
Abstract
Fibrolamellar hepatocellular carcinoma (FLC) is a rare form of liver cancer affecting children and young adults that is driven by a chimeric protein, DNAJ-PKAc. The development of molecular inhibitors of DNAJ-PKAc has been hampered by unacceptable on-target toxicity, but the chimera results in a tumor-specific antigen (neoantigen) that may be targeted immunologically. Here we conducted a phase 1 clinical trial of a therapeutic peptide vaccine targeting DNAJ-PKAc (FLC-Vac), in combination with nivolumab and ipilimumab, in children and adults with advanced FLC, who had not previously received immune checkpoint therapy. The primary objectives were safety and T cell responses after week 10 (priming phase). Of the 16 patients enrolled, 12 completed the vaccine priming phase and were evaluable for both immunological and clinical endpoints. The median age was 24 years (range 12–47 years). Grade 3 treatment-related adverse events were reported by six patients (37.5%). DNAJ-PKAc-specific T cell responses were detected in 9 of 12 patients after treatment. In the subset of patients who completed the initial priming phase the disease control rate was 75% (9/12), with three partial responses (25%). All patients with clinical responses also had DNAJ-PKAc-specific T cell responses, from which we identified multiple class-II-restricted T cell receptors with specificity for DNAJ-PKAc. Correlates of response included both functional neoantigen reactivity and changes in T cell receptor repertoire features over time. Immune escape in two patients corresponded with immune exhaustion rather than neoantigen escape or human leukocyte antigen loss. Our findings demonstrate the potential for therapeutic vaccines targeting ‘undruggable’ oncogenic drivers and suggest a rubric for evaluating effective anti-neoantigen immunity. ClinicalTrials.gov identifier: NCT04248569.
Similar content being viewed by others

Main
Fibrolamellar hepatocellular carcinoma (FLC) is a type of liver cancer that typically affects children and young adults. There is no standard systemic therapy for FLC1,2, and the median survival for patients with unresectable FLC is approximately 12 months3. The FLC transcriptome is characterized by an in-frame fusion (DNAJB1::PRKACA), yielding a chimeric protein, DNAJ-PKAc4. Molecularly targeted therapies have revolutionized the management of some fusion-driven cancers (for example, ALK, ROS1 and NTRK)5; but efforts to develop specific inhibitors for FLC have been limited by unacceptable on-target toxicity1. Alternative strategies to target DNAJ-PKAc in FLC are urgently needed.
Neoantigens derived from oncogenic fusions can induce antitumor immunity6. Recent data indicate that DNAJ-PKAc is immunogenic7,8. A patient with FLC who participated in a personalized vaccine program developed immunity against DNAJ-PKAc after vaccination that was associated with prolonged disease-free survival after undergoing resection of FLC tumors. The DNAJB1::PRKACA fusion breakpoint occurs within an intron, making the sequence of the chimera consistent across patients. This allows a single vaccine to be broadly applicable for patients with FLC. Here, we prospectively evaluated the safety and immunogenicity of a vaccine targeting DNAJ-PKAc (containing a synthetic long peptide spanning the chimera, with a poly-ICLC adjuvant), combined with immune checkpoint inhibitors (ICI) therapy, in children and adults with FLC.
Results
From April 2020 to September 2022, 16 patients with unresectable FLC were enrolled (Fig. 1a). The clinical characteristics of the enrolled patients are summarized in Table 1. The median age was 23.5 years (range 12–47 years). The majority of patients (63%, n = 10) had received at least one line of prior systemic therapy (range of one to four lines).

a, A consort diagram of enrolled patients. n values represent the number of patients at each stage of the protocol. The plus–minus sign signifies the safety population and included all patients who received at least one dose of the study drugs. The pilcrow (¶) signifies the evaluable population and included all patients who received at least one dose of vaccine and had baseline and post-treatment T cell measures at 10 weeks. Four patients were not evaluable as patients came off the study before obtaining post-treatment T cell measures at 10 weeks. b, The study schema. c, The change from baseline in the target lesion diameter according to RECIST, version 1.1. d, A spider plot of radiographic responses to treatment. Tumor responses were measured at regular intervals, and the values shown are the largest percentage change in the sum of the longest diameters from the baseline measurements of each measurable tumor. Each line represents one patient. e,f, Kaplan–Meier curves of overall survival (OS) (e) and PFS (f). The 95% CIs for point estimates are shown in red shading. NA, not achieved. g, Radiologic scans of two representative responders (P14 and P2) during FLC-Vac with ipilimumab and plus nivolumab therapy. ITT, intention to treat; ORR, overall response rate; PD, progressive disease; PR, partial response; SD, stable disease; SLD, sum of longest diameters of the target lesions. The orange arrows point to the tumor sites.
Efficacy
The first three patients (age ≥18 years) received one cycle of vaccine monotherapy before starting combination therapy with nivolumab and ipilimumab (Fig. 1b). As no unacceptable toxicities occurred during the vaccine lead-in, the trial progressed to full enrollment. Of the 16 enrolled patients, four discontinued treatment before the 10-week post-treatment T cell assessments and were not evaluable for the primary immunological endpoints (Fig. 1a). In the intent-to-treat population (n = 16), the objective response rate (ORR) was 19% (3/16), and the disease control rate (DCR) was 62.5% (10/16) (Extended Data Fig. 1a–c). Among the four early discontinuations, three were due to clinical progression, one due to elevated liver function tests (this patient subsequently achieved a response in combination with gemcitabine and oxaliplatin chemotherapy).
Because vaccine-induced immunity typically takes several weeks to manifest, we focused our analysis on the 12 patients who completed the priming phase. In this subset, the ORR was 25%, and the DCR was 75% (9/12) (Fig. 1c,d). The median progression-free survival (PFS) was 8.5 months (95% confidence interval (CI) 4.1– not reached (n.r.)), and the median overall survival was not reached (n.r.) (95% CI 21.5–n.r.) (Fig. 1e–g and Extended Data Fig. 1d,e). All three responding FLC cancers were mismatch repair proficient, had low tumor mutational burden (<10 mutations per megabase) and absent PDL1 expression (PDL1 <1%). Two responders discontinued therapy to undergo resection of residual disease, subsequently achieving no evidence of disease. The third, a 13-year-old, completed 2 years of therapy with an ongoing response.
Safety
Toxicity was evaluated in all patients who received at least one dose of study drug(s) (n = 16, Table 2). The most common vaccine-related adverse events were injection site reactions (16/16, 100%), headaches (8/16, 50%) and fatigue (7/16, 45%), all grade ≤2. Grade 3 treatment-related adverse effects were reported by six patients (6/16, 37.5%) and were broadly consistent with ipilimumab plus nivolumab in other tumor types. No grade 4 or 5 events were observed.
