
Abstract
Human iPSC-derived models currently used in eye research usually replicate later events of tissue differentiation rather than early steps involving concurrent development of diverse embryonic cell types. Here we present a multi-timepoint morphological and transcriptomic analysis of a 2D model mirroring the early stages of human whole eye development, self-formed ectodermal autonomous multi-zone (SEAM) of ocular cells. SEAM cultures maintained a reproducible growth profile over the standard 28-day differentiation process with quantifiable morphological changes accompanying generation of key ocular cell types over time. Bulk and single-cell RNA-seq analyses at Days 0, 14 and 28 identified dynamic transcriptomic changes indicative of the emerging cell types, including rare stem cell-like populations analogous to those comprising the ciliary marginal zone, transit-amplifying cells, limbal epithelial stem cells, and corneal stromal stem cells. Integrated developmental trajectory analysis highlighted intermediate differentiation states underlying SEAM maturation. Cluster-specific interrogation of eye disease-associated genes demonstrated dynamic temporal patterns and enrichment in relevant developing cell types. These analyses establish a comprehensive baseline of SEAM formation, supporting the potential of the model to facilitate mechanistic studies of genetic variants that may uniquely impact humans, thus improving the success rate in resolving cases presenting with a broad range of developmental eye phenotypes.
Similar content being viewed by others


Introduction
Human eye development is a complex process mediated by a coordinated series of cellular and molecular events influenced by reciprocal interactions between emerging ocular tissues derived from different primordial cell lineages1,2. As each ocular tissue is essential for proper function of the eye, defects in their induction, proliferation, or differentiation can result in a range of disorders compounded by partial or complete vision loss. Genetic or environmental disruption early in eye development typically results in severe structural anomalies such as microphthalmia, anophthalmia, or coloboma (MAC)3. Whereas improper development at later stages tend to cause more isolated phenotypes, such as anterior segment dysgenesis, infantile glaucoma, cataracts, corneal opacities, retinal dystrophies, and other early-onset ocular disorders4. While there are numerous well-described genes known to be associated with ocular conditions, many cases remain unexplained, requiring further investigation into novel mechanisms.
Studies in vertebrate models provide insight into the conserved mechanisms governing ocular development. Nevertheless, these models are not sufficient to evaluate less conserved non-coding genetic variants, or those that display functional divergence. Advances in the generation of human organoids derived from human induced pluripotent stem cells (iPSCs) provide a platform to investigate such variants, however, currently available models are more suited for studies into specialized tissue differentiation rather than the concurrent development of diverse embryonic cell types of the whole eye. The recently presented 2D model representing whole eye development derived from human iPSCs, known as self-formed ectodermal autonomous multi-zone (SEAM) of ocular cells5,6 mimics the earlier stages of human whole eye development, which is supported by the initial expression studies of several cell type-specific markers, as well as the recently published single cell analyses of SEAM cultures at several time points7.
Here we provide an independent in-depth temporal evaluation of the morphological and transcriptomic changes that accompany SEAM development over the standard 28-day differentiation process, with an emphasis on the intermediate (Day 14) and late (Day 28) stages of differentiation. By integrating bulk and single-cell RNA sequencing with dynamic changes in morphology and growth, zone-specific spatial expression of ocular markers, and enrichment of biological processes and signaling pathways this study offers novel insight into the progression of SEAM formation as a model for the early stages of human eye development. Moreover, single cell data were further interrogated to identify expression patterns of genes associated with ocular disorders, supporting the use of this model for future studies of normal developmental processes, as well as mechanistic assessment of pathogenic and candidate genetic variants associated with complex ocular phenotypes.
Results
Temporal profile of SEAM growth and differentiation
SEAM cultures maintained a consistent and reproducible growth profile over the course of the 28-day differentiation process (Fig. 1A) consistent with that originally described for this culture system5,6. Zones were easily distinguished by differences in cell morphology and density, from Zone 1 characterized by the most densely packed cells to Zone 4 showing more sparse cells with a stellate morphology (Fig. 1B). The innermost zone (Zone 1) formed first after 5 days in differentiation medium, followed by a visibly distinct Zone 2 by Day 7. By Day 14, external borders between Zones 1, 2, and 3 were clearly visible, distinguishing the emerging Zone 4 as loosely packed mesenchymal cells that spanned between SEAMs. All zones became more defined as the cultures matured through Day 28 (Fig. 2A).

Overview of human iPSC-derived SEAM differentiation. (a) Workflow for generating SEAM cultures from human iPSCs. SEAM cultures form 4 concentric zones over the course of a 28-day differentiation protocol, each consisting of cells analogous to ocular cell types found in the developing human eye. Immunostaining and transcriptomic profiling were performed at Day 0, 14 and 28. (b) Individual zones (dotted lines) and lens clusters are visually distinguishable at 28 days under differentiation conditions. Scale bar, 200 μm.

Temporal profile of SEAM growth and differentiation. (a) Representative images of an individual SEAM (SEAM 9) undergoing differentiation at Day 0 (asterisk), 4, 7, 14, 21 and 28. (b) Overall area of individual SEAMs assessed across several time points (Day 0: 2224.8 μm2 ± 1264.1; Day 7: 35,644.0 μm2 ± 8232.6; Day 14: 36,319.7 μm2 ± 8763.7; Day 21: 25,369.5 μm2 ± 11,124.4; Day 28: 31,499.7 μm2 ± 19,229.3). SEAM 9 indicated with an arrow for reference. (c) Comparison of the overall area of each distinct zone (Zones 1, 2 and 3) at each time point. (d) Schematic illustrating SEAM growth over the course of the 28-day differentiation process. n = 12 SEAMs. One-way ANOVA, values represent mean ± SD; **** p < 0.0001. Scale bar, 200 μm.
To address observed variability in comparative SEAM size (Supplemental Fig. 1A), overall area (Fig. 2B), width (Supplemental Fig. 1B), and height (Supplemental Fig. 1C) of individual SEAMs, as well as the area of each distinct zone (Fig. 2C), were measured at several time points (Day 0, 7, 14, 21 and 28; n = 12 SEAMs). When normalized to initial colony size at Day 0, there was a 20.54 ± 12.93-fold increase in the overall area of individual SEAMs by Day 7 which remained consistent through Day 28. Moreover, the area of the zones as they formed and matured remained consistent over time (Fig. 2C), where the formation of Zone 2 appeared to define the concentric space in which cells of all zones would continue developing through Day 28. Given minimal changes in overall size at Day 7 compared to Day 28, we propose that once differentiation is initiated and the primitive Zone 1 and presumptive Zone 2 are established by Day 7, subsequent development of ocular cell types comprising Zones 2 and 3 are specified based on internal reciprocal cell–cell interactions (Fig. 2D).
Spatiotemporal expression of ocular markers highlights developing cell types
Next, the ability of the SEAM cultures to generate various cell types found within the developing human eye was tested using PAX6 (pan-ocular marker), TP63 (ocular surface ectoderm), VSX2 (retinal progenitor cells, RPC), MITF (retinal pigmented epithelium, RPE), and CRYAA/B (lens), similar to as described earlier5,6, but extending these studies to Day 14 (Fig. 3). As expected, PAX6 was expressed in cells within Zones 1, 2 and 3 at both timepoints, while TP63 was expressed in cells comprising Zones 3 and 4 (Fig. 3A and E). PAX6/TP63 double-positive cells are indicative of Zone 3 specifically and represent corneal cell types arising from the surface ectoderm. Notably, while all zones are present at both timepoints, they are visibly less developed at Day 14 (Fig. 3A) compared to Day 28 (Fig. 3E). Zone 2 has been previously shown to be analogous to the developing optic cup, comprising the neuroretina and RPE5,6. At Day 14, both VSX2-positive cells (Fig. 3B) and MITF-positive cells (Fig. 3C) were found diffusely throughout the presumptive Zone 2. In contrast, at Day 28, VSX2 expression was restricted to the inner margin of Zone 2 (Fig. 3F), while MITF-positive cells (Fig. 3G) were distinctly observed in the outer margin, a tissue organization that recapitulates human optic cup development. In vertebrates, the lens development is induced through the interaction between the optic cup and the surface ectoderm, represented in the SEAM cultures at Day 14 by Zones 2 and 3, respectively. Concurrently, CRYAA/B-positive lens cells were first observed at Day 14 forming small clusters at the presumptive margin of Zones 2 and 3 (Fig. 3D), and further expanded through Day 28 (Fig. 3H).

