
Abstract
Neurodevelopmental disorders (NDD) are characterized by impairments in brain development that affect motor and cognitive systems. Early identification of NDD risk factors is crucial for prevention. Metabolomic profiling holds promise for early detection but remains underexplored in longitudinal studies. Here, we map the longitudinal metabolomic profiles in 3212 samples covering 683 metabolites from 581 mother-child pairs at six timepoints in the Danish COPSAC2010 cohort to assess childhood NDD risk over a 10-years follow-up period. Cross-validated sPLS models identify metabolic biomarkers and patterns associated with NDD, with maternal gestational week 24 metabolic profile showing the best prediction. Metabolite trajectories prior 18-month of age are linked to NDD risk. Quinolinate is consistently associated with NDD across timepoints and mediated the effect of maternal inflammation on NDD. In the single metabolite analysis, no metabolites remained significant after multiple testing correction. In this work, we provide valuable insights into the role of the longitudinal metabolome in neurodevelopment.
Similar content being viewed by others

Introduction
Neurodevelopmental disorders (NDD) are characterized by impairments in brain development that affect motor and cognitive systems. These difficulties may in turn compromise social abilities, learning abilities, and behavior later in life1,2. Attention-deficit/hyperactivity disorder (ADHD) and autism spectrum disorder (ASD) are the two most prevalent NDD3. Approximately 15–20% of children are affected by NDD, with reported prevalence rates of 5–11% for ADHD, and 0.70–3% for ASD4. NDD manifests in early childhood, and the fetal environment may play a critical role5. The negative effects of NDD often persist throughout life, posing a significant public health concern and highlighting the necessity to better understand and support healthy neurodevelopment, especially in early life4,6.
Both ADHD and ASD are highly heritable disorders (up to 88% for ADHD and up to 91% for ASD) involving complex and heterogeneous neurodevelopmental, cognitive, and behavioral processes7,8,9,10,11. Metabolites are both intermediate and end products of cellular processes, acting as pivotal connections from the genome to the proteome, metabolome, and the clinical phenotype. Studies suggest that individuals with NDD, autism or ADHD symptoms show differences in their metabolome compared to neurotypical participants12,13, particularly in amino acids, sphingolipids, and vitamin profiles13,14,15. Increased plasma levels of sphingosine-1-phosphate and sphinganine-1-phosphate have been reported in ADHD patients15, while reduced levels of glutathione and tryptophan have been associated with autism and autism severity14. Additionally, lower plasma levels of cortisol, triacylglycerols and phospholipids and elevated levels of γ-tocopherol have been observed in children aged 6 to 14 with ADHD16,17. Alterations in the kynurenine pathway—such as increased kynurenic acid and 3-hydroxyanthranilic acid—have also been associated with ADHD-like behaviors in mouse models of ADHD and ASD18. In children with ASD, lower levels of linolenic acid and long-chain polyunsaturated fatty acids, higher levels of perfluorodecanoic acid, and alterations of a range of triglycerides have been reported19,20. Exposures before onset of disease are more likely to be causally involved in the disease than cross-sectional findings21,22,23. However, previous metabolomics studies predominantly focused on animal models or data from children at specific timepoints in case-control designs, rather than examining the pregnancy period or tracking metabolic changes longitudinally through childhood. Additionally, limited sample sizes and the absence of robust clinical validations restrict the generalizability of current findings.
The fetal neurodevelopmental period represents a time of significant brain growth and differentiation24. This period of accelerated neurodevelopment thus represents a window of increased vulnerability, with early adversities potentially increasing the risk of developing NDD25. Key neurodevelopmental processes, such as neuronal apoptosis, myelination, synaptogenesis, neurogenesis and proliferation occur during the second and third trimesters25,26,27, but our understanding of the mechanisms underlying abnormal neurodevelopment and NDD is still limited. Metabolic alterations during pregnancy have been observed in rodent models of NDD, such as ASD28,29. Similarly, maternal metabolome changes during pregnancy have previously been associated with the risk of ASD assessed during childhood30,31. We hypothesize that metabolic perturbations during pregnancy and early childhood may contribute to the risk of developing NDD, and longitudinal studies tracking metabolic alterations across these developmental stages are therefore essential.
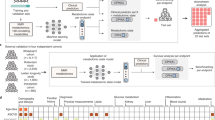
In this work, we analyzed >3000 plasma metabolomic profiles consisting of 683 metabolites from 581 mother-child pairs in the Danish COPSAC2010 cohort. Plasma samples were collected from pregnancy through age 10 as part of the cohort’s original design, with a particular focus on early life to capture critical developmental windows. Samples were collected at 6 timepoints from gestational week 24, week 1 postpartum, child age 6 months, 18 months, 6 years and age 10 years to investigate potential metabolite biomarkers, patterns and trajectories related to the risk of NDD at age 10.
Results
Baseline characteristics
A total of 604 children of the COPSAC2010 cohort participated in the follow-up at age 10 years. Of the 604 children, 593 underwent psychopathological assessment. The numbers of participants included at each time point are shown in Supplementary Fig. 1. Of the 593 children, 581 (98%) of their mothers had metabolomic profiling at gestational week 24 and 579 (98%) at one week postpartum, 520 (88%), 535 (90%), 477 (80%) and 520 (88%) of the children had metabolomic profiling at 6 months, 18 months, 6 years, and 10 years, respectively. Figure 1a, b presents the distribution of metabolome data across all timepoints, analyzed using principal component analysis (PCA), as well as the study design including participants and classes of studied metabolites.

a PCA for all metabolome data from all time points included in this study. b Number of participants included in each time point and classes of studied metabolites. TCA citric acid cycle, PCM partially characterized molecules, OP oxidative phosphorylation, PC principal component, ADHD attention-deficit/hyperactivity disorder, Any NDD any neurodevelopmental disorders. Source data are provided as a Source data file.
Of the 581 children with both psychopathological assessments and maternal metabolome data available from gestational week 24, 86 (15%) met criteria for at least one NDD, 63 (11%) had ADHD, and 16 (3%) had autism by age 10. A total of 59 (11%) had other psychiatric disorders than NDD (e.g., anxiety disorders, enuresis, and encopresis), and 454 (78%) had no mental disorders and were considered neurotypical participants. More boys (n = 61, 71%) than girls (n = 25, 29%) fulfilled criteria for any NDD (P < 0.001). Mothers (P < 0.001) and fathers (P = 0.04) of children with any NDD had lower educational attainment (Table 1). Baseline characteristics of children with and without ADHD or autism are presented in Supplementary Data 1 and 2.
Association between metabolome profile and NDD at age 10
For any NDD, the sparse partial least squares (sPLS) metabolome model showed a cross-validated median repeat area under the curve (AUC) of 0.60 (range of repeats 0.59–0.62) for gestational week 24, 0.57 (0.52–0.60) for one week postpartum, 0.54 (0.50–0.56) for 6 months, 0.61 (0.58–0.63) for 18 months, 0.41 (0.37–0.45) for 6 years, and 0.55 (0.51–0.59) for 10 years. Among these, metabolome data from gestational week 24 and child age 18 months of children demonstrated the highest AUC value compared to the other time points (Fig. 2a–g).

a–f AUC (area under the curve) from repeated tenfold CV (cross-validation) of an sPLS (sparse partial least squares) model for risk of any NDD (neurodevelopmental disorders) at different time points. The box plots represent the diagnosis predictions for the median cross-validated model. The center of the boxes represents the median, their bounds represent the 25th and 75th percentiles, and the lower and upper ends of whiskers represent the smallest and largest values, respectively, no further than 1.5 × IQR (interquartile range) from the respective end of the box plot. Sample sizes were N = 581 at gestational week 24, N = 579 at one week postpartum, and N = 520, 535, 477, and 520 at 6 months, 18 months, 6 years, and 10 years, respectively. g ROC curve of sPLS models of any NDD. ROC (receiver operating characteristic) curves were constructed for sPLS models at each timepoint, with the best-performing resample selected based on AUC. h Representation of the top 50 metabolites related to risk of any NDD, contributing to the optimal set of loadings for the 10-times repeated 10-fold cross-validation sPLS model (n = 100 separate model runs) using the entire samples of week 24 (N = 581 samples). Bars depict the median across repeats ± SD. Source data are provided as a Source data file.
