
Abstract
Cancer cell plasticity enables the acquisition of new phenotypic features and is implicated as a major driver of metastatic progression1,2. Metastasis occurs mostly in the absence of additional genetic alterations3,4,5, which suggests that epigenetic mechanisms are important6. However, they remain poorly defined. Here we identify the chromatin-remodelling enzyme ATRX as a key regulator of colonic lineage fidelity and metastasis in colorectal cancer. Atrx loss promotes tumour invasion and metastasis, concomitant with a loss of colonic epithelial identity and the emergence of highly plastic mesenchymal and squamous-like cell states. Combined analysis of chromatin accessibility and enhancer mapping identified impairment of activity of the colonic lineage-specifying transcription factor HNF4A as a key mediator of these observed phenotypes. We identify squamous-like cells in human patient samples and a squamous-like expression signature that correlates with aggressive disease and poor patient prognosis. Collectively, our study defines the epigenetic maintenance of colonic epithelial identity by ATRX and HNF4A as suppressors of lineage plasticity and metastasis in colorectal cancer.
Similar content being viewed by others

Main
Metastasis poses a substantial challenge in cancer management because it significantly affects patient prognosis and response to therapy7,8. The dissemination and spread of cancer cells to distant organs is associated with increased cellular heterogeneity and phenotypic plasticity that mostly occurs in the absence of additional genetic alterations3,4,5. This finding indicates that additional layers of regulation, particularly epigenetic alterations, are responsible for driving the phenotypic changes required for metastasis6. Epigenetic modifications, including DNA methylation, post-translational histone modifications and chromatin compaction, collectively influence gene expression. Epigenetic modification is typically involved in the maintenance of cellular identity. However, changes in epigenetic modification patterns can influence lineage determination, thereby providing cells with a heightened degree of plasticity. For example, widespread reprogramming of chromatin modification has been reported in pancreatic cancer metastasis, and alterations in chromatin accessibility have been identified across multiple tumour state transitions, including metastasis6,9. Together, these results suggest that epigenetic alterations can enhance plasticity to enable cancer cells to undergo phenotypic transitions that facilitate invasion and colonization at distant sites10.
Epigenetic modifying enzymes are among the most mutated families of genes in cancer11,12. However, comprehensive studies investigating the specific impact of mutated epigenetic modifiers on cellular plasticity and metastasis are limited13. Here we investigated the function of the chromatin-remodelling helicase ATRX in colorectal cancer (CRC). We show that loss of Atrx promotes metastasis and is associated with the emergence of highly plastic, mesenchymal and squamous-like cell states. Transcriptional analysis identifies loss of expression of genes associated with colonic epithelial identity, which suggests that loss of lineage fidelity is a key step that controls metastatic progression. Mechanistically, Atrx loss leads to a loss of chromatin accessibility and enhancer activity at targets of the colonic-specifying transcription factor (TF) HNF4A, which we functionally identify as a key mediator of colonic lineage fidelity. In patient samples, expression of a squamous-like gene expression signature is associated with aggressive, metaplastic CRC subtypes and predicts poor patient outcomes. Together, our data indicate that loss of colonic epithelial identity and acquisition of highly plastic squamous-like cell states are key drivers of CRC metastasis.
Atrx loss induces CRC metastasis
Analysis of the International Cancer Genome Consortium database revealed a prevalence of mutations in epigenetic regulators in CRC, including ATRX, what was mutated in around 7% of samples (Extended Data Fig. 1a). Immunohistochemistry (IHC) analysis of a tissue microarray (TMA) of CRC cases revealed that loss of ATRX expression is associated with late-stage, metastatic disease (Extended Data Fig. 1b–d). Consistent with this finding, ATRX mutation is more prevalent in the highly aggressive CRIS-B transcriptional CRC subtype14, in which it correlated with poor prognosis (Extended Data Fig. 1e,f). Together, these data suggest that loss of ATRX function has a role in promoting aggressive disease and metastasis.
To investigate this possibility, we used CRISPR–Cas9 genome editing to disrupt Atrx in a mouse Apcfl/flKrasG12DTrp53fl/fl (AKP) CRC organoid line15. AKP organoids have low metastatic potential, which facilitates analyses of the effects of additional perturbations on this sytem15. An analysis of pan-cancer mutational data indicated a preponderance of stop codon mutations in the 5′ end of the SNF2 and helicase chromatin-remodelling domains of ATRX (in particular, R1426*) (Extended Data Fig. 1g). Therefore, we used a single guide RNA (sgRNA) targeted to this region to knock out Atrx in AKP organoids (AKP AtrxKO) (Extended Data Fig. 1g,h). An AKP control line was also generated using a non-targeting gRNA to control for effects of transduction and Cas9 expression.
We first investigated the effects of Atrx loss on CRC metastasis. Tail-vein inoculation or intrasplenic transplantation of dissociated AKP AtrxKO organoids led to a significant increase in lung and liver metastatic burden, respectively, compared with AKP controls (Fig. 1a–h). Increased metastatic potential was confirmed with an independent clonal AtrxKO organoid line (AKP AtrxKO2), which also displayed an increase in lung metastasis after tail-vein injection (Extended Data Fig. 1h–l). To determine whether this increase was associated with changes in primary tumour phenotypes, we subcutaneously transplanted the same organoid lines into mice. Although there was no difference in tumour size (Extended Data Fig. 2a,b), histological analyses uncovered considerable differences in tumour histology. AKP tumours had a classical glandular morphology, whereas AKP AtrxKO tumours displayed areas of poor differentiation and evidence of tumour cells invading into the surrounding stroma (Extended Data Fig. 2c–e). AKP control tumour cells were predominantly positive for the expression of the epithelial marker E-cadherin, whereas Atrx loss led to consistent loss of E-cadherin positivity at invasive regions (Extended Data Fig. 2f,g). Transcriptional and protein analyses also identified an induction of mesenchymal markers in AKP AtrxKO tumours (Extended Data Fig. 2h–j). This phenotype, indicative of epithelial-to-mesenchymal transition (EMT), was further investigated in vitro. TGFβ can act as an inducer of EMT and is associated with aggressive, late-stage CRC14. Treatment of AKP AtrxKO organoids with TGFβ led to the emergence of spreading, spindle-like cells reminiscent of mesenchymal cells and the induction of EMT marker expression compared with controls (Fig. 1i–k). This effect was confirmed with the AKP AtrxKO2 clonal line (Extended Data Fig. 2k–l) and was observed across a range of TGFβ concentrations but not in response to other EMT inducers such as TNF or IFNγ (Extended Data Fig. 3a–c). This result suggests that Atrx deletion leads to a high level of sensitivity to TGFβ-driven EMT induction. We also analysed the response of these organoid lines to epigenetic modifying drugs to determine whether Atrx deletion sensitizes cells to such agents. Treatment with inhibitors of both BET (JQ1) and HDAC (FK228) impaired cell viability in both lines, with no increased sensitivity in AtrxKO cells (Extended Data Fig. 3d,e).

a, Representative images of lung metastases (stained with haematoxylin and eosin) from mice injected with AKP control or AKP AtrxKO organoids through the tail vein. Metastatic nodules are indicated with black arrowheads. b, Quantification of the number of lung metastases per mouse (n = 8 mice each). c, Quantification of total lung tumour burden per mouse (n = 8 mice each). d, Summary data indicating the presence or absence of lung metastases. The number of mice with or without lung metastases is indicated on the graph (n = 8 mice each). e, Representative images of liver metastases (stained with haematoxylin and eosin) from mice injected with AKP control or AKP AtrxKO organoids through intrasplenic injection. Metastatic nodules are indicated with black arrowheads. f, Quantification of the number of liver metastases per mouse (n = 8 mice each). g, Quantification of the total liver tumour burden per mouse (n = 8 mice each). h, Summary data indicating the presence or absence of liver metastases. The number of mice with or without liver metastases is indicated on the graph (n = 8 mice each). i, Fluorescence microscopy of calcein-stained AKP control and AKP AtrxKO organoids after treatment with 5 ng ml–1 TGFβ (TGFβ1). Spindle-like organoid structures are indicated with white arrowheads. Zoomed areas are outlined by the white boxes. j, Quantification of the percentage of AKP control and AKP AtrxKO organoids adopting a spindle-like morphology after TGFβ treatment (n = 7 (control) and 6 (KO) independent experiments). P = 0.000027. k, RT–qPCR analysis of EMT markers in AKP control and AKP AtrxKO organoids untreated or treated with 5 ng ml–1 TGFβ (n = 3 independent experiments). Gene expression was normalized to Actb, and levels relative to untreated AKP control were calculated using the ΔΔCt method. Data are the mean ± s.d. (b,c,f,g,j,k). P values were calculated using two-tailed Mann–Whitney tests (b,c,f,g), two-sided Fisher’s exact tests (d,h), two-tailed Student’s t-tests (j) or ordinary one-way analysis of variance ANOVA with multiple comparisons (k). Scale bars, 2.5 mm (a,e) or 1,000 μm (i).
To determine whether Atrx loss promotes metastatic dissemination from primary tumours, mice were implanted with AKP control or AKP AtrxKO organoids into the colonic submucosa. Although no differences in overall survival or primary tumour growth were observed, AKP AtrxKO tumours led to significantly more metastases than controls, with metastases observed in the liver, lymph nodes and diaphragm (Extended Data Fig. 3f–k). Moreover, analyses of primary tumour histology demonstrated loss of glandular morphology in AKP AtrxKO tumours (Extended Data Fig. 3k–m). To further extend these findings, we deleted Atrx in the BrafV600ETrp53fl/flNotchICD (BPN) mouse CRC model (Extended Data Fig. 4a,b). This BrafV600E-driven model is transcriptionally distinct from the AKP model and resembles the CMS4 CRC subtype16. Similar to the AKP model, Atrx deletion increased the induction of TGFβ-mediated EMT (Extended Data Fig. 4c–f). After orthotopic transplantation, BPN AtrxKO tumours completely lost their glandular morphology and exhibited a poorly differentiated phenotype (Extended Data Fig. 4g,h). Atrx deletion also led to metastatic progression in about 25% of mice (Extended Data Fig. 4i,j). Together, these data demonstrate that Atrx loss in multiple mouse CRC models promotes an aggressive tumour phenotype associated with sensitivity to EMT induction, tumour invasion and metastatic progression.
Atrx protects colonic epithelial identity
To mechanistically define how Atrx loss mediates tumour metastasis, we performed RNA sequencing (RNA-seq) analysis of AKP control and AKP AtrxKO organoid lines treated or untreated with TGFβ. Consistent with the observed EMT phenotype, gene set enrichment analysis (GSEA) identified significant enrichment of multiple EMT-related signatures in TGFβ-treated AKP AtrxKO organoids, which was confirmed by quantitative PCR with reverse transcription (RT–qPCR) (Extended Data Fig. 4k–l and Supplementary Table 1). Untreated organoids did not show the same global enrichment for EMT signatures or EMT marker expression after TGFβ treatment (Fig. 2a, Extended Data Fig. 4l and Supplementary Table 2). However, untreated AKP AtrxKO organoids expressed several genes that are expressed in mesenchymal cells, such as Twist1, Itga5, Tfap2c and Irx2. This result suggests that despite being insufficient to induce EMT, Atrx loss leads to induction of a partial mesenchymal-like phenotype (Fig. 2a and Extended Data Fig. 4l). Notably, Atrx loss precipitated a discernible lack of expression of gene signatures associated with colonic epithelial identity (Extended Data Fig. 4m); for example, targets of the colon-specifying TFs HNF4A, CDX1 and CDX2, and genes more broadly associated with colonic function, such as Aqp1, Cftr and Tff3 (Fig. 2a and Extended Data Fig. 4n). Loss of expression of key colonic TFs was also seen in the BPN model (Extended Data Fig. 4o). We expanded our analysis using TissueEnrich, which not only confirmed the loss of colonic epithelial gene expression but also revealed a shift towards the expression of genes typically associated with squamous tissues, specifically the oesophagus, skin and adipose tissue (Fig. 2b). Such genes included highly specific markers of squamous tissues, such as Ly6d, Krt5, Cav1, Cdkn1c and Elf5, and adipose-specific genes encoding molecules involved in lipid catabolism, storage and localization, such as Plin5, Acacb, Cidea and Dgat2 (Fig. 2b, Extended Data Fig. 4p and Supplementary Table 3). Among this lineage reprogramming effect, the expression of more broadly expressed epithelial markers, such as Epcam, Cdh1 and Cldn4, was maintained (Fig. 2a). The preservation of these more ubiquitous epithelial markers suggests that there is a selective loss of colonic lineage specification, whereas broader epithelial characteristics are maintained.

a, Heatmap of RNA-seq data from AKP control and AKP AtrxKO organoids with or without TGFβ treatment. Representative genes marking colonic epithelial, squamous and mesenchymal lineages are shown. log2 fold change values relative to untreated AKP control organoids are indicated by the colour intensity. Genes of multiple lineages co-expressed in AKP AtrxKO organoids are highlighted as ‘hybrid phenotype’. b, TissueEnrich analysis of genes upregulated and downregulated in AKP AtrxKO organoids compared with AKP controls. Dashed line indicates P = 0.05. c, UMAP plot of AKP control (23,579 cells) and AKP AtrxKO (25,757 cells) single cells coloured by genotype. d, UMAP plot coloured and numbered by cluster in AKP control and AKP AtrxKO single cells. e, UMAP plots coloured by the expression of genes used for defining colonic differentiation and EMT in AKP control and AKP AtrxKO single cells. Colour scale indicates expression levels. f, UMAP plot coloured by the expression of genes used for defining squamous differentiation in AKP control and AKP AtrxKO single cells. Colour scale indicates expression levels. g, TissueEnrich analyses of genes enriched in single-cell RNA-seq clusters 4 and 15. Dashed line indicates P = 0.05. h, Dot plot of signature scores across all clusters coloured by the average expression and sized by the percentage of cells expressing the signature. Cluster 4 oesophagus and cluster 15 skin signatures are derived from TissueEnrich analyses. Significance was calculated using hypergeometric tests (one-sided) with Benjamini–Hochberg multiple-testing correction (b,g).