Spatiotemporal expression of ocular markers highlights developing cell types. (a–h) Markers indicative of ocular cell type-specific differentiation (PAX6, TP63, VSX2, MITF and CRYAA/B) were selected for immunostaining at both Day 14 (a–d) and Day 28 (e–h). (a’–h’) Brightfield images show presumptive zones at Day 14, and fully formed zones at Day 28. n = 12 SEAMs. Scale bar, 200 μm.
SEAM development is regulated by specific factors
Total RNA sequencing was utilized to elucidate gene expression changes accompanying SEAM formation. Transcriptomic comparisons were carried out at Day 14 and 28 relative to undifferentiated human iPSCs (Day 0). Overall, 7593 differentially expressed (DE) transcripts were detected, and grouped based on temporal patterning of Z-scores using K-mean clustering, thus identifying 7 distinct trends (T) characterized by peak expression of corresponding DE genes (DEGs) (Fig. 4A; Supplementary Table 1). T1-T4 were associated with upregulation of gene expression over time. T1 included 510 (6.7%) DEGs with peak expression at Day 14, T2 comprised 1441 (19.0%) DEGs upregulated at both timepoints, T3 included 1232 (16.2%) DEGs with expression gradually peaking by Day 28, while T4 included 567 (7.5%) DEGs with upregulation specifically at Day 28. In contrast, T5 (344 DEGs; 4.5%) and T6 (3276 DEGs; 43.1%) were associated with downregulation of gene expression over time. Lastly, T7 included a subset of genes (223 DEGs, 2.9%) expressed in undifferentiated iPSCs, but then markedly downregulated at Day 14, followed by upregulation at Day 28.

SEAM development is regulated by specific factors. (a) Temporally clustered DEGs reveal 7 distinct gene expression trends corresponding to the intermediate (Day 14) and late (Day 28) stages of ocular differentiation in comparison to undifferentiated human iPSCs (Day 0). Each row represents a single gene. Each column represents one of two biological replicates for each time point. Red and blue indicate standard deviation above and below the mean, respectively. (b, c) Overrepresentation analysis of temporally clustered DEGs identified significantly enriched (FDR < 0.05) biological processes (b) and canonical pathways (c) associated with each trend. Dot size represents the percentage of cells associated with each term. Color scale represents adjusted p values. (d) Heatmap showing the top 10 significant DE transcription factor-related genes in each trend (adjusted p-value < 0.05, LFC ≥ 1.2). Each row represents a single gene. Each column represents one of two biological replicates for each time point. Red and blue indicate standard deviation above and below the mean, respectively. KE, KEGG pathway50; PID, Pathway Interaction Database; REA, Reactome; WP, WikiPathways.
Analysis for enriched biological processes (Fig. 4B) and canonical pathways (Fig. 4C) associated with DEGs in each trend identified unique and shared biological events underlying the progression of SEAM formation (see Supplemental Data for details). Briefly, ‘eye/visual system development’, ‘nervous system development’ and pathways driving ECM organization were significant terms shared across all trends displaying upregulation of gene expression over time. Notably, T1 DEGs were uniquely enriched for ‘Neural crest cell development’, ‘neural retina development’ and MAPK, IGF and TGFβ signaling pathways, likely promoting the initial transition from a progenitor state toward an intermediate stage of differentiation, while enrichment of cell type-specific processes were associated with DEGs displaying peak expression by Day 28 (T3 and T4), including ‘lens development’, ‘pigment cell differentiation’, and ‘keratinocyte differentiation’.
To determine potential regulators driving the progression, the transcriptomic data was compared to a published catalog of known human transcription factors8. 599 DEGs encoding transcription factors were identified as likely drivers of gene expression changes across the trends (Fig. 4D, Supplementary Table 2). In respect to trends specifically upregulated over time, POM/neural crest-related factors (ie. TFAP2A, TBX3, DLX3, GATA6) were among the most significantly upregulated DEGs at Day 14 (T1), while eye field transcription factors that define the eye primordium during early development (NR2E1, LHX2, SIX3/6, PAX6, RAX) were enriched in DEGs upregulated at both timepoints (T2), likely driving ocular cell type specification in the context of the SEAM cultures, as well as promoting the initial stages of RPC development2,9. Concurrently, regulators of neuronal development (ASCL1, POU3F3, NEUROD1/4, VSX2) were among the significant DEGs gradually upregulated by Day 28 (T3). HOX gene expression appeared to predominate transcriptional regulators with peak expression at Day 28 (T4), but also featured ATOH7 and NEUROG2, known regulators of RPC specification and retinal ganglion cell (RGC) differentiation10,11,12, as well as EGR1 and Ets factor EHF, both of which are associated with corneal epithelial cell differentiation13,14.
Single cell analyses of SEAM cultures identify emerging ocular cell types
In order to resolve heterogeneity and identify the emergence of various cell types within the SEAM cultures, single cell sequencing was performed at Day 14 (8,983 cells; Figs. 5 and 6) and Day 28 (8,429 cells; Figs. 7 and 8). Annotation was performed based on comparison of differential gene expression profiles of each cluster (Supplementary Table 3) against publicly available scRNA-seq datasets representing various human ocular tissues15,16,17,18,19,20,21,22,23,24,25. Twelve clusters were identified at Day 14 and fourteen at Day 28, including 9 clusters that were shared between the two timepoints, displaying similar gene expression signatures, and those that were unique (3 clusters at Day 14; 5 clusters at Day 28) together accentuating the progression of maturation of cell types within the cultures over time.

Single cell analyses of SEAM cultures at Day 14. (a) UMAP plot visualization of ocular cell types identified 12 transcriptionally distinct clusters. (b) Clusters represent nascent and mature cell populations that correspond to presumptive zones observed in the SEAM cultures at Day 14. (c) Feature plots highlight expression of well-defined markers indicative of cell type-specific differentiation. (d) Concurrent expression of several genes indicative of a ciliary marginal zone (CMZ) cell identity support the presence of a CMZ-like cell population in Clusters 2 (‘Early optic cup’) and 4 (‘Late optic cup’) at Day 14. (e) Heatmap showing the top 15 significant DE transcription factor-related genes in each cluster (adjusted p-value < 0.05, LFC ≥ 1.2). Significant DE transcription factors were not detected for ‘Developing lens’ (Cluster 11). Red and blue indicate standard deviation above and below the mean, respectively.

Cell type-specific differentiation programs at Day 14 are mediated by signaling interactions across clusters. (a, b) Gene Set Enrichment Analysis (GSEA) analysis of cluster-specific DEGs identified significantly enriched (FDR < 0.05) biological processes (a) and canonical pathways (b) associated with each cell population. Dot size represents the percentage of cells associated with each term. Color scale represents adjusted p values. (c) Global ligand-receptor interaction plots infer cell–cell communication networks across clusters that are common between Day 14 and Day 28, or predominantly enriched at Day 14 (VEGF signaling, box). KE, KEGG pathway50; PID, Pathway Interaction Database; REA, Reactome; WP, WikiPathways.

