
Abstract
Lung adenocarcinoma (LUAD) is an aggressive cancer defined by oncogenic drivers and metabolic reprogramming. Here we leverage next-generation spatial screens to identify glycogen as a critical and previously underexplored oncogenic metabolite. High-throughput spatial analysis of human LUAD samples revealed that glycogen accumulation correlates with increased tumour grade and poor survival. Furthermore, we assessed the effect of increasing glycogen levels on LUAD via dietary intervention or via a genetic model. Approaches that increased glycogen levels provided compelling evidence that elevated glycogen substantially accelerates tumour progression, driving the formation of higher-grade tumours, while the genetic ablation of glycogen synthase effectively suppressed tumour growth. To further establish the connection between glycogen and cellular metabolism, we developed a multiplexed spatial technique to simultaneously assess glycogen and cellular metabolites, uncovering a direct relationship between glycogen levels and elevated central carbon metabolites essential for tumour growth. Our findings support the conclusion that glycogen accumulation drives LUAD cancer progression and provide a framework for integrating spatial metabolomics with translational models to uncover metabolic drivers of cancer.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
All MALDI files in imzML format generated in this study are available at https://sunlabresources.rc.ufl.edu. Proteomics datasets are available on ProteomeXchange at https://doi.org/10.25345/C5N01058W and accession number PXD060090. Source data are provided with this paper.
References
Imielinski, M. et al. Mapping the hallmarks of lung adenocarcinoma with massively parallel sequencing. Cell 150, 1107–1120 (2012).
Russell, P. A. et al. Does lung adenocarcinoma subtype predict patient survival?: a clinicopathologic study based on the new International Association for the Study of Lung Cancer/American Thoracic Society/European Respiratory Society international multidisciplinary lung adenocarcinoma classification. J. Thorac. Oncol. 6, 1496–1504 (2011).
Dragani, T. A. et al. Lung adenocarcinoma diagnosed at a younger age is associated with advanced stage, female sex, and ever-smoker status, in patients treated with lung resection. Cancers 15, 2395 (2023).
Singh, S. S., Dahal, A., Shrestha, L. & Jois, S. D. Genotype driven therapy for non-small cell lung cancer: resistance, pan inhibitors and immunotherapy. Curr. Med. Chem. 27, 5274–5316 (2020).
Zagryazhskaya, A., Gyuraszova, K. & Zhivotovsky, B. Cell death in cancer therapy of lung adenocarcinoma. Int. J. Dev. Biol. 59, 119–129 (2015).
Keibler, M. A. et al. Metabolic requirements for cancer cell proliferation. Cancer Metab. 4, 16 (2016).
Pavlova, N. N. & Thompson, C. B. The emerging hallmarks of cancer metabolism. Cell Metab. 23, 27–47 (2016).
Kimmelman, A. C. & White, E. P. Autophagy and tumor metabolism. Cell Metab. 25, 1037–1043 (2017).
Sullivan, M. R. et al. Quantification of microenvironmental metabolites in murine cancers reveals determinants of tumor nutrient availability. eLife 8, e44235 (2019).
Prabakaran, S. Kras rewires metabolic networks. Sci. Signal. 9, ec56 (2016).
Liu, J., Zhang, C., Hu, W. & Feng, Z. Tumor suppressor p53 and its mutants in cancer metabolism. Cancer Lett. 356, 197–203 (2015).
DeBerardinis, R. J. & Chandel, N. S. Fundamentals of cancer metabolism. Sci. Adv. 2, e1600200 (2016).
Martínez-Reyes, I. & Chandel, N. S. Cancer metabolism: looking forward. Nat. Rev. Cancer 21, 669–680 (2021).
Nwosu, Z. C. et al. Uridine-derived ribose fuels glucose-restricted pancreatic cancer. Nature https://doi.org/10.1038/s41586-023-06073-w (2023).
Sanderson, S. M., Gao, X., Dai, Z. & Locasale, J. W. Methionine metabolism in health and cancer: a nexus of diet and precision medicine. Nat. Rev. Cancer 19, 625–637 (2019).
Young, L. E. et al. In situ mass spectrometry imaging reveals heterogeneous glycogen stores in human normal and cancerous tissues. EMBO Mol. Med. 14, e16029 (2022).
Hawkinson, T. R. & Sun, R. C. Matrix-assisted laser desorption/ionization mass spectrometry imaging of glycogen in situ. Mass Spectr. Imag. Small Mol. Methods Protoc. 2437, 215–228 (2022).