DNAJ-PKAc-specific T cell responses as a prerequisite for clinical activity
To investigate the mechanisms underlying the observed clinical activity, we used ex vivo IFNγ ELISpot assays to measure T cell responses against DNAJ-PKAc in the peripheral blood at multiple time points. Positive responses were defined as a ≥2.5-fold increase in IFN-γ-producing DNAJ-PKAc-specific T cells compared with an unstimulated control. Responses against the full-length vaccinated peptide were observed in 8.3% of patients (1/12) at baseline and 75% (9/12) at post-vaccination time points (Fig. 2a and Extended Data Fig. 2), including all patients with clinical objective responses. Of the nine post-treatment responses against the full-length peptide, five out of nine also recognized at least one nine-amino-acid minimal epitope (Fig. 2b and Extended Data Fig. 2). The intensity and timing of peak ELISpot responses varied by clinical outcome (Fig. 2c). Patients with partial responses showed delayed peaks compared with those with progressive or stable disease, possibly reflecting initial immune sequestration within the tumor. We did not observe any correlation between the number of DNAJ-PKAc fusion neoepitopes predicted for patients’ human leukocyte antigen (HLA) type and their ELISpot response or clinical outcome (Supplementary Tables 1–3), indicating that predicted neoepitope presentation (or lack thereof) is not sufficient to explain functional or clinical responses.

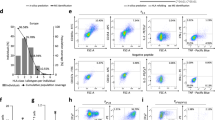
a, Pie charts illustrating the distribution of clinical responses among ELISpot nonresponders and ELISpot responders. b, Quantified ELISpot responses (mean ± s.d. of three replicate wells per condition) against the full 24-amino-acid peptide (peptide 9), eight overlapping 9-amino-acid peptides (peptides 1–8) spanning the chimera, negative controls (medium-only and unstimulated) and positive control (CEF peptide pool, consists of epitopes from cytomegalovirus, Epstein-Barr virus and influenza (flu)). An unpaired two-sided t-test comparing the mean ELISpot repose to peptide versus unstimulated control was performed. For P1 week 10, Pep9 ***P = 0.0001. For P13 week 32, Pep2 *P = 0.0161, Pep5 **P = 0.0031, Pep7 *P = 0.0474, Pep9 ***P = 0.0008. Example images of ELISpot assay results for patient P1 are shown with the unstimulated condition and the response after stimulation with peptide 9. Spot formation indicates antigen-specific T cell activity. c, A graph depicting the timing of peak ELISpot responses (left) and the intensity of ELISpot responses over time (right) for each patient, grouped by clinical response. d, Pie charts illustrating the CyTOF-derived T cell responses for each patient categorized by T cell type (TH, TC and combined TH + TC responses) (left) and the T cell response types among clinical responders (right). e, An example heat map illustrating CyTOF data from patient P8 clustered by T cell population for each simulation condition. The groups include unstimulated samples, samples stimulated by KRAS peptide (negative control) and the FLC 24mer peptide, all at baseline (B) and peak ELISpot response (P) time points. f, The number of statistically expanded TCR clones per patient after one cycle of treatment (C2D1 versus C1D1) (left) or two or more cycles of treatment (C3D1+ versus C1D1) (right), colored by whether patient received FLC-Vac monotherapy (turquoise) or vaccine combined with ICB (pink) during the first cycle of treatment. Two-sided Wilcoxon rank-sum test, **P = 0.009, not significant (n.s.) P = 1. ICB, immune checkpoint blockade. g, TCRdist networks of statistically expanded TCR clones after one cycle of treatment (C2D1, left) or two or more cycles of treatment (C3D1+, right). Each node represents a unique TCR clone, and two nodes are connected by an edge when their TCRdist ≤120; the node size corresponds to node degree (number of neighbors), and the nodes are colored by patient. The dashed lines mark clusters attributed to response to influenza epitope M158 (A*02:01-GILGFVFTL). h, A quantification of network connectivity by number of edges in the network for each patient, grouped by clinical response, after one cycle of treatment (C2D1) versus two or more cycles of treatment. i, Left: a decision tree predicting response to therapy based on integrated results from change in TCRdist clustering network connectivity over time and ELISpot response. Right: Receiver operating characteristic curves for classification of responders (PR only) versus nonresponders (SD + PD) based on ELISpot response (blue), change in TCRdist clustering network connectivity using the threshold determined by the decision tree (red) or both metrics together (black). The area under the curve (AUC) is given for each classifier, as well as the P value from a two-sided Fisher’s exact test on the confusion matrix for each classifier. PD, progressive disease; Pos, positive; PR, partial response; SD, stable disease; TH, T helper; TC, cytotoxic T cells.
To further characterize peptide-specific T cell subsets, we performed in vitro stimulations using the FLC peptide or an irrelevant peptide corresponding to a neoantigen that is not known to be expressed by FLC tumors (KRAS G12D). CyTOF analysis revealed notable post-treatment increases in effector (CD3+CD8+), effector memory (CD62L−CD44+) and central memory (CD62L+CD44+) T cell populations, potentially enabling durable responses in some patients (Fig. 2d,e, Extended Data Fig. 3, Supplementary Fig. 1 and Supplementary Tables 4 and 5a,b). Although therapeutic cancer vaccines can induce exhaustion in some contexts, we did not observe an increase in PD1+TIM3+ T cells from baseline to early on treatment or any appreciable change over time (Extended Data Fig. 3b). These data suggest that the vaccine expands polyfunctional, antigen-experienced and memory T cell responses, predominantly driven by CD4+ T helper cells.
We also profiled changes in patients’ peripheral blood T cell receptor (TCR) repertoires at multiple time points using TIRTL-seq9, a high-throughput paired TCR sequencing technology, to identify response-associated features (Extended Data Fig. 4). Dynamic repertoire changes were observed in all 12 patients (Extended Data Fig. 5). Patients receiving vaccine monotherapy for the first cycle (patient (P)6, P14 and P16) showed fewer expanded TCR clones at C2D1 compared with those receiving concurrent vaccine and ICIs, suggesting that the addition of ICIs to vaccine therapy may augment clonal expansions beyond those elicited by vaccine alone. After two or more cycles of treatment, both groups had similar numbers of expanded clones (Fig. 2f). To assess if expanding clones in peripheral blood also infiltrated tumors, paired pre- and post-treatment biopsies were analyzed. Although biopsies from distinct lesions showed little overlap, all six patients with paired samples exhibited increased tumor infiltration of peripherally expanding clones post-treatment (Supplementary Fig. 2), indicating that peripheral changes reflect intratumoral TCR repertoire shifts.
