
Abstract
Early exposure to environmental contaminants such as bisphenol A (BPA) poses significant health risks due to their potential to disrupt development. However, many studies focus on immediate effects, leaving the long-term impact poorly understood. Here, we investigate the long-term effects of transient BPA exposure during critical periods of visual system development in zebrafish (Danio rerio). We exposed zebrafish to BPA at 72 hours post-fertilization (hpf) and 7 days post-fertilization (dpf) and assessed retinal anatomy, optomotor responses, retinal physiology, and molecular signaling pathways during juvenile or adult stages. Photopic ERGs from adults revealed changes to bipolar and photoreceptor cell responses that were differentially affected by exposure age and BPA concentration. Exposure to low (0.001 µM) BPA at 72 hpf, not 7 dpf, altered expression of estrogen-responsive genes cyp19a1b and esr2 in adult retinas, as well as the thyroid-responsive gene thrb. We also identified changes in phosphorylation levels of the intracellular kinases Akt, ERK, and JNK in adult retinas and found differences depended upon BPA concentration and exposure age. This study demonstrates that brief early life exposure to BPA can have profound and lasting effects on the visual system, identifying a novel effect of this well-known chemical.
Similar content being viewed by others


Introduction
Estradiol (E2), the biologically relevant estrogen, has many more properties than simply the determination and control of reproductive functions. It is now recognized that precise estrogen signaling is crucial for proper central nervous system development, maturation, and function1,2. While estradiol is known to have pleiotropic actions in the brain, its effects also extend to the retina3. As a result, any interference with the precise and highly regulated estrogen signaling mechanisms, particularly during development, can lead to deleterious consequences on the visual system.
Exposure to endocrine disrupting compounds (EDCs) is one way estrogenic signaling can be altered as most EDCs have estrogenic or anti-estrogenic activity4. EDCs are natural or synthetic chemicals ubiquitously found in many everyday products such as plastic bottles and containers, cosmetics, detergents, food, toys, and pesticides4. These compounds are known to have significant adverse effects on human health as exposure, particularly developmental exposure, has been associated with cognitive deficits, neurological disorders, reproductive abnormalities, metabolic diseases/disorders, obesity, and diabetes5. Additionally, these compounds pose great ecological risks as they commonly enter aquatic ecosystems where they persist in the environment and bioaccumulate in aquatic organisms4.
Bisphenol A (BPA) is a familiar and widespread EDC used as a building block for plastic polycarbonate and epoxy resins. BPA is commonly found in plastic water bottles and food containers, paints, adhesives, dental sealants, and household products6. Consequently, human exposure is frequent, resulting in detectable levels of BPA in ~90% of the human population7. Further, BPA is lipophilic8 and able to cross both the blood brain barrier9 and the placenta, the latter resulting in measurable levels in fetal blood samples10,11. BPA disrupts estrogen signaling and/or causes physiological effects at very low (nM) doses12 that correspond to concentrations found in aquatic environments and wildlife13. Its harmful effects have raised considerable public health concerns, resulting in the US Food and Drug Administration, European Commission, and Health Canada banning its use in baby bottles6. Additionally, many countries have also banned the use of BPA in food contact materials14, leading to the development of related bisphenols (i.e., BPS, BPF)15,16.
BPA is classically described as a weak estrogen receptor (ER) agonist17,18, binding to all ERs19,20, including zebrafish ERs21,22,23, and initiating estrogen signaling pathways14. In this way, BPA administration upregulates expression of aromatase (estrogen synthase), ERα, and ERβ in brain. BPA is also known to bind to and antagonize thyroid hormone receptors (TRs) by preventing triiodothyronine (T3) binding and subsequent ligand-dependent transcription of TRs24. Indeed, zebrafish larvae chronically administered BPA for the first 8 days postfertilization (dpf) showed decreased T4 and rT3 levels25. In addition, BPA can directly alter neuronal physiology by changing voltage-gated26 and/or ligand-gated27,28,29 ion channel activity. These physiological effects are concentration-dependent, with low BPA concentrations often reported to be most effective27,29.
There is a body of literature highlighting the effects of BPA on brain development and neuronal function, and many developmental and behavioral deficits from epidemiological reports are suggested to be associated with effects on neurogenesis30,31. For example, ingested BPA alters behavior in rodents32,33 and zebrafish larvae embryonically exposed to BPA are hyperactive34,35 with multiple adverse neurobiological effects. Transient developmental exposure to BPA alters learning, memory36 and aggression35 in adult zebrafish.
In contrast, what effects BPA poses on the visual system have received less attention. Reduced eye size, decreased expression of genes involved in visual perception37, reduced retinal layer thickness and decreased responses to white and red light25 are reported in zebrafish larvae after BPA exposure. Adult male zebrafish exposed to low dose (0.22-2 nM) BPA for 7-weeks display differences in color preference38. Zebrafish exposed to BPS, a BPA analog, for the first 5 dpf showed altered retinal structure characterized by increased empty areas and a more sparse retinal ganglion cell layer39 and BPS exposure from 2-5 dpf reduced responses to L- and S-type cone signals which was correlated with changes in morphology of red and UV cones40. Extended BPS exposure to 120 days decreased tracking ability and retinal ganglion cell layer thickness and caused an irregular arrangement of retinal photoreceptor cells41. These studies reveal immediate deleterious effects of chronic developmental exposure to bisphenols.
Here, we examine persistent effects of short-term (24 hour, hr) developmental BPA exposure on the visual system in zebrafish (Danio rerio). We focus specifically on the zebrafish retina, which has similar organization, cell types, and synaptic contacts as other vertebrates. We extend our previous work, which identified changes in vision-based optomotor responses in larval zebrafish 1-week and 2-weeks after developmental exposure to BPA42, to ask if transient, developmental exposure to BPA causes persistent effects in retina that can be measured in adulthood (3-4 months postexposure). Our short (24 hr) exposure occurred at 72 hours postfertilization (hpf) and 7 days postfertilization (dpf) as these ages (1) represent critical periods in zebrafish visual system development43,44,45 and (2) correspond with development of estrogen signaling pathways46,47. At each timepoint, visually guided optomotor responses and retinal anatomy were assessed; photopic electroretinograms and molecular analysis of retinal tissue were also performed on adults developmentally exposed to BPA. We hypothesized that exposures to environmentally relevant concentrations of BPA during critical periods in visual system development would directly affect the retina and be correlated with altered visual behavior and physiology. Our data reveal persistent age- and concentration-dependent effects of BPA exposure on the developing visual system. To our knowledge, this is the first study to report that a transient exposure to environmentally relevant BPA concentrations can adversely affect adult retinal function.
Results
We utilized a 24 hr exposure paradigm (Fig. 1) based on the rapid uptake of BPA across the gills48, with measurable internal concentrations within hours and peak values after 24 hr49. To assess the efficacy of a 24 hr BPA exposure, we (1) recorded electroretinograms (ERGs) in adult retinal eye cups and (2) examined changes in gene expression from the same retinal tissue. BPA can alter nerve cell activity by inhibiting Ca+2 currents, increasing K+ currents26, potentiating glycine currents27, increasing Ca+2 influx through NMDA receptors28, or altering GABAA responses29. ERGs, which are used to clinically diagnose retinal diseases, capture the responses of photoreceptors (a-wave), ON-bipolar cells (b-wave), and OFF-bipolar cells (d-wave) (Fig. 2A). In other tissues, BPA is reported to increase expression of the estrogen-responsive genes cyp19a1b (Aromatase B, neural aromatase), esr1 (ERα), and esr2 (ERβ) and decrease expression of thyroid receptors TRa (thraa, thrab) and TRb (thrb).

To assess long-term effects of developmental exposure to BPA, zebrafish larvae, aged either 72 hours postfertilization (hpf) or 7 days postfertilization (dpf) were exposed to 0.1 µM BPA, 0.001 µM BPA, DMSO (vehicle control, 0.0003%), or water for 24 hours (hr). After exposure, larvae were transferred to system water and raised in control conditions until either 1- or 2-weeks post exposure, 1-month (4-weeks) postexposure, or 3–4 months of age. At the 1- and 2-week time points, retinal anatomy was assessed to build on our previous data42. At the 1-month time point, behaviors were assessed by measuring optomotor responses. At the 3–4 month time point, optomotor responses were also recorded, followed by electroretinogram (ERGs) recordings from excised retinal eyecups. Retinal tissue was then collected for molecular analysis and carcasses were processed to quantify BPA levels in body tissues. For comparison, acute effects of BPA exposure were assessed in a separate set of adult fish. These fish were exposed to one of the four treatment groups for 24 hr, when ERGs were recorded, and tissue was collected.

A–G Mean photopic electroretinogram (ERG) recordings from retinal eyecup preparations from adults exposed to water, vehicle (0.0003% DMSO), low BPA (0.001 µM), or high BPA (0.1 µM) for 24 hr. The tissue was stimulated by 300 msec full-field white light pulses at 7 irradiance levels ranging from ND 3.0 (brightest) - 6.0 (dimmest) in 0.5 increments. Traces shown in A represent the mean response to the brightest stimulus (3.0 ND) from all eye cups in each treatment group. The red square pulse denotes duration of the light stimulus. Water = 7 eyes; DMSO = 6 eyes; low BPA = 7 eyes; high BPA = 3 eyes. ERG a-wave, ON-bipolar cell b-wave, and OFF-bipolar d-wave components (noted on vehicle trace in (A)) were quantified. B a-wave amplitude and C a-wave trough time (p = 0.08) were not altered by BPA exposure. D b-wave amplitude was reduced following low BPA treatment (p < 0.001) though (E) time to b-wave peak was not changed (p = 0.162). Low BPA decreased d-wave amplitude (F) but increased (delayed) peak time (G) (p < 0.001). Data in B–G are presented as box and whisker plots: median line is in the center of the box and the ‘x’ is the mean. Significant differences are indicated with asterisks (α = 0.05).
Adults acutely exposed to BPA show tissue accumulation, altered retinal physiology, and changes in gene expression
We observed no mortality in adult zebrafish resulting from a 24 hr exposure to either 0.1 µM (high) BPA, 0.001 µM (low) BPA, or 0.0003% DMSO (vehicle). Analysis of ERG recordings (Fig. 2) identified decreased (p < 0.001) ON-bipolar cell b-wave amplitudes in response to low BPA (17 ± 1.84 µV) compared to high BPA (29.2 ± 1.98 µV), vehicle (33.4 ± 5.32 µV) and water (35.6 ± 3.34 µV) controls (Fig. 2D). OFF-bipolar cell d-wave amplitude was also reduced as a result of low BPA exposure (7.6 ± 0.67 µV) compared to vehicle (13.3 ± 1.07 µV) and water (21.4 ± 2.49 µV) controls (p < 0.001) (Fig. 2F); low BPA also increased (p < 0.001) d-wave peak time (Fig. 2G; 134 ± 8.52 ms) compared to all other groups (water: 92.9 ± 8.45 ms | vehicle: 94.3 ± 5.31 ms | high BPA: 82.4 ± 8.83 ms). No changes between treatment and control groups for a-wave trough time (p = 0.08; Fig. 2C) or b-wave peak time (p = 0.162; Fig. 2E) were noted, though a-wave amplitude was found to be different between water and vehicle controls (p = 0.02; Fig. 2B). Because of the observed difference between vehicle and water controls, suggesting an effect of vehicle50,51,52,53,54,55, we discuss all treatment effects relative to vehicle controls. However, figures and figure captions indicate all significant differences to provide a complete picture of BPA’s effects.
We also examined changes in expression of estrogen- and thyroid-responsive genes in retinal homogenates after a 24 hr exposure using qPCR (Fig. 3A). Our data identified an average 30-fold increase in cyp19a1b expression in the low BPA treatment group, though high variation in fold changes (between approximately 9 and 50) was observed and the increase was not statistically significant. We also observed concentration-dependent uptake of BPA in adult carcasses after a 24 hr exposure (Fig. 3B). Together, these results indicate that BPA exposure for 24 hr is sufficient for tissue uptake and is associated with physiological and genomic effects in retinal tissue. The changes in ERG b- and d-waves further suggest that retinal bipolar cells may be the cell type most sensitive to BPA exposure.

Adult zebrafish were exposed to low BPA (0.001 µM), high BPA (0.1 µM), water, or vehicle (DMSO, 0.0003%) for 24 hr. A qPCR analysis of retinal homogenates tested if the 24 hr exposure regime altered expression of estrogen responsive (cyp19a1b, esr1, esr2a) and thyroid responsive (thrab, thrb) genes. Low BPA increased thrab expression compared to vehicle (p ≤ 0.01). White bars—water, light gray bars—vehicle, dark gray bars—low BPA, black bars—high BPA. B After removal of internal organs, carcasses were flash frozen for LC/MS analysis. Concentration dependent accumulation was evident, and no BPA was detected in either water or vehicle exposed carcasses. C Adults exposed to high BPA at either 72 hpf (light gray bars) or 7 dpf (dark gray bars) had measurable levels of BPA within their carcasses. BPA was not detected in adults developmentally exposed to low BPA at either age. No BPA was found in carcasses of adults that were developmentally exposed to either vehicle or water.
Modest and time-dependent effects of BPA on retinal anatomy and OMRs
Our previous study identified differences in visually guided optomotor responses (OMRs) and eye diameter in zebrafish larvae 1- and 2-weeks post exposure42. Here, we expand that work and build on the acute effects observed after a 24 hr exposure to (1) explore additional visual system consequences of developmental BPA exposure, (2) assess if effects persist after an even greater postexposure interval (i.e., into adulthood), and (3) explore potential cellular mechanisms targeted by BPA (Fig. 1).
To begin, we asked if the previously identified BPA-induced differences in OMRs 1-week and 2-weeks postexposure were associated with anatomical changes in retina. Measurements of H & E-stained retinal sections (Supplementary Fig. 1) did not reveal any effects of BPA on retinal anatomy 1- or 2-weeks postexposure in comparison with vehicle controls (p > 0.05 for all; Supplementary Fig. 2).
When the postexposure assessment was extended to 1-month (4-weeks) fish exposed to low BPA at 7 dpf displayed a decrease in INL thickness compared to vehicle controls (p = 0.001; N = 3 per treatment) (Fig. 4A). Low BPA exposure at 72 hpf also reduced thickness of individual retinal layers, though these reductions were not significant, and no further differences were evident across treatments or exposure ages. We also observed no significant differences in OMRs between vehicle control and BPA treated fish at the 4-week postexposure time point (Supplementary Fig. 3A) and differences in retinal layer measurements were significant only for the reduced INL measurement in larvae treated with low BPA when they were 7 dpf. The absence of significant differences in OMRs was opposite what we observed at earlier post exposure time points42. In contrast, retinal layer measurements taken 1-, 2-, and 4-weeks after BPA exposure all display modest effects localized to the INL, where bipolar cell bodies are located.