Zhou, Z. et al. Clear cell adenocarcinoma of the urinary bladder is a glycogen-rich tumor with poorer prognosis. J. Clin. Med. 9, 138 (2020).
Zhou, Z. et al. Clinical features, survival and prognostic factors of glycogen-rich clear cell carcinoma (GRCC) of the breast in the US population. J. Clin. Med. 8, 246 (2019).
Sun, R. C. et al. Nuclear glycogenolysis modulates histone acetylation in human non-small cell lung cancers. Cell Metab. 30, 903–916 (2019).
Liu, Q. et al. Glycogen accumulation and phase separation drives liver tumor initiation. Cell 184, 5559–5576 (2021).
Curtis, M. et al. Fibroblasts mobilize tumor cell glycogen to promote proliferation and metastasis. Cell Metab. 29, 141–155 (2019).
Deng, Y. et al. Spatial profiling of chromatin accessibility in mouse and human tissues. Nature 609, 375–383 (2022).
Rappez, L. et al. SpaceM reveals metabolic states of single cells. Nat. Methods 18, 799–805 (2021).
Cao, J. et al. Deciphering the metabolic heterogeneity of hematopoietic stem cells with single-cell resolution. Cell Metab. 36, 209–221 (2024).
Sun, C. et al. Spatially resolved multi-omics highlights cell-specific metabolic remodeling and interactions in gastric cancer. Nat. Commun. 14, 2692 (2023).
Buglakova, E. et al. Spatial single-cell isotope tracing reveals heterogeneity of de novo fatty acid synthesis in cancer. Nat. Metab. https://doi.org/10.1038/s42255-024-01118-4 (2024).
Schwaiger-Haber, M. et al. Using mass spectrometry imaging to map fluxes quantitatively in the tumor ecosystem. Nat. Commun. 14, 2876 (2023).
Conroy, L. R. et al. Spatial metabolomics reveals glycogen as an actionable target for pulmonary fibrosis. Nat. Commun. 14, 2759 (2023).
Hirase, H., Akther, S., Wang, X. & Oe, Y. Glycogen distribution in mouse hippocampus. J. Neurosci. Res. 97, 923–932 (2019).
Nakamura-Tsuruta, S. et al. Comparative analysis of carbohydrate-binding specificities of two anti-glycogen monoclonal antibodies using ELISA and surface plasmon resonance. Carbohydr. Res. 350, 49–54 (2012).
Donohue, K. J. et al. Gys1 antisense therapy prevents disease-driving aggregates and epileptiform discharges in a Lafora disease mouse model. Neurotherapeutics 20, 1808–1819 (2023).
Oe, Y., Baba, O., Ashida, H., Nakamura, K. C. & Hirase, H. Glycogen distribution in the microwave‐fixed mouse brain reveals heterogeneous astrocytic patterns. Glia 64, 1532–1545 (2016).
Markussen, K. H. et al. The multifaceted roles of the brain glycogen. J. Neurochem. 168, 728–743 (2023).
Lu, B. et al. Metabolic crosstalk: molecular links between glycogen and lipid metabolism in obesity. Diabetes 63, 2935–2948 (2014).
Nilsson, L. H. & Hultman, E. Liver glycogen in man—the effect of total starvation or a carbohydrate-poor diet followed by carbohydrate refeeding. Scand. J. Clin. Lab. Invest. 32, 325–330 (1973).
Charrez, B., Qiao, L. & Hebbard, L. The role of fructose in metabolism and cancer. Horm. Mol. Biol. Clin. Invest. 22, 79–89 (2015).
Goncalves, M. D. et al. High-fructose corn syrup enhances intestinal tumor growth in mice. Science 363, 1345–1349 (2019).
Escrich, R. et al. A high-corn-oil diet strongly stimulates mammary carcinogenesis, while a high-extra-virgin-olive-oil diet has a weak effect, through changes in metabolism, immune system function and proliferation/apoptosis pathways. J. Nutr. Biochem. 64, 218–227 (2019).
Wu, B. et al. Dietary corn oil promotes colon cancer by inhibiting mitochondria-dependent apoptosis in azoxymethane-treated rats. Exp. Biol. Med. 229, 1017–1025 (2004).
He, X.-Y. et al. Chronic stress increases metastasis via neutrophil-mediated changes to the microenvironment. Cancer Cell 42, 474–486 (2024).