We then examined whether characteristics of expanding TCR clones differed between responders and nonresponders. It is well established that effective immune responses against immunodominant epitopes frequently involve groups of TCRs with shared sequence features10, so we first quantified TCR sequence similarity of longitudinally expanded clones within and across patient repertoires using TCRdist11, then constructed similarity networks visualizing clusters of TCRs with closely related amino acid sequences (Fig. 2g). While a previous study8 observed little TCR clustering in endogenous antitumor responses in FLC, we observed numerous clusters of related TCRs among those that expanded over the course of combination immunotherapy therapy in our patients. Some clusters could be attributed to TCRs recognizing nontumor epitopes, such as clusters corresponding to the influenza epitope A*02:01-GILGFVFTL (M158) identified in a patient who developed a viral respiratory infection during treatment (Fig. 2g, dashed outline), while others probably reflect antitumor responses. Clustering was more pronounced at later time points, especially in responders. Connectivity within TCR networks (edges per patient) remained stable in progressive disease (except P16) but increased over time in partial responders and some patients with stable disease (Fig. 2h).
Finally, we developed a hierarchical decision tree to stratify responders using ELISpot response and changes in TCRdist network connectivity measured as the number of edges in each network (Fig. 2i). Responders were characterized by: (1) increase in edges ≥110 and (2) positive ELISpot response. Whereas ELISpot response alone was a poor classifier (area under the receiver operating characteristic curve (AUROC) of 0.667, Fisher’s exact test P value = 0.509), change in TCRdist edges performed reasonably well as a single metric (AUROC of 0.889, Fisher’s exact test P value = 0.045). Classifier performance was further improved by combining both components (AUROC of 0.944, Fisher’s exact test P value = 0.018). These findings suggest productive responses to combination immunotherapy involve both increased TCR clustering and functional vaccine reactivity.
Characterization of DNAJ-PKAc-specific T cell responses
To understand mechanisms of response, we conducted an in-depth analysis of DNAJ-PKAc-specific T cell responses in responding P12 (Fig. 3a–c). At baseline, no DNAJ-PKAc-specific response was detected via IFNγ ELISpot, whereas after treatment, ELISpot showed reactivity to the vaccinated peptide (Extended Data Fig. 2). CyTOF analysis implicated activated CD4+ T helper and CD8+ cytotoxic T cells as the predominant responders (Extended Data Fig. 3a). We cloned and screened 33 TCRs from P12, two of which were fusion-specific (P12-TCR2 and P12-TCR3) (Fig. 3d). Single-cell RNAseq and paired TCR sequencing of P12 T cells expanded and restimulated with the vaccinated peptide identified activation-induced marker (AIM)-positive clones expressing genes such as CD40LG, IFNG and TNF. P12-TCR2 and P12-TCR3 localized to a cluster enriched in these activation markers, along with three additional TCRs (P12-TCR6, P12-TCR8 and P12-TCR13) (Fig. 3e,f), which were also reactive to the 24mer peptide presented by autologous lymphoblastoid cell lines (LCLs) (Fig. 3g,h).

a, Patient P12 achieved a partial response (−51% decrease), with reductions in all target lung lesions and abdominal nodes, but later developed oligoprogression of a cardiophrenic node (lesion 4), resulting in discontinuation of study therapy. The orange arrows point to the tumor sites. b, Change in target lesions from baseline by RECIST 1.1. c, Vitamin B12 binding capacity, a FLC serum tumor marker, declined on study therapy. ULN, upper limit of normal. d, A scatterplot showing relative peripheral blood frequencies of unique TCR clones detected by TIRTL-seq for P12 at baseline (C1D1, x axis) and after two cycles of treatment (C3D1, y axis). Statistically expanded clones at C3D1 relative to baseline are outlined in red, statistically contracted clones are outlined in blue, and stable clones are outlined in gray. The cloned and screened TCRs that were not DNAJ-PKAc-specific are colored black, while fusion-specific clones P12-TCR2 and P12-TCR3 are colored yellow and cyan, respectively. e, The same scatterplot as d, filtered to show only clones that appeared in the scRNA sequencing dataset and expressed at least one of IFNG, TNF and/or CD40LG (AIM+). The colors same as in d. f, A uniform manifold approximation and projection representation of P12 scRNA sequencing data highlighting TCR clonotypes of interest (upper left, cells colored by TCR clonotype ID) and expression levels of CD8A, CD4, IFNG, TNF and CD40LG (right and bottom, cells colored by normalized expression level of relevant gene). All TCRs of interest localize to activated CD4 cluster. g, Representative flow cytometry plots for fusion specificity screening of P12-TCR2 TCR-expressing Jurkat cells against autologous LCLs pulsed with 24mer DNAJ-PKAc fusion peptide, with or without HLA-blocking antibodies. DP, DNAJ-PKAc fusion peptide. h. Quantification of flow cytometry results for screening of all five TCRs of interest against autologous LCLs pulsed with 24mer DNAJ-PKAc fusion peptide, with or without HLA-blocking antibodies. The results shown for duplicate experiments, normalized to unstimulated negative and PMA-ionomycin positive controls (see Supplementary Methods for formula).
HLA restriction analysis showed that all five TCRs recognized DNAJ-PKAc via HLA-DR alleles. P12-TCR2, P12-TCR3 and P12-TCR8 recognized the peptide presented by donors with DRB3*01:01, while P12-TCR6 and P12-TCR13 were restricted to DRB1*13:01 (Extended Data Fig. 6a). These TCRs recognized distinct overlapping 15mers derived from the fusion, highlighting responses to multiple class-II-restricted neoepitopes (Extended Data Fig. 6b). To determine whether the combined criteria of longitudinal expansion and AIM reactivity would accurately identify fusion-specific clones, we screened an additional TCR selected in this manner, P12-TCR14, and confirmed that it was also fusion-specific and restricted to HLA-DRB3*01:01 (Extended Data Fig. 6c,d).
Notably, P12-TCR2 clustered with two longitudinally expanded TCRs from P8, who also expressed DRB3*01:01 (Extended Data Fig. 7a and Supplementary Table 2). Reconstruction and screening of these clustered TCRs, P8-TCR1 and P8-TCR2, confirmed that the latter was fusion-specific and restricted to DRB3*01:01 (Extended Data Fig. 7b). When we performed TCRdist clustering of all TCR clones from TIRTL-seq (rather than just expanded clones), we then identified a TCR from P12 (P12-TCR21) that differed from the fusion-specific P8-TCR2 by a single amino acid in the TCRβ chain, effectively a public DNAJ-PKAc-specific clone (Extended Data Fig. 7c). We also observed further clustering with other DRB3-restricted fusion-specific TCRs, including P12-TCR3, P12-TCR2/P8-TCR2 and P12-TCR8/P12-TCR14 clusters (Extended Data Figs. 7c and 8). The vast majority of TCRs in these clusters arose in patients who express a DRB3 allele (Supplementary Table 2 and Extended Data Fig. 7d). Further, each of these clusters exhibited strong similarities in V/J gene segment usage and CDR3 sequences (Extended Data Fig. 7e,f), suggesting similar neoepitope responses in patients with shared HLA types. Together, these findings demonstrate that FLC-Vac combined with ICIs elicited polyclonal T cell responses targeting diverse fusion neoepitopes with features that spanned multiple patients.
