
Abstract
Effective countermeasures against the adverse cardiovascular effects of circadian misalignment, such as effects experienced due to night work or jet lag, remain to be established in humans. Here, we aim to test whether eating only during daytime can mitigate such adverse effects vs. eating during the night and day (typical for night shift workers) under simulated night work (secondary analysis of NCT02291952). This single-blind, parallel-arm trial randomized 20 healthy participants (non-shift workers) to simulated night work with meals consumed during night and day (Nighttime Meal Control Group) or only during daytime (Daytime Meal Intervention Group). The primary outcomes were pNN50 (percentage consecutive heartbeat intervals >50 ms), RMSSD (root mean square of successive heartbeat differences), and LF/HF (low/high cardiac frequency). The secondary outcome was blood concentrations of prothrombotic factor plasminogen activator inhibitor-1 (PAI-1). These measures were assessed under Constant Routine conditions, before (baseline) and after (postmisalignment) simulated night work. The meal timing intervention significantly modified the impact of simulated night work on cardiac vagal modulation and PAI-1 (pFDR = 0.001). In the Control Group, the postmisalignment Constant Routine showed a decrease in pNN50 by 25.7% (pFDR = 0.008) and RMMSD by 14.3% (pFDR = 0.02), and an increase in LF/HF by 5.5% (pFDR = 0.04) and PAI-1 by 23.9% (pFDR = 0.04), vs. the baseline Constant Routine. In the Intervention Group, there were no significant changes in these outcomes. For exploratory outcomes, the intervention significantly modified the impact of simulated night work on blood pressure (P < 0.05), with no significant change in the Control Group, and a significant reduction by 6-8% (P < 0.01) in the Intervention Group; without significant effects for heart rate or cortisol. These findings indicate that daytime eating, despite mistimed sleep, may mitigate changes in cardiovascular risk factors and offer translational evidence for developing a behavioral strategy to help minimize the adverse changes in cardiovascular risk factors in individuals exposed to circadian misalignment, such as shift workers.
Similar content being viewed by others

Introduction
Shift work is prevalent worldwide, with ~15% of the workforce performing night shift work in industrialized countries1, and it increases the risk for cardiovascular disease (CVD)2,3,4,5,6,7. For instance, longer duration of night shift work was associated with increased coronary heart disease (CHD) risk in individuals followed over 24 years8. Importantly, this increased risk cannot be fully explained by differences in lifestyle and socioeconomic status9. Such observations highlight the urgent need to understand the underlying mechanisms and develop evidence-based countermeasures to mitigate these adverse health effects. Circadian misalignment, the misalignment between the central circadian pacemaker and the behavioral sleep/wake cycle (typical in shift workers) causes negative cardiometabolic changes10,11,12,13,14.
The adverse effects of circadian misalignment on cardiovascular risk factors in humans can be determined using stringently controlled laboratory protocols, including Forced Desynchrony (FD) and simulated shift work protocols15. In a FD protocol, in which participants were exposed to seven 28-h “days” under dim light (<3 lx), waketime blood pressure was higher during circadian misalignment. These misalignment-induced effects on blood pressure were not explained by changes in waketime heart rate, cortisol, nor sympathoadrenal measures (e.g., urinary epinephrine and norepinephrine), which remained unchanged11. While FD protocols provide experimental evidence for the adverse circadian misalignment effects on cardiovascular function, its translational value may be limited because shift workers do not live on 28-h “days”. Therefore, simulated shift work protocols offer a closer approximation to what shift workers typically experience while controlling for behavioral/environmental factors that directly affect cardiovascular function. For instance, a randomized crossover clinical trial showed that circadian misalignment increased 24-h systolic and diastolic blood pressure. At the same time, it decreased waketime cardiac vagal modulation (e.g., pNN50, percentage of consecutive heartbeat intervals differing by >50 ms, and RMSSD, root mean square of successive differences between normal heartbeats)16. These experimental human studies indicate that exposure to circadian misalignment may act as an underlying mechanism for the increased risk of cardiovascular disease in shift workers17. Since shift work is not foreseen to disappear, treatment interventions against the long-term adverse cardiovascular events are urgently needed. Preclinical work indicates that a well-consolidated 24-hour cycle of feeding and fasting can sustain cardiac health18. Food access limited to daytime protected against cardiac tissue aging in flies on either a normal or a fat-supplemented diet. Therefore, circadian organization is fundamental for normal health and longevity, while circadian disruption is implicated in the etiology of cardiac disease. Despite these recent breakthroughs, effective countermeasures against adverse cardiovascular effects due to circadian misalignment remain to be established in humans.
Here, we aim to identify whether circadian alignment of eating (despite the mistiming of sleep and of other behaviors) prevents the adverse effects of circadian misalignment on cardiovascular risk factors.
Results
Healthy participants [7 females, 12 males; age: 26.5 ± 4.1 y; body-mass index: 22.7 ± 2.1 kg/m2] (Table 1) underwent a randomized single-blind parallel clinical trial (Fig. 1) consisting of a 14-day circadian laboratory protocol to test the effects of simulated night work with either nighttime and daytime or only daytime eating on circadian rhythms of cardiovascular function compared to simulated day work (Fig. 2). No important harms nor unintended adverse effects were observed in the meal timing groups. The simulated night work was induced using a Forced Desynchrony protocol (FD; four 28-h “days”, see Methods). In the Nighttime Meal Control (NMC) Group, participants had a typical 28-h FD protocol with all behaviors scheduled across a 28-h cycle, including the fasting/eating cycle, which resulted in meals consumed during the daytime and nighttime (typical among shift workers). In the Daytime Meal Intervention (DMI) Group, participants underwent a modified 28-h FD protocol, with all behaviors scheduled on a 28-h cycle, except for the fasting/eating cycle, which was scheduled according to a 24-h cycle, resulting in meals consumed only during the daytime. We assessed the cardiovascular outcomes during Constant Routine protocols (CR, i.e., at least 32-hours of constant wakefulness, semi-recumbent posture, dim light conditions [<3 lx], and hourly isocaloric snacks19), before (baseline CR) and after simulated night work (post-misalignment CR). The CR approach allowed disentangling the contribution of the endogenous circadian control from the acute effects of fasting/eating, sleep/wake, rest/activity, body posture, and dark/light cycles. Because the effects of behaviors and environment are minimized during the CR protocol, it is ideally suited to assess the cardiovascular outcomes, which could have otherwise been affected during the FD study segment. Throughout the baseline and post-misalignment CR protocols, we repeatedly assessed cardiac vagal modulation by heart rate variability analyses (pNN50, RMSSD, LF/HF ratio, which are sensitive to circadian misalignment effects14), circulating concentrations of plasminogen activator inhibitor-1 (PAI-1, a key prothrombotic factor under circadian control20), blood pressure, heart rate, and plasma cortisol.