Zebrafish larvae aged 72 hpf or 7 dpf were exposed to water, vehicle (DMSO, 0.0003%), low BPA (0.001 µM) or high BPA (0.1 µM) for 24 hr and then returned to system water. Measurements of H&E-stained zebrafish retinas taken A 4-weeks or B 3–4 months postexposure. The dashed line separates data from 72 hpf (below the line) and 7 dpf (above the line) exposure groups. N = 3 per treatment group. Missing measurements due to tissue loss. For all graphs, bars show mean ± SE, individual data points are black dots. Significance is indicated by asterisks (α = 0.05).
We next asked if zebrafish adults (3-4 months postexposure) displayed any differences in OMR and/or retinal anatomy resulting from developmental BPA exposure. Anatomical measurements of H&E-stained adult retinal sections (N = 3 per treatment, Supplementary Fig. 4) again identified minimal effects of BPA. However, adults exposed to high and low BPA at 72 hpf exhibited reduced IPL thickness (p = 0.042) (Fig. 4B). Behaviorally, adults exposed to low BPA at 72 hpf had reduced OMRs compared to all other groups (water: 5 ± 1.44, n = 6; low BPA: 1.6 ± 0.53, n = 7 | vehicle controls: 2.3 ± 0.58, n = 8 | high BPA: 3.8 ± 0.67, n = 18), though these differences were not significant (H = 7.03, df, = 3, p = 0.071) (Supplementary Fig. 3B). Adults exposed to BPA at 7 dpf also showed a non-significant reduction in OMRs that was concentration dependent (low BPA: 2.4 ± 0.53 (n = 13) | vehicle: 4 ± 0.63 (n = 8) | high BPA: 1.5 ± 0.5 (n = 5) | water controls: 1.5 ± 0.51 (n = 14)) (Supplementary Fig. 3B).
We found no significant effect of treatment on survival to adulthood for either exposure age (Supplementary Fig. 5). However, if median survival is examined, exposure to low BPA at 72 hpf resulted in the lowest survival to adulthood as did exposure to vehicle at 7 dpf. This is consistent with our previous study demonstrating that, at the 2-week timepoint, exposure to low BPA at 72 hpf and vehicle at 7 dpf reduced larval survival, while low BPA at 7 dpf had no effect42.
Comparing the anatomical and behavioral data obtained 1-month vs 3-4 months postexposure identified comparable BPA-induced effects. Anatomical changes, though modest overall, were observed in the INL (1-month) or IPL (3-4 months), consistent with results from 1-week and 2-week postexposure tissues42. In general, low BPA exposure at 72 hpf caused more significant effects than either high BPA exposure or exposure at 7 dpf. No BPA-induced changes in OMRs were observed at either exposure age or postexposure assessment time.
BPA alters retinal physiology 3–4 months postexposure
To identify if developmental BPA exposure compromises adult retinal physiology, we examined photopic ERGs, evoked by a white light stimulus at 3.0 ND, from adult (3-4 months postexposure) eye cups of 72 hpf (Fig. 5) or 7 dpf (Fig. 6) treated fish to assess photoreceptor (a-wave), ON-bipolar (b-wave) and OFF-bipolar (d-wave) cell responses. Representative intensity-response curves are provided in Supplementary Fig. 6. In 72 hpf-exposed adults, both low BPA (0.92 ± 0.2 µV) and high BPA (0.86 ± 0.15 µV) increased a-wave trough amplitudes compared to vehicle (1.9 ± 0.28 µV; p < 0.001) (Fig. 5A) with no effect (p = 0.055) on a-wave trough time (Fig. 5B). b-wave amplitude was increased (p < 0.001) in adults exposed to low BPA at 72 hpf (17.2 ± 1.39 µV) compared to responses from vehicle (13 ± 1.42 µV) and high BPA (9.7 ± 0.42 µV) treatment groups; in contrast, high BPA reduced peak b-wave amplitude compared to vehicle (Fig. 5C; p < 0.001). There was no difference in b-wave peak time between BPA and vehicle treatment groups (Fig. 5D). d-wave amplitude in 72 hpf-exposed (Fig. 5E) adults was reduced in the low BPA group (4.7 ± 0.23 µV; p = 0.003) compared to vehicle controls (10.4 ± 2.13 µV). d-wave peak time (Fig. 5F) was also reduced (faster) in the low BPA group (93.4 ± 4.64 ms) relative to vehicle controls (116.9 ± 7.23 ms; p = 0.005).

A–F Mean photopic ERG recordings (top) from retinal eyecup preparations from adults exposed to water, vehicle (0.0003% DMSO), low BPA (0.001 µM), or high BPA (0.1 µM) when they were 72 hpf. Traces shown represent the mean response to the brightest stimulus (3.0 ND) from all eye cups in a given treatment group. Water = 5 eyes; vehicle = 10 eyes; low BPA = 6 eyes; high BPA = 18 eyes. The tissue was stimulated by 300 msec full-field white light pulses at 7 irradiance levels ranging from ND 3.0 (brightest)–6.0 (dimmest) in 0.5 increments. Red square pulse on all graces represents the photocell. White-light ERG responses had clear a-wave, b-wave, and d-wave components (noted on vehicle trace at top), which were evoked on an infrared (IR, RG780 filter) background. A a-wave amplitude, B a-wave trough time, C b-wave amplitude, D time to b-wave peak, E d-wave amplitude, F d-wave peak time. Data in A–F are presented as box and whisker plots: median line is in the center of the box and the ‘x’ is the mean. Significant differences are indicated with asterisks (α = 0.05).

A–F Mean ERG recordings (top) from retinal eyecup preparations from adults exposed to water, vehicle (0.0003% DMSO), low BPA (0.001 µM), or high BPA (0.1 µM) when they were 7 dpf. The tissue was stimulated by 300 msec full-field white light pulses at 7 irradiance levels ranging from ND 3.0 (brightest)–6.0 (dimmest) in 0.5 increments. These ERGs were in response to 10 replicates of a 7-step irradiance response series covering a 3-log unit range. Traces shown represent the mean response to the brightest stimulus (3.0 ND) from all eye cups in a given treatment group. Water = 11 eyes; Vehicle = 6 eyes; low BPA = 13 eyes; high BPA = 5 eyes. ERG a-wave, b-wave, and d-wave components are noted on the vehicle trace at the top. A a-wave amplitude. B a-wave trough time. C b-wave amplitude. D time to b-wave peak. E d-wave amplitude. F d-wave peak time. Data in A–F are presented as box and whisker plots: median line is in the center of the box and the ‘x’ is the mean. Significant differences are indicated with asterisks (α = 0.05).
Differences in photopic ERGs recorded from adults exposed to BPA when they were 7 dpf were also evident (Fig. 6). In this group, no differences in a-wave (Fig. 6A) or b-wave amplitudes (Fig. 6C) were identified relative to vehicle controls. Measurements of a-wave trough time were reduced in the high BPA (36.3 ± 9.21 ms) group relative to vehicle controls (87.5 ± 13.4 ms) (Fig. 6B) and low BPA (90.3 ± 8.82 ms; p = 0.003). In contrast, time to the b-wave (Fig. 6D) or d-wave peak (Fig. 6F) did not differ from vehicle (p = 0.403). d-wave amplitude was reduced in high BPA (1.3 ± 0.17 µV) treated fish compared to the low BPA (4 ± 0.25 µV) and vehicle control (4.5 ± 0.42 µV; p < 0.001) groups (Fig. 6E).
To identify potential physiological mechanisms underlying the observed BPA-induced changes in ERG responses, we used whole-cell patch clamp techniques to record voltage-gated currents. Acute BPA exposure is reported to inhibit voltage-gated Ca+2 currents but increase K+ currents26. We identified a decrease in depolarization elicited IK and ICa currents in zebrafish bipolar cells in adults exposed to BPA at 72 hpf (Fig. 7). IK amplitude was decreased (p ≤ 0.026; Fig. 7A, B) in response to a voltage step to −60 mV and to voltage steps equal to or more depolarizing than +10 mV (Vhold = −60 mV). For the most depolarizing steps (+40 to +60 mV), current amplitude in both BPA groups was reduced compared to vehicle controls (p ≤ 0.008). At slightly less depolarizing voltage steps (to +20 and +30 mV), high BPA (n = 3) was still different from vehicle controls (n = 6) (p = 0.016 and p = 0.013, respectively). ICa amplitude was also decreased in both BPA groups and often not evident (Fig. 7C, D). Calcium current amplitude was consistently reduced in low BPA treated tissue, with differences from vehicle controls noted in response to voltage steps to −20 mV (0.022) and −10 mV (p = 0.018).

Whole-cell patch clamp techniques in the zebrafish retinal slice preparation were used to record depolarization elicited potassium (IK) and calcium (ICa) currents from retinal bipolar cells. A Representative whole cell currents from vehicle (N = 6; 0.003% DMSO), low BPA (N = 3; 0.001 µM) and high BPA (N = 3; 0.1 µM) treated tissues. Responses were elicited from Vhold = −60 mV and in response to voltage steps from −80 to +60 mV in 10 mV increments. Brackets denote differences among treatment groups. B Current-voltage relationship depicting mean (±SE) current amplitudes at each voltage step. Differences in mean current amplitude were observed in response to a voltage step to −60 mV (high BPA vs. low BPA and vehicle, p = 0.005), to +10 mV (p = 0.026), +20 mV (p = 0.016, high BPA vs. vehicle), +30 mV (p = 0.013, high BPA vs. vehicle), and +40, +50, or +60 mV (p < 0.008, vehicle vs. both low and high BPA). C Representative whole-cell Ca+2 current elicited by a voltage step to −20 mV (Vhold = −60 mV), when the greatest amplitude differences were noted. Vehicle (N = 2), low BPA (N = 3), high BPA (N = 4). D Current-voltage relationship depicting mean ( ± SE) current amplitudes evoked in response to voltage steps from −40 to +10 mV, in 10 mV increments from a holding potential of −60 mV. Differences in mean current amplitudes were noted in response to voltage steps equal to or more depolarizing than −30 mV (p < 0.002). For all these membrane potentials, the calcium current was significantly reduced/absent in the BPA treatment groups.
Voltage-gated currents recorded from bipolar cells in adults exposed at 7 dpf (Fig. 8) also showed a reduction in ICa, but concentration-dependent effects on IK. Here, IK amplitude from high BPA treated tissue was consistently the largest amplitude (Fig. 8A, B) with significant differences noted in response to voltage steps equal to or more depolarizing than −20 mV. In contrast, the lowest amplitude for IK was found in low BPA treated retinas. Depolarizing voltage steps to −20 mV (p = 0.004) and −10 mV (p = 0.003) revealed differences between amplitudes from high BPA treated cells vs. vehicle control. A voltage step to 0 mV (p = 0.005) found high BPA current amplitude to be significantly different from both vehicle and low BPA values. For ICa, however, responses observed in adults treated at 7 dpf were similar to those identified in adults exposed to BPA at 72 hpf: calcium current amplitude was reduced/not evident in BPA-treated groups (Fig. 8C, D). Vehicle controls showed the largest calcium current amplitude, and these values were significantly different from amplitudes measured for both low and high BPA treatment groups at all voltage steps more depolarizing than −40 mV (p ≤ 0.002).

Whole-cell patch clamp techniques in the zebrafish retinal slice preparation were used to record voltage gated potassium (IK) and calcium (ICa) currents of retinal bipolar cells. A Representative whole cell currents from vehicle (N = 4), low BPA (N = 5), and high BPA (N = 4) treated tissues. Responses were elicited from Vhold = −60 mV and in response to voltage steps from −80 to +60 mV in 10 mV increments. Brackets denote differences among treatments. B Current-voltage relationship depicting mean ( ± SE) current amplitudes at each voltage step. Differences were observed for voltage steps to −20 mV or higher (asterisks). Steps to −20 mV (p = 0.004) or −10 mV (p = 0.003) identified a difference between high BPA vs. vehicle controls. A voltage step to 0 mV (p = 0.005) showed differences between high BPA and all other treatment groups. Voltage steps to +10 mV or more depolarized levels (p < 0.004) revealed a difference between low vs. high BPA treatment groups. C Representative whole-cell Ca+2 current elicited by a voltage step to −30 mV (Vhold = −60 mV), when the greatest amplitude differences were noted. Vehicle (N = 3), low BPA (N = 3), high BPA (N = 2). Whole cell Ca+2 currents elicited by voltage steps from −50 to 0 mV in 10 mV increments from Vhold = −60 mV. D Current-voltage relation showing peak amplitude of the calcium current. Low and high BPA exposure reduced ICa amplitude.
Taken together, our electrophysiology data indicates that developmental BPA exposure had persistent effects on adult retinal function. We also identified differential responses based on exposure age and with respect to BPA concentration. Both low (0.001 µM) and high (0.1 µM) BPA altered physiological responses in adults if exposure occurred at 72 hpf. In contrast, when exposure occurred at 7 dpf, the high BPA treatment group was more effective. This finding was consistent with LC-MS analysis of carcasses collected from adults developmentally exposed to high BPA (Fig. 3C) which revealed measurable accumulation of BPA in carcasses from fish exposed to high BPA only. Further, specific components of outer retinal signaling (i.e., photoreceptor, ON-bipolar, and OFF-bipolar responses) were differentially affected by BPA. While 24 hr acute exposure to adults did not alter a-wave responses (Fig. 2), exposure to BPA during development did affect a-wave parameters, suggesting a direct effect on photoreceptor development. Changes to b-wave and d-wave responses suggest bipolar cells, which are postsynaptic to photoreceptors, are also sensitive to BPA exposure. BPA’s effects on these second-order neurons could be due to a direct effect on the bipolar cells or an indirect effect of BPA on photoreceptors. Together, these data suggest potentially distinct cellular/molecular effects of BPA depending on exposure age, concentration, and cell type.
E2 and TH target gene expression are altered by low concentrations of BPA 3–4 months postexposure
It is widely accepted that BPA can exert its effects via classic ER-mediated genomic signaling mechanisms, binding to and activating nuclear ERα/β to alter transcription of E2-responsive genes. To determine if persistent BPA-induced alterations to retinal physiology are associated with perturbations to E2 genomic signaling as we observed in adults following a 24 hr BPA exposure (Fig. 3A), we examined expression of E2-regulated genes in retinal tissue: cyp19a1b (Aromatase B, neural aromatase), esr1 (ERα), and esr2 (ERβ). RT-qPCR analysis of retinal tissue from adults exposed at 72 hpf (Fig. 9A) revealed a dramatic upregulation of cyp19a1b in the low BPA group that was different from all other treatments (p ≤ 0.0038). Neither esr1 (ERα) nor esr2a (ERβ2) mRNA levels were altered by BPA relative to vehicle controls, though esr1 exhibited a BPA concentration-dependent trend of increased expression. Unable to obtain or design primers specifically targeting esr2b (ERβ1), we performed Western blots with retinal homogenates using a general esr2 (ERβ) antibody to potentially reveal ER isoform-specific differences in expression when compared with qPCR data. Retinal esr2 protein expression exhibited similar patterns in response to BPA as esr2a mRNA (Fig. 9B), suggesting either antibody detection of esr2a protein or similar effects to both esr2 isoforms. At the protein level, however, the decrease in esr2 protein in the low BPA group relative to both vehicle controls and high BPA was statistically significant (vehicle: p = 0.0048 | high BPA: p = 0.034).

