
Abstract
Parkinson’s disease (PD) is a complex neurodegenerative disorder with growing evidence suggests peripheral immunity plays a role in its pathogenesis. However, the specific peripheral immune cell types and gene expression profiles associated with PD remain unclear. In this study, we integrated single-cell expression quantitative trait loci (sc-eQTL) data from 14 immune cell types in the OneK1K cohort with large-scale genome-wide association study (GWAS) data for PD. Using Mendelian randomization (MR) and Bayesian colocalization analyses, we identified 28 immune-cell-specific eGenes with significant associations to PD risk, among which 24 showed strong or moderate evidence of shared genetic signals. Notable candidates included FDFT1, ARSA, CTSB, and HLA-DQA1, each displaying cell-type-specific associations in CD4+ T cells, CD8+ T cells, B cells, and monocytes. Replication using an independent sc-eQTL dataset from the DICE project confirmed consistent findings for several eGenes. Additional validation through peripheral blood single-cell RNA sequencing (scRNA-seq) revealed distinct expression patterns and significant changes in PD patients. Phenome-wide association studies (PheWAS) showed multiple associations with immune-related traits and minimal associations with unrelated traits, indicating a favorable safety profile for therapeutic targeting. Drug repurposing analysis identified several candidate compounds, including felodipine, amodiaquine, alprazolam, and tetrandrine, some of which are predicted to cross the blood-brain barrier. Molecular docking simulations further supported strong binding interactions between these compounds and PD-associated targets such as CTSB and ARSA. This integrative approach highlights key immune-cell-specific genes involved in PD and proposes several repurposable drugs with central nervous system potential, paving the way for more targeted therapeutic strategies.
Similar content being viewed by others


Introduction
Parkinson’s disease (PD) is a common neurodegenerative disorder characterized by motor symptoms such as tremor, rigidity, and bradykinesia, as well as non-motor features like cognitive decline and mood disturbances1. It affects approximately 1% of the population over 60 years old and poses a growing global health burden due to aging populations2,3.
Although the etiology of PD remains incompletely understood, considerable progress has been made in identifying therapeutic targets4,5. Current treatments, including dopamine replacement therapy (e.g., levodopa), MAO-B inhibitors, and deep brain stimulation, primarily focus on symptom management5. However, these approaches do not halt or reverse disease progression, underscoring the need for novel drug targets and disease-modifying strategies.
Emerging evidence suggests a critical role of the immune system in PD pathogenesis6,7,8. Postmortem studies reveal activated microglia and T cell infiltration in PD brains9. Moreover, elevated levels of peripheral cytokines and altered immune cell profiles have been observed in PD patients10. Immunomodulatory agents are now under investigation, and genetic studies have implicated immune-related loci in PD risk11,12. Despite these insights, the specific immune cell types and molecular mechanisms contributing to PD remain poorly defined. Understanding immune-mediated regulatory pathways may reveal novel therapeutic targets and enable more precise interventions with fewer off-target effects.
Human genetics has increasingly been used to inform drug development by identifying genes causally linked to disease, thereby improving the likelihood of clinical trial success13. Genome-wide association studies (GWAS) have uncovered over 90 independent loci associated with PD risk, implicating biological pathways such as lysosomal function, endocytosis, and synaptic signaling14,15,16,17. However, pinpointing the effector genes and relevant tissues or cell types remains challenging due to the complexity of linkage disequilibrium (LD) and regulatory mechanisms. Mendelian randomization (MR), which leverages genetic variants as instrumental variables (IVs), enables causal inference between gene expression and disease18. Several studies have applied genetic approaches to identify druggable PD targets from GWAS, including polygenic priority score (PoPS)-based prioritization, transcriptomics integrated with MR, multi-omics aging analyses, and large-scale MR of druggable genes proposing repurposing opportunities19,20,21,22. While these studies have advanced drug target prioritization, most relied on bulk-tissue or protein-level QTLs, which may obscure cell-type-specific effects crucial for understanding disease mechanisms and developing precise interventions.
Recently, single-cell eQTL (sc-eQTL) datasets have enabled unprecedented resolution of gene regulatory architecture across distinct immune cell subsets23,24. This provides a powerful opportunity to refine causal gene prioritization and therapeutic targeting in complex diseases like PD.
In this study, we aimed to systematically identify immune-cell-specific gene expression programs that causally influence PD risk using single-cell transcriptome-wide Mendelian randomization (scTWMR) and Bayesian colocalization analysis. By integrating sc-eQTL data with large-scale GWAS of PD, we sought to uncover immune-mediated regulatory mechanisms and evaluate their potential as drug targets. We further prioritized genes based on pleiotropy profiles using phenome-wide association studies (PheWAS) and predicted candidate compounds through drug signature enrichment and molecular docking. This integrative approach provides novel insights into immune contributions to PD and lays the groundwork for precision drug discovery targeting specific immune pathways (Fig. 1).

a Summary of input data including sc-eQTL data from 14 immune cell types (OneK1K cohort) and GWAS summary statistics of PD. b Primary analyses including two-sample MR to infer causal associations, Bayesian colocalization to detect shared causal variants, and scRNA-seq of peripheral immunological features. c Follow-up analyses including PheWAS for pleiotropic effects, drug prediction from DSigDB, and molecular docking simulations.
Results
Selection of genetic instruments for immune cell-specific eGenes
To construct a reliable set of genetic instruments for MR, we implemented a rigorous filtering pipeline using cis-expression quantitative trait loci (cis-eQTL) data from diverse immune cell types. Initially, 26,597 eGenes were detected across multiple immune subsets. To ensure independence among genetic variants and reduce LD effects, we applied a stringent clumping threshold (r2 < 0.001), which narrowed the set to 8733 distinct eGenes. The relative distribution of these genes among the 14 immune cell types remained largely unchanged after clumping, with CD4 naïve/central memory T cells (CD4 NC) and natural killer cells (NK) comprising the largest shares. Sample sizes across cell types ranged from 643 to 982 individuals, with the largest cohorts observed for CD4 NC, CD8 naïve and central memory T cells (CD8 NC), CD4+ T cells with an effector memory or central memory phenotype (CD4 ET), CD8+ T cells with an effector memory or central memory phenotype (CD8 ET), as well as immature and naïve B cells (B IN) and memory B cells (B Mem). Other innate immune populations, including classical monocytes (Mono C, n = 969), DC (n = 968), natural killer recruiting cells (NK R, n = 967), nonclassical monocytes (Mono NC, n = 932), and NK (n = 928), also had robust sample coverage. CD8+ T cells with expression of S100B (CD8 S100B, n = 981) were included as a functionally specialized cytotoxic T cell subset, while CD4+ T cells expressing SOX4 (CD4 SOX4) and plasma cells (Plasma) had smaller sample sizes of 857 and 643, respectively. To minimize weak-instrument bias, we retained instruments with an F-statistic > 10. Most eGenes were linked to a single instrument, while a subset of eGenes had multiple independent SNPs, providing robust instruments for causal inference in the MR framework (Fig. 2).