The clinical trial flow chart depicts the number of participants in the screening, allocation, and data analyses.

The 14-day stringently controlled circadian laboratory study included a parallel-design, randomized controlled trial with participants randomized to the Nighttime Meal Control (NMC) group or the Daytime Meal Intervention (DMI) group (see SI appendix for details). The study design is presented in relative clock time (for a participant with a habitual wake-up time of 7 a.m.).
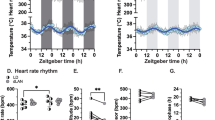
Heart rate variability
Our results showed that the meal timing intervention significantly modified the impact of simulated night work on pNN50 (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work interaction: pFDR = 0.008). The pNN50 levels decreased in the NMC group by 25.7% after exposure to simulated night work (post-misalignment CR), compared to that before (i.e., baseline CR) (95% confidence intervals [CI], −33.9% to −17.5%; Tukey-Kramer post-hoc test adjusted for multiple comparisons, P = 0.001; Fig. 3A). Conversely, no significant change was observed in the DMI group (95% CI, −8.9% to 4.9%; P = 0.23; Fig. 3B). Similarly, the meal timing intervention significantly modified the impact of simulated night work on RMSSD (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work intervention: pFDR = 0.02). The RMSSD levels significantly decreased in the NMC group by 14.3% (95% CI, −18.4% to −10.1%; P = 0.001; Fig. 3C), after simulated night work compared to baseline. Conversely, no significant change was observed in the DMI group (95% CI, −2.1% to 2.2%; P = 0.31; Fig. 3D). Likewise, the meal timing intervention significantly modified the impact of simulated night work on LF/HF (i.e., the ratio of low frequency and high cardiac frequency heart rate variability, a measure to estimate cardiac autonomic modulation) (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work interaction: pFDR = 0.04). The LF/HF ratio levels significantly increased in the NMC group by 5.5% (95%CI, 3.5% to 6.9%; P = 0.03; Fig. 3E), after simulated night work compared to baseline. Conversely, no significant change was observed in the DMI group (95% CI, −0.3% to 1.7%; P = 0.29; Fig. 3F). Of note, the meal timing intervention did not significantly modify the impact of simulated night work on the endogenous circadian rhythms (i.e., circadian phase and amplitude) of pNN50, RMSSD, and LF/HF ratio (cosinor mixed-model analyses; interaction of meal timing group, pre/post simulated night work, and circadian effect: pFDR = 0.34).

The meal timing intervention significantly modified the impact of simulated night work on pNN50 (A, B), RMSSD (C, D), LF/HF (E, F) and PAI-1 (G, H) levels. For all results, cosinor mixed-model analyses were used, and the interaction of meal timing group vs. pre/post simulated night work is reported in the figures. Cosine curves make use of precise circadian phase of each measurement for each participant. Circles indicate data grouped into 15°-circadian windows (~1-hour resolution, except for PAI-1 that is presented with 2-h resolution) with SEM error bars and the top x axes were scaled to the approximate group-averaged clock time of the circadian CBT minimum (reflected by 0o) for reference (i.e., relative clock time). Data correspond to the average (mean ± SEM) across participants per pre/post simulated night work condition and per meal timing group (n = 10 in the NMC group and n = 9 in the DMI group).
PAI-1
The meal timing intervention significantly modified the impact of simulated night work on circulating PAI-1 concentrations (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work interaction: pFDR = 0.02). The PAI-1 concentrations significantly increased in the NMC group by 23.9% (95%CI, −2.3% to −45.6%; P = 0.001; Fig. 3G), after simulated night work compared to baseline. Conversely, no significant change was observed in the DMI group (95% CI, −2.1% to 2.2%; P = 0.24; Fig. 3H).
Cortisol and Heart rate
The meal timing intervention did not significantly modify the effect of simulated night work on cortisol levels (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work interaction: P = 0.31; Fig. S1). Neither in the NMC group (95% CI, −5.4% to 0.9%) nor in the DMI group (95% CI, −2.1% to 5.6%) was there a significant change in cortisol levels (Tukey-Kramer post-hoc test adjusted for multiple comparisons, P = 0.29 and P = 0.33, respectively). Similarly, the meal timing intervention did not significantly modify the effect of simulated night work on the heart rate (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work interaction: P = 0.36; Fig. S2). Neither in the NMC group (95% CI, −0.4% to 2.8%) nor in the DMI group (95% CI, −1.2% to 2.1%) was there a significant change in heart rate (Tukey-Kramer post-hoc test adjusted for multiple comparisons, P = 0.39 and P = 0.32, respectively).
Blood pressure
The meal timing intervention significantly modified the impact of simulated night work on the SBP and DBP levels (cosinor mixed-model analyses; meal timing group vs. pre/post simulated night work interaction: respectively, P = 0.03 and P = 0.02, respectively; Fig. S3). In the NMC group, SBP and DBP levels did not significantly change after exposure to the simulated night work, compared to baseline (SBP: 95% CI, −1.3% to 2.8%; DBP: 95% CI, −1.1% to 0.7%; Tukey-Kramer post-hoc test adjusted for multiple comparisons, P = 0.22). Conversely, in the DMI group, blood pressure levels significantly decreased by 6-8% after exposure to the simulated night work, compared to baseline (SBP: −6.1%, 95% CI, −8.9% to −4.5%; DBP: −8.0%, 95% CI, −9.1% to −6.9%; Tukey-Kramer post-hoc test adjusted for multiple comparisons, P = 0.005 and P = 0.001). Of note, the meal timing intervention did not significantly modify the impact of simulated night work on the endogenous circadian rhythms (i.e., circadian phase and amplitude) of cortisol, heart rate, SBP, and DBP (cosinor mixed-model analyses; interaction of meal timing group, pre/post simulated night work, and circadian effect: P = 0.45, P = 0.51, P = 0.31 and P = 0.34, respectively).
Discussion
We found evidence for an increase in cardiovascular risk factors after simulated night work with nighttime and daytime eating, whereas a Daytime Meal Intervention minimized the adverse effects of simulated night shift work on cardiovascular risk factors.