The final sPLS model of any NDD selected 200 metabolites. Figure 2h displays the top 50 metabolites with the highest absolute numerical loading scores from the model at gestational week 24 ranked from high to low. Adipoylcarnitine (C6-DC), glutamine conjugate of C6H10O2, and caprate (10:0) from gestational week 24 were the strongest positively associated metabolites with the risk of any NDD, while 4-hydroxy-2-oxoglutaric acid, branched-chain 14:0 dicarboxylic acid, and gentisate were the metabolites most strongly negatively associated, showing protective effects against any NDD. Metabolites associated with any NDD identified by sPLS at one week postpartum and at 18 months of age were shown in Supplementary Fig. 3a, b.
For ADHD, the sPLS model showed an AUC of 0.60 (0.57–0.62) for week 24, 0.57 (0.55–0.61) for one week postpartum, 0.53 (0.49–0.58) for 6 months, 0.60 (0.56–0.64) for 18 months, 0.46 (0.43–0.49) for 6 years, and 0.51 (0.45–0.53) for 10 years (Supplementary Fig. 2). Among these, the metabolome data from pregnancy week 24 and child age 18 months demonstrated equal performance with AUC values of 0.6 and showed higher AUC than the other timepoints. Adipoylcarnitine (C6-DC) at week 24 of pregnancy was the metabolite most strongly positively associated with the risk of ADHD, while threonate was the strongest negatively associated metabolite. Metabolites associated with ADHD at one week postpartum (e.g., 5, 6-dihydrouridine, urate, and 3,4-dihydroxybutyrate) were shown in Supplementary Fig. 3c. At 18 months, only quinolinate was selected by the model and positively associated with the ADHD risk (Supplementary Fig. 3d).
In the nested five-fold CV model, only the gestational week 24 metabolome achieved comparable performance, with an AUC of 0.60 (SD = 0.05, 0.57–0.67) for any NDD and 0.59 (SD = 0.04, 0.54–0.65) for ADHD (Supplementary Results in Supplementary Information).
Association between metabolome-NDD risk scores and risk factors
The final sPLS model of gestational week 24 was selected based on the highest median AUC value and lowest variance across repeated cross-validation rounds, and the predicted values from the median repeat were used to extract the predicted metabolome-NDD risk scores. Our association analysis identified several potential risk factors linked to the gestational week 24 metabolome-NDD risk score. Specifically, pre-pregnancy BMI, gestational diabetes, an unhealthy Western dietary pattern derived from Food Frequency Questionnaires (FFQs)32, maternal inflammation measured as hs-CRP levels and maternal inflammation PC1 derived from 92 proteins33, antibiotic use during pregnancy, low family income, and low maternal and paternal educational attainment were associated with an increasing gestational week 24 metabolome-NDD risk score at a false discovery rate (FDR) 5% significance level (Supplementary Fig. 4a). Additionally, we examined the association between quinolinate levels at gestational week 24 and the same potential risk factors mentioned above. Consistently, hs-CRP levels, Western dietary pattern, pre-pregnancy BMI, and maternal inflammation PC1 were all positively associated with quinolinate levels (Supplementary Fig. 4b).
Metabolic trajectories from gestational week 24 and childhood NDD risk
We conducted longitudinal analyses using the metabolomic profiles at six timepoints. Since the metabolome during gestational week 24 provided the strongest association for both any NDD and ADHD using the sPLS full and nested model, we analyzed the differences in predicted risk scores based on the week 24 metabolome for these outcomes between children with vs. without NDD (Fig. 3). As the prediction model was built based on gestational week 24 metabolome, risk scores for the week 24 metabolome were calculated using the best fold from nested outfold CV of the testing set (n = 116). The analysis identified difference in metabolic risk scores over time for any NDD and ADHD. Significant differences in these scores emerged early in childhood, particularly before age 18 months, between children diagnosed with and without NDD or ADHD.

Risk scores from 6 months to 10 years were predicted based on a full model trained on the maternal week 24 metabolome. Sample sizes were N = 579 at one week postpartum, and N = 520, 535, 477, and 520 at 6 months, 18 months, 6 years, and 10 years, respectively. As the prediction model was built based on week 24 metabolome, risk scores for the week 24 metabolome were calculated using the scores predicted on the testing set (n = 116 samples) using the outfold of nested five-fold CV (cross-validation). Smoothed loess curves were used to depict trends over time. The Y axis represents the mean score difference between the NDD and ADHD risk scores calculated as: mean risk score (any NDD/ADHD = Yes)-mean risk score (any NDD/ADHD = No). The mean difference between any NDD and ADHD risk scores for individuals diagnosed with any NDD and ADHD and those without the diagnoses was calculated using the Wilcoxon rank-sum test, with a confidence level set at 95%. Two-sided P < 0.05 was considered significant. ADHD attention-deficit/hyperactivity disorder, Any NDD any neurodevelopmental disorders. Source data are provided as a Source data file.
There were significant differences between children with vs. without NDD at one week postpartum (p = 0.02), whereas this was non-significant at 6 months (P = 0.10), and 18 months (P = 0.08). For ADHD, there were significant differences at 1 week postpartum (P = 0.002), and 6 months (P = 0.04), whereas the difference disappeared by 18 months (P = 0.12). Additionally, we correlated the predicted metabolome-NDD risk scores for any NDD and ADHD based on the maternal week 24 metabolome with the NDD outcomes (Supplementary Data 3). Using all timepoints of children, the results demonstrated a trend of association for the metabolome-any NDD risk score (aOR = 1.04, 95% CI [1.00–1.07], P = 0.058) and the metabolome-ADHD risk score (aOR = 1.06 [1.01–1.11], P = 0.012), whereas there were significant associations using the timepoints until age 18 months for the metabolome-any NDD risk score (aOR = 1.06 [1.00–1.13], P = 0.046) and the metabolome-ADHD risk score (OR = 1.11 [1.02–1.20], P = 0.021), indicating that the early metabolic timepoints may be more important. We further adjusted for maternal exposures, including maternal smoking, maternal antibiotic use and alcohol use during pregnancy, which did not substantially modify the results (Supplementary Data 3).
Single metabolite analysis
We thereafter associated each individual metabolite at each timepoint to the risk of any NDD, ADHD, and autism, including conservative adjustment for potential confounders and multiple testing adjustments. No metabolites reached significance when adjusted for FDR 5%, while some metabolites exhibited nominal significance across the neurodevelopmental outcomes as outlined below.
In the maternal week 24 plasma metabolome (Fig. 4a–c), 54 metabolites were nominally associated with the risk of any NDD, exhibiting ORs ranging from 0.25 to 16.28 after adjusting for covariates. Sulfate, sphingomyelin (d18:1/18:1, d18:2/18:0), C-glycosyltryptophan, O-sulfo-tyrosine, and N-acetylneuraminate had the highest ORs. For ADHD, 39 metabolites achieved nominal significance in analyses adjusted for covariates (aOR range 0.13–25.83), with sulfate, C-glycosyl tryptophan, N-acetylneuraminate, 5-oxoproline, and quinolinate being the top positively associated metabolites with the highest ORs. For autism, 32 metabolites reached nominal significance after covariate adjustment (aOR range 0.01–11.88), with vanillylmandelate, N(1) + N(8)-acetylspermidine, creatine, sphingomyelin (d18:1/21:0, d17:1/22:0, d16:1/23:0), and picolinoyl glycine being the top positively associated metabolites with the highest OR.