Distribution of immune cell-specific eGenes based on instrument count.
Immune-cell type specific effects of gene expression on PD risk
Detailed information on the instruments used in the MR analysis is summarized in Supplementary Table S1. MR analysis revealed 28 eGenes with distinct immune-cell-type-specific gene expression effects on PD after false discovery rate (FDR) correction (Fig. 3, Supplementary Table S2). Notably, FDFT1 showed consistent associations across multiple cell types, including CD4+ and CD8+ T cell subsets as well as B Mem, highlighting its broad regulatory role. eGenes such as HLA-DQA1, HLA-DQA2, and CTSB were also significantly linked to PD in CD8+ T cells, NK, and Mono, suggesting immune-related and lysosomal pathways in PD pathogenesis. DGKQ displayed the strongest association specifically in NK, indicating a novel and highly cell-specific risk mechanism. Other notable eGenes like ARSA, FAHD1, and KRTCAP3 exhibited significant associations in CD4+ or CD8+ T cell subsets, reinforcing the importance of adaptive immune cell regulation. Overall, these findings support the hypothesis that PD susceptibility is mediated through immune-cell-specific gene expression patterns.

Each dot represents an eGene-cell type pair tested for causal association with PD risk. Red points indicate associations that passed the FDR threshold. Notable eGenes with significant associations across specific cell types are annotated.
Colocalization analysis prioritizes immune cell-specific PD-associated eGenes
Bayesian colocalization analysis identified a set of 24 immune-cell-specific eGenes with evidence of shared causal variants for both gene expression and PD risk (Fig. 4, Supplementary Table S3). We applied a two-tier confidence framework: strong evidence (PP.H4 > 80%) and moderate evidence (50% < PP.H4 ≤ 80%). Only strong-evidence eGenes were advanced for downstream functional investigation and therapeutic prioritization, whereas moderate-evidence eGenes were reported separately and interpreted with caution. Among the strong-evidence set, FDFT1 stood out with robust colocalization across six immune cell types (CD4 NC, CD4 ET, CD8 NC, CD8 ET, CD8 S100B, and B Mem), supported by high PP.H4 values and consistent risk effects. Other notable eGenes included SPNS1, KRTCAP3, and ZSWIM7, each showing distinct cell-type-specific associations, as well as HLA-DQA1 with moderate evidence. The chord diagram illustrates the diversity of eGene-cell-PD connections, highlighting both common and unique patterns (Supplementary Fig. S1). Overall, CD4 NC and CD8 NC contributed the largest number of prioritized eGenes, underscoring their central role in the immune-mediated genetic architecture of PD (Table 1).

Colocalization strength is indicated by color: red bars represent eGenes of tier 1 with strong evidence (PP.H4 > 80%), and blue bars indicate eGenes of tier 2 with moderate evidence (50% < PP.H4 ≤ 80%).
Replication of prioritized eGenes in an independent sc-eQTL dataset
To assess reproducibility, we conducted a replication analysis using the DICE project. Due to SNP availability and cell-type specificity constraints, MR analysis was feasible for only four eGenes: KRTCAP3 in CD8+ naïve T cells, and ZSWIM7, FAHD1, and NEIL2 in CD4+ naïve T cells. Of these, all except NEIL2 demonstrated significant associations with PD and effect directions consistent with those observed in the primary analysis (Supplementary Fig. S2).
Peripheral blood scRNA-seq evidence for prioritized eGenes in PD
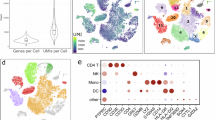
To further evaluate the disease relevance of our prioritized eGenes, we analyzed an independent peripheral blood single-cell RNA sequencing (scRNA-seq) dataset, which profiled six individuals, includung two healthy controls (HC), two early-stage PD patients, and two late-stage PD patients (Fig. 5A). After quality control, 58,808 cells were retained and clustered into nine major immune cell types (Fig. 5B, C), with CD8+ T cells, CD4+ T cells, and NK comprising the largest proportions in both PD and HC groups (Fig. 5D). Several of our genetically prioritized eGenes showed robust expression in distinct immune cell populations (Fig. 5E). Notably, HLA-DQA1 was predominantly expressed in B IN and B Mem, while CTSB was highly expressed in Mono C and Mono NC, as well as DC (Fig. 5F). Differential expression analysis revealed cell-type-specific transcriptional alterations in PD (Fig. 5G, Supplementary Table S4). Statistical testing identified significant expression changes for HLA-DQA1, DDRGK1, and ZNF391 in CD8+ T cells, and for KRTCAP3 in CD4+ T cells, with effect directions consistent with our primary genetic findings (Fig. 5H).

A UMAP projection of all cells from six samples, colored by sample identity. B UMAP projection colored by unsupervised clustering results. C Annotation of nine major immune cell types based on canonical marker genes. D Proportion of each major immune cell type in HC and PD groups. E Dot plot showing the expression levels and detection rates of genetically prioritized eGenes across major immune cell types. F Representative UMAP feature plots illustrating cell-type-specific expression patterns of HLA-DQA1 and CTSB. G Proportion of significantly upregulated and downregulated genes per immune cell type. H Cell-type-specific differential expression of prioritized eGenes.
Phenome-wide association analysis of prioritized eGenes and IVs
To assess the broader phenotypic spectrum, potential immune relevance, and safety profile of our top-prioritized targets, we performed PheWAS at both the gene and SNP levels. At the gene level, we examined the prioritized eGenes with colocalization evidence using the AstraZeneca PheWAS Portal. Most eGenes showed no significant associations at the genome-wide level, suggesting a low risk of widespread off-target effects. Notable exceptions included ARSA, which was linked to several continuous nervous system traits spanning neurological and ocular categories; NEIL2, associated with circulatory system traits; and SPNS1, which exhibited a significant binary trait association with abnormal clinical and laboratory findings (Fig. 6, Supplementary Table S5–7). At the SNP level, we queried the IVs used for MR analyses in the Open Targets Platform and FinnGen. Multiple immune-related traits were significantly associated with the IVs, including lymphocyte count, C-reactive protein (CRP) levels, autoimmune thyroiditis (AIT), rheumatoid arthritis (RA), and spondyloarthritis (SpA) (Fig. 7, Supplementary Table S8–9). Additionally, we observed significant associations with several non-immune-related traits of potential clinical or mechanistic relevance to PD, such as type 2 diabetes (T2D), obesity, gout, and senile cataract (Supplementary Table S10–11).

A ARSA. B NEIL2. C SPNS1. Dot color indicates phenotype category; triangle direction reflects effect size direction.

