
Abstract
The disruption of intestinal homeostasis represents a hallmark of different diseases. Here, we report a montmorillonite-based oral fermentation system (MOFS) that achieves long-lasting in-situ preparation of beneficial bioproducts for intestinal homeostasis restoration. MOFS is fabricated by loading metabolic substrates in the lamellar structure of montmorillonite via cation replacement and adsorbing the substrate-related bacteria through electrostatic interactions. The water-impermeable property and mucoadhesive ability of montmorillonite endow MOFS with high oral availability and extended retention in the gut. The continuous release of loaded substrates and the formation of biofilms by carried bacteria enable long-term yet efficient synthesis of beneficial metabolites in the intestine, which comprehensively improve gut microbiota composition, suppress intestinal inflammation, and increase the integrity of the gut barrier. In murine models of imbalanced intestinal homeostasis, MOFS demonstrates superior efficacies by alleviating the associated symptoms, even in comparison to clinical therapeutics.
Similar content being viewed by others

Introduction
Intestinal homeostasis, or the healthy and balanced state of the intestine, is established by intricate interactions between the intestinal barrier, the gut microbiota, and the host immune system1. Under normal physiological conditions, the intestinal barrier ensures the selective permeability of dietary nutrients and the physical separation of the gut microbiota from the host immune system1. Meanwhile, the gut microbiota actively shapes the host immune system and modulates intestinal barrier functions2,3. For instance, the gut symbiont Clostridium sporogenes (C. sporogenes) metabolizes tryptophan to indole and its derivatives, which have been reported to promote the expression of tight junction proteins to enhance intestinal barrier functions and stimulate the host immune system to play anti-inflammatory effects4. In addition, the host immune system reinforces intestinal barrier functions by priming protective immune responses against pathogens and maintaining tolerance to commensal microbiota and food antigens5,6. Usually, the breakdown of intestinal homeostasis may occur upon subjecting to external attacks7,8. As a hallmark of multiple diseases, this often causes the pathogenesis of inflammatory bowel diseases (IBD), acquired immunodeficiency syndrome, viral hepatitis, kidney injury, diabetes, and even cancers1,9. Therefore, strategies capable of restoring intestinal homeostasis hold great significance for disease treatment.
Conventional approaches to protect and restore intestinal homeostasis mainly focus on the intervention of the gut microbiota, the intestinal barrier, or the host immune system by small molecular therapeutics or macromolecular biologicals10,11. For example, a domain-binding small molecule divertin has been documented to reverse the intestinal barrier by blocking myosin light chain kinase recruitment12. Moreover, biopharmaceuticals, such as Janus kinase inhibitors and ustekinumab, have been applied clinically to restore intestinal immune homeostasis13,14. Unfortunately, the application of these oral therapeutics always faces a short half-life due to the instability in the intraluminal environments of the gastrointestinal tract. To overcome these challenges, a variety of advanced drug modification methods and delivery systems have been developed during the past few years15,16,17. For example, sodium butyrate formulated with enteric coating or encapsulation has been prepared to avoid the absorption of butyrate in the stomach, while increasing its concentration in the colon, where it plays a therapeutic effect by recovering microbial balance18. In addition, hydrophilic polysaccharide inulin has been conjugated with hydrophobic compound ursodeoxycholic acid to self-assemble into oral nanoscale devices for enhancing T-cell immunity modulation19. Despite these improvements, previous therapeutics have an overall low oral bioavailability caused by limited gut retention. More importantly, they are difficult to comprehensively restore intestinal homeostasis, as which highly depends on the multiple interactions among the gut microbiota, the intestinal barrier, and the host immune system.
Here, we present a montmorillonite-based oral fermentation system (MOFS) that provides durable in-situ synthesis of beneficial bioproducts for intestinal homeostasis restoration (Fig. 1a). MOFS is constructed by carrying metabolic substrates in the interlayer of montmorillonite (MMT) through cation replacement and then loading the substrate-associated bacteria via electrostatic interactions. As a proof-of-concept study, we fabricate MOFS using tryptophan and lactulose as the substrates and employing Enterococcus faecalis (E. faecalis), a lactic acid bacterium that metabolizes tryptophan and lactulose separately to indole and lactic acid, as the substrate-related microbe. The water-impermeable property of MMT endows MOFS with strong resistance to harsh gastrointestinal environments. Meanwhile, the mucoadhesive capacity of MMT enables the immobilization of MOFS on the mucus layer. In addition to the resulting high oral availability and prolonged retention in the intestine, the continuous release of carried tryptophan and lactulose and the formation of E. faecalis biofilms ensure long-lasting yet efficient production of lactic acid and indole in the gut (Fig. 1b). These beneficial bioproducts synergistically regulate the microbiota structure, mucosal immune responses, and barrier functions of the intestine. In two murine models of infective enteritis and IBD associated with Candida albicans (C. albicans) infection, MOFS effectively mitigates the related symptoms by restoring intestinal homeostatic balance (Fig. 1c). Given its flexibility of using cation replacement and electrostatic interactions to combine other metabolic substrates and bacterial strains, we anticipate the broad application of MOFS for in-situ biosynthesis of diverse therapeutics for treating different diseases.

a Preparation of MOFS by cation replacement and absorption. b High resistance of MOFS and its long-lasting in situ synthesis of beneficial bioproducts in the gut. c MOFS-mediated protection of intestinal hemostasis by improving gut microbiota composition, suppressing intestinal inflammation, and increasing the integrity of the gut barrier. Created in BioRender. Liu, J. (2026) https://BioRender.com/dprgxuh.
Results and discussion
Design, preparation and characterization of MOFS
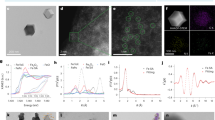
We chose MMT, a clinically used nanosheet-structured layered clay, as a carrier material due to its distinctive physicochemical characteristics, such as high stability, mucoadhesive ability, high adsorption capacity, water impermeability, and high cation-exchange capacity20. MMT is a 2:1 smectite with a permanent layer charge arising from isomorphic substitution21. Negative surfaces are balanced by exchangeable interlayer cations (e.g., Na⁺, Ca2+), conferring a high cation-exchange capacity21. It has been indicated that the abundant lipophilic components of the bacterial cell walls, such as lipoproteins and liposaccharides, make them easily adsorbed by MMT22. Indole, a principal intercellular signaling molecule in gut microbial ecosystems, plays dual regulatory roles in intestinal physiology by modulating the expression of tight junction proteins and orchestrating immune responses through aryl hydrocarbon receptor-mediated pathways23. Lactic acid refers to a readily accessible fuel for intestinal epithelia and an important metabolic buffer that affects the cell microenvironment, accounting for its key role in maintaining gut homeostasis24,25. Given these facts, we selected indole and lactic acid as targeted beneficial bioproducts for restoring intestinal homeostasis. Indole and lactic acid are mainly derived from the metabolism of tryptophan and lactulose by gut microorganisms, such as E. faecalis, Streptococcus rosenbergii, and C. sporogenes26,27. As a symbiotic specie, E. faecalis is widely used in dairy productions27. Thus, we chose tryptophan and lactulose as the substrates and E. faecalis as the substrate-related microbe to fabricate MOFS. The ability of E. faecalis to metabolite tryptophan and lactulose to indole and lactic acid was tested using Kovac’s reagent and methyl red, respectively. As shown in Supplementary Fig. 1a, there was a cherry red ring occurred on the top of the culture medium, suggesting the presence of indole. This could be explained by the combination of indole with p-dimethylaminobenzaldehyde in Kovac’s reagent. Similarly, after adding methyl red to the culture medium, the color changed from yellow to red, indicating the production of lactic acid that triggered an acidic environment and the change of the medium color (Supplementary Fig. 1b). We then optimized the conditions for the fabrication of MOFS by measuring the viability and proliferation of E. faecalis after incubation with different contents of MMT, tryptophan, and lactulose, respectively. As plotted in Supplementary Fig. 1c, the viability of E. faecalis increased with the amount of MMT, reaching a significant proliferation at 25 mg/ml. E. faecalis presented high viability in the medium supplemented with 0.6 mg/ml of tryptophan and 0.6 mg/ml of lactulose (Supplementary Fig. 1d–f). To ensure sufficient adsorption of bacteria by MMT, the incubation time was optimized. By measuring the number of adsorbed bacteria in MMT, a high ratio of 99% E. faecalis was obtained after a 4-hour incubation (Supplementary Fig. 1g). In comparison with free E. faecalis (Bac), scanning electron microscope (SEM) images showed that bacteria adsorbed in MMT (MMT@Bac) were covered by lamellar MMT (Fig. 2a). The adsorption of E. faecalis by MMT was further confirmed by laser scanning confocal microscopy (LSCM). We labeled MMT with Rhodamine B (RhB) by 1-(3-dimethylaminopropyl)−3-ethylcarbodiimide (EDC) and N-hydroxy succinimide (NHS). E. faecalis adsorbed in the resulting RhB-labeled MMT (RhB-MMT) was further stained with 4’,6-diamidino-2-phenylindole (DAPI). As expected, LSCM images displayed a consistent colocalization between E. faecalis and MMT (Fig. 2b and Supplementary Fig. 2). Note that there were rare free E. faecalis appeared around MMT, indicating efficient adsorption of E. faecalis by MMT.

a SEM images of Bac, MMT, and MMT@Bac. Scale bar, 1 μm. The experiment was independently repeated at least twice. b LSCM images of MMT@Bac. The red channel indicates MMT stained with RhB. The blue channel indicates bacteria labeled with DAPI. Scale bar, 10 μm. The experiment was independently repeated at least twice. c XRD patterns of MMT, MMT@T, MMT@L, and MMT@Bac@TL. The formula is the Bragg’s Law: 2dsinθ = nλ (d is the distance between crystal planes); θ is the Bragg angle, which is the angle of incidence (and reflection) of the X-ray upon the crystal plane; λ is the wavelength of the incident X-ray; n is the order of reflection, representing the integer multiples of the wavelength). d, e Release of tryptophan (d) and lactulose (e) from MMT (n = 3 independent samples). f ζ-potentials of MMT, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL measured by DLS (n = 6 independent samples). g, h Survival rate of E. faecalis in Bac, MMT@Bac, and MMT@Bac@TL treated with SGF (g) and SIF (h) at the indicated time points (n = 3 independent samples). i, j Counts of E. faecalis in the Bac, MMT@Bac, and MMT@Bac@TL groups after incubating in the separated segment of the ileum (i) and colon (j), respectively (n = 3 independent samples). k, n, q, Amounts of E. faecalis in the mucus layer of the ileum (k), cecum (n), and colon (q), respectively (n = 3, 4 or 5 independent samples). l, o Amounts of E. faecalis in the contents of the ileum (l) and cecum (o), respectively (n = 4 or 5 independent samples). m, p, r, Total bacterial amounts in the ileum (m), cecum (p), and colon (r), respectively (n = 4 or 5 independent samples). Data are represented as means ± SD. p-values were determined using one-way ANOVA. p-values < 0.05 are shown. Source data are provided as a Source Data file.