Acquired resistance driven by T cell exhaustion
Acquired resistance to immunotherapy often involves loss of antigen presentation, but DNAJ-PKAc persisted in all six patients with post-treatment tissue and one on-treatment sample (Fig. 4a). IHC confirmed retained HLA class I and II expression on tumor cells post- or on-treatment (Fig. 4b), supported by whole transcriptome sequencing showing no allele-specific HLA or TAP1 loss (Fig. 4c, Extended Data Fig. 9a and Supplementary Tables 6 and 7). No new oncogenic mutations or immune bypass mechanisms were detected (Extended Data Fig. 9b–g). Imaging mass cytometry and IHC of P12’s resistant lesion revealed a distinct tumor immune microenvironment with increased granulocytes, loss of B cell infiltration and an increase in CD16+ CD86− and CD206− macrophages, as compared with pretreatment and regressing lesions (Fig. 4d and Extended Data Fig. 10). T cells were confined to the tumor periphery and expressed higher levels of exhaustion markers (TIM3 and LAG3) (Fig. 4e and Supplementary Tables 8–10). These findings suggest impaired T cell trafficking and exclusion, along with enhanced T cell exhaustion and myeloid reprogramming, may have contributed to oligoprogression. Further supporting these resistance mechanisms, responder P14 experienced lung recurrence 2 years post study. Tumor profiling showed post-treatment expression of DNAJ-PKAc and intact HLA class II expression (Fig. 4a,b). Although the patient had progression on nivolumab, CyTOF analysis revealed a primarily CD4-driven response against DNAJ-PKAc, and a decision to rechallenge with nivolumab and ipilimumab (which robustly invigorates CD4 T cells)12,13 subsequently led to a complete response (Fig. 4f).

a, Tumor samples (n = 7) from the latest available time point with viable tissue were stained using RNAscope and probes for DapB (negative control) or DNAJ-PKAc fusion of interest and show retention of the target antigen post treatment. b, IHC demonstrating expression of HLA-I and HLA-II pre- and post-treatment in all samples. c, A heat map depicting log2 fold change in RNA expression of genes associated with antigen presentation machinery in patients with paired pre- and on-treatment tissue-based RNA sequencing (n = 6). No significant changes in antigen presentation machinery were observed. Color scale represents the log10 fold change in gene expression (post-treatment transcripts per million (TPM) / pre-treatment TPM). d, Imaging mass cytometry of samples from primary tumor, a responding lesion and a resistant lesion from P12, revealing an increase in the density of granulocytes and loss of B cells in the resistant tumor lesion. i, representative image 1; ii, representative image 2; Gran, granulocytes, Mac, macrophages. Scale bars, 200 µm. e, IHC showing an increase in immune exhaustion markers in the resistant lesion of P12 (mean ± s.d. for five replicate tissue areas). One-way analysis of variance, not significant (n.s.) P > 0.05, *P < 0.05, **P < 0.01, ***P < 0.001. For central CD3, primary versus response P = 0.0275, responding versus resistant P = 0.0037. For central TIM3, primary versus resistant P = 0.0076, responding versus resistant P = 0.0007. For central LAG3, primary versus resistant P = 0.0057, responding versus resistant P = 0.0108. f, Radiologic scans from P14 showing renewed response after rechallenge with ipilimumab (left) and CyTOF analysis from the same patient (right). The orange arrows point to the tumor sites.
At this report’s writing, 3 out of 3 responding patients remain without evidence of active cancer. Two responding patients discontinued study therapy to pursue resection of metastatic foci of disease and are without evidence of cancer. The third responding patient’s sole residual measurable lesion is no longer enhancing, consistent with tumor scar rather than active malignancy. These findings underscore the potential to overcome vaccine therapy resistance by targeting T cell exhaustion and restoring immune function.
Discussion
ICIs have limited efficacy in most pediatric tumor types, which are often characterized by a dominant genomic alteration and a low tumor mutational burden14. Therapeutic cancer vaccines are one promising strategy for stimulating antitumor immunity in tumor types for which such immunity does not endogenously occur15. Here, we show the potential for off-the-shelf therapeutic vaccines targeting shared neoantigens derived from currently ‘undruggable’ oncogenic drivers. Our clinical results are encouraging in the context of highly refractory cancer. In the largest prospective clinical trial of FLC so far evaluating multiple potential therapies, there were no responses, and all patients experienced progression by 6 months16. By contrast, in the present study, 6 of 12 patients who completed the 10-week priming period achieved disease control lasting ≥6 months.
Although it is not possible to definitively attribute the observed clinical responses to a complementary immune effect of the FLC vaccine and ICIs, ICIs have shown limited activity as monotherapy in FLC17,18,19. Similarly, therapeutic cancer vaccines have generally failed to elicit meaningful clinical responses as monotherapies in advanced cancers, probably due to immune exhaustion of vaccine-expanded T cells20. Single-cell RNA sequencing of tumor-infiltrating T cells revealed upregulation of exhaustion markers, underscoring the rationale for combining neoantigen vaccines with ICIs to generate robust and durable antitumor immunity21,22. In our study, we observed that ICIs were required to induce large T cell expansions detected in peripheral blood, which may be involved in antitumor activity. However, given that several patients with vaccine-induced T cell responses failed to achieve clinical benefit, additional therapies may be required to overcome the immunosuppressive tumor microenvironment. In particular, myeloid-targeted agents could be considered, based on our observation that changes in myeloid populations correlated with the development of acquired resistance in one patient.
An important limitation of the study is that four patients discontinued treatment before completing the 10-week priming phase and were therefore not assessable for immunologic response. These patients came off study early for a variety of different reasons including early disease progression or ICI-related toxicities, before vaccine-induced antitumor T cell immunity could be generated. This is not unexpected in the context of an aggressive disease such as FLC, particularly in patients who had received prior therapy. The early discontinuation of these patients impacted our ability to fully assess immune responses across the entire cohort. Notably, at least one of these patients went on to respond robustly to subsequent therapy, suggesting that while they did not complete the priming phase, their immune systems may still have been modulated in ways that could influence later treatment outcomes. Overall, the observation that half of the evaluable patients experienced prolonged disease control supports the feasibility of vaccine-primed immune engagement, also raising the possibility that immunologic priming may have delayed or downstream effects that warrant further exploration. A randomized study to determine the additive contribution and toxicity of each component of this therapeutic combination may be needed in the future.
A current challenge for immunotherapy is defining predictive biomarkers for responders and nonresponders to vaccines and checkpoint blockades. A report following ICI treatment of melanoma suggested that responders had more diverse TCR epitope-specific repertoires than nonresponders23. Here, we suggest a combined heuristic for stratifying response potential based on a TCR repertoire similarity measure (utilizing the entire expanding repertoire) combined with epitope-specific responses measured in functional assays. The repertoire similarity metric appeared to be particularly important in this heuristic, correctly stratifying 10 out of 12 patients even without the addition of functional response data. Increased TCR similarity among expanding TCR clones may correlate with enhanced antigen specificity, which could explain the importance of this feature in conjunction with functional reactivity to the vaccine antigen.