RT-qPCR and Western Blot results in retinal homogenates of adult fish exposed to water, vehicle (0.0003% DMSO), low BPA (0.001 µM), or high BPA (0.1 µM) when they were 72 hpf. A RT-qPCR of estrogenic genes cyp19a1b, esr1, and esr2a. N = 3-5 fish per treatment for each gene (cyp19a1b: vehicle = 3, High BPA = 3, Low BPA = 3, Water = 4, | esr1: vehicle = 4, High BPA = 5, Low BPA = 5, Water = 5 | esr2a: vehicle = 4, High BPA = 4, Low BPA = 4, Water = 4). B Western Blot using an antibody against ERβ and β-actin as a loading control and quantification. Fold changes were calculated from densitometry analysis and normalizing values to loading control and water. N = 2 fish per treatment. C RT-qPCR of thyroid receptor genes thrab and thrb. N = 3 fish per treatment for each gene. For RT-qPCR, values were normalized to rpl13a housekeeping gene and water control. For all graphs, box plots reflect fold change. Asterisks indicate statistical significance: *p < 0.05, **p < 0.01, ***p < 0.001.
In contrast, BPA exposure at 7 dpf did not affect cyp19a1b, esr1, or esr2a gene expression (Supplementary Fig. 7A), though trends were observed. The high BPA treatment group displayed decreased ER expression (Supplementary Fig. 7A), and both the high and low BPA groups exhibited a trend of decreased cyp19a1b expression. esr2 protein expression in 7 dpf treated adult retinas (Supplementary Fig. 7B) again exhibited a similar pattern to esr2a mRNA, but here assessment at the protein level revealed a significant increase in the low BPA group (p ≤ 0.002) in contrast to the decrease observed when BPA exposure occurred at 72 hpf.
BPA also antagonizes triiodothyronine (T3)-dependent activation of thyroid hormone receptors (TRs), particularly TRβ, either directly by binding to TR56 or indirectly via non-genomic pathways57. Given the critical roles of thyroid hormones and TRs in retinal development58,59, and a recent publication showing that chronic BPA exposure affects thyroid hormone levels in zebrafish larvae25, we were motivated to assess the effect of BPA on TR transcription in retina. Exposure to low BPA at 72 hpf decreased thrb (TRβ) expression in retina of adults compared to vehicle control (p = 0.00078) and high BPA (p = 0.00097) groups (Fig. 9C). A similar reduction in thrab expression was also observed in response to low BPA, though it was not significant (Fig. 9C). Consistent with our estrogenic gene expression findings, BPA exposure at 7 dpf had no effect on thrab or thrb expression in adult retina (Supplementary Fig. 7C; high BPA data not available), though increasing trends in thrb expression were noted for the low BPA group.
Taken together, this data indicates that developmental BPA exposure has long-term effects on the expression of retinal cyp19a1b, esr2, and thrb—essential components of two hormone pathways highly involved in retinal development, maintenance, and function. Of note, significant transcriptional changes were identified when exposure occurred at 72 hpf, but not 7 dpf, and changes were predominantly induced by the lower BPA concentration.
Developmental BPA exposure activates non-genomic ER signaling
In addition to classical genomic signaling, BPA can also bind extranuclear membrane-bound ERs (mERs)60 and a G-protein coupled estrogen receptor (GPER)20 to initiate nongenomic signaling mechanisms. Activation of these membrane receptors triggers downstream changes in mitogen-activated protein kinase (MAPK) and phosphatidyl-inositol-3-kinase/AKT (PI3K/AKT) pathways, ultimately influencing gene transcription61,62. Significantly, BPA can activate these pathways at concentrations far below (pM - nM range)60 those required for nuclear ER binding63 and activation64. Considering the transcriptional effects we observed exclusively in response to low BPA and the key functions of MAPK and PI3K/AKT signaling in retinal development and cell survival, we next examined whether the long-term effects of BPA in retina are associated with changes in these nongenomic pathways.
Using retinal homogenates collected from adult zebrafish developmentally exposed to BPA, we used Western blots to assess activation (phosphorylation levels) of the three MAPK families (ERK, JNK, P38) and AKT, the primary regulator of downstream PI3K/AKT pathway effectors. At 72 hpf, both BPA concentrations decreased phosphorylated ERK (p-ERK) levels (Fig. 10A) relative to vehicle controls (high BPA: p = 0.00045 | low BPA: p = 0.00012), with low BPA causing a more pronounced reduction. Low BPA also increased JNK phosphorylation (p-JNK; Fig. 10B, p = 0.048). Interestingly, high BPA at 72 hpf had no effect on p-JNK, with levels similar to vehicle controls. We found no significant effect of high BPA at 72 hpf on phosphorylated P38 levels (p-P38; Fig. 10C) relative to vehicle controls, though an increased trend was observed. Due to undetectable total P38 expression in response to low BPA and water treatments, we were unable to quantitatively normalize p-P38 to total protein levels. However, qualitative comparison of Western blots suggests that low BPA increased p-P38 relative to vehicle control and high BPA. Consistent with JNK and P38, high BPA at 72 hpf had no effect on p-AKT levels (Fig. 10D). Again, low BPA at 72 hpf reduced expression below detectable levels, thus preventing analysis of low BPA effects on p-AKT.

Western Blot and quantification for MAPK protein and AKT phosphorylation in retinal homogenates of adult fish exposed to water, vehicle (0.0003% DMSO), low BPA (0.001 µM), or high BPA (0.1 µM) when they were 72 hpf (left) (A–D) or 7 dpf (right) (E–H). A, E ERK, p-ERK. B, F JNK, p-JNK. C, G P38, p-P38. D, H AKT, p-AKT. N = 2 fish per treatment for each blot. For all blots, β-actin was used as a loading control. Quantification was performed using values obtained from densitometry analysis and normalizing phosphorylated protein to total protein and water control. Box plots show fold change. Asterisks indicate statistical significance: *p < 0.05, **p < 0.01, ***p < 0.001.
Assessment of MAPK and AKT phosphorylation in retinas of adults exposed at 7 dpf also revealed BPA-induced alterations, though treatment responses differed. Low BPA at 7 dpf decreased p-ERK (Fig. 10E) with respect to all other treatments (vehicle: p = 0.025 | water: p = 0.0025 | high BPA: p = 0.0076) whereas high BPA had no effect. p-JNK, in contrast, was prominently increased by high BPA exposure at 7 dpf (Fig. 10F) compared to all other treatments (vehicle: p = 0.0017 | water: p = 0.0023 | low BPA: p = 0.0035). Neither concentration of BPA at 7 dpf affected p-P38 levels (Fig. 10G). Interestingly, low and high BPA at 7 dpf had pronounced, opposite effects on p-AKT expression (Fig. 10H): high BPA increased p-AKT relative to other treatments (vehicle: p < 0.001 | water: p = 0.0017 | low BPA: p < 0.001) while low BPA reduced levels (p < 0.001).
These findings indicate that BPA exposure during development has persistent effects on ERK, JNK, and AKT pathway activation in the retina, with effects depending on the age of exposure and BPA concentration. Low BPA decreased ERK phosphorylation in adult retinas when exposure occurred at 72 hpf and 7 dpf, while JNK and AKT phosphorylation was differentially affected by low vs high BPA depending on the exposure age. Overall, this data suggests BPA exposure can target non-genomic signaling pathways in retina, providing a potential mechanism underlying its long-term effects, especially at low concentrations.
Discussion
Environmental contaminants are a continued health concern, with exposure occurring through ingestion and/or inhalation8. Developmental exposure to these contaminants, even at low levels, changes neurogenesis65 with subsequent sequelae reported in both humans and animal models. One such compound is BPA, an endocrine disruptor and ubiquitous global contaminant8,66 with measurable levels reported in ≥86% of urine and/or plasma samples worldwide8. Some published studies using zebrafish report that BPA exposure causes hyperactivity in larvae34,35 whereas others report reduced responses in the dark9,25. Behavioral deficits have been observed in adults that were exposed during development35, suggesting both short- and long-term effects of exposure. Developmental BPA exposure can also target sensory systems, leading to malformed otoliths67, inhibited regeneration of lateral line neuromasts68, reduced responses to green and red light25, and reduced habituation and prepulse inhibition in response to an auditory startle stimulus69. While these studies clearly show deleterious effects of BPA exposure, few examine if early life exposure leads to persistent effects by measuring outcomes after a recovery/washout period. Here, we report the novel finding that a transient 24 hr BPA exposure during a critical period of visual system development leads to persistent molecular changes in gene expression and intracellular signaling that are coupled to perturbations in adult retinal function.
Zebrafish have 3 ER types, one isoform of ERα and two isoforms of ERβ (ERβ1 and ERβ2) which are encoded by different genes: esr1, esr2b, and esr2a respectively. All are sensitive to E223,70,71, able to bind BPA18,72, and have specific expression patterns during development73. Zebrafish also express GPER74. BPA treated larvae show reduced eye size25 and decreased expression of genes involved in visual perception37,75 suggesting BPA exposure can specifically, and adversely, affect the visual system. Published studies of adult zebrafish exposed to BPA report a significant upregulation of aromatase B (cyp19a1b) expression in brain18,34,37,72,76 consistent with its role as a weak estrogen agonist18,77. To determine if BPA has a similar role in zebrafish retina, we examined the effect of BPA exposure on estrogen responsive genes (cyp19a1b, esr1 = ERα, and esr2a = ERβ2) in retinal homogenates from adults assessed immediately after a 24 hr exposure or after developmental exposure to BPA at 72 hpf or 7 dpf. We observed no significant effect of BPA exposure on any of the genes examined when assessed in adults immediately after a 24 hr exposure. Average fold inductions of AroB and ERa were increased relative to vehicle controls, but these differences were not statistically significant, likely due to a large variation in fold changes among biological replicates. However, we found that exposure to low BPA at 72 hpf increased cyp19a1b and decreased esr2 protein levels in adult retinal homogenates. We also observed increased esr1 and reduced esr2a expression that were not significant. These results support 48-96 hpf being an ‘estrogen sensitive window’ for zebrafish76 and agree with the reported BPA-induced overexpression of cyp19a1b. The observed decrease in esr2a, though nonsignificant, is consistent with reduced esr2 protein levels and with other studies demonstrating an antagonistic effect of BPA on ERβ78 due to BPA interfering with the ligand binding domain and preventing the necessary conformation for transcriptional action7,79. Low BPA exposure at 7 dpf, in contrast, did not affect expression of cyp19a1b, esr1, or esr2a in adult retinal tissue though there was an increase in esr2 protein in retinal homogenates, as in ref. 21. Overall, these results are consistent with BPA acting as a weak ER agonist and are the first to report prolonged estrogenic effects of BPA in retinal tissue.
A surprising finding from our qPCR results was a lack of significant differences in esr1 expression at either age or BPA concentration. Several publications note BPA is an ERα agonist that induces ERE-mediated transcriptional activation of esr114,22. There are three possibilities for the differential response of BPA on esr1 and esr2 in our retinal homogenates: (1) absence of ERα in retinal tissue, (2) differential affinity for ERα and ERβ by BPA, and (3) posttranslational modification of activated ER. Initial characterization of zebrafish estrogen receptors did not report esr1 in retinal tissue70, though both esr2a and esr2b were present. A subsequent study, however, identified all three ER types in adult zebrafish retinas71. Splice variants of ERα resulting in short (ERαS) and long (ERαL) isoforms have also been reported in zebrafish and BPA displays preferential selectivity for ERαS, which is not the isoform found in retina80. Thus, the reduced expression of esr1 in our retinal homogenates may reflect overall reduced levels of ERα in zebrafish retina, though we were able to measure expression of esr1 in our homogenates. Second, BPA is reported to have a higher affinity for ERβ than ERα in vivo21,22,64. Thus, it is possible that the low concentrations used here were insufficient for ERα binding but sufficient to interact with ERβ and/or suppress esr2 transcription. Finally, the different actions of BPA on ERα and ERβ in our retinal homogenates may be due to posttranslational modification through phosphorylation or altered recruitment of cofactors61. Phosphorylation of ERs can occur with or without ligand binding81 and ERα transcription can be modulated by phosphorylation from intracellular kinases such as Akt and MAPKs70, increasing or decreasing ERα -ERE affinity depending on the kinase and phosphorylation site82. Therefore, it is possible that transcriptional activity of ERα to initiate target gene expression, including its own, was inhibited by phosphorylation82. Multiple phosphorylation sites have been identified on both ERα and ERβ81,82, supporting this hypothesis, though these sites have yet to be reported in zebrafish.
In addition to persistent changes in genomic signaling pathways, we also observed differential phosphorylation of Akt and the MAPK associated kinases ERK and JNK in adult retinas following developmental BPA exposure. Adult retinal homogenates exposed to low BPA at 72 hpf had decreased p-ERK and p-Akt protein levels but increased p-JNK. Low BPA exposure at 7 dpf also significantly reduced p-ERK and p-Akt levels in adult retinal homogenates, while high BPA exposure at this age significantly increased adult p-JNK and p-Akt levels. Activated kinases, such as ERK and Akt, can regulate ER activity through phosphorylation81, supporting the observed differential effects of BPA on ER-mediated gene expression of estrogen target genes. The intracellular MAPK/ERK signaling pathway is also associated with development, proliferation, and differentiation of cells83; while the JNK signaling pathway is associated with apoptosis and proinflammatory cytokines84. These different signaling pathways, which are associated with ERα activation and ERβ inhibition, respectively, depend on cell context with the balance of receptor activation determining the overall response of the cell78. Our results, therefore, suggest that exposure to low BPA at 72 hpf has the potential to cause increased apoptosis/inflammation (JNK pathway) and decreased differentiation (ERK and Akt pathways). Retinal layer measurements made in adults exposed to low BPA at 72 hpf identified reductions in inner plexiform layer thickness which could be due to differential activation of MAPK pathways following BPA exposure. In contrast, BPA exposure at 7 dpf revealed concentration-dependent effects: low BPA exposure decreased p-ERK and p-Akt (associated with reduced differentiation); high BPA increased p-JNK (associated with increased apoptosis) in retinal tissues. However, these molecular changes were not correlated with changes in either retinal morphology or thickness measurements of adult retinas exposed to BPA at 7 dpf.