To further investigate changes in cell state induced by Atrx loss, we carried out single-cell RNA-seq of AKP control and AKP AtrxKO organoids. Harmony integration and uniform manifold approximation and projection (UMAP) revealed clear separation between the two conditions, which indicated that Atrx loss induces considerable changes in the cell transcriptional state (Fig. 2c). Clustering analysis identified 18 cell clusters (Fig. 2d and Extended Data Fig. 5a and Supplementary Table 4), for which we mapped expression of key lineage markers. Consistent with the bulk RNA-seq results, Hnf4a and Cdx1 expression was lost in AtrxKO cells, whereas Twist1 expression was increased (Fig. 2e). Notably, Epcam expression was maintained, which reinforced the idea that broad epithelial identity is maintained despite extensive lineage reprogramming.
Given the observed loss of colonic epithelial identity and gain of squamous gene expression observed in the bulk RNA-seq data, we next sought to determine whether Atrx deletion induces an adoption of non-intestinal lineage programs. Cluster analysis confirmed the emergence of multiple squamous-like and other non-canonical cell states after Atrx loss. Cluster 15 expressed markers of squamous-like epithelia, including Ly6d, Trp63, Krt5, Krt14, Krt79 and Krtdap (Fig. 2f–h and Supplementary Table 4). Cluster 4 also expressed squamous-like markers, such as Ly6d, Sprr1a and Mxd1 (Fig. 2f–h and Supplementary Table 4). TissueEnrich analysis of markers of these clusters identified cluster 15 as being strongly associated with skin and oesophageal epithelia, whereas cluster 4 showed a weaker association with these cell states, which was probably due to the lack of keratin expression (Fig. 2g). In AtrxKO cells, we also observed broad upregulation of osteoblast-like markers such as Id1, Id3, Bmp2 and Wif1, which indicated a shift towards a mesenchymal-like phenotype (Fig. 2h and Extended Data Fig. 5b). Expression of osteoblast markers was particularly enriched in clusters 13 and 15, which indicated the presence of mixed-lineage populations17 (Fig. 2h and Extended Data Fig. 5b). Overall, the presence of multiple non-canonical, squamous and mesenchymal-like lineages, which have been recently linked to metastasis17, highlights the induction of broad cellular reprogramming in AtrxKO cells, a finding indicative of increased phenotypic plasticity.
We next determined the relationship of these non-canonical lineages to previously identified CRC cell states linked to tumour progression. Canonical, LGR5 stem cell and tumour intestinal stem cell signatures were depleted in AtrxKO cells, consistent with the loss of colonic identity17 (Fig. 2h and Extended Data Fig. 5c). Conversely, we found upregulation of fetal intestine, epithelial-specific high-risk and injury-repair programs, which have previously been associated with tumour progression and metastasis17,18,19 (Fig. 2h and Extended Data Fig. 5d). These programs were enriched in the squamous-like clusters 4 and 15, which indicated the co-expression of fetal and injury repair and non-canonical cell-lineage states (Fig. 2h). Together, these results suggest that rather than shifting towards a single alternative lineage, AtrxKO cells adopt a range of non-canonical cell states with distinct clusters exhibiting mixed features of squamous, mesenchymal and fetal intestinal identities. The emergence of these hybrid cell states, which have been linked to tumour progression and metastasis17, suggests that Atrx plays a key role in maintaining colonic epithelial identity and restricting lineage plasticity in CRC.
Atrx suppresses multilineage plasticity
To dissect how Atrx loss leads to lineage plasticity, we first investigated the induction of mesenchymal gene expression. Co-immunofluorescence staining for CDH1 and TWIST1 revealed that AtrxKO organoids express both these epithelial and mesenchymal markers (Extended Data Fig. 6a). Further characterization using fluorescent-activated cell sorting (FACS) for markers of epithelial (EPCAM) and mesenchymal (ITGA5) cells confirmed the expression of both these lineage markers (Extended Data Fig. 6b–d). Notably, EPCAM expression was maintained in AtrxKO cells, which indicated the transition towards a hybrid EMT cell state rather than full EMT induction (Extended Data Fig. 6e). Subsequent cell sorting on the basis of ITGA5 expression showed that ITGA5+ cells displayed increased sensitivity to TGFβ-induced EMT and elevated expression of EMT markers, which confirmed the establishment of a hybrid EMT phenotype (Extended Data Fig. 6f–j).
We next investigated the characteristics of the squamous-like cell states induced by Atrx loss. FACS analysis showed that expression of the squamous cell marker LY6D was significantly induced in AKP AtrxKO organoids (Fig. 3a–c). Transcriptional analysis of sorted LY6D+ cells confirmed the expression of markers of squamous epithelium, such as Krt5 and Krt13 (Fig. 3d). These cells had increased expression of Emp1, a marker of metastatic CRC18, and, consistent with a shift away from a colonic epithelial expression profile, they had reduced expression of the canonical colonic stem cell marker Lgr5 (Extended Data Fig. 7a). To determine whether squamous-like cells were present in vivo, we stained for KRT5 in both subcutaneous transplants and lung metastases from our mouse models. AKP AtrxKO tumours contained significantly more KRT5+ cells than control tumours, and metastatic lesions were further enriched compared with subcutaneous tumours (Fig. 3e–g and Extended Data Fig. 7b). Further analysis of these cells revealed an abnormal phenotype compared with CRC cells not expressing KRT5. Individual KRT5+ cells displayed an elongated morphology, distinct from the typical columnar morphology of colonic epithelium (Extended Data Fig. 7c). Moreover, clusters of KRT5+ cells acquired a stratified-like phenotype characteristic of squamous epithelia (Extended Data Fig. 7c). We also observed the occurrence of structures resembling keratin pearls, which are more commonly associated with cancers of squamous cell origin (Extended Data Fig. 7c). Together, these findings indicate that Atrx loss leads to the emergence of cells with squamous-like characteristics. Co-immunofluorescence staining for EPCAM and KRT5 in subcutaneous tumours demonstrated an apparent continuum of cell states. We found areas of exclusive KRT5 or EPCAM positivity, but also cells expressing both markers, a result indicative of phenotypic transitions between columnar epithelial and squamous-like states (Fig. 3h). To better understand the relationship between the observed squamous and mesenchymal hybrid cells, LY6D and ITGA5 co-staining was used. This revealed the presence of populations that expressed the individual marker and a highly mixed-lineage cell population that expressed both the squamous and mesenchymal markers (Fig. 3i,j and Extended Data Fig. 7d).

a, FACS plots for analysing and sorting EPCAM+LY6D+ cells from AKP control and AKP AtrxKO organoids. b, Quantification of the percentage of cells in each EPCAM and LY6D population (n = 3 independent experiments each). c, Quantification of the percentage of LY6D+ cells in AKP control and AKP AtrxKO organoids (n = 3 independent experiments each). P = 0.000019. d, RT–qPCR analysis of squamous cell markers in LY6D– and LY6D+ cells sorted from AKP AtrxKO organoids (n = 3 independent experiments each). Gene expression was normalized to Actb, and levels relative to LY6D– cells were calculated using the ΔΔCt method. e, Representative images of KRT5-stained subcutaneous tumours and lung metastases from mice injected with AKP control or AKP AtrxKO organoids. f, Quantification of the percentage of KRT5+ cells in subcutaneous tumours from mice injected with AKP control or AKP AtrxKO organoids (n = 5 mice each). g, Quantification of the percentage of KRT5+ cells in subcutaneous tumours and lung metastases from mice injected with AKP AtrxKO organoid cells (n = 5 (subcutaneous) and 6 (lung metastasis) mice). h, Representative images of EPCAM and KRT5 co-immunofluorescence in AKP AtrxKO subcutaneous tumours. White arrows indicate cells exclusively expressing EPCAM (in EPCAM only panel, magenta), KRT5 (in KRT5 only panel, green) or co-expressing EPCAM and KRT5 (merge). n = 3 biologically independent samples. i, FACS plots for analysing and sorting LY6D+ITGA5+ cells from AKP control and AKP AtrxKO organoids. j, Quantification of the percentage of cells in each LY6D and ITGA5 population (n = 3 independent experiments each). k, Schematic of the strategy used to determine the plasticity of different ITGA5 and LY6D expressing cell populations in AKP AtrxKO organoids. l, Quantification of the percentage of cells in each LY6D and ITGA5 population 9 days after plating. The original plated population is noted on the x axis (n = 4 independent experiments each). m, Representative images of KRT5-stained subcutaneous tumours from mice injected with different LY6D and ITGA5 populations derived from AKP AtrxKO organoid cells. The population transplanted is indicated above each image (n = 5 mice each). Scale bars, 50 µm. Data are the mean ± s.d. (c,d,f,g). P values were calculated using two-tailed Student’s t-test (c,d) or two-tailed Mann–Whitney test (f,g). Scale bars, 50 µm (e,h,m). The schematic in k was created using BioRender (https://biorender.com).
Our observations that Atrx loss induces cellular plasticity and mixed-lineage cell states prompted us to explore the stability of the hybrid-lineage populations that emerge. To this end, we sorted the various hybrid epithelial, mesenchymal and squamous cell populations, replated them and reanalysed them 9 days later (Fig. 3k). Regardless of the initial population plated, a resurgence of the other cell populations was observed (Fig. 3l and Extended Data Fig. 7e). This result was validated in vivo, as transplant experiments revealed a similar, dynamic re-emergence of KRT5+ squamous-like cell populations regardless of the cell population transplanted (Fig. 3m and Extended Data Fig. 7f). Thus, Atrx loss confers a high level of plasticity and gives CRC cells the ability to readily shift between columnar epithelial, mesenchymal and squamous-like cell states.
Owing to its previously described function in regulating EMT in cancer, we next tested the role of TGFβ signalling in mediating these lineage transitions. First, we tested the short-term effects of TGFβ treatment on organoid growth. AKP AtrxKO organoids were partially resistant to the growth-suppressive effects of this treatment (Extended Data Fig. 8a,b). To test the functional requirement for TGFβ signalling in mediating non-canonical lineage plasticity, we deleted Tgfbr2 in our AKP AtrxKO model to disrupt TGFβ signalling (Extended Data Fig. 8c). This modification resulted in a reduction in the expression of both squamous (Krt5 and Ly6d) and EMT-associated (Twist1) markers, along with a decrease in the proportion of LY6D+, ITGA5+ and LY6D+ITGA5+ double-positive cells (Extended Data Fig. 8d–f). Notably, the expression of colon-specifying genes such as Hnf4a, Cdx1 and Cdx2 was unchanged, which suggested that loss of colonic identity is mostly independent of TGFβ signalling (Extended Data Fig. 8d). Moreover, AKP AtrxKO cells lacking Tgfbr2 were unable to undergo EMT in response to TGFβ stimulation (Extended Data Fig. 8g,h). Although these results highlight an important role for TGFβ in promoting plasticity, the persistence of some LY6D+ populations suggests that alternative mechanisms may contribute to these transitions. Thus, Atrx loss leads to an impairment of colonic lineage fidelity and an induction of mesenchymal and squamous-like plasticity in a manner that partially depends on TGFβ signalling.
Hnf4a loss perturbs colonic identity
To unravel the molecular mechanisms underpinning the role of ATRX in maintaining lineage fidelity, we investigated its chromatin-remodelling function by carrying out assay for transposase-accessible chromatin with sequencing (ATAC–seq) on AKP control and AKP AtrxKO organoids. Principal component analysis revealed prominent separation of organoid genotypes in the first component (Extended Data Fig. 9a), which indicated the presence of differences in ATAC–seq enrichment patterns between the sample groups (Supplementary Table 6). The differences included notable, significant losses at sites associated with colonic epithelial gene expression (Extended Data Fig. 9b). We also observed gain of accessibility at some mesenchymal and squamous cell markers, consistent with induction of these intermediate cell states (Extended Data Fig. 9c,d). To investigate how these chromatin changes induce the phenotypic alterations we observed, we carried out TF-binding site analysis on sites of altered accessibility. This analysis revealed a marked enrichment in binding sites of the HNF4A, HNF4G, GATA2, GATA3 and GATA5 TFs in regions with decreased accessibility (Extended Data Fig. 9e). To further quantify this differential TF activity, we performed diffTF analysis on our combined ATAC–seq and RNA-seq datasets20. This analysis confirmed the loss of HNF4A and HNF4G and GATA binding site accessibility and reduced transcriptional output associated with these TFs (Fig. 4a). Complementary mapping of enhancer activity using H3K27ac CUT&RUN revealed a similar pattern, with widespread loss of enhancer activity after Atrx loss (Fig. 4b, Extended Data Fig. 9f and Supplementary Table 7). By comparing these datasets, we revealed that the majority of sites that lose H3K27ac overlapped with regions displaying reduced chromatin accessibility (Fig. 4c). Again, this was apparent at sites associated with colonic epithelial gene expression (Fig. 4d). The integration of these datasets suggests that Atrx loss disrupts the chromatin landscape, specifically at sites associated with epithelial gene expression, by directly affecting chromatin accessibility and enhancer activity.