Finally, our study identified specific TCRs targeting DNAJ-PKAc, which opens a potential future avenue for adoptive T cell therapy. Notably, we identified large TCR clusters containing both validated fusion-specific TCRs and other closely related TCRs arising in patients who shared expression of HLA-DRB3 (9 out of 12 patients in our study). This response was striking for its public motifs shared across a majority of our cohort, a feature common to antiviral epitopes but rarely described thus far for tumor antigens. Many of these clustered TCRs were only detected at low levels in peripheral blood, potentially explaining why they were not sufficient to induce clinical responses in all DRB3-positive patients. Nevertheless, their presence highlights how cell therapy employing fusion-specific TCRs could provide meaningfult therapeutic benefit to a large portion of the patient population. Notably, the only epitope-specific T cell responses we confirmed targeting the fusion were from CD4+ T cells. The mechanisms by which CD4+ T cells might control epithelial-derived tumors are not fully resolved but could include direct cytolytic killing in cases where the tumor is class II positive, or through activating and inducing other antitumor responses including NK cells24,25.
Bespoke therapeutic vaccines tailored against the neoepitopes present in an individual cancer have recently demonstrated promise in multiple tumor types26,27,28,29. In this study, we show the potential for off-the-shelf therapeutic vaccines targeting shared neoantigens derived from a currently ‘undruggable’ oncogenic fusion event. Our findings contribute to the growing body of evidence supporting the feasibility of immunologically targeting shared (‘public’) neoantigens derived from oncogenic driver events—particularly in cancers such as FLC, which exhibit low tumor mutational burden and a correspondingly restricted repertoire of targetable neoantigens. Unlike many immunogenic passenger mutations, neoantigens arising from driver oncogenes are more likely to be persistently expressed through the continuum of disease progression, owing to the essential fitness advantage they confer to the cancer30. This sustained expression, combined with their immunogenic potential, renders driver-derived neoantigens potentially attractive targets for immunotherapy. Recent studies have demonstrated the clinical efficacy of both adoptive cell therapies and vaccines targeting driver-derived neoantigens, including a trial in renal cell carcinoma in which vaccinated patients exhibited prolonged relapse-free survival26,31,32,33,34. A key theoretical advantage of targeting oncogenic drivers immunologically is the potential to circumvent resistance mechanisms that often arise quickly with the use of targeted therapies, such as bypass signaling. Moreover, as compared with adoptive cell therapies targeting a neoepitope in the context of a single HLA molecule, immune escape should be less likely to occur with a vaccine-induced polyclonal CD4 response24. Our findings are consistent with these hypotheses, though based on a limited number of observations. In our study, we did not detect loss of DNAJ-PKAc or HLA in any patient post treatment. Among two patients who experienced eventual progression after an initial clinical response, we observed features suggestive of T cell exhaustion, and in one case, a potential restoration of efficacy with ICI rechallenge. Although preliminary, these results highlight a potential approach to address tumors with currently ‘undruggable’ oncogenic drivers.
Methods
Study design and treatment
We enrolled patients ≥12 years of age with unresectable FLC without prior exposure to an ICI. Patients received the FLC vaccine (consisting of a 24-amino-acid peptide and poly-ICLC adjuvant) on weeks 1, 2, 3, 4, 7 and 10 and nivolumab, 3 mg kg−1, followed by ipilimumab, 1 mg kg−1, every 3 weeks for four doses during the priming phase. Afterward, patients received nivolumab at a fixed dose of 480 mg every 4 weeks for up to 2 years and booster vaccinations approximately every three cycles for up to 1 year from the initial vaccination until disease progression, unacceptable toxicity or withdrawal. Ipilimumab plus nivolumab was selected as a backbone given its clinical availability for liver cancers (for example, hepatocellular carcinoma) and effects on CD4+ T cell populations, which have historically been the target of peptide vaccines12,13. Ipilimumab plus nivolumab was provided by Bristol Myers Squibb. The disease assessment, management of toxic effects and study design are detailed in the Supplementary Information (ClinicalTrials.gov identifier: NCT04248569).
Trial oversight
All the patients provided written informed consent. The study was conducted in accordance with the principles of the Declaration of Helsinki. The authors vouch for the accuracy and completeness of the data and for the fidelity of the trial to the protocol. The scientific review committee and institutional review board at the Johns Hopkins Sidney Kimmel Cancer Center reviewed the protocol and all amendments. All patients provided written informed consent before enrollment.
Study endpoints
The primary trial endpoint were safety, defined as the frequency of toxicities by grade assessed by documenting the drug-related adverse events using the NCI CTCAE Version 5.0 and neoepitope-specific T cells against DNAJ-PKAc in the peripheral blood at week 10. Validation data for the ELISpot assay used for the coprimary endpoint are provided (see ELISPOT Validation Report in Supplementary Information). Secondary endpoints include ORR, duration of response, DCR, PFS and overall survival.
Assessment
Disease assessments (by computed tomography and magnetic resonance imaging were performed every 8 weeks (±7 days) for the first 6 months and then every 12 weeks thereafter until disease progression, according to the definitions of RECIST, version 1.1. Adverse events, vital signs and laboratory tests were assessed at each visit and graded based on the NCI CTCAE Version 5.0. Survival, subsequent treatment and disease status were assessed every 12 weeks (window, ±2 weeks) after discontinuing treatment or disease progression (whichever occurred first) until the end of the trial, death, loss to follow-up or withdrawal of consent.
Statistical analysis
The primary endpoint for safety was the percent of adverse events by grade and for immunogenicity was the fold change in IFN-γ-producing DNAJ-PKAc-specific CD8+ and CD4+ T cells at week 10 after vaccination compared with unstimulated. Validation data for the ELISPOT assay is provided in a supplemental validation report. The evaluable population of the endpoint of T cell response consisted of all patients who received at least one dose of vaccine and had baseline and post-treatment T cell measures at 10 weeks. We anticipated that our regimen would result in at least a 2.5-fold increase of IFN-γ-producing DNAJ-PKAc-specific T cells with treatment. We assumed, conservatively, that the coefficient of variation of IFN-γ-producing DNAJ-PKAc -specific T cell density was 100%, and the correlation of the pre-and post-treatment measure is weak at 0.2. A sample size of 12 evaluable patients provided 81% power to detect a 2.5-fold increase based on a two-sided paired-sample t-test at a significance level of 0.05. Full statistical details are provided in the statistical analysis plan (see Protocol, Section 12, in Supplementary Information). Additional methods regarding correlative analyses are discussed in the Supplementary Information.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The study protocol is available in the Supplementary Information. The raw RNA sequencing data, 10x genomics scRNA sequencing data and raw TIRTL-seq are publicly available in the dbGaP repository (accession number phs003970). The remaining data that support the findings of this study are available within the Article and its Supplementary Information. All other data supporting the findings of this study are available from the corresponding authors upon reasonable request.