Ghaddar, N. et al. The integrated stress response is tumorigenic and constitutes a therapeutic liability in KRAS-driven lung cancer. Nat. Commun. 12, 4651 (2021).
Worby, C. A., Gentry, M. S. & Dixon, J. E. Laforin, a dual specificity phosphatase that dephosphorylates complex carbohydrates. J. Biol. Chem. 281, 30412–30418 (2006).
Zacharek, S. J. et al. Lung stem cell self-renewal relies on BMI1-dependent control of expression at imprinted loci. Cell Stem Cell 9, 272–281 (2011).
McCorvie, T. J. et al. Molecular basis for the regulation of human glycogen synthase by phosphorylation and glucose-6-phosphate. Nat. Struct. Mol. Biol. 29, 628–638 (2022).
Xie, H. et al. Glycogen metabolism is dispensable for tumour progression in clear cell renal cell carcinoma. Nat. Metab. 3, 327–336 (2021).
Paglia, G. et al. Ion mobility derived collision cross sections to support metabolomics applications. Anal. Chem. 86, 3985–3993 (2014).
Goldsmith, E., Fletterick, R. & Withers, S. The three-dimensional structure of acarbose bound to glycogen phosphorylase. J. Biol. Chem. 262, 1449–1455 (1987).
Adisakwattana, S., Lerdsuwankij, O., Poputtachai, U., Minipun, A. & Suparpprom, C. Inhibitory activity of cinnamon bark species and their combination effect with acarbose against intestinal α-glucosidase and pancreatic α-amylase. Plant Foods Hum. Nutr. 66, 143–148 (2011).
Martin, A. E. & Montgomery, P. A. Acarbose: an α-glucosidase inhibitor. Am. J. Health Syst. Pharm. 53, 2277–2290 (1996).
Commisso, C. et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 497, 633–637 (2013).
Varani, J. Healthful eating, the Western style diet and chronic. Approaches Poult. Dairy Vet. Sci. 387, 1377–1396 (2017).
Carbone, D. P., Gandara, D. R., Antonia, S., Zielinski, C. & Paz-Ares, L. G. Non–small-cell lung cancer: role of the immune system and potential for immunotherapy. J. Thorac. Oncol. 10, 974–984 (2015).
Salem, A. et al. Targeting hypoxia to improve non–small cell lung cancer outcome. J. Natl Cancer Inst. 110, 14–30 (2018).
White, E. S. Lung extracellular matrix and fibroblast function. Ann. Am. Thorac. Soc. 12, S30–S33 (2015).
Zhang, H. et al. Lkb1 inactivation drives lung cancer lineage switching governed by polycomb repressive complex 2. Nat. Commun. 8, 14922 (2017).
DePaoli-Roach, A. A. et al. Laforin and malin knockout mice have normal glucose disposal and insulin sensitivity. Hum. Mol. Genet. 21, 1604–1610 (2012).
Ganesh, S. et al. Targeted disruption of the Epm2a gene causes formation of Lafora inclusion bodies, neurodegeneration, ataxia, myoclonus epilepsy and impaired behavioral response in mice. Hum. Mol. Genet. 11, 1251–1262 (2002).
Xu, C. et al. Loss of Lkb1 and Pten leads to lung squamous cell carcinoma with elevated PD-L1 expression. Cancer Cell 25, 590–604 (2014).
Fillmore, C. M. et al. EZH2 inhibition sensitizes BRG1 and EGFR mutant lung tumours to TopoII inhibitors. Nature 520, 239–242 (2015).
Clarke, H. A. et al. Spatial metabolome lipidome and glycome from a single brain section. Preprint at bioRxiv https://doi.org/10.1101/2023.07.22.550155 (2023).
Stanback, A. E. et al. Regional N-glycan and lipid analysis from tissues using MALDI-mass spectrometry imaging. STAR Protoc. 2, 100304 (2021).
Lee, J. H. et al. Lung stem cell differentiation in mice directed by endothelial cells via a BMP4–NFATc1–thrombospondin-1 axis. Cell 156, 440–455 (2014).
Navarrete-Perea, J., Yu, Q., Gygi, S. P. & Paulo, J. A. Streamlined tandem mass tag (SL-TMT) protocol: an efficient strategy for quantitative (phospho) proteome profiling using tandem mass tag-synchronous precursor selection-MS3. J. Proteome Res. 17, 2226–2236 (2018).