a Maternal week 24 plasma metabolites associated with NDD in offspring (adjusted). b Maternal week 24 plasma metabolites associated with ADHD in offspring (adjusted). c Maternal week 24 plasma metabolites associated with autism in offspring, adjusted. d Associations between child age 18 months plasma metabolites and NDD at age 10 years. e Associations between child age 18 months plasma metabolites and ADHD at 10 years. f Associations between child age 18 months plasma metabolites and autism at 10 years. Two-sided P < 0.05 was considered significant. All associations are nominally significant, while none of them remained significant after FDR 5% correction. Top 20 metabolites with the highest absolute OR from each analysis were shown. g Associations between highlighted metabolites and neurodevelopmental outcomes. Note: N = 581 for w24, N = 579 for w1, N = 520, N = 535, N = 477, N = 520 for 6 months, 18 months, 6 years, and 10 years of children, respectively. Error bars represent covariate-adjusted log(OR) with 95%CI. Covariates adjusted for week 24 were maternal BMI, maternal age, smoking, antibiotic use, pre-eclampsia, diabetes and alcohol intake during pregnancy, household income, maternal and paternal educational attainment at birth, gestational age, child sex, and birth weight. Covariates of 18-months children: sex, maternal and paternal educational attainment, household income, and BMI Z-score of children. Logistic regression was applied and adjusted for multiple comparisons using FDR (a–g). P values and the FDR-corrected P values are available in the Source data file. “*” indicates nominal significance of P < 0.05 after covariates adjustment (a–g). w24 gestational week 24, w1 week 1 postpartum, 6mth 6 months of children, 18 mth 18 months of children, 6yr 6 years of children, 10yr 10 years of children, NDD neurodevelopmental disorder, ADHD attention deficit/hyperactivity disorder, OR odd ratio, CI confidence interval. Source data are provided as a Source data file.
In the child age 18 months, plasma metabolome (Fig. 4d–f), 27 metabolites were associated with risk of any NDD (aOR range 0.05–4.42) after adjusting for covariates. Hydroxyasparagine, N-acetylputrescine, 2’-O-methylcytidine, 3-methoxytyrosine, and quinolinate had the highest ORs, positively associated with any NDD. For ADHD, 30 metabolites achieved nominal significance (aOR range 0.04–8.31) with hydroxyasparagine, glucose, 2’-O-methylcytidine, 3-methoxytyrosine, and 1, 5-anhydroglucitol (1, 5-AG) being the top positively associated metabolites with the highest ORs. For autism, 56 metabolites were nominally significant (aOR range 0.01–9.96), with citrate, cortisone, and imidazole lactate being the top positively associated metabolites with the highest ORs. Associations between each single metabolite from the other time points and risk of different NDD outcomes are shown in Supplementary Fig. 5 and Supplementary Data 4. We additionally adjusted for the randomized controlled trials (RCTs) of fish oil and vitamin D supplementation during pregnancy, which did not significantly alter the findings. (Supplementary Fig. 6).
We identified three metabolites: quinolinate, ferulic acid 4-sulfate, and 8-methoxy kynurenate, which were consistently associated with at least one outcome: any NDD, ADHD, or autism, measured during week 24 of pregnancy, as well as at 6 and 18 months of age (Fig. 4g). Interestingly, levels of quinolinate at all the six time points were positively associated with at least one of the outcomes of any NDD, ADHD, or autism across all timepoints, particularly ADHD. Ferulic acid 4-sulfate levels from gestational week 24 and child age 6 months were negatively associated with the risk of any NDD, and levels from 6 months and 18 months also showed a negative association with the risk of ADHD and autism, respectively. Similarly, 8-methoxy kynurenate levels at 6 and 18 months were positively associated with the risk of any NDD and autism, respectively. Further adjusting for breastfeeding status at one week postpartum showed similar associations between the metabolome and NDD risk (Supplementary Fig. 7).
For ADHD and autism dimensional psychopathology, we observed a similar trend to NDD diagnosis for these metabolites, though with fewer significant data points (Supplementary Fig. 8).
To explore the impact of single versus multiple NDD diagnoses, we performed logistic regression analyses associating each individual metabolite from gestational week 24 and child age 6 months with the binary outcome (0 = ADHD only, 1 = multiple NDD diagnoses) (Supplementary Fig. 9). Notably, metabolites, such as quinolinate, bilirubin and its degradation products, and several sphingomyelins were positively associated with ADHD-only cases and negatively associated with multiple NDD diagnoses.
Enrichment analysis
An enrichment analysis was conducted using the top 100 metabolites identified by the sPLS model at gestational week 24 to investigate their association with any NDD and ADHD. Figure 5 illustrates the Sankey and bubble diagrams of enrichment of sub-pathways for key metabolites associated with any NDD. The analysis revealed several pathways significantly enriched in long-chain saturated fatty acid, fatty acid-dicarboxylate, medium-chain fatty acid, chemicals, lactoyl amino acid, and vitamin A metabolites. Notably, the long-chain saturated fatty acid pathway (fold enrichment = 5.65, Pfdr = 0.018) was among the most significantly enriched pathways for any NDD. For ADHD, the enriched pathways were similar to those for any NDD (Supplementary Fig. 10), with Vitamin A metabolism showing the highest fold of enrichment and lowest p-value after FDR correction.

Each left-side node represents a metabolite, and each right-side node represents a sub-pathway. All metabolites are uniquely assigned to one pathway. Node colors are arbitrarily assigned for visual clarity and do not reflect statistical significance or effect direction. All edges (links) have equal thickness and indicate membership relationships between metabolites and their respective pathways. Bubble plot displaying pathway-level enrichment statistics. Each point corresponds to one pathway. X-axis representing the enrichment magnitude of each pathway relative to metabolites. Y-axis: pathways with larger fold enrichment are plotted lower. Dot size: encodes p-value tier. One-sided P < 0.05 was considered significant, and raw P values were corrected for multiple testing using the FDR (false discovery rate) procedure across all tested sub-pathways. NDD neurodevelopmental disorder. Source data are provided as a Source data file.
Meditation analysis
We lastly performed mediation analyses to investigate whether quinolinate explained the association between NDD risk factors, such as maternal inflammatory proteins, unhealthy Western diet and maternal BMI, and NDD. Causal mediation analyses indicated that maternal inflammatory proteins (Supplementary Fig. 11), such as CCL3 (proportion mediated: 21.9%, P = 0.048), were mediated through maternal quinolinate levels during pregnancy. The inflammatory protein profile captured in PC1 of a PCA and individual proteins, including VEGFA, CD5, and IL-12B, showed non-significant results for mediation. Maternal BMI (proportion mediated: 17.6%, P = 0.042) and unhealthy Western diet (proportion mediated: 10.0%, P = 0.004) were also found to be mediated through maternal quinolinate levels.
Discussion
This study examines longitudinal mother-child pairs’ global metabolome profiles from pregnancy to middle childhood in relation to NDD risk at age 10 years. The study demonstrated that metabolic disturbances, especially during week 24 of pregnancy, are associated with an increased risk of offspring NDD, emphasizing the fetal environment’s pivotal role in shaping neurodevelopment. Further, sPLS-derived metabolome scores and enrichment analyses identified early life metabolic trajectories from pregnancy until age 18 months, with pathways including lactoyl amino acid, vitamin A metabolism, fatty acids, and xenobiotics playing a role in the risk of NDD. Finally, single metabolite analyses revealed three metabolites (quinolinate, ferulic acid 4-sulfate, and 8-methoxy kynurenate) at week 24 of pregnancy, child age 6 and 18 months that were consistently nominally associated with risk of NDD, whereof quinolinate levels at all timepoints until age 10 years showed nominal significant associations. Mediation analyses suggested that quinolinate mediates the association between maternal inflammatory proteins, an unhealthy diet, maternal BMI, and NDD. However, none of the metabolites remained significant after FDR correction, indicating that metabolic perturbations may be subtle. The modest AUC values from the sPLS model further suggest that factors beyond maternal metabolic profiles likely play a role in the neurodevelopment of children.
We investigated the predictive performance of sPLS models using maternal and child metabolome profiles at six timepoints in relation to the risk of any NDD at age 10. Of all timepoints, the gestational week 24 metabolome showed the highest AUC for predicting both any NDD and ADHD in the cross-validated sPLS models and nested CV models. This suggests that the maternal metabolic profile is associated with NDD risk in offspring, which aligns with previous studies emphasizing the importance of the maternal metabolic state in the prenatal period for fetal neurodevelopment5,24,25,34. Maternal metabolome alterations during pregnancy have been linked to ASD (Norwegian Autism Birth Cohort, N = 408)30, and disturbed lipid metabolism during late pregnancy and in placental serum profiles have been associated with non-typical disorders, which is related to ADHD (N = 141)35,36. Further, a previous study (N = 114) suggested that mid-pregnancy metabolic changes may be associated to ASD risk31, but hitherto our study is to explore longitudinal, global metabolic profiles from pregnancy through childhood in relation to multiple NDD outcomes in a large cohort.