While BPA binding to extranuclear ER can trigger MAPK pathway activation78, the observed activation of the intracellular kinase pathways observed here more probably occurred via BPA binding to GPER, the G-protein coupled ER, as membrane bound ERs have not been identified in zebrafish74. In transfected cells79 and mammals66, BPA has a higher affinity for GPER than either ERα or ERβ, and BPA binding to GPER activates Akt and ERK pathways79, consistent with changes we observed. However, BPA binding to zebrafish GPER is inhibitory21, which could have caused the reductions in p-ERK and p-Akt identified in retinal homogenates in response to low BPA exposure. Further, BPA antagonizing ERβ (observed in our genomic data) is associated with increased apoptosis78, which is correlated with the observed increase in p-JNK levels and suggests persistent, overlapping genomic and nongenomic effects of low BPA in zebrafish retina. Adults exposed to high BPA showed similar responses, though effects were more pronounced at the 7 dpf exposure age. Though we did not assess estrogen related receptors (ERRs), BPA could also be mediating non-genomic effects by binding to ERRγ66 which can also trigger MAPK pathways28. BPA binding to both ERRγ and GPER in zebrafish may be a mechanism whereby some MAPK pathways are activated, and others are not. Future experiments are needed to address this hypothesis.
Overall, our molecular data supports BPA as an estrogen agonist that can persistently activate both genomic and nongenomic signaling pathways after developmental exposure. Low BPA exposure at 72 hpf, an age of high neurogenesis within the zebrafish visual system, causes long-term alterations to both genomic signaling and nongenomic pathways, whereas high BPA exposure at 7 dpf, when most retinal neurogenesis is complete, targets nongenomic pathways.
We assessed the impact of developmental BPA exposure on later visual function both behaviorally and physiologically. We began by recording OMRs, a vision-based behavior initiated in the retina. OMR circuitry involves projections of directionally selective ganglion cells to the pretectum, linking visual and motor circuits85 with spinal projection neurons mediating the movement associated with the OMR86. Our previous work showed that BPA exposure at 72 hpf and 7 dpf enhanced optomotor responses in zebrafish larvae when measured 1- and 2-weeks after exposure42. Here, however, we find that extending the postexposure time to either 1-month or 3-4 months decreased OMRs, ultimately revealing no difference from controls. This response difference is consistent with previous reports in zebrafish identifying initial BPA-induced hyperactivity in response to acute/early exposure34,35 but later reductions in locomotor behavior35. While it is possible that BPA clearance over time is responsible for effects observed at earlier (1-2 weeks) vs. later (1-month or 3-4 months) timepoints, we believe this to be unlikely for two reasons. First, laboratory studies indicate BPA elimination in zebrafish is rapid, occurring through intestinal and gill (renal) pathways49 and via conversion in the liver48. This elimination involves an initial rapid phase (within 1 hr), followed by a slower phase (139 hr)87. Elimination, therefore, would be reached even before the first postexposure time point. Second, LC/MS analysis of adult zebrafish carcasses found measurable BPA levels in the high BPA exposure groups, suggesting some bioaccumulation.
Our morphological data identified only modest effects of early BPA exposure at both the 4-week and 3-month time points. These findings were surprising and contrasted with studies showing BPA-induced morphological differences. For example, exposing zebrafish larvae to BPA until 8 dpf reduced both RPE and INL thicknesses at 4 dpf, which preceded locomotor differences recorded at 7 dpf25. Exposure to BPS until 6 dpf changed the structure of the optic nerve, decreased expression of gfap, and caused a decrease in the GCL39. BPS exposure for 120 days decreased IPL and GCL thickness, increased thickness of the RPE and changed expression of opsin genes41. Differences in cone signals and morphology were also observed following exposure to BPS from 2-5 dpf40. The difference between these reports and our data could be twofold. First, these published studies assessed bisphenol-induced effects immediately after a chronic exposure that lasted several days. Our study, in contrast, used only 24 hr exposure with assessments occurring several weeks/months later. While the transient exposure was efficacious, the longer post-exposure interval may have dampened BPA-induced effects. The second consideration is the concentration used. Changes in retinal morphology are seen in response to 10 and 100 µg/L BPS40,41, ≤ 3 mg/L BPS39, and 2.8 mg/L BPA25. Our BPA concentrations of 0.001 µM (0.22829 µg/L) and 0.1 µM (22.829 µg/L) are well below many of the values in the literature. Consequently, the modest effects we observed may reflect differences in concentration and/or duration of exposure to BPA.
Estrogenic compounds, in general, show concentration dependent effects on ion channels, with low doses increasing and high doses decreasing responses88. Consistent with our molecular data, we found that BPA exposure at 72 hpf induced greater changes in adult ERGs compared to exposure at 7 dpf. Adults exposed to low BPA at 72 hpf had increased amplitudes for photoreceptor a-wave and ON-bipolar cell b-wave responses but reduced and faster OFF-bipolar cell d-waves. High BPA exposure at 72 hpf also increased photoreceptor a-wave amplitudes in adult ERGs, but reduced b-wave amplitudes; no changes were observed for OFF-bipolar d-waves. Adult ERGs from animals exposed to BPA at 7 dpf were characterized by a faster photoreceptor a-wave, increased b-wave amplitude, and a faster, reduced d-wave in the high BPA treatment group only. Comparison of ERG responses of adults acutely exposed to BPA for 24 hr vs. adults developmentally exposed to BPA identified a consistent reduction in both b-wave and d-wave amplitudes, particularly between the acute low BPA and 72 hpf low BPA treatment groups. Importantly, these results show that bipolar cell responses were altered at all ages and BPA concentrations tested, suggesting they may be the retinal cell type most sensitive to BPA.
It is also worth noting that we often observed significant functional differences between high and low BPA treatment groups, though neither value may have been different from vehicle controls. Differences in ERG wave components were evident between the two BPA groups in adults developmentally exposed at 7 dpf. b-wave differences between BPA groups were also evident in ERGs recorded from adults acutely exposed to BPA for 24 hr. BPA exposure at 72 hpf resulted in concentration dependent differences in retinal anatomy at 1-week post exposure and OMR responses at 4-weeks. Although not statistically significant compared to vehicle controls, these differences further highlight the unique biological effects of BPA at different concentrations.
To identify a potential mechanism for the BPA-induced ERG b- and d-wave differences, we recorded voltage-gated current responses from bipolar cells using whole cell patch clamp techniques in zebrafish retinal slices. We focused on depolarization-elicited K+ and Ca+2 currents because these voltage-gated currents are prominent in zebrafish retinal bipolar cells89 and BPA has specific effects on both current types26. Recordings from adults exposed to high BPA at 72 hpf identified decreased amplitudes of depolarization-elicited K+ and Ca+2 currents in response to both low and high BPA exposure. Decreased IK and ICa amplitudes would reduce overall responses and slow/reduce repolarization which agrees with the decreased b-wave and d-wave amplitudes observed in the high BPA exposure group. Voltage-gated currents recorded from bipolar cells in adults exposed at 7 dpf revealed high BPA exposure increased IK but decreased ICa amplitudes. These differences would cause faster repolarization of the cells and quicker, smaller responses, which agrees with the increased ERG b-wave responses and faster d-wave peak times observed for the high BPA group.
To further examine changes in bipolar cell responses, we examined differences in dopamine and GABA systems in retinas of adults exposed to BPA at 72 hpf because of the consistent physiological differences noted at this exposure age. BPA exposure, as well as exposure to the bisphenol analogs BPAF, BPF, and BPS, can change levels of neurotransmitters16, suggesting a potential mechanism for the observed physiological effects. These results show that high BPA exposure at 72 hpf increased levels of glutamic acid decarboxylase (GAD, a marker for the inhibitory neurotransmitter GABA) in adult retinal homogenates; whereas low BPA exposure decreased GAD levels (Supplementary Fig. 8B, C). In contrast, increased levels of tyrosine hydroxylase (TH) were evident, regardless of BPA concentration (Supplementary Fig. 8A, C). Zebrafish bipolar cells have GABA-evoked inhibitory currents90 due to inputs from presynaptic horizontal and/or amacrine cells. Dopamine is released from interplexiform and/or amacrine cells in inner retina and can impact both bipolar and photoreceptor cell responses91. High BPA-induced increases in GAD levels/GABA synthesis would decrease bipolar cell responses, which was observed in both patch clamp and ERG recordings from this treatment group (72 hpf exposed adults). The low BPA-induced reduction in GAD levels suggests reduced inhibition, which is consistent with increased ERG a-wave and b-wave responses in this treatment group. Zebrafish bipolar cell IK currents are decreased following dopamine application92 and dopamine can decreases ON-BC excitation93 by strengthening GABAergic surround responses91, both of which are consistent with our physiological data. Our results indicate low BPA increased b-wave amplitudes and high BPA decreased b-wave responses, suggesting concentration-dependent effects. Further, BPA exposure at 72 hpf leads to reduced inner retinal thickness and changes in GABAergic and dopaminergic circuits in adult zebrafish retinas which, in turn, affects bipolar cell responses, ultimately altering ERGs.
Disruption of estrogen signaling in zebrafish retina using known estrogen receptor modulators such as tamoxifen and 4-OH-A (formestane) alter eye development and retinal thickness94, suggesting proper estrogenic signaling is important for normal eye development. Developmental exposure to 4-OH-A at the same ages used in this study reduces adult OMRs95, supporting the need for estrogen signaling in retinal development. Our physiological responses from adult retinas further show that developmental BPA exposure alters light-evoked and voltage-gated responses in retinal bipolar cells. The effect of BPA depends on both concentration and age of exposure. Whether these effects occur because BPA is directly interacting with the ion channels on bipolar cells themselves26 or indirectly altering neuronal activity via intracellular pathways and/or non-genomic estrogenic signaling88 is not known, though our data supports both hypotheses. However, the long interval between exposure age and adult testing suggests a direct effect of BPA is unlikely and points to BPA-induced persistent changes at the molecular level that ultimately compromise retinal function. Another possibility is that BPA targets other retinal cells that are presynaptic to bipolar cells, suggesting a more generalized retinal effect of exposure. The observed changes in GAD and TH levels in retinal homogenates because of BPA exposure support this hypothesis.
While our data reveals a persistent effect of developmental BPA exposure on estrogen signaling in zebrafish retina that is correlated with reduced physiological responses, we can’t rule out the possibility that BPA is also interacting with other signaling pathways.
BPA can also bind to thyroid receptors (TR), androgen receptors (AR)48, and retinoic acid receptors (RAR)37. BPA binding to AR increases cyp19a1b expression in brains of zebrafish larvae34 and ar transcripts are present in developing retina96, suggesting a potential mechanism in this tissue. However, a BPA-induced increase in cyp19a1b was not suppressed by co-application with an AR blocker72 and BPA does not impair androgen effects in cell lines7, suggesting a more indirect mechanism of BPA on AR signaling. Retinoic acid signaling establishes dorsal-ventral patterning in zebrafish retina97 and is involved in photoreceptor development98. A reported decrease in eye size observed in zebrafish larvae after BPA exposure was associated with altered retinoic acid signaling37.
Thyroid signaling is also important for proper retinal development58,59. Specifically, thyroid receptor β2 (TRβ2) is required for the correct expression of opsins in cone photoreceptors58 and loss of this receptor shifts spectral sensitivity curves and retinal ERG responses in zebrafish99,100. As in mammals, zebrafish have TRα and TRβ receptors. However, zebrafish have a single gene encoding TRβ (thrb) and two genes encoding TRα (thrab, thraa)101,102 and all isoforms are found in retina. Further, the thrb gene encodes 3 variants, one of which is TRβ2103.
BPA affects TH signaling as an antagonist of TR56,57 which prevents the binding of T3 to the receptor48. BPA has a greater affinity for TRβ, particularly at low (0.001-0.1 µM) concentrations25,104 with low BPA reducing the recruitment/binding of cofactors necessary for transcription105. In agreement with these reports, we found that exposure to low (0.001 µM) BPA at 72 hpf, but not 7 dpf, significantly reduced thrb expression in adult retinal homogenates. We do not know if this effect is due to a direct interaction of BPA with TRβ or an indirect effect initiated via nongenomic thyroid hormone receptor signaling105,106. Low BPA concentrations can activate these pathways106 and thyroid mediated nongenomic signaling involves MAPK/ERK and Akt pathways105. Given the role of TRβ in proper photoreceptor development, decreased thrb expression may contribute to altered photoreceptor a-wave responses we observed in adult ERGs from this treatment group. Current experiments are more specifically probing the cross-reactivity of BPA on retinal ER and TR to assess the role of each hormone in BPA-induced effects.
Chronic exposure to the BPA derivative TBBPA until 5 dpf upregulated TRα and dio1 and downregulated tpo expression, changes that were correlated with decreased eye size and decreased optokinetic responses107. A subsequent study with TBBPA identified decreased thickness of the retinal RPE108, suggesting specific effects in retina. However, TBBPA increased whole body T3 levels in 5 dpf zebrafish larvae, but did not alter TH levels in the eyes108 suggesting a general effect of this EDC109. BPS exposure until 5 dpf revealed an interaction with TRβ receptors leading to the induction of cyp26a1, an enzyme involved in retinal metabolism40. Finally, chronic exposure to BPA until 8 dpf decreased T4 and rT3 levels at 5 dpf, with decreased eye diameter and changes to retinal layers observed at 4 dpf25.
Together these findings indicate that bisphenols can impact TH signaling, though the specific impact in retina seems less clear. Finally, BPA exposure has recently been shown to alter genes associated with lipid, cholesterol, and glucose metabolic pathways21,37 in zebrafish. Cholesterol is the single starting molecule for the synthesis of steroid hormones, such as estrogen. BPA affecting cholesterol metabolism, therefore, would have downstream effects on estrogen synthesis.
A limitation of this study is the reliance on bulk retinal homogenates for RT-qPCR and Western blot analyses as retinal layer- or cell-type specific effects of BPA exposure are likely and suggested by our physiology findings. Furthermore, while our work highlighted transcriptional outcomes and intracellular signaling pathways, we did not investigate epigenetic modifications, which are key regulators of developmental transitions and events and could underly the development deficits observed here. BPA has been shown to influence DNA methylation110,111,112 and histone modifications/histone modifying enzymes in zebrafish112,113 and other species114, which could play a role in mediating the observed effects on gene expression. Another limitation is that the temporal resolution of our study captures snapshots at only a few timepoints in development and our molecular analyses are limited to adulthood. Therefore, they may miss significant transient or dynamic changes at different or earlier stages of visual system development. Addressing these limitations in future research would provide a more comprehensive understanding of BPA’s effects on retinal development and function.