Li, J. et al. TMTpro-18plex: the expanded and complete set of TMTpro reagents for sample multiplexing. J. Proteome Res. 20, 2964–2972 (2021).
Schweppe, D. K. et al. Full-featured, real-time database searching platform enables fast and accurate multiplexed quantitative proteomics. J. Proteome Res. 19, 2026–2034 (2020).
Schweppe, D. K. et al. Characterization and optimization of multiplexed quantitative analyses using high-field asymmetric-waveform ion mobility mass spectrometry. Anal. Chem. 91, 4010–4016 (2019).
Rad, R. et al. Improved monoisotopic mass estimation for deeper proteome coverage. J. Proteome Res. 20, 591–598 (2020).
Beausoleil, S. A., Villén, J., Gerber, S. A., Rush, J. & Gygi, S. P. A probability-based approach for high-throughput protein phosphorylation analysis and site localization. Nat. Biotechnol. 24, 1285–1292 (2006).
Juras, J. A. et al. In situ microwave fixation provides an instantaneous snapshot of the brain metabolome. Cell Rep. Methods 3, 100455 (2023).
Tang, Z. et al. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 45, W98–W102 (2017).
Acknowledgements
This study was supported by National Institute of Health (NIH) grants R01AG066653, R01CA266004, R01AG078702, R01CA288696, V-Scholar Grant, and RM1NS133593 to R.C.S., R35NS116824 to M.S.G., R35GM142701 to L.C., T32HL134621 to H.A.C., and R01CA237643 and P20GM121327-03 to C.F.B. This research was also supported by the Biospecimen Procurement and Translational Pathology Shared Resource Facility of the University of Kentucky Markey Cancer Center (P30CA177558) to D.B.A. and supported in part by an NIH award, S10 OD030293, for MRI/S instrumentation. Z.L. is supported by the MBI Gator NeuroScholar Program. Large language models, for example, ChatGPT, were used to make minor grammatical improvements in the text. We thank N. R. Gough (BioSerendipity, LLC) for critical discussions and editorial assistance.
Author information
Authors and Affiliations
Contributions
Conceptualization, R.C.S.; methodology, R.C.S.; investigation, H.A.C., T.R.H., C.J.S., T.M., R.A.R., L.W., J.E.C., L.E.A.Y., J.A.J., M.D.B., A.N.J., A.M., J.F.L., E.C.M., X.M., L.C., Z.L., Y.X., Y.H., X.H., A.R., M.E.M., P.K.S., J.B., G.L.A., B.M.E., C.W.V.K., C.F.B., D.B.A., M.S.G. and R.C.S.; writing—original draft, R.C.S.; writing—review and editing, R.C.S., H.A.C., C.J.S., T.R.H., C.F.B., L.E.A.Y., C.W.V.K., M.S.G. and D.B.A.; funding acquisition, M.S.G. and R.C.S.; resources, M.S.G. and R.C.S; supervision, M.S.G. and R.C.S.
Corresponding authors
Ethics declarations
Competing interests
R.C.S. has received research support and consultancy fees from Maze Therapeutics. R.C.S. is a member of the Medical Advisory Board for Little Warrior Foundation. M.S.G. has research support and research compounds from Maze Therapeutics, Valerion Therapeutics and Ionis Pharmaceuticals. M.S.G. also received consultancy fee from Maze Therapeutics, PTC Therapeutics and the Glut1-Deficiency Syndrome Foundation. D.B.A. receives book royalty from Wolters Kluwer. The other authors declare no competing interests.
Peer review
Peer review information
Nature Metabolism thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Alfredo Giménez-Cassina, in collaboration with the Nature Metabolism team.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
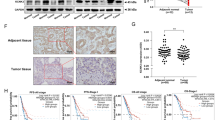
Extended Data Fig. 1 Validation of MALDI glycogen imaging with anti-glycogen antibody staining in lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) tissues.
a, Representative images of MALDI glycogen imaging (left) and corresponding anti-glycogen antibody staining (right) in two LUAD and one LUSC patient tissue samples. b, Detailed view of various tumor regions stained with anti-glycogen antibody showing predominately tumors positive for glycogen similar to MALDI analysis. Labels indicate tumor (T), stroma (S), normal adjacent tissue (N), vessels (V), necrosis (Ne), smooth muscle (SM), cartilage (C), and chondrocytes (Cc). c, Tissue overlays combining anti-glycogen antibody staining and MALDI glycogen imaging performed in SCILs software. The upper panels show the antibody staining with outlined regions for zoomed-in views, and the lower panels display the corresponding MALDI imaging overlays. c and c represent three biological replicates (individual patients). Black boxes indicate areas magnified in the zoomed-in images to illustrate the concordance between glycogen imaging and antibody staining.