Maternal metabolic disturbances may indirectly influence fetal neurodevelopment by modulating placental functions and nutrient transport37,38. Our results suggest that the metabolic profile during pregnancy offers insights into the intrauterine environment and fetal programming. However, modest AUC values of 0.6 indicate that factors beyond maternal metabolic profiles are involved in the development of NDD in the offspring. This warrants further research to validate and expand our findings and potentially incorporate environmental factors and possibly other omics data layers in a multi-omics integrative approach. The lower AUC values at subsequent timepoints during childhood compared to gestational week 24 suggest that the predictive power of metabolic profiles for NDD decreases, particularly after age 18 months, possibly due to the increasing influence of genetic and environmental factors2,39.
Using sPLS-derived metabolome–NDD risk scores based on maternal metabolome data at 24 weeks of pregnancy, we identified metabolic trajectories potentially associated with any NDD and ADHD prior to 18 months of age. These findings suggest that metabolic alterations in early life are crucial for NDD risk. Further, metabolome-NDD risk scores derived from maternal metabolome underscore the importance of prenatal metabolic regulation and the need for early NDD risk identification. Maternal metabolic perturbations during pregnancy may initiate a cascade of biological changes that persist into later life, contributing to NDD in offspring. Gestational week 24 represents a critical window in fetal brain development, occurring shortly after peak neurogenesis, when processes like neuronal proliferation, migration, early synapse formation and cortical layering are highly active and sensitive to maternal metabolic and inflammatory cues5,25. This can lead to long-term consequences for neurodevelopmental programming.
In contrast, postnatal periods reflect distinct and temporally specific neurodevelopmental processes. Between birth and 6 years, the brain undergoes rapid synaptogenesis, myelination, and cortical thickening, with gray matter volume peaking around age 6. From 6 to 10 years, these processes slow and give way to synaptic pruning and cortical refinement. Therefore, the same metabolite may interact with different molecular or cellular pathways depending on the developmental stage, potentially explaining the weaker associations observed at later time points. Incorporating a developmental perspective provides important biological context to our findings and highlights the significance of prenatal metabolic exposures—particularly during mid-gestation—as early-life determinants of NDD risk, as supported by recent literature outlining normative trajectories in brain growth and structure across developmental stages40,41.
Our pathway enrichment analysis utilized the top 100 significant metabolites associated with NDD from the sPLS model. Notably, the long-chain saturated fatty acids (LC-SFAs) were among the most significantly enriched for any NDD, and the vitamin A metabolism pathway was among the most significantly enriched for ADHD. The LC-SFAs pathway supports energy storage, cell membranes structure, and bioactive lipids synthesis42. Stearate (C18:0) and myristate (C14:0), key LC-SFAs, are common in the human diet43. The relationship between LC-SFAs and NDD appears to be less directly studied. However, in vitro studies have shown that LC-SFAs can induce adverse metabolic effects, such as lipotoxicity and apoptotic cell death44, as well as upregulate the expression of cyclooxygenase-2 (COX-2) via activation of toll-like receptor 4 (TLR4)45. TLR4 involved in the maturation of immune responsiveness in infants and adults and cortical development in vitro, suggesting its role in neurodevelopmental processes46,47. COX-2 has been identified as a potential neuroinflammatory marker, may also contribute to the NDD risk48. Studies have suggested that unhealthy dietary patterns high in saturated fat may be associated with ADHD symptoms49. Our finding that maternal Western dietary patterns during pregnancy are associated with increased NDD risk scores, along with the previously reported association with ADHD risk32, further strengthens the relevance of the LC-SFA pathway enrichment results. Metabolites, such as myristate (14:0), stearate (18:0), enriched in the LC-SFAs pathway, were positively associated with any NDD and ADHD risk in the sPLS model, suggesting their potential role in increasing risk.
Vitamin A is essential for brain health, and synaptic plasticity underlying learning and memory50. Vitamin A deficiency is linked to cognitive impairments and may contribute to psychiatric disorders like ADHD50,51,52. Maternal deficiency has also been associated with autistic-like behaviors via disrupted purine and tryptophan metabolism in rat model29. Beta-carotene diol and beta-cryptoxanthin, key carotenoids in vitamin A metabolism, are precursors of vitamin A53. Vitamin A levels have been associated with ADHD in children aged 6–952. In our model, both metabolites were negatively associated with any NDD and ADHD risk, consistent with previous studies54,55.
The fatty acid dicarboxylate pathway was also enriched for NDD risk in our results. Metabolites, including saturated long-chain dicarboxylic acids (e.g., dodecanedioate (C12-DC), sebacate (C10-DC), tetradecanedioate (C14-DC)), were associated with an increased NDD risk. Notably, dicarboxylic acid metabolism has also been linked to autism previously56.
In our single metabolite analysis, no associations remained significant after FDR correction. However, most of the nominally significant metabolites were also selected by the sPLS final models, underscoring their relevance. Among these, three metabolites, namely, quinolinate, ferulic acid 4-sulfate, and 8-methoxy kynurenate from gestational week 24–18 months of the child were consistently associated with at least one NDD outcome at age 10. Notably, quinolinate showed nominal significant associations with NDD risk across all timepoints. As a secondary exploration, preliminary findings of metabolic profile differences between ADHD—only cases and multiple NDD diagnoses suggest the possibility of divergent metabolic signatures between children with isolated versus comorbid neurodevelopmental conditions. However, these results require validation in larger cohorts. For autism outcome, previous studies have linked maternal metabolomic profiles to autism risk, though findings vary by metabolomics platforms, sample sizes, and timing of sample collection30,31. While most metabolites identified by Ritz et al. and Che et al. were not significant in our cohort, we observed a consistent association of pseudouridine with autism at 6 months, aligning with their cord blood findings and supporting its potential relevance30,31.
Quinolinate (quinolinic acid), a kynurenine pathway metabolite, processes tryptophan into nicotinamide adenine dinucleotide (NAD+) and other neuroactive metabolites57,58,59. It redirects tryptophan catabolism to replenish NAD+ in response to inflammation and infection59. While direct evidence linking elevated quinolinate level to NDD risk is limited, it is neurotoxic at high levels60. As an N-methyl-D-aspartate (NMDA) receptor agonist, quinolinate influence synaptic transmission and plasticity in the brain61,62. NMDA receptor dysfunction has been associated with ADHD, autism, and schizophrenia63,64,65, and overactivation can causes excitotoxicity, damaging nerve cells through excessive neurotransmitter stimulation66. Thus, maintaining balance among kynurenine metabolites is critical for brain function, and its disruptions is implicated in neuroinflammation, neurodegenerative and psychiatric disorders60,62,67,68,69,70. Our findings suggest a role for quinolinate in early brain development, through mechanisms behind remain unclear. 8-methoxy kynurenate, a kynurenic acid derivative, also participates in tryptophan and kynurenine metabolism, and may influence neurobiology and neuroprotection that linked to neuropsychiatric disorders71,72, though studies are limited.
In our study, quinolinate was found to be associated with maternal inflammation, Western dietary pattern, and pre-pregnancy BMI. We proposed that quinolinate mediates the association between maternal risk factors, such as maternal inflammatory proteins and BMI, and ADHD using mediation analysis. The effect of CCL3, previously linked to NDDs33, was significantly mediated by maternal quinolinate levels. VEGFA, CD5, IL-12B, and the inflammatory protein profile (PC1) showed non-significant mediation effects. Similarly, the effects of maternal BMI and unhealthy Western diet patterns were mediated through quinolinate. Metabolic disturbances during pregnancy may increase inflammation, oxidative stress, and nutrient dysregulation, adversely affecting fetal brain development37,73. These findings suggest that quinolinate may serve as a biochemical link between maternal inflammation, metabolism, and fetal neurodevelopment, highlighting its potential as a potential biomarker or intervention target. It would be valuable in future work to explore whether other inflammatory proteins act independently or via pathways such as the kynurenine pathway.
Ferulic acid 4-sulfate, a phenolic compound involved in the gut-brain axis, may facilitate communication between gut microbiota and the brain74. As a sulfate form of ferulic acid, it possesses antioxidant, anti-inflammation, and potential neuroprotective effects75,76,77. Although specific research on the sulfate form is limited, its systemic effects suggest a role in reducing neuroinflammation and oxidative damage, potentially lowering NDD risk75. In our study, higher levels of ferulic acid 4-sulfate during pregnancy and in infants were associated with reduced NDD risk at age 10, supporting its potential protective role.