a, DiffTF analysis of combined ATAC–seq and RNA-seq data with selected TFs highlighted. Shaded areas indicate gain (red) or loss (blue) of TF activity in AKP AtrxKO organoids compared with AKP controls. TFs highlighted in green are associated with transcriptional activation and in red with transcriptional repression. b, Volcano plot of H3K27ac CUT&RUN data in AKP control and AKP AtrxKO organoids. Each data point represents a H3K27ac binding peak. Significantly altered sites (false discovery rate (FDR) ≤ 0.05) are highlighted in pink. c, Table outlining the overlap between ATAC–seq accessibility changes and altered H3K27ac peaks in AKP control and AKP AtrxKO organoids. H3K27ac losses are mostly associated with reduced chromatin accessibility. d, Representative Integrative Genomics Viewer (IGV) browser tracks of AKP control and AKP AtrxKO ATAC–seq accessibility and H3K27ac CUT&RUN data. Cdx1 and Hnf4a gene loci are shown. Regions shaded grey have significant loss of chromatin accessibility and corresponding depletion of H3K27ac. e, Table outlining the overlap between RNA-seq gene expression changes in AKP AtrxKO and AKP Hnf4aKO organoids. f, Fluorescence microscopy of phalloidin-stained AKP control or AKP Hnf4aKO organoids after treatment with TGFβ (5 ng ml–1). Scale bars, 400 μm. g, Quantification of the percentage of AKP control and AKP Hnf4aKO organoids adopting a spindle-like morphology after TGFβ treatment (n = 3 independent experiments each). h, Representative IHC images of β-catenin-stained subcutaneous tumours from mice injected with AKP control or AKP Hnf4aKO organoid cells (n = 5 mice each). β-catenin staining is used to identify tumour cells. Scale bars, 100 µm (overview) and 50 μm (zoom). For a, two-sided P values for each transcription factor was calculated with Welch two-sample t-tests using the bootstrap approach. Adjusted P values were calculated using the Benjamini–Hochberg method for multiple-testing correction. For g, data are mean ± s.d., and P values were calculated using two-tailed Student’s t-tests.
The HNF4 TFs have a crucial role in maintaining normal colonic identity21,22, which suggests that loss of this activity may result in the impaired lineage fidelity observed after Atrx loss. To functionally test this possibility, we carried out HNF4A gain and loss of function experiments. Consistent with the proposal that HNF4A activity maintains colonic epithelial identity, HNF4A overexpression in AKP AtrxKO organoids rescued the expression of some of the colonic lineage markers analysed (Extended Data Fig. 9g). This partial rescue suggests that HNF4A activity maintains colonic epithelial identity, probably in cooperation with additional factors. To further investigate this possibility, we generated AKP Hnf4aKO organoids (Extended Data Fig. 9h,i) and performed RNA-seq, which revealed widespread changes in gene expression (Extended Data Fig. 9j and Supplementary Table 8). Notably, we observed loss of colon-specific genes such as Cdx1, Muc4, Lyz1 and Tff3, alongside upregulation of genes associated with non-canonical lineages, including Twist1, Krt4, Krt17, Krt79, Wif1 and Id3 (Extended Data Fig. 9j and Supplementary Table 8). Comparative analysis of gene expression changes in AtrxKO and Hnf4aKO organoids demonstrated a strong overlap in downregulated genes, which indicated that ATRX and HNF4A have a key role in maintaining colonic identity (Fig. 4e).
Functionally, Hnf4aKO organoids exhibited increased sensitivity to TGFβ-induced EMT (Fig. 4f,g). Moreover, subcutaneous transplantation of these organoids gave rise to tumours that had lost their glandular architecture (Fig. 4h and Extended Data Fig. 9k–l). Together, these findings indicate that loss of HNF4A activity at colonic specifying genes results in loss of colonic epithelial identity, which in turn may create a permissive environment for the emergence of the highly plastic cell states that emerge after Atrx loss.
Squamous phenotype in metastatic disease
To assess the clinical relevance of our findings, we investigated the consequences of loss of colonic identity and squamous-like transitions in human CRC samples. We first asked whether the relationship among ATRX, HNF4A, CDX2 and LY6D expression identified in our mouse model exists in human tumour samples. Staining of a CRC tissue microarray (TMA) consisting of stage I–III primary tumours using antibodies for these four proteins revealed a significant correlation between the expression of ATRX, HNF4A and CDX2, a result in keeping with our mouse model findings (Fig. 5a and Extended Data Fig. 10a–c). Moreover, consistent with a role for ATRX, HNF4A and CDX2 in maintaining lineage fidelity and suppressing squamous-like plasticity, we observed low expression of these proteins in tumours containing LY6D+ cells (Fig. 5b).

a, Representative IHC staining of a human CRC TMA for ATRX, HNF4A, CDX2 and LY6D. Examples of positive and negative staining are shown. Scale bar, 100 µm. b, Quantification of ATRX, HNF4A and CDX2 histoscore (H-score) values in LY6D– (<2% cells LY6D+) and LY6D+ (>2% cells LY6D+) tumour cores. n = 500 (ATRX, HNF4A) and 509 (CDX2) biologically independent samples. c, Representative IHC image of a LY6D-stained human stage IV primary tumour. Scale bars, 50 µm. d, Summary data indicating the percentage of human primary tumours at stages I–III versus stage IV positive for LY6D. The percentages are based on >2% cells LY6D+ and <2% cells LY6D–. P = 0.000027. e, Representative IHC image of LY6D-stained human liver metastasis. Scale bars, 100 µm. f, Summary data indicating the percentage of LY6D+ cells in matched human primary tumours and liver metastases. n = 17 biologically independent matched samples (7 data points are visible as 11 primary tumour samples have the same value (0) and are overlapping). g, UMAP plot of Juanito scRNA-seq dataset overlayed with iCMS designation. For comparison, AtrxKO, AtrxWT and AtrxKO/AtrxWT transcriptional expression scores are overlayed in the same data. h, Survival plot of Marisa CRC patient dataset separated on Atrx-based gene expression clusters. i, Volcano plot of HOMER TF enrichment analysis of TFs with differential motif accessibilities between HiSquam and HiCol signature tumours. Selected TF motifs in regions with reduced accessibility in HiSquam tumours highlighted in blue and TF motifs in regions with increased accessibility in HiSquam tumours highlighted in red. Data are the mean ± s.d. (b). P values were calculated using two-tailed Mann–Whitney tests (b), two-sided Fisher’s exact tests (d), two-tailed Wilcoxon matched-pairs signed-rank tests (f), log-rank (Mantel–Cox) tests (h) or two-sided Fisher’s exact test and adjusted for multiple comparisons with the Benjamini–Yekutieli method (i). NS, not significant.
We next examined how these lineage transitions evolve in advanced disease by staining matched primary and liver metastasis samples for ATRX, HNF4A, CDX2, LY6D and KRT5. Compared with our stage I–III tumour TMA dataset, squamous-like cells were more prevalent in stage IV tumours, which suggested that squamous-like plasticity becomes more prominent in later stage disease (Fig. 5c,d and Extended Data Fig. 10d,e). However, there was no difference in squamous cell abundance between primary tumours and their matched liver metastases, which indicated that this phenotype is already established before metastatic dissemination (Fig. 5e,f and Extended Data Fig. 10f,g). We also found a modest reduction in HNF4A and ATRX expression in liver metastases compared to their matched primary tumours, which provided further support for a progressive loss of colonic identity in advanced disease (Extended Data Fig. 10h–k).
To functionally validate these observations, we knocked out ATRX in human patient-derived CRC organoids and assessed their phenotype (Extended Data Fig. 10l). Consistent with our findings in mouse models and patient samples, deletion of ATRX led to a reduction in the expression of the colonic epithelial marker genes HNF4A and CDX1 and an increase in the squamous-like marker KRT5 (Extended Data Fig. 10m). Moreover, ATRXKO organoids showed an enhanced propensity for TGFβ-induced EMT, thereby providing direct functional evidence of a role for ATRX in maintaining colonic identity and restricting lineage plasticity in human CRC (Extended Data Fig. 10n,o).
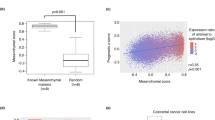
We next asked whether squamous-like gene expression was associated with disease phenotype and clinical outcome. The recently defined intrinsic consensus molecular subtypes (iCMS) separate CRC cases on the basis of epithelial gene expression patterns23. Two iCMS exist: iCMS2, which is associated with relatively good prognosis, and iCMS3, which shows evidence of metaplasia and is associated with poor prognosis23. We first analysed the prevalence of ATRX mutations in these subtypes and found an enrichment of ATRX alterations in iCMS3 tumours (Extended Data Fig. 11a). We next generated transcriptional signatures indicative of the squamous-like phenotype observed after Atrx loss in our CRC model (AtrxKO signature) and the colonic epithelial-like phenotype that is lost (AtrxWT signature). The AtrxKO signature contained markers of squamous cells (LY6D, KRT5, KRT13, SPRR1A and TRIM29) and those associated with the basal–squamous pancreatic ductal adenocarcinoma subtype and/or metaplastic CRC (CAV1, AQP3, FAM83A, F3 and AQP5)24,25. By contrast, the AtrxWT signature contained markers of colonic epithelium (HNF4A, ASCL2, CDX1 and CDX2). Analyses of previously published patient data26 using AtrxKO and AtrxWT signatures separated patients into three distinct clusters: those with high colonic epithelial and low squamous-like expression (HiCol cluster); those with reduced colonic epithelial and increased squamous-like expression (intermediate cluster); and those with high squamous-like and low colonic epithelial expression (HiSquam cluster) (Extended Data Fig. 11b). Analyses of these patients determined that intermediate and HiSquam cluster expression was significantly enriched in patients with right-sided (proximal) disease, mismatch-repair deficiency and BRAF mutation (Extended Data Fig. 11c–e). Patients with an advanced tumour stage were also enriched in HiSquam cluster expression, but this did not reach significance (Extended Data Fig. 11f). Consistent with these findings, intermediate and HiSquam cluster expression was associated with iCMS3 tumours (Extended Data Fig. 11g,h). Moreover, when mapped onto single tumours cells23, the ATRXKO score was enriched in iCMS3 cells and the ATRXWT score mapped to iCMS2 tumour cells (Fig. 5g). These analyses suggested an enrichment of lineage plasticity in aggressive, right-sided disease. To further investigate this finding, we analysed primary patient biopsy samples for the presence of cells with squamous-like phenotypes. FACS analysis confirmed the presence of tumour cells expressing both LY6D and EPCAM, thereby confirming the presence of this hybrid squamous-like state in primary human CRC samples (Extended Data Fig. 11i and Supplementary Table 9). Consistent with the transcriptional datasets, analysis of multiple samples revealed that the proportion of LY6D+ cells was significantly increased in right-sided disease (Extended Data Fig. 11j). To determine the prognostic relevance of lineage plasticity, we stratified patients on the basis of expression of our AtrxKO and AtrxWT expression signatures. Patients with high squamous-like and low colonic epithelial gene expression (HiSquam cluster) had significantly poorer overall survival than patients in the other two transcriptional clusters (Fig. 5h). Thus, the transcriptional phenotype controlled by Atrx can define tumour molecular phenotypes that are associated with aggressive disease and poor patient prognosis.
We then asked whether these transcriptional signatures are associated with alterations in chromatin structure by analysing previously published ATAC–seq data from primary human CRC27. To this end, we used our AtrxKO and AtrxWT signatures to stratify patients from The Cancer Genome Atlas (TCGA) with available RNA and ATAC–seq data into high, medium and low on the basis of the single-sample GSEA scores of the signatures. We then compared chromatin accessibility differences between the groups, HiSquam (high for AtrxKO and low for AtrxWT) and HiCol (low for AtrxKO and high for AtrxWT), and identified a set of genomic regions with altered accessibility across tumour samples (Extended Data Fig. 12a). TF motif enrichment analysis revealed loss of accessibility at sites that contained motifs for epithelial-specifying genes, including HNF4A and CDX2 in patients with HiSquam tumours (Fig. 5i and Supplementary Table 10). In line with our Atrx loss-of-function model, these results demonstrate that loss of activity of colonic lineage-specifying transcriptional regulators is associated with increased lineage plasticity and the expression of squamous-like phenotypic markers in human samples.
Discussion
Cellular plasticity is a key mechanism that drives cancer metastasis. As metastatic disease often arises in the absence of additional genetic perturbations, epigenetic mechanisms have been proposed as pivotal mediators. Here we identified the chromatin-remodelling enzyme ATRX as a crucial regulator of CRC plasticity and metastasis. Loss of Atrx gives rise to a highly plastic cell state, whereby cancer cells acquire both mesenchymal and squamous-like characteristics. Although EMT is a well-recognized aspect of tumour progression, the role of squamous trans-differentiation is poorly understood. Squamous cell transition states have been identified in cancers such as pancreatic, bladder and lung28,29,30, but have only recently been described in CRC. A recent study17 identified non-intestinal cell states, including squamous cells marked by KRT5 expression, in individuals with poor prognosis, metastatic CRC. Thus, the acquisition of non-canonical lineages seems to be an emerging phenomenon in this disease. In this context, our study provides mechanistic insight into how these cell states arise, and places the loss of colonic lineage fidelity and the activity of lineage specifying TFs at the heart of this regulation.