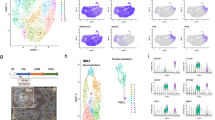
Single cell analyses of SEAM cultures at Day 28. (a) UMAP plot visualization of ocular cell types identified 14 transcriptionally distinct clusters. (b) Groups of clusters represent cell type-specific populations expected within each of the fully developed zones observed at Day 28. (c) Feature plots highlight expression of well-defined markers indicative of cell type-specific differentiation. (d) Concurrent expression of PAX6, TP63, CXCL14 and TXNIP highlight a putative population of limbal epithelial stem cells (LESC) predominantly enriched in Cluster 7 (‘Limbal epithelial stem cells’) at Day 28. (e) Heatmap showing the top 15 significant DE transcription factor-related genes in each cluster (adjusted p-value < 0.05, LFC ≥ 1.2). Red and blue indicate standard deviation above and below the mean, respectively.

Cell type-specific differentiation programs at Day 28 are mediated by signaling interactions across clusters. (a, b) Gene Set Enrichment Analysis (GSEA) analysis of cluster-specific DEGs identified significantly enriched (FDR < 0.05) biological processes (a) and canonical pathways (b) associated with each cell population. Dot size represents the percentage of cells associated with each term. Color scale represents adjusted p values. (c) Global ligand-receptor interaction plots infer cell–cell communication networks across clusters that are common between Day 14 and Day 28, or uniquely enriched at Day 28 (KLK signaling, box). KE, KEGG pathway50; PID, Pathway Interaction Database; REA, Reactome; WP, WikiPathways.
Single cell analyses of Day 14 SEAM cultures
The 12 clusters (Fig. 5A) included ‘Developing anterior segment’ (Cluster 0, 2,264 cells, 25.2%), ‘Neural stem/progenitor cells’ (Cluster 1, 1,611 cells, 17.9%), ‘Early optic cup’ (Cluster 2, 1,177 cells, 13.1%), ‘Early anterior segment’ (Cluster 3, 1,164 cells, 13.0%), ‘Late optic cup’ (Cluster 4, 1,128 cells, 12.6%), ‘Early transit-amplifying cells’ (Cluster 5, 516 cells, 5.7%), ‘Presumptive epithelial progenitors’ (Cluster 6, 350 cells, 3.9%), ‘Proliferating optic cup progenitors’ (Cluster 7, 348 cells, 3.9%), ‘Developing stroma and endothelial cells’ (Cluster 8, 226 cells, 2.5%), ‘Developing retina’ (Cluster 9, 96 cells, 1.1%), ‘Undifferentiated iPSCs’ (Cluster 10, 67 cells, 0.7%), and ‘Developing Lens’ (Cluster 11, 36 cells, 0.4%). Clusters 1, 2, 4, 7 and 9 are likely to reside in Zones 1 and 2, while clusters 0, 3, 5, 6, 8 and 11- in Zones 3 and 4 (Fig. 5B); this is supported by specific expression of PAX6, TP63, VSX2, MITF and CRYAA/B whose protein products were also observed in the corresponding zones at Day 14 (Figs. 3 and 5C and D). Expression of well-established cell type-specific markers were used to distinguish cell populations in each cluster (Supplementary Table 4), which are briefly described below. Extended cluster-specific descriptions can be found in Supplemental Data. In addition, 359 significantly upregulated transcription factors (adjusted p-value < 0.05, LFC ≥ 1.2) were identified across all clusters, highlighting crucial drivers regulating cluster-specific gene expression at this early stage of SEAM differentiation (Fig. 5E; Supplementary Table 5).
Clusters 1 and 9 displayed gene signatures associated with neural stem/progenitor cells and developing neuroretinal cell types, respectively. Cluster 9 featured expression of neuronal (TUBB3, MAP2) and RGC (GAP43, SNCG) markers, but those associated with amacrine, bipolar or horizontal cells were not detected. A range of differentiation states characterized clusters associated with developing optic cup derivatives (‘Proliferating optic cup progenitors’ (Cluster 7), ‘Early optic cup’ (Cluster 2), ‘Late optic cup’ (Cluster 4)) based on differential expression levels of RPC and RPE markers. Clusters 2 and 4 contained retinal stem/progenitor cells displaying a transcriptional signature reminiscent of those comprising the ciliary marginal zone (CMZ; Fig. 5D). Cluster 4 also featured high co-expression of VSX2 and MITF, a mixed RPC/RPE profile unique to Day 14. Clusters 0, 3, 5, 6, 8 and 11 likely represent emerging cell types associated with the developing anterior segment, including cornea and lens. Cluster 5 displayed high expression of PAX6, Keratin 19 (KRT19) and several cell-cycle dependent markers associated with early transit-amplifying cells, a precursor of corneal epithelial cell types. Clusters 3 and 0 featured POM/neural crest cell marker expression and displayed a mixture of anterior segment-related cell types, including corneal epithelial cells, iris, ciliary body, stroma and endothelial cells, where Cluster 0 displayed a more advanced and less proliferative profile, comparatively. Cluster 8 distinctly exhibited significant upregulation of corneal stroma and endothelial markers, as well as keratinocyte progenitor/corneal stromal stem cell (CSSC) markers (ABCG2, PDGFRB/CD140b, NOTCH1). Lastly, ‘Developing lens’ (Cluster 11) featured modest CRYAB expression and low levels of PAX6 and PROX1, however other advanced lens markers were not detected at Day 14.
Enrichment analysis of cluster-specific biological processes (Fig. 6A) and canonical pathways (Fig. 6B), coupled with ligand-receptor interactions between cells in each cluster (Fig. 6C), provided insight into the reciprocal cell–cell interactions mediating both shared and unique biological events underlying cell type-specific differentiation at Day 14. Of note, ‘cilium organization’ and ‘ciliopathies’ were enriched in Clusters 1, 2, 4 and 7, highlighting putative cell types within the SEAMs for further investigating primary cilia function during ocular development in vitro, and how ciliary dysregulation might contribute to ocular disease26. Enrichment of several biological processes related to ‘Neural crest development’ were observed in anterior segment-related Clusters 0 and 3. This is in concordance with gene expression patterning enriched in T1 (Peak at Day 14) from the global transcriptomic analysis (Fig. 3B), and correlates with previously observed data7, supporting an important role for neural crest-related gene expression at this intermediate stage of ocular differentiation. ‘Response to retinoic acid (RA)’ was an enriched term identified in Clusters 4 and 8, correlating with inferred ligand-receptor interactions detected between these same clusters, as well as others (Fig. 6C). These intracellular RA signaling networks mimic those described in vivo during vertebrate eye development as RA is synthesized in the neuroretina, RPE, lens and corneal ectoderm and interacts with RA receptors expressed in the lens, choroid, RPE, and inner layer of the retina, as well as the POM and its derivatives27,28. Finally, VEGF signaling and ‘vasculature development’ were enriched in Cluster 8. Ligand-receptor interaction plots show VEGF ligands secreted by cells across multiple clusters converging with a distinct impact on differentiation of cell types in Cluster 8, likely those associated with the initial stages of pericyte and vasculature development uniquely prevalent at Day 14, as well as ‘Developing lens’ (Cluster 11), potentially contributing to growth, differentiation, and/or survival of lens cells29,30.