On the basis of these results, we prepared MOFS by mixing MMT with tryptophan and lactulose at a weight content ratio of 40:1:1 and subsequently incubating with 8 × 108 colony forming units (CFUs)/ml of E. faecalis for 4 hours. To facilitate comparison, the obtained MMT carrying bacteria, tryptophan, and lactulose was defined as MMT@Bac@TL. Meanwhile, MMT@Bac, MMT loaded with tryptophan (MMT@T), MMT loaded with lactulose (MMT@L), MMT loaded with E. faecalis and tryptophan (MMT@Bac@T), and MMT loaded with E. faecalis and lactulose (MMT@Bac@L) were prepared as controls, respectively. MMT@Bac@TL showed limited fluctuations in the viability of carried bacteria, demonstrating both the preparation procedure and the components of MMT, tryptophan, and lactulose had a negligible impact on E. faecalis vitality (Supplementary Fig. 1h). The characteristic X-ray powder diffractometer (XRD) reflection of MMT appeared at 2θ = 6.34° (1.25 nm). In MMT@T and MMT@L, the basal space of MMT increased slightly from 1.25 to 1.31 and 1.33 nm, due to the insertion of tryptophan and lactulose. This indicated that tryptophan and lactulose were intercalated inside the interlayer space of MMT (Fig. 2c). The cation exchange capacity of MMT enabled high loading contents of tryptophan and lactulose, which were calculated to 91% and 93% (Supplementary Fig. 1i), respectively. To verify the releases of tryptophan and lactulose, MMT@Bac@TL were incubated in simulated intestinal fluid (SIF), and the accumulated release at different time points was measured using an ultraviolet-visible spectrophotometer. After incubation for 12 hours, the total releases of tryptophan and lactulose separately reached 62% and 83% (Fig. 2d and e). We further measured the ζ-potentials of MMT, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL, respectively. In contrast to the ζ-potential of MMT (− 8.0 mV), the ζ-potential of MMT@Bac largely decreased (− 27.8 mV), but this reduction was mitigated as the loading of tryptophan and lactulose (Fig. 2f). Collectively, these results proved MMT@Bac@TL could be simply constructed by taking advantage of MMT to adsorb both the metabolic substrates and the substrate-related bacteria.
High oral availability and extended gut retention of MOFS
Living bacteria suffer from deactivation and disintegration following oral ingestion due to the presence of gastric acid and intestinal juice stimulations28,29. We speculated that the water-impermeable property of MMT might protect the loaded bacteria from harsh gastrointestinal environments. To verify this, we examined the protective effect of MMT during gastrointestinal transit by counting the numbers of survived bacteria at different time points after incubation in simulated gastric fluid (SGF) and SIF, respectively. As expected, the presence of MMT could increase the survival of E. faecalis in gastric acid. After a 15-minute culture in SGF, 91% of the bacteria in the Bac group were deactivated. In contrast, the influence on E. faecalis viability was negligible in the MMT@Bac@TL group (Fig. 2g). It was found that MMT@Bac also had a higher survival rate compared to the Bac group, indicating the protective effect of MMT toward E. faecalis. The ability of MMT to protect loaded bacteria was further confirmed by incubation with SIF, as both MMT@Bac and MMT@Bac@TL enabled a greatly increased survival rate compared to the Bac group (Fig. 2h). MMT is a clay containing multiple isomeric substituents, where the aluminum and silicon atoms at the crystal edges undergo partial hydrolysis in water, forming silanol and aluminum hydroxyl groups30,31. These unsaturated edge sites are more reactive than saturated base sites and can bind to mucin in the mucus layer of the intestinal wall through hydrogen bonding, van der Waals forces, and hydrophobic interactions30,31. To examine the retention on the mucus layer, an equal area of ileum and colon segments was sampled to expose the mucus layer, followed by incubation with Bac, MMT@Bac, and MMT@Bac@TL, respectively. The numbers of E. faecalis in the different segments were counted after an overnight culture. As shown in Fig. 2i, compared to free bacteria, MMT@Bac@TL exhibited ~ 3-fold increment in adhesion to the mucus layer of the ileum compared to Bac or MMT@Bac. Likewise, MMT@Bac@TL presented the highest number of bacteria adhered to the mucus layer of the colon, further supporting its strongest adhesive ability (Fig. 2j).
The oral availability and gut retention of MMT@Bac@TL were also evaluated in vivo. To quantify the number of survived E. faecalis, the contents of the lumen were separately collected from different intestinal segments for bacterial plate counting 4 hours after oral administration of 4 × 107 CFUs of Bac, MMT@Bac, and MMT@Bac@TL in mice, respectively. Interestingly, the count of survived E. faecalis in the luminal contents collected from mice orally delivered with MMT@Bac@TL was substantially higher than those in the Bac and MMT@Bac groups. As plotted in Supplementary Fig. 3a, the amounts of living E. faecalis in the mucus layer from cecum and colon segments were increased by 48 times compared to either Bac or MMT@Bac. Similarly, it was found that in the luminal contents of the cecum and colon segment, the number of survived E. faecalis in the MMT@Bac@TL group reached 2 × 106 CFUs/g, which was higher than those of Bac (~ 3-fold) and MMT@Bac (~ 8-fold) (Supplementary Fig. 3b). As calculated, MMT@Bac@TL exhibited the highest total survival of delivered bacteria 4-hour post administration (Supplementary Fig. 3c). We further examined the long-term adhesive ability of MMT@Bac@TL in mouse intestine and found that the amounts of living E. faecalis in the luminal content and the mucus layer of the entire intestine from mice orally delivered with MMT@Bac@TL were higher than those with Bac and MMT@Bac 72 hours after oral ingestion (Supplementary Fig. 3d, e). Correspondingly, the total number of survived bacteria in the entire intestine was the highest in the MMT@Bac@TL group (Supplementary Fig. 3f). We also counted the number of survived bacteria in the luminal content of each segment, including the ileum, cecum, and colon (Fig. 2k–r). The plotted results indicated that rare living E. faecalis was detected in each location in the Bac and MMT@Bac groups. While there were a large number of survived bacteria in these locations in the MMT@Bac@TL group. Particularly, the total survivals were over 106 CFUs/g in the ileum (Fig. 2m), the cecum (Fig. 2p), and the colon (Fig. 2r), respectively. Give MMT is known to bind mucin in the mucus layer30,31, the differential retention of MMT@Bac@TL across gut segments might be resulted from the structural and thickness variations in the mucus layer32. In addition, pH gradients along the gastrointestinal tract and gut motility could also play a role by influencing the binding33. Taken together, these results demonstrated the high oral availability and extended gut retention of MMT@Bac@TL.
MOFS-enabled long-lasting in-situ synthesis of beneficial bioproducts
To determine the metabolic efficiency of MMT@Bac@TL, the levels of indole and lactic acid were measured using corresponding test kits after incubation for the indicated time points. The levels of indole in the MMT@Bac@TL group after 72 hours of incubation were ~ 1.7- and ~ 2.3-fold higher than those of Bac + T (a simple mixture of E. faecalis and tryptophan) and MMT@Bac@T, respectively (Fig. 3a). Meanwhile, the concentration of lactic acid produced by MMT@Bac@TL reached ~ 0.5 mmol/L after 24 hours of incubation. Differently, only ~ 0.1 mmol/L lactic acid was produced by Bac + L (a simple mixture of E. faecalis and lactulose) or MMT@Bac@L (Fig. 3b). The higher productions of indole and lactic acid in MMT@Bac@TL compared to the simple mixture of bacteria with metabolic substrates (Bac + T or Bac + L) might be ascribed to the catalytic characteristic of MMT34,35. In addition, the synergistic effects of tryptophan and lactulose on bacterial growth and metabolism contributed to the MMT@Bac@TL-mediated high production of indole and lactic acid. Lactulose serves as a prebiotic, promoting the growth of lactic acid bacteria36. In turn, this facilitates the metabolism of tryptophan, resulting in increased indole production. Meanwhile, indole has been shown to modulate the growth, viability, biofilm formation, motility, and stress resistance in different bacteria37. We next evaluated the influence of metabolic substrate supplements on the metabolic profile of MMT@Bac@TL in vivo. Mice were orally delivered with Bac, MMT@Bac, or MMT@Bac@TL, and the concentrations of indole and lactic acid in different intestinal segments were measured 4 hours after gavage. As shown in Fig. 3c-e, the concentration of indole in the MMT@Bac@TL group was the highest among all the groups. The concentration of indole in the cecum was increased by ~ 2.7- and ~ 2.0 times compared to Bac or MMT@Bac (Fig. 3c). The total levels of indole in the cecum and colon segment sampled from mice treated with Bac and MMT@Bac were only ~ 0.7 mmol/gprot, which was increased by ~ 2 times in the MMT@Bac@TL group 4 hours after oral administration (Fig. 3d). Accompanied by promoted production of indole, the concentration of lactic acid in the MMT@Bac@TL group was increased by ~ 4- and ~ 2-fold compared to those in the Bac and MMT@Bac group 12 hours after oral administration, respectively (Fig. 3f). Moreover, we monitored the metabolites over a time course. The concentration of indole in the MMT@Bac@TL group was increased by ~ 1.4-fold compared to those in the Bac and MMT@Bac group 24 hours after oral administration, respectively (Supplementary Fig. 4a). Indole levels at 48 hours after oral administration and lactate levels at 24 hours and 48 hours showed no significant differences (Supplementary Fig. 4b–d). Notably, both the levels of indole and lactic acid in the MMT@Bac@TL group were increased by ~ 2-fold compared to the Bac group 72 hours after oral gavage. In contrast, the administration of MMT@Bac failed to enhance lactic acid production throughout the time course (Fig. 3g, h). These dynamic changes in indole and lactic acid production observed over time in the MMT@Bac@TL group might be associated with transitions, substrate availability, and microbial adaptation of the sample in the gut. Collectively, these data showed that MMT@Bac@TL enabled long-lasting in situ biosynthesis in the intestine, and that the production of beneficial metabolites could be amplified by co-loading tryptophan and lactulose.

a, b Levels of indole (a) and lactic acid (b) after 72 or 24 hours of incubation (n = 4 or 5 independent samples). c Concentration of indole in the cecum (n = 4 or 5 independent samples). d,Total indole level in the cecum and colon (n = 4 or 5 independent samples). e Concentration of indole in the entire intestine 4 hours after oral administration (n = 4 or 5 independent samples). f Concentration of lactic acid in faces 12 hours after oral administration (n = 4 or 5 independent samples). g Concentration of indole in the entire intestine (n = 4 or 5 independent samples). h Concentration of lactic acid in the colon 72 hours after oral gavage (n = 4 or 5 independent samples). i, j Biofilm mass of Bac, MMT@Bac, and MMT@Bac@TL after 6 hours (i) or 48 hours (j) of incubation in SIF (n = 4 or 5 independent samples). k Confocal images of the biofilms formed by Bac, MMT@Bac, and MMT@Bac@TL, respectively. Biofilms are identified by Con A (Red). Bacteria are labeled with DAPI. A section of the ileum was placed on a concave glass slide. Then, Bac, MMT@Bac, and MMT@Bac@TL re-suspended in 0.5 ml SIF were incubated on the ileum section for 72 hours. Scale bar, 30 μm. Data are represented as means ± SD. p-values were determined using one-way ANOVA. p values < 0.05 are shown. Source data are provided as a Source Data file.