Sankey plots illustrating significant immune-related phenotypic associations for SNPs corresponding to prioritized eGenes. A Direct immune measurements, including lymphocyte count and measurement of C-reactive protein (CRP), cathepsin B (CTSB), leukocyte immunoglobulin-like receptor subfamily B member 4 (LILRB4), and collectin-12 (CL-12). B Immune-related diseases, including anti-citrullinated protein antibody-positive rheumatoid arthritis (ACPA + RA), polyarthropathies, seropositive rheumatoid arthritis (SPRA), coeliac disease (CD), ankylosing spondylitis (AS), subacute thyroiditis (SAT), spondyloarthritis (SpA), thyroiditis, psoriasis vulgaris (PV), juvenile arthritis (JIA). Cell types represent the source of the eGene, with links to the corresponding SNP and associated traits. *measurement. #count.
Drug repurposing opportunities for immune cell-specific PD targets
Drug enrichment analysis using the Drug Signatures Database (DSigDB) database via the Enrichr platform identified several promising compounds targeting prioritized PD-associated eGenes, with potential for therapeutic repurposing (Table 2, Supplementary Table S12). High-ranking candidates included leupeptin, DNQX, and β-solamarine, all showing strong enrichment for CTSB and ARSA, with high combined scores exceeding 900, suggesting potent target engagement. Notably, FDFT1 was targeted by drugs like mitoxantrone, tetrandrine, and pravastatin, the latter already being an approved cholesterol-lowering agent, thus representing a viable repositioning candidate. CTSB, a lysosomal protease linked to PD pathology, was targeted by multiple compounds with diverse indications such as amodiaquine (antimalarial), trifluridine (antiviral), and alprazolam (anxiolytic), while ARSA was enriched for several drug classes including receptor antagonists and opioid analgesics. To further assess their translational potential, we evaluated the predicted blood-brain barrier (BBB) permeability of all candidate compounds using the SwissADME platform (Supplementary Fig. S4-7). Several agents, including amodiaquine, alprazolam, methadone hydrochloride, and felodipine, were predicted to be BBB-permeant, indicating potential central nervous system (CNS) activity, whereas others are more likely to act peripherally. This distinction provides pharmacokinetic context for prioritizing drugs in future CNS-targeted versus peripheral immune modulation strategies.
Molecular docking supports drug-target binding interactions
To further evaluate the binding interactions and druggability of prioritized compounds, molecular docking simulations were conducted for all BBB-permeant candidates and the top six drug-target pairs identified in the previous analysis (Supplementary Table S14). Among the BBB-permeant ligands, felodipine (CTSB, -6.61 kcal/mol) and amodiaquine (CTSB, −6.27 kcal/mol) exhibited the most favorable binding energies, followed by alprazolam (CTSB, -5.80 kcal/mol) and methadone hydrochloride (ARSA, −5.68 kcal/mol) (Fig. 8). Notably, several non-BBB-permeant compounds also demonstrated strong predicted binding, including β-solamarine (CTSB, −7.56 kcal/mol) and gambierol (FAHD1, −8.23 kcal/mol), suggesting potential utility in peripheral modulation strategies. Other non-permeant candidates, such as leupeptin (CTSB, −6.34 kcal/mol) and DNQX (ARSA, −5.25 kcal/mol), showed moderate binding energies (Supplementary Fig. S8).

A Amodiaquine and CTSB. B Methadone hydrochloride and ARSA. C Alprazolam and CTSB. D Felodipine and CTSB. Left: protein-ligand complex; center: close-up of binding site; right: 2D interaction map highlighting key residues and binding types.
Discussion
PD is a progressive neurodegenerative disorder that imposes a substantial burden on individuals and healthcare systems worldwide4. With its prevalence rising sharply in aging populations, PD affects approximately 1% of individuals over the age of 60, yet current therapies primarily address symptoms without altering disease progression2,3. Despite advances in genetic studies and the identification of over 90 PD-associated loci, translating these findings into precise, cell-type-specific therapeutic targets remains a major challenge25. Particularly, the role of immune dysregulation in PD pathogenesis has gained increasing attention, but the specific immune cell subtypes and gene regulatory mechanisms involved are not fully understood. In this study, we integrated scTWMR with Bayesian colocalization to systematically identify immune cell-specific genes with potential causal links to PD. We prioritized key eGenes, including FDFT1, ARSA, CTSB, and HLA-DQA1, and further validated their expression patterns through scRNA-seq analysis. Using drug repurposing approaches, we identified promising compounds such as felodipine, alprazolam, and tetrandrine, several of which are predicted to cross the BBB. Molecular docking simulations supported strong binding interactions between these compounds and their corresponding targets. Collectively, these findings highlight a set of druggable immune-related targets in PD and provide a foundation for the following discussion on their biological functions, immune cell specificity, and therapeutic relevance.
Among the eGenes prioritized through our scTWMR and colocalization analysis, FDFT1 emerged as a robust candidate with significant protective associations across six immune cell subsets, including CD8 NC, CD8 ET, CD8 S100B, CD4 ET, CD4 NC, and B Mem. FDFT1 (Farnesyl-diphosphate farnesyltransferase 1) encodes a rate-limiting enzyme in the cholesterol biosynthesis pathway, catalyzing the formation of squalene26. Cholesterol metabolism is closely linked to neuronal membrane structure, synaptic plasticity, and neuroinflammation. Abnormal or excessive cholesterol synthesis can trigger lipid peroxidation and mitochondrial dysfunction, thereby promoting α-synuclein aggregation and dopaminergic neuron degeneration27. Disrupted cholesterol homeostasis may alter lipid raft composition, affecting immune receptor signaling and activation states. Thus, dysregulation of FDFT1 in immune cells could exacerbate PD pathology through immune-metabolic coupling mechanisms.
HLA-DQA1, part of the major histocompatibility complex (MHC) class II, plays a pivotal role in antigen presentation to CD4+ T cells28. Its expression was significantly associated with decreased PD risk in CD8 NC and CD8 ET, while showing a risk effect in NK. This dual-directional association indicates cell-type-specific regulatory mechanisms. Given that HLA class II variants have been recurrently implicated in GWAS of PD and in autoimmune disorders such as type 1 diabetes (T1D) and multiple sclerosis (MS), the involvement of HLA-DQA1 suggests potential shared mechanisms between PD and autoimmunity29,30,31. It is plausible that altered expression of HLA-DQA1 in cytotoxic CD8+ T cells might reduce aberrant antigen-driven inflammation, thereby attenuating neurodegeneration. Conversely, its elevated expression in NK may enhance interactions with other immune cells, increase recognition of stressed or damaged neurons, and amplify cytotoxic responses, ultimately contributing to sustained neuroinflammation and neuronal loss.
KRTCAP3 (Keratinocyte-associated protein 3) remains poorly characterized but has been implicated in cell structural stability and metabolic regulation32. Interestingly, our results showed divergent effects depending on the immune cell type: increased expression in CD4 ET was associated with lower PD risk, while higher risk was associated with expression in CD8 NC and CD4 NC. Although the functional role of KRTCAP3 remains unclear, emerging evidence suggests a potential role for KRTCAP3 in energy metabolism and lipid homeostasis32,33. Aberrant expression in T cell subsets could influence stress responses and immune competence. Moreover, genetic links between KRTCAP3 and obesity/metabolic syndrome, both recognized PD risk factors, support its possible involvement in PD pathogenesis through immune-metabolic pathways34.