Single cell analyses of Day 28 SEAM cultures
14 transcriptomically distinct clusters were identified at Day 28 (Fig. 7A), including ‘Neural stem/progenitor cells’ (Cluster 0, 1,411 cells, 16.7%), ‘Mature retina’ (Cluster 1, 1,331 cells, 15.8%), ‘RPE/melanocytes’ (Cluster 2, 1,090 cells, 12.9%), ‘Late anterior segment’ (Cluster 3, 1,120 cells, 13.3%), ‘Retinal progenitor cells’ (Cluster 4, 992 cells, 11.8%), ‘Early anterior segment’ (Cluster 5, 647 cells, 7.7%), ‘Early optic cup’ (Cluster 6, 586 cells, 7.0%), ‘Limbal epithelial stem cells (LESC; Cluster 7, 438 cells, 5.2%), ‘Stroma and endothelial cells’ (Cluster 8, 261 cells, 3.1%), ‘Mature transit-amplifying cells’ (Cluster 9, 166 cells, 2.0%), ‘Immature neuronal cells’ (Cluster 10, 162 cells, 1.9%), ‘Intermediate anterior segment’ (Cluster 11, 146 cells, 1.7%), ‘Lens’ (Cluster 12, 43 cells, 0.5%), and ‘Undifferentiated iPSCs’ (Cluster 13, 36 cells, 0.4%). Similar to Day 14, clusters were further assigned to zones (Zone 1: Clusters 0, 1, 10; Zone 2: Clusters 2, 4, 6, 10; Zones 3/4: Clusters 3, 5, 7, 8, 9, 11; Fig. 7B) based on the observed correlation between expression of cluster-specific genes and spatial distribution of the previously reported markers5 (Fig. 3), as well as other characteristic markers of cell type-specific differentiation (Fig. 7C and D; Supplementary Table 4). Extended cluster-specific descriptions can be found in Supplemental Data. Additionally, 745 significantly upregulated transcription factors (adjusted p-value < 0.05, LFC ≥ 1.2) were identified as potential regulators of cell type-specific gene expression across the clusters at this later stage of SEAM differentiation (Fig. 7E; Supplementary Table 5).
Clusters representing progressive neuroretinal cell differentiation were identified (‘Neural stem/progenitor cells’ (Cluster 0), ‘Immature neuronal cells’ (Cluster 10), ‘Mature retina’ (Cluster 1), where Cluster 1 featured gene expression associated with advanced neuronal cell types, including RGCs, amacrine, bipolar and horizontal cells. Cells in Cluster 10 comprised both Clusters 1 and 4, featuring lower expression of neuronal and RPC markers, comparatively, and specific upregulation of immature neuronal markers (ASCL1, ELAVL2/4) and ATOH7, a transcriptional regulator of RGC specification11,12. Transcriptionally distinct clusters representing RPCs (Cluster 4; VSX2, RAX, SOX2) and RPE/melanocytes (Cluster 2; MITF, OTX2, RLBP1) were exclusively identified at Day 28, correlating with visual spatial regionalization of these cell types (Fig. 3F and G). Critical developmental intermediates of corneal epithelial cell differentiation were also identified at Day 28, including a uniquely discrete population of LESCs (Cluster 7; PAX6, TP63, CXCL14, TXNIP16,31,32; Fig. 7D), in conjunction with mature TA cells (Cluster 9; RRM2, UBE2C, MIK67, PBK33,34. Anterior segment-related Clusters 3 and 5 displayed expression of POM/neural crest cell markers, and those indicative of mixed corneal epithelial, stromal and endothelial cell types at progressive levels of differentiation similar to that observed at Day 14. In comparison, Cluster 11 featured a distinct, intermediate gene signature characterized by high S100A8 expression coupled with keratin and mucin expression suggestive of basal conjunctival epithelial cells31,35, as well as modest stromal, endothelial, iris and ciliary body gene expression, albeit lacking specific POM/neural crest marker expression. Comparable with Day 14, Cluster 8 exhibited significant upregulation of corneal and limbal stroma (LUM, FBLN1) and endothelial markers (TAGLN/SM22α, ACTA2), including newly apparent vessel endothelial markers (PECAM/CD31, POSTN)16,24, as well as those associated with CSSCs (ABCG2, PDGFRB/CD140b, ALCAM/CD166) and keratocytes (PDK4)36,37. Finally, Cluster 12 featured a robust gene profile corresponding to lens cell identity with high expression of FOXE3, PITX3, PROX1, PAX6 and numerous lens crystallins, highlighting the progression of lens development in the context of the SEAM culture system.
Enriched biological processes (Fig. 8A) and canonical pathways (Fig. 8B) were further identified in each cluster. As expected, similar enrichments as observed at Day 14 were detected in complementary clusters (eg. ‘neuron differentiation’, ‘ciliary organization’ and ‘ciliopathies’, VEGF signaling associated with ‘vasculature development’, ‘keratinization’), while clusters associated with more specialized cell types at Day 28 were uniquely enriched for more advanced biological processes (eg. ‘lens development’, ‘pigment cell differentiation’, ‘keratinocyte differentiation’). These data were complemented by ligand-receptor interactions (Fig. 8C). Of note, kallikrein (KLK) signaling was identified uniquely at Day 28. KLKs have also been shown to modulate keratin gene expression under the influence of calcium, vitamin D3 and RA signaling38. Moreover, KLK overexpression led to suppressed WNT and EGFR signaling, resulting in deficient cornea wound healing in a murine model of diabetic keratopathy, and decreased migration and proliferation of human corneal epithelial cells in vitro39. Thus, the SEAM cultures provide an avenue for further investigating the potential role of KLK signaling in corneal epithelial cell homeostasis and/or keratinocyte differentiation in the developing eye.
Temporal modeling of ocular differentiation in SEAMs using integrated data across timepoints
Both time points were merged to reconstruct differentiation trajectories for the SEAM cultures as they transitioned from Day 14 to 28, generating a new UMAP (Fig. 9A) annotated using the same methods as described above (Supplementary Table 6). Estimations of differentiation were cross-validated using two independent analyses: diffusion pseudotime (Fig. 9B) and RNA velocity (Fig. 9C). Pseudotime analysis revealed that differentiation from Day 14 to 28 is correlated with cell state transitions from least differentiated (blue; Fig. 9B) to most differentiated (yellow; Fig. 9B), branching into at least 2 distinct trajectories: (1) associated with developing neuroretina and optic cup derivatives and (2) associated with formation of cell types within the anterior segment. Accordingly, overlaying RNA velocity vector fields (Fig. 9C) supported segregation of differentiation trajectories initiating from the least developed cell types comprising ‘General proliferating progenitors’ (Cluster 5, 8.0%). In one branch, precursors of neuroretinal/optic cup derivatives (‘Early optic cup’ (Cluster 1, 18.2%)) flows into ‘Neural stem/progenitor cells’ (Cluster 6, 8.0%) and more developed RPCs identified within ‘Late optic cup’ (Cluster 2, 14.6%) likely contributing to the formation of retinal cell types comprising ‘Mature retina’ (Cluster 4, 8.7%), which share gene expression analogous to that observed at Day 28 (Fig. 7A, Cluster 1). In the second branch, corneal progenitors (‘Transit-amplifying cells’ (Cluster 7, 4.0%)) flow toward ‘Early anterior segment’ (Cluster 0, 21.9%) characterized by high POM marker expression and keratin expression indicative of simple non-stratified corneal epithelial cells, in addition to expression of LESC markers similarly observed at Day 28 (Fig. 7A, D, Cluster 7). These nascent cells transition into ‘Intermediate anterior segment’ (Cluster 8, 3.4%), featuring increased gene expression associated with both non-stratified and stratified corneal epithelia and developing stromal/endothelial cells. This cluster partially shares a gene signature with the developing anterior segment derivatives observed at Day 14 (Fig. 5A, Clusters 0 and 8) and the more mature derivatives identified at Day 28 (Fig. 7A, Clusters 3 and 8). However, in this merged analysis, cells representing ‘Late anterior segment’ were further segregated into two clusters characterized by differential transcriptomic signatures enriched for either stratified corneal epithelial cells and lens (Cluster 3, 10.8%), or POM-derived endothelial cells and pericytes (Cluster 9, 2.5%), highlighting some of the more developed cell types in the SEAM cultures that are present predominantly at Day 28.