It has been reported that bacteria resided in biofilms express distinct properties compared to their free-living counterparts, including increased metabolic activities38,39. For instance, intestinal bacteria associated with biofilms can metabolize diet and mucin-related carbohydrate complexes into short-chain fatty acids, such as acetates, propionates, and butyrates38,40. As efficient surface adhesion is a prerequisite for biofilm formation, we speculated that the mucoadhesion ability of MMT@Bac@TL might promote the formation of E. faecalis biofilms, thereby enhancing bacterial metabolism. To confirm this hypothesis, we next assessed the formation of biofilms by MMT@Bac@TL using Congo red test. Typically, on Congo red plates, bacterial strains that produce rough and black colonies are recognized as a sign of biofilm formation. After 24 and 48 hours of incubation, the groups containing MMT, including MMT@Bac and MMT@Bac@TL, changed to brown or black, while Bac remained in red (Supplementary Fig. 5a). We further quantified the formation of biofilms using Congo red staining after 24 and 48 hours of incubation in SIF. In the MMT@Bac@TL group, the absorbance at 495 nm showed approximately 3-fold (24 hours) and 2-fold (48 hours) increments compared to the Bac group, respectively (Supplementary Fig. 5b). The enhanced biofilm-forming ability of MMT@Bac@TL was also confirmed by crystal violet staining. After culturing each group in a 96-well plate for 6 or 48 hours, an absorbance measurement at 540 nm was performed to quantify biofilm production. Consistent with the results of the Congo red assay, MMT@Bac@TL showed ~ 18 and ~ 34-fold increments compared to the Bac group after 6 and 48 hours of incubation, respectively (Fig. 3i, j). The enhanced biofilm formation observed in MMT@Bac could be attributed to MMT’s high surface area, which provides an adhesive scaffold that stabilized the development and maturation of biofilm matrix41. In addition, we measured the biofilm production in the intestinal environment by culturing Bac, MMT@Bac, and MMT@Bac@TL in both isolated intestinal segments and SIF. Following 48 and 72 hours of incubation, the tissues were stained with DAPI and Alexa Fluor 633-concanavalin A (Con A) to observe biofilm formation. As shown in Fig. 3k and Supplementary Fig. 6a, the mean fluorescence intensity (MFI) of Con A in the MMT@Bac@TL group was significantly higher than other groups, indicating a denser biofilm layer on the inner surface of the intestine. The Con A MFI of MMT@Bac@TL was ~ 40-fold higher than MMT@Bac and ~ 14-fold higher than Bac (Supplementary Fig. 6b). Meanwhile, the formation of biofilms was further confirmed by SEM. Unlike the smooth surface of bare bacteria, MMT@Bac displayed a rough surface due to the presence of MMT. An enhanced bacterial aggravation on a rougher surface was observed in the MMT@Bac@TL group compared to MMT@Bac as the formation of biofilms (Supplementary Fig. 6c). Moreover, we evaluated the biofilm formation by MMT@Bac@TL in vivo. Mice were orally administered with Bac, MMT@Bac, or MMT@Bac@TL, and the colon segments were collected 6 days post-treatments. As depicted in Supplementary Fig. 7, compared to either Bac- or MMT@Bac-treated mice, more biofilms were observed on the colon sections sampled from MMT@Bac@TL-treated mice. Both in vitro and in vivo experiments revealed that the superior biosynthesis efficiency of MMT@Bac@TL was benefited from the formation of biofilms, which might be associated with the upregulated energy metabolism of bacteria in biofilms. An upregulated energy metabolism characterized by enhanced glycolysis and tricarboxylic acid cycle activity has been observed in biofilms38,39. The enhanced energy metabolism supplies additional adenosine triphosphate (ATP) and central carbon intermediates to fuel anabolic pathways, such as amino acid, lipid, and exopolysaccharide biosynthesis42. Thus, we further evaluated potential metabolic pathways underlying the biofilm-mediated increase in metabolite production by comparing pyruvate and ATP levels between MMT@Bac@TL with and without biofilms. Given that biofilm formation was observed after 3 days of SIF culture, we cultured MMT@Bac@TL in PBS using the same time schedule as a biofilm-negative control. As shown in Supplementary Fig. 8, compared to MMT@Bac@TL cultured in PBS, the MMT@Bac@TL in SIF presented increases in the levels of pyruvate (~ 10-fold) and ATP ( ~ 2-fold).
Positive modulation of the gut microbiota by MOFS
The gut microbiota is considered as one of the key elements contributing to the regulation of intestinal homeostasis7. The imbalance of gut homeostasis caused by the changes in microbial structure, namely gut dysbiosis, is linked to a broad spectrum of diseases1. Probiotic bacteria play significant roles in maintaining a healthy microbial structure by secreting beneficial metabolites3. For example, indole and lactic acid can downregulate the expressions of virulence genes and/or movement/invasion-related genes, thereby reducing the virulence and biofilm formation of pathogens3,37,43. Given the enhanced production of metabolites, we next investigated the effects of MMT@Bac@TL on the modulation of the gut microbiota. We first cultured a classical probiotic strain of Escherichia coli Nissle 1917 (EcN) in a culture medium supplied with Bac, MMT@Bac, or MMT@Bac@TL. EcN cultured in pure culture medium was set as a control. Plate counting of EcN was conducted after 2 and 4 hours of incubation at 37 °C on Luria-Bertani (LB) agar plates containing 50 μg/ml kanamycin to eliminate E. faecalis. Compared to other groups, the supplement of MMT@Bac@TL significantly promoted the growth of EcN (Fig. 4a, b). After 2 hours of culture, the growth rate of EcN in the medium group reached only ~ 64%. The supplement of MMT@Bac substantially increased the growth rate. The growth rate was 144-fold higher in the MMT@Bac@TL group in contrast to the MMT@Bac group. This effect of MMT@Bac@TL on EcN growth was observed with incubation extending to 4 hours. We further tested the effect of MMT@Bac@TL on the growth of pathogenic Salmonella Typhimurium (S. typhimurium). As shown in Fig. 4c, MMT@Bac@TL presented a 25-fold increment in the inhibition of S. typhimurium growth. The supplement of Bac could not inhibit the growth of S. typhimurium. Although MMT@Bac showed an inhibitory effect on S. typhimurium, with the culture time prolonged to 6 hours, only MMT@Bac@TL could effectively inhibit the growth of S. typhimurium (Fig. 4d). The different effects of MMT@Bac@TL on EcN and S. typhimurium might be relevant to the context where the interspecies signaling molecule indole can modulate the expression of crucial regulatory genes in S. typhimurium44,45. On the other hand, lactic acid can cause a drop in pH, inhibiting the growth of pathogens24. To further understand the influence of MMT@Bac@TL on the gut microbiota, we studied the variation of microbial composition in mice after suffering from antibiotic cocktails. As illustrated in Supplementary Fig. 9, following 2-week treatments with a cocktail of neomycin sulfate, vancomycin, ampicillin, and metronidazole, mice were orally administered with phosphate-buffered saline (PBS), Bac, MMT@Bac, or MMT@Bac@TL at a dose per day for 7 days. Mice without any treatment were set as a healthy control. The similarity of microbial communities among groups was confirmed using principal component analysis (PCA). As validated in Fig. 4e, the microbial structure of mice treated with MMT@Bac@TL was closer to that of healthy mice. Taxonomic analysis and linear discriminant analysis effect size (LEfSe) indicated that oral administration of MMT@Bac@TL enriched the abundances of Bacilli, Synergistota, and Staphylococcales (Fig. 4f–i and Supplementary Fig. 10). It has been identified that most species from Bacill class show a potent effect against pathogens by producing bacteriocin46. Synergistota has been identified as a species capable of producing butyrate, which has received considerable attention for its multiple beneficial effects on intestinal homeostasis47. Although the strains from the Staphylococcales order are recognized as pathogens, a group of antimicrobial peptides and anti-inflammatory peptides produced by Staphylococcus aureus (S. aureus) have been identified48. For example, the extracellular adherence protein expressed by S. aureus has been demonstrated to be an anti-inflammatory factor that inhibits host leukocyte recruitment, T-cell proliferation, and delayed-type hypersensitivity responses49. Overall, these data verified the capability of MMT@Bac@TL to positively regulate the gut microbiome.

a, b Growth rate of EcN after 2 (a) or 4 (b) hours of incubation with culture medium, Bac, MMT@Bac, and MMT@Bac@TL, respectively (n = 5 or 6 independent samples). c, d Growth rate of S. typhimurium after 4 (c) or 6 (d) hours of incubation with culture medium, Bac, MMT@Bac, and MMT@Bac@TL, respectively (n = 4, 5 or 6 independent samples). Antibiotic-treated mice were administered with PBS, Bac, MMT@Bac, and MMT@Bac@TL at a dose per day for 7 days, respectively. Then, the cecum was collected for 16S rRNA gene sequencing. Mice without any treatment were set as a healthy control. e Results of the PCA of the gut microbiota at class level (n = 5 independent samples). p-value was analyzed by non-parametric statistical test ANOSIM. f Cladogram of LEfSe from phylum to species levels (n = 5 independent samples). LEfSe analyses were performed with the Kruskal-Wallis rank sum test. The threshold of the LDA score is 3.5. g Abundances of the gut microbiota at the class level. h, i Relative abundances of Bacill in class level (h) and Synergistota in phylum level (i) (n = 4 or 5 independent samples). Data are represented as means ± SD. p-values were determined using one-way ANOVA. p-values < 0.05 are shown. Source data are provided as a Source Data file.
Suppression of intestinal inflammation by MOFS
The advances in understanding the compositions and functions of individual microbial species and complex microbial communities highlight the importance of microbial metabolism to the host immune system5. Recent studies report that lactic acid acts as an essential signaling molecule that shapes the innate and adaptive immune responses by upregulating the expressions of anti-inflammatory genes or M2-like genes25. It has also been documented that as important metabolites of lactobacillus species, indole and its derivatives enhance the M2 polarization of macrophages, subsequently reducing intestinal inflammation50. Given the critical role of macrophages in the maintenance of intestinal homeostasis, we first evaluated the effect of MMT@Bac@TL on the polarization of macrophages. J774A.1 cells treated with lipopolysaccharide (LPS) were cultured with blank medium, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL, respectively. Cells without any treatment were set as a control. After 24 hours of culture, the percentage of M2 macrophages in the MMT@Bac@TL group was ~ 2-fold higher than those in other groups (Fig. 5a–d). Furthermore, the M2/M1 classification ratio also showed a significant increase, indicating a notable M2 polarization effect (Fig. 5e). The augmented M2 polarization was followed by a notable increase in the levels of cytokines and chemokines involved in regulating intestinal homeostasis (Fig. 5f–h). Although its association with inflammation responses, interleukin-1β (IL-1β) is crucial for maintaining the homeostasis of intestinal immune responses, such as immune globulin A (IgA) production51. Vascular endothelial growth factor (VEGF) plays a vital role in epithelial cell restitution by preserving mucosal homeostasis following mucosal damage52. Likewise, transforming growth factor-β (TGF-β) is a key regulator of the regenerative process of the intestine53. As evidenced, the levels of IL-1β, VEGF, and TGF-β secreted by macrophages cultured with MMT@Bac@TL were the highest. In addition, interleukin-4 (IL-4) was 30% increased by MMT@Bac@TL compared to the medium group, contributing to an increased ratio of IL-4 to IL-6 (Supplementary Fig. 11a, b). Meanwhile, the ratio of interleukin-10 (IL-10) to nitric oxide (NO) in the MMT@Bac@TL group was 17% higher than in the medium group (Supplementary Fig. 11c). IL-4 and IL-10 are key mediators of mucosal immune regulation to control inflammation54,55. In contrast, increases in IL-6 and NO are commonly involved in the aggravation of immunopathology during gut inflammation56,57. Dendritic cells (DCs) are extensively involved in maintaining intestinal homeostasis by mediating the tolerance to food antigens, limiting the reactivity to the gut microbiota, and responding to intestinal pathogens58. In addition to macrophage polarization, MMT@Bac@TL facilitated the maturity of DCs (Supplementary Fig. 12). We further examined the effects of MMT@Bac@TL on the intestinal immune system in vivo. Consist with the effects of MMT@Bac@TL on macrophages and DCs, MMT@Bac@TL promoted the M2 polarization of macrophages and the maturation of DCs in the mesenteric lymph nodes (MLNs) (Fig. 5i, j and Supplementary Fig. 13a, b). DCs are known to regulate the production of IgA by plasma cells59. Indeed, accompanied by increased DCs, there were more IgA+ B cell populations in the Peyer’s patches (PPs) sampled from MMT@Bac@TL-treated mice compared to other treatments (Fig. 5k and Supplementary Fig. 13c). Note that IgA plays a pivotal role in gut homeostasis through orchestrating the mucosal responses, namely protecting the mucosal surface from pathogen invasion while tolerating commensal bacteria6,60 Briefly, MMT@Bac@TL presented a favorable ability to regulate gut immune responses for maintaining intestinal homeostasis.