ZSWIM7 (Zinc finger SWIM-type containing 7) is an important regulator in the DNA homologous recombination repair pathway, essential for meiosis and double-strand break repair35. Previous genetic studies have identified ZSWIM7 as a susceptibility genes for PD in brain tissues36,37. Our findings revealed significant associations with PD in CD4 NC, CD8 NC, and CD8 ET. Impaired DNA damage response and repair mechanisms are widely observed in neurodegenerative diseases38. In immune cells, such defects can lead to apoptosis, altered immune responses, and the establishment of chronic inflammatory states. The immune cell-specific role of ZSWIM7 may involve maintaining genomic stability, thus influencing immune cell lifespan and functionality, and indirectly promoting neuroinflammation in PD.
SPNS1 (Spinster homolog 1) encodes a lysosome-associated transmembrane transporter involved in lipid transport and autophagy39. Previous experimental models have shown that SPNS1 deficiency causes lysosomal dysfunction and neurodegenerative phenotypes40. Defective autophagy is a hallmark of PD pathology, with lysosomal impairment contributing to α-synuclein accumulation and dopaminergic neuron loss41. Our findings consistently revealed its negatively association with PD risk in CD8 NC and CD8 ET, suggesting a potential protective role. In immune cells, SPNS1 likely facilitates autophagic clearance of damaged mitochondria and aggregated proteins. In CD8+ T cells specifically, proper SPNS1 function may sustain energy metabolism and stress adaptability, dampening chronic inflammation and reducing CNS-targeted immune damage.
Beyond these core eGenes, we also identified several additional candidates with cell type-specific PD associations. ARSA (Arylsulfatase A), associated with PD in CD4 NC, encodes a lysosomal enzyme involved in sphingolipid metabolism and α-synuclein clearance42. DDRGK1 in CD8 NC may modulate endoplasmic reticulum stress responses and UFMylation, thereby influencing immune homeostasis43. NEIL2 (CD4 NC), a DNA glycosylase, participates in oxidative damage repair and may protect immune cell genomic integrity under chronic inflammation44. ZNF391 (CD8 ET) and FAM86B1 (CD8 S100B), though poorly characterized, show immune cell-specific associations that point to potential roles in underexplored regulatory pathways. FAHD1 (CD4 NC), a mitochondrial enzyme, could influence immune cell activation by regulation of the tricarboxylic acid (TCA) cycle and energy metabolism45.
Based on our drug repurposing, several promising small-molecule ligands targeting prioritized PD-associated eGenes were identified, with distinct pharmacological classes and mechanistic links to PD pathology. Four candidate drugs were predicted to be BBB-permeant, making them suitable for direct CNS-targeted strategies. Among these, amodiaquine, alprazolam, and felodipine are all predicted to be CTSB-targeting agents, positioning them as promising candidates for mechanism-based therapy in PD. Activation of CTSB has been shown to enhance lysosomal function, stimulate glucocerebrosidase activity, and promote clearance of fibrillar α-synuclein in dopaminergic neurons, thereby mitigating key pathogenic processes in PD46. Amodiaquine, initially identified as a weak Nurr1 modulator47,48, may confer dual neuroprotective effects by engaging both lysosomal degradation pathways and Nurr1-mediated transcriptional programs. Alprazolam, in addition to its potential CTSB-modulating properties, has demonstrated clinical efficacy in reducing anxiety-triggered freezing of gait and improving gait stability in PD, suggesting combined disease-modifying and symptomatic benefits49,50. Felodipine has robust epidemiological and preclinical support, including reduced PD risk in a nationwide cohort51, induction of brain autophagy and α-synuclein clearance at clinically relevant concentrations52, and restoration of dopamine synthesis alongside mitigation of oxidative stress, neuroinflammation, mitochondrial dysfunction, and motor impairment53,54. Methadone hydrochloride, an ARSA-targeting agent, does not bind to α-synuclein or induce its pathogenic compaction55, suggesting a low risk of directly exacerbating α-synuclein aggregation. Combined with ARSA’s role in lipid metabolism and lysosomal function, methadone may offer a safe pharmacological avenue for modulating neurodegenerative pathways in PD. Together, these BBB-permeant candidates represent compelling mechanistically driven options for PD therapy, with the potential to target core pathogenic pathways while offering both disease-modifying and symptomatic benefits.
Given the emerging recognition that PD pathogenesis involves not only CNS-specific mechanisms but also peripheral immune dysregulation6,56, particularly after BBB disruption facilitates peripheral immune cell infiltration into the brain57,58, compounds with poor BBB permeability may still hold substantial therapeutic relevance. Our study identified several candidate agents with limited CNS penetration, which could modulate peripheral immune responses, potentially influencing neuroinflammation and PD progression. CTSB-targeting ligands such as leupeptin and β-solamarine could alter lysosomal activity in peripheral immune cells, thereby affecting antigen processing and inflammatory signaling59,60. Similarly, ARSA-targeting compounds (DNQX, 4-amino-1,8-naphthalimide, 3,5-dinitrocatechol) may influence sphingolipid metabolism in immune cell membranes, potentially impacting immune activation states42. Gambierol, a potent voltage-gated potassium channel inhibitor61, may exert peripheral neuroprotective effects in PD through FAHD1-mediated mitochondrial regulation and modulation of immune-related signaling. Thus, although these agents lack robust BBB permeability, their capacity to reprogram peripheral immune function offers a plausible complementary or alternative route for PD intervention, especially in early or peripheral inflammation-driven stages of disease.
This study presents several key innovations that distinguish it from prior investigations into PD genetics and drug discovery. First, we employed sc-eQTL data across 14 well-characterized immune cell types, allowing unprecedented resolution in identifying cell-type-specific gene expression patterns causally linked to PD. Unlike traditional bulk-tissue eQTL studies that average signals across heterogeneous cell populations, our approach revealed fine-grained regulatory mechanisms that would otherwise be masked62. Second, by integrating scTWMR and Bayesian colocalization, we provided robust causal inference, reducing confounding from LD or pleiotropy63. Third, we combined scRNA-seq transcriptomic profiling to characterize the upregulation and downregulation of prioritized eGenes within peripheral blood immune cells of PD patients, thereby directly validating their immune cell specificity in disease context. Fourth, the integration of PheWAS-based safety profiling with molecular docking-guided drug prediction enabled both therapeutic target validation and the prioritization of compounds with favorable safety and binding characteristics64. This comprehensive pipeline, from genetic association to functional prediction, offers a holistic translational framework for drug repurposing in PD. To our knowledge, this is one of the first studies to systematically link sc-eQTL driven immune regulation to PD risk and simultaneously derive pharmacological insights from it, offering novel directions for precision neuromodulation therapy.