Environmental factors influencing neurodevelopmental outcomes start before conception and persist throughout life78,79. Xenobiotics, such as erythritol, which reflect environmental exposures during pregnancy, were associated with NDD risk in both single-metabolite and sPLS models, suggesting that the prenatal exposome may play a role in neurodevelopmental outcomes and warrants further investigation. Given that obesity, gestational diabetes, dietary patterns, smoking, pre-eclampsia, and stress during pregnancy contribute to the NDD risk32,79,80. Our analyses were adjusted for most of these factors. The results showed that maternal metabolome-NDD risk scores were associated with gestational diabetes, maternal BMI, maternal diet, and family income, aligning with previous studies. We also found that parental educational attainment and antibiotic use during pregnancy were associated with the metabolome-NDD risk scores, highlighting areas for further research. Importantly, our results remained largely unchanged after confounder adjustments, such as our micronutrient RCTs, prenatal exposures, and breastfeeding status.
The study benefits from a large participant pool from the general Danish population, providing broad representation and applicability. Comprehensive data collection over 10 years, including detailed metabolic profiles across multiple time points and a high retention rate within the COPSAC cohort, enables robust adjustment for potential confounders and highlights the role of early-life metabolomics, particularly during pregnancy in NDD inception. The longitudinal metabolomic profiles with a large number of common metabolites present at all timepoints is the biggest strength and novelty of the current study design. Another strength is the detailed neuropsychiatric assessment, which includes interviews, performance-based measures, and questionnaires. However, several limitations should be considered. The cohort’s sample size, predominantly white demographic, higher average educational attainment, and missing data may limit the generalizability of the findings. Additionally, the lack of data on parental mental health is significant, given the heritability of NDDs. Polygenetic risk scores explain only a small portion of the variance accounted for by family history of mental disorders. Furthermore, the sensitivity analysis used unbiased internal validation, which, although less robust than external replication, was the best option available for this study.
This longitudinal study analyzing metabolic profiles from pregnancy to middle childhood indicates that metabolic disturbances in early life may increase the NDD risk of offspring. We observed metabolic trajectories during pregnancy and early life up to 18 months that showed potential associations with NDD risk. Additionally, we identified three specific metabolites that were consistently associated with NDD risk with nominal significance, with quinolinate showing a consistent nominal association across all observed timepoints. Quinolinate may mediate the relationship between maternal risk factors, such as inflammatory proteins, BMI and diet, and the NDD risk. These findings highlight the role of the fetal environment in shaping neurodevelopment. However, no metabolites passed FDR correction, and modest AUCs from sPLS suggest that other factors beyond maternal metabolism contribute to NDD risk. Further research is needed to identify molecular pathways for targeted prevention and risk identification to improve neurodevelopment. Investigating the underlying mechanisms are warranted to fully elucidate their role in dysregulation and potential as therapeutic targets.
Methods
Study population
The COpenhagen Prospective Study on Neuro-PSYCHiatric Development (COPSYCH) enrolled 10-year-old children from the population-based COPSAC2010 cohort81,82, including 700 mother-child pairs. The children were assessed clinically for neuropsychiatric outcomes from January 2019 to December 2021. Exclusion criteria for the present study included offspring with a birth weight <1500 grams or a gestational age below 28 weeks, as well as twin B in pairs due to lack of independence. The biological sex was defined by the sex assigned at birth. The study was conducted in accordance with the Declaration of Helsinki and approved by the Danish Ethics Committee (H-B-2008-093) and the Danish Data Protection Agency (2015-41-3696). Written informed consent was obtained from all participants prior to any study-related procedures. Study design details of the COPSAC2010 cohort are available in Supplementary Methods (Supplementary information).
Psychopathological outcomes of NDD
We focused on the most prevalent categorical NDDs outcomes in this cohort. Primary outcomes were any NDD, ADHD, and autism. The prevalence of all research diagnoses in this study is shown in Supplementary Table 1. Research diagnoses followed the International Classification of Disorders 10th revision of mental and behavioral disorders (ICD-10) criteria83,84, based on clinical information from the Kiddie Schedule for Affective Disorders and Schizophrenia Present and Lifetime version (K-SADS-PL) interviews and additional clinical information85, as detailed in the COPSYCH study protocol82.
Secondary outcomes were dimensional psychopathology using K-SADS-PL. Dimensional psychopathology of NDDs includes autistic and ADHD symptoms component scores, the ADHD rating scale (ADHD-RS) and the social responsiveness scale 2 (SRS-2). Parents completed the ADHD-RS and the SRS-2 questionnaires to assess ADHD symptoms and autistic traits86,87,88 (detailed Supplementary Methods in Supplementary information).
Covariates
Thirteen covariates were included in the analyses using maternal week 24 metabolome: pre-pregnancy maternal body mass index (BMI), maternal age, smoking, antibiotic use, pre-eclampsia, maternal diabetes, and alcohol intake during pregnancy, household income level, maternal and paternal educational attainment at birth, gestational age, child sex, and birth weight. For analyses using the maternal one week postpartum metabolome, we included: pre-pregnancy BMI, maternal age, maternal diabetes, mother smoking during pregnancy, household income level, maternal and paternal educational attainment, gestational age, child sex, and birth mode (cesarean or vaginal birth). Sex, maternal and paternal educational attainment, household income (at birth), and WHO BMI Z-score of children were included as covariates in analyses using childhood metabolomes. We additionally adjusted for the RCTs of fish oil and vitamin D interventions at each time point89,90.
Metabolomic and proteomics profiling
Metabolomic profiling was run on blood plasma samples from pregnancy week 24 and one week postpartum of mothers, and at 6 months, 18 months, 6 years, and 10 years for children. Metabolome data from one week postpartum were analyzed in this study under the hypothesis that they may reflect both the prenatal and postnatal environment.
Blood samples (n = 3212) were collected in EDTA tubes at the research clinic, centrifuged for 10 min at 4000 rpm, and plasma was stored at −80 °C until analysis. Untargeted metabolomics of plasma samples was performed by Metabolon, Inc., in Durham, NC, USA, using their HD4 platform. Sample preparation used the automated Hamilton MicroLab STAR system. Recovery standards were added for quality control (QC), and metabolites were extracted with methanol following vigorous shaking (2 min, GenoGrinder 2000) and centrifugation to precipitate proteins. Extracts were divided into four aliquots and analysed across four LC-MS/MS platforms. After drying with TurboVap and storage under nitrogen overnight, samples were analysed using a Waters ACQUITY UPLC system coupled to a Thermo Fisher Q-Exactive Quadrupole-Orbitrap mass spectrometer with HESI-II source. Four chromatographic methods were applied: two reverse-phase UPLC-ESI(+) for hydrophilic and hydrophobic compounds, one reverse-phase UPLC-ESI(−), and one HILIC UPLC-ESI(−). Data were acquired using alternating full-scan MS and data-dependent MSn with dynamic exclusion (70–1000 m/z). Metabolites were identified and semi-quantified using the area under the curve following established QC procedures. Compounds were identified by comparison to library entries of purified standards or recurrent unknown entities.
All samples were normalized using anchor samples with metabolites present in every data set. Only bridging metabolites were included in this study, resulting in a total of 683 metabolites common for all timepoints. All Batch-norm datasets were normalized to the reference set using median ratios of non-missing anchor sample pairs (≥6 per metabolite). Metabolites with >30% missingness were excluded, missing values were imputed with the minimum observed value, and data were log-transformed before analysis. A comprehensive description of the metabolomics protocol was outlined previously91, and more details of data extraction, data preprocessing, and annotation are described in the Supplementary Methods. Xenobiotics, as exogenous compounds distinct from endogenous metabolites, were also included in the metabolomics dataset due to their potential relevance to metabolic function and early-life exposures.