After 24 hours of LPS (1 µg/ml) stimulation, J774A.1 cells were incubated with culture medium, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL for 24 hours. Cells without any treatment were set as a control. a, b Flow cytometric analyses of (a) M2 and (b) M1 macrophages. c, d Percentages of M2 (c) and M1 (d) macrophages. e Ratio of M2 to M1 macrophages (n = 5 or 6 independent samples). f–h Concentrations of IL-1β (f), TGF-β (g), and VEGF (h) in the supernatant (n = 4, 5 or 6 independent samples). Mice pretreated with an antibiotic cocktail were administered with PBS, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL one dose per day for 7 days. i, j Percentages of M2 macrophages (i) and mature DCs (j) in the MLNs (n = 6 independent samples). k Percentage of IgA+ B cells in the PPs (n = 4-6 independent samples). Data are represented as means ± SD. p-values were determined using one-way ANOVA. p-values < 0.05 are shown. Source data are provided as a Source Data file.
Protection of intestinal barrier integrity by MOFS
The composition of the gut microbiome influences the intestinal immune system, which in turn affects the integrity of the gut barrier2. In light of this, we then evaluated the capability of MMT@Bac@TL to protect the intestinal barrier from external insults. The integrity of the intestinal barrier is typically related to the mucus layer and tight junctions1. Under normal physiological conditions, goblet cells continually produce mucin to replenish and maintain the mucus layer. Recent studies disclose that the gut microbiota and its metabolites modulate the functions of goblet cells61. For instance, indole has been demonstrated to enhance mucus secretion by upregulating IL-10 and MUC gene expression62. We hence cultured HT29-MTX-E12 cells, a mucus-producing intestinal cell line, with 50 μg/ml LPS for 6 hours and subsequently treated them with 1 × 105 CFUs of Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL for an additional 6 hours, respectively. The level of mucus was then assessed using Alcian Blue/Periodic acid-Schiff (AB/PAS) staining, which can distinguish neutral mucin from acid mucin, thereby providing a comprehensive analysis of the mucus composition. As illustrated in Fig. 6a, the proportion of mucus was significantly reduced due to LPS stimulation. As expected, incubation with MMT@Bac@TL showed the highest accumulation of mucus compared to other treatments. It was worth noting that MMT@Bac@TL exhibited an ~ 1.1-fold increase in acidic mucin production compared to the medium group (Fig. 6b). Meanwhile, the level of neutral mucin was decreased under the treatment of MMT@Bac@TL in contrast to the medium group (Fig. 6c). This resulted in an increased ratio of acidic mucin to neutral mucin, which was close to the normal condition (Fig. 6d). It has been identified that secreted mucin becomes more acidic and develop higher N- and O- linked glycosylation levels during maturation, which are implicated in the protection of the mucus layer from pathogen invasion63,64. In addition to the mucus layer, numerous studies have found that indole and lactic acid can enhance endothelial cell barrier functions by inducing the expression of genes involved with endothelial cell structures, including tight junctions, adherent junctions, and actin cytoskeleton65. As a widely used in vitro model of the intestinal barrier, Caco-2 cell monolayer was treated with 50 μg/ml LPS for 12 hours and then incubated with 1 × 104 CFUs of Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL for 7 hours, respectively. We first measured the effect of MMT@Bac@TL on cell permeability by quantifying the fluorescence intensity of FITC-dextran (4 kDa) that permeated from the upper to the lower chamber of the transwell device. As shown in Supplementary Fig. 14, compared to the cell culture medium-treated group, the fluorescence intensity of FITC-dextran was lower in the MMT@Bac@TL group, indicating decreased cell permeability. Given that tight junction barrier loss is an important contributor to increased barrier permeability, we next measured the expressions of tight junction proteins zona occludens 1 (ZO-1) and occludin under different treatments. The cells were stained with specific antibodies against the tight junction protein ZO-1 and occludin, which are critical components of the intestinal barrier. Expectedly, LPS stimulation reduced the expression of ZO-1 and occludin, which were notably recovered after incubation with MMT@Bac@TL. In contrast, other treatments presented negligible effects in the protection of tight junctions, particularly the expression of occludin (Fig. 6e and Supplementary Fig. 15a). Furthermore, in the antibiotic cocktail-induced mouse model of gut dysbiosis, we verified the potential of MMT@Bac@TL to protect the intestinal barrier. Compared to healthy mice, the antibiotic cocktail led to a marked depletion of goblet cells. In stark contrast to the sparse mucus observed in other treatment groups, the reduced expression of mucus was reversed by the MMT@Bac@TL treatment, which resulted in a level closely aligning with patterns found in healthy mice (Fig. 6f, h). We also validated the capability of MMT@Bac@TL to repair tight junctions in the colon of mice treated with antibiotics. Compared to the healthy control, antibiotic cocktail-induced gut dysbiosis had a more pronounced effect on the tight junction protein ZO-1 than on occludin. It is indicated that pro-inflammatory and contractile cues associated with dysbiosis rapidly remodel ZO-1 continuity and localization at cell-cell borders66,67. By contrast, occludin homeostasis is more strongly influenced by context-dependent endocytosis/turnover pathways68. Notably, both decreased occludin and ZO-1 expressions triggered by antibiotic cocktail-induced gut dysbiosis could be greatly reversed by MMT@Bac@TL (Fig. 6g, i, j and Supplementary Fig. 15b). Together, MMT@Bac@TL could protect the intestinal barrier from damage under external attacks.

a Mucus released by HT29-MTX-E12 cells under 6 hours of incubation with culture medium, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL, respectively. Cells without any treatment were set as a control. Scale bar, 200 µm. b, c Levels of acidic (b) and neutral (c) mucin secreted by HT29-MTX-E12 cells under different treatments (n = 6 independent samples). d Ratio of acidic to neutral mucin (n = 6 independent samples). e Confocal images of tight junction expression in LPS pretreated Caco-2 cells after treatment with culture medium, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL for 7 hours, respectively. Cells without any treatment were set as a control. Red, green, and blue indicate the signals of occludin, ZO-1, and cell nuclei, respectively. Scale bar, 35 µm. f Images of goblet cells of the colon identified by AB-PAS staining. Scale bar, 60 µm. g Confocal images of colon sections stained with occludin (Red), ZO-1 (Green), and DAPI (Blue). Scale bar, 60 µm. h Mean gray value of goblet cells measured by Image-J (n = 3 independent samples). i, j MFI of occludin (i) and ZO-1 (j) measured by Image-J (n = 3 independent samples). Data are represented as means ± SD. p-values were determined using one-way ANOVA. p-values < 0.05 are shown. Source data are provided as a Source Data file.
Potential of MOFS in treating S. typhimurium-induced enteritis
Encouraged by the beneficial effects of MMT@Bac@TL on intestinal homeostasis, we then evaluated the potential of MMT@Bac@TL in treating enteritis caused by S. typhimurium infection, which is a frequently occurred clinical scenario. A typical syndrome associated with infectious enteritis is the presence of intestinal homeostatic imbalance. However, currently available therapeutics are difficult to comprehensively restore intestinal homeostasis. To develop a murine model of enteritis, mice were infected with 5 × 107 CFUs of S. typhimurium via oral gavage 24 hours after pretreatment with streptomycin (Fig. 7a). The mice were orally administered with MMT, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL one dose per day for 6 days, respectively. In each treatment group, equivalent MMT (0.05 mg), E. faecalis (1 × 107 CFUs), tryptophan (0.0013 mg), or lactulose (0.0013 mg) was applied. Infected mice treated with PBS were used as a negative control. The untreated mice served as a healthy control. Meanwhile, MMT, a type of medical clay used to eliminate pathogens69, was used as a positive control. As shown in Fig. 7b and c, S. typhimurium infection-induced decrease of colon length was effectively restored by MMT@Bac@TL. In contrast, other treatments failed to control this symptom. Specifically, the colon length of MMT@Bac@TL-treated mice (9.4 ± 0.1 cm) was longer than those of mice treated with PBS (7.7 ± 1.1 cm), MMT (8.2 ± 0.6 cm), Bac (7.4 ± 1.2 cm), MMT@Bac (8.0 ± 0.9 cm), MMT@Bac@T (8.3 ± 0.8 cm), and MMT@Bac@L (8.3 ± 0.9 cm), highlighting its superior treatment efficacy. To confirm that the improved effects were attributed to the enhanced production of beneficial metabolites, we measured the concentrations of indole and lactic acid in the gut. As plotted in Fig. 7d, compared to PBS, the treatments of Bac, MMT@Bac, and MMT@Bac@T could only induce a 3.0-fold increase in indole production. The indole level in the ileum sampled from mice treated with MMT@Bac@TL group was ~ 7.3 times higher than that in the PBS group. We also noted that the concentration of lactic acid in the MMT@Bac@TL group reached approximately 63 mmol/g in the intestine (Fig. 7e), which was increased by ~ 1.4 times compared to the PBS, MMT, Bac, or MMT@Bac group. After confirming its metabolic efficiency, we next tested the effect of MMT@Bac@TL on the inhibition of S. typhimurium colonization and translocation. As shown in Fig. 7f, the number of S. typhimurium in the intestine declined markedly by MMT@Bac@TL. Accompanied by a decreased colonization, rare S. typhimurium was detected in the kidney and the spleen (Fig. 7g, h). Differently, the rest treatments could not effectively inhibit the colonization of S. typhimurium in the gut, and a large number of S. typhimurium (~1010 CFUs) were translocated to the kidney and the spleen. The clearance of S. typhimurium by MMT@Bac@TL further alleviated intestinal inflammation and eventually protected the gut from damage. As illustrated in Fig. 7i and Supplementary Fig. 16a, myeloperoxidase (MPO) staining indicated that treatment with MMT@Bac@TL significantly reduced neutrophil granulocyte infiltration in the colon compared to other groups. Meanwhile, severe pathological injury on the muscular layer caused by S. typhimurium infection was mitigated by MMT@Bac@TL (Fig. 7j and Supplementary Fig. 16b). The above results verified the therapeutic potential of MMT@Bac@TL in intervening enteritis, which was ascribed to the modulation of intestinal homeostasis mediated by higher levels of beneficial metabolites.