Temporal modeling of ocular differentiation in SEAMs using integrated data across timepoints. (a) Distribution of cells originally from Day 14 (pink) and Day 28 (light blue) after merging single cell data from both time points to generate a new UMAP describing dynamic changes in cell differentiation potential. (b) Pseudotime analysis of merged data reveals two distinct differentiation trajectories from least differentiated (blue; root cell indicated by red asterisk) to most differentiated (yellow). (c) Merged UMAP features 10 transcriptionally distinct clusters. RNA velocity vectors convey predicted differentiation directionality originating in Cluster 5 (‘General proliferating progenitors’) toward retinal derivatives (Cluster 4, ‘Mature retina’ and Cluster 2, ‘Late optic cup’) and anterior segment derivatives (Cluster 3 ‘Corneal epithelial cells/lens’ and Cluster 9 ‘Endothelial cells/pericytes’).
SEAM cultures show robust expression of disease-associated ocular genes in relevant clusters
Single cell data was further interrogated to identify expression patterns for known genes associated with a broad range of human ocular disorders, such as corneal opacities, anterior segment dysgenesis, cataract, inherited retinal disease (IRD)/optic atrophy, albinism and Microphthalmia/Anophthalmia/Coloboma (MAC) spectrum and albinism (Fig. 10). The observed patterns matched the expected genes’ profiles based on the current knowledge about the associated human ocular features as well as known expression patterns from animal models3,4. Additionally, dynamic changes in level/distribution of the disease-associated genes between Day 14 and 28 correlated with the progressive development of associated embryonic structures during the SEAM differentiation process, emulating expression changes that likely coincide with the early stages of human eye development. Overall, stronger expression was detected at Day 28 with a clear enrichment in the cornea/anterior segment-associated clusters for corneal dystrophy/anterior segment dysgenesis genes; lens cluster- for cataract-linked genes; retinal clusters- for IRD/optic atrophy phenotypes; and RPE- for albinism. MAC genes demonstrated the most complex temporal and spatial pattern, with an equally strong presence at both stages and diverse expression in multiple ocular clusters consistent with the early onset of MAC spectrum and the broad range of affected tissues typically observed.

SEAM cultures show robust expression of disease-associated ocular genes in relevant clusters. Scaled expression of genes associated with several ocular malformations visualized across clusters at Day 14 and Day 28. Dot size represents the percentage of cells associated with each term. Representative feature plots highlight the overall concurrent expression of genes in each category denoting potentially affected cell populations within the SEAM cultures.
Discussion
Human organoids and ocular tissues derived from human iPSCs emerged as promising platforms for disease modeling and mechanistic studies40,41,42, however these models usually represent fully differentiated cell types, rather than the dynamic processes occurring during early stages of eye development. The recently published SEAM culture system5,6 offers a promising model that recapitulates the formation of diverse cell types developing concurrently within the whole embryonic eye. In this study, we present a detailed characterization of SEAM development at several time points along the 28-day differentiation process, with an emphasis on an intermediate (Day 14) and late (Day 28) stage of SEAM formation. This analysis revealed: (1) consistent and quantifiable changes in morphology and zone formation over time, (2) global gene expression trends characterized by enriched biological processes and pathways, and their respective transcriptional drivers together mediating the progression of SEAM development, (3) a full range of specific ocular cell types generated during this process, including rare stem cell-like populations such as those that transcriptomically resemble cells found in the ciliary marginal zone (CMZ), transit-amplifying (TA) cells, limbal epithelial stem cells (LESCs), and corneal stromal stem cells (CSSCs), (4) inferred signaling interactions between clusters facilitating cell type-specific differentiation, (5) an integrated developmental trajectory highlighting intermediate states of ocular differentiation in vitro, and (6) cluster-specific expression patterns of key genes associated with ocular disease. Establishing a temporal morphological and transcriptomic baseline for normal SEAM development through these time points enables future investigations into how disease-causing variants may disrupt the early stages of human eye development leading to complex ocular phenotypes.
The morphological evaluation demonstrates that SEAM formation is consistent and reproducible over time, despite comparative variability in size between individual SEAMs. In addition, immunostaining and single cell analyses at Day 14 and 28 demonstrate changes in marker distribution and transcriptomic profiles accompanying the distinct and progressive regionalization of the early SEAM cultures into primitive neural retina/RPCs and RPE, and gradual formation of lens clusters at the interface between Zones 2 and 3. Lastly, we identified Zone 3 at Day 14 and tracked its dynamic expansion toward Day 28 allowing for interrogation of the progressive development of cell types found in the embryonic anterior segment. Using these benchmarks, it is expected that changes in zone area and/or the transcriptomic composition of the populations generated within can be quantified and compared between normal and disease-state providing insight into both primary and secondary consequences of a gene variant on eye development within the context of the SEAM model.
Bulk RNA-seq analyses identified seven gene expression trends that synergistically promote the transition from a progenitor state (Day 0) toward the intermediate (Day 14) and late (Day 28) stage of differentiation. Global transcriptomic changes correlate with well-described, sequential patterning events that occur during vertebrate eye development in vivo. Uniquely, the presented analyses identify biological processes and pathways for further investigation within the SEAM cultures to clarify their roles during ocular differentiation, including those related to primary cilia organization and function in ocular cell types, and KLK signaling, among others.
Toward discerning heterogeneity in the SEAM cultures at Day 14 and 28, the conducted single cell analyses identified a range of developmental and advanced cell types correlating with those reported in similar studies of human fetal15,16,17,20 and adult15,16,19,21,24 ocular tissues. Shared cell populations highlighted a continuum of differentiation stages accompanying formation of cell types found within the developing retina, RPE, and anterior segment of the eye, supported by pseudotime and RNA velocity analyses. Unique to Day 14 were nascent cell populations transitioning between progenitor and differentiated cell states, including ‘Proliferating optic cup progenitors’, a subsequent mixed RPC/RPE profile denoted as ‘Late optic cup’, and ‘Presumptive epithelial progenitors’. Clusters unique to Day 28 included transcriptomically distinct RPCs and RPE, as well as newly developed transitional cell populations differentiating towards retinal (‘Immature neuronal cells’ (Cluster 10)), corneal (‘LESC’ (Cluster 7)), and anterior segment-related (‘Intermediate anterior segment’ (Cluster 11)) cell fates. Finally, these analyses identified several stem cell-like populations indicative of rare, transitional states difficult to capture in vivo, including populations containing CMZ-like cells, LESCs, TA cells, and CSSCs.
The CMZ is described as a region containing retinal stem/progenitor cells that contributes to neurogenesis in the neural retina, with the competency to generate all retinal cell types43,44. It is well-characterized in non-mammals45,46,47, mammals48, and more recently, single cell transcriptomic and histological analyses of human fetal eyes have identified CMZ-like regions, providing insight into the gene signatures for subpopulations of early and late neurogenic progenitors and their derivatives at varying stages of differentiation17,18,20. The CMZ-like cell populations described in this present study correlate with those defined in these published analyses. In addition, pseudotime and RNA velocity analyses support the differentiation trajectory of the CMZ-like cells identified in ‘Early optic cup’ and their progression toward a mixed RPC/RPE gene signature indicative of the developing optic cup (‘Late optic cup’), and advanced cell types (RGCs, horizontal, bipolar, amacrine cells) of the neural retina (‘Mature retina’). While these combined data support the presence of a CMZ-like population, cell lineage tracing combining cell barcoding with single cell analyses would provide functional validation of the cells and their competency within the context of the SEAM culture system.