Exposure to simulated night work with nightime and daytime eating decreased cardiac vagal modulation, with vagal (parasympathetic) activity typically considered cardioprotective16, and these findings are consistent with a previous clinical trial that induced circadian misalignment, i.e., 12-h inverted behavioral and environmental cycles for 3 days16. Circadian clocks within the suprachiasmatic nucleus in the hypothalamus (SCN) and the heart set daily rhythms in sinoatrial and atrioventricular node activity, with a consequential time-of-day dependent vulnerability to ventricular arrhythmia21. Exposure to circadian misalignment might alter the electrophysiological cardiac milieu, potentially increasing the susceptibility to adverse cardiac events. This may provide a mechanism underlying the elevated risk of cardiovascular disease and cardiac events in shift workers22,23 Additionally, circadian misalignment decreased PAI-1 concentration by 11% during misalignment in a previous simulated shift work study16, while we show an increase by 24% after misalignment24. Mice exposed to chronic circadian phase shifts had an increase in PAI-1 mRNA expression and PAI-1 levels and decreased tissue-type plasminogen activator mRNA expression in the liver25. Such findings and ours suggest that circadian disruption might induce hypofibrinolysis and increase cardiovascular risk by inducing PAI-1 gene expression following circadian misalignment.
Our results indicate no significant circadian misalignment nor meal timing effects on cortisol levels (Fig. S1), in line with previous results where acute11,12, and chronic26 circadian misalignment had a limited impact on cortisol levels. Similarly, we observed no significant circadian misalignment nor meal timing effects on heart rate (Fig. S2). While not directly comparable to our current study, time-restricted eating (i.e., 8-hour eating window from 7 a.m. to 3 p.m.) did not significantly affect heart rate, compared with a control eating pattern (i.e., a self-selected ≥12-hour window)27. Of note, the daytime meal intervention significantly reduced blood pressure levels by 6-8% after exposure to the simulated night work, compared to baseline, an effect that was not observed in the nighttime meal control (Fig. S3). The beneficial effect of appropriately aligned meal timing has been shown in a randomized control trial with shift workers who underwent 12 weeks of 10-h time-restricted eating compared to a standard of care (i.e., nutritional counseling and Mediterranean diet at usual eating times)28. Accordingly, participants consistently showed decreased diastolic blood pressure, compared to those who received standard of care. Similarly, a 14-week parallel-arm, randomized clinical trial weight-loss program with either early TRE (8-hour eating window from 7a.m.–3p.m.) or control eating (i.e., a self-selected ≥12-hour window) showed decreased diastolic blood pressure27. While our DMI findings bear resemblance to these TRE clinical trials, direct comparisons are challenging, as those studies, e.g., modified the fasting durations as part of the intervention, recruited participants who were overweight, included different sample sizes, and were long-term field studies. Of note, we did not observe endogenous circadian rhythms in blood pressure during baseline CR in either group. Circadian blood pressure rhythms have been described in previous laboratory studies29,30,31,32,33. While those previous studies used a Dynamap blood pressure monitor typically used for in-hospital spot-checks, our study used a Spacelabs blood pressure monitor typically used for 24-h automated blood pressure monitoring at home, which limits a direct comparison.
Preclinical work suggests dramatic effects of circadian rhythm disorganization on cardiovascular integrity34. A point mutation in the circadian regulatory gene, casein kinase-1, results in an intrinsic circadian period of 22 h (instead of approximately 24 h in wildtypes), early-onset cardiomyopathy, extensive fibrosis, and severely impaired contractility34. Intriguingly, under light cycles of 22 h, which match their intrinsic period (i.e., in resonance), their cyclic behavioral patterns and heart structure and function normalized, indicating the importance of synchrony between the internal circadian clock with environmental and behavioral cycles34. Moreover, preclinical work suggests a robust 24-h feeding/fasting cycle can maintain cardiac health in a high-fat fed aging Drosophila model, as compared to ad libitum fed flies that showed drastic deterioration of cardiac function, including increased arrhythmia index and reduced heart contractility18. In mice models, time-restricted feeding with food available only during the active nighttime reduced sympathetic activity during the resting daytime35. Here, we show that daytime meal timing avoids the adverse effects of simulated night work on cardiovascular function in humans. Critically, in our laboratory protocol, the design of the meal timing groups was identical (i.e., caloric and macronutrient intake, physical activity, posture, scheduled sleep schedule, and lighting conditions) except for the timing of meals. Moreover, our findings were unlikely due to group differences in participant sociodemographics and study-related characteristics (Table 1). Short sleep duration is associated with increased morbidity and mortality, particularly from cardiovascular disorders, such as CHD, arrhythmias, and hypertension36. As sleep structure before the baseline and simulated night work conditions did not differ between the meal timing groups24, differences in prior sleep are unlikely to mediate our reported effects.
Eating patterns are increasingly varied. Typical breakfast, lunch, and dinner meals during the daytime are no longer the social norm because skipping meals and snacking are becoming increasingly prevalent, as well as eating during the night37. Nighttime eating was associated with increased CHD risk compared with those who did not eat during the night after adjustment for sociodemographic data, diet, lifestyle, and CHD risk factors38. Data from 103,389 adults in the NutriNet-Santé study indicated that a later first meal (after 9 a.m. relative to before 8 a.m.) and a later last meal (after 9 p.m. relative to before 8 p.m.) was associated with elevated CVD risk39. Night shift workers show changes in meal patterns, skipping meals and consuming them during the night40. Therefore, meal timing is a potential modifiable lifestyle factor involved in the higher CVD risk in night workers. In this context, our study demonstrates that a meal-timing-based behavioral intervention can help mitigate the adverse cardiovascular effects induced by a mistimed sleep/wake schedule, as typically experienced by night workers. Limitations in our study include the restricted age range of participants (18–30 years), which limits its relevance to middle-aged and older populations. Moreover, caution is warranted as our study has a limited sample size (due to the stringent participant selection criteria and the intensive and controlled study design) and includes individuals who do not engage in actual shift work. Additionally, we performed an FD protocol to induce circadian misalignment, and while laboratory approaches offer the greatest experimental control, its findings have limited direct translation to night shift workers, as they do not live under such carefully controlled behavioral and environmental conditions. Future clinical trials with larger samples, including individuals undergoing real-life shift work schedules (e.g., permanent, rotating or irregular night shifts, morning shifts, and evening shifts), are needed to establish whether our reported beneficial effects on cardiovascular risk factors translate to the shift work population.