a Schedule of MMT@Bac@TL-mediated treatment of S. typhimurium-induced enteritis. At 24 hours after infection with 5 × 107 CFUs of S. typhimurium, the mice were oral administered with 0.2 ml of PBS, MMT, Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL one dose per day for 6 days, respectively. Healthy mice without any treatment were set as a control. Created in BioRender. Liu, J. (2026) https://BioRender.com/06e34rd. b, c Photographs (b) and average lengths (c) of the colon sampled from treated mice (n = 6 independent samples). d, e Level of indole (d) and lactic acid (e) in the gut (n = 5 or 6 independent samples). f–h Counts of S. typhimurium in the intestine (f), kidney (g), and spleen (h), respectively (n = 5 or 6 independent samples). i MPO staining of the colon sections. Blue arrows indicate MPO-positive cells. Scale bar, 130 μm. j H&E staining of the colon sections. Black arrows indicate the pathological changes of the muscular layer. Scale bar, 130 μm. Data are represented as means ± SD. p-values were determined using one-way ANOVA. p-values < 0.05 are shown. Source data are provided as a Source Data file.
Value of MOFS in treating colitis associated with C. albicans infection
C. albicans represented the most abundant genera in IBD patients70. It has been reported that candidalysin produced by C. albicans promotes the pathological process of IBD by inducing intestinal inflammation71. In addition to its role against pathogen infection, indole offers a viable alternative to eliminate C. albicans and its biofilms72. Lactic acid-producing bacteria are known to block C. albicans filamentation, an important virulence trait, by affecting the intestinal microenvironment43. In addition, E. faecalis was identified to secrete Fsr-dependent proteases, which inhibit hyphal morphogenesis and biofilm formation of the fungus, thereby reducing the virulence72. We hence speculated that MMT@Bac@TL could also remove pathogenic fungi in the gut during the treatment of enteritis. To clarify this, we first verified the bactericidal effect of MMT@Bac@TL on C. albicans in a rose bengal medium, the selective medium of C. albicans. After incubating C. albicans with MMT@Bac@TL at a 1:10 ratio (C. albicans to E. faecalis) for 3 hours, the CFUs of C. albicans in the MMT@Bac@TL group were reduced by 2 times compared to the medium group (Supplementary Fig. 17). The bactericidal effect of MMT@Bac@TL was increased up to 21-fold with incubation time prolonging to 24 hours. For greater relevance, we further compared the bactericidal effect of MMT@Bac@TL on pathogens with that of clinically available probiotics, including EcN and Bacillus subtilis (Bs). C. albicans were co-cultured with EcN, Bs or MMT@Bac@TL in yeast extract peptone dextrose (YPD) medium at a ratio of 1:10. The fungi cultured with medium were set as a control. After 12 hours of co-culture, samples were serially diluted, plated on YPD plates and incubated at 37 °C for colony enumeration of C. albicans. As shown in Supplementary Fig. 18a, MMT@Bac@TL exhibited comparable effects in inhibiting C. albicans to those of clinically available probiotics. Encouraged by the bactericidal effect of MMT@Bac@TL on fungi, we further evaluated the bactericidal effect of MMT@Bac@TL on S. typhimurium SL1344, one of the representative pathogenic bacteria. As shown in Supplementary Fig. 18b, Bs has a negligible effect on inhibiting the growth of S. typhimurium. The counts of S. typhimurium were ~ 3-fold and ~ 2-fold lower in EcN and MMT@Bac@TL, respectively, compared to the medium group. Considering that the anti-inflammatory efficacy of living bacteria is another factor in the therapy of colitis, in addition to antimicrobial efficacy, we next compared MMT@Bac@TL with clinically available probiotics. J774A.1 cells pre-treated with LPS (1 µg/ml) for 24 hours were subsequently incubated with 102 CFUs EcN, Bs, or MMT@Bac@TL for 24 hours. The cells treated with culture medium served as the control. As shown in Supplementary Fig. 18c, the ratio of M2 to M1 macrophages was over 2-fold increase in MMT@Bac@TL compared to other treatments, indicating a potent anti-inflammatory effect of MMT@Bac@TL.
Having confirmed the antimicrobial and anti-inflammatory effects of MMT@Bac@TL, we next explored the therapeutic value of MMT@Bac@TL in a colitis model with C. albicans infection. As shown in Fig. 8a, mice were orally administered with streptomycin (20 mg/kg) and penicillin (20000 U/kg) for 2 days to clear the intestinal microbiota and then given 3% DSS water (w/v) freely over 23 days. During this period, mice were gavaged with C. albican (1 × 108 CFUs) every other day and simultaneously treated with MMT@Bac@TL containing 1 × 107 CFUs E. faecalis, 0.05 mg MMT, 0.0013 mg tryptophan, and 0.0013 mg lactulose every other day. Healthy mice and infected mice treated with PBS, 5-aminosalicylic acid (5-ASA, 6 mg/ml, 0.1 ml per day), and equivalent MMT, Bac, MMT@Bac, MMT@Bac@T, and MMT@Bac@L were applied as controls. After 12 doses of treatment, mice were euthanized, and samples were collected for further analyses. As presented in Fig. 8b, c, the average length of the colon sampled from MMT@Bac@TL-treated mice (8.6 ± 1.1 cm) was longer than those of mice treated with PBS (7.2 ± 0.7 cm), MMT (7.1 ± 1.0 cm), Bac (7.7 ± 0.9 cm), and MMT@Bac (7.5 ± 1.1 cm). The homogenates of the cecum were prepared for plate counting of C. albicans. The CFUs of C. albicans in the PBS group were as high as 2 × 104 CFUs/g (Fig. 8d). Notably, C. albicans was substantially lowered in the entire gut after treatment with MMT@Bac@TL ( ~ 2.0 × 103 CFUs). In contrast, there were ~ 2 × 104, 3 × 104, and 2 × 104 CFUs of C. albicans detected in the MMT, Bac, and MMT@Bac groups, respectively. We further used fluorescent in situ hybridization (FISH) to visualize the localization of C. albicans in the colon. Different from other treatments, a rare fluorescent signal of C. albicans was observed in the MMT@Bac@TL group (Fig. 8e and f). Meanwhile, MMT@Bac@TL induced effective defense responses against C. albicans infection as the percentage of IgA+ B cells in the MMT@Bac@TL group was 2-fold higher than that in the PBS group (Fig. 8g). IgA+ B cells have been demonstrated to play a nonredundant role against fungal infections. In addition, treatment with MMT@Bac@TL avoided the occurrence of intestinal damage and reduced the inflammation responses. While the disruption of the muscular layer and the elicitation of inflammation were obvious in the groups of 5-ASA, MMT, and Bac compared to the PBS group (Fig. 8h–j and Supplementary Fig. 19). In brief, these results elucidated the value of MMT@Bac@TL to treat fungal infection-associated IBD.

a Experimental design for evaluating the therapeutic potential of MMT@Bac@TL in treating colitis with fungal infection. Mice were orally delivered with 0.2 ml of streptomycin (20 mg/kg) and penicillin (20000 U/kg), one dose per day for 2 days to eliminate the intestinal microbiota. Then, the mice were given 3% DSS water freely for 23 days. During this time frame, the mice were gavaged with 1 × 108 CFUs of C. albican in combination with PBS, 5-ASA (0.6 mg), MMT (0.05 mg), 1 × 107 CFUs of Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL every other day (a dose per day). After 12 doses, the mice were euthanized, and samples were collected for subsequent measurements. Created in BioRender. Liu, J. (2026) https://BioRender.com/06e34rd. b, c Photographs (b) and average lengths (c) of the colon sampled from treated mice (n = 4, 5, 6 or 7 independent samples). d Counts of C. albicans in the cecum (n = 4, 5, 6 or 7 independent samples). e, f Confocal images (e) and MFI (f) of C. albicans in colon sections stained with FISH (Red) and DAPI (Blue) (n = 3 independent samples). Scale bar, 30 μm. g Percentage of IgA+ B cells in the PPs (n = 4, 5, 6 or 7 independent samples). h Percentage of MPO-positive area measured by Image-J (n = 3 independent samples). i, j Pathological score (i) of H&E staining (j) of the colon (n = 3 independent samples). Scale bar, 75 μm. Pathological changes on the mucosa and muscular layer were indicated by black arrows. Data are represented as means ± SD. p-values were determined using one-way ANOVA. p-values < 0.05 are shown. Source data are provided as a Source Data file.
In summary, we report the design and preparation of MMT@Bac@TL that enables long-lasting in-situ synthesis of beneficial bioproducts for restoring intestinal homeostasis. MMT@Bac@TL is constructed by adsorption of E. faecalis onto the surface of MMT, which is preloaded with metabolic substrates of tryptophan and lactulose in the interlayer space through cation exchange. The water-impermeable and mucoadhesive property of MMT endows MMT@Bac@TL with high resistance and durable retention, achieving an efficient in-situ biosynthesis of beneficial metabolites of indole and lactic acid in the gut. These metabolites contribute to gut microbiota regulation, immune response balance, and intestinal barrier protection. Specifically, in an antibiotic-induced mouse model of gut dysbiosis, MMT@Bac@TL significantly inhibits the invasion of pathogenic bacteria and enhances the abundance of beneficial bacteria, recovering a normal composition and function of the gut microbiome. Meanwhile, it increases the percentages of M2 macrophages, IgA+ B cells, and mature DCs, which are conducive to intestinal homeostasis. Moreover, MMT@Bac@TL improves intestinal barrier integrity by increasing tight junction expression and mucus production. We also demonstrate the protective effects of MMT@Bac@TL in maintaining intestinal homeostasis, enabling satisfactory efficacies in treating infective enteritis and IBD with C. albicans infection. This work provides a unique platform for oral drug delivery, with an appealing characteristic of long-lasting in situ synthesis of bioproducts. Before considering for future translation, the safety, dose, and administration frequency of MMT@Bac@TL need to be systemically examined and optimized. In our future work, we will pursue the versatility of this platform to carry other metabolic substrates and microbial species, with the aim of generating diverse therapeutic agents and exploring advanced approaches for treating different diseases.