Plasma proteins at week 24 of pregnancy were measured using the Olink Target 96 inflammation panels (Olink). Blood plasma samples were thawed, randomized across nine 96-well plates, refrozen, and transferred to Olink on dry ice before being screened with the Olink target 96 Inflammation assay. A total of 92 inflammatory proteins were quantified from our samples. Each sample was tested once, and all samples were fully randomized, without stratification, across nine plates. All samples were analyzed in nine different plates. The results were presented as log2-transformed normalized protein expression (NPX) values. Four internal controls were added to each sample to monitor the quality of assay performance, as well as the quality of individual samples. All plates and samples passed QC thresholds, defined as an internal control SD of <0.2 NPX from the median between plates and <0.3 NPX between samples within plates, and the samples with QC warnings were excluded from the analyses33. A PCA was performed on the 92 proteins, with PC1 explaining 30.7% of the variance. Inflammatory protein PC1 and key proteins associated with NDD risk, identified in our previous study33—VEGFA, CD5, IL-12B, CCL3, FGF-23, and MCP-1—were included in the mediation analysis in this study.
Statistics
All statistical analyses were conducted using R version 4.2.1. R package “caret” and “mixOmics (6.20.0)” were used in sPLS92,93. We applied a FDR of 5% to account for multiple testing. Two-sided P < 0.05 was considered significant. Children without ADHD, autism or any NDD were used as controls, depending on the specific outcome being modeled. ADHD, autism, and any NDD outcomes were examined separately. The number of cases for each diagnosis is shown in Fig. 1b.
In this study, we applied univariable (logistic and linear regression) and multivariable (sPLS) analyses. Univariable models assessed associations between each metabolite and any NDD, ADHD, and autism. Autism was analyzed only in univariable models because the small sample size (n = 16) precluded reliable multivariable sPLS modeling and subsequent pathway-level analyses. sPLS models focused on ADHD and NDD outcomes, as the low prevalence of autism prevented robust 10-fold cross-validation and nested validation essential for model reliability. The emphasis on ADHD and NDD outcomes was guided by statistical feasibility rather than biological rationale.
First, the sPLS model with 10 times repeated and 10 times cross-validations were performed to predict NDD outcomes from the metabolome data at the six timepoints. For each timepoint, the metabolites loading scores were shown when the area under the receiver operating characteristic curve (AUC) was higher than 0.55. We aimed to identify the timepoint that best predicted NDD and determine the most strongly associated metabolites. This derived a metabolome-NDD risk score for each individual based on the best performed model with highest median AUC and lowest variance across repeated cross-validation rounds (Supplementary Methods in Supplementary Information). Due to the limited number of autism cases (N = 16), we were unable to perform proper 10-fold cross-validation with 10 repetitions and nested cross-validation. Therefore, only ADHD and any NDD outcomes were included in the sPLS modeling and the following analyses involving the sPLS model.
Second, logistic and linear regression analyses were performed to investigate the association between each single metabolite and the NDD outcomes. These analyses were performed unadjusted and adjusted for covariates. As a further exploration of whether single versus multiple NDD diagnoses are associated with metabolomic profiles, we extracted all NDD cases and categorized them into two groups: children diagnosed with only ADHD (n = 48) and those with multiple NDD diagnoses (n = 38). In the logistic regression analysis, individuals with only ADHD were labeled as 0, and those with multiple NDD diagnoses were labeled as 1.
Third, pathway enrichment analyses were conducted by initially converting a list of the top 100 significant metabolites associated with NDD in the sPLS model into a data frame. The list of the top 100 metabolites was then merged with the corresponding counts of metabolites associated with each pathway. Enrichment ratios were calculated as the ratio of selected to total metabolites. Statistical significance was assessed using the hypergeometric test, with p-values adjusted for multiple testing using 5% FDR to identify significantly enriched metabolic pathways among the selected metabolites.
We performed mediation analysis to assess whether the effect of NDD-related risk factors, such as inflammatory proteins and maternal BMI, on ADHD is mediated by the highlighted metabolite. The mediate function from the “mediation (4.5.0)” R package was applied with 1000 bootstrap simulations to estimate the average causal mediation effect, the average direct effect, and the proportion of the total effect mediated. Statistical significance was assessed using bootstrap-derived p-values, and the mediation pathways were visualized with annotated effect estimates, including the proportion mediated and corresponding p-values. We focused on six inflammatory proteins—VEGFA, CCL3, CD5, IL-12B, FGF-23, and MCP-1, together with a maternal protein inflammatory profile PC1 that we previously identified as being associated with NDD risk33. Given the limited number of mediation tests and the hypothesis-driven nature of the analyses, we did not apply FDR correction here.
Sensitivity analysis of sPLS
To address potential overfitting from traditional tenfold cross-validation for sPLS, we additionally conducted a sensitivity analysis using a nested five-fold cross-validation approach, as outlined in our previous study (detailed in Supplementary Methods in Supplementary Information)94.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Source data are provided with this paper. Participant-level personally identifiable data are protected under the Danish data protection act and European Regulation 2016/679 of the European Parliament and of the Council (GDPR) that prohibit distribution even in pseudo-anonymized form. However, participant-level data can be made available under a data transfer agreement as part of a collaboration effort with COPSAC (chawes@copsac.com). Source data are provided with this paper.
Code availability
The custom code employed in this research is freely accessible to the public for transparency and reproducibility purposes. Related codes can be found at https:// github already.https://github.com/tiwa1125/Longitudinal_metabolome and Zenodo (https://doi.org/10.5281/zenodo.17340755).
References
Thapar, A., Cooper, M. & Rutter, M. Neurodevelopmental disorders. Lancet Psychiatry 4, 339–346 (2017).
Parenti, I., Rabaneda, L. G., Schoen, H. & Novarino, G. Neurodevelopmental disorders: from genetics to functional pathways. Trends Neurosci. 43, 608–621 (2020).
Verhoef, E. et al. Discordant associations of educational attainment with ASD and ADHD implicate a polygenic form of pleiotropy. Nat. Commun. 12, 6534 (2021).
Antshel, K. M., Zhang-James, Y., Wagner, K. E., Ledesma, A. & Faraone, S. V. An update on the comorbidity of ADHD and ASD: a focus on clinical management. Expert Rev. Neurother. 16, 279–293 (2016).
Bale, T. L. et al. Early life programming and neurodevelopmental disorders. Biol. Psychiatry 68, 314–319 (2010).
Sayal, K., Prasad, V., Daley, D., Ford, T. & Coghill, D. ADHD in children and young people: prevalence, care pathways, and service provision. Lancet Psychiatry 5, 175–186 (2018).
Tick, B., Bolton, P., Happé, F., Rutter, M. & Rijsdijk, F. Heritability of autism spectrum disorders: a meta-analysis of twin studies. J. Child Psychol. Psychiatry 57, 585–595 (2016).
Wang, K., Gaitsch, H., Poon, H., Cox, N. J. & Rzhetsky, A. Classification of common human diseases derived from shared genetic and environmental determinants. Nat. Genet 49, 1319–1325 (2017).
Sandin, S. et al. The heritability of autism spectrum disorder. JAMA 318, 1182–1184 (2017).
Faraone, S. V. & Larsson, H. Genetics of attention deficit hyperactivity disorder. Mol. Psychiatry 24, 562–575 (2019).
Larsson, H., Chang, Z., D’Onofrio, B. M. & Lichtenstein, P. The heritability of clinically diagnosed attention deficit hyperactivity disorder across the lifespan. Psychol. Med. 44, 2223–2229 (2014).
Needham, B. D. et al. Plasma and fecal metabolite profiles in autism spectrum disorder. Biol. Psychiatry 89, 451–462 (2021).
Wang, L.-J. et al. Novel plasma metabolite markers of attention-deficit/hyperactivity disorder identified using high-performance chemical isotope labelling-based liquid chromatography-mass spectrometry. World J. Biol. Psychiatry 22, 139–148 (2021).
Adams, J. B. et al. Nutritional and metabolic status of children with autism vs. neurotypical children, and the association with autism severity. Nutr. Metab. 8, 34 (2011).
Brunkhorst-Kanaan, N. et al. Sphingolipid and endocannabinoid profiles in adult attention deficit hyperactivity disorder. Biomedicines 9, 1173 (2021).
Ma, L., Chen, Y.-H., Chen, H., Liu, Y.-Y. & Wang, Y.-X. The function of hypothalamus–pituitary–adrenal axis in children with ADHD. Brain Res. 1368, 159–162 (2011).