Methods
The protocol was approved by the Partners HealthCare’s institutional review board (IRB) and performed in accordance with the principles of the Declaration of Helsinki, and participants provided written informed consent. IRB guidelines were followed. Laboratory protocols were conducted at the Center for Clinical Investigation at Brigham and Women’s Hospital, Boston, United States, between 19 March 2015 and 29 August 2018. The study ended when the laboratory protocol for the final randomized participant was completed. Due to the complex study design, this is a single study performed in a human clinical trial. This randomized clinical trial provides proof-of-evidence for the beneficial effect of meal timing as a countermeasure against the adverse cardiovascular effects of circadian misalignment in humans, and future studies are needed to replicate our findings.
Participants and study randomization
Participants admitted to the laboratory protocols were free from medical conditions, including current and previous history of cardiometabolic disorders. Participants were not engaged in shift work, had not crossed more than one time zone in the previous 3 months, and did not have sleep disorders, including insomnia. Medical suitability was assessed using clinical history, biochemical and toxicology blood and urine screenings, and physical and psychological exams. Biochemical blood panels at screening included a comprehensive metabolic panel, TSH, complete blood count, and HbA1c, all of which had to be within typical ranges for study inclusion. Participants were not taking medications (excepting oral contraceptives), caffeine, smoking, or using recreational drugs (verified with urine toxicological panel). Participants underwent a randomized, parallel, controlled, single-blinded trial24, in which they were randomly assigned to one of two meal timing groups. The Nighttime Meal Control Group (NMC) completed a protocol that included simulated day work with day eating [baseline] followed by simulated night work with daytime and nighttime eating, typical for shift workers. The Daytime Meal Intervention Group (DMI) completed a protocol that included simulated day work with day eating (baseline) followed by simulated night work with daytime eating. Participants were randomized using minimization (Minim.exe, MS-DOS free access program for minimizing participants into the arms of a clinical trial) to minimize imbalance between meal timing groups. Minimization was performed—in decreasing sequence of importance—by sex, body mass index (BMI), and age (these factors were dichotomized, i.e., female or male, 18.5–24.9 kg/m2 or 25–29.9 kg/m2, and 18-26 years or 27-35 years, respectively).
Twenty healthy normotensive participants (mean age, 26.6 y [SD, 4.2 y, range: 18–35 y], eight females, BMI range: 18.5–29.9 kg/m2, hemoglobin A1C range: 4.9–5.4%) were randomized to the laboratory protocol: ten were allocated to the NMC Group and ten to the DMI Group (Fig. 1). Four females commenced the laboratory protocol on menstrual cycle days 1-5 (two per meal timing group) and four during days 14–19 (two per meal timing group). We excluded data from one participant in the DMI Group due to their inability to consume all food during the simulated night work. The final study sample included ten participants for the NMC Group (mean age, 27.0 years [SD, 4.4 y], 4 females, BMI: 22.5 kg/m2 [SD, 3.5]), and nine for the DMI Group (mean age, 26.2 years [SD, 4.1 y], 3 females, BMI: 23.1 kg/m2 [SD, 3.1]). Additional study-related characteristics between groups (e.g., diet, chronotype, among others) are shown in Table 1. No statistical differences in participant sociodemographics and study-related characteristics occurred between groups. Other aspects of this study, that was designed to test separate, independent hypotheses, have previously been published24,41,42,43.
Study design
Ambulatory segment
To ensure the initial stability of circadian rhythms and sleep patterns, participants were requested to maintain a regular sleep/wake schedule for two weeks prior to admission to the laboratory. The personalized laboratory sleep/wake schedule was determined as the participants’ habitual bedtime with 8-h time in bed. To ensure compliance with this schedule, each participant was monitored with at-home wristworn wearables (Actiwatch, Respironics). Participants also called into a time-stamped voice-mailbox when they were going to bed and getting out of bed. If more than one deviation (>1 h) per week from the target sleep/wake time was detected, participants were excluded from the study. In addition, during the three days before the laboratory study, participants received all meals (three meals and one snack) from the Metabolic Kitchen to meet dietary requirements (Harris-Benedict formula with activity factor 1.4) and controlled macronutrient distribution (45–50% carbohydrate, 15–20% protein, 30–35% fat, with 150 mEq Na+ (±20%), 100 mEq K+ (±20%)), to match the subsequent laboratory diet. Participants had to consume the provided meals at their habitual eating times to standardize the amount, type, and timing of food intake before the laboratory protocol. The latter was ensured by participants calling into a time-stamped voice-mailbox when they began each meal and by a daily food log that included food content and times. Participants were requested to consume their meals at the same time as in the laboratory relative to waketime (+/−1h). Participants who did not comply with the dietary requirements were excluded from the laboratory study. Moreover, participants were required to refrain from exercise (running/jogging, swimming, cycling, weight lifting, circuit training, and yoga) for 3 days prior to the laboratory protocol.