Methods
Materials and strains
L (-)-Tryptophan (TP2893390) and 5-ASA (01122621) were purchased from Shanghai Titan Scientific Co., Ltd. MMT (M916670), lactulose (L812332), ampicillin (A830931), neomycin sulfate (N799581), metronidazole (M813525), and vancomycin (V871983) were purchased from Macklin Biochemical Co., Ltd. RhB (A5102) was purchased from Tokyo Chemical Industry Co., Ltd. Antibodies including anti-mouse CD11c-FITC (117305), CD11b-PerCP Cy5.5 (101227), F4/80-phycoerythrin (PE, 123110), CD86-allophycocyanin (APC, 105011), CD86-PE/Cyanine7 (PE Cy7, 105014) CD206-PE-Cy7 (141719), CD206-Alexa Fluor® 647 (AF647, 141712), CD3-APC Cy7 (100221), CD45R/B220-PE-Cy7 (103221), and CD80-PE (104707) were purchased from Biolegend. Con A(C21402) and IgA-PE (12-5994-81) were purchased from Thermo Fisher Scientific. LPS (ST1470), DAPI (C1002), and NO detection kit (S0021S) were purchased from Beyotime Biological Technology Co. LB broth powder (A507002) was obtained from Sangon Biotech (Shanghai) Co., Ltd. E. faecalis agar (HB0268-2) and trypticase soy broth (TSB, HB4114) were purchased from Qingdao Hi-Tech Industrial Park Hope Bio-Technology Co., Ltd. E. faecalis was obtained from the American Type Culture Collections. EcN was purchased from Ningbo Bionice (China). S. typhimurium was purchased from the China General Microbiological Culture Collection Center. C. albicans was purchased from Shanghai Bioresource Collection Center. Mouse macrophage cell line J774A.1, human colon carcinoma cell line Caco-2, and DC 2.4 dendritic cell line were purchased from iCell Bioscience Inc. Mucus-producing intestinal cell line HT29-MTX-E12 was kindly provided by Wei Wu (Fudan University, Shanghai, China). Fetal bovine serum (FBS, 10099-141), penicillin-streptomycin solution (15140122), and Dulbecco’s Modified Eagle Medium (DMEM, 11965092) were purchased from Gibco. Nonessential amino acid solution (N1250) and crystal violet (G1062) were obtained from Beijing Solarbio Science & Technology Co., Ltd. HT-29-MTX-E12 cells were cultured with DMEM containing 10% FBS, 1% penicillin-streptomycin solution, and 1% nonessential amino acid. Caco-2, J774A.1, and DC 2.4 cells were cultured with DMEM containing 10% FBS and 1% penicillin-streptomycin solution. IL-4 (AF02165-A), IL-6 (AF02163-A), and IL-10 (AF02176-A) enzyme-linked immunosorbent assay (ELISA) kits were purchased from Hunan Aifang Biotechnology Co., Ltd. VEGF (EK283), IL-1β (EK201B), and TGF-β1 (EK981) ELISA kits were obtained from Multi Sciences (Lianke) Biotech Co., Ltd.
Preparation of MMT@Bac@TL
To prepare MMT loaded with tryptophan and lactulose, MMT (0.125 g), tryptophan (0.0032 g), and lactulose (0.0032 g) were suspended in 5 ml ddH2O and then incubated at 37 °C with a shaking speed of 200 rpm for 24 h. An E. faecalis colony was taken from an LB agar plate and cultured in LB liquid medium overnight (200 rpm, 37 °C). Then, E. faecalis were washed with ddH2O twice and resuspended in 980 µl PBS. Afterward, MMT loaded with tryptophan and lactulose (900 μl) and E. faecalis (100 μl) were mixed and cultured at room temperature for 4 hours with shaking at 700 rpm. Similarly, MMT@Bac, MMT@Bac@T, and MMT@Bac@L are prepared in the same manner.
Characterization of MMT@Bac@TL
The morphologies of bacteria, MMT, and MMT@Bac were measured by SEM (HITACHI Regulus 8230). For SEM measurement, samples were dropped on a silicon wafer separately after serial dehydration in 30, 50, 70, 80, 90, and 100% ethanol for 10 minutes. XRD measures were performed by Bruker X-ray diffractometer (D8 ADVANCE Da Vinci). MMT, MMT@T, MMT@L, and MMT@Bac@TL were collected by centrifugation (7000 rpm, 10 minutes) and washed 3 times with ddH2O. The samples were then frozen at − 80 °C for 24 hours, followed by freeze-drying in a lyophilizer. The dry samples were analyzed for the changes in the d001 value by XRD. Bacterial numbers were determined via making serious dilutions of E. faecalis, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL suspensions, culturing them on selective EF agar plates overnight (200 rpm, 37 °C), and bacterial counts were measured using an automated colony counting system (Interscience Scan 500). The ζ-potential of MMT, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL was determined by dynamic light scattering (DLS).
Fluorescence labeling of MMT@Bac
EDC (0.20 g, D1601), NHS (0.12 g, B0249), and RhB (0.01 g) were activated in 10 ml of ddH2O for 1 hour. Then, MMT (0.1 g) was added into the reaction solution. The mixture was stirred at room temperature for 24 hours. Finally, RhB-MMT was collected after washing with ddH2O. E. faecalis was mixed with DAPI (5 µg/ml) and incubated for 30 minutes at room temperature. Afterward, RhB-MMT (900 μl) and DAPI-labeled E. faecalis (100 μl) were mixed and cultured at room temperature for 4 hours with shaking at 700 rpm. E. faecalis and MMT were applied as controls for the above characterization. E. faecalis, MMT, and MMT@Bac were characterized by LSCM (Leica TCS SP8).
Resistance assessment in vitro
A 300 µl suspension of E. faecalis, MMT@Bac, or MMT@Bac@TL (4 × 10⁵ CFUs) was mixed with 700 µl of SGF (pH 1.2) or SIF (pH 6.8). For SGF, 1 g of pepsin (88918 F) and 384 µl of hydrochloric acid (10011061) were added to 100 ml of ddH₂O, and the pH was adjusted to 1.2. For SIF, 1.36 g of potassium dihydrogen phosphate (82821B) and 2 g of trypsin (88927B) were dissolved in 100 ml of ddH₂O. The pH was adjusted to 6.8 with 0.2 mol sodium hydroxide solution (G19852A). The mixtures were then further incubated at 37 °C with gentle shaking. At predetermined time points, 50 μl solution was withdrawn from the medium and washed with PBS. After spreading on solid LB plates, the bacteria were incubated overnight at 37 °C for counting.
Ex vivo adhesion of bacteria on the ileum
Freshly murine ileum or colon tissue was divided into ~ 2 cm segments and everted in order to expose the intestinal mucus layer. The sectioned ileum and colon segments were dripped with 10 µl of PBS containing an equal number of E. faecalis, MMT@Bac, or MMT@Bac@TL (1 × 108 CFUs/ml) for 30 minutes. At the indicated time points, the intestinal segments were flushed gently with PBS. After spreading on solid LB plates, the bacteria were incubated overnight at 37 °C for counting.
Crystal violet staining
To quantify biofilms using crystal violet, the wells were washed 3 times with 250 μl of PBS to remove the planktonic cells. Then, the samples were mixed with 300 μl of methanol for 20 minutes to fix biofilm. Subsequently, a 60 min staining process was performed using 0.1% crystal violet. The nonspecific stain was removed by washing the samples with 250 µl of ddH2O. The plates were left to dry and then subjected to an elution process with 30% acetic acid for 1 hour at room temperature. Absorbance was measured at 540 nm using a microplate reader (Tecan Infinite M200 PRO).
Congo red staining
The Congo red broth and agar assays were prepared as described elsewhere73. Briefly, TSB medium was prepared with an additional 4% sucrose (G66841D) and 0.08% Congo red dye (A600324). 20 µl of Bac, MMT@Bac, or MMT@Bac@TL (1 × 108 CFUs) was streaked onto a Congo red-containing TSB plate for the agar test. One colony of bacteria on a TSB plate was inoculated on the Congo red-containing TSB for the broth test. The colors of colonies were observed after 24- or 48-hour incubation. Brown to black colors were considered a positive sign of biofilm formation.
Effects of MMT@Bac@TL on the growth of bacteria
To evaluate the regulatory effects of MMT@Bac@TL on microorganisms, a series of in vitro experiments was conducted. To examine the effects of E. faecalis, MMT@Bac, or MMT@Bac@TL on the growth of bacteria and fungi, EcN, C. albicans, or S. typhimurium was cultured overnight at 37 °C, and bacterial quantities were determined by plate counting. EcN (2 × 105 CFUs), C. albicans (1 × 106 CFUs), or S. typhimurium (2 × 106 CFUs) was then mixed with E. faecalis, MMT@Bac or MMT@Bac@TL in a 1:1, 1:10, or 10:1 ratio (CFUs) and incubated at 37 °C with shaking. Samples were collected at 2, 4, and 6 hours or 3, 6, 12, and 24 hours, serially diluted, and spread onto selective plates.
Mucus level assay
HT29-MTX-E12 cells seeded into the 96-well plate were treated with 50 μg/ml LPS for 6 hours. Then, the cells were treated with 1 × 105 CFUs of Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL for 6 hours, respectively. For the acidic mucus assay, cells were fixed with 4% paraformaldehyde for 30 minutes and then incubated with 150 μl of AB solution (pH 2.5) for 30 minutes in the dark. Then, 300 μl of supernatant was collected and transferred into a 96-well plate and assayed at 630 nm using a microplate reader. Likewise, for the neutral mucus measurement, 100 μl of 10% Triton-100 was added into the plate and incubated at room temperature for 30 minutes. Subsequently, supernatant (15 μl) was collected and transferred into a 96-well plate. After adding 60 μl of periodic acid, the plate was incubated in the dark for 90 minutes, followed by incubation with 50 μl of PAS solution in the dark for 40 minutes. The resultant samples were analyzed at 550 nm using a microplate reader (BioTek).
Tight junction protein expression level
Expression levels of tight junction proteins were measured both in vitro and in vivo. For the cell sample, Caco-2 cells were pretreated with 50 μg/ml of LPS for 12 hours and then incubated with 1 × 104 CFUs of Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL for 7 hours, respectively. To evaluate the intestinal physical barrier, tissue sections were fixed and permeabilized prior to immunofluorescence staining. The cells and tissue sections were sequentially incubated with primary antibodies against occludin (sc-133256; Santa Cruz Biotechnology) and ZO-1 (AF5145; Affinity Biosciences), followed by species-matched secondary antibodies conjugated with fluorescent probes. Imaging was performed using LSCM (Leica TCS SP8).
In vitro immunomodulatory ability of MMT@Bac@TL
In this experiment, J774A.1 cells were pretreated with 1 μg/ml LPS for 24 hours, followed by co-incubation with 102 CFUs of E. faecalis, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL for an additional 24 hours. The supernatants were then collected to measure the cytokine levels. The cytokines were detected using ELISA kits. Subsequently, the cells were harvested and washed twice with precooled PBS for flow cytometry.
Immune response analysis
Flow cytometry was then employed to evaluate the polarization of the macrophages. The J774A.1 cells were washed and incubated with anti-CD11b, anti-F4/80, and anti-CD86 antibodies for 30 minutes at room temperature. Afterward, the cells were then stained with anti-CD206 antibodies after fixation and permeabilization with Fixation/Permeabilization Concentrate and Diluent kit (Thermo Fisher Scientific, 00-5521-00). Data were acquired using flow cytometry (BD LSRFortessa). DC 2.4 cells were treated with 1 × 104 CFUs of Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL for 18 hours, respectively. The samples were then incubated with extracellular antibodies (CD86). The samples were then washed twice with PBS and analyzed by flow cytometry (BD LSRFortessa). Data were analyzed using FlowJo (Tree Star).