Mechanistically, we showed that loss of chromatin accessibility and enhancer activity at colonic lineage specifying genes mediates loss of colonic epithelial identity. Across all our analyses, the loss of colonic identity was highly notable, which suggests that this is the initiating event controlled by ATRX that, when lost, enables the induction of plasticity. Indeed, the gain in non-canonical lineages, both mesenchymal and squamous, was less evident at the chromatin level. This finding indicates that rather than being driven towards an alternative lineage, once colonic identity is ‘softened’, the promotion of non-canonical lineages may occur in a stochastic manner, perhaps influenced by environmental factors. This hypothesis is supported by previous studies of pancreatic cancer that have highlighted the importance of the tumour microenvironment in mediating cellular transitions25. Our functional studies defined HNF4A as a key protector of the colonic lineage, thereby further emphasizing the importance of lineage-defining TFs in protecting against the acquisition of non-canonical cell states. We provided evidence of a similar mechanism in patient samples, whereby a squamous-like gene expression signature based on genes that are overexpressed after Atrx loss predicts aggressive disease and poor prognosis. Notably, tumours that exhibited this squamous-like gene expression pattern had reduced expression of colonic epithelial genes and showed changes in chromatin accessibility indicative of loss of HNF4A activity. Similar findings in other cancers, such as pancreatic, indicate that loss of activity of lineage-specifying TFs may be a ubiquitous mechanism that enables cellular plasticity in aggressive cancers31. Together, our study shed light on the mechanisms that facilitate metastasis and promote cellular plasticity. Further understanding of how these processes contribute to cancer progression will pave the way for potential opportunities for therapeutic intervention.
Methods
Mouse studies
All in vivo experiments were performed in accordance with the UK Home Office regulations (project licence: PP9016178, PP7510272 and PP3908577) and were subject to local ethics review by the animal welfare and ethics board of the University of Edinburgh and University of Glasgow. Mice were housed under a 12-h light–dark cycle at a temperature ranging from 19 to 23 °C. Ambient humidity was 55 ± 10%. Standard diet and water were available ad libitum. Work was performed on the C57BL/6J and the CD1 nude background available from Charles River Laboratories. For the lung metastatic model, 500,000 AKP control or AKP AtrxKO cells were resuspended in 100 µl PBS and injected into the tail vein of female CD1 nude mice. For the liver metastatic model, C57BL/6J mice were anaesthetized with isoflurane and splenic access was achieved using laparotomy. Around 500,000 cells were injected in 50 µl PBS, following which, the incision was closed with staples. Subcutaneous injections into female CD1 nude mice were performed with 50,000 AKP control, AKP AtrxKO or AKP Hnf4aKO cells resuspended in 100 µl basement membrane extract (BME). Subcutaneous injections into female CD1 nude mice were performed with 12,000 flow cytometry-sorted cells resuspended in 100 µl BME.
Colonic submucosal injections were performed using a Karl Storz TELE PACK VET X LED endoscopic video unit. AKP control, AKP AtrxKO, BPN control and BPN AtrxKO organoids were dissociated by mechanical pipetting and then were washed with PBS before being injected orthotopically in female CD1 nude mice. Approximately 500 organoids in 70 μl PBS were injected in a single injection. The BPN organoid line (male) was generated in the laboratory of O.J.S.
In accordance with the respective licence and study protocol, mice were humanely euthanized at the reported experimental end points (15 mm diameter for subcutaneous injection) as defined in the relevant licensing documents.
In accordance with the 3Rs (replacement, reduction and refinement), the smallest sample size was chosen that could give a significant difference.
C57BL/6J or CD1 nude mice 6–12 weeks of age were randomly grouped for transplantation experiments. All mice received the same number of cells of a different genotype or phenotype. Experimental groups were determined by the genotype of injected cells (for example, AKP control versus AKP AtrxKO). Investigators were blinded to the genotype of tumours when monitoring for clinical signs, when carrying out histological analyses and during data collection. IHC analysis of tumour histology was carried out using QuPath software, with the investigator blinded to the tumour genotype.
Histopathology, IHC and immunofluorescence
Tissues were fixed overnight in formaldehyde 4% stabilized, buffered (VWR) and then transferred to 70% ethanol. Tissue processing was done using a Tissue-TeK VIP infiltration processor (Sakura) followed by embedding in paraffin wax. Tissues were sectioned at 5 µm thickness, dried at 37 °C and then deparaffinized and rehydrated. For each sample, one section was stained with haematoxylin and eosin. Additional sections were used for either standard IHC or immunofluorescence analyses. The following primary antibodies were used: KRT5 (rabbit; Abcam 52635; 1:200 or chicken; BioLegend 905903; 1/200); LY6D (rabbit; Atlas HPA024755; 1:200); TWIST (mouse; Santa Cruz 81417, 1:200); EPCAM (rabbit; Abcam 71916; 1:200); HNF4A (rabbit; CST 3113; 1:500); ATRX (mouse; Sigma MABE1798; 1:500); β-catenin (mouse; BD 610154; 1:50); E-cadherin (rabbit; CST 3195, 1:200); and CDX2 (mouse; Atlas AMAb 91828, 1:1,000). IHC secondary detection was achieved using EnVision+ System-HRP labelled polymer (Dako). Positive signals were visualized using DAB substrate (Epredia and 2b scientific) for 5–10 min. Sections were counterstained with haematoxylin. For immunofluorescence studies, the following secondary antibodies (1:400) were used: anti-rabbit-594 (Invitrogen, A21207); anti-chicken-488 (Invitrogen, A78948); anti-streptavidin-647 (Invitrogen, S32357); and anti-rabbit-488 (Abcam, 150073) Representative images are shown for each stain. The slides were digitized using a Nanozoomer digital slide scanner (Hamamatsu) with NDP.view2plus software and analysed using QuPATH (v.0.2.3). The commercial colon cancer human tissue array used was CO804b (Biomax) (Extended Data Fig. 1b–d). The TMA in Fig. 5 and Extended Data Fig. 10 includes patients with stage I–III CRC who underwent potentially curative resection between 1997 and 2013 at Glasgow Royal Infirmary. Three cores were taken from each donor block at the invasive edge, capturing a small amount of tumour and the environment around it. Patient tissue access was authorized by the NHS Greater Glasgow and Clyde Biorepository under their NHS Research Ethics Committee, with ethics approval granted in biorepository application number 845, West of Scotland Ethics 22/WS/0207 in accordance with recognized ethical guidelines as described in the Declaration of Helsinki. Overall, 17 patients undergoing synchronous resection of primary CRC and CRC liver metastases with curative intent between April 2002 and June 2010 at Glasgow Royal Infirmary were analysed. Patient tissue access was authorized by the NHS Greater Glasgow and Clyde Biorepository under their NHS Research Ethics Committee, with ethical approval granted in biorepository application number 357, West of Scotland Ethics 22/WS/0207 in accordance with recognized ethical guidelines as described in the Declaration of Helsinki.
H-scores were generated for HNF4A, ATRX and CDX2 tumour cell expression using QuPath-0.2.3. Tumour cores with >2% tumour cells positive for LY6D and KRT5 expression were designated as LY6D+ and KRT5+, respectively. Tumour cores with <2% tumour cells positive for LY6D or KRT5 expression were designated as LY6D– and KRT5–, respectively. Analysis was performed using QuPath-0.2.3. H-scores were generated for CDH1 cell expression using QuPath-0.2.3.
Mouse organoid studies
Organoids were resuspended in Cultrex reduced growth factor BME type 2 (Bio-Techne), plated in a 10 µl drop on a culture plate and cultured in organoid culture medium containing advanced DMEM–F12 (Gibco) medium supplemented with 100 units ml–1 penicillin, 100 µg ml–1 streptomycin, 2 mM l-glutamine, 10 mM HEPES (all from Life Technologies), 1 ml Primocin (Invivogen), N2, B27 (both from Gibco), 50 ng ml–1 EGF (Peprotech) and 1% Noggin conditioned medium. The Noggin-producing cell line was a gift from H. Clevers’ group (Hubrecht Institute). Organoids were passaged by mechanical fragmentation with a p1000 and p200 pipette. All organoids were grown in a humidified incubator at 37 °C supplemented with 5% CO2. Single cells were generated by incubating organoids in TrypLE Express (Life Technologies) for 15 min at 37 °C and passed through a 40 µm strainer. Cell counting was performed using a Countess II automated cell counter (Invitrogen). To study the effect of TGFβ, dissociated cells (10,000 (or 5,000 for the Tgfbr2KO) cells in 10 µl BME) were cultured in organoid culture medium in the presence of 5 ng ml–1 TGFβ (Peprotech). TGFβ treatment in Extended Data Fig. 3 was performed using a different range of concentrations as specified in the figure. On day 3 the medium was replaced with fresh organoid culture medium containing TGFβ. Cells were collected on day 13 for RNA extraction. Alternatively, cells were stained with calcein (Abcam) or phalloidin-568 (VWR) within 14 days of treatment.
For ITGA5+ cell sorting and TGFβ treatment, AKP control and AKP AtrxKO organoids were digested as described above, and ITGA5 FACS was carried out as described in the FACS section below. In brief, 10,000 ITGA5+ or ITGA5– cells were plated in 10 µl BME and treated with 5 ng ml–1 TGFβ as described above. On day 13, organoids were fixed with 4% paraformaldehyde, stained with phalloidin-568 and analysed for the presence of spindle-like structures.
For the tail-vein injections, organoids were dissociated in TrypLE Express (Life Technologies) supplemented with a Rho kinase inhibitor (Y-27632, Tocris) for 15 min at 37 °C, passed through a 40 µm strainer, counted and resuspended in PBS. For splenic injections, organoids were collected, washed in PBS, mechanically fragmented by vigorous pipetting and then incubated for 7 min at 37 °C in 0.25% trypsin in EDTA–PBS. After quenching of trypsinization by immersion in 10% FBS, cells were passed through a 40 µm strainer, counted using a haemocytometer and resuspended in PBS to achieve a final volume of 1 × 107cells per ml. Cells were routinely tested for mycoplasma contamination.
Human patient-derived organoids
The patient-derived organoids used in this study were generated by F.V.N.D. and M.G.D. MD20043 is an 81-year-old man with stage 4 rectal cancer: T3, N2, M1 (where ‘T’ is tumour, ‘N’ is nodes and ‘M’ is metastases). Ethics approval for human CRC organoid derivation was carried out under NHS Lothian Ethical Approval Scottish Colorectal Cancer Genetic Susceptibility Study 3 (SOCCS3) (REC reference: 11/SS/0109). The patient provided fully informed consent for the use of their tissues.
Human organoid culture medium and ATRX KO generation
Human carcinoma organoids were cultured in advanced DMEM–F12 (Gibco) medium supplemented with 100 units ml–1 penicillin, 100 µg ml–1 streptomycin, 2 mM l-glutamine, 10 mM HEPES (all from Life Technologies), 1 ml Primocin (Invivogen), 1% Noggin conditioned medium (The Noggin-producing cell line was a gift from H. Clevers’ group, Hubrecht Institute), B27 (Gibco), 50 ng ml–1 EGF (Peprotech), 10 nM gastrin (Sigma), 10 nM PGE2 (Tocris), 10 mM nicotinamide (Sigma), 10 µM SB202190 (Sigma), 600 nM A83-01 (Biotechne) and 12.5 mM N-acetylcysteine (Sigma).
For transduction conditions, human organoids were pretreated with IntestiCult (Stem Cell Tech) organoid growth medium and 1 mM valproic acid (Merk) for 48 h after 2 days post-split (day 1). On transduction day (day 1), human organoids were digested into a single-cell suspension in TrypLE Express (Life Technologies) with 10 µM Y-27632 (Tocris) for 8 min at 37 °C with mechanical dissociation every 4 min. Single-cell suspensions were combined with viral particles containing a non-targeting or ATRXKO sgRNA (hATRX: 5′-GCTATAAACAGAAAAAGAAA-3′; pLentiV2-Addgene) and placed on a BME layer. On day 2, medium was changed into IntestiCult supplemented with 1 mM valproic acid and 10 µM Y-27632. On day 3, antibiotic selection started with IntestiCult supplemented with 10 µg ml–1 blasticidin (Gibco) for 3 weeks. IntestiCult medium was used exclusively during transduction and the single-cell stage of organoid development for clone selection. Otherwise, the above-described medium was used for maintaining the organoids.
In vitro drug treatment and cell-proliferation assay
Organoids were dissociated in TrypLE Express (Life Technologies) for 15 min at 37 °C, passed through a 40 µm strainer and resuspended in BME. Next, 1,000 single cells in 10 µl BME were plated in 24-well plates and treated with 250 nM JQ1 (Stratech), 0.25 ng ml–1 IFNγ (Thermo Fisher Scientific), 10 nM FK228 (Stratech) or (0.5 ng ml–1) TNF (Peprotech) for 7 days.
For Resazurin cell viability assays, Resazurin (R&D systems) was added at a volume equal to 10% of the cell culture medium volume and incubated at 37 °C. Fluorescence was read using 544 nm excitation and 590 nm emission wavelengths.