Laboratory protocol
Participants remained in individual laboratory suites in an environment free of time cues. Throughout the study, when participants were not involved in a study task, they could undertake leisure activities, such as reading, writing, watching movies, crafts, etc. We monitored each participant’s activity for compliance by means of closed-circuit TV and wrist-worn actigraphy. Days 1–2 comprised the laboratory adaptation days. Days 3-4 included a baseline Constant Routine (CR) protocol (Fig. 2), during which participants spent 32 h continuously awake in a constant semi-recumbent body posture, without physical exertion, in dim light (~3 lx in the horizontal angle of gaze) and eating hourly isocaloric snacks. This allowed a baseline assessment of endogenous circadian rhythms of physiologic markers by eliminating or minimizing the influences of behavioral and environmental factors on a given rhythm19. Following the baseline CR, participants had a 12-h sleep opportunity to recover. On Days 5 and 6, participants had further recovery from the baseline CR. On Day 7, participants underwent a 28-h Forced Desynchrony protocol (FD) to induce circadian misalignment, with 28-h sleep/wake cycles under dim light (~3 lx), to which the central circadian pacemaker in humans cannot entrain44. We used a 28-h FD protocol to assess the impact of circadian misalignment on cardiometabolic function11. During each 28-h cycle, the ratio of scheduled wakefulness (18 h:40 min) and sleep (9 h:20 min) was maintained at 2:1, to match the self-selected 8-h habitual time in bed per 24-h. The participants’ sleep episodes (0 lx) were split into three identical blocks, each separated by 1 h of wakefulness in dim light (~3 lx) while remaining at rest in a semi-recumbent posture in bed. This allowed the participants to consume food during the circadian day when otherwise they would be sleeping. Importantly, participants woke during each sleep episode irrespective of meal consumption to ensure that both study groups had three equal sleep blocks during each of the FD’days’. On the first 28-h sleep/wake cycle, participants had normal circadian alignment (waking up at their habitual wake-time, e.g., 7a.m.; baseline). In contrast, on the fourth sleep/wake cycle, participants were 12-h misaligned (wake up at, e.g., 7p.m.; simulated night work) as compared to the first cycle in both meal timing groups. The NMC Group had a typical 28-h FD protocol with all behaviors, including the fasting-eating cycle, maintained on a 28-h cycle. Because of that, three meals and a snack were scheduled at fixed times relative to scheduled wake-time (at 0 h:10 min, 4 h:10 min, 8 h:10 min, and 12 h:10 min since scheduled awakening, during baseline and simulated night work). Thus, each meal was shifted 4 h later each cycle, in alignment with the sleep/wake cycle. Participants in the NMC Group thus consumed food during both the daytime and nighttime, which is typical behavior of shift workers. In contrast, the DMI Group had a modified 28-h FD protocol, with all behaviors identically scheduled on a 28-h cycle, except for the fasting/eating cycle that was maintained on a 24-h cycle. Accordingly, participants had standardized meals at 0 h:10 min, 4 h:10 min, 8 h:10 min, and 12 h:10 min relative to the habitual wake time of the participant, regardless of the sleep/wake cycle. This meal timing approach in the DMI Group allowed alignment of the fasting/eating cycle to the ~24-h central circadian cycle and ensured meal consumption only during the daytime and at the same clock time during each FD cycle. After the four “days” of a 28-h FD protocol, participants underwent a post-misalignment CR (Days 11/12/13) that allowed assessing the impact of prior circadian misalignment on endogenous circadian rhythms. During the post-misalignment CR, participants spent 40-h under the same experimental conditions described for the baseline CR. Thereafter, participants were scheduled to a 12-h sleep opportunity to allow them to recover partially from the post-misalignment CR protocol, and then were discharged from the study.
Diet
CR protocol
Participants received an isocaloric diet (i.e., CR snacks) calculated according to the Harris-Benedict equation with an activity factor of 1.2 (as participants had decreased activity). The diet consisted of 45–50% carbohydrate, 15–20% protein, 30–35% fat, with 150 mEq Na+ (±20%), 100 mEq K+ (±20%), and at least 2.5 L of water per 24 h. CR snacks comprised two alternating CR options (e.g., CR snack A, then CR snack B, then CR snack A, and so forth). This was based on a food preference form for each participantʼs two CR preselected CR snack choices (2 of 6 snack choices with different ingredients but the same macronutrient composition). CR snacks were calculated with the same two snack options and the same caloric level throughout both CR protocols per participant. Participants had 10–15 minutes to consume the CR snacks and were instructed to consume all food provided (verified by checking their food trays). During the CR protocol, actual energy consumption in the NMC Group was 99.9% (SEM, 0.01) and 99.9% (SEM, 0.04%) during baseline CR and post-misalignment CR, respectively. In the DMI Group, it was 98.6% (SEM, 1.9%) and 99.9% (SEM, 0.01%) during baseline CR and post-misalignment CR, respectively.
Other segments of the laboratory protocol
During the FD protocol, participants received meals (breakfast, lunch, snack and dinner) standardized across days based on a food preference form for each participant. Meals were calculated according to a 28-h day for the NMC Group and to a 24-h day for the DMI Group, during the four “days” in the FD protocol. Diet was calculated according to the Harris-Benedict equation with an activity factor of 1.4, and consisted of 45–50% carbohydrate, 15–20% protein, 30–35% fat, with 150 mEq Na+ (±20%), 100 mEq K+ (±20%), and at least 2.5 L of water per 24 h. The energy content of the meals/snack (% of total “day’s” calorie intake) was as follows: Breakfast: 33.3% (±35 kcals); Lunch: 23.4% (±20 kcals); Snack: 10% ( ± 10 kcals); Dinner: 33.3% (±35 kcals). Breakfast and Dinner Test meals (used for glucose tolerance assessment published separately24) were preselected (one of two test meal choices, based on a food preference form for each participant).
During the other segments of the laboratory protocol, participants received an isocaloric diet, calculated according to the Harris–Benedict equation with an activity factor of 1.4. The diet had the same macronutrient composition as for the CR and FD segments, which was 45–50% carbohydrate, 15–20% protein, 30–35% fat, with 150 mEq Na+ (±20%), 100 mEq K+ (±20%), and at least 2.5 L of water per 24 h. Participants were instructed to consume all food provided (verified by checking their food trays).
Previously reported primary and secondary outcomes of the study
The primary and secondary outcomes of this study are reported in clinicaltrials.gov (clinical trial registration: NCT02291952). Accordingly, the primary outcomes of this study as reported in ref. 24 were changes in glucose tolerance, plasma insulin levels, and plasma leptin levels. These primary outcomes were published in24, except for the leptin results, due to funding constraints and issues regarding preprocessing of the blood assays. The secondary outcomes of this study were: changes in circadian phase markers, such as from core body temperature and cortisol, in the circadian rhythm in resting energy expenditure, in hunger and appetite, mood, and cognitive performance, in microbiota, gene expression, epigenetic or proteomic markers, and in sleep. Most of these secondary outcomes have been published16,41,42,43.
Current primary and secondary outcomes of the study
The study is a secondary analysis of our randomized clinical trial, and here we assessed the following cardiovascular measurements:
Primary outcomes
Heart rate variability (pNN50, RMSSD, and LF/HF ratio). Electrocardiographic (ECG) measurements were performed continuously during the CR protocols, and data were binned into every 10-min.
Secondary outcome
PAI-1 concentration. Assays were obtained using blood samples every 2 hours during the baseline and post-misalignment CR protocols.
Exploratory outcomes
1) Blood pressure and heart rate. Measurements were obtained with a Spacelabs 90217 ambulatory blood pressure monitor (Spacelabs Medical, Inc.) placed on the non-dominate arm. 2) Cortisol concentration. Assays were obtained using blood samples every hour during the baseline and post-misalignment CR protocols.
