weiss, A. & Ganem, D. Deregulated signal transduction by the K1 gene product of Kaposi’s sarcoma-associated herpesvirus. Proc. Natl. Acad. Sci. USA 96, 5704–5709 (1999).
Fruehling, S. & Longnecker, R. The immunoreceptor tyrosine-based activation motif of Epstein-Barr virus LMP2A is essential for blocking BCR-mediated signal transduction. Virology 235, 241–251 (1997).
Steinbrück, L. et al. K1 and K15 of Kaposi’s sarcoma-associated herpesvirus are partial functional homologues of latent membrane protein 2A of Epstein-Barr virus. J. Virol. 89, 7248–7261 (2015).
Minamitani, T. et al. Mouse model of Epstein-Barr virus LMP1- and LMP2A-driven germinal center B-cell lymphoproliferative disease. Proc. Natl. Acad. Sci. USA 114, 4751–4756 (2017).
Wang, L. W., Jiang, S. & Gewurz, B. E. Epstein-Barr Virus LMP1-mediated oncogenicity. J. Virol. 91, https://doi.org/10.1128/JVI.01718-16 (2017).
Guo, L. et al. Epstein-Barr virus oncoprotein LMP1 mediates survivin upregulation by p53 contributing to G1/S cell cycle progression in nasopharyngeal carcinoma. Int. J. Mol. Med. 29, 574–580 (2012).
Li, L. et al. Viral oncoprotein LMP1 disrupts p53-induced cell cycle arrest and apoptosis through modulating K63-linked ubiquitination of p53. Cell Cycle 11, 2327–2336 (2012).
Shao, J. Y. et al. Epstein-Barr virus LMP1 status in relation to apoptosis, p53 expression and leucocyte infiltration in nasopharyngeal carcinoma. Anticancer Res. 24, 2309–2318 (2004).
Vrazo, A. C., Chauchard, M., Raab-Traub, N. & Longnecker, R. Epstein-Barr virus LMP2A reduces hyperactivation induced by LMP1 to restore normal B cell phenotype in transgenic mice. PLoS Pathog. 8, e1002662 (2012).
Ma, S. D. et al. Latent membrane protein 1 (LMP1) and LMP2A collaborate to promote Epstein-Barr virus-induced B cell lymphomas in a cord blood-humanized mouse model but are not essential. J. Virol. 91, https://doi.org/10.1128/JVI.01928-16 (2017).
Hernández Borrero, L. J. & El-Deiry, W. S. Tumor suppressor p53: Biology, signaling pathways, and therapeutic targeting. Biochim. Biophys. Acta Rev. Cancer 1876, 188556 (2021).
Adler, H., Messerle, M., Wagner, M. & Koszinowski, U. H. Cloning and mutagenesis of the murine gammaherpesvirus 68 genome as an infectious bacterial artificial chromosome. J. Virol. 74, 6964–6974 (2000).
Tischer, B. K., Smith, G. A. & Osterrieder, N. En passant mutagenesis: a two step markerless red recombination system. Methods Mol. Biol. 634, 421–430 (2010).
Stahl, J. A. et al. Amplification of JNK signaling is necessary to complete the murine gammaherpesvirus 68 lytic replication cycle. J. Virol. 86, 13253–13262 (2012).
Zaqout, S., Becker, L. L. & Kaindl, A. M. Immunofluorescence staining of paraffinsections step by step. Front. Neuroanat. 14, 582218 (2020).
Virgin, H. W., Presti, R. M., Li, X. Y., Liu, C. & Speck, S. H. Three distinct regions of the murine gammaherpesvirus 68 genome are transcriptionally active in latently infected mice. J. Virol. 73, 2321–2332 (1999).
Rozenblatt-Rosen, O. et al. Interpreting cancer genomes using systematic host network perturbations by tumour virus proteins. Nature 487, 491–495 (2012).
Dobin, A. et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21 (2013).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Law, C. W., Chen, Y., Shi, W. & Smyth, G. K. voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 15, R29 (2014).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 102, 15545–15550 (2005).
Mootha, V. K. et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 34, 267–273 (2003).
Howe, D. G. et al. Model organism data evolving in support of translational medicine. Lab Anim. 47, 277–289 (2018).
Acknowledgements
We thank Andrea Harris for the UAMS Flow Cytometry Core. This work was supported in part by grant R01CA167065 from the NIH National Cancer Institute and start-up funds from the UAMS College of Medicine and Arkansas Biosciences Institute to J.C.F. S.J.M. was supported through a diversity supplement to R01CA167065 and R25GM083247 from the National Institute of General Medical Sciences (NIGMS) of the National Institutes of Health. The Flow Cytometry and Microscopy Core and the work described here were also supported in part by the Center for Microbial Pathogenesis and Host Inflammatory Responses award P20GM103625 from the NIH National Institute of General Medical Sciences Centers of Biomedical Research Excellence. S.M.O. was supported by Translational Research Institute (TRI) grant TL1TR003109 through the NIH National Center for Advancing Translational Sciences. M.M. is supported by K22CA241355 from the National Cancer Institute with additional support from P30GM145393 from the National Institute of General Medical Sciences. The funders had no role in study design, data collection, and interpretation, or the decision to submit the work for publication. The model in Fig. 8 was created in BioRender. Lab, F. (2024) https://BioRender.com/b70y129.
Author information
Authors and Affiliations
Contributions
S.M.O., J.M.S., and J.C.F designed the study, analyzed data, and wrote the manuscript. G.L. constructed and tested retroviral vectors. S.J.M. performed IFNAR1 experiments, analyzed data, and wrote the manuscript. E.S. participated in the study design. D.G.O. generated MHV68 BAC recombinants. I.N. performed RNA-seq raw data analysis. M.M. performed RNA-seq pathway analyses and wrote the manuscript. D.G. and J.S. assisted with flow cytometry data analysis. J.C.F. acquired funding. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Owens, S.M., Sifford, J.M., Li, G. et al. Intrinsic p53 activation restricts gammaherpesvirus driven germinal center B cell expansion during latency establishment. Nat Commun 16, 951 (2025). https://doi.org/10.1038/s41467-025-56247-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-56247-5