Extended Data Fig. 2 MALDI glycogen imaging workflow and profiling of human tissue microarray (TMA) and mouse GEMM tumors.
a, Schematic of the MALDI glycogen imaging workflow. Tissue sections are subjected to antigen retrieval, followed by treatment with isoamylase and application of the CHCA matrix. The isoamylase enzyme digests glycogen into glucose polymers, which are then detected and visualized using MALDI mass spectrometry. Created with BioRender.com. b, Representative H&E staining (top row) and MALDI glycogen imaging (bottom row) of lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) tissue cores from a TMA, image represents single run from three repeats, Scale bar: 2 mm. c, Representative H&E staining (top row) and MALDI glycogen imaging (bottom row) of normal lung tissue and various genetically engineered mouse model (GEMM) tumors. The GEMM tumors include K (Kras), KP (Kras; Pten), KL (Kras; Lkb1), and E (Egfr). Scale bar represents 3 mm. d, Quantification of total glycogen levels in normal lung tissue and GEMM tumors, as measured by MALDI glycogen imaging. Bar graph shows the relative abundance of glycogen in normal, K, KP, KL, and E tissues (n = 6/group; mean +/−s.e.m. p-values indicated; one-way ANOVA and Tukey’s multiple comparison test).
Extended Data Fig. 3 Determination of absolute glycogen concentration using spotted standards by MALDI imaging.
a, Scanned image showing the location of spotted glycogen standards (0.06, 0.18, 0.55, 1.6, and 5 ng) next to a tissue section on a microscope slide. b, Mass spectra of different glycogen concentrations spotted on the slide, indicating the glucose polymer 7 m/z peaks corresponding to varying amounts of glycogen. c, XY plots showing the relationship between glycogen concentration (ng) and relative intensity per pixel for each spotted standard. d, Log-transformed plots of relative intensity versus glycogen concentration for the standards for different glucose chain length indicated above. The linear regression lines indicate the strong correlation used for glycogen quantification. R² values for each plot are shown, line equations are displayed on top. e, Absolute quantification of glycogen in four human lung adenocarcinoma (LUAD) and one lung squamous cell carcinoma (LUSC) tissue sections (n = 3 ROIs of 500 pixels/tissue; mean +/− s.e.m. p-values indicated; one-way ANOVA adjusted for Tukey’s multiple comparisons).
Extended Data Fig. 4 Glycogen accumulation correlates with tumor grade and predicts survival outcomes.
a, Representative immunohistochemical staining of glycogen in well differentiated (top) and poorly differentiated (bottom) tumour regions. T, tumour tissue; S, stromal tissue. b, Quantification of normalized glycogen intensity (adjusted for stromal regions) across tumour samples stratified by differentiation status: well, moderate, and poor in 247 LUAD patients (mean +/- s.e.m. p-values indicated). c, Kaplan-Meier survival curves illustrating the relationship between glycogen levels and patient survival. Top: Patients divided by the median glycogen expression level (low vs. high). Bottom: Survival analysis stratified into glycogen low (quartile), mid (50th percentile), and high (quartile) groups. Log-rank test p-value indicates a significant association between high glycogen content and decreased survival (p < 0.0001). d, Predictive performance of a random forest model for glycogen intensity in relation to survival outcomes, shown through the area under the receiver operating characteristic (AUROC = 0.846) and the area under the precision-recall curve (AUPRC = 0.888).