Despite its strengths, this study has several limitations that warrant consideration. First, our analyses were based solely on data from cohorts of European ancestry, which may limit the generalizability of our findings to other populations. Differences in genetic architecture, allele frequencies, and LD patterns across ancestries can influence both immune gene regulation and the prioritization of causal genes, potentially affecting their relevance as therapeutic targets. However, with the rapid advancement of sequencing technologies and expanding efforts in diverse biobanks, future availability of diverse ancestry data will enable robust cross-ancestry validation of our findings and help ensure their broader applicability. Second, while MR and colocalization analyses provide strong causal inference at the genetic level, our proposed immune function mechanisms for key genes rely on bioinformatic inference. Additionally, the statistical power for smaller cell types was limited by their smaller sample sizes in the underlying sc-eQTL dataset, and these results should be interpreted cautiously until validated in larger cohorts. Experimental validation, such as CRISPR-Cas9 mediated knockout, gene overexpression, and functional assays in relevant immune cell or animal models, was beyond the scope of this study but will be essential in future work to confirm the biological mechanisms and therapeutic effects of our prioritized targets. Importantly, our current framework provides a high-confidence, cell-type-specific shortlist of candidate genes and drugs that could guide such downstream experimental efforts. Third, although some predicted compounds showed strong enrichment for prioritized targets, there is currently insufficient clinical evidence for their immune-modulating effects specifically in PD; such results should therefore be interpreted with caution and regarded as hypothesis-generating until further experimental or clinical validation is available. These limitations highlight the importance of complementing computational frameworks with empirical research in future translational efforts.
Methods
Study design and ethics
Overview of the study design and analysis pipeline were outlined in Fig. 1. This study was designed following the Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomization (STROBE-MR) guidelines to ensure transparency and scientific rigor in the application of MR18. The research framework integrated sc-eQTL data with large-scale GWAS results, aiming to investigate the causal influence of immune-cell-specific gene expression on PD risk. The workflow included instrument selection from sc-eQTL datasets, MR analysis, Bayesian colocalization, PheWAS evaluation, and compound screening through drug enrichment and molecular docking simulations. MR analysis relies on three fundamental assumptions: firstly, that the genetic variants selected as instruments are significantly correlated with the exposure; secondly, that these variants are not linked to potential confounders; and thirdly, that the association with the outcome is mediated solely through the exposure pathway. To uphold these assumptions, rigorous data harmonization and instrument filtering procedures were applied. All datasets utilized were obtained from publicly available resources and previously published studies, including GWAS summary statistics of PD and sc-eQTL data. No new human or animal subjects were involved, and no individual-level or identifiable data were accessed. Therefore, additional ethical approval and informed consent were not required. Ethical clearances for the original data sources had been secured by their respective research teams23. All analytical procedures were conducted in alignment with recognized ethical and regulatory standards.
Exposure data and instrument selection
To construct valid genetic instruments reflecting immune-cell-specific gene expression, we utilized cis-eQTL data from the OneK1K cohort23. This dataset offers high-resolution regulatory information across 14 distinct immune cell types. Only conditionally independent cis-eQTLs meeting genome-wide significance (P < 5.0 × 10−8) were selected to minimize the risk of false-positive associations and enhance reliability. In addition, sc-eQTL data from the DICE project were used as an independent replication resource65. A multi-step quality control process was employed to ensure instrument validity. First, LD pruning was performed using data from the 1000 Genomes Project (European population) with a stringent r2 threshold of <0.001 within a 10 Mb window66. This step aimed to remove correlated SNPs and retain only independent variants. Second, harmonization between exposure and outcome was carried out. This included aligning effect alleles, removing ambiguous or strand-incompatible variants, and retaining palindromic SNPs only when allele frequency data enabled unambiguous matching. To further strengthen the instruments, we calculated F-statistics for all SNPs and excluded those with F < 10 to avoid weak instrument bias67. After filtering and validation, a final set of 9117 SNPs representing 8733 unique immune-cell-specific genes (eGenes) was retained for downstream MR analysis.
Outcome data
To assess PD risk, we utilized publicly available summary statistics from a large-scale GWAS meta-analysis conducted by the International Parkinson’s Disease Genomics Consortium (IPDGC), which included 33,674 clinically diagnosed cases, and 449,056 controls, all of European ancestry14. The strong genetic correlation between proxy and clinical cases (rG > 0.8) justified their inclusion, increasing analytical power. This study identified 90 genome-wide significant signals across 78 loci, including numerous novel associations. A polygenic risk score (PRS) derived from 1,805 SNPs captured up to 36% of the heritable risk depending on disease prevalence assumptions. Importantly, functional analyses linked these risk variants to lysosomal and neurotransmitter-related pathways in the brain, offering mechanistic insights and supporting their use in downstream MR analysis.
Sc-eQTL MR analysis
To assess the causal relationship between immune-cell-specific gene expression and PD, we applied MR using the TwoSampleMR R package (v0.6.6)68. Each eGene identified from the sc-eQTL dataset was analyzed based on the number of independent SNP available as instruments. When an eGene was linked to a single instrumental variant, we employed the Wald ratio method; for eGenes with multiple independent instruments, the inverse-variance weighted (IVW) approach was used to estimate causal effects69. To address the issue of multiple comparisons across thousands of eGenes, we applied FDR correction, ensuring that only statistically significant gene-trait associations were retained for further interpretation. eGenes surpassing the FDR threshold were considered robust candidates and subsequently included in downstream analyses, such as colocalization, protein interaction exploration, and drug discovery assessments. This systematic MR framework enabled us to identify immune cell-specific genes that may play a causal role in PD susceptibility.
Colocalization analysis
To determine whether the same genetic variants influenced both gene expression and PD risk, we performed colocalization analysis using the coloc R package (v5.2.3), which applies a Bayesian framework to estimate the probability of shared causal variants70. For each eGene tested, we calculated the posterior probabilities for five possible hypotheses, placing particular emphasis on PP.H4, the probability that a single variant is responsible for both the expression and disease association signals. We classified eGenes into two confidence tiers: strong evidence (PP.H4 > 80%) and moderate evidence (50% < PP.H4 ≤ 80%)71. Only strong-evidence eGenes were prioritized for downstream functional analyses and therapeutic targeting, whereas moderate-evidence eGenes were reported separately and interpreted with caution. This tiered approach addresses potential false positives and reduces the impact of LD or coincidental overlap, ensuring that our main conclusions rely on the most credible immune-cell-specific regulatory signals for PD.
Peripheral blood scRNA-seq data pre-processing and annotation
We analyzed an independent scRNA-seq dataset from Xiong et al. (GSE223138)7, comprising two healthy controls, two early-stage PD patients, and two late-stage PD patients. Raw count matrices and metadata were processed in Seurat (v5.1.0) under R (v4.2.1)72,73. Quality control retained cells with <10% mitochondrial gene expression and ≥300 detected genes, and genes expressed in at least three cells with total counts between 600 and 7500. Filtered data were normalized and scaled using a linear regression model with the “Log-normalization” method, and the top 2000 variable genes were identified using “FindVariableFeatures”. Dimensionality reduction was performed with Principal Component Analysis (PCA), and batch effects were corrected using “Harmony”. Cell clustering was conducted with “FindClusters” at a resolution of 1. Cluster annotation relied on highly expressed, uniquely patterned genes and known canonical markers (Supplementary Fig. S3). For each immune cell type, differential expression between PD and HC was assessed using the Wilcoxon rank-sum test. Genes were classified as upregulated, downregulated, or not significant based on P-values and the direction of log2 fold change.