RNA extraction, RT–qPCR and RNA-seq
Total RNA was isolated using a RNeasy Mini kit (Qiagen) accordingly to the manufacture’s protocol. RNA was then subjected to DNA-free DNase treatment (Invitrogen). cDNA was generated using 1 µg RNA by reverse transcription using qScript cDNA SuperMix (Quantabio). RT–qPCR was performed using SYBR Select master mix (Applied Biosystems). Ct values were normalized to β-actin or 18S rRNA. The ΔΔCt method was used to calculate relative gene expression values. Oligonucleotides used in this study are listed in Supplementary Table 11. For the RNA-seq experiments, RNA integrity was evaluated using an Agilent 2200 Bioanalyser. Truseq mRNA-seq libraries were prepared from total AKP and AKP ATRXKO RNA, and these were then sequenced using NovaseqS1 Illumina sequencing at Edinburgh Genomics Facility. AKP and AKP Hnf4aKO libraries were prepared from 100 ng of each total RNA sample using a NEBNEXT Ultra II Directional RNA Library Prep kit (NEB 7760) and the Poly-A mRNA magnetic isolation module (NEB E7490) according to the provided protocol. Sequencing was performed on a NextSeq 2000 platform (Illumina, 20038897) using NextSeq 2000 P3 reagents (200 cycles) (20040560). RNA-seq analysis was carried out using the RaNA-seq pipeline with default settings32.
Western blotting
Cells were lysed using RIPA buffer (Sigma) supplemented with 1% phosphatase and protease inhibitors (Sigma). Protein concentration was measured using a BCA Protein Assay kit (Pierce). A total of 20 µg protein lysate was resuspended in 4× LDS sample buffer (Invitrogen) supplemented with sample reducing agent (Invitrogen) and denatured at 100 °C for 5 min. Proteins were separated by electrophoresis on NuPAGE 3–8% Tris-acetate protein gels (Invitrogen) using Tris-acetate buffer and blotted onto an activated PVDF or nitrocellulose (Cytiva) membrane at 100 V for 1.15 h. Membranes were incubated in blocking solution (5% milk, 0.1% Tween-20–PBS) for 1 h at room temperature, and then in primary antibody. The following primary antibodies were used: β-actin (Cell Signalling Technology, 1:5,000); ATRX (MABE1798, Sigma; 1:500); and HNF4A (C11F12, Cell Signalling Technology, 1:,1,000). After 3× 10-min washes in 0.1% Tween-20–PBS, the membrane was incubated in HRP-linked secondary antibody for 1 h at room temperature. The following secondary antibodies were used: anti-rabbit or anti-mouse IgG HRP-linked (Cell Signalling Technology, 1:5,000). Following 3× 10-min washes in 0.1% Tween-20–PBS, antibody signals were detected by using ECL Plus Western blotting substrate (Pierce) and visualized using an ImageQuant 800 (GE Healthcare). Full scans are provided in Supplementary Information 1 and 2.
CRISPR–Cas9 genome editing
sgRNAs (mAtrx: 5′-ACGGCGCATTAAGGTTCAAG-3′; mHnf4a 5′-CGGGCCACCGGCAAACACTA-3′, mTgfbr2: 5′-AAGCCGCATGAAGTCTGCG-3′, non-targeting controls 5′- GCTTTCACGGAGGTTCGACG-3′ or 5′-ATGTTGCAGTTCGGCTCGAT-3′) were cloned individually into lentiCRISPR v.2 plasmids (Addgene) following Addgene’s protocol. Lentiviral particles were generated using HEK293T cells (provided by J. C. Acosta (IGMM, Edinburgh), originally obtained from the American Type Culture Collection): 10 µg gene-specific lentiviral vector was mixed with 7.5 µg lentiviral packaging vector psPAX2 and 2.5 µg envelope-protein-producing vector pCMV-VSV-G (both from Addgene) and transfected into HEK293T cells in a 10 cm2 dish using polyethylenimine as the transfection reagent (Polysciences). After 48 h, the supernatant medium was filtered using a 0.45 µm syringe filter and concentrated using a Lenti-X concentrator (Takara Bio). Lentiviral transduction was carried out as previously described33. In brief, AKP organoids were expanded and cultured in organoid culture medium supplemented with 10 µM Rho kinase inhibitor (Y-27632, Tocris) and 1 mM valproic acid (Sigma) for 48 h. Spheroids were enzymatically dissociated with StemPro Accutase cell dissociation reagent (Gibco) supplemented with 10 µM Y-27632 for 3 min at 37 °C. Dissociated organoids were then washed twice with advanced DMEM–F12 medium. Cells were counted using a Countess II Automated cell counter (Invitrogen). Next, 5 × 105 cells were plated on a 150 µl bed of BME in a 6-well plate in the presence of Y-27632, 1 mM valproic acid and 4 µg ml–1 polybrene (Sigma). Virus was removed 24 h after transduction, and adhered organoids were overlaid with 150 µl BME. To select transduced cells, 2 µg ml–1 of puromycin or 10 µg ml–1 blasticidin (both from Gibco) was added to the organoid culture medium supplemented with 10 µM Y-27632. Multiple deleted clones were generated. Editing of clonal lines was confirmed by genomic sequencing or western blotting.
TissueEnrich analysis
TissueEnrich analysis34 (https://tissueenrich.gdcb.iastate.edu/) was carried out using the web-based tool with the following settings: gene symbol, Homo sapiens, Human Protein Atlas, All. Lists of gene upregulated or downregulated by >2 fold in the AKP versus AKP AtrxKO dataset were used for input. In Fig. 2g, TissueEnrich analysis was performed on the list of genes upregulated in scRNA-seq cluster 4 and cluster 15 (fold change > 1.5).
ATRX mutation analysis in CRC transcriptional subtypes
For ATRX mutation enrichment analysis, we used previously designated CRIS subtypes14 and analysed TCGA mutational data for ATRX mutations for which the CRIS designation is known. The number of tumours carrying ATRX mutations was calculated for the CRIS-B subtype and compared with all other subtypes using Fishers’ exact tests. For survival analysis, only CRIS-B tumours were analysed using TCGA survival data downloaded from the TCGA data portal. For iCMS, the same analysis was carried out but with TCGA data separated on the basis of iCMS designation.
HNF4A overexpression
HNF4A overexpression was generated using the pGCDNsam-HNF4A-IRES-GFP plasmid (Addgene). In brief, AKP Atrx organoids were transduced as above. To select transduced cells, FACS analysis for GFP positivity was performed.
Collection of human samples and processing for FACS
Normal colorectal mucosa and tumour were sampled from freshly resected surgical specimens from patients diagnosed with CRC. Ethics approval was carried out under NHS Lothian Ethical Approval Scottish Colorectal Cancer Genetic Susceptibility Study 3 (SOCCS3 REC: 11/SS/0109, IRAS: 9556). All patients provided fully informed consent for the use of their tissues. Tissues were cut into small pieces and then incubated in Advanced DMEM–F12 supplemented with 1 mg ml–1 collagenase type IV (Sigma), 0.5 mg ml–1 hyaluronidase (Sigma) and 10 µM Y-27632 (Tocris) at 37 °C with vigorous shaking until the tissue was completely disaggregated (60–90 min). The digested reaction was then filtered through a 70 µm cell strainer. The filtered cells were centrifuged at 500g for 5 min, washed twice in Advanced DMEM–F12 and once in 0.1% BSA in PBS. Single-cell suspensions were then analysed by FACS.
FACS
Pelleted organoids were resuspended in 1 ml TrypLE Express (Gibco) and incubated at 37 °C for 15 min. Cells were vigorously dissociated by pipetting, resuspended in 10 ml advanced DMEM–F12, passed through a 40 µm cell strainer and centrifuged at 300g for 5 min at 4 °C. Single cells were washed with 0.1% BSA in PBS and stained with the following antibodies: EPCAM–APC (BioLegend, 118213; 1:200); LY6D–PE (BioLegend, 138603; 1:200) or LY6D–APC (Miltenyi, 130-115-313; 1:50); and ITGA5–PE (BioLegend, 103805; 1:200). Human single-cell suspensions were stained with EPCAM–APC (BioLegend, 324207; 1:50) and LY6D–FITC (Cusabio Biotech, CSB-PA613492LC01HU; 1:50). Cells were then washed twice in 0.1% BSA in PBS before being subjected to FACS (BD FACSARIA II/BD LSR-Fortessa X-20). Single viable cells were gated by negative staining for DAPI. The gating strategy is provided in Supplementary Information 3. Analyses were performed using FlowJo (v.10.8) software.
scRNA-seq data processing
Raw sequencing reads were processed and aligned to the mouse reference genome (mm10) using the 10x Genomics CellRanger pipeline (v.7.2.0). The gene expression matrices obtained from CellRanger were analysed using the R package Seurat (v.5). Cell barcodes with <200 unique genes and >10% mitochondrial gene expression were removed. The filtered matrices were normalized to the total unique molecular identifier counts per cell, and cell cycle effects were regressed out. Data integration was performed with Harmony, and the subsequent Louvain clustering resulted in 18 clusters. Differentially expressed genes between clusters were obtained using Wilcoxon rank-sum tests implemented by the FindAllMarkers function in Seurat.
ATAC–seq
ATAC–seq was performed using an Active Motif commercially available kit as per the manufacturer’s instructions. In brief, 50,000 cells were lysed with ATAC lysis using a pestle and dounced slowly for 25 strokes on ice followed by Tn5 tagmentation for 30 min at 37 °C. Size selection was performed using the SPRIselect protocol (Beckam). After indexing and PCR amplification, DNA libraries were multiplexed and sequenced with an Illumina Nextseq 2000 on a 100-cycles kit by Edinburgh Clinical Research Facility–Wellcome Trust CRF.
ATAC–seq analysis
Raw sequencing output fastq files were inputed into an nf-core ATAC–seq pipeline using default pipeline settings (https://nf-co.re/atacseq/2.1.2)35. ATAC–seq library quality was assessed for the presence of contaminating mitochondrial DNA sequences. Pipeline output bigWig files were used for creating locus-specific genome images from the IGV browser (https://www.igv.org/). ATAC–seq peaks were analysed for differential enrichment between samples using DESeq2 (ref. 36). Principal component analysis was performed on tables of ATAC-seq read counts (DESeq2 output) using base R scripts. Subsequently, DESeq2 ATAC–seq read counts were input into the monaLisa R package37 to determine the presence of differentially enriched TF-binding or DNA-binding site motifs between sample groups.
CUT&RUN
Cells were digested with TrypLE Express at 37 °C for 15 min. Next, 1 × 106 viable cells were fixed in PBS containing 0.1% formaldehyde at room temperature for 1 min. Formaldehyde was quenched with glycine at 0.125 M, followed by 5 min of incubation at room temperature. Fixed cells were pelleted and washed twice with room temperature wash buffer (20 mM HEPES, 150 mM NaCl, 0.5 mM spermidine (Sigma-Aldrich), 1% Triton X-100, 0.05% SDS and 1× protease inhibitor cocktail (Roche)). Cells were pelleted and resuspended in 100 µl wash buffer. The cell suspension was added at a 10:1 ratio to concanavalin A beads (CST) activated as per the manufacturer’s instructions, gently mixed and incubated at room temperature for 20 min. The cell–bead slurry was placed on a magnetic rack until clear, the supernatant removed and samples resuspended in cold antibody buffer (20 mM HEPES, 150 mM NaCl, 0.5 mM spermidine, 0.0025% digitonin (Thermo Fisher), 2 mM EDTA, 1% Triton X-100, 0.05% SDS and 1× protease inhibitor cocktail (Roche)). The following antibodies were added and incubated at 4 °C overnight: XP isotype control DA1E (rabbit; CST 1:10) or anti-histone H3, acetyl K27 (rabbit; Abcam 1:250). Samples were washed twice with digitonin buffer (20 mM HEPES, 150 mM NaCl, 0.5 mM spermidine, 0.0025% digitonin, 1% Triton X-100, 0.05% SDS and 1× protease inhibitor cocktail (Roche)) and resuspended in digitonin buffer. 1× CUTANA pAG-MNase (EpiCypher) was added and incubated for 10 min at room temperature. Samples were washed twice with digitonin buffer. Chromatin was cleaved by the addition of 2 mM CaCl2 and incubation for 2 h at 4 °C. Digestion was halted by the addition of stop buffer (340 mM NaCl, 20 mM EDTA, 4 mM EGTA and 50 µg ml–1 glycogen (ThermoFisher)) and chromatin released by incubation at 37 °C for 10 min. The supernatant was collected and SDS added to a final concentration of 0.09%. Proteinase K (CST) was added and incubated overnight at 55 °C. DNA was purified using MaXtract High Density columns (Qiagen) as per the manufacturer’s instructions and precipitated with GlycoBlue (ThermoFisher) as per the manufacturer’s instructions. DNA was eluted in TE buffer (ThermoFisher). Samples were quantified and assessed for size distribution using an Agilent 2100 Electrophoresis Bioanalyser Instrument with a DNA HS kit (Agilent Technologies). Libraries were generated using a Simple-ChIP DNA library prep kit (CST) and quantified by fluorometry using a Qubit dsDNA HS assay and assessed for size distribution on an Agilent Bioanalyser with a DNA HS kit. Next, 100 bp paired-end sequencing was performed on a NextSeq 2000 platform (Illumina) using a NextSeq 1000/2000 P1 reagents (300 cycles) kit.
CUT&RUN data analysis
The nf-core CUT&RUN analysis pipeline (https://doi.org/10.5281/zenodo.7715959) was used to process CUT&RUN data using default parameters. Reads were normalized to spike in DNA (CST) and aligned to the mm10 reference genome. Regions in the mm10 ENCODE blacklist were removed38. MACS2 (ref. 39) was used for peak calling and peak calls were normalized to IgG controls. Differential analysis was conducted using DiffBind40 with default parameters. BEDTools41 was used to identify overlapping regions of significant difference in the H3K27ac CUT&RUN with the ATAC dataset.