Another limitation was the ability to obtain enough tissue for all analyses. While there were no statistical differences in survival to adulthood, there were large variations in survival with each spawn and/or treatment, particularly the high BPA exposure group. Previous reports indicating chronic 5 day larval exposure to 25 µM69 or 40 µM9 BPA did not impact survival agree with our data. However, qualitative differences, particularly in the high BPA treatment, suggest there may be subtle effects of exposure.
DMSO was used in this study as a carrier solvent to dissolve BPA directly into the tank water. To ensure the vehicle alone did not cause estrogenic effects, a DMSO treatment (0.0003%) was included for each analysis. DMSO consistently showed effects different from the water control both at the mRNA and protein level of estrogenic target genes/proteins. This was unexpected as DMSO is widely used as a solvent in exposure studies, including those examining estrogen signaling in zebrafish. However, there are reports in the literature of DMSO effects. For example, in dogs, 20 µL of DMSO used as a vehicle solvent for coumestrol increased progesterone levels and altered vaginal epithelial cells55,115. In fish, 0.1% DMSO increased expression of ERα and ERβ in cultured salmon hepatocytes54. Increased aromatase expression53 and StAR protein levels and P450scc mRNA116 were elevated in response to in vivo exposure in Atlantic salmon, suggesting estrogen stimulating effects. Larval zebrafish chronically exposed to 0.1-1% DMSO showed increased swimming speeds but an overall decrease in time spent active at the highest concentration51. Similarly, concentrations of DMSO ranging from 0.01-1% were reported to cause hyperactivity in zebrafish larvae, with morphological abnormalities noted only at the 1% exposure group50. We found differences in our OMR data between vehicle and water controls, in agreement with low DMSO concentration effects on behavior50. Hillare et al.52 report that DMSO exposure up to 2% did not influence survival from 2-96 hpf; however, concentrations ≥0.01% levels of heat shock protein (hsp) 70 were elevated, indicating a stress response52,117. In our data, we found differences in vehicle vs. water controls in some of our results for the functional assays (OMR, ERG) and the molecular data, in agreement with the above reports. Interestingly, we see differences at a DMSO concentration well below what has been previously reported (0.0003%), suggesting sensitivity to DMSO for some assessments, but not all. These differences may also be due to age/condition of the animals when exposure occurred55,115.
Our results indicate that a brief exposure to BPA during key stages of visual system development has long-term implications to genomic and non-genomic E2 signaling mechanisms in the retina. A transient exposure to BPA caused upregulation of AroB and downregulation to ERβ and TR gene expression in the adult retina, with a BPA concentration of 0.001 µM producing more significant transcriptional effects than a higher BPA concentration (0.1 µM). Changes in the activation of cytoplasmic E2-initiated signaling cascades were also observed, with decreased phosphorylation of ERK, JNK, and Akt. The impacts of BPA were more prominent when exposure occurred at 72 hpf compared to 7 dpf, suggesting the developing retina is more susceptible to long-term dysregulation of E2 signaling by BPA.
We propose the following mechanism for the adverse effects of early exposure to BPA on retinal function. Exposure to low BPA at 72 hpf, when the retina has formed44 and is becoming functional43, initially affects eye size, retinal layering and optomotor responses42. However, these anatomical and behavioral differences dampen with age, though long-term changes in genomic signaling characterized by increased cyp19a1b expression, decreased ERβ protein, and decreased thrb expression persist to adulthood. Low BPA exposure at this age activated the JNK pathway, a regulator of apoptosis, and reduced p-ERK levels, which likely affects cell differentiation/proliferation (Fig. 11A). GABAergic amacrine cells have processes within the IPL that are associated with bipolar cell terminals and the smaller IPL may contribute to the overall decrease in retinal GAD levels. Reduced GAD levels suggest decreased inhibitory signaling in the retina, causing larger photopic responses. TH levels, in contrast, were increased in adult retinal homogenates, which would reduce bipolar cell responses. ERG responses identified increased photoreceptor and ON-bipolar cell responses, but reduced OFF-bipolar cell responses, which were correlated with BPA-induced changes in voltage-gated K+ and Ca+2 currents recorded from retinal bipolar cells. When BPA exposure occurred slightly later in development - 7 dpf - when most retinal neurogenesis is complete and vision behaviors can be reliably recorded118, differences in retinal morphology were not evident and initial differences in OMRs diminished with age. In adults, most effects were observed in the high BPA treatment group, consistent with measurable BPA levels in these carcasses (Fig. 3C). At this exposure age, non-genomic estrogenic pathways associated with apoptosis were activated (Fig. 11B), though we observed no anatomical differences in the high BPA treatment group. Nonetheless, bipolar cell responses were affected as noted by increased IK but decreased ICa amplitudes, increased ERG b-wave amplitudes, and reduced, but faster, ERG d-wave responses.

Schematic diagrams show the effects of low BPA (left) and high BPA (right) on estrogen signaling pathways when exposure occurred at 72 hpf (A) and 7 dpf (B). A Exposure to low (0.001 µM) BPA at 72 hpf activated both genomic and non-genomic estrogen signaling. Low BPA binding to intracellular ER increased cyp19a1b expression. In contrast, low BPA binding to ERβ was antagonistic, decreasing ERβ protein levels. Low BPA binding to GPER (dark blue) reduced p-ERK levels (−); while low BPA binding to membrane bound ER increased p-JNK levels (+). Functionally, low BPA exposure was associated with increased ERG responses, changes in GAD and TH protein levels, and reduced OMRs. High BPA (0.1 µM) exposure at 72 hpf (right) in contrast, lead to reduced p-ERK levels only, suggesting a specific effect on non-genomic pathways. This result was associated with increased GAD and TH levels in retinal homogenates, increased photoreceptor, but decreased ON-bipolar cell responses. B Exposure to low (0.001 µM) BPA at 7 dpf activated both genomic and non-genomic estrogen signaling. Low BPA binding to intracellular ER increased ERβ protein levels. Low BPA binding to GPER (dark blue), in contrast, reduced p-ERK (−) and p-AKT (−) levels. High BPA exposure at 7 dpf (right) activated non-genomic pathways only, increasing p-JNK (+) and p-AKT (+) levels. Functionally high BPA exposure at 7 dpf caused physiological changes in adult zebrafish retinas by increasing ON-bipolar cell responses, reducing OFF-bipolar cell responses and quickening both OFF-bipolar and photoreceptor responses. High BPA exposure was also associated with the lowest positive OMRs in adults. For both A and B, the timeline at the top indicates exposure age (yellow box) and when assessments were made (3–4 months of age).
Though use of BPA has been banned in baby bottles6 and food contact materials14, it is still a chemical of concern. While many studies show that chronic (several day) exposure during development induces morphological, behavioral, and genomic changes, very few have examined the effect of a brief exposure and/or the impact of a recovery/washout period on assessed outcomes. This work fills that gap in knowledge by demonstrating that a brief early-life BPA exposure, even at low concentrations, is sufficient to cause persistent adverse effects. The visual deficits we observed correlate with BPA-induced changes in estrogen and thyroid signaling pathways, despite the absence of obvious morphological abnormalities, suggesting that subtle molecular disruptions can have significant functional consequences that may go undetected with anatomical assessments. Further, the adverse effects observed here occurred at BPA levels that were environmentally relevant and well below those used with chronic developmental studies, underscoring the continued relevance of bisphenol exposure studies.
Our findings underscore the potential for transient low-dose BPA exposure to disrupt development, reinforcing the need to re-evaluate acceptable exposure levels and reduce ongoing environmental contamination. This work highlights the importance of continued public health efforts to limit early-life exposure to endocrine-disrupting chemicals like BPA, which may have lasting effects on ecological and human health.
Methods
Animals
Adult, wildtype, mixed sex zebrafish, Danio rerio, were maintained in the laboratory at 28-29 °C on a 14 hr light: 10 hr dark photoperiod in an Aquatic Habitats (Pentair, Apopka, FL) recirculating system and fed daily. Embryos were obtained from in-house spawning of adults or purchased from a commercial supplier (LiveAquaria, Rhinelander, WI). To obtain embryos, adults were placed into a breeding chamber the night before, with multiple males and females ( ≤ 8-10 of each, at a 1:2 male-to-female ratio) used for each spawn. The following morning, 30-90 minutes after light-ON, embryos were collected, staged, and placed in petri dishes of system water. Petri dishes were housed in an incubator (Heratherm; Thermo Fisher, Waltham, MA) at the same temperature and photoperiod as the adults. Standard environmental parameters are 27-30 C, pH 6.8-7.6, 1000-1200 µS/cm conductivity, 0-20 ppm nitrate. We have complied with all relevant ethical regulations for animal use. All procedures were approved by the Institutional Animal Care and Use Committee of American University.
Exposure ages, treatment groups, tissue collection
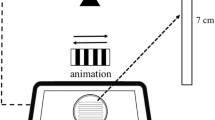
At 72 hours postfertilization (hpf) and 7 days (d)pf, zebrafish were placed into one of the experimental treatments for 24 hr (Fig. 1). A 24 hr treatment duration was chosen based on our previous work42,95 and because of the stability of BPA6 during this time period. 72 hpf and 7 dpf were selected as the exposure ages because they correspond to specific events in visual system development, with the 70-74 hpf developmental timepoint being associated with functional retinal transduction pathways44, innervation of the optic tectum45, and the onset of visually guided behaviors43. At 7 dpf, visually guided OMRs can be reliably recorded118 and estradiol synthesis and signaling are present46,47. An additional set of experiments used adult zebrafish acutely exposed to BPA for 24 hr. These adults were used to assess the level of bioaccumulation of BPA after a 24 hr exposure (Fig. 3C) and as a comparison between acute and prolonged effects of BPA on vision physiology.
BPA (bisphenol A or 2,2-Bis(4-hydroxyphenyl)propane; TCI, Tokyo) was initially mixed as a 313 µM stock solution, solubilized in DMSO (dimethylsulfoxide; VWR, Swedesboro, NJ) and diluted with system water to the final concentration in the experimental chamber. BPA was administered to the fish directly through tank water. Exposure occurred in 100 mm glass petri dishes with an average of 30 larvae per dish, with 2 dishes (technical replicates) per exposure. Biological replicates used larvae from at least 2 different spawns. Individual treatment groups were: (1) DMSO vehicle control (0.0003%, corresponding to the DMSO concentration in the high BPA group), (2) water control, (3) 0.001 µM (low) BPA, and (4) 0.1 µM (high) BPA. These nominal concentrations were selected because they are comparable to levels in the environment119 and human tissue10. These concentrations have also been used by others14,34,35, including ourselves42 allowing us to compare our results with published studies. Consistent with these studies, we have omitted use of E2 and/or EE2 in our experiments to focus specifically on BPA-induced effects.
After each larval exposure, fish were returned to system water and raised to either 1-month (4-weeks) of age or adulthood (3-4 months of age; Fig. 1) when they were tested using the optomotor response (OMR). All larvae were housed in petri dishes with daily water changes until 2 weeks of age, when they were placed into the AHAB recirculating system. One month old and adult fish were collected from these AHAB tanks. We maintained fish these fish in the recirculating system at densities of 12-15 fish per 1.5 L tank120. After recording OMRs, adult zebrafish were anesthetized in a 0.02% tricaine solution for several minutes until gill movements slowed, removed from the solution, blotted dry on a Kimwipe, and decapitated. Retinal tissue for RNA and protein extraction was collected after ERG recordings. Retinas were removed from dark-adapted eye cups, flash frozen and stored at −80 °C or placed in RNAlater™ Stabilization Solution (Invitrogen, Waltham, MA) at −20 °C. Whole eyes not used for RNA or protein extraction were subjected to electroretinogram recordings (ERGs). The remaining skeletal components of the head and the body trunk were flash frozen for LC/MS analysis. The above experimental pipeline was applied to fish acutely exposed as adults.
Optomotor responses (OMRs)
OMRs were elicited using a rotating black and white pinwheel stimulus projected onto a computer screen directly below the experimental container95,121. Animals were placed in the experimental container (a clear glass cylindrical tank) in pairs and allowed to acclimate for 5 min, during which time general swimming behavior was observed and baseline activity recorded. The pinwheel stimulus rotated in one direction (clockwise or counterclockwise) for 30 sec each, with a 30 sec control period of gray light shown in between. The stimulus sequence was run 2x for each fish pair and the swimming behavior/response of the fish recorded using a digital video camera (Canon Vixia HFR700). A positive response occurred when the fish swam in the direction of the stimulus. Recordings were performed in an isolation chamber to prevent the fish from responding to other cues. OMRs were scored by examination of video playback by two trained, blinded observers. For adults, the number of complete revolutions made while the stimulus was present was calculated. For 1-month old fish, scan sampling was used with the position of the fish determined every 6 sec when the stimulus was present121. Results are presented as the percentage of larval fish displaying a positive optomotor response.
Retinal anatomy
Immediately after behavioral and/or physiological assessment, a subset of exposed larvae (N = 3 per treatment for each exposure age and postexposure timepoint) were euthanized in 0.02% tricaine and fixed in 4% paraformaldehyde, followed by 70% ethanol, for anatomical analysis. Adult eyes to be analyzed were removed from euthanized adults and placed in 4% paraformaldehyde and then 70% ethanol.
Whole larvae (2-weeks and 4-weeks postexposure) were photographed using a Leica MZ10F stereo microscope equipped with a Leica DFC700T digital camera and LASX software. Larvae, or adult eyes were shipped, in ethanol, to Histoserv, Inc. for dehydration, paraffin embedding, sectioning, mounting, and staining with H&E (https://www.histoservinc.com/). Stained retinal layers were imaged in-house using an Olympus BX51 compound microscope with an infinity 1 Lumenera digital camera and Lumenera software. The sections with an optic nerve or most prominent lens were chosen for each larva. For adults, only retinal sections were measured.
Digital images of retinal sections were measured using Image J. Thickness of the different retinal layers in central retina were measured. Each measurement was made three times, then averaged, to reduce error.
Photopic electroretinograms (ERGs)
Removed eyes were placed scleral side down on a piece of filter paper. Using iris scissors (Fine Scientific Tools; Foster City, CA), eye cups were generated by removing the lens and cornea. The filter paper + eye was transferred to the recording chamber and superfused with oxygenated MEM solution (Millipore, Burlington, MA) equilibrated with 95% O2/5% CO2. The perfusion needle was placed near the eye with a perfusion rate of 0.3 ml/min. Once in the recording chamber, the eye cup was visualized and positioned using an IR camera (Teledyne QImaging, Surrey, British Columbia) mounted on an Olympus BX51WI compound microscope and Metamorph imaging software (Molecular Devices, San Jose, CA).