HRV preprocessing
Electrocardiographic (ECG) measurements were performed continuously during the CR protocols, and data were binned into 10-min bins. R-wave peak detection was done using ARISTOTLE45 following the same procedures as outlined in previously published work46,47. We established an analysis pipeline using MATLAB (Ver. R2022a; The MathWorks Inc., Natick, MA, USA) for HRV analyses. Specifically, consecutive R peaks were used to construct the RR interval time series. As the R peak detection was based on an automated approach, anomalous intervals due to false detections could not be avoided. They are usually manifested as spikes in the time series. To diminish the influence of these spikes on subsequent analyses, an impulse rejection filter was first applied, and if spikes were ever identified, they were replaced by the median value of the surrounding five samples48. Time- and frequency-domain analyses of RR interval time series were done per 10-min fixed time window. Time-domain measures included the root mean square of the successive differences in RR intervals (RMSSD) and the proportion of the number of times that successive normal sinus intervals differed greater than 50 ms (pNN50). Greater values of RMSSD or pNN50 mean higher vagal modulation. No results were rendered if the 10-min time series contained less than 20 RR intervals. For frequency-domain analysis, each 10-min time series was first evenly resampled. The power spectral density (psd) was computed using the Welch’s method with an added Hanning window. The power of low-frequency (LF; 0.04–0.15 Hz) and high-frequency (HF; 0.15–0.4 Hz) bands were then calculated. Note that for a reliable estimation of the psd, at least two consecutive segments, each with 150-s data, were required. The PSD was performed separately to each 150-s data, and the final psd for the 10-min time series was estimated by averaging all available psd’s. If less than two consecutive 150-s data were available, no frequency-domain results were rendered.
Blood pressure and heart rate
Blood pressure and heart rate measurements started shortly after constant routine protocol started until scheduled recovery sleep time. Measurements were obtained with a Spacelabs 90217 ambulatory blood pressure monitor (Spacelabs Medical, Inc.) placed on the non-dominate arm. Such method has been validated according to the Association for the Advancement of Medical Instrumentation’s standards49. Due to data loss, we had full time-series data for seven participants in the NMC group and six participants in the DMI group for the blood pressure and heart rate measurements.
Venous blood collection and processing
PAI-1 and cortisol assays were obtained using blood samples every 2-hours and one hour, respectively, during the baseline and post-misalignment CR protocols. On admission to the laboratory, an 18-gauge intravenous catheter was inserted into the participant’s forearm. The catheter was connected to a triple-stopcock manifold (Cobe Laboratories Inc., Lakewood, CO) via an intravenous loop with a 12-foot small-lumen extension cable (Sorex Pharmaceuticals, Salt Lake City, UT) through which blood sampling could continue in the next room without disturbing sleep. Between samples, infusion of a solution of 0.45% saline with 5000 IU/liter heparin at one drop every 5 to 10 sec maintained patency. Blood was transferred to 5-cm3 vacutainer tubes and centrifuged at 4 °C, pipetted into polystyrene tubes, and frozen at −80 °C until analysis. Participants’ hematocrit and hemoglobin were measured on each CR day and on FD day 1 and day 4 (when blood measurements took place) to assess whether levels remained within normal range.
Statistical analysis
We performed statistical analyses using SAS version 9.4 (SAS Institute, Cary, NC, USA). The sample size derived from the difference in the effect of misalignment on glucose tolerance (primary endpoint: 3 h postprandial plasma glucose profiles; for details, see24) between the meal timing groups. To determine a large effect size (d = 1.5) with −80% power, eight participants per group were required (total sample = 16). We compared participants’ characteristics with Yates’s chi-squared tests or two-tailed t-tests for independent groups, and their demographics and study-related characteristics did not significantly differ between the meal timing groups (Table 1).
Females and males were included in the study with a 1:1 ratio. Biological sex was self-reported. This study was not designed to test for sex-differences and the sample size calculation did not include sex as a potential factor of interest, as there are no sex differences reported for meal timing studies in humans. Thus, we did not test for interaction effects of sex or menstrual phase because of the limited sample size. Regarding race, we included 1 Black, 1 Asian American, and 8 White participants in the control group, and 1 Black and 8 White participants in the intervention group. Regarding ethnicity, we included 10 non-Hispanics in the control group, and 7 non-Hispanics and 2 Hispanics in the intervention group. The race and ethnicity ratio are in accordance with the population demographics census of 2014 for the state of Massachusetts, United States. Race and ethnicity were self-reported. This study was not designed to test for race and ethnicity differences and the sample size calculation did not factor these as a potential covariate of interest, as there are no race and ethnicity differences reported for meal timing studies in humans. Thus, we did not test for such differences in the effects of the intervention because of the limited sample size.
We assessed cardiac vagal modulation by heart rate variability analyses, blood pressure, and blood assays of cortisol and PAI-1 before and after the simulated night work (i.e., pre/post night work, i.e., baseline CR and post-misalignment CR, respectively). This CR approach allowed disentangling the contribution of endogenous circadian control from the acute effects of fasting/eating, sleep/wake, rest/activity, and dark/light cycles19. We first performed tests of normality (the Kolmogorov–Smirnov test and the Shapiro–Wilk test). As the outcomes had non-normal data distribution, we normalized the cardiovascular data using an average of each participant’s levels measured throughout the Baseline CR. This approach also allowed minimizing interindividual differences in physiologic measures. We applied a mean ± 3 SD (standard deviation) filter to remove outliers on the raw data. Data from the first 5 h after starting the CRs were excluded from analysis, as is standard, to allow for stabilization of circadian rhythms. The effects of the circadian cycle and circadian alignment condition were assessed by cosinor analyses using mixed model analyses of variance (PROC MIXED, SAS), which were applied to all cardiovascular outcomes. These cosinor mixed-models included “Circadian effect” (a fundamental circadian component of ∼24-h), “Time since scheduled waketime” (a linear effect of hours into the CR protocol), “pre/post simulated night work” (baseline CR vs. post-misalignment CR), “meal timing group” (NMC and DMI), and the interaction of meal timing group and pre/post night work. Additionally, we tested whether the meal timing intervention modified the impact of simulated night work on the endogenous circadian cardiovascular rhythms (i.e., circadian phase and amplitude) using cosinor mixed-model analyses that included the interaction of meal timing group, pre/post night work, and circadian effect. In the results section, we also report the percentage of change from baseline to post-simulated night work, per group. This corresponds to the post-hoc results from the interaction of meal timing group and pre/post night work, and are presented for each group separately, as we used a parallel design (i.e., while each participant serves as their baseline per meal timing condition, different participants were enrolled for the two meal timing groups).