Code availability
This study did not generate new unique code.
References
Dinh, T. A. et al. A framework for fibrolamellar carcinoma research and clinical trials. Nat. Rev. Gastroenterol. Hepatol. 19, 328–342 (2022).
O’Neill, A. F. et al. Fibrolamellar carcinoma: an entity all its own. Curr. Probl. Cancer 45, 100770 (2021).
Stipa, F. et al. Outcome of patients with fibrolamellar hepatocellular carcinoma. Cancer 106, 1331–1338 (2006).
Honeyman, J. N. et al. Detection of a recurrent DNAJB1–PRKACA chimeric transcript in fibrolamellar hepatocellular carcinoma. Science 343, 1010–1014 (2014).
Schram, A. M., Chang, M. T., Jonsson, P. & Drilon, A. Fusions in solid tumours: diagnostic strategies, targeted therapy, and acquired resistance. Nat. Rev. Clin. Oncol. 14, 735–748 (2017).
Yang, W. et al. Immunogenic neoantigens derived from gene fusions stimulate T cell responses. Nat. Med. 25, 767–775 (2019).
Bauer, J. et al. The oncogenic fusion protein DNAJB1–PRKACA can be specifically targeted by peptide-based immunotherapy in fibrolamellar hepatocellular carcinoma. Nat. Commun. 13, 6401 (2022).
Kirk, A. M. et al. DNAJB1–PRKACA fusion neoantigens elicit rare endogenous T cell responses that potentiate cell therapy for fibrolamellar carcinoma. Cell Rep. Med. 5, 101469 (2024).
Pogorelyy, M. V. et al. TIRTL-seq: deep, quantitative and affordable paired TCR repertoire sequencing. Nat. Methods https://doi.org/10.1038/s41592-025-02907-9 (2025).
Bradley, P. & Thomas, P. G. Using T cell receptor repertoires to understand the principles of adaptive immune recognition. Annu Rev. Immunol. 37, 547–570 (2019).
Dash, P. et al. Quantifiable predictive features define epitope-specific T cell receptor repertoires. Nature 547, 89–93 (2017).
Franken, A. et al. CD4+ T cell activation distinguishes response to anti-PD-L1+ anti-CTLA4 therapy from anti-PD-L1 monotherapy. Immunity 57, 541–558.e7 (2024).
Nakazawa, M. et al. Anti-CTLA4 therapy leads to early expansion of a peripheral Th17 population and induction of Th1 cytokines. Cancer Immunol. Res. https://doi.org/10.1158/2326-6066.CIR-24-1055 (2025).
Thatikonda, V. et al. Comprehensive analysis of mutational signatures reveals distinct patterns and molecular processes across 27 pediatric cancers. Nat. Cancer 4, 276–289 (2023).
Ott, P. A. et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 547, 217–221 (2017).
El Dika, I. et al. A multicenter randomized three-arm phase II study of (1) everolimus, (2) estrogen deprivation therapy (EDT) with leuprolide + letrozole, and (3) everolimus + EDT in patients with unresectable fibrolamellar carcinoma. Oncologist 25, 925–e1603 (2020).
O’Neill, A. F. et al. Clinical and immunophenotype correlating with response to immunotherapy in paediatric patients with primary liver carcinoma. A case series. eBioMedicine. https://doi.org/10.1016/j.ebiom.2024.105147 (2024).
Abou-Alfa, G. K. et al. Evaluation of neratinib (N), pembrolizumab (P), everolimus (E), and nivolumab (V) in patients (pts) with fibrolamellar carcinoma (FLC). JCO 39, 310–310 (2021).
Chen, K. Y. et al. Clinical outcomes in fibrolamellar hepatocellular carcinoma treated with immune checkpoint inhibitors. Cancers 14, 5347 (2022).
Martini, D. J. & Wu, C. J. The future of personalized cancer vaccines. Cancer Discov. https://doi.org/10.1158/2159-8290.CD-25-0300 (2025).
Hilf, N. et al. Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature 565, 240–245 (2019).
Keskin, D. B. et al. Neoantigen vaccine generates intratumoral T cell responses in phase Ib glioblastoma trial. Nature 565, 234–239 (2019).
Puig-Saus, C. et al. Neoantigen-targeted CD8+ T cell responses with PD-1 blockade therapy. Nature 615, 697–704 (2023).
Alspach, E. et al. MHC-II neoantigens shape tumour immunity and response to immunotherapy. Nature 574, 696–701 (2019).
Huff, A. L. et al. CD4 T cell-activating neoantigens enhance personalized cancer vaccine efficacy. JCI Insight 8, e174027 (2023).
Braun, D. A. et al. A neoantigen vaccine generates antitumour immunity in renal cell carcinoma. Nature https://doi.org/10.1038/s41586-024-08507-5 (2025).
Rojas, L. A. et al. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature 618, 144–150 (2023).
Yarchoan, M. et al. Personalized neoantigen vaccine and pembrolizumab in advanced hepatocellular carcinoma: a phase 1/2 trial. Nat. Med. 30, 1044–1053 (2024).
Weber, J. S. et al. Individualised neoantigen therapy mRNA-4157 (V940) plus pembrolizumab versus pembrolizumab monotherapy in resected melanoma (KEYNOTE-942): a randomised, phase 2b study. Lancet 403, 632–644 (2024).
Hoyos, D. et al. Fundamental immune–oncogenicity trade-offs define driver mutation fitness. Nature 606, 172–179 (2022).
Chandran, S. S. et al. Immunogenicity and therapeutic targeting of a public neoantigen derived from mutated PIK3CA. Nat. Med. 28, 946–957 (2022).
Tran, E. et al. T-cell transfer therapy targeting mutant KRAS in cancer. N. Engl. J. Med. 375, 2255–2262 (2016).
Pant, S. et al. Lymph-node-targeted, mKRAS-specific amphiphile vaccine in pancreatic and colorectal cancer: the phase 1 AMPLIFY-201 trial. Nat. Med. 30, 531–542 (2024).
Leidner, R. et al. Neoantigen T-cell receptor gene therapy in pancreatic cancer. N. Engl. J. Med. 386, 2112–2119 (2022).