A gene signature indicative of LESCs was uniquely identified at Day 28, while those associated with TA cells were found shared between the two time points. In vivo, both LESCs and TA cells reside in the basal corneal epithelium, providing a pool of progenitors supporting the continuous generation of corneal epithelial cells in a sequential manner. LESCs proliferate and give rise to intermediary TA cells, which migrate to the central cornea, proliferate, and differentiate into the various corneal epithelial cell types. LESCs and TA cells are notoriously difficult to isolate and characterize given the heterogeneity of the limbal and peripheral corneal epithelial basal layer, and partially overlapping gene signatures31,33,34. As the understanding of these niche stem cell populations increases, combinations of markers considered to be indicative of putative LESCs or TA cells have been formulated16,31,33,34, and utilized to identify these populations at Day 14 (Early TA cells: Cluster 5) and Day 28 (LESC: Cluster 7; Mature TA cells: Cluster 9). We propose that the absence of a definitive LESC population at Day 14 may be due to unresolved transcriptomic similarities between early TA cells and LESCs, or LESCs and other developing corneal epithelial progenitor populations at this time point. Nevertheless, SEAM cultures provide a platform for further interrogating the mechanisms underlying LESC differentiation, enabling development of more efficient methods for identifying this rare population in patients presenting with limbal stem cell deficiency for autologous ex vivo expansion and transplantation applications.
Finally, a small population of putative CSSCs/keratinocyte progenitors was detected within stroma/endothelial-related clusters at both Day 14 and 28. Serving as a reservoir of progenitor cells, CSSCs contribute to corneal clarity by replenishing damaged or aged corneal keratocytes, which are responsible for maintaining structural integrity of the stromal matrix through continued ECM production and organization49. Characteristic markers of CSSCs include PAX6 and ABCG2, a key keratocyte progenitor and stem cell marker, as well as mesenchymal stem cell markers CD73 (NT5E), CD140b (PDGFRB), CD166 (ALCAM), and PDK436,37, all of which are detected in these clusters, albeit within a larger cell population displaying mixed cell types. Advances in single cell sequencing techniques with higher resolution may enable distinct separation of these stem cells from their derivatives, allowing further investigation into the mechanisms underlying their regenerative capacity and keratocyte differentiation within the context of a system modeling development of the whole eye.
The differential gene expression analyses reported here revealed distinct and dynamic gene signatures indicative of multiple cell types generated over time within the SEAM cultures. Nevertheless, some clusters represented mixed cell populations too transcriptionally similar to separate into distinct cell types (i.e. corneal and stromal cells, and their derivatives such as iris, ciliary body and pericytes) or cell subtypes (i.e. conjunctival, limbal and basal corneal epithelial cells). This may be an inherent consequence of capturing the early stages of eye development characterized by the presence of multiple cell populations at different levels in the differentiation process. Future studies utilizing high-resolution spatial transcriptomics may not only provide clearer segregation of distinct cell types within closely related transcriptomic backgrounds, but also their localization within the SEAM cultures for visualizing changes in developmental maturation between the two time points.
The 2D SEAMs represent an example of the intrinsic capacity for spontaneous development and self-organization of various ocular cell types similar to that observed in organoid cultures, modeling the early stages of embryonic eye development. Accordingly, gene expression signatures of cell types identified within the SEAMs after the 28-day differentiation process had characteristics of developmental intermediates rather than fully matured cell types of the adult eye. Having the ability to capture the intermediate stages of cell type-specific differentiation uniquely enables interrogation of the molecular mechanisms underlying embryonic eye development. Moreover, methods for further differentiation into advanced cell types from the SEAM cultures at Day 28 were described previously5,6, including those for extended corneal epithelial differentiation from cells isolated from Zones 3 and 4, enrichment of RPE cell colonies derived from Zone 2, and encouragement of neural crest cell differentiation by lowering the initial seeding density. These studies demonstrate the stem/progenitor cell capabilities of the cells originally isolated from the SEAMs at Day 28 and emphasizes the developmental potential of cells in vitro for establishing additional protocols to investigate the advanced stages of ocular tissue-specific differentiation within the SEAM culture system in future studies.
This work captures the key in vitro features mediating the progression of SEAM formation over the 28-day differentiation process, serving as a resource for further functional studies elucidating the mechanisms of embryonic eye development and for evaluating the effects of genetic variants associated with human ocular disease. The significance of a model targeting earlier stages of embryonic eye development is accentuated by the high rate of unsolved cases affected by a broad range of developmental ocular disorders, many of which likely due to unique genetic variants/mechanisms that may only be interrogated in a human-based model. We envision these analyses will direct thorough evaluation of SEAMs generated using patient-derived iPSCs enabling clarification and resolution for these types of cases.
Methods
Generation and morphological assessment of SEAM cultures
SEAM cultures were generated according to previously described protocols5,6. SEAM cultures were derived from human iPS cell line 201B7 obtained from the RIKEN Bio Resource Center (Tsukuba, Japan). Human iPS cultures were propagated in StemFit Basic03 medium (Ajinomoto, Tokyo, Japan) on 6-well plates coated with Laminin iMatrix-511 E8 fragment (0.5 μg/cm2; AMSBio) for a minimum of three passages before initiation of differentiation. To induce differentiation of SEAM cultures, human iPSCs were seeded (350 cells/cm2) across LN511E8-coated (0.5 μg/cm2) 6-well plates and cultivated in StemFit Basic03 medium for 7 days prior to changing to differentiation medium. Differentiation medium consisted of GMEM (Life Technologies) supplemented with 10% knockout serum replacement (Life Technologies), 1 mM sodium pyruvate (Sigma), 0.1 mM non-essential amino acids (Life Technologies), 2 mM L-glutamine (Life Technologies), 1% penicillin–streptomycin solution (Life Technologies), and 55 μM 2-mercaptoethanol (Life Technologies). Differentiation medium was exchanged every other day. SEAM cultures were cultivated for 28 days under these differentiation conditions. Brightfield images of live cultures were acquired with a KEYENCE BZ-X710 All-in-one fluorescence microscope (KEYENCE, Osaka, Japan) equipped with a high resolution 2.8-megapixel monochrome CCD camera. To generate a growth profile individual SEAMs were tracked and imaged at several time points using the multipoint function of the KEYENCE imaging software. Measurements were taken using ImageJ software (Version 1.54g; http://www.imagej.org). Statistical analyses and graphical representations for the data presented in Fig. 2 and Supplemental Fig. 1 were computed using GraphPad Prism software (Version 9.3.1 for Windows 64-bit; http://www.graphpad.com).
Immunofluorescence staining
SEAM cultures were fixed at Day 14 and 28 with 4% paraformaldehyde (PFA) for 30 min at room temperature and rinsed three times with PBS prior to immunostaining. Cultures were washed with TBS with 0.3% Triton-X (TBST), blocked in 5% normal donkey serum/TBST for 1 h and incubated with primary antibodies overnight at 4 °C. The following primary antibodies were used at a dilution of 1:200: anti-VSX2 (sc-365519; Santa Cruz Biotechnology), anti-MITF (X2398M; Exalpha Biologicals), anti-PAX6 (PRB-278P; BioLegend), anti-TP63 (sc-25268; Santa Cruz Biotechnology), anti-Crystallin (ADI-SPA-224-F, Enzo). Cultures were incubated with secondary antibodies conjugated to AlexaFluor 488 or AlexaFluor 568 (donkey anti-mouse AF488 #A21202, donkey anti-rabbit AF568 #10042; Life Technologies) at a dilution of 1:500, and counterstained with 30 μM DAPI (Life Technologies). Single-focal-plane images of EYFP and rhodamine signals were acquired with a Zeiss AxioZoom.V16 fluorescence stereomicroscope equipped with an AxioCam 820 Mono camera, a Zeiss ApoTome 3, an ApoZ 1.5 × FWD 30 mm objective, and appropriate filter sets. Fluorescent images were pseudocolored, as indicated in figure legends, and merged using ZEN 3.9 software (Zeiss).