Missing data were not included in the cosinor mixed-model analyses (pNN50: 0.81%; RMSSD: 0.81%; LF/HF: 0.82%, and PAI-1: 1.12%). Participant was included as a random factor. Post-hoc comparisons used the Tukey-Kramer test to adjust for multiple testing. To control overall type I error in null hypothesis testing when conducting multiple comparisons, P-values from the mixed-model analysis were adjusted using False Discovery Rates (pFDR) (PROC MULTTEST, SAS) for the primary (i.e., pNN50, RMSSD, LF/HF ratio) and secondary (PAI-1) cardiovascular outcomes. Unless specified, data are the mean and standard error of the mean. The significance for all statistical tests was set as P < 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data that support the findings of this study are available on request from the corresponding authors (S.L.Chellappa@soton.ac.uk; fscheer@bwh.harvard.edu). As per the NIH Policy on Data Sharing, we will make the datasets that underlie the results reported in this article available to other investigators following publication of the final study results. All proposals for data will be reviewed and will ensure that the proposals are complete and valid and that the data are available, consistent with participant privacy and informed consent by the corresponding author. Responses will be provided within three months. Such datasets will not contain identifying information per the regulations outlined in HIPAA. Per standard Partners HealthCare System policies, we will require from any investigator or entity requesting the data a data-sharing agreement that provides for: (1) a commitment to using the data only for research purposes and not to identify any individual participant; (2) a commitment to securing the data using appropriate computer technology; and (3) a commitment to destroying or returning the data after analyses are completed.
References
Survey NNHI. Public-use data file and documentation. Available from ftp.cdc.gov/pub/Health_Statistics/ NCHS/Dataset_Documentation/NHIS/2010/samadult_freq.pdf. Accessed Feb 19 2024., (2010).
Ellingsen, T., Bener, A. & Gehani, A. A. Study of shift work and risk of coronary events. J. R. Soc. Promot. Health 127, 265–267 (2007).
Ha, M. & Park, J. Shiftwork and metabolic risk factors of cardiovascular disease. J. Occup. Health 47, 89–95 (2005).
Hermansson, J., Hallqvist, J., Karlsson, B., Knutsson, A. & Gillander Gadin, K. Shift work, parental cardiovascular disease and myocardial infarction in males. Occup. Med. 68, 120–125 (2018).
McNamee, R. et al. Shiftwork and mortality from ischaemic heart disease. Occup. Environ. Med. 53, 367–373 (1996).
Oishi, M. et al. A longitudinal study on the relationship between shift work and the progression of hypertension in male Japanese workers. J. Hypertens. 23, 2173–2178 (2005).
Torquati, L., Mielke, G. I., Brown, W. J. & Kolbe-Alexander, T. Shift work and the risk of cardiovascular disease. A systematic review and meta-analysis including dose-response relationship. Scand. J. Work Environ. Health 44, 229–238 (2018).
Vetter, C. et al. Association Between Rotating Night Shift Work And Risk Of Coronary Heart Disease Among Women. JAMA 315, 1726–1734 (2016).
Karlsson, B., Knutsson, A. & Lindahl, B. Is there an association between shift work and having a metabolic syndrome? Results from a population based study of 27,485 people. Occup. Environ. Med. 58, 747–752 (2001).
Morris, C. J. et al. Endogenous circadian system and circadian misalignment impact glucose tolerance via separate mechanisms in humans. Proc. Natl Acad. Sci. USA 112, 13 (2015).
Scheer, F. A., Hilton, M. F., Mantzoros, C. S. & Shea, S. A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl Acad. Sci. USA 106, 4453–4458 (2009).
Morris, C. J., Purvis, T. E., Mistretta, J. & Scheer, F. A. Effects of the internal Circadian system and circadian misalignment on glucose tolerance in chronic shift workers. J. Clin. Endocrinol. Metab. 101, 1066–1074 (2016).
Grimaldi, D., Carter, J. R., Van Cauter, E. & Leproult, R. Adverse impact of sleep restriction and circadian misalignment on autonomic function in healthy young adults. Hypertension 68, 243–250 (2016).
Chellappa, S. L., Vujovic, N., Williams, J. S. & Scheer, F. Impact of Circadian disruption on cardiovascular function and disease. Trends Endocrinol. Metab. 30, 767–779 (2019).
Qian, J. & Scheer, F. Circadian system and glucose metabolism: implications for physiology and disease. Trends Endocrinol. Metab. 27, 282–293 (2016).
Morris, C. J., Purvis, T. E., Hu, K. & Scheer, F. A. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc. Natl Acad. Sci. USA 113, 8 (2016).
Morris, C. J., Purvis, T. E., Mistretta, J., Hu, K. & Scheer, F. Circadian misalignment increases C-reactive protein and blood pressure in chronic shift workers. J. Biol. Rhythms 32, 154–164 (2017).
Gill, S., Le, H. D., Melkani, G. C. & Panda, S. Time-restricted feeding attenuates age-related cardiac decline in Drosophila. Science 347, 1265–1269 (2015).
Duffy, J. F. & Dijk, D. J. Getting through to circadian oscillators: why use constant routines? J. Biol. Rhythms 17, 4–13 (2002).
Scheer, F. A. & Shea, S. A. Human circadian system causes a morning peak in prothrombotic plasminogen activator inhibitor-1 (PAI-1) independent of the sleep/wake cycle. Blood 123, 590–593 (2014).
Hayter, E. A. et al. Distinct circadian mechanisms govern cardiac rhythms and susceptibility to arrhythmia. Nat. Commun. 12, 2472 (2021).
Vyas, M. V. et al. Shift work and vascular events: systematic review and meta-analysis. BMJ 345, e4800 (2012).
Meloni, M., Setzu, D., Del Rio, A., Campagna, M. & Cocco, P. QTc interval and electrocardiographic changes by type of shift work. Am. J. Ind. Med. 56, 1174–1179 (2013).
Chellappa, S. L. et al. Daytime eating prevents internal circadian misalignment and glucose intolerance in night work. Sci. Adv. 7, eabg9910 (2021).
Oishi, K. & Ohkura, N. Chronic circadian clock disruption induces expression of the cardiovascular risk factor plasminogen activator inhibitor-1 in mice. Blood Coagul. Fibrinol. 24, 106–108 (2013).
Wright, K. P. Jr. et al. Influence of sleep deprivation and circadian misalignment on cortisol, inflammatory markers, and cytokine balance. Brain Behav. Immun. 47, 24–34 (2015).