Acknowledgements
We are grateful to the patients and their families for their courage, commitment and invaluable contributions to this clinical trial. We thank M. Brancati, A. Ferguson, C. Salamone, C. D. Cannon, K. Munjal, P. Griffith, A. M. Salazar, A. Thomas, A. Thompson, C. Shubert, S. C. Yang, J. Saunders, J. Leatherman, J. Durham and additional physicians, nurses, staff and caregivers who cared for the participants during this clinical trial. We also thank providers at other institutions who contributed to the care of patients who participated in this study, including P. Kent and T. Stockwell (FibroFighters), W. Harris (University of Washington), A. O’Neill (Dana-Farber/Boston Children’s), J. Gordan (University of California San Francisco), D. Hsieh (UT Southwestern), S. Armstrong (Iowa University), S. Lee (MD Anderson) and A. Noonan (Ohio State). We thank K. Losert and J. Hopper from the Fibrolamellar Cancer Foundation for their support of this project. We thank R. Ashmun (St. Jude Flow Cytometry Core) and G. Lennon (St. Jude Immunology Flow Core) for cell sorting; the St. Jude Hartwell Center for sequencing; and the St. Jude Vector Core Lab for providing lentiviral backbones and packaging plasmids. This work was supported by R01-CA265009 to M.Y. and P.G.T. (grant nos. P30CA006973 and S10OD034407 to W.J.H), the Fibrolamellar Cancer Foundation, ASCO Career Development Award, BMS Rare Cancers, The Johns Hopkins Sidney Kimmel Cancer Core Grant (grant no. P30 CA006973), Poole Family Charitable Foundation in honor of Tessa Wiseman, the St. Jude Cancer Center (grant no. P30 CA021765), the Mark Foundation, the TIRTL Blue Sky Initiative and ALSAC at St. Jude. The funders had no role in the conduct of the trial or in the analysis of clinical or immunological endpoints.
Author information
Authors and Affiliations
Contributions
Author contributions: M.B., A.M.K., EJ., P.G.T. and M.Y. contributed to the conception and design of the work, acquisition, analysis and interpretation of data, draft of the manuscript and substantial revision to the manuscript. B.H.L., Z.K., K.J.B., W.J.H., H.W., M.F., R.A.A., C.T., J.N. and M.V.P. contributed to the analysis and interpretation of data and substantial revision to the manuscript. S.A., N.A.C., B.S., H.-C.K., J.H., H.Q., S.M.S., A.H., M.N., S.A.S. J.C.C., N.Z. and A.L.H. contributed to the analysis of data and substantial revision to the manuscript. The authors assume responsibility for the completeness and accuracy of the data.
Corresponding authors
Ethics declarations
Competing interests
Bristol Myers Squibb provided nivolumab and ipilimumab and support for the clinical trial. M.B. reports advisory board from AstraZeneca. A.M.K holds a patent for a previously described fusion-specific TCR in FLC (US patent number 20220251167). W.J.H. reports patent royalties from Rodeo/Amgen, received research funding from Sanofi, NeoTX, Riboscience (to Johns Hopkins), and speaking/travel honoraria from Exelixis and Standard BioTools. S.M.S received travel honoraria from Standard Biotools. J.C.C. reports patents related to TCR amplification, cloning and/or applications thereof and holds a patent for a previously described fusion-specific TCR in FLC (US patent numbers 20220251167, 20220326216, 20220136051). R.A.A. reports consulting arrangment with Bristol Myers Squib, AstraZeneca, Merck SD and Jazz Pharmaceuticals N.Z. reports research support from Bristol Myers Squibb, is a consultant for Genentech and receives other support from Adventris Pharmaceuticals. A.L.H. is a paid consultant of Adventris Pharmaceuticals, Inc. E.J. reports other support from Abmeta and Adventris, personal fees from Dragonfly, Neuvogen, STIMIT Tx, Mestag, Candel therapeutics, HDTbio, and grants from Lustgarten, Genentech, BMS, NeoTx, and Break Through Cancer. E.J. is a founder of and holds equity in Adventris Pharmaceuticals. She also serves as a consultant to the entity. M.V.P. has patents related to TCR amplification, cloning and/or applications thereof (US patent numbers 20250116673, 20250066852). P.G.T. is on the Scientific Advisory Board of Immunoscape and Shennon Bio; has received research support and personal fees from Elevate Bio; and consulted for CellCarta, RAVentures, 10X Genomics, Illumina, Pfizer, Cytoagents, Sanofi, Merck and JNJ. M.Y. receives grant/research support (to Johns Hopkins) from Bristol Myers Squibb, Exelixis, Incyte and Genentech; receives honoraria from Exelixis, AstraZeneca, Lantheus, Genentech and Incyte; is the coinventor of patents related to therapeutic cancer vaccines (managed by Johns Hopkins, US patent number 20230173049); and is a cofounder with equity of Adventris Pharmaceuticals. M.B., A.M.K, J.C.C., M.V.P., P.G.T. and M.Y. are coinventors on a pending patent application for the fusion-specific TCRs described herein. B.H.L., Z.K., K.J.B., S.A., N.A.C, B.S., H.W., H.-C.K., J.H., H. Q., A.H., M.N., S.A.S., M.F., R.A.A., C.T., A.G. and J.N. have no reported disclosures related to the current work.
Peer review
Peer review information
Nature Medicine thanks Alessandro Granito, David Pinato and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Ulrike Harjes, in collaboration with the Nature Medicine team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Efficacy in the intention to treat population.
a. Duration and type of response. The median duration of response was calculated with the use of the Kaplan–Meier method; responses were evaluated in accordance with Response Evaluation Criteria in Solid Tumors, version 1.1. b. The change from baseline in the target lesion diameter according to RECIST, version 1.1 (n = 16). c. Spider plot of radiographic responses to treatment. Tumor responses were measured at regular intervals, and the values shown are the largest percentage change in the sum of the longest diameters from the baseline measurements of each measurable tumor. Each line represents one patient (n = 16). d, e. Kaplan–Meier curves of OS (d) and PFS (e). The 95% CIs for point estimates are shown in red shading. DCR, disease control rate; ITT, intention to treat; ORR, overall response rate; OS, overall survival; PD, progressive disease; PFS, progression-free survival; PR, partial response; SD, stable disease: SLD, sum of longest diameters of the target lesions.
Extended Data Fig. 2 ELISpot responses for all patients.
Quantified ELISpot responses (mean ± SD of 3 replicate wells per condition) against the full 24-amino-acid peptide (Peptide 9), eight overlapping 9-amino-acid peptides (Peptides 1-8) spanning the chimera, negative controls (Medium-Only and Unstimulated), and positive control (CEF peptide pool, consists of epitopes from cytomegalovirus, Epstein-Barr virus, and influenza (flu) at baseline, week 10, and a subsequent timepoint where available. Unpaired two-sided t test comparing mean ELISpot repose to peptide vs unstimulated control; exact p-values for significant results are shown on the appropriate graphs. Line graphs show ELISpot response over time to each peptide or control stimulation, highlighting the unstimulated control (green) and Peptide 9 (blue).
Extended Data Fig. 3 CyTOF results for all patients.
a. Cytometry by time of flight (CyTOF) analysis of baseline (B) and post-treatment (P) PBMCs cultured with vehicle, an irrelevant peptide (KRAS G12D), or the full length FLC peptide, shows that treatment was associated with an expansion of CD4+ T helper cells, as well as activated CD8+ cytotoxic T cells. b. Quantification of expanded T helper cells and cytotoxic T cells from FLC peptide stimulated populations with TIM3hiPD1hi and TIM3loPD1hi expression. Paired two-sided t-test, ns p > 0.05. ThEFF, effector T cells, ThEM, effector memory T cells. UnB, unstimulated Baseline, Un P, unstimulated post- treatment.