TCGA patient stratification based on ATRX transcriptional signatures
Bulk RNA transcripts per kilobase million values for patients with colorectal adenocarcinoma from TCGA27 were obtained from the Genomic Data Commons Portal and were log2 normalized. Patients who had ATAC–seq data (n = 36) were stratified into high, medium or low for AtrxKO and AtrxWT signatures. This was performed by calculating the single-sample GSEA score using GSVA in R and selecting the samples above the third, second or first quantile for the respective category. These patients were also assigned an iCMS class as previously described23.
Differential analysis of TCGA bulk ATAC–seq data
Normalized ATAC–seq peak counts were obtained from the Genomic Data Commons Portal for 77 samples from 36 patients with colorectal adenocarcinoma; details on the normalization can be found the original publication27. In brief, the peak counts matrix was normalized using ‘cpm(matrix, log = TRUE, prior.count = 5)’ in edgeR followed by quantile normalization using normalize.quantiles of preprocessCore in R. A SummarizedExperiment object was created with the normalized count matrix and metadata for the patients. A two-tailed t-test was used to identify the peaks that have a significantly different mean count between the samples from patients categorized as HiSquam (high for AtrxKO and low for AtrxWT) or HiCol (low for AtrxKO and high for AtrxWT). The selection of differential peaks was based on FDR < 0.01 and Δlog2counts > 1 cut-off values. Counts from all samples for the set of significantly differential peaks were plotted into a heatmap using ComplexHeatmap in R.
HOMER TF motif enrichment
TF motif enrichment analysis was performed on the set of peaks that were differentially accessible for patients categorized as HiSquam or HiCol by using HOMER. The peaks were first annotated using ChIPeakAnno and then formatted into HOMER input style. The analysis was performed with the command findMotifsGenome.pl, genome “hg38” and “-size 200 -mask” as options. TF motif enrichment was presented as previously described42.
Analysis of bulk gene expression data with Atrx WT and Atrx KO signature genes
Normalized gene expression data of samples from patients with CRC were obtained from the Gene Expression Omnibus (accession GSE39582), and 557 patients with relapse survival information were used for the downstream analysis. HiCol, intermediate and HiSquam groups were identified by hierarchical clustering using AtrxWT and AtrxKO signature genes. The relapse-free survival of three patient groups was determined by Kaplan–Meier survival with log-rank tests.
Identification of Atrx WT and Atrx KO signatures from single-cell resolution
To understand the association between ATRXKO-driven gene expression and iCMS classification at the single-cell level, we performed the analysis using scRNA-seq data after downloading from syn26844071 (ref. 23) and used tumour cells with iCMS classification from primary tumour tissue samples after excluding patients with low numbers of cells. The dimension reduction analysis was performed using 715 iCMS-associating genes, and patient batch was corrected using Harmony43. AtrxWT and AtrxKO scores for each cell were calculated using the average expression level of each signature and then subtracted with the aggregated expression of the control gene score sets. The control gene score was calculated using the average expression of 100 randomly selected genes, replicated 10 times.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 9. Statistical tests used are indicated in figure legends and exact P values shown throughout.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The CUT&RUN, ATAC–seq, scRNA-seq and RNA-seq data generated in this study have been deposited into the Genome Sequence Archive (https://ngdc.cncb.ac.cn/gsa/) website with the following accession numbers: CRA024850 (H3K27ac AKP versus AKP ATRX); CRA024849 (ATAC-seq AKP versus AKP ATRX); CRA024816 (scRNA-seq AKP versus AKP ATRX); CRA024804 (RNA-seq AKP versus AKP HNF4A); and CRA024763 (RNA-seq AKP versus AKP ATRX). Source data are provided with this paper.
References
Hanahan, D. Hallmarks of cancer: new dimensions. Cancer Discov. 12, 31–46 (2022).
Pérez-González, A., Bévant, K. & Blanpain, C. Cancer cell plasticity during tumor progression, metastasis and response to therapy. Nat. Cancer 4, 1063–1082 (2023).
Birkbak, N. J. & McGranahan, N. Cancer genome evolutionary trajectories in metastasis. Cancer Cell 37, 8–19 (2020).
Martínez-Jiménez, F. et al. Pan-cancer whole-genome comparison of primary and metastatic solid tumours. Nature 618, 333–341 (2023).
Priestley, P. et al. Pan-cancer whole-genome analyses of metastatic solid tumours. Nature 575, 210–216 (2019).
Terekhanova, N. V. et al. Epigenetic regulation during cancer transitions across 11 tumour types. Nature 623, 432–441 (2023).
Alexander, S. & Friedl, P. Cancer invasion and resistance: interconnected processes of disease progression and therapy failure. Trends Mol. Med. 18, 13–26 (2012).
Lambert, A. W., Pattabiraman, D. R. & Weinberg, R. A. Emerging biological principles of metastasis. Cell 168, 670–691 (2017).
McDonald, O. G. et al. Epigenomic reprogramming during pancreatic cancer progression links anabolic glucose metabolism to distant metastasis. Nat. Genet. 49, 367–376 (2017).
Cammareri, P. & Myant, K. B. Be like water, my cells: cell plasticity and the art of transformation. Front. Cell Dev. Biol. 11, 1272730 (2023).
Garraway, L. A. & Lander, E. S. Lessons from the cancer genome. Cell 153, 17–37 (2013).
Shen, H. & Laird, P. W. Interplay between the cancer genome and epigenome. Cell 153, 38–55 (2013).
Cui, J. et al. MLL3 loss drives metastasis by promoting a hybrid epithelial–mesenchymal transition state. Nat. Cell Biol. 25, 145–158 (2023).
Isella, C. et al. Selective analysis of cancer-cell intrinsic transcriptional traits defines novel clinically relevant subtypes of colorectal cancer. Nat. Commun. 8, 15107 (2017).
Roper, J. et al. In vivo genome editing and organoid transplantation models of colorectal cancer and metastasis. Nat. Biotechnol. 35, 569–576 (2017).
Torang, A. et al. Enterocyte-like differentiation defines metabolic gene signatures of CMS3 colorectal cancers and provides therapeutic vulnerability. Nat. Commun. 16, 264 (2025).
Moorman, A. et al. Progressive plasticity during colorectal cancer metastasis. Nature 637, 947–954 (2025).
Cañellas-Socias, A. et al. Metastatic recurrence in colorectal cancer arises from residual EMP1+ cells. Nature 611, 603–613 (2022).
Mustata, R. C. et al. Identification of Lgr5-independent spheroid-generating progenitors of the mouse fetal intestinal epithelium. Cell Rep. 5, 421–432 (2013).
Berest, I. et al. Quantification of differential transcription factor activity and multiomics-based classification into activators and repressors: diffTF. Cell Rep. 29, 3147–3159 (2019).
Chen, L. et al. The nuclear receptor HNF4 drives a brush border gene program conserved across murine intestine, kidney, and embryonic yolk sac. Nat. Commun. 12, 2886 (2021).
Chen, L. et al. A reinforcing HNF4–SMAD4 feed-forward module stabilizes enterocyte identity. Nat. Genet. 51, 777–785 (2019).
Joanito, I. et al. Single-cell and bulk transcriptome sequencing identifies two epithelial tumor cell states and refines the consensus molecular classification of colorectal cancer. Nat. Genet. 54, 963–975 (2022).
Chen, B. et al. Differential pre-malignant programs and microenvironment chart distinct paths to malignancy in human colorectal polyps. Cell 184, 6262–6280 (2021).
Raghavan, S. et al. Microenvironment drives cell state, plasticity, and drug response in pancreatic cancer. Cell 184, 6119–6137 (2021).
Marisa, L. et al. Gene expression classification of colon cancer into molecular subtypes: characterization, validation, and prognostic value. PLoS Med. 10, e1001453 (2013).
Corces, M. R. et al. The chromatin accessibility landscape of primary human cancers. Science https://doi.org/10.1126/science.aav1898 (2018).
Bailey, P. et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 531, 47–52 (2016).
Han, X. et al. Transdifferentiation of lung adenocarcinoma in mice with Lkb1 deficiency to squamous cell carcinoma. Nat. Commun. 5, 3261 (2014).
Wang, M. et al. Acquired semi-squamatization during chemotherapy suggests differentiation as a therapeutic strategy for bladder cancer. Cancer Cell 40, 1044–1059 (2022).
Brunton, H. et al. HNF4A and GATA6 loss reveals therapeutically actionable subtypes in pancreatic cancer. Cell Rep. 31, 107625 (2020).
Prieto, C. & Barrios, D. RaNA-seq: interactive RNA-seq analysis from FASTQ files to functional analysis. Bioinformatics 36, 1955–1956 (2020).
Hall, A. E. et al. RNA splicing is a key mediator of tumour cell plasticity and a therapeutic vulnerability in colorectal cancer. Nat. Commun. 13, 2791 (2022).
Jain, A. & Tuteja, G. TissueEnrich: tissue-specific gene enrichment analysis. Bioinformatics 35, 1966–1967 (2019).
Ewels, P. A. et al. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 38, 276–278 (2020).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Machlab, D. et al. monaLisa: an R/Bioconductor package for identifying regulatory motifs. Bioinformatics 38, 2624–2625 (2022).
Amemiya, H. M., Kundaje, A. & Boyle, A. P. The ENCODE blacklist: identification of problematic regions of the genome. Sci. Rep. 9, 9354 (2019).
Zhang, Y. et al. Model-based analysis of ChIP–seq (MACS). Genome Biol. 9, R137 (2008).
Ross-Innes, C. S. et al. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer. Nature 481, 389–393 (2012).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Xu, B. et al. Acute depletion of CTCF rewires genome-wide chromatin accessibility. Genome Biol. 22, 244 (2021).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289–1296 (2019).
Acknowledgements
This work was funded by the Cancer Research UK (CRUK) under a Career Development Fellowship (A19166 to K.B.M.) and a Small Molecule Drug Discovery Project Award (A25808 to K.B.M.), a European Research Council under Starting Grant (COLGENES–715782 to K.B.M.) and the MRC (project grant MR/X008762/1 to K.B.M.). C.W.S. is funded by a UKRI Future Leaders Fellowship (MR/W007851/1). O.J.S. is funded by a CRUK ACRCelerate: Colorectal Cancer Stratified Medicine Network Accelerator Award (A26825). We thank technical staff at the University of Edinburgh’s Institute of Genetics and Cancer (IGC) for providing support for some of the experiments; staff at the Flow Cytometry service for support with FACS; the animal technicians at the Biomedical Research Facility (BRF) facility for animal husbandry support; NHS Lothian staff and patients for support obtaining human tissue biopsies; B. Callaghan for helping N.P. with cell proliferation assays; J. Roper and O. Yilmaz for the gift of AKP organoids; and G. Brien for advice on CUT&RUN assays.
Author information
Authors and Affiliations
Contributions
The project was conceived and experiments planned by M.R., P.C. and K.B.M. Experiments were conducted by P.C., M.R., N.P., J.F., M.W., C.V.B., S.Ö.-G.P., N.J.D., J.Q., A.B.A. and K.B.M. Bioinformatic analyses were conducted by Y.H. and F.D.V.-B. (human data), D.S.D. (ATAC–seq), N.P. (CUT&RUN), N.T.Y. and Y.Z. (scRNA-seq), S.B.M., P.D.D. and K.P. J.Q. stained the in-house TMAs. C.V.B. performed mouse work, prepared tissue histology and lentivirus. P.C., N.J.D. and C.V.B. performed the orthotopic transplantations. J.F. and M.W. performed mouse work. R.G., M.G.D. and F.V.N.D. provided the patient-derived samples. R.R.M., O.J.S., J.E., M.G.D., F.V.N.D., S.T., C.W.S. and K.B.M. supervised the project. The manuscript was prepared by P.C. and K.B.M., and all authors read and approved it.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Eduard Batlle and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Loss of ATRX expression is associated with metastasis.
(a) International Cancer Genome Consortium data of top 20 mutated cancer genes with high functional impact mutations in colorectal cancer (CRC). (b) Representative ATRX staining of human normal and CRC tissue microarray. Examples of positive and negative staining are shown. Scale bars, 500 µm. (c) Quantification of ATRX expression using immunohistochemistry H-score method analysed using QuPath. Samples are separated into normal, non-metastatic (stage I and II) and metastatic (stage III and IV) groups, n = 8 vs 47 vs 25 tumours. (d) Summary data indicating presence (H-score > 10) or absence (H-score <10) of ATRX staining in non-metastatic and metastatic samples. Number of tumours in each group indicated on graph, n = 47 vs 25 tumours. (e) Summary data indicating presence or absence of ATRX mutation in CRIS-B vs all other CRIS transcriptional subtypes. Data extracted from TCGA dataset where CRIS tumour annotation is known. Number of tumours in each group indicated on graph, n = 43 vs 278 tumours. (f) Overall survival data of patients with CRIS-B tumours separated on presence or absence of ATRX mutation. Data extracted from TCGA dataset, n = 37 vs 6 patients. For (c) data are mean ± SD. P values were calculated using ordinary one-way ANOVA with multiple comparisons. For (d) and (e) p values were calculated using two-sided Fisher’s exact test. For (f) P value was calculated using Log-rank (Mantel-Cox) test. (g) Lollipop plot of TCGA PanCancer mutational data for ATRX. ATRX mutations were analysed using cBioPortal (07/12/23) with TCGA PanCancer Atlas Studies selected. (h) Western-blot analysis of AKP ATRXKO organoids for ATRX and β-actin. n = 2 technical replicates. (i) Representative images of haematoxylin and eosin (H&E) stained lung metastases in mice injected with AKP Control or AKP AtrxKO2 organoid cells via tail vein. Scale bars, 500 µm. (j) Quantification of number of lung metastases per mouse, n = 7 vs 8 mice. (k) Quantification of total lung tumour burden per mouse, n = 7 vs 8 mice. (l) Summary data indicating presence or absence of lung metastases. Number of mice with lung metastases or no metastases indicated on graph, n = 7 vs 8 mice. For (j) and (k) data are mean ± SD. P values were calculated using two-tailed Mann-Whitney test. For (l) p value was calculated using two-sided Fisher’s exact test.