ERGs evoked by a white-light stimulus were recorded by inserting a tungsten recording electrode (FHC, Bowdoin, ME) directly into the vitreal chamber in the center of the eye cup. We selected excised eyecups, rather than recordings in intact fish, as the different ERG components in zebrafish have been cataloged using this preparation122. After dark adapting (10-20 min), the tissue was stimulated by 300 msec full-field white light pulses at 7 irradiance levels ranging from ND 3.0 (brightest) - 6.0 (dimmest) in 0.5 increments. Responses were amplified using a DAM80 amplifier (World Precision Instruments, Sarasota, FL) using a bandpass from 0.1 Hz to 1 kHz and a Digidata 1440 A (Axon Instruments, San Jose, CA) with a sampling rate of 2 kHz. Traces averaged 4× for each stimulus condition were boxcar filtered over a period corresponding to one cycle of 60 Hz line frequency. Mean response component amplitudes and peak times were obtained from the different treatment groups. Data were collected with pCLAMP 10 software (Axon Instruments) and analyzed using Origin 2021.
Zebrafish white-light ERG responses have clear ON-bipolar b-wave and OFF-bipolar d-wave components122,123 which were evoked on an infrared (IR, RG780 filter) background. The white-light protocol generated a response dataset with 70, 4-trace-average ERGs. These ERGs were in response to 10 replicates of a 7-step irradiance response series covering a 3-log unit range. This protocol was run 1-2 times per eyecup, each recording lasting 25 minutes, resulting in an overall minimum total of 20-180 ERGs at each brightness level generated from 2-18 eyes per treatment group. The white-light source was a 150 W Xenon arc lamp, as imaged through UV compliant optics, providing a flat energy spectrum across the visual sensitivity range of zebrafish from 650 nm to 370 nm. The maximum delivered irradiance (at 3.0 ND) measured at the objective was 1.8 µW/cm2 (Tektronix digital light meter).
Statistical comparisons across treatments were performed as previously described123. Specifically, the responses to the brightest stimulus (3.0 ND) from all white-light datasets were combined into a single, cumulative dataset for each treatment group. N = total number of ERG responses in the cumulative dataset. Amplitudes of b- and d-waves were measured as means within fixed, characteristic time intervals (pre-stimulus baselines set to ‘0’), which were selected to include b-wave or d-wave peaks for consistent measurements of response amplitude. ERG b-wave amplitude (µV) and peak times (ms) were measured in the interval 51-200 ms after stimulus onset. For the d-wave, amplitude and peak times were measured in the interval 25-250 ms after stimulus offset. In the absence of a peak during the interval time, no peak time was recorded.
Whole-cell patch clamp
Whole cell patch clamp techniques in zebrafish retinal slices were used to record voltage-gated currents of retinal bipolar cells, as in ref. 89. Briefly, adult zebrafish were anesthetized in 0.02% tricaine solution until gill movements slowed, decapitated, and eyes enucleated and placed vitreal side down on filter paper (0.45 µm pore size; Millipore). Retinal slices ( ~ 100 µm thick) were anchored in the recording chamber between Vaseline strips and covered with standard Ringer’s solution (120 mM NaCl, 2 mM KCl, 3 mM CaCl2, 1 mM MgCl2, 3 mM D-glucose, and 4 mM HEPES, adjusted to pH 7.4-7.5 with NaOH). Slices were viewed using an Olympus BX60 fixed stage microscope fitted with Hoffman Modulation Contrast optics and a 40x water immersion lens. Thin walled filamented borosilicate glass pipettes (World Precision Instruments) were pulled to the desired tip diameter ( ~ 1 µm; <10 MΩ) with a Flaming Brown P-80 micropipette puller (Sutter Instruments, Novato, CA) and filled with intracellular solution (12 mM KCl, 104 mM K-gluconate, 0.1 mM CaCl2, 4 mM HEPES, and 1 mM EDTA, adjusted to pH 7.4-7.5 with KOH). Bipolar cells were identified prior to recording by position within the mid/distal inner nuclear layer. Voltage-gated currents were elicited in response to voltage steps from −80 mV to +60 mV in 10 mV increments (Vhold = −60mV) using an Axopatch 1D patch clamp amplifier and pCLAMP 11 software (Axon Instruments). Traces/data were analyzed using Clampfit or OriginLab software.
RNA isolation and RT-qPCR
Total RNA was extracted from pooled retinal tissue from 2 eyes of a single adult fish (individual fish served as a biological replicate) using the RNeasy Plus Mini Kit (Qiagen, Germantown, MD) according to the manufacturer’s protocol for animal tissue. Briefly, approximately 25 mg of tissue was lysed and homogenized in Buffer RLT Plus containing 1% β-mercaptoethanol using an RNase-free microcentrifuge pestle. Lysates were passed through gDNA Eliminator spin columns, 1 volume of 70% ethanol was added to the flow-through, and samples were applied to RNeasy spin columns. A series of wash steps were performed, RNA was eluted with 40 µL of RNase-free water, and samples were stored at −80 °C. To assess the quality of total RNA preparations, diluted samples were visualized by gel electrophoresis on a 2% agarose gel stained with ethidium bromide to confirm the presence of 28S and 18S ribosomal RNA (rRNA) bands. Additionally, RNA yield and purity was quantified by measuring the concentration and A260/A280 absorbance ratios using a NanoVue Plus™ spectrophotometer (Biochrom, Holliston, MA). RNA samples with confirmed rRNA bands and A260/A280 ratios between 1.8 and 2.1, an indication of pure RNA, were selected for use in downstream applications.
Gene-specific primer pairs used in the study (Supplementary Table 1) were designed or confirmed from sequences established in the literature6,124,125 to span exon-exon junctions or large introns to avoid possible amplification of genomic DNA. Lyophilized primers were resuspended in TE buffer (1 M Tris, 0.5 M EDTA) upon arrival and diluted 1:10 in sterile water to create a 10 μM working solution.
The isolated RNA was used in quantitative reverse transcription PCR (RT-qPCR) to assess expression of genes listed in Supplementary Table 1. Reactions were performed with the qScript One-Step SYBR Green RT-qPCR, Low ROX Kit (Quantabio, Beverly, MA) in an AriaMx Real-time PCR System (Agilent, Savage, MD). Reactions were prepared in a 96-well plate at 10 µL total volume with 1X One-Step Master Mix, 200-400 nM forward and reverse primer, 20-35 ng total RNA diluted in sterile water, and 1X qScript One-Step Reverse Transcriptase. The thermal cycling conditions consisted of the following: cDNA synthesis for 10 minutes at 50 °C, initial denaturation and Taq Polymerase activation for 5 minutes at 95 °C, followed by 40 cycles of denaturation at 95 °C for 10 seconds and annealing/extension at 60 °C for 30 seconds. At the end of the cycling, a melt curve analysis was performed at 95 °C for 1 minute, 55 °C for 30 seconds, and 95 °C for 30 seconds to confirm primer specificity and absence of primer dimers.
The housekeeping gene (hkg) ribosomal protein L13A (rpl13a) was used as an endogenous control to normalize expression levels target genes. Rpl13a has been previously identified as the most stable hkg in zebrafish for chemical treatment studies using qPCR analysis126,127. All samples were run in technical triplicate and each developmental exposure group (i.e., adult, 72 hpf, 7 dpf exposure) were run on independent plates containing all treatments, tissue types, three genes of interest, and the hkg. The RT-qPCR design was replicated with biological replicates for each treatment condition, treating each plate as a statistical block128.
Ct values from technical replicates were averaged after calculating a coefficient of variation (CV) for each qPCR reaction to assess any inconsistency across replicates. If the CV was greater than 5%, the values were excluded from the analysis and the reaction was repeated. Results were presented with the comparative Ct method (∆∆CT) for relative quantification of gene expression. The ∆∆CT was transformed to 2-∆∆Ct to obtain the expression fold change for each treatment relative to the water control. The technical replicate mean ∆Ct was used to test statistical significance across biological replicates.
Western blots
Adult retinal tissue pooled from 1-3 fish (tissue from 2-6 eyes) were homogenized in Pierce™ RIPA Buffer (ThermoFisher) lysis reagent (1 µL/mg tissue) supplemented with 1X Halt™ Protease Inhibitor Cocktail (ThermoFisher) using a sterile microcentrifuge pestle and cordless pestle motor (VWR). Samples were incubated on ice for 35 min, centrifuged for 20 min at 4 °C at 12,000 rpm, and protein concentrations were quantified using the Pierce™ BCA Protein Assay Kit (ThermoFisher) according to the manufacturer’s microplate procedure protocol.
Proteins were diluted to 25 μg in Pierce™ RIPA Buffer lysis reagent with 1X Laemmli Sample Buffer (Bio-Rad) containing 10% β-mercaptoethanol and incubated for 10 min at 70 °C. Proteins from each treatment were loaded in technical duplicate, resolved by SDS-PAGE on NuPAGE™ 4 to 12%, Bis-Tris, 1.0 mm polyacrylamide gels (Invitrogen) and transferred to a PVDF membrane using the iBlot™ 2 Transfer Stacks, PVDF, mini and Gel Transfer Device (Invitrogen). Prior to antibody application, membranes were stained with Ponceau S to confirm successful transfer and equal loading of the protein.
Membranes were blocked in 5% non-fat dry milk with TBST for 30 min at room temperature and incubated overnight at 4 °C with primary antibodies (Supplementary Table 2) diluted in either 5% milk-TBST or 5% bovine serum albumin (BSA)-TBST for phospho-specific antibodies. The membrane was washed three times with TBST and incubated with either an anti-rabbit (Cell Signaling, 7074) or anti-mouse (Cell Signaling, 7076) horseradish peroxidase (HRP) conjugated secondary antibody diluted to 1:2000 for 50 min at room temperature. After an additional three TBST washes, protein was visualized with SuperSignal™ West Dura Extended Duration Chemiluminescent Substrate (ThermoFisher) reagents following the manufacturer’s instructions and imaged using a ChemiDoc-It® Imaging System (UVP, Upland, CA) with VisionWorks software. After visualization, excess chemiluminescent reagents were washed as before and the membrane was stripped, blocked, and re-probed with additional antibodies. β-actin primary antibody was applied to each membrane as a loading control. Western blots were quantified using densitometry analysis in Image J software and values were normalized to water controls prior to statistical analysis.
Liquid chromatography/mass spectroscopy (LC/MS)
To verify that a 24 hr exposure was sufficient for BPA uptake and to quantify BPA accumulation, light chromatography/mass spectroscopy (LC/MS) was performed on adult fish head/body carcasses prepared as described above.
Sample preparation
Fish wrapped in aluminum foil were removed from the freezer and thawed to room temperature. Individual fish were divided using a razor blade into 0.5 cm sections and added to a 40 mL glass tube along with 0.1 mL of DI water (18 MΩ-cm Milli-Q, EMD Millipore, Burlington, MA) whereupon tissues were homogenized into a paste using an Omni TH (Omni Int., Kennesaw, GA) tissue homogenizer. A 1 g aliquot (weighed precisely) of homogenate was placed in a 2 mL PP snap cap microtube. The tissue sample was spiked with the internal standard (IS, 125 ng of BPA-d6) and surrogate standard (SS) mix (125 ng each of 17β-estradiol-d5 and estrone-13C3) prior to the addition of 0.5 mL of acetonitrile (containing 1% formic acid). The mixture was vortexed for 20 s and placed in a 40 kHz Branson Ultrasonics bath for 30 min before adding 150 ng of MgSO4 + 38 mg CH3COONa + 0.5 mL of DI water. The mixture was vortexed for 20 s and re-sonicated for 15 min. The tube was centrifuged at 5000 rpm on a microcentrifuge for 5 min. The upper acetonitrile layer was transferred by glass pipette to a 2 mL dSPE tube containing 100 mg of PSA (UCT, Bristol, PA). Additional acetonitrile was added as needed (100-200 μL). The dSPE tube was vortexed for 20 s and placed in the ultrasonic bath for 15 min, after which the tube was centrifuged at 5000 rpm for 5 min. The supernatant was transferred by glass pipette to a 5 mL glass syringe and pushed via glass plunger through a pre-rinsed 13 mm 0.2 μm (pore dia.) PVDF syringe filter into a 1.5 mL amber autosampler vial. BPA was subsequently analyzed using LC-MS/MS.
LC-MS/MS analysis
BPA was quantified using a Shimadzu Model 8050 TQ liquid chromatograph-mass spectrometer (LC-MS/MS) configured with a SIL-20ACXR autosampler (Columbia, MD). The LC-MS/MS interface was operated in DUIS mode using both positive and negative ionization at a scan speed of 30,000 u/s at 0.1 u step size, coupled with polarity switching of 5 msec. LC-MS/MS separation of endocrine disrupting compounds (EDCs) was performed using a 50 mm × 2.1 mm (id), 1.8 μm particle (dia) Titan C18 reversed-phase UHPLC column (Supelco, Bellefonte, PA) in combination with a guard column equipped with a guard column insert containing C18 stationary phase complementary with the analytical column. A binary mobile phase was used consisting of DI water (solvent A), and acetonitrile (solvent B), both containing 5 mM ammonium acetate as a phase modifier. Operating conditions for the LC-MS/MS are listed in Supplementary Table 3.
LC-MS/MS quantitation of BPA was accomplished using the MRM mode and included 2 MRM ions, including quantifier (212.1 m/z) and confirmation (133.0 m/z) ions. The MRM ions were established through automated MRM optimization procedures following manual precursor ion identification using the full scan mode. Quantitation was performed using a 10-point internal calibration standard (ranging from 0.05 to 450 ng/mL) based on the primary MRM ion abundance of BPA to that of the associated IS (215.5 m/z). The tissue extracts were diluted as necessary with acetonitrile/DI water to fit the calibration range. Data analysis and quantitation was performed using LabSolutions software (ver. 5.91).
Quality assurance
Surrogate spike recoveries (N = 5) for 17β-estradiol-d5 and estrone-13C3 were 79 ± 18% and 91 ± 19% in tissues, respectively. Matrix recoveries were performed in BPA spiked tissues and averaged 104 ± 10%. The quantitation limit (QL) for BPA was 5.3 ng/g wet weight and was determined according to Eq. (1) as:
where Sy was the regression standard deviation at the y-intercept, m was the slope of the calibration curve and Ms was the sample mass (g) of tissue. Method blanks were prepared using vials and sorbents without tissues. BPA was not detected in the method blanks. Concentrations of BPA in samples are expressed as ng/g wet tissue weight.
All glassware used for sample storage and preparation were cleaned by washing with soap, rinsing with DI and fired at 400 °C overnight to ignite any interfering organic residues on surfaces that may have interfered with quantitative analysis. All laboratory materials were made of glass, stainless steel, or Teflon to minimize contamination. The Teflon materials were cleaned the same way as glass, but without firing. All non-glass items were rinsed with methanol and air dried before use.
Gas chromatography/mass spectrometry (GC/MS)
GC/MS was used to measure whether BPA accumulation was evident in adult carcasses that were developmentally exposed to BPA at ages 72 hpf or 7 dpf.