Extended Data Fig. 4 Study sample timepoints for all patients.
a. Timepoints used for TIRTL-seq for each of the 12 evaluable patients are represented in green, and the timepoint from P12 used for scRNAseq is represented in cyan. b. Tumor biopsies available for each patient. Pre-treatment biopsies are indicated in pink, on-treatment in yellow, post-treatment in dark blue, and no biopsy available in black.
Extended Data Fig. 5 Longitudinal changes in peripheral blood TCR repertoires.
Scatterplots showing relative peripheral blood frequencies of unique TCR clones detected by TIRTL-seq for all 12 evaluable patients at baseline (C1D1, x-axis for all plots) and after one cycle of treatment (C2D1, y-axis on left plots for each patient) or two or more cycles of treatment (C3D1+, y-axis on left plots for each patient). Clones that were statistically expanded relative to baseline are outlined in red, statistically contracted clones are outlined in blue, and stable clones are outlined in gray. Patient labels are color coded by clinical response (red=PD, blue=SD, green=PR) and the treatment received during the first cycle of therapy (FLC-VAC alone or combined FLC-VAC+ICB) is indicated.
Extended Data Fig. 6 Fusion-specificity of TCRs identified in responder patient P12.
a. Quantification of flow cytometry results for screening of all five P12 fusion-specific TCRS against healthy donor PBMCs expressing different combinations of patient class II HLA alleles pulsed with 24mer vaccinated DNAJ-PKAc fusion peptide. HLA-DRB1*13:01 and HLA-DRB3*01:01 expression for each healthy donor is noted below the plot; see Supplementary Table 2 for full list of patient class II HLAs expressed by each healthy donor. Results shown for one experiment, normalized to unstimulated negative and PMA-ionomycin positive controls (see Supplementary Methods for formula). b. Quantification of flow cytometry results for screening of all five TCRs of interest against autologous LCLs pulsed with individual 15mer DNAJ-PKAc fusion peptides tiling across the fusion breakpoint. Results shown for duplicate experiments, represented as mean fold change relative to unstimulated control (peptide-specific %NFAT-GFP+ divided by unstimulated %NFAT-GFP+). c. Scatterplot showing relative peripheral blood frequencies of unique TCR clones detected by TIRTL-seq for P12 at baseline (C1D1, x-axis) and after two cycles of treatment (C3D1, y-axis) as shown in Fig. 3d, with additional fusion-specific TCR P12-TCR14 highlighted in pink. d. Representative flow cytometry plots for fusion specificity screening of P12-TCR14 TCR-expressing Jurkat cells against autologous LCLs (DRB1*13+ and DRB3*01+) and DRB1*13:01- DRB3*01+ LCLs pulsed with 24mer DNAJ-PKAc fusion peptide.
Extended Data Fig. 7 TCR similarity clustering of DRB3-restricted DNAJ-PKAc-specific TCRs across multiple DRB3+ patients, including a public DNAJ-PKAc-specific TCR.
a. TCRdist clustering network for longitudinally expanded clones after two or more cycles of treatment (C3D1+) as shown in Fig. 2h, highlighting DRB3*01:01-restricted P12-TCR2 clustered with two TCRs from P8 (P8-TCR1 and P8-TCR2). Highlighted nodes are colored by patient and outlined by TCR ID. b. Representative flow cytometry plots for fusion specificity screening of P8-TCR1 and P8-TCR2 TCR-expressing Jurkat cells against autologous LCLs pulsed with 24mer DNAJ-PKAc fusion peptide. c. Network of TCRs clustered (TCRdist≤120) with known DRB3-restricted fusion-specific TCRs, including P12-TCR2/P8-TCR2, P12-TCR3, and P12-TCR8/P12-TCR14. Nodes are colored by patient and known TCRs are outlined by TCR ID. Node size corresponds to node degree (number of neighbors). Inset shows CDR3 sequence logos colored by amino acid chemistry for effectively public P8-TCR2 and P12-TCR21, which differ by a single amino acid in the TCRβ chain with conserved chemistry. d. Same clustering network as c with nodes colored by patient DRB3 expression. e. V/J gene segment usage for P12-TCR3 cluster (top), P12-TCR2/P8-TCR2 cluster (middle), and P12-TCR8/P12-TCR14 cluster (bottom). The thickness of each path is proportional to the frequency of TCR clones within the cluster using a particular gene segment, and gene segments are colored by their frequency within the cluster using a fixed rank-order color scheme (that is red is most frequent, green is second most frequent, etc.). f. CDR3 sequence logos for CDR3α (left) and CDR3β (right) for P12-TCR3 cluster (top), P12-TCR2/P8-TCR2 cluster (middle), and P12-TCR8/P12-TCR14 cluster (bottom), colored by amino acid chemistry.
Extended Data Fig. 8 TCR similarity clustering with fusion-specific TCRs.
Heatmap of TCRs clustered (TCRdist≤120) with known fusion-specific TCRs, including P12-TCR2, P12-TCR3, P12-TCR6, P12-TCR8, P12-TCR13, P12-TCR14, and P8-TCR2. For each TCR, the annotation bars represent, from left to right, the patient ID, the timepoint where the TCR was sequenced, whether the TCR is a known fusion-specific TCR (green boxes mark the known TCRs listed above), and to which cluster (see Extended Data Fig. 7) the TCR belongs. The heatmap is clustered and colored by TCRdist score; the pairwise TCRdist score between each pair of TCRs is shown on the heatmap.
Extended Data Fig. 9 Whole transcriptome sequencing of primary, responsive, and resistant lesions.
Heatmaps with hierarchical clustering based on log transformed expression levels of a. antigen presentation machinery, b. adaptive immune response, c. innate immune response, d. checkpoint modulator, e. chemokine, f. cytokine, g. repair and replication deficit, and h. tumor associated antigen genes in response in primary, responding, and resistant patient specimens.
Extended Data Fig. 10 Myeloid reprogramming in a resistant lesion.
a. Representative results from imaging mass cytometry for select markers that denote macrophage subtypes in primary, responding, or resistant lesions from one patient. Scale bar: 100 μm. b. Scaled expression profiles for myeloid subtypes sub-clustered. c. Composition of macrophage subtypes shown as stacked barplots (left) and individual barplots (right). d. Per-cell correlation of myeloid marker expression in all macrophages. e. Violin plots showing per-cell macrophage expression of functional markers (metal intensity).
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–3, Methods and ELISpot Validation Report.
Supplementary Tables 1–13 (download XLSX )
Supplementary Tables 1–13.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Baretti, M., Kirk, A.M., Ladle, B.H. et al. A therapeutic peptide vaccine for fibrolamellar hepatocellular carcinoma: a phase 1 trial. Nat Med 31, 4246–4255 (2025). https://doi.org/10.1038/s41591-025-03995-y
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41591-025-03995-y