Sample preparation for single cell or total RNA sequencing
In preparation for sequencing, SEAM cultures at Day 0, 14 and 28 were lifted in 0.05% Trypsin-EDTA (Life Technologies) followed by gentle mechanical dissociation in Accumax (STEMCELL Technologies). Dissociated cells were filtered through a 70 μm mesh membrane (Miltenyi Biotec) to achieve a single cell suspension. Magnetically labelled apoptotic and dead cells were removed using a Dead Cell Removal kit (Miltenyi Biotec) and MACS separation column. Cell concentration and viability were determined using a CellDrop FL Automated Cell Counter (DeNovix) prior to proceeding with either the 10X Genomics single cell protocol or total RNA extraction.
Approximately 10,000 cells were used to generate a single cell library at each time point using the Chromium Next GEM Single Cell 3’ Reagent kit (v3.1, Dual Index; 10X Genomics), following the manufacturer’s protocol. Single cell libraries were quantified using an Agilent TapeStation System (Agilent Technologies) prior to being submitted to Psomagen, Inc. (Rockville, MD) for sequencing (Illumina NovaSeq X Plus 10B (2 × 150 bp)). Total RNA was extracted from the remaining cell suspension using a Direct-Zol RNA Miniprep Kit (Zymo, Irvine, CA), which included a DNase treatment step. Total RNA from two biological replicates from each time point were sent to Psomagen, Inc. (Rockville, MD) for library preparation and sequencing. Only samples with an RNA integrity number (RIN) ≥ 7 were considered for cDNA library preparation. cDNA libraries were generated using TruSeq Stranded Total RNA Gold Sample Prep Kits, (Illumina), which included an rRNA suppression step. cDNA libraries were indexed for multiplexing and sequenced on the Illumina NovaSeq X Plus platform (2 × 150bp; 50M read depth). Detailed methods for processing of the bulk and single cell sequencing data are described in Supplemental Methods.
Data availability
The experimental data that support the findings of this study are available in Supplemental Materials. Bulk and single cell RNA sequencing data has been deposited in the Gene Expression Omnibus (GEO) under the accession code GSE290847.
References
Adler, R. & Canto-Soler, M. V. Molecular mechanisms of optic vesicle development: complexities, ambiguities and controversies. Dev. Biol. 305, 1–13. https://doi.org/10.1016/j.ydbio.2007.01.045 (2007).
Diacou, R. et al. Cell fate decisions, transcription factors and signaling during early retinal development. Prog Retin Eye Res 91, 101093. https://doi.org/10.1016/j.preteyeres.2022.101093 (2022).
Reis, L. M. & Semina, E. V. Conserved genetic pathways associated with microphthalmia, anophthalmia, and coloboma. Birth Defects Res. C Embryo Today 105, 96–113. https://doi.org/10.1002/bdrc.21097 (2015).
Reis, L. M., Seese, S. E., Costakos, D. & Semina, E. V. Congenital anterior segment ocular disorders: Genotype-phenotype correlations and emerging novel mechanisms. Prog Retin Eye Res 102, 101288. https://doi.org/10.1016/j.preteyeres.2024.101288 (2024).
Hayashi, R. et al. Co-ordinated ocular development from human iPS cells and recovery of corneal function. Nature 531, 376–380. https://doi.org/10.1038/nature17000 (2016).
Hayashi, R. et al. Coordinated generation of multiple ocular-like cell lineages and fabrication of functional corneal epithelial cell sheets from human iPS cells. Nat. Protoc. 12, 683–696. https://doi.org/10.1038/nprot.2017.007 (2017).
Howard, L. et al. Single-cell transcriptomics reveals the molecular basis of human iPS cell differentiation into ectodermal ocular lineages. Commun. Biol. 7, 1495. https://doi.org/10.1038/s42003-024-07130-4 (2024).
Lambert, S. A. et al. The human transcription factors. Cell 172, 650–665. https://doi.org/10.1016/j.cell.2018.01.029 (2018).
Zuber, M. E., Gestri, G., Viczian, A. S., Barsacchi, G. & Harris, W. A. Specification of the vertebrate eye by a network of eye field transcription factors. Development 130, 5155–5167. https://doi.org/10.1242/dev.00723 (2003).
Ma, W. & Wang, S. Z. The final fates of neurogenin2-expressing cells include all major neuron types in the mouse retina. Mol. Cell. Neurosci. 31, 463–469. https://doi.org/10.1016/j.mcn.2005.10.018 (2006).
Feng, L. et al. MATH5 controls the acquisition of multiple retinal cell fates. Mol. Brain 3, 36. https://doi.org/10.1186/1756-6606-3-36 (2010).
Brzezinski, J. A. T., Prasov, L. & Glaser, T. Math5 defines the ganglion cell competence state in a subpopulation of retinal progenitor cells exiting the cell cycle. Dev. Biol. 365, 395–413. https://doi.org/10.1016/j.ydbio.2012.03.006 (2012).
Hsueh, Y. J., Kuo, P. C. & Chen, J. K. Transcriptional regulators of the DeltaNp63: their role in limbal epithelial cell proliferation. J. Cell. Physiol. 228, 536–546. https://doi.org/10.1002/jcp.24160 (2013).
Stephens, D. N. et al. The Ets transcription factor EHF as a regulator of cornea epithelial cell identity. J. Biol. Chem. 288, 34304–34324. https://doi.org/10.1074/jbc.M113.504399 (2013).
Collin, J. et al. Single-cell RNA sequencing reveals transcriptional changes of human choroidal and retinal pigment epithelium cells during fetal development, in healthy adult and intermediate age-related macular degeneration. Hum. Mol. Genet. 32, 1698–1710. https://doi.org/10.1093/hmg/ddad007 (2023).
Collin, J. et al. A single cell atlas of human cornea that defines its development, limbal progenitor cells and their interactions with the immune cells. Ocul. Surf. 21, 279–298. https://doi.org/10.1016/j.jtos.2021.03.010 (2021).
Dorgau, B. et al. Single-cell analyses reveal transient retinal progenitor cells in the ciliary margin of developing human retina. Nat. Commun. 15, 3567. https://doi.org/10.1038/s41467-024-47933-x (2024).
Eldred, K. C. et al. Ciliary marginal zone of the developing human retina maintains retinal progenitor cells until late gestational stages. bioRxiv (2024). https://doi.org/10.1101/2024.09.03.611053.
Gautam, P. et al. Multi-species single-cell transcriptomic analysis of ocular compartment regulons. Nat. Commun. 12, 5675. https://doi.org/10.1038/s41467-021-25968-8 (2021).
Hu, Y. et al. Dissecting the transcriptome landscape of the human fetal neural retina and retinal pigment epithelium by single-cell RNA-seq analysis. PLoS Biol. 17, e3000365. https://doi.org/10.1371/journal.pbio.3000365 (2019).
Menon, M. et al. Single-cell transcriptomic atlas of the human retina identifies cell types associated with age-related macular degeneration. Nat Commun 10, 4902. https://doi.org/10.1038/s41467-019-12780-8 (2019).