Extended Data Fig. 2 Atrx deletion leads to tumour invasion.
(a) Subcutaneous tumour volume growth of mice injected with AKP Control or AKP AtrxKO organoid cells. (b) Volumes of tumours generated via subcutaneous injection of AKP Control or AKP AtrxKO, n = 5 vs 5 tumours. (c) Representative images of H&E-stained subcutaneous tumours in mice subcutaneously injected with AKP Control or AKP AtrxKO organoid cells. Scale bars, 100 μm overview and 50 μm zoom. (d) Representative images of β-catenin-stained subcutaneous tumours in mice subcutaneously injected with AKP Control or AKP AtrxKO organoid cells. β-catenin staining is used to identify tumour cells. Black arrows indicate β-catenin positive tumour cells invading into surrounding stroma. Scale bars, 100 μm overview and 50 μm zoom. (e) Quantification of invasive area of AKP Control vs AKP AtrxKO tumours, n = 5 vs 5 tumours. (f) Representative images of CDH1 stained subcutaneous tumours in mice subcutaneously injected with AKP Control or AKP AtrxKO organoid cells. Tumour core and invasive region of AKP AtrxKO tumour shown. Scale bars, 100 μm overview and 50 μm zoom. (g) Quantification of CDH1 staining intensity (H-score analysed with QuPath) in AKP Control vs AKP AtrxKO tumours, n = 5 vs 5 tumours. (h) Representative images of TWIST1 stained subcutaneous tumours in mice subcutaneously injected with AKP Control or AKP AtrxKO organoid cells. Scale bars, 100 μm overview and 50 μm zoom. (i) Quantification of TWIST1 staining in AKP Control vs AKP AtrxKO tumours, n = 5 vs 5 tumours. (j) RT-qPCR analysis of EMT marker expression in AKP Control vs AKP AtrxKO tumours, n = 5 vs 5 tumours. For (b), (e) and (i) data are mean ± SD. P values were calculated using two-tailed Mann-Whitney test. For (g) data are mean ± SD. P values were calculated using ordinary one-way ANOVA with multiple comparisons. For (j) gene expression was normalised to Actb and levels relative to AKP Control tumours calculated using the ΔΔCt method. Data are mean ± SD. P values were calculated using two-tailed student’s ttest. (k) Fluorescence microscopy of phalloidin stained AKP Control and AKP AtrxKO2 organoids following treatment with 5 ng/ml TGF-beta. Scale bars, 200 μm. (l) Quantification of the percentage of AKP Control vs AKP AtrxKO2 organoids adopting spindle-like morphology following TGF-beta treatment, n = 3 vs 3 independent experiments. Data are mean ± SD. P value was calculated using two-tailed student’s ttest.
Extended Data Fig. 3 Deletion of Atrx promotes metastasis.
(a) Fluorescence microscopy of phalloidin stained AKP Control and AKP AtrxKO organoids following concentration gradient of TGF-beta (TGFβ). Scale bars, 400 μm. (b) Quantification of the percentage of AKP Control vs AKP AtrxKO organoids adopting spindle-like morphology following TGF-beta gradient treatment, n = 3 vs 3 independent experiments. Data are mean ± SD. P values were calculated using ordinary two-way ANOVA with Sidak’s multiple comparisons. (c) Representative images of AKP Control and AKP AtrxKO organoids treated with TNFα (0.5 ng/mL), IFNγ (0.25 ng/mL) or control for 8 days, n = 3 vs 3 independent experiments. (d) Representative images of AKP Control and AKP AtrxKO treated with JQ1 (250 nm), FK228 (10 nM) or control for 8 days. n = 3 vs 3 independent experiments. (e) Relative cell viability following JQ1 or FK228 treatment in AKP Control vs AKP AtrxKO organoids, n = 3 vs 3 independent experiments. Data are mean ± SD. P values (DMSO vs JQ1 AKP control (p = 0.00000043), DMSO vs JQ1 AKP ATRX (p = 0.00000037). DMSO vs FK228 AKP control (p = 0.00000003), DMSO vs FK228 AKP ATRX (p = 0.00000003) were calculated using ordinary two-way ANOVA with multiple comparisons. (f) Cartoon illustrating organoids orthotopic colonic injection. Created with BioRender. (g) Representative colonoscopy images of mice orthotopically transplanted with AKP Control or AKP AtrxKO organoids, 7-weeks post transplantation, n = 9 vs 10 mice. (h) Survival plot for mice orthotopically transplanted with AKP Control or AKP AtrxKO aged until clinical end-points. (i) Total tumour area of primary tumours per mouse of the shown genotypes, n = 8 vs 9 mice. (j) Summary data indicating absence, presence, and site of metastases per genotype. Number of mice with metastases or no metastases indicated on graph, n = 9 vs 10 mice. (k) Representative images of H&E-stained primary tumours and liver metastasis in mice orthotopically transplanted with AKP Control and AKP AtrxKO organoid cells. n = 9 vs 10 biologically independent samples. Scale bars, 1 mm for primary tumours and 500 µm for liver metastasis. (l) Representative β-catenin-stained images of primary tumours in mice orthotopically transplanted with AKP Control or AKP AtrxKO organoids. β-catenin is used to identify tumour cells. Magnification highlights the distinct morphological features. Scale bars, 100 µm overview, 50 µm zoom. (m) Quantification of glandular morphology areas of AKP Control vs AKP AtrxKO primary tumours, n = 9 vs 10 mice. For (h) p value was calculated using Long-rank (Mantel-Cox) test. For (i) and (m) data are mean ± SD. P values were calculated using two-tailed Mann-Whitney test. For (j) p value is calculated using Fisher’s exact test.
Extended Data Fig. 4 Atrx deletion in BPN model results in tumour invasion.
(a) Targeted DNA sequencing of targeted Atrx locus in BPN AtrxKO organoid lines. (b) Representative pictures of BPN Control and BPN AtrxKO organoid culture. Scale bars, 1000 µm. (c) RT-qPCR analysis of the EMT marker Twist1 in BPN Control vs BPN AtrxKO organoid culture, n = 3 vs 3 independent experiments. (d) Fluorescence microscopy of phalloidin stained BPN Control and BPN AtrxKO organoids following treatment with 5 ng/ml TGF-beta. Zoomed area outlined in white box. Scale bars, 400 μm overview and 200 μm zoom. (e) Average number of clusters of BPN Control and BPN AtrxKO organoids per well exhibiting a spindle-like morphology following TGF-beta treatment, n = 5 vs 4 independent experiments. (f) RT-qPCR analysis of EMT and epithelial marker expression in BPN AtrxKO organoids either untreated or treated with 5 ng/ml TGF-beta, n = 3 vs 3 independent experiments. (g) Representative H&E and β-catenin-stained images of primary tumours in mice orthotopically transplanted with BPN Control or BPN AtrxKO organoid cells. β-catenin is used to identify tumour cells. Magnification highlights the distinct morphological features. Scale bars, 1 mm overview and 100 µm zoom, n = 6 vs 8 mice. (h) Quantification of tumour areas with glandular morphology in BPN Control and BPN AtrxKO primary tumours, n = 6 vs 8 mice. (i) Representative H&E-stained image of BPN AtrxKO metastasis. Scale bars, 1 mm. (j) Summary data indicating presence or absence of metastases. Number of mice with metastases or no metastases indicated on graph, n = 6 vs 8 mice. For (c) and (f) gene expression was normalised to Actb and relative levels were calculated using the ΔΔCt method. Data are mean ± SD. For (c), (e) and (f) p values were calculated using two-tailed student’s ttest. For (e) p = 0.000023. For (h) p value was calculated using two-tailed Mann-Whitney test. (k) Normalised enrichment scores of significantly enriched gene sets in TGF-beta treated AKP Control vs AKP AtrxKO organoids. (l) RT-qPCR analysis of representative EMT markers induced in AKP AtrxKO organoids following treatment with TGF-beta, n = 3 vs 3 independent experiments. (m) Normalised enrichment scores of significantly enriched gene sets in untreated AKP Control vs AKP AtrxKO organoids. (n) RT-qPCR analysis of representative genes important for colonic lineage specification and function, n = 3 vs 3 independent experiments. (o) RT-qPCR analysis of representative genes important for colonic lineage specification and function in BPN Control vs BPN AtrxKO organoids, n = 3 vs 3 independent experiments. For (l) gene expression was normalised to 18S rRNA and levels relative to untreated AKP control organoids calculated using the ΔΔCt method. For Twist1, Irx2 and Tfap2c gene expression was normalised to Actb and levels relative to untreated AKP control organoids calculated using the ΔΔCt method. For (n) gene expression was normalised to Actb and levels relative to untreated AKP Control organoids calculated using the ΔΔCt method. For (o) gene expression was normalised to Actb and levels relative to BPN Control organoids calculated using the ΔΔCt method. For (l) and (n) data are mean ± SD. P values were calculated using ordinary one-way ANOVA with multiple comparisons. For (l) Fgf2 p = 0.00000006, Serpine2 p = 0.000006, Col1a1 p = 0.000005, Col6a1 p = 0.000045, Col6a2 p = 0.000004, Pcolce p = 0.000015, Twist1 control p = 0.00000026, Twist1 + TGFβ1 p = 0.00000004. For (n) Hnf4a control p = 0.00000001, Cdx1 control p = 0.000000000000051, Cftr control p = 0.00000008. For (o) data are mean ± SD. P values were calculated using two-tailed student’s ttest. (p) Representative images of LY6D, KRT5 and EPCAM stained oesophagus and colon from Human Protein Atlas (Human Protein Atlas proteinatlas.org).
Extended Data Fig. 5 scRNAseq of AKP vs AKP AtrxKO organoids.
(a) Heatmap of single-cell RNAseq analysis showing the top 5 differentially expressed genes per cluster in AKP Control and AKP AtrxKO organoids. (b) UMAP plot of AKP Control and AKP AtrxKO cells coloured by expression levels of the osteoblast signature. The zoomed area of cluster 13 and 15 are outlined in black boxes. (c) UMAP plots of AKP Control and AKP AtrxKO cells coloured by expression levels of the Lgr5 and tumour intestinal stem cells (ISC) signatures. (d) UMAP plots of AKP Control and AKP AtrxKO cells coloured by expression levels of the Fetal, Epithelial-Specific High-Risk (EpiHR) or Injury Repair signatures.
Extended Data Fig. 6 Atrx deletion results in hybrid-EMT phenotype.
(a) Representative images of CDH1/TWIST1 co-immunofluorescence in AKP Control vs AKP AtrxKO organoids. n = 1 biological sample examined over three independent experiments. Scale bars, 100 µm. (b) FACS plots for analysing and sorting EPCAM+ve/ITGA5+ve cells from AKP Control or AKP AtrxKO organoids. (c) Quantification of percentage of cells in each EPCAM/ITGA5 population, n = 5 vs 5 independent experiments. (d) Quantification of percentage of ITGA5+ve cells in AKP Control vs AKP AtrxKO organoids, n = 5 vs 5 independent experiments. (e) Quantification of percentage of EPCAM+ve cells in AKP Control vs AKP AtrxKO organoids, n = 5 vs 5 independent experiments. (f) Fluorescence microscopy of phalloidin stained ITGA5+ve and ITGA5-ve cells sorted from AKP Control or AKP AtrxKO organoids following treatment with 5 ng/ml TGF-beta, n = 3 vs 3 independent experiments. Spindle-like structures indicated with white arrows. Scale bars, 1000 μm. Zoomed area outlined in white box. (g) Quantification of presence of spindle-like organoid structures formed following treatment of ITGA5+ve and ITGA5-ve cells sorted from AKP Control or AKP AtrxKO organoids with 5 ng/ml TGF-beta, n = 3 vs 3 independent experiments. (h) RT-qPCR analysis of Itga5 expression in ITGA5+ve vs ITGA5-ve cells sorted from AKP AtrxKO organoids, n = 3 vs 3 independent experiments. (i) RT-qPCR analysis of various mesenchymal marker gene expression in ITGA5+ve vs ITGA5-ve cells sorted from AKP AtrxKO organoids, n = 3 vs 3 independent experiments. (j) RT-qPCR analysis of various colonic epithelial marker gene expression in ITGA5+ve vs ITGA5-ve cells sorted from AKP AtrxKO organoids, n = 3 vs 3 independent experiments. For (d) and (e) data are mean ± SD. P values were calculated using two-tailed student’s ttest. For (d) p = 0.000032. For (g) data are mean ± SD. P values were calculated using two-way ANOVA with Sidak’s multiple comparisons test. For (h) gene expression was normalised to Actb and levels relative to ITGA5-ve cells calculated using the ΔΔCt method. Data are mean ± SD. P value was calculated using two-tailed student’s ttest. For (i) and (j) gene expression was normalised to Actb and levels relative to ITGA5-ve cells calculated using the ΔΔCt method. Data are mean ± SD. P values were calculated using two-way ANOVA with Sidak’s multiple comparisons test. For (i) Twist1 p = 0.00000006, Col16a1 p = 0.00000003, Snai1 p = 0.00000012.