Spahis, S. et al. Lipid profile, fatty acid composition and pro- and anti-oxidant status in pediatric patients with attention-deficit/hyperactivity disorder. Prostaglandins Leukot. Essent. Fat. Acids 79, 47–53 (2008).
Murakami, Y., Imamura, Y., Saito, K., Sakai, D. & Motoyama, J. Altered kynurenine pathway metabolites in a mouse model of human attention-deficit hyperactivity/autism spectrum disorders: a potential new biological diagnostic marker. Sci. Rep. 9, 13182 (2019).
Ahrens, A. P. et al. Infant microbes and metabolites point to childhood neurodevelopmental disorders. Cell 187, 1853–1873 (2024).
Yap, C. X. et al. Interactions between the lipidome and genetic and environmental factors in autism. Nat. Med. 29, 936–949 (2023).
Engelhard, M. et al. Patterns of health services use before age 1 in children later diagnosed with ADHD. J. Atten. Disord. 25, 1639 (2021).
Polańska, K., Jurewicz, J. & Hanke, W. Exposure to environmental and lifestyle factors and attention-deficit/hyperactivity disorder in children—a review of epidemiological studies. Int. J. Occup. Med. Environ. Health 25, 330–55 (2012).
Yap, C. X. et al. Autism-related dietary preferences mediate autism-gut microbiome associations. Cell 184, 5916–5931 (2021).
Buss, C., Entringer, S. & Wadhwa, P. D. Fetal programming of brain development: intrauterine stress and susceptibility to psychopathology. Sci. Signal. 5, 1–7 (2012).
Graham, A. M., Marr, M., Buss, C., Sullivan, E. L. & Fair, D. A. Understanding vulnerability and adaptation in early brain development using network neuroscience. Trends Neurosci. 44, 276–288 (2021).
Vasung, L. et al. Development of axonal pathways in the human fetal fronto-limbic brain: histochemical characterization and diffusion tensor imaging. J. Anat. 217, 400–417 (2010).
Tau, G. Z. & Peterson, B. S. Normal development of brain circuits. Neuropsychopharmacol 35, 147–168 (2010).
Bruce, M. R. et al. Altered behavior, brain structure, and neurometabolites in a rat model of autism-specific maternal autoantibody exposure. Mol. Psychiatry 28, 2136–2147 (2023).
Cheng, B. et al. Vitamin A deficiency from maternal gestation may contribute to autistic-like behaviors and gastrointestinal dysfunction in rats through the disrupted purine and tryptophan metabolism. Behav. Brain Res. 452, 114520 (2023).
Che, X. et al. Metabolomic analysis of maternal mid-gestation plasma and cord blood in autism spectrum disorders. Mol. Psychiatry 28, 2355–2369 (2023).
Ritz, B. et al. Untargeted metabolomics screen of mid-pregnancy maternal serum and autism in offspring. Autism Res. 13, 1258–1269 (2020).
Horner, D. et al. A western dietary pattern during pregnancy is associated with neurodevelopmental disorders in childhood and adolescence. Nat. Metab. 7, 586–601 (2025).
Wang, T. et al. Maternal inflammatory proteins in pregnancy and neurodevelopmental disorders at age 10 years. JAMA Psychiatry 82, 514–525 (2025).
Mitchell, A., Dunn, G. A. & Sullivan, E. L. The influence of maternal metabolic state and nutrition on offspring neurobehavioral development: a focus on preclinical models. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 7, 450–460 (2022).
Parenti, M. et al. Neurodevelopment and metabolism in the maternal-placental-fetal unit. JAMA Netw. Open 7, e2413399 (2024).
Hertz-Picciotto, I. et al. A prospective study of environmental exposures and early biomarkers in autism spectrum disorder: design, protocols, and preliminary data from the MARBLES study. Environ. Health Perspect. 126, 117004 (2018).
Bell, A. W. & Ehrhardt, R. A. Regulation of placental nutrient transport and implications for fetal growth. Nutr. Res. Rev. 15, 211–230 (2002).
Gaccioli, F., Lager, S., Powell, T. L. & Jansson, T. Placental transport in response to altered maternal nutrition. J. Dev. Orig. Health Dis. 4, 101–115 (2013).
Manchia, M. & Fanos, V. Targeting aggression in severe mental illness: the predictive role of genetic, epigenetic, and metabolomic markers. Prog. Neuro Psychopharmacol. Biol. Psychiatry 77, 32–41 (2017).
Bethlehem, R.aI. et al. Brain charts for the human lifespan. Nature 604, 525–533 (2022).
Johnson, M. H. Functional brain development in humans. Nat. Rev. Neurosci. 2, 475–483 (2001).
Cockcroft, S. Mammalian lipids: structure, synthesis and function. Essays Biochem. 65, 813–845 (2021).
Shramko, V. S., Polonskaya, Y. V., Kashtanova, E. V., Stakhneva, E. M. & Ragino, Y. I. The short overview on the relevance of fatty acids for human cardiovascular disorders. Biomolecules 10, 1127 (2020).
Krogmann, A. et al. Inflammatory response of human coronary artery endothelial cells to saturated long-chain fatty acids. Microvasc. Res. 81, 52–59 (2011).
Lee, J. Y., Sohn, K. H., Rhee, S. H. & Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through toll-like receptor 4. J. Biol. Chem. 276, 16683–16689 (2001).
Liao, S.-L., Yeh, K.-W., Lai, S.-H., Lee, W.-I. & Huang, J.-L. Maturation of toll-like receptor 1–4 responsiveness during early life. Early Hum. Dev. 89, 473–478 (2013).
Zurolo, E. et al. Activation of toll-like receptor, RAGE and HMGB1 signalling in malformations of cortical development. Brain 134, 1015–1032 (2011).
El-Ansary, A. et al. Alpha-synuclein, cyclooxygenase-2 and prostaglandins-EP2 receptors as neuroinflammatory biomarkers of autism spectrum disorders: use of combined ROC curves to increase their diagnostic values. Lipids Health Dis. 20, 155 (2021).
Hernández, R. & Margarita, A. Diet and sleep in children and adolescents with Attention-Deficit Hyperactivity Disorder. Phd thesis, Universitat de Barcelona (2017).
Wołoszynowska-Fraser, M. U., Kouchmeshky, A. & McCaffery, P. Vitamin A and retinoic acid in cognition and cognitive disease. Annu. Rev. Nutr. 40, 247–272 (2020).
Ramakrishna, T. Vitamins and brain development. Physiol. Res. 48, 175–187 (1999).
Li, H.-H. et al. Serum levels of vitamin A and vitamin D and their association with symptoms in children with attention deficit hyperactivity disorder. Front. Psychiatry 11, 599958 (2020).
Gürbüz, M. & Aktaç, Ş Understanding the role of vitamin A and its precursors in the immune system. Nutr. Clin. Métab. 36, 89–98 (2022).
Moeini, F., Mostaghimi, M., Honarvar, M. R. & Sharifi, A. Comparison of dietary intake of vitamin A in children with autism spectrum disorders with healthy children in Gorgan city in 2021: a case-control study. Pajouhan Sci. J. 21, 97–103 (2023).
Miodownik, C. & Lerner, V. The neuroprotective efficacy of vitamins. in Brain Protection in Schizophrenia, Mood and Cognitive Disorders (ed Ritsner, M. S.) 505–553 https://doi.org/10.1007/978-90-481-8553-5_17 (Springer Nature, 2010).
El-Ansary, A. et al. The role of lipidomics in autism spectrum disorder. Mol. Diagn. Ther. 24, 31–48 (2020).
Stone, T. W. & Darlington, L. G. Endogenous kynurenines as targets for drug discovery and development. Nat. Rev. Drug Discov. 1, 609–620 (2002).
Ruddick, J. P. et al. Tryptophan metabolism in the central nervous system: medical implications. Expert Rev. Mol. Med. 8, 1–27 (2006).
Moffett, J. R. et al. Quinolinate as a marker for kynurenine metabolite formation and the unresolved question of NAD+ synthesis during inflammation and infection. Front. Immunol. 11, 31 (2020).
Stone, T. W. & Darlington, L. G. The kynurenine pathway as a therapeutic target in cognitive and neurodegenerative disorders. Br. J. Pharmacol. 169, 1211–1227 (2013).