Sample preparation
Preparation of samples followed procedures was modified from129 which incorporated the QuEChERS method of130. In brief, fish tissue was homogenized (Omni tissue grinder) and 1 g (to 0.1 mg) of fish was added to a 40-ml glass centrifuge tube. Analytes were extracted with 15 ml of 1% (v/v) acetic acid in acetonitrile added to the 40 ml tube and shaken on vortex mixer for 1 min. Next, NaCl (0.2 g), anhydrous MgSO4 (0.3 g) and anhydrous Na2SO4 (1.7 g) were added to tube (as salting-out agents) and vortexed again for 1 minute. The tube was then centrifuged for 8 min at 3,400 rpm. The supernatant decanted into a new 40 ml centrifuge tube and MgSO4 (0.5 g), primary secondary amine (0.1 g) and C18 sorbents (0.4 g) added. The tube was centrifuged for 8 min at 3,400 rpm (again) and decanted into a 200 ml round bottom flask and 2 ml of toluene added for vapor exchange. The mix was rotavapped to 0.5 ml and analyzed using a GC/MS.
GC/MS analysis
Extract (1 ml) was injected immediately after rotovapping into a Thermo Trace GC/Polaris Q GC/MS containing a 30-m Restek Rxi-5ms fused silica column. Extracts from water blanks were also injected as were toluene blanks every 10th injection. Both blanks did not detect BPA. Standard curves were created using stock solutions of between 0.0002 and 0.00005 M 99% BPA and peak areas were used for determining concentration. The heating program used started at 50 °C, ramped at 10 °C/min to 250 °C and held at that temperature for 2 minutes. After injection, there was a 4-minute delay before the MS was activated to allow the solvent to pass.
Statistics and reproducibility
For each analysis performed, at least 2 biological replicates (fish or eyes) were used; typical samples sizes were 3-5 biological replicates. These values were chosen to be consistent with previous and published data and to allow statistical differences to be noted. Specific sample sizes are given in the text/captions. Statistical analyses of anatomical, ERG, patch clamp, and OMR data were performed using SPSS ver. 27 software (IBM Corporation), and graphs were generated in Excel or Origin. Statistical differences among treatments were assessed using either a parametric one-way ANOVA (anatomical measurements, ERGs) followed by a Tukey posthoc test, or a nonparametric Kruskal-Wallis (H) test (optomotor responses) followed by Dunn’s test to identify the source of the significance. Statistical analyses of RT-qPCR and immunoblotting data were assessed in RStudio ver. 1.3.1073 with one-way ANOVAs and Tukey post hoc tests. Significance for all tests was evaluated at an α-level of 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Data are available on Open Science Framework (https://osf.io/7r5fx/)131.
References
McCarthy, M. Estradiol and the developing brain. Physiol. Rev. 88, 91–127 (2008).
McEwen, B. & Alves, S. Estrogen actions in the central nervous system. Endocr. Rev. 20, 279–307 (1999).
Cascio, C., Deidda, I., Russo, D. & Guarneri, P. The estrogenic retina: the potential contribution to healthy aging and age-related neurodegenerative diseases in retina. Steroids 103, 31–41 (2015).
Campbell, C. et al. Biologically directed environmental nomitoring, fate, and transport of estrogenic endocrine disruptin compounds in water: a review. Chemosphere 65, 1265–1280 (2006).
Yilmaz, B., Terekeci, H., Sandal, S. & Kelestimur, F. Endocrine disrupting chemicals: exposure, effects on human health, mechanisms of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 21, 127–147 (2020).
Qiu, W. et al. Actions of bisphenol A and bisphenol S on the reproductive neuroendocrine system during early development in zebrafish. Endocrinology 157, 636–647 (2016).
Acconcia, F., Pallottini, V. & Marino, M. Molecular mechanisms of action of BPA. Dose Response 13, 1559325815610582 (2015).
Vasiljevic, T. & Harner, T. Bisphenol A and its analogues in outdoor and indoor air: properties, sources and global levels. Sci. Total Environ. 789, 148013 (2021).
Kim, S. et al. Neurochemical and behavioral analysis by acute exposure to bisphenol A in zebrafish larvae model. Chemosphere 239, 124751 (2020).
Chapin, R. et al. NTP-CERHR expert panel report on the reproductive and developmental toxicity of bisphenol A. Birth Defects Res. B 83, 157–395 (2008).
Corbel, T. et al. Bidirectional placental transfer of Bisphenol A and its mail metabolite, Bisphenol-A-glucuronide, in the isolated perfused human placenta. Reprod. Toxicol. 47, 51–58 (2014).
Ben-Jonathan, N. & Steinmetz, R. Xenoestrogens: the emerging story of bisphenol A. TEM 9, 124–128 (1998).
Corrales, J. et al. Global assessment of bisphenol A in the environment. Dose Response 13, 1559325815598308 (2015).
Cano-Nicolau, J. et al. Estrogenic effects of several BPA analogs in the developing zebrafish brain. Front. Neurosci. 10, 112 (2016).
dos Santos, B., Ivantsova, E., Guzman, A. & Martyniuk, C. Critical review of the toxicity mechanisms of bisphenol F in zebrafish (Danio rerio): knowledge gaps and future directions. Chemosphere 297, 134132 (2022).
Gu, J. et al. A systemtic comparison of neurotoxicity of bisphenol A and its derivates in zebrafish. Sci. Total Environ. 805, 150210 (2022).
Gould, J. et al. Bisphenol A interacts with the estrogen receptor alpha in a distinct manner from estradiol. Mol. Cell Endocrinol. 142, 203–214 (1998).
Pinto, C. et al. Differential activity of BPA, BPAF, and BPC on zebrafish estrogen receptors in vitro and in vivo. Toxicol. Appl. Pharm. 380, 114709 (2019).
Vandenberg, L. et al. Hormones and endocrine-disrupting chemicals: low-dose effects and nonmonotonic dose responses. Endocr. Rev. 33, 378–455 (2012).
Thomas, P. & Dong, J. Binding and activation of the seven-transmembrane estrogen receptor GPER30 by environmental estrogens: a potential novel mechanism of endocrine disruption. J. Steriod Biochem. Mol. Biol. 102, 175–179 (2006).
Huang, W. et al. Identification of receptors for eight endocrine disrupting chemicals and their underlying mechanisms using zebrafish as a model organism. Ecotoxicol. Environ. Saf. 204, 111068 (2020).
Mu, X. et al. Developmental effects and estrogenicity of bisphenol A alternatives in a zebrafis embryo model. Environ. Sci. Technol. 52, 3222–3231 (2018).
Pinto, C. et al. Selectivity of natural, synthetic and environmental estrogens for zebrafish estrogen receptors. Toxicol. Appl. Pharm. 280, 60–69 (2014).
Grontved, L. et al. Transcriptional activation oby thyroid hormone receptor through ligand-dependent receptor recruitment and chromatin remodeling. Nat. Commun. 6, 7048 (2015).
Volz, S., Poulsen, R., Hansen, M. & Holbech, J. Bisphenol A alters retinal morphology, visually guided behavior, and thyroid hormone levels in zebrafish larvae. Chemosphere 348, 140776 (2024).
Soriano, S. et al. Effects of bisphenol A on ion channels: experimental evidence and molecular mechanisms. Steroids 111, 12–20 (2016).
Nguyen, H., Jang, S., Park, S., Cho, D. & Han, S. Potentiation of the glycine response by bisphenol A, an endocrine disruptor, on the substantia gelatinosa neurons of the trigeminal subnucleus caudalis in mice. Chem. Res. Toxicol. 33, 782–786 (2020).
Zhong, X. et al. Rapid effect of bisphenol A on glutamate-induced Ca+2 influx in hippocampal neurons of rats. Mol. Cell Endocrinol. 485, 35–43 (2019).
Choi, I.-S. et al. Multiple effects of bisphenol A, and endocrine disruptor, on GABAA receptors in acutely dissociated rat CA3 pyramidal neurons. Neurosci. Res. 59, 8–17 (2007).
Inadera, H. Neurological effects of bisphenol A and its analogues. Int. J. Med. Sci. 12, 926–936 (2015).
Rubin, B. Bisphenol A: an endocrine disruptor with widespread exposure and multiple effects. J. Steriod Biochem. Mol. Biol. 127, 27–34 (2011).
Kim, M. et al. Exposure to bisphenol A appears to impair hippocampal neurogenesis and spatial learning and memory. Food Chem. Toxicol. 49, 3383–3389 (2011).
Wolstenholme, J. et al. Gestational exposure to bisphenol A produces transgenerational changes in behaviors and gene expression. Endocrinology 153, 3828–3838 (2012).
Kinch, C., Ibhazehiebo, K., Jeong, J.-H., Habibi, H. & Kurrasch, D. Low-dose exposure to bisphenol A and replacement bisphenol S induces precocious hypothalamic neurogenesis in embryonic zebrafish. Proc. Natl. Acad. Sci. USA 112, 1475–1480 (2015).
Weber, D., Hoffmann, R., Hoke, E. & Tanguay, R. Bisphenol A exposure during early development induces sex-specific changes in adult zebrafish social interactions. J. Toxicol. Environ. Health A 78, 50–66 (2015).
Saili, K. et al. Neurodevelopmental low-dose bisphenol A exposure leads to early life-stage hyperactivity and learning deficits in adult zebrafish. Toxicology 291, 83–92 (2012).
Martinez, R. et al. Dose-dependent transcriptomic responses of zebrafish eleutheroembryos to bisphenol A. Environ. Poll. 243, 988–997 (2018).
Li, X. et al. Impact of low-dose chronic exposure to bisphenol A (BPA) on adult male zebrafish adaptation to the environmental complexity: disturbing the color preference patterns and reliving the anxiety behavior. Chemosphere 186, 295–304 (2017b).
Gu, J. et al. Neurobehavioral effects of bisphenol S exposure in early life stages of zebrafish larvae (Danio rerio). Chemosphere 217, 629–635 (2019).
Qiu, L. et al. Mechanism of bisphenol S exposure on color sensitivity of zebrafish larvae. Environ. Pollut. 316, 120670 (2023).
Liu, W.-L. et al. Long-term exposure to bisphenol S damages the visual system and reduces the tracking capability of male zebrafish (Danio rerio). J. Appl. Toxicol. 38, 248–258 (2018).
Crowley-Perry, M., Barberio, A., Zeino, J., Winston, E. & Connaughton, V. Zebrafish optomotor response and morphology are altered by transient, developmental exposure to bisphenol-A. J. Dev. Biol. 9, 14 (2021).
Easter, S. & Nicola, G. The development of vision in the zebrafish (Danio rerio). Dev. Biol. 180, 646–663 (1996).
Schmitt, E. & Dowling, J. Early retinal development in the zebrafish, Danio rerio: ligth and electron microscopic analysis. J. Comp. Neurol. 404, 515–536 (1999).
Stuermer, C. Retinotopic organizaiton of the developing retinotectal projection in the zebrafish embryo. J. Neurosci. 8, 4513–4530 (1988).
Mouriec, K. et al. Early regulation of brain aromatase (cyp19a1b) by estrogen receptors during zebrafish development. Dev. Dyn. 238, 2641–2651 (2009b).
Le Page, Y., Vosges, M., Servili, A., Brion, F. & Kah, O. Neuroendocrine effects of endocrine disruptors in teleost fish. J. Toxicol. Environ. Health B 14, 370–386 (2011).
Canesi, L. & Fabbri, E. Environmental effects of BPA: focus on aquatic species. Dose Response 2015, 1–14 (2015).
Chen, Q.-L. et al. Enhanced uptake of BPA in the presence of nanoplastics can lead to neurotoxic effects in adult zebrafish. Sci. Total Environ. 609, 1312–1321 (2017).
Chen, T.-H., Wang, Y.-H. & Wu, Y.-H. Developmental exposures to ethanol or dimethylsulfoxide at low concentrations alter locomotor activity in larval zebrafish: implications for behavioral toxicity bioassays. Aquat. Toxicol. 102, 162–166 (2011).
Christou, M., Kavaliauskis, A., Ropstad, E. & Fraser, T. DMSO effects larval zebrafish (Danio rerio) behavior, with additive and interaction effects when combined with positive controls. Sci. Total Environ. 709, 134490 (2020).
Hallare, A., Nagel, K., Kohler, H.-R. & Triebskorn, R. Comparative embryotoxicity and proteotoxicity of three carrier solvents of zebrafish (Danio rerio) embryos. Ecotoxicol. Environ. Saf. 63, 378–388 (2006).
Lyssimachou, A., Jenssen, B. & Arukwe, A. Brain cytochrome P450 aromatase gene isoforms and activity levels in Atlantic Salmon after waterborne exposure to nominal environmental concentrations of the pharmaceutical ethynylestradiol and antifoulant tributylin. Toxicol. Sci. 91, 82–92 (2006).
Mortensen, A. & Arukwe, A. Dimethylsulfoxide is a potent modulator of estrogen receptor isoforms and xenoestrogen biomarker responses in primary culture of salmon hepatocytes. Aquat. Toxicol. 79, 99–103 (2006).
Pena-Corona, S. et al. Effect of a single application of coumestrol and/or dimethyl sulfoxide, on sex hormone levels and vaginal cytology of anestrus bitches. Veterinaria Mexico OA 6, 1–15 (2019).
Moriyama, K. et al. Thyroid hormone action is disrupted by bisphenol A as an antagonist. J. Clin. Endocrinol. Metab. 87, 5185–5190 (2002).
Sheng, Z.-G. et al. Low concentrations of bisphenol A suppress thyroid hormone receptor transcription through a nongemonic mechanism. Toxicol. Appl. Pharm. 259, 133–142 (2012).
Volkov, L. et al. Thyroid hormone receptors mediate two distinct mechanisms of long-wavelength vision. Proc. Natl. Acad. Sci. USA 117, 15262–12569 (2020).
Roberts, M. R., Srinivas, M., Forrest, D., Morreale de Escobar, G. & Reh, T. A. Making the gradient: thyroid hormone regulates cone opsin expression in the developing mouse retina. Proc. Natl. Acad. Sci. USA 103, 6218–6223 (2006).
Nadal, A. et al. Nongenomic actions of estrogens and xenoestrogens by binding at a plasma membrane receptor unrelated to estrogen receptor alpha and estrogen receptor beta. Proc. Natl. Acad. Sci.USA 97, 11603–11608 (2000).
Fuentes, N. & Silveyra, P. Chapter three—Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 116, 135–170 (2019).
Stefkovich, M., Arao, Y., Hamilton, K. & Korach, K. Experimental models for evaluating non-genomic estrogen signaling. Steroids 133, 34–37 (2019).