Phenome-wide association analysis across multiple resources
To comprehensively assess the broader phenotypic effects, immune relevance, and potential safety considerations of our prioritized targets, we conducted a two-part phenome-wide association analysis using complementary resources. We first queried the AstraZeneca PheWAS Portal (https://azphewas.com)74, which reports associations for rare variant-enriched genes in UK Biobank. For each prioritized eGene, we retrieved all available trait associations. Associations surpassing the genome-wide threshold (P < 1 × 10−8) were considered statistically significant, and those with P-values between 1 × 10−8 and 1 × 10−6 were considered suggestive. This analysis allowed us to directly evaluate whether our candidate eGenes have known links to immune-related or other phenotypic traits in the general population. Next, we performed SNP-level PheWAS for the IVs corresponding to our prioritized targets from the MR analysis. We first queried the Open Targets Platform (https://platform.opentargets.org)75, which integrates associations from 95% GWAS credible sets across multiple consortia, enabling systematic cross-trait evaluation. We then queried FinnGen (https://r12.finngen.fi/)76, which contains data from 500,348 individuals, including 2466 binary endpoints and 3 quantitative traits. Statistical significance was defined by Bonferroni correction. Significant associations then were systematically classified into immune-related and non-immune-related traits to facilitate interpretation within the context of immune function and potential on-target safety considerations.
Drug prediction based on DsigDB
To identify potential therapeutic compounds targeting the eGenes implicated in PD, we performed drug enrichment analysis using the DSigDB via the Enrichr web platform77,78. This database connects thousands of bioactive molecules with gene expression signatures, enabling the prediction of compounds that may modulate the activity of our prioritized eGenes. All eGenes identified through MR and colocalization were input into the tool, and enrichment scores were computed based on the overlap between gene sets and compound targets. Significance was evaluated using P-values and combined scores, with higher scores indicating stronger associations. The resulting list highlighted candidate drugs with potential for repurposing or further investigation as targeted treatments for PD-related immune gene regulation (Supplementary Table S12).
BBB permeability prediction
For candidate compounds predicted to target the prioritized PD-associated eGenes, we then assessed their potential to penetrate the BBB, which is a critical consideration for drugs intended to exert therapeutic effects within the CNS79. BBB permeability was predicted using the SwissADME online platform (http://www.swissadme.ch), which applies a combination of physicochemical descriptors and predictive models, including the BOILED-Egg method, to classify compounds as BBB permeant or non-permeant80. SMILES structures of each candidate compound were retrieved from the PubChem database and submitted to SwissADME (Supplementary Table S13).
Molecular docking simulation
The two-dimensional structures of small-molecule ligands were first retrieved from the PubChem database and subsequently converted into three-dimensional conformations. These structures were energy-minimized using ChemOffice 20.0 and saved in .mol2 format. Receptor proteins were selected from the RCSB Protein Data Bank (PDB), ensuring high-resolution crystal structures (resolution <2.0 Å)81. Using PyMOL (v2.6.0), crystallographic water molecules, phosphate ions, and unrelated ligands were removed, while active site residues were retained, and the cleaned structures were exported as .pdb files. Protein preparation steps, including protonation, hydrogenation, and side-chain optimization, were performed using Molecular Operating Environment (MOE) 2019.01 software. The binding pockets were defined using the “Site Finder” module with a constraint radius of 6 Å. Ligands were minimized using the AMBER10:EHT force field, with a convergence gradient threshold of 0.05 kcal·mol−1·Å−1. Docking was then carried out using 50 rigid conformation searches. The London dG scoring function was applied to evaluate binding poses, and free energy calculations were refined using the GBVI/WSA dG scoring algorithm. Key interactions, including hydrogen bonding, hydrophobic contacts, and π-π stacking, were visualized and analyzed using PyMOL (v2.6.0) and Discovery Studio 2019 to assess binding modes and energies.
Ethics approval and consent to participate
Ethical approval was obtained for each of the original GWAS and scRNA-seq studies used in this work. No individual-level or identifiable data were accessed. Therefore, no new ethical review board approval was required.
Data availability
GWAS summary statistics of PD were obtained from the IPDGC, which includes genetic data from 33,674 clinically diagnosed cases and 449,056 controls of European ancestry. The sc-eQTL data covering 14 immune cell types were sourced from the OneK1K cohort, providing high-resolution gene regulatory profiles across diverse immune subsets. For replication analyses, additional sc-eQTL data were retrieved from the DICE project. The independent scRNA-seq dataset used for transcriptomic validation is available in the GEO under accession number GSE223138. All data used are fully de-identified and comply with ethical standards for secondary analysis. Complete details, including dataset accession numbers and links, are provided in the Methods section and Supplementary Information. No new individual-level or proprietary datasets were generated in this study. All data that support the findings of this study are available online and can be further obtained from the corresponding author Yanggang Hong upon reasonable request.
Code availability
The analytical workflow of this study was implemented using open-source computational tools and publicly available R packages. The scRNA-seq data were processed and analyzed using Seurat (v5.1.0), and batch correction was performed with Harmony. MR analyses were conducted using the TwoSampleMR package (v0.6.6), and Bayesian colocalization was performed using the coloc R package (v5.2.3). Additional packages such as ggplot2, dplyr, and tidyverse were employed for data visualization and manipulation. Molecular docking simulations were carried out using PyMOL (v2.6.0), ChemOffice 20.0, and MOE 2019.01 software. The SwissADME platform was used to predict BBB permeability. Custom R and shell scripts developed for SNP filtering, harmonization, MR model setup, and result integration are available upon reasonable request to the corresponding author. All code was executed under R (v4.2.1), and full documentation is available to ensure reproducibility. Researchers interested in replicating or extending the analysis are encouraged to contact the corresponding author Yanggang Hong for script sharing and guidance.
References
Tolosa, E., Garrido, A., Scholz, S. W. & Poewe, W. Challenges in the diagnosis of Parkinson’s disease. Lancet Neurol. 20, 385–397 (2021).
Ascherio, A. & Schwarzschild, M. A. The epidemiology of Parkinson’s disease: risk factors and prevention. Lancet Neurol. 15, 1257–1272 (2016).
Collaborators, G. B. D. P.sD. Global, regional, and national burden of Parkinson’s disease, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 17, 939–953 (2018).
Poewe, W. et al. Parkinson disease. Nat. Rev. Dis. Prim. 3, 17013 (2017).
Vijiaratnam, N., Simuni, T., Bandmann, O., Morris, H. R. & Foltynie, T. Progress towards therapies for disease modification in Parkinson’s disease. Lancet Neurol. 20, 559–572 (2021).