In vivo survival assessment after oral ingestion
Male Balb/c mice at 6 to 8 weeks were fed 4 × 107 CFUs of Bac, MMT@Bac, or MMT@Bac@TL by oral gavage. At 4, 24, 48 or 72 hours after administration, the mice were euthanized. To evaluate the survival of the bacteria, tissues including the ileum, colon, and cecum, as well as contents, were separately collected. The samples were grounded and diluted with 1 ml of PBS. The suspensions (50 μl) were withdrawn and spread on solid E. faecalis agar plates. The bacteria were incubated overnight at 37 °C before counting.
Levels of indole and lactic acid in the gut
To analyze the content of intraluminal indole, the suspensions of the ileum, colon, and cecum were processed for the indole assay kit (MIK7664, MesGen Biotech) in accordance with the manufacturer’s instructions. Specifically, after homogenizing and centrifuging the intestinal segments, the supernatant (25 μl) was mixed with 25 μl of reaction reagent and 50 μl of PBS. The samples were then incubated at 37 °C for 25 minutes. The resultant samples were analyzed at 565 nm using a microplate reader. To analyze intestinal lactic acid levels, the samples were processed using a lactic acid assay kit (A019-2-1, Nanjing Jiancheng Bioengineering Institute) according to the manufacturer’s instructions. The tissue supernatants (1 μl) were mixed with 50 μl of enzyme working solutions and 10 μl of color reagents. The samples were then incubated at 37 °C for 10 minutes. After the reaction, the stop solution (100 μl) was added and mixed well. The absorbance was measured at 530 nm. ddH2O or 3 mmol/l standard solution was used as the negative or positive control, respectively.
Recovery of the microbiota following antibiotics
Mice were given oral gavage of a broad-spectrum antibiotic solution every 24 hours for 14 days. The broad-spectrum antibiotic solution was comprised of ampicillin (1 mg/kg), vancomycin (0.5 mg/kg), metronidazole (1 mg/kg), and neomycin sulfate (1 mg/kg). The solutions were prepared freshly every 24 hours. Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL was orally gavaged with an equivalent of 4 × 107 CFUs of E. faecalis. Colon and cecum contents of all mice were collected and stored at − 80 °C for the following experiments. The colon tissue was fixed with 4% paraformaldehyde and then used for subsequent analysis.
Analysis of the biofilm in vivo
To observe the production of E. faecalis biofilms within the intestinal tract, intestinal segments were fixed with Carnoy’s fixative, embedded in paraffin, and sectioned. The extracellular matrix of the bacteria was stained with Con A. After dewaxing and rehydration, colon tissue sections (5 μm thickness) underwent antigen retrieval treatment using 1 × All-purpose Powerful Antigen Retrieval Solution (Beyotime, P0088) under 98 °C for 20 minutes. After PBS washes, the sections were incubated with Alexa Fluor 633-conjugated Con A (20 μg/ml) for 1 hour at room temperature, then rinsed 3 times with PBS. Finally, the specimens were stained with DAPI (5 μg/ml) and sealed with an antifade mounting medium. The slides were imaged with LSCM (TCS SP8, Leica).
Biofilm formation in isolated intestinal segments
Healthy Balb/c mice were used to harvest standardized ileal segments (2 cm) from the same anatomical location. The ileal segments were then opened longitudinally and placed mucosal side up on grooved slides. We then applied 1 ml of SIF containing 1 × 107 CFUs of Bac, MMT@Bac, or MMT@Bac@TL to measure biofilm formation under an intestinal simulation environment. After 72 hours of incubation at 37 °C, the SIF was removed, and the mucosal surface was gently rinsed with PBS in 48-well plates. Tissues were then fixed in 4% paraformaldehyde for 30 minutes at room temperature and stained with Con A (20 μg/ml) and DAPI (5 μg/ml) for imaging.
Treatment of S. typhimurium-induced enteritis
Streptomycin (20 mg) was administered after the mice were deprived of water and food for 6 hours. After 7 days of adaptive feeding, mice were treated with streptomycin for 24 hours and orally inoculated with 5 × 107 CFUs of S. typhimurium. S. typhimurium was grown for 8 hours in LB supplemented with kanamycin. Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, or MMT@Bac@TL with equivalent CFUs (1.0 × 107 CFUs) was administered intragastrically for treatment on the following 6 days. Mice treated with MMT (0.05 mg) were used as a benchmark. Then, the major organs, including the intestine, kidney, and spleen, were sampled for further analysis.
Treatment of colitis with fungal infection
Balb/c mice were gavaged with 0.2 ml of water containing 20 mg/kg of streptomycin, 20000 U/kg of penicillin for 2 doses. Then, mice were given drinking water containing 3% (w/v) DSS ad libitum to induce colitis. On the same day, mice in the DSS groups were orally administered with 1 × 108 CFUs of C. albicans every other day and simultaneously treated with Bac, MMT@Bac, MMT@Bac@T, MMT@Bac@L, and MMT@Bac@TL containing 1 × 107 CFUs of E. faecalis and 0.05 mg MMT every other day from day 3 to 25. Healthy mice and infected mice treated with PBS, 5-ASA (30 mg/kg), and equivalent MMT, Bac, MMT@Bac, MMT@Bac@T, and MMT@Bac@L were applied as controls. After 12 doses of treatment, mice were euthanized, and samples were collected for further analysis. Homogenates of the cecum were prepared for C. albicans plate counting. The colon was then sampled for further study. Flow cytometric analysis was performed to quantify the frequencies of IgA+ B cells in the PPs.
FISH detection for C. albicans
To observe C. albicans colonization in the intestine, intestinal segments were fixed in 4% paraformaldehyde, paraffin-embedded, and sectioned. FISH was performed in a hybridization buffer with a fungal 28S rRNA DNA oligonucleotide that is coupled to 5 µmol Cy3 labeled probe (5’-CTCTGGCTTCACCCTATTC-3’). The tissue sections were incubated in the hybridization solution (G3045, Servicebio) at 37 °C for 1 hour for pre-hybridization. Slides were washed for 10 minutes at 37 °C in 2 ×, 1 ×, 0.5 × SSC (G3015, Servicebio). Subsequently, samples were counterstained with DAPI for 1 hour at 37 °C. C. albicans was quantified manually using the cell counter plug-in for Fiji.
Histopathological analysis
The collected intestinal tissues were fixed in 4% paraformaldehyde, paraffin-embedded, and sectioned. The obtained sections were deparaffinized and stained with H&E (C0105S, Beyotime Biotechnology) and AB-PAS (MST-8050, Fuzhou Maixin Biotechnology) according to the manufacturer’s instructions to evaluate pathological damage in the intestinal regions. To assess inflammatory infiltration in the intestine, the deparaffinized sections of intestinal samples underwent antigen retrieval using 1 × All-purpose Powerful Antigen Retrieval Solution, followed by blocking endogenous peroxidase activity with 3% H2O2. The sections were then incubated with the primary anti-MPO antibody (GB11224, Servicebio) and the corresponding secondary HRP antibody (G1215, Servicebio). Visualization was performed using a 3, 3’-diaminobenzidine (DAB) substrate kit according to the manufacturer’s instructions.
Microbiota 16S sequencing and analysis
The 16S rRNA gene sequencing and analysis were conducted at Biomarker Technologies. Total DNA was extracted from the contents of the cecum. Sequencing was performed on an Illumina HiSeq platform by targeting hypervariable V3-V4 regions of the bacterial 16S rRNA gene. Data analysis was performed on BMKCloud. Beta diversity was analyzed using PCA. Microbial composition was analyzed at the class level. LEfSe was analyzed at the phylum and species levels.
Statistical analysis
All statistical analyses were performed with GraphPad Prism 9 and IBM SPSS Statistics 27 software. Data points were excluded only when identified as statistical outliers by the standardized boxplot-based analysis using Tukey’s method. Experimental data were analyzed using one-way analysis of variance (ANOVA) with Fisher’s LSD post-test for multiple comparisons. Student’s t-test was used for the two-group comparisons. Data were expressed as means ± standard deviation (SD). p-value < 0.05 was considered statistically significant.
Ethical statement
Male Balb/c mice aged 6 to 8 weeks were purchased from Jiesijie Laboratory Animal Technology. Mice were housed under standard conditions (12-hour light/dark cycle, 22 ± 2 °C, 55 ± 10% humidity) with ad libitum access to water and standard diet. All animal experiments were carried out according to the institutional guidelines and approved by the Animal Care and Use Committee in Shanghai Yishang Biotechnology Company (IACUC-2022-Mi-203).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The 16S ribosomal RNA gene-sequencing data are available from the National Center for Biotechnology Information under accession code PRJNA1364733. The remaining data generated in this study are provided in the paper, the Supplementary Information, or the Source data. Source data are provided in this paper.
References
Neurath, M. F., Artis, D. & Becker, C. The intestinal barrier: a pivotal role in health, inflammation, and cancer. Lancet Gastroenterol. Hepatol. 10, 573–592 (2025).
Chen, Y. et al. Reinforcement of the intestinal mucosal barrier via mucus-penetrating PEGylated bacteria. Nat. Biomed. Eng. 8, 823–841 (2024).
Yang, W. & Cong, Y. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol. Immunol. 18, 866–877 (2021).
Du, L., Qi, R., Wang, J., Liu, Z. & Wu, Z. Indole-3-propionic acid, a functional metabolite of Clostridium sporogenes, promotes muscle tissue development and reduces muscle cell inflammation. Int. J. Mol. Sci. 22, 12435 (2021).
Zheng, D., Liwinski, T. & Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 30, 492–506 (2020).
Lu, Q. et al. A host-adapted auxotrophic gut symbiont induces mucosal immunodeficiency. Science 385, eadk2536 (2024).
Sosnowski, K. & Przybyłkowski, A. Ethanol-induced changes to the gut microbiome compromise the intestinal homeostasis: a review. Gut Microbes 16, 2393272 (2024).
Li, T. et al. A gut microbiota-bile acid axis promotes intestinal homeostasis upon aspirin-mediated damage. Cell Host Microbe 32, 191–208.e199 (2024).
Soranno, D. E. et al. A review of gut failure as a cause and consequence of critical illness. Crit. Care 29, 91 (2025).
Lee, Y., Kamada, N. & Moon, J. J. Oral nanomedicine for modulating immunity, intestinal barrier functions, and gut microbiome. Adv. Drug Deliv. Rev. 179, 114021 (2021).
Kotla, N. G. & Rochev, Y. IBD disease-modifying therapies: insights from emerging therapeutics. Trends Mol. Med. 29, 241–253 (2023).
Graham, W. V. et al. Intracellular MLCK1 diversion reverses barrier loss to restore mucosal homeostasis. Nat. Med. 25, 690–700 (2019).
Schwartz, D. M. et al. JAK inhibition as a therapeutic strategy for immune and inflammatory diseases. Nat. Rev. Drug Discov. 16, 843–862 (2017).
Wang, Z. et al. Synergistic role of gut-microbial L-ornithine in enhancing ustekinumab efficacy for Crohn’s disease. Cell Metab. 37, 1089–1102 (2025).
Zhu, Q. et al. Oral delivery of proteins and peptides: Challenges, status quo and future perspectives. Acta Pharm. Sin. B 11, 2416–2448 (2021).
Cao, Z. & Liu, J. Surface nanocoating of bacteria as a versatile platform to develop living therapeutics. Nat. Protoc. 19, 3162–3190 (2024).
Zhou, J. et al. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat. Commun. 13, 3432 (2022).