Extended Data Fig. 5 Impact of different diets on glycogen and glucose metabolism in C57BL/6 J mice.
a, Top: Schematic of the experimental design where C57BL/6 J mice were fed with vehicle, corn oil, high-fructose corn syrup (HFCS), or a combination (Combo) diet for 2 weeks. Created with BioRender.com. Bottom: MALDI glycogen imaging of lung tissues from mice subjected to different dietary treatments. Scale bar: 2 mm. b, Glycogen structure analysis in the lung tissues of WT mice treated with different diets. The graph shows the relative abundance of glycogen chain lengths in mice fed with corn oil, HFCS, or the Combo diet (n = 3 mice; mean +/− s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons). c, Glucose tolerance test results for mice on Day 1 (left) and Day 14 (right) post-treatment. The graphs show changes in blood glucose levels (mg/dL) over time following glucose gavage. No significant differences (ns) were observed among the different diet groups (n = 3 mice; mean +/− s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons). d, Fasting blood glucose levels (mg/dL) in the same cohort of mice. No significant differences (ns) were observed among the different diet groups (n = 3 mice; mean +/− s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons). e, Body weight (g) of mice over the 14-day treatment period, showing no significant differences among the diet groups (n = 3 mice; mean +/− s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons). f, Changes in blood glucose levels (mg/dL) over the 14-day treatment period, showing no differences among the diet groups (n = 3 mice; mean +/− s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons). g, Liver glycogen imaging using CL6 staining in mice fed with vehicle, corn oil, HFCS, or the Combo diet. Scale bar: 2 mm. h, Quantification of total glycogen in liver tissues from mice subjected to different dietary treatments (n = 3 mice; mean +/− s.e.m. p-values indicated; one-way ANOVA adjusted for Tukey’s multiple comparison). i, Glycogen chain length analysis in normal adjacent tissues from KP and KP Combo-treated mice related to Fig. 3. The graph shows the relative intensity of glycogen chain lengths, (n = 6 mice; mean +/− s.e.m. two-way ANOVA for Tukey’s multiple comparison).
Extended Data Fig. 6 Laforin expression, phosphorylated glycogen analysis, and glycogen dynamics in LKO animals.
a, Representative IHC images of tissue cores and zoomed-in views stained with antibodies against Laforin in LUSC (leftmost), Laforin in LUAD (second from left), GP (glycogen phosphorylase), and GYS (glycogen synthase,), image represents single replicates from three repeats. b, Phosphorylated glycogen chain length analysis in LUAD and LUSC tissue cores from TMA (n = 67 patients for LUAD, 52 for LUSC, mean +/− s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons). c, Glycogen chain length analysis in the lungs of WT and LKO (Laforin knockout) mice over 1-, 2, and 4- months (n = 4 animals/group mean +/−s.e.m. p-values indicated; two-way ANOVA adjusted for Tukey’s multiple comparisons).
Extended Data Fig. 7 Lung stem cell growth in vitro, H&E and MALDI glycogen imaging of different mouse models.
a, Schematic of isolating bronchoalveolar stem cells and differentiation to bronchiolar and alveolar organoids in matrigel. b, Representative bright field images of bronchiolar and alveolar organoids derived from WT and laforin−/− (LKO) animals. c, Number of colonies and colony size from bronchiolar and alveolar organoids derived from WT and laforin−/− (LKO) animals. Values are presented as mean +/− s.e.m. p-values were calculated using two-tailed t-test. d, Additional representative H&E images and MALDI imaging of an adjacent tissue section showing tumor formation and glycogen levels LSL-KrasG12D/p53fl/fl (KP) and LSL-KrasG12D/p53fl/fl /LKO(KPL) (n = 3 each). e, Additional representative H&E images and MALDI imaging of an adjacent tissue section showing tumor formation and glycogen levels LSL-KrasG12D/p53fl/fl (KP); LSL-KrasG12D/p53fl/fl/Gysfl/fl (KPG); LSL-KrasG12D/p53fl/fl/Gysfl/fl:Vehicle (KPG:V); LSL-KrasG12D/p53fl/fl/Gysfl/fl:Combination diet (KPG/C)animals (n = 2 each). f, Distribution of tumor grades across KP and KPL cohorts of mice (n = 3 mice; mean +/− s.e.m. p-values indicated; one-way ANOVA adjusted for Tukey’s multiple comparisons). g. Kaplan Meier survival analysis for mice in different groups. Log-rank test stated.