Extended Data Fig. 7 Emergence of squamous-like cells following Atrx deletion.
(a) RT-qPCR analysis of Emp1 and Lgr5 expression in LY6D+ve vs LY6D-ve cells sorted from AKP AtrxKO organoids, n = 3 vs 3 independent experiments. (b) RT-qPCR analysis of squamous markers Ly6d and Krt5 expression in tumours from mice injected with AKP Control or AKP AtrxKO organoids, n = 5 vs 5 tumours. (c) Representative images of KRT5 staining of AKP AtrxKO subcutaneous tumours (left) and lung metastases (right). Examples of the different morphologies of KRT5+ cells observed in these tumours are shown. In subcutaneous example 1, stratified epithelium (indicated by a black *) and elongated cells (indicated by black arrow) are shown. In subcutaneous tumour example 2 a keratin pearl is shown. In lung metastasis example 1 elongated cells are shown (indicated by black arrows). In lung metastasis example 2 stratified regions are shown. (d) Quantification of percentage of ITGA5+ve single positive, LY6D+ve single positive, and ITGA5+ve/LY6D+ve double positive cells in AKP Control vs AKP AtrxKO organoids, n = 3 vs 3 independent experiments. For (a) gene expression was normalised to Actb and levels relative to LY6D-ve cells calculated using the ΔΔCt method. Data are mean ± SD. P values were calculated using two-tailed student’s ttest. For (b) gene expression was normalised to Actb and levels relative to AKP Control tumours calculated using the ΔΔCt method. Data are mean ± SD. P values were calculated using two-tailed student’s ttest. For Ly6d p = 0.00005. For (d) data are mean ± SD. P values were calculated using two-tailed student’s ttest. For ITGA5+ cells (left panel) p = 0.000044. (e) Quantification of percentage of ITGA5-ve/LY6D-ve double negative, LY6D+ve single positive, ITGA5+ve single positive, and ITGA5+ve / LY6D+ve double positive cells in cultures of AKP AtrxKO organoids 9 days after plating of different sorted populations. The plated population is indicated on the x-axis and the population being analysed indicated in the graph title and y-axis, n = 4 vs 4 independent experiments. (f) Quantification of percentage of KRT5+ cells in tumours derived from mice injected with ITGA5-ve/LY6D-ve double negative, LY6D+ve single positive, ITGA5+ve single positive, or ITGA5+ve / LY6D+ve double positive AKP AtrxKO cells, n = 5 vs 5 mice. For (e) and (f) data are mean ± SD. P values were calculated using one-way ANOVA with multiple comparisons test.
Extended Data Fig. 8 Tgfbr2 deletion suppresses plasticity in AtrxKO organoids.
(a) Representative images of AKP Control and AKP AtrxKO cells treated or untreated with 5 ng/ml TGF-beta (TGFβ) for 72 h, n = 3 vs 3 independent experiments. Scale bars, 200 μm. (b) Relative cell viability of AKP Control vs AKP AtrxKO single cells treated or untreated with TGF-beta, n = 3 vs 3 independent experiments. (c) Targeted DNA sequencing of targeted Tgfbr2 locus in AKP AtrxKO organoid lines. (d) RT-qPCR analysis of representative genes important for squamous, EMT and colonic lineage specification and function in AKP Control, AKP AtrxKO Control and AKP AtrxKO Tgfbr2KO organoids. (e) FACS plots for analysing LY6D+ve/ITGA5+ve cells from AKP AtrxKO Control and AKP AtrxKO Tgfbr2KO organoids, n = 3 vs 3 independent experiments. (f) Quantification of percentage of cells in each LY6D / ITGA5 population, n = 3 vs 3 independent experiments. (g) Fluorescence microscopy of phalloidin stained AKP AtrxKO Control and AKP AtrxKO Tgfbr2KO organoids following treatment with 5 ng/ml TGF-beta, n = 3 vs 3 independent experiments. Scale bars, 200 μm. (h) Quantification of presence of spindle -like organoid structures formed following treatment of AKP AtrxKO Control and AKP AtrxKO Tgfbr2KO organoids with 5 ng/ml TGF-beta, n = 3 vs 3 independent experiments. For (b) data are presented as mean values ± SD. P values were calculated using one-sided ANOVA multiple comparisons test. For (d) n = 1 biologically independent sample examined over 3 independent experiments. Gene expression was normalised to Actb and levels relative to AKP Control calculated using the ΔΔCt method. Data are mean ± SD. P values were calculated using one-sided ANOVA multiple comparisons test. For (h) p value was calculated using two-tailed student’s ttest.
Extended Data Fig. 9 HNF4A mediates Atrx loss phenotypes.
a) PCA plot of ATAC-seq data comparing AKP Control vs AKP AtrxKO organoids, n = 3 vs 3 independent experiments. (b) Representative IGV browser tracks of ATAC peaks at colonic epithelial genes. Significantly altered peaks are outlined with a red box. Blue track refers to AKP control, while red track refers to AKP AtrxKO organoids. (c) Representative IGV browser tracks of ATAC peaks at mesenchymal genes. Significantly altered peaks are outlined with a red box. Blue track refers to AKP control, while red track refers to AKP AtrxKO organoids. (d) Representative IGV browser tracks of ATAC peaks at squamous genes. Significantly altered peaks are outlined with a red box. Note lack of significantly different peaks at Krt5 and Ly6d despite altered expression. Blue track refers to AKP control, while red track refers to AKP AtrxKO organoids. (e) Heatmap showing transcription factor binding sites enriched in regions that lose accessibility in AKP AtrxKO organoids. Brown shade gradient indicates regions where accessibility is lost in AKP AtrxKO organoids. (f) PCA plot of H3K27ac CUT&RUN comparing AKP Control vs AKP AtrxKO organoids, n = 3 vs 3 independent experiments. (g) RT-qPCR analysis of colonic epithelial cell marker expression in AKP Control, AKP AtrxKO and AKP AtrxKO Hnf4a overexpressing organoids, n = 3 vs 3 independent experiments. (h) RT-qPCR analysis of Hnf4α in AKP Control vs AKP Hnf4aKO organoids, n = 3 vs 3 independent experiments (i) Western-blot analysis of AKP Control and AKP Hnf4aKO organoids for HNF4A and β-actin. n = 4 vs 4 technical replicates. (j) Volcano plot depicting differentially expressed genes in AKP Control vs AKP Hnf4a KO organoids following RNAseq analysis. Red dots represent genes expressed at higher levels in the AKP Hnf4a KO organoids, while blue dots represent genes expressed at higher levels in the AKP Control, n = 3 vs 3 technical replicates. (k) Representative H&E images of subcutaneous tumours in mice subcutaneously injected with AKP Control or AKP Hnf4aKO organoid cells, n = 5 vs 5 mice. Magnification highlights the distinct morphological features. Scale bars, 1 mm overview and 250 µm zoom. (l) Quantification of non-glandular morphology area of AKP Control vs AKP Hnf4aKO subcutaneous tumours, n = 5 vs 5 mice. For (e) p values were calculated using one-sided Fisher’s exact test. For (g) and (h) gene expression was normalised to Actb and levels relative to AKP Control calculated using the ΔΔCt method. Data are mean ± SD. For (g) p values were calculated using one-way ANOVA with multiple comparisons test. For (h) p value was calculated using two-tailed student’s ttest. For (l) data are mean ± SD. P values were calculated using two-sided Mann-Whitney test.
Extended Data Fig. 10 Squamous-like plasticity observed in stage IV CRC samples.
(a) Scatter plot of individual tumour H-score values for HNF4A and ATRX of stained human CRC tissue microarray. (b) Scatter plot of individual tumour H-score values for CDX2 and ATRX of stained human CRC tissue microarray. (c) Scatter plot of individual tumour H-score values for CDX2 and HNF4A of stained human CRC tissue microarray. For (a), (b) and (c) samples staining positive for LY6D ( > 2% cells LY6D+ve) highlighted in red. Data is individual value X/Y scatter plot. P values were calculated using two-sided Pearson’s correlation test. For (a) p < 0.000000000000001, (b) p = 0.0000000009, (c) p < 0.000000000000001. (d) Representative IHC staining of a human primary stage IV CRC tissue microarray for KRT5. (e) Summary data indicating the presence of KRT5 positive cells (>2% cells KRT5+ve) in human stage I-III vs stage IV primary CRC. (f) Representative IHC staining of a human matched liver metastasis tissue microarray for KRT5 expression. (g) Summary data indicating the percentage of KRT5 positive cells in matched human primary tumours and liver metastases. (h) Representative image of HNF4A IHC staining in matched human stage IV primary tumour and matched liver metastasis. Scale bars, 100 µm. (i) Summary data indicating the percentage of HNF4A positive cells in matched human primary tumours and liver metastases. (j) Representative image of ATRX IHC staining in matched human stage IV primary tumour and liver metastasis. Scale bars, 100 µm. (k) Summary data indicating the percentage of ATRX positive cells in matched human primary tumours and liver metastasis. For (e) p value was calculated using two-sided Fisher’s exact test. For (g), (i) and (k) p values were calculated using two-tailed Wilcoxon matched-pairs signed rank test. n = 17 vs 17 matched human primary stage IV tumours and liver metastases. (l) Targeted DNA sequencing of targeted ATRX locus in human CRC organoid line. (m) RT-qPCR analysis of colonic epithelial and squamous marker gene expression in human ATRXKO CRC organoids and control (NT), n = 3 vs 3 independent experiments. (n) Fluorescence microscopy of phalloidin stained human control (NT) and ATRXKO CRC organoids following treatment with 5 ng/ml TGF-beta, n = 3 vs 3 independent experiments. Zoomed area outlined in white box. Scale bars, 400 μm overview, 200 μm zoom. (o) Quantification of the percentage of human control (NT) and ATRXKO CRC organoids adopting spindle-like morphology following TGF-beta treatment. For (m) gene expression was normalised to ACTB. For (m) and (o) n = 1 biologically independent sample examined over 3 independent experiments. Data are mean ± SD. P values were calculated using two-tailed student’s ttest.
Extended Data Fig. 11 ATRX mutation correlates with iCMS3 transcriptional subtype.
(a) Summary data indicating presence or absence of ATRX mutation in iCMS2 vs iCMS3 transcriptional subtypes. Data extracted from TCGA dataset. Number of tumours in each group indicated on graph, n = 348 vs 244 tumours. (b) Heatmap clustering of Marisa CRC patient dataset of 557 tumours based on squamous-like (AtrxKO) and colonic epithelial (AtrxWT) gene expression signatures. (c) Percentage of tumours located in the distal (left sided) or proximal (right sided) colon in different Atrx expression clusters. One-tailed Chi-squared test, p value = 1.72e-16. (d) MMR status of tumours in HiCol, Intermediate and HiSquam expression clusters. (e) Mutational status of tumours in HiCol, Intermediate and HiSquam expression clusters. (f) TNM stage of tumours in HiCol, Intermediate and HiSquam expression clusters. (g) Correlation of AtrxWT (colonic epithelial-like) and AtrxKO (squamous-like) gene expression signatures in GSE39582 dataset. iCMS status overlayed in purple (iCMS2) and orange (iCMS3). (h) Percentage of tumours designated as iCMS2 (i2) or iCMS3 (i3) transcriptional subtype in different Atrx expression clusters. Chi-squared test, p value = 2.2e-16. (i) FACS plots for analysing EPCAM+ve/LY6D+ve cells from normal colon and colon cancer samples. Box highlights the presence of EPCAM+ve/LY6D+ve cells. (j) Quantification of EPCAM+ve/LY6D+ve cells from normal colon and colon cancer samples, separated into left and right sided groups. For (a) p value was calculated using two-sided Fisher’s exact test. For (c), (d), (e), (f) and (h) p values were calculated using one-tailed Chi-Square test. For (g) Pearson’s correlation coefficient and two-sided p-value are described. The blue line and gray shaded area represent the regression line and 95% confidence interval. For (j) n = 5 (normal colon), n = 2 (left sided tumour), n = 3 (right sided tumour) biological independent samples. Data are mean ± SD. P values were calculated using ordinary one-way ANOVA with multiple comparisons.
Extended Data Fig. 12 AtrxKO expression signature is associated with loss of HNF4A activity.
Heatmap of z-scores of ATAC-seq log2 normalised counts across 77 samples from 37 CRC patients from the TCGA. ATAC-seq peaks shown correspond to the significantly differential accessible regions in patients with high AtrxKO expression score vs high AtrxWT patients. FDR was calculated using two tailed ttest.
Supplementary information
Supplementary Information (download PDF )
Supplementary Information 1–3.
Supplementary Tables (download ZIP )
Supplementary Tables 1–11.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cammareri, P., Raponi, M., Hong, Y. et al. Loss of colonic fidelity enables multilineage plasticity and metastasis. Nature 644, 547–556 (2025). https://doi.org/10.1038/s41586-025-09125-5
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41586-025-09125-5
This article is cited by
-
KMT2D loss drives adeno-to-squamous transition and sensitizes TKI-resistant lung cancer to AURKA inhibition
Cell Death & Differentiation (2026)
-
A robust classifier for the intrinsic consensus molecular subtypes in colorectal cancer
Journal of Translational Medicine (2025)