Lv, Y. et al. Sodium butyrate-loaded mcrospheres with enhanced bioavailability for targeted treatment of intestinal barrier injury. Adv. Healthc. Mater. 14, e2402773 (2024).
Zhu, R. et al. Inulin-based nanoparticle modulates gut microbiota and immune microenvironment for improving colorectal cancer therapy. Adv. Funct. Mater. 34, 2407685 (2024).
Jafari, M. T., Saraji, M. & Sherafatmand, H. Polypyrrole/montmorillonite nanocomposite as a new solid phase microextraction fiber combined with gas chromatography-corona discharge ion mobility spectrometry for the simultaneous determination of diazinon and fenthion organophosphorus pesticides. Anal. Chim. Acta 814, 69–78 (2014).
Millman, E., Chatterjee, A., Parker, K. M. & Catalano, J. G. Cation exchange to montmorillonite induces selective adsorption of amino acids. Geochim. Cosmochim. Acta 372, 181–195 (2024).
Sun, Y. & Lei, A. Ca2+-Facilitated adhesion of bacteria on the Na-montmorillonite surface. ACS Omega 8, 3385–3395 (2023).
Roager, H. M. & Licht, T. R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 9, 3294 (2018).
Wang, C., Chang, T., Yang, H. & Cui, M. Surface physiological changes induced by lactic acid on pathogens in consideration of pKa and pH. Food Control 46, 525–531 (2014).
Llibre, A., Kucuk, S., Gope, A., Certo, M. & Mauro, C. Lactate: A key regulator of the immune response. Immunity 58, 535–554 (2025).
Lee, Y.-S. et al. Microbiota-derived lactate accelerates intestinal stem-cell-mediated epithelial development. Cell Host Microbe 24, 833–846 (2018).
Zhu, P. et al. Fabrication of hyaluronic acid-inulin coated Enterococcus faecium for colon-targeted delivery to fight Fusobacterium nucleatum. Carbohydr. Polym 329, 121797 (2024).
Caillard, R. & Lapointe, N. In vitro gastric survival of commercially available probiotic strains and oral dosage forms. Int. J. Pharm. 519, 125–127 (2017).
Zmora, N. et al. Personalized gut mucosal colonization resistance to empiric probiotics is associated with unique host and microbiome features. Cell 174, 1388–1405.e1321 (2018).
Huang, B. et al. Inflammation-oriented montmorillonite adjuvant enhanced oral delivery of anti-TNF-α nanobody against inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 121, e2320482121 (2024).
Lin, Y. et al. A soil-inspired dynamically responsive chemical system for microbial modulation. Nat. Chem. 15, 119–128 (2023).
Pelaseyed, T. et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol. Rev. 260, 8–20 (2014).
Lee, J., Menon, N. V., Truong, H. D. & Lim, C. T. Dynamics of spatial organization of bacterial communities in a tunable flow gut microbiome-on-a-chip. Small 21, 2410258 (2025).
Gong, F., Ito, K., Nakamura, Y. & Yanofsky, C. The mechanism of tryptophan induction of tryptophanase operon expression: tryptophan inhibits release factor-mediated cleavage of TnaC-peptidyl-tRNA(Pro). Proc. Natl. Acad. Sci. USA 98, 8997–9001 (2001).
Kaur, N. & Kishore, D. Montmorillonite: An efficient, heterogeneous and green catalyst for organic synthesis. J. Chem. Pharm. Res. 4, 991–1015 (2012).
Tolonen, A. C. et al. Synthetic glycans control gut microbiome structure and mitigate colitis in mice. Nat. Commun. 13, 1244 (2022).
Zarkan, A., Liu, J., Matuszewska, M., Gaimster, H. & Summers, D. K. Local and universal action: the paradoxes of indole signalling in bacteria. Trends Microbiol. 28, 566–577 (2020).
Zhang, Y., Cai, Y., Zhang, B. & Zhang, Y.-H. P. J. Spatially structured exchange of metabolites enhances bacterial survival and resilience in biofilms. Nat. Commun. 15, 7575 (2024).
Malviya, J. et al. Metabolomic profiling of bacterial biofilm: trends, challenges, and an emerging antibiofilm target. World J. Microb. Biot. 39, 212 (2023).
Jandl, B., Dighe, S., Gasche, C., Makristathis, A. & Muttenthaler, M. Intestinal biofilms: pathophysiological relevance, host defense, and therapeutic opportunities. Clin. Microbiol. Rev. 37, e0013323 (2024).
Ma, W. et al. Bacillus subtilis biofilm development in the presence of soil clay minerals and iron oxides. NPJ Biofilms Microbiomes 3, 4 (2017).
Lu, H., Que, Y., Wu, X., Guan, T. & Guo, H. Metabolomics deciphered metabolic reprogramming Required for Biofilm Formation. Sci. Rep. 9, 13160 (2019).
Wang, F., Wang, Z. & Tang, J. The interactions of Candida albicans with gut bacteria: a new strategy to prevent and treat invasive intestinal candidiasis. Gut Pathogens 15, 30 (2023).
Zhang, Z. J., Pedicord, V. A., Peng, T. & Hang, H. C. Site-specific acylation of a bacterial virulence regulator attenuates infection. Nat. Chem. Biol. 16, 95–103 (2020).
Vega, N. M., Allison, K. R., Samuels, A. N., Klempner, M. S. & Collins, J. J. Salmonella typhimurium intercepts Escherichia coli signaling to enhance antibiotic tolerance. Proc. Natl. Acad. Sci. USA 110, 14420–14425 (2013).
Zhu, J. et al. Mechanisms of probiotic Bacillus against enteric bacterial infections. One Health Adv. 1, 21 (2023).
Eshleman, E. M. et al. Microbiota-derived butyrate restricts tuft cell differentiation via histone deacetylase 3 to modulate intestinal type 2 immunity. Immunity 57, 319–332 (2024).
Gallardo-Becerra, L., Cervantes-Echeverría, M., Cornejo-Granados, F., Vazquez-Morado, L. E. & Ochoa-Leyva, A. Perspectives in searching antimicrobial peptides (AMPs) produced by the microbiota. Microb. Ecol. 87, 8 (2023).
Tomlinson, K. L. et al. Staphylococcus aureus stimulates neutrophil itaconate production that suppresses the oxidative burst. Cell Reports 42, 112064 (2023).
Keskey, R. C. et al. Enterobactin inhibits microbiota-dependent activation of AhR to promote bacterial sepsis in mice. Nat. Microbiol. 10, 388–404 (2025).
Jung, Y. et al. IL-1β in eosinophil-mediated small intestinal homeostasis and IgA production. Mucosal Immunol. 8, 930–942 (2015).
Johnson, K. E. & Wilgus, T. A. Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair. Adv. Wound Care 3, 647–661 (2014).
Chen, L. et al. TGFB1 induces fetal reprogramming and enhances intestinal regeneration. Cell Stem Cell 30, 1520–1537.e1528 (2023).
Branchett, W. J., Saraiva, M. & O’Garra, A. Regulation of inflammation by Interleukin-10 in the intestinal and respiratory mucosa. Curr. Opin. Immunol. 91, 102495 (2024).
Hegarty, L. M., Jones, G.-R. & Bain, C. C. Macrophages in intestinal homeostasis and inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 20, 538–553 (2023).
Mudter, J. & Neurath, M. F. Il-6 signaling in inflammatory bowel disease: pathophysiological role and clinical relevance. Inflamm. Bowel Dis. 13, 1016–1023 (2007).
Muro, P. et al. The emerging role of oxidative stress in inflammatory bowel disease. Front. Endocrinol. 15, 1390351 (2024).
Zheng, T. et al. Protein kinase p38α signaling in dendritic cells regulates colon inflammation and tumorigenesis. Proc. Natl. Acad. Sci. USA 115, E12313–e12322 (2018).
Tezuka, H. & Ohteki, T. Regulation of IgA production by intestinal dendritic cells and related cells. Front. Immunol. 10, 1891 (2019).
Lin, S. et al. Mucosal immunity–mediated modulation of the gut microbiome by oral delivery of probiotics into Peyer’s patches. Sci. Adv. 7, eabf0677 (2021).
Chen, Z. et al. Foxo1 controls gut homeostasis and commensalism by regulating mucus secretion. J. Exp. Med. 218, e20210324 (2021).
Jiang, H., Chen, C. & Gao, J. Extensive summary of the important roles of indole propionic acid, a gut microbial metabolite in host health and disease. Nutrients 15, https://doi.org/10.3390/nu15010151 (2022).
Matsuzawa, M. et al. The protective role of conjunctival goblet cell mucin sialylation. Nat. Commun. 14, 1417 (2023).
Zhao, T. et al. Impact of structurally diverse polysaccharides on colonic mucin O-glycosylation and gut microbiota. NPJ Biofilms Microbiomes 9, 97 (2023).
Kayama, H., Okumura, R. & Takeda, K. Interaction Between the Microbiota, Epithelia, and Immune Cells in the Intestine. Annu. Rev. Immunol. 38, 23–48 (2020).
Yu, D. et al. MLCK-dependent exchange and actin binding region-dependent anchoring of ZO-1 regulate tight junction barrier function. Proc. Natl. Acad. Sci. USA 107, 8237–8241 (2010).
He, W.-Q. et al. Contributions of myosin light chain kinase to regulation of epithelial paracellular permeability and mucosal homeostasis. Int. J. Mol. Sci. 21, 993 (2020).
Marchiando, A. M. et al. Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. J. Cell Biol. 189, 111–126 (2010).
Fatale, S. et al. Montmorillonite: An advanced material with diverse pharmaceutical and medicinal applications. Ann. Pharm. Fr. 83, 588–604 (2025).
Iliev, I. D. & Cadwell, K. Effects of intestinal fungi and viruses on immune responses and inflammatory bowel diseases. Gastroenterology 160, 1050–1066 (2021).
Li, X. V. et al. Immune regulation by fungal strain diversity in inflammatory bowel disease. Nature 603, 672–678 (2022).
Oh, S., Go, G. W., Mylonakis, E. & Kim, Y. The bacterial signalling molecule indole attenuates the virulence of the fungal pathogen Candida albicans. J. Appl. Microbiol. 113, 622–628 (2012).
Tursi, S. A. et al. Salmonella Typhimurium biofilm disruption by a human antibody that binds a pan-amyloid epitope on curli. Nat. Commun. 11, 1007 (2020).
Acknowledgements
This work was financially supported by the National Key Research and Development Program of China (2021YFA0909400, J.L.) and the National Natural Science Foundation of China (22425505, J.L.; 22575151, S.L.; 22505158, Z.F.; 22305152, H.C.; 32301099, L.W.).
Author information
Authors and Affiliations
Contributions
J.L. supervised the project. J.L., S.L., S.X., and L.W. conceived and designed the experiments with Y.Z. and Z.F. Y.Z., Z.F., S.L., J.C., Y.C., H.C., and M.Z. performed all experiments. All authors analyzed and discussed the data. Y.Z., S.L., and J.L. wrote the paper. Y.P. discussed and edited the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Qian Chen, and the other anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zhang, Y., Fu, Z., Chen, Y. et al. A montmorillonite-based oral fermentation system enables long-lasting in-situ biosynthesis to restore intestinal homeostasis. Nat Commun 17, 2418 (2026). https://doi.org/10.1038/s41467-026-69071-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-026-69071-2