Masuya, T., Iwamoto, M., Liu, X. & Matsushima, A. Discovery of novel oestrogen receptor alpha agonists and antagonists by screening a revisited previleged structure moiety for nuclear receptors. Sci. Rep. 9, 9954 (2019).
Iwamoto, M. et al. Bisphenol A derivatives act as novel coactivator-binding inhibitors for estrogen receptor beta. J. Biol. Chem. 297, 101173 (2021).
Rice, D. & Barone, S. Jr Critical periods of vulnerability for the developing nervous system: evidence from humans and animal models. Environ. Health Perspect. 108, 511–533 (2000).
Ribeiro, E., Ladeira, C. & Viegas, S. Occupational exposure to bisphenol A (BPA): a reality that still needs to be unveiled. Toxics 5, 5030022 (2017).
Gibert, Y. et al. Bisphenol A induced otolith malformations during vertebrate embryogenesis. BMC Dev. Biol. 11, 4 (2011).
Froehlicher, M. et al. Estrogen receptor subtype B2 is involved in neuromast development in zebrafish (Danio rerio) larvae. Dev. Biol. 330, 32–43 (2009).
Scaramella, C. et al. Bisphenol A exposure induces sensory processing deficits in larvae zebrafish during development. eNEURO 9, 1–13 (2022).
Menuet, A. et al. Molecular characterization of three estrogen receptor forms in zebrafish: binding characteristics, transactivation properties, and tissue distributions. Biol. Reprod. 66, 1881–1892 (2002).
Chandrasekar, G., Archer, A., Gustafsson, J.-A. & Lendahl, M. Levels of 17B-estradiol receptors expressed in embryonic and adult zebrafish following in vivo treatment of natural or synthetic ligands. PLoS ONE 5, e9678 (2010).
Le Fol, V. et al. In vitro and in vivo estrogenic activity of BPA, BPF and BPS in zebrafish-specific assays. Ecotoxicol. Environ. Saf. 142, 150–156 (2017).
Celeghin, A. et al. The knockdown of maternal estrogen receptor 2a (esr2a) mRNA affects embryo transcript contents and larval development in zebrafish. Gen. Comp. Endocrinol. 172, 120–129 (2011).
Chen, Y. et al. Interaction of nuclear ERs and GPER in vitellogenesis in zebrafish. J. Steriod Biochem. Mol. Biol. 189, 10–18 (2019).
Wu, C.-C. et al. The phenotypic and transcriptomic effects of developmental exposure to nanomolar levels of estrone and bisphenol A in zebrafish. Sci. Total Environ. 757, 143736 (2021).
Chung, E., Genco, M., Megrelis, L. & Ruderman, J. Effects of bisphenol A and triclocarban on brain-specific expression of aromatase in early zebrafish embryos. Proc. Natl. Acad. Sci. USA 108, 17732–17737 (2011).
Gertz, J., Reddy, T., Varley, K., Garabedian, M. & Myers, R. Genistein and bisphenol A exposure cause estrogen receptor 1 to bind thousands of sites in a cell type-specific manner. Genome Res. 22, 2153–2162 (2012).
Marino, M., Pellegrini, M., Rosa, P. L. & Acconcia, F. Susceptibility of estrogen receptor rapid responses to xenoestrogens: physiological outcomes. Steroids 77, 910–917 (2012).
Shanle, E. & Xu, W. Endocrine disrupting chemicals targeting estrogen receptor signaling: identification and mechanisms of action. Chem. Res. Toxicol. 24, 6–19 (2011).
Cotter, K., Yershov, A., Novillo, A. & Callard, G. Multiple structurally distinct ERa mRNA variants in zebrafish are differentially expressed by tissue type, stage of development and estrogen exposure. Gen. Comp. Endocrinol. 194, 217–229 (2013).
Sanchez, M., Picard, N., Sauve, K. & Tremblay, A. Challenging estrogen receptor B with phosphorylation. Trends Endocrinol. Metab. 21, 104–110 (2009).
Ascenzi, P., Bocedi, A. & Marino, M. Structure-function relationship of estrogen receptor a and b: impact on human health. Mol. Asp. Med. 27, 299–402 (2006).
Guo, Y.-J. et al. ERK/MAPK signalling pathway and tumorigenesis (review). Exp. Ther. Med. 19, 1997–2007 (2020).
Kim, E. & Choi, E.-J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 1802, 396–405 (2010).
Naumann, E. et al. From whole-brain data to functional circuit models: the zebrafish optomotor response. Cell 167, 947–960 (2016).
Portugues, R. & Engert, F. The neural basis of visual behaviors in the larval zebrafish. Curr. Opin. Neurobiol. 19, 644–647 (2009).
Lindholst, C., Wynne, P., Marriott, P., Pedersen, S. & Bjerregaard, P. Metabolism of bisphenol A in zebrafish (Danio rerio) and rainbow trout (Oncorhynchus mykiss) in relation to estrogenic response. Comp. Biochem. Physiol. C135, 169–177 (2003).
Subbamanda, Y. & Bhargava, A. Intercommunication between voltage-gated calcium channels and estrogen receptor/estrogen signaling: insights into physiological and pathological conditions. Cells 11, 3850 (2022).
Connaughton, V. & Maguire, G. Differential expression of voltage-gated K+ and Ca+2 currents in bipolar cells in the zebrafish retinal slice. Eur. J. Neurosci. 10, 1350–1362 (1998).
Connaughton, V., Nelson, R. & Bender, A. Electrophysiological evidence of GABAA and GABAC receptors on zebrafish retinal bipolar cells. Vis. Neurosci. 25, 139–153 (2008).
Goel, M. & Mangel, S. Dopamine-mediated circadian and ligh/dark-adaptive modulation of chemical and electrical synapses in outer retina. Front. Cell Neurosci. 15, 647541 (2021).
Yu, C. & Li, L. Dopamine modulates voltage-activated potassium currents in zebrafish retinal ON bipolar cells. J. Neurosci. Res. 82, 368–376 (2005).
Korshunov, K., Blakemore, L. & Trombley, P. Illuminating and sniffing out the neuromodulatory roles of dopamine in the retinal and olfactory bulb. Front. Cell Neurosci. 14, 275 (2020).
Hamad, A., Kluk, M., Fox, J., Park, M. & Turner, J. The effects of aromatase inhibitors and selective estrogen receptor modulators on eye development in the zebrafish (Danio rerio). Curr. Eye Res. 32, 819–827 (2007).
Gould, C., Wiegand, J. & Connaughton, V. Acute developmental exposure to 4-hydroxyandrostenedione has a long term effect on visually-guided behaviors. Neurotoxicol. Teratol. 64, 45–49 (2017).
Gorelick, D., Iwanowicz, L., Hung, A., Blazer, V. & Halpern, M. Transgenic zebrafish reveal tissue-specific differences in estrogen signalling in response to environmental water samples. Environ. Health Perspect. 122, 356–362 (2014).
Hyatt, G. et al. Retinoic acid establishes ventral retinal characteristics. Development 122, 195–204 (1996).
Hyatt, G., Schmitt, E., Fadool, J. & Dowling, J. Retinoic acid alters photoreceptor development in vivo. Proc. Natl. Acad. Sci. USA 93, 13298–13303 (1996).
Deveau, C. et al. Thyroid hormone receptor beta mutations alter photoreceptor development and function in Danio rerio (zebrafish). PLoS Genet. 16, e1008869 (2020).
Nelson, R. et al. The developmental progression of eight opsin spectral signals recorded from the zebrafish retinal cone layer is altered by the timing and cell type expression of thyroxin receptor B2 (TRB2) gain-of-function transgenes. eNEURO 9, 1–25 (2022).
Darras, V., Van Herc, S., Heijlen, M. & De Groef, B. Thyroid hormone receptors in two model species for vertebrate embryonic development: chicken and zebrafish. J. Thyroid Res. 2011, 402320 (2011).
Marelli, F. et al. Patterns of thyroid hormone receptor expression in zebrafish and generation of a novel model of resistance to thyroid hormone action. Mol. Cell Endocrinol. 424, 102–117 (2016).
Vancamp, P., Houbrechts, A. & Darras, V. Insights from zebrafish deficiency models to understand the impact of local thyroid hormone regulator action on early development. Gen. Comp. Endocrinol. 279, 45–52 (2019).
Kim, M. & Park, Y. Bisphenols and thyroid hormone. Endocrinol. Metab. 34, 340–348 (2019).
Hammes, S. & Davis, P. Overlapping nongemonic and genomic actions of thyroid hormone and steroids. Best. Pr. Res. Clin. Endocrinol. Metab. 29, 581–593 (2015).
Koutaki, D., Paltoglou, G., Vourdoumpa, A. & Charmandari, E. The impact of bisphenol A on thyroid function in neonates and children: a systematic review of the literature. Nutrients 14, 168 (2022).
Baumann, L., Ros, A., Rehberger, K., Neuhauss, S. & Segner, H. Thyroid disruption in zebrafish (Danio rerio) larvae: different molecular response patterns lead to impaired eye development and visual functions. Aquat. Toxicol. 172, 44–55 (2016).
Pannetier, P. et al. Reversibility of thyroid hormone system-disrupting effects on eye and thyroid follicle development in zebrafish (Danio rerio) embryos. Environ. Toxicol. Chem. 42, 1276–1292 (2023).
Baumann, L., Segner, H., Ros, A., Knapen, D. & Vergauwen, L. Thyroid hormone disruptors interfere with molecular pathways of eye development and function in zebrafish. Int. J. Mol. Sci. 20, 1543 (2019).
Liang, L. et al. Bisphenol A causes reproductive toxicity, decreases DNMT1 transcription, and reduces global DNA methylation in breeding zebrafish (Danio rerio). Epigenetics 11, 526–538 (2016).
Liu, Y. et al. Global DNA methylation in gonads of adult zebrafish (Danio rerio) under bisphenol A exposure. Ecotoxicol. Environ. Saf. 130, 124–132 (2016).
Torres, T., Ruivo, R. & Santos, M. Epigenetic biomarkers as tools for chemical hazard assessment: gene expression profiling using the model Danio rerio. Sci. Total Environ. 773, 144830 (2021).
Yu, H., Song, W., Chen, X., Zhao, Q. & Du, X. Transcriptomic analysis reveals up-regulated histone genes may play a key role in zebrafish embryo-larvae response to bisphenol A (BPA) exposure. Ecotoxicol. Environ. Saf. 252, 114578 (2023).
Qin, T. et al. Epigenetic alteration shaped by the environmental chemical bisphenol A. Front. Genet. 11, 618966 (2021).
Pena-Corona, S. et al. Administration of coumestrol and/or dimethyl sulfoxide affects vaginal epithelium and sex hormones in female dogs. Ind. J. Anim. Res. 6, 1–15 (2022).
Lyssimachou, A. & Arukwe, A. Alteration of brain and interrenal StAR protein, P450scc, and Cyp11B mRNA levels in Atlantic salmon after nominal waterborne exposure to the synthetic pharmaceutical estrogen ethynylestradiol. J. Toxicol. Environ. Health A 70, 606–613 (2007).
Hallare, A., Kohler, H.-R. & Triebskorn, R. Developmental toxicity and stress protein responses in zebrafish embryos after exposure to diclofenac and its solvent, DMSO. Chemosphere 56, 659–666 (2004).
Neuhauss, S. Behavioral genetic approaches to visual system development and function in zebrafish. J. Neurobiol. 54, 146–160 (2003).
Kolpin, D., Furlong, E., Meer, M., Thurman, E. & Zaugg, S. Pharmaceuticals, hormones, and other organic wastewater contaminants in US streams, 1999-2000: a national reconnaissance. Environ. Sci. Technol. 36, 1202–11 (2002).
Castranova, D. et al. The effect of stocking densities on reproductive performance of laboratory zebrafish (Danio rerio). Zebrafish 8, 141–146 (2011).
LeFauve, M. & Connaughton, V. Impacts of heavy metal exposure alters visually-guided behaviors in zebrafish. Curr. Zool. 63, 221–227 (2017).
Nelson, R. & Singla, N. A spectral model for signal elements isolated from zebrafish photopic electroretinogram. Vis. Neurosci. 26, 349–363 (2009).
Tanvir, Z., Nelson, R., DeCicco-Skinner, K. & Connaughton, V. One month of hyperglycemia alters spectral responses of the zebrafish photopic electroretinogram. Dis. Models Mech. 11, dmm035220 (2018).
Lazcano, I. et al. Knock-down of specific thyroid hormone receptor isoforms impairs body plan development in zebrafish. Front. Endocrinol. 10, 156 (2019).
Jeffery, J. et al. Cep55 regulates embryonic growth and development by promoting Akt stability in zebrafish. FASEB J. 29, 1999–2009 (2015).
Tang, R., Dodd, A., Lai, D., McNabb, W. & Love, D. Validation of zebrafish (Danio rerio) reference genes for quantitative real-time RT-PCR normalization. Acta Biochim. Biophys Sin. 39, 384–390 (2007).
Xu, H. et al. Genome-wide identification of suitable zebrafish Danio rerio reference genes for normalization of gene expression data by RT-qPCR. J. Fish. Biol. 88, 2095–2110 (2016).
Rieu, I. Real-time quantitative RT-PCR: design, calculations, and statistics. Plant Cell 21, 1031–1033 (2009).
Arya, G. et al. Pharmaceutical chemicals, steroids and xenoestrogens in water, sediments and fish from the tidal freshwater Potomac River (Virginia, USA). J. Environ. Sci. Health A 52, 7 (2017).
Lehotay, S. et al. Fast easy multiresidue method employing acetonitrile extraction partitioning and dispersive solid phase extraction for determination of pesticide residues in produce. J. AOAC Int. 85, 1148–1166 (2022).
Cohen, A. et al. Source data—Persistent molecular effects of developmental BPA exposure [data set]. Open Science Framework, https://doi.org/10.17605/OSF.IO/7R5FX (2025).
Acknowledgements
The work was supported by NEI R15EY029866-01 and a NASA DC Space Grant (to V.P.C.).
Author information
Authors and Affiliations
Contributions
A.C. contributed molecular (qPCR and Western) data and its analysis, wrote the original manuscript, participated in revision of manuscript drafts; A.S. contributed behavioral data; J.S.J. contributed physiological data; R.H. and E.B. contributed anatomical data; G.F. contributed LC/LC-MS data; S.M. contributed GC-MS data; M.D.-P. contributed larval data; V.P.C. contributed experimental design, data analysis, oversight of all experimental personnel and protocols, edited original and revised versions of the manuscript, and obtained funding for the project.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Biology thanks Arash Salahinejad and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Christina Karlsson Rosenthal.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Cohen, A., Sherffius, A., Jensen, J.S. et al. Loss of adult visual responses by developmental BPA exposure is correlated with altered estrogenic signaling. Commun Biol 8, 847 (2025). https://doi.org/10.1038/s42003-025-08245-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s42003-025-08245-y