Jamshed, H. et al. Effectiveness of early time-restricted eating for weight loss, fat loss, and cardiometabolic health in adults with obesity: a randomized clinical trial. JAMA Intern Med 182, 953–962 (2022).
Manoogian, E. N. C. et al. Feasibility of time-restricted eating and impacts on cardiometabolic health in 24-h shift workers: The Healthy Heroes randomized control trial. Cell Metab. 34, 1442–1456 e1447 (2022).
Scheer, F. A. et al. Impact of the human circadian system, exercise, and their interaction on cardiovascular function. Proc. Natl Acad. Sci. USA 107, 20541–20546 (2010).
Hu, K., Scheer, F. A., Laker, M., Smales, C. & Shea, S. A. Endogenous circadian rhythm in vasovagal response to head-up tilt. Circulation 123, 961–970 (2011).
Shea, S. A., Hilton, M. F., Hu, K. & Scheer, F. A. Existence of an endogenous circadian blood pressure rhythm in humans that peaks in the evening. Circ. Res 108, 980–984 (2011).
Scheer, F., Chellappa, S. L., Hu, K. & Shea, S. A. Impact of mental stress, the circadian system and their interaction on human cardiovascular function. Psychoneuroendocrinology 103, 125–129 (2019).
Qian, J., Scheer, F. A., Hu, K. & Shea, S. A. The circadian system modulates the rate of recovery of systolic blood pressure after exercise in humans. Sleep 43, zsz253 (2020).
Martino, T. A. et al. Circadian rhythm disorganization produces profound cardiovascular and renal disease in hamsters. Am. J. Physiol. Regul. Integr. Comp. Physiol. 294, 13 (2008).
Hou. T. et al. Time-restricted feeding protects the blood pressure circadian rhythm in diabetic mice. Proc. Natl. Acad. Sci. USA 118, e2015873118 (2021).
Tobaldini, E. et al. Short sleep duration and cardiometabolic risk: from pathophysiology to clinical evidence. Nat. Rev. Cardiol. 16, 213–224 (2019).
St-Onge, M. P. et al. Meal timing and frequency: implications for cardiovascular disease prevention: a scientific statement From the American Heart Association. Circulation 135, e96–e121 (2017).
Cahill, L. E. et al. Prospective study of breakfast eating and incident coronary heart disease in a cohort of male US health professionals. Circulation 128, 337–343 (2013).
Palomar-Cros, A. et al. Dietary circadian rhythms and cardiovascular disease risk in the prospective NutriNet-Sante cohort. Nat. Commun. 14, 7899 (2023).
Kosmadopoulos, A. et al. Effects of shift work on the eating behavior of police officers on patrol. Nutrients 12, 999 (2020).
Chellappa, S. L. et al. Proof-of-principle demonstration of endogenous circadian system and circadian misalignment effects on human oral microbiota. FASEB J. 36, e22043 (2022).
Qian, J. et al. Daytime eating prevents mood vulnerability in night work. Proc. Natl Acad. Sci. USA 119, e2206348119 (2022).
Scheer, F. & Chellappa, S. L. Endogenous circadian rhythms in mood and well-being. Sleep. Health 10, 149–153 (2024).
Czeisler, C. A. et al. Stability, precision, and near-24-hour period of the human circadian pacemaker. Science 284, 2177–2181 (1999).
Moody, G. & Mark, R. G. Development and evaluation of a 2-lead ECG analysis program. Comput Cardiol. 4, 39–44 (1982).
Gao, L. et al. Resting Heartbeat Complexity Predicts All-Cause and Cardiorespiratory Mortality in Middle- to Older-Aged Adults From the UK Biobank. J. Am. Heart Assoc. 10, e018483 (2021).
Gao, L. et al. Nocturnal heart rate variability moderates the association between sleep-wake regularity and mood in young adults. Sleep 42, zsz034 (2019).
Shi, B., Motin, M. A., Wang, X., Karmakar, C. & Li, P. Bivariate entropy analysis of electrocardiographic RR-QT Time Series. Entropy 22, 1439 (2020).
Baumgart, P. & Kamp, J. Accuracy of the SpaceLabs Medical 90217 ambulatory blood pressure monitor. Blood Press Monit. 3, 303–307 (1998).
Acknowledgements
This study was funded by Grant number NIH R01HL118601 (ClinicalTrials.gov number: NCT02291952), 1UL1TR001102 and 1UL1TR002541-01. S.L.C. was supported by R01HL118601 and the Alexander Von Humboldt Foundation. J.Q. is supported by NIH R00HL148500. K.H. is supported by NIH RF1AG064312 and R01AG083799. F.A.J.L.S. was supported by NIH R01HL118601, R01DK099512, R01DK102696, R01DK105072 and R01HL140574. The funder had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish it.
Author information
Authors and Affiliations
Contributions
Conceptualization: F.A.J.L.S. Funding acquisition: F.A.J.L.S. Investigation: S.L.C., J.Q. and N.V. Visualization: S.L.C., L.G., J.Q., N.V., P.L., K.H. and F.A.J.L.S. Project administration: S.L.C., J.Q. and N.V. Supervision: F.A.J.L.S. Data curation: S.L.C., L.G., P.L., K.H. and F.A.J.L.S. Formal analysis: S.L.C. Validation: F.A.J.L.S. Writing—review and editing: S.L.C., L.G., J.Q., N.V., P.L., K.H. and F.A.J.L.S.
Corresponding authors
Ethics declarations
Competing interests
F.A.J.L.S. served on the Board of Directors for the Sleep Research Society and has received consulting fees from the University of Alabama at Birmingham and Morehouse University. F.A.J.L.S. interests were reviewed and managed by Brigham and Women’s Hospital and Partners HealthCare in accordance with their conflict-of-interest policies. F.A.J.L.S. consultancies are not related to the current work. The other authors declare that they have no competing interests.
Peer review
Peer review information
Nature Communications thanks Tami Martino and the other anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’snote Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Chellappa, S.L., Gao, L., Qian, J. et al. Daytime eating during simulated night work mitigates changes in cardiovascular risk factors: secondary analyses of a randomized controlled trial. Nat Commun 16, 3186 (2025). https://doi.org/10.1038/s41467-025-57846-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-57846-y
This article is cited by
-
Night shift work, dietary patterns, and coronary heart disease
European Journal of Epidemiology (2026)
-
Factors influencing heart rate variability in nurses following night shifts: a prospective observational clinical study
BMC Nursing (2025)
-
Circadian nutrition and obesity: timing as a nutritional strategy
Journal of Health, Population and Nutrition (2025)