Extended Data Fig. 8 Comparative proteomics, phosphoproteomic, and spatial metabolomics analysis in KP and KPL tumors.
a, left: Principal component analysis (PCA) of 5993 proteins from proteomics data, showing no major changes between KP and KPL tumors. right: Volcano plot of proteomics data, with log2 fold change (log2FC) on the x-axis and -log10 p-value on the y-axis, indicating no significant differential expression between KP and KPL tumors. b, Left: Principal component analysis (PCA) of 5323 proteins from phosphoproteomic data, showing no major changes between KP and KPL tumors. Right: Volcano plot of phosphoproteomic data, with log2 fold change (log2FC) on the x-axis and -log10 p-value on the y-axis, indicating no significant differential phosphorylation between KP and KPL tumors. c, H&E-stained images of adjacent tissue slices used on MALDI analysis from KP (left) and KPL (right) tumors, image represents single analysis from three repeats. d, MALDI imaging of various metabolites in the lungs of KP and KPL GEMM tumors. Metabolites analyzed include glutamate, glycerol-3-phosphate (G3P), aspartate, glutathione (GSH), aconitate, ascorbic acid, uridine monophosphate (UMP), cyclic adenosine monophosphate (cAMP), glutamine, citric acid, adenosine monophosphate (AMP), ADP, malate, succinate, adenosine triphosphate (ATP), pyruvate, glucose/inositol, inosine, lactate, and arachidonic acid.
Extended Data Fig. 9 Multiplexed spatial metabolomics and glycogen analysis with 13C-glucose tracer enrichment.
a, Schematic of the multiplexed spatial metabolomics and glycogen workflow. Cells grown in chamber are grown in 12C- or 13C-glucose. After enrichment period, cells in chamber wells are imaged for metabolomics with NEDC, followed by antigen retrieval, isoamylase treatment, and CHCA matrix application for glycogen imaging. Created with BioRender.com. b, Layout of tracer enrichment in a chamber well format. Cells are exposed to 12C-glucose or 13C-glucose for 24, 48, and 72 hours in different wells. c, Metabolic pathway of 13C-glucose showing its incorporation into glycolysis and the TCA cycle, and 13C-glycogen formation. Key metabolic intermediates such as pyruvate (P), phosphoenolpyruvate (PEP), citrate (Cit), and oxaloacetate (OAA) are highlighted, along with enzymes pyruvate carboxylase (PC) and pyruvate dehydrogenase (PDH). d, Enrichment of various isotopologues in metabolites of glycolysis, TCA cycle, nucleotides, and glycogen. Fraction labeled graphs for m42 (glycogen), m6 (glucose and G6P), m5 (AMP), m3 (pyruvate, PEP, 3PG, lactate), and m2 (citric acid, GSH, glutamate, glutamine, malate) are shown at 0, 24, 48, and 72 hours. (n = 6697 individual pixels measured by MALDI imaging. e, Representative examples of 13C isotopologues detected through MALDI imaging, with 12C-glucose wells as controls. Metabolites visualized include citric acid, AMP, pyruvate, and glutamate.
Extended Data Fig. 10 Ion mobility and MALDI imaging of 12C and 13C-labeled glycogen species.
a, Mass spectra showing both 12C and 13C-labeled glycogen-released glucose chain lengths. b, Schematic of ion mobility drift separation for 13C-labeled species. The diagram illustrates how 12C and 13C glycogen species are separated based on their drift times in the ion mobility spectrometer. Created with BioRender.com. c, 2D plot of ion mobility drift time versus m/z (mass-to-charge ratio), with a zoomed-in view highlighting the separation between 12C-GP7 and 13C-GP7 species. The plot shows similar drift times for both 12C and 13C glycogen species, indicating effective alignment. d, Overlay of mass spectra from unlabeled (12C) and 13C-glucose labeled wells (24 hours) for glycogen. The overlay highlights the differences in peak intensities, indicating the incorporation of 13C into the glycogen molecules. e, Detailed overlay of 12C and 13C-labeled glycogen species, showing the enrichment of fully labeled 13C-GP7 at m/z 42. Representative MALDI images are included to show the spatial distribution of glycogen species at 0 (12 C), 24, 48, and 72 hours of 13C labeling.
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–8 and Tables 1–4.
Source data
Source Data Figs. 1–5 (download ZIP )
Statistical source data for Figs. 1–5.
Source Data Extended Data Figs. 2–9 (download ZIP )
Statistical source data for Extended Data Figs. 2–10.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Clarke, H.A., Hawkinson, T.R., Shedlock, C.J. et al. Glycogen drives tumour initiation and progression in lung adenocarcinoma. Nat Metab 7, 952–965 (2025). https://doi.org/10.1038/s42255-025-01243-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s42255-025-01243-8
This article is cited by
-
Decoding cancer across scales with metabolomics
Nature Reviews Cancer (2026)
-
Catabolic rewiring in cancer impacts dietary interventions
Communications Biology (2025)