Atashrazm, F. et al. Reduced glucocerebrosidase activity in monocytes from patients with Parkinson’s disease. Sci. Rep. 8, 15446 (2018).
Xiong, L. L. et al. Single-cell RNA sequencing reveals peripheral immunological features in Parkinson’s Disease. NPJ Parkinsons Dis. 10, 185 (2024).
Moquin-Beaudry, G. et al. Mapping the peripheral immune landscape of Parkinson’s disease patients with single-cell sequencing. Brain 148, 2847–2860 (2025).
Tansey, M. G. et al. Inflammation and immune dysfunction in Parkinson disease. Nat. Rev. Immunol. 22, 657–673 (2022).
Tan, E. K. et al. Parkinson disease and the immune system - associations, mechanisms and therapeutics. Nat. Rev. Neurol. 16, 303–318 (2020).
Kong, X. et al. The anti-inflammatory effects of itaconate and its derivatives in neurological disorders. Cytokine Growth Factor Rev. 78, 37–49 (2024).
Chitramuthu, B. P., Bennett, H. P. J. & Bateman, A. Progranulin: a new avenue towards the understanding and treatment of neurodegenerative disease. Brain 140, 3081–3104 (2017).
Weiner, H. L. Immune mechanisms and shared immune targets in neurodegenerative diseases. Nat. Rev. Neurol. 21, 67–85 (2025).
Nalls, M. A. et al. Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: a meta-analysis of genome-wide association studies. Lancet Neurol. 18, 1091–1102 (2019).
Rizig, M. et al. Identification of genetic risk loci and causal insights associated with Parkinson’s disease in African and African admixed populations: a genome-wide association study. Lancet Neurol. 22, 1015–1025 (2023).
Kim, J. J. et al. Multi-ancestry genome-wide association meta-analysis of Parkinson’s disease. Nat. Genet 56, 27–36 (2024).
Chang, D. et al. A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nat. Genet 49, 1511–1516 (2017).
Skrivankova, V. W. et al. Strengthening the reporting of observational studies in epidemiology using Mendelian randomization: the STROBE-MR statement. JAMA 326, 1614–1621 (2021).
Lange, L. M. et al. Prioritizing Parkinson’s disease risk genes in genome-wide association loci. NPJ Parkinsons Dis. 11, 77 (2025).
Lyu, Q., Chen, R., Qiu, Z., Wang, C. & Liu, R. Druggable targets for Parkinson’s disease: transcriptomics based Mendelian randomization study. Sci. Rep. 14, 25763 (2024).
Wang, Z. et al. Multi-omics analysis reveals the genetic aging landscape of Parkinson’s disease. Sci. Rep. 14, 31167 (2024).
Storm, C. S. et al. Finding genetically-supported drug targets for Parkinson’s disease using Mendelian randomization of the druggable genome. Nat. Commun. 12, 7342 (2021).
Yazar, S. et al. Single-cell eQTL mapping identifies cell type-specific genetic control of autoimmune disease. Science 376, eabf3041 (2022).
Nathan, A. et al. Single-cell eQTL models reveal dynamic T cell state dependence of disease loci. Nature 606, 120–128 (2022).
Blauwendraat, C., Nalls, M. A. & Singleton, A. B. The genetic architecture of Parkinson’s disease. Lancet Neurol. 19, 170–178 (2020).
Sun, Q. et al. Cholesterol mediated ferroptosis suppression reveals essential roles of Coenzyme Q and squalene. Commun. Biol. 6, 1108 (2023).
Mallick, K., Paul, S., Banerjee, S. & Banerjee, S. Lipid droplets and neurodegeneration. Neuroscience 549, 13–23 (2024).
Ting, J. P. & Trowsdale, J. Genetic control of MHC class II expression. Cell 109, S21–S33 (2002).
Wissemann, W. T. et al. Association of Parkinson disease with structural and regulatory variants in the HLA region. Am. J. Hum. Genet 93, 984–993 (2013).
Ettinger, R. A. et al. Technical validation and utility of an HLA Class II Tetramer assay for type 1 diabetes: a multicenter study. J. Clin. Endocrinol. Metab. 109, 183–196 (2023).
Moutsianas, L. et al. Class II HLA interactions modulate genetic risk for multiple sclerosis. Nat. Genet 47, 1107–1113 (2015).
Hong-Le, T. et al. Genetic mapping of multiple traits identifies novel genes for adiposity, lipids, and insulin secretory capacity in outbred rats. Diabetes 72, 135–148 (2023).
Keele, G. R. et al. Genetic fine-mapping and identification of candidate genes and variants for adiposity traits in outbred rats. Obesity (Silver Spring) 26, 213–222 (2018).
Kim, R. & Jun, J. S. Impact of overweight and obesity on functional and clinical outcomes of early Parkinson’s disease. J. Am. Med Dir. Assoc. 21, 697–700 (2020).
Prakash, R. et al. Distinct pathways of homologous recombination controlled by the SWS1-SWSAP1-SPIDR complex. Nat. Commun. 12, 4255 (2021).
Yao, S. et al. A transcriptome-wide association study identifies susceptibility genes for Parkinson’s disease. NPJ Parkinsons Dis. 7, 79 (2021).
Dang, X., Zhang, Z. & Luo, X. J. Mendelian randomization study using dopaminergic neuron-specific eQTL nominates potential causal genes for Parkinson’s disease. Mov. Disord. 37, 2451–2456 (2022).
Maiuri, T. et al. DNA damage repair in Huntington’s disease and other neurodegenerative diseases. Neurotherapeutics 16, 948–956 (2019).
Beenken, A. et al. Spns1 is an iron transporter essential for megalin-dependent endocytosis. Am. J. Physiol. Ren. Physiol. 327, F775–F787 (2024).
Ha, H. T. et al. Lack of SPNS1 results in accumulation of lysolipids and lysosomal storage disease in mouse models. JCI Insight 9, e175462 (2024).
Deneubourg, C. et al. The spectrum of neurodevelopmental, neuromuscular and neurodegenerative disorders due to defective autophagy. Autophagy 18, 496–517 (2022).
Lee, J. S. et al. Arylsulfatase A, a genetic modifier of Parkinson’s disease, is an alpha-synuclein chaperone. Brain 142, 2845–2859 (2019).
Panichnantakul, P. et al. Protein UFMylation regulates early events during ribosomal DNA-damage response. Cell Rep. 43, 114738 (2024).
Mandal, S. M. et al. Role of human DNA glycosylase Nei-like 2 (NEIL2) and single strand break repair protein polynucleotide kinase 3’-phosphatase in maintenance of mitochondrial genome. J. Biol. Chem. 287, 2819–2829 (2012).
Cappuccio, E. et al. FAHD1 and mitochondrial metabolism: a decade of pioneering discoveries. FEBS J. 292, 2973–2991 (2025).
Jones-Tabah, J. et al. The Parkinson’s disease risk gene cathepsin B promotes fibrillar alpha-synuclein clearance, lysosomal function and glucocerebrosidase activity in dopaminergic neurons. Mol. Neurodegener. 19, 88 (2024).