Nguyen, Q. H. et al. Single-cell RNA-seq of human induced pluripotent stem cells reveals cellular heterogeneity and cell state transitions between subpopulations. Genome Res. 28, 1053–1066. https://doi.org/10.1101/gr.223925.117 (2018).
van Zyl, T. et al. Cell atlas of aqueous humor outflow pathways in eyes of humans and four model species provides insight into glaucoma pathogenesis. Proc. Natl. Acad. Sci. U. S. A. 117, 10339–10349. https://doi.org/10.1073/pnas.2001250117 (2020).
van Zyl, T. et al. Cell atlas of the human ocular anterior segment: Tissue-specific and shared cell types. Proc. Natl. Acad. Sci. U. S. A. 119, e2200914119. https://doi.org/10.1073/pnas.2200914119 (2022).
Lukowski, S. W. et al. A single-cell transcriptome atlas of the adult human retina. EMBO J. 38, e100811. https://doi.org/10.15252/embj.2018100811 (2019).
Chandra, B., Tung, M. L., Hsu, Y., Scheetz, T. & Sheffield, V. C. Retinal ciliopathies through the lens of Bardet-Biedl Syndrome: Past, present and future. Prog Retin Eye Res 89, 101035. https://doi.org/10.1016/j.preteyeres.2021.101035 (2022).
Cvekl, A. & Wang, W. L. Retinoic acid signaling in mammalian eye development. Exp. Eye Res. 89, 280–291. https://doi.org/10.1016/j.exer.2009.04.012 (2009).
Mori, M., Ghyselinck, N. B., Chambon, P. & Mark, M. Systematic immunolocalization of retinoid receptors in developing and adult mouse eyes. Invest. Ophthalmol. Vis. Sci. 42, 1312–1318 (2001).
Garcia, C. M. et al. The function of VEGF-A in lens development: Formation of the hyaloid capillary network and protection against transient nuclear cataracts. Exp. Eye Res. 88, 270–276. https://doi.org/10.1016/j.exer.2008.07.017 (2009).
Shui, Y. B. et al. Vascular endothelial growth factor expression and signaling in the lens. Invest. Ophthalmol. Vis. Sci. 44, 3911–3919. https://doi.org/10.1167/iovs.02-1226 (2003).
Li, D. Q. et al. Single-cell transcriptomics identifies limbal stem cell population and cell types mapping its differentiation trajectory in limbal basal epithelium of human cornea. Ocul. Surf. 20, 20–32. https://doi.org/10.1016/j.jtos.2020.12.004 (2021).
Ojeda, A. F., Munjaal, R. P. & Lwigale, P. Y. Expression of CXCL12 and CXCL14 during eye development in chick and mouse. Gene Expr. Patterns 13, 303–310. https://doi.org/10.1016/j.gep.2013.05.006 (2013).
Li, J. M. et al. Single-cell transcriptomics identifies a unique entity and signature markers of transit-amplifying cells in human corneal limbus. Invest. Ophthalmol. Vis. Sci. 62, 36. https://doi.org/10.1167/iovs.62.9.36 (2021).
Kaplan, N. et al. Single-cell RNA transcriptome helps define the limbal/corneal epithelial stem/early transit amplifying cells and how autophagy affects this population. Invest. Ophthalmol. Vis. Sci. 60, 3570–3583. https://doi.org/10.1167/iovs.19-27656 (2019).
Merjava, S., Neuwirth, A., Tanzerova, M. & Jirsova, K. The spectrum of cytokeratins expressed in the adult human cornea, limbus and perilimbal conjunctiva. Histol. Histopathol. 26, 323–331. https://doi.org/10.14670/HH-26.323 (2011).
Du, Y., Funderburgh, M. L., Mann, M. M., SundarRaj, N. & Funderburgh, J. L. Multipotent stem cells in human corneal stroma. Stem Cells 23, 1266–1275. https://doi.org/10.1634/stemcells.2004-0256 (2005).
Funderburgh, M. L., Du, Y., Mann, M. M., SundarRaj, N. & Funderburgh, J. L. PAX6 expression identifies progenitor cells for corneal keratocytes. FASEB J. 19, 1371–1373. https://doi.org/10.1096/fj.04-2770fje (2005).
Morizane, S., Yamasaki, K., Kabigting, F. D. & Gallo, R. L. Kallikrein expression and cathelicidin processing are independently controlled in keratinocytes by calcium, vitamin D(3), and retinoic acid. J. Invest. Dermatol. 130, 1297–1306. https://doi.org/10.1038/jid.2009.435 (2010).
Liang, W. et al. Pathogenic role of diabetes-induced overexpression of Kallistatin in corneal wound healing deficiency through inhibition of canonical Wnt signaling. Diabetes 71, 747–761. https://doi.org/10.2337/db21-0740 (2022).
Fligor, C. M. et al. Three-dimensional retinal organoids facilitate the investigation of retinal ganglion cell development, organization and neurite outgrowth from human pluripotent stem cells. Sci. Rep. 8, 14520. https://doi.org/10.1038/s41598-018-32871-8 (2018).
Foster, J. W. et al. Cornea organoids from human induced pluripotent stem cells. Sci. Rep. 7, 41286. https://doi.org/10.1038/srep41286 (2017).
Worsdorfer, P. et al. Generation of complex human organoid models including vascular networks by incorporation of mesodermal progenitor cells. Sci. Rep. 9, 15663. https://doi.org/10.1038/s41598-019-52204-7 (2019).
Fischer, A. J., Bosse, J. L. & El-Hodiri, H. M. The ciliary marginal zone (CMZ) in development and regeneration of the vertebrate eye. Exp. Eye Res. 116, 199–204. https://doi.org/10.1016/j.exer.2013.08.018 (2013).
Moshiri, A., McGuire, C. R. & Reh, T. A. Sonic hedgehog regulates proliferation of the retinal ciliary marginal zone in posthatch chicks. Dev. Dyn. 233, 66–75. https://doi.org/10.1002/dvdy.20299 (2005).
Fischer, A. J. & Reh, T. A. Identification of a proliferating marginal zone of retinal progenitors in postnatal chickens. Dev. Biol. 220, 197–210. https://doi.org/10.1006/dbio.2000.9640 (2000).
Link, B. A. & Darland, T. Genetic analysis of initial and ongoing retinogenesis in the zebrafish: comparing the central neuroepithelium and marginal zone. Prog. Brain Res. 131, 565–577. https://doi.org/10.1016/s0079-6123(01)31044-0 (2001).
Perron, M., Kanekar, S., Vetter, M. L. & Harris, W. A. The genetic sequence of retinal development in the ciliary margin of the Xenopus eye. Dev. Biol. 199, 185–200. https://doi.org/10.1006/dbio.1998.8939 (1998).
Marcucci, F. et al. The ciliary margin zone of the mammalian retina generates retinal ganglion cells. Cell. Rep. 17, 3153–3164. https://doi.org/10.1016/j.celrep.2016.11.016 (2016).
Pinnamaneni, N. & Funderburgh, J. L. Concise review: Stem cells in the corneal stroma. Stem Cells 30, 1059–1063. https://doi.org/10.1002/stem.1100 (2012).
Kanehisa, M. & Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. https://doi.org/10.1093/nar/28.1.27 (2000).
Acknowledgements
This work was funded by the National Institutes of Health (R01 EY015518, R01 EY034398 and R21 EY035121 to EVS).
Author information
Authors and Affiliations
Contributions
Maria R Replogle: conceptualization, investigation, data collection, data analysis, data interpretation, writing—original draft; Xiaolong Ma: computer data analysis, statistical analysis, writing- review and editing; Chien-Wei Lin: computer data analysis, statistical analysis, supervision, writing- original draft; Elena V Semina: conceptualization, supervision, funding acquisition, data interpretation, writing—original draft.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Replogle, M.R., Ma, X., Lin, CW. et al. Examination of an iPSC model of human eye development reveals progressive emergence of critical embryonic cell types. Sci Rep 15, 23009 (2025). https://doi.org/10.1038/s41598-025-06602-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-06602-9