Heyes, M. P. et al. Quinolinic acid and kynurenine pathway metabolism in inflammatory and non-inflammatory neurological disease. Brain 115, 1249–1273 (1992).
Lima, S., Gawandi, V., Momany, C. & Phillips, R. S. Crystal structure of the Homo sapiens kynureninase-2-amino-3-hydroxyhippuric acid inhibitor complex. FASEB J. 20, A895–A895 (2006).
Chang, J. P.-C., Lane, H.-Y. & Tsai, G. E. Attention deficit hyperactivity disorder and N-methyl-D-aspartate (NMDA) dysregulation. Curr. Pharm. Des. 20, 5180–5185 (2014).
Dessus-Gilbert, M.-L. et al. NMDA antagonist agents for the treatment of symptoms in autism spectrum disorder: a systematic review and meta-analysis. Front. Pharmacol. 15, 1395867 (2024).
Sandström, K. O. et al. Add-on MEmaNtine to dopamine antagonism to improve negative symptoms at first psychosis-the AMEND trial protocol. Front. Psychiatry 13, 889572 (2022).
Stone, T. W. Neuropharmacology of quinolinic and kynurenic acids. Pharmacol. Rev. 45, 309–379.
Sapko, M. T. et al. Endogenous kynurenate controls the vulnerability of striatal neurons to quinolinate: Implications for Huntington’s disease. Exp. Neurol. 197, 31–40 (2006).
Ting, K. K., Brew, B. J. & Guillemin, G. J. Effect of quinolinic acid on human astrocytes morphology and functions: implications in Alzheimer’s disease. J. Neuroinflammation 6, 36 (2009).
Hestad, K., Alexander, J., Rootwelt, H. & Aaseth, J. O. The role of tryptophan dysmetabolism and quinolinic acid in depressive and neurodegenerative diseases. Biomolecules 12, 998 (2022).
Guillemin, G. J. Quinolinic acid: neurotoxicity. FEBS J. 279, 1355–1355 (2012).
Muneer, A. Kynurenine pathway of tryptophan metabolism in neuropsychiatric disorders: pathophysiologic and therapeutic considerations. Clin. Psychopharmacol. Neurosci. 18, 507–526 (2020).
Schwarcz, R. & Stone, T. W. The kynurenine pathway and the brain: challenges, controversies and promises. Neuropharmacology 112, 237–247 (2017).
Bilbo, S. D. & Schwarz, J. M. The immune system and developmental programming of brain and behavior. Front. Neuroendocrinol. 33, 267–286 (2012).
Matsumura, Y., Kitabatake, M., Kayano, S. & Ito, T. Dietary phenolic compounds: their health benefits and association with the gut microbiota. Antioxidants 12, 880 (2023).
Thapliyal, S. et al. A review on potential footprints of ferulic acid for treatment of neurological disorders. Neurochem. Res. 46, 1043–1057 (2021).
Di Giacomo, S. et al. Recent advances in the neuroprotective properties of ferulic acid in Alzheimer’s disease: a narrative review. Nutrients 14, 3709 (2022).
Zduńska, K., Dana, A., Kolodziejczak, A. & Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Skin Pharmacol. Physiol. 31, 332–336 (2018).
Bilbo, S. D., Block, C. L., Bolton, J. L., Hanamsagar, R. & Tran, P. K. Beyond infection-maternal immune activation by environmental factors, microglial development, and relevance for autism spectrum disorders. Exp. Neurol. 299, 241–251 (2018).
Carlsson, T., Molander, F., Taylor, M. J., Jonsson, U. & Bölte, S. Early environmental risk factors for neurodevelopmental disorders–a systematic review of twin and sibling studies. Dev. Psychopathol. 33, 1448–1495 (2021).
Kim, B. ora et al. Prenatal exposure to paternal smoking and likelihood for autism spectrum disorder. Autism 25, 1946–1959 (2021).
Bisgaard, H. et al. Deep phenotyping of the unselected COPSAC 2010 birth cohort study. Clin. Exp. Allergy 43, 1384–1394 (2013).
Mohammadzadeh, P. et al. Effects of prenatal nutrient supplementation and early life exposures on neurodevelopment at age 10: a randomised controlled trial-the COPSYCH study protocol. BMJ Open 12, e047706 (2022).
Bell, C. C. DSM-IV: diagnostic and statistical manual of mental disorders. JAMA 272, 828–829 (1994).
World Health Organization. The ICD-10 Classification of Mental and Behavioural Disorders: Clinical Descriptions and Diagnostic Guidelines. (2009).
Kaufman, J. & Schweder, A. E. The Schedule for Affective Disorders and Schizophrenia for School-Age Children: Present and Lifetime Version (K-SADS-PL) (eds. Hilsenroth, M. J. & Segal, D. L.) 247–255 (John Wiley & Sons, Inc., 2004).
DuPaul, G. J., Power, T. J., Anastopoulos, A. D. & Reid, R. ADHD Rating Scale—IV: Checklists, Norms, and Clinical Interpretation Vol. 79, (The Guilford Press, 1998).
Makransky, G. & Bilenberg, N. Psychometric properties of the parent and teacher ADHD rating scale (ADHD-RS): measurement invariance across gender, age, and informant. Assessment 21, 694–705 (2014).
Bruni, T. P. Test review: social responsiveness scale–second edition (SRS-2). J. Psychoeduc. Assess. 32, 365–369 (2014).
Bisgaard, H. et al. Fish oil–derived fatty acids in pregnancy and wheeze and asthma in offspring. N. Engl. J. Med. 375, 2530–2539 (2016).
Chawes, B. L. et al. Effect of vitamin D3 supplementation during pregnancy on risk of persistent wheeze in the offspring: a randomized clinical trial. JAMA 315, 353–361 (2016).
Evans, A. M., DeHaven, C. D., Barrett, T., Mitchell, M. & Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 81, 6656–6667 (2009).
Rohart, F., Gautier, B., Singh, A. & Cao, K.-A. L. mixOmics: an R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 13, e1005752 (2017).
Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 28, 1–26 (2008).
Leal Rodríguez, C. et al. The infant gut virome is associated with preschool asthma risk independently of bacteria. Nat. Med. 30, 138–148 (2023).
Acknowledgements
We thank the children and families of the COPSAC2010 cohort study for all their support and commitment. We acknowledge and appreciate the efforts of the COPSAC research team. All funding received by COPSAC is listed on www.copsac.com. T.W., M.A., and B.C. are supported by the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement no. 946228). The COPSYCH study was funded by The Lundbeck Foundation (grant no R269-2017-5), and The Capital Region Research Foundation has provided core support to the COPSAC research Center.
Author information
Authors and Affiliations
Contributions
T.W. performed all analyses and has written the first draft of the manuscript, under supervision from B.E. and B.C. T.W. and B.C. formulated the research question, concept and design. P.M. and J.B.R. were the primary clinical investigators in the COPSYCH 10-year follow-up visit and substantial contributors to the data collection. J.R.M., R.V., N.B., L.C., M.H., M.A., J.B.R., P.M., M.W., P.E., M.A.R., J.T., K.B., B.H., and B.C. rigorously reviewed and revised the manuscript, providing substantial intellectual input and contributing to the analysis plan and interpretation of results. Approval of the final manuscript, ensuring its accuracy and integrity, was unanimous among all authors. The corresponding author had full access to the data and had final responsibility for the decision to submit for publication. No honorarium, grant, or other form of payment was given to any of the authors to produce this manuscript.
Corresponding authors
Ethics declarations
Competing interests
B.H.E. is part of the Advisory Board of Boehringer Ingelheim, Lundbeck Pharma A/S; and has received lecture fees from Boehringer Ingelheim, Otsuka Pharma Scandinavia AB, and Lundbeck Pharma A/S. J.T. has received a lecture fee from AstraZeneca. The remaining authors declare no conflicts of interest, whether potential, perceived, or actual, pertaining to the content of this manuscript.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, T., Jepsen, J.R.M., Vinding, R. et al. Longitudinal metabolome profiling from pregnancy through childhood and risk of neurodevelopmental disorders at age 10. Nat Commun 17, 1369 (2026). https://doi.org/10.1038/s41467-025-68115-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-68115-3