Sai, M. et al. Development of Nurr1 agonists from amodiaquine by scaffold hopping and fragment growing. Commun. Chem. 7, 149 (2024).
Sellnow, R. C. et al. Striatal Nurr1 facilitates the Dyskinetic state and exacerbates levodopa-induced Dyskinesia in a rat model of Parkinson’s disease. J. Neurosci. 40, 3675–3691 (2020).
Ferreira, J. J. et al. MDS evidence-based review of treatments for essential tremor. Mov. Disord. 34, 950–958 (2019).
Al Jaja, A. et al. Alprazolam reduces freezing of Gait (FOG) and improves FOG-related gait deficiencies. Parkinsons Dis. 2024, 3447009 (2024).
Lee, Y. C. et al. Antihypertensive agents and risk of Parkinson’s disease: a nationwide cohort study. PLoS One 9, e98961 (2014).
Siddiqi, F. H. et al. Felodipine induces autophagy in mouse brains with pharmacokinetics amenable to repurposing. Nat. Commun. 10, 1817 (2019).
Fernandes, H. J. R. et al. Single-cell transcriptomics of Parkinson’s disease human in vitro models reveals dopamine neuron-specific stress responses. Cell Rep. 33, 108263 (2020).
Abou-Hany, H. O., El-Sherbiny, M., Elshaer, S., Said, E. & Moustafa, T. Neuro-modulatory impact of felodipine against experimentally-induced Parkinson’s disease: Possible contribution of PINK1-Parkin mitophagy pathway. Neuropharmacology 250, 109909 (2024).
Kakish, J., Lee, D. & Lee, J. S. Drugs that bind to alpha-synuclein: neuroprotective or neurotoxic?. ACS Chem. Neurosci. 6, 1930–1940 (2015).
Papagiannakis, N. et al. Lysosomal alterations in peripheral blood mononuclear cells of Parkinson’s disease patients. Mov. Disord. 30, 1830–1834 (2015).
Pan, Y. & Nicolazzo, J. A. Impact of aging, Alzheimer’s disease and Parkinson’s disease on the blood-brain barrier transport of therapeutics. Adv. Drug Deliv. Rev. 135, 62–74 (2018).
Sweeney, M. D., Sagare, A. P. & Zlokovic, B. V. Blood-brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat. Rev. Neurol. 14, 133–150 (2018).
Seglen, P. O. Inhibitors of lysosomal function. Methods Enzymol. 96, 737–764 (1983).
Sun, L. et al. A lysosomal-mitochondrial death pathway is induced by solamargine in human K562 leukemia cells. Toxicol. Vitr. 24, 1504–1511 (2010).
Perez, S. et al. Effect of gambierol and its tetracyclic and heptacyclic analogues in cultured cerebellar neurons: a structure-activity relationships study. Chem. Res. Toxicol. 25, 1929–1937 (2012).
Ding, R. et al. scQTLbase: an integrated human single-cell eQTL database. Nucleic Acids Res. 52, D1010–D1017 (2024).
Ying, H. et al. Single-cell transcriptome-wide Mendelian randomization and colocalization reveals immune-mediated regulatory mechanisms and drug targets for COVID-19. EBioMedicine 113, 105596 (2025).
Bush, W. S., Oetjens, M. T. & Crawford, D. C. Unravelling the human genome-phenome relationship using phenome-wide association studies. Nat. Rev. Genet 17, 129–145 (2016).
Schmiedel, B. J. et al. Impact of genetic polymorphisms on human immune cell gene expression. Cell 175, 1701–1715.e1716 (2018).
Gkatzionis, A., Burgess, S. & Newcombe, P. J. Statistical methods for cis-Mendelian randomization with two-sample summary-level data. Genet Epidemiol. 47, 3–25 (2023).
Pierce, B. L., Ahsan, H. & Vanderweele, T. J. Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int J. Epidemiol. 40, 740–752 (2011).
Hemani, G. et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife 7, e34408 (2018).
Burgess, S. et al. Guidelines for performing Mendelian randomization investigations: update for summer 2023. Wellcome Open Res. 4, 186 (2019).
Giambartolomei, C. et al. Bayesian test for colocalisation between pairs of genetic association studies using summary statistics. PLoS Genet 10, e1004383 (2014).
Hong, Y. et al. Mapping the immune-genetic architecture of aging: a single-cell causal framework for biomarker discovery and therapeutic targeting. Ageing Res. Rev. 111, 102835 (2025).
Satija, R., Farrell, J. A., Gennert, D., Schier, A. F. & Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 33, 495–502 (2015).
Hao, Y. et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 42, 293–304 (2024).
Wang, Q. et al. Rare variant contribution to human disease in 281,104 UK Biobank exomes. Nature 597, 527–532 (2021).
Ochoa, D. et al. Open Targets Platform: supporting systematic drug-target identification and prioritisation. Nucleic Acids Res. 49, D1302–D1310 (2021).
Kurki, M. I. et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature 613, 508–518 (2023).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–W97 (2016).
Yoo, M. et al. DSigDB: drug signatures database for gene set analysis. Bioinformatics 31, 3069–3071 (2015).
Banks, W. A. From blood-brain barrier to blood-brain interface: new opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 15, 275–292 (2016).
Daina, A., Michielin, O. & Zoete, V. SwissADME: a free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 7, 42717 (2017).
Berman, H. M. et al. The protein data bank. Nucleic Acids Res. 28, 235–242 (2000).
Acknowledgements
The authors sincerely thank all researchers who generously provided the GWAS summary statistics and scRNA-seq datasets that were essential to this study. The authors also gratefully acknowledge BioRender for providing some of the materials used in the figures. Yanggang Hong extends his heartfelt thanks to Dr. Jiajun Li from Wenzhou Medical University, as well as Dr. Chen Yang, Dr. Haigang Geng, and Dr. Xifu Cheng from the Shanghai Cancer Institute, Renji Hospital, Shanghai Jiao Tong University School of Medicine, for their valuable technical support during the manuscript revision stage. This study was supported by the Natural Science Foundation of China [No. 82301672], Basic Public Welfare Projects of Zhejiang Province [No. LQ23H010002], and Science and Technology Bureau Project of Wenzhou [No. Y20240023].
Author information
Authors and Affiliations
Contributions
Yanggang Hong: conceptualization, investigation, formal analysis, data curation, validation, visualization, writing - original draft, writing - review & editing. Jingxuan Zhou: investigation, writing - original draft. Yirong Wang: writing - original draft. Sihan Song: writing - original draft. Han Chen: writing - original draft. Yuze Mi: investigation, writing - original draft. Xiucui Li: validation, writing - review & editing, funding acquisition.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hong, Y., Zhou, J., Wang, Y. et al. Peripheral immune cell-specific genes in Parkinson’s disease uncovered by multi-omics with therapeutic implications. npj Parkinsons Dis. 11, 302 (2025). https://doi.org/10.1038/s41531-025-01148-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41531-025-01148-z
