
Abstract
Placental dysfunction is linked to neurodevelopmental disorders, with males showing greater vulnerability to perinatal inflammation-mediated brain injuries. Using our transgenic mouse model, Akr1c14cyp19aKO (plKO), we investigate how reduced placental allopregnanolone (ALLO), an anti-inflammatory neurosteroid, contributes to sex-specific brain injury. plKO mice display sex-divergent cerebellar myelination and male-specific autism-like behaviors. Here we show that placental ALLO insufficiency triggers sex-divergent neuroinflammatory responses and microglial dysfunction. Sex-divergent differential expression of inflammatory genes and distinct inflammatory cytokine/chemokine patterns are seen in the placenta and the brain. Prostaglandin E2 (PGE2)-EP4 signaling is identified as a key regulator and, consistent with male plKO cerebellar hypermyelination, male microglial myelin phagocytosis is impaired by SIRPα-CD47 signaling changes. Postnatal manipulation of these critical pathways can normalize cerebellar myelin content and rescue abnormal behavior in male plKO mice. Sex-divergent microglial dysfunction and prostaglandin signaling drive male-biased neurodevelopmental impairments in our model, suggesting new therapeutic targets to improve brain development following placental dysfunction.
Similar content being viewed by others

Introduction
Many neurodevelopmental disorders exhibit sex bias in frequency, severity, and phenotypic presentation1, but the mechanisms by which sex influences brain development are not well understood. Placental dysfunction, affecting over 10% of pregnancies, can severely impact fetal neurodevelopment, leaving many infants vulnerable to long-term cognitive and motor deficits2,3,4. The endocrine function of the placenta may be a key driver of developmental differences, both through local, placental regulation as well as via long-range hormonal signaling in the developing fetal brain. Our approach is to identify placentally produced hormones that shape fetal and long-term brain development so that they can be targeted for replacement. ALLO is one such hormone, produced from progesterone in large quantities by the placenta during the second half of gestation5,6. It is a positive allosteric modulator of the GABAA receptor that regulates a variety of neurodevelopmental processes, such as neurogenesis and myelination7,8,9. ALLO also exhibits non-glucocorticoid10 anti-inflammatory properties by inhibiting toll-like receptors (TLRs) and regulating cytokine expression11,12,13,14,15,16,17,18. Placental ALLO may maintain the immune balance of the intrauterine environment and protect the fetus against exogenous stressors6,19,20.
To directly investigate the consequences of reduced placental ALLO on brain development, we created a transgenic mouse model, termed plKO, in which the gene Akr1c14, which encodes the key ALLO synthesis enzyme 3α-hydroxysteroid dehydrogenase (3α-HSD), is specifically ablated in placental trophoblast cells expressing Cyp19a-cre21, resulting in a significant 50% reduction in placental ALLO production5. The plKO model primarily serves as a tool to investigate the fundamental role of placental ALLO during neurodevelopment while also modeling clinical conditions where placental ALLO production is compromised22,23. ALLO’s role in supporting healthy pregnancies has been established, with reduced ALLO levels linked to conditions such as preeclampsia22 and intrauterine growth restriction23. In animal models, ALLO levels rise in response to prenatal stressors such as umbilical cord occlusion24 and result in potentiated cell death and myelination defects when pharmacologically inhibited6. Assessment of our plKO mice revealed broad but regionally specific sex differences across brain regions. In the cerebellum (CB), males display an increased rate of oligodendrocyte (OL) maturation and thicker myelin, while females show the converse5. These divergent cerebellar phenotypes were paralleled by sex-specific neurobehavioral impairments in plKO mice, with males exhibiting autistic-like behavioral traits, including reduced sociability and increased stereotyped behaviors5. Female plKOs show different neurodevelopmental impairments, including female-predominant reduction in pyramidal neuron populations in the primary somatosensory cortex (S1) and accompanying deficits in novel object recognition25.
Since ALLO production by the placenta peaks in late gestation, it is an attractive neuroprotective target for corrective treatment following pregnancies with adverse outcomes. However, to develop effective interventions, there is an increased need to understand the complex interplay between placental pathology, sex, and neurological disease. The current study aims to decipher the mechanisms driving the sex-dependent neurodevelopment trajectories of the plKO brain in the postnatal period.
In this work, we show that placental ALLO insufficiency triggers sex-specific neuroinflammatory responses and microglial dysfunction. These alterations contribute to divergent myelination patterns (hypermyelination in males, hypomyelination in females) and male-specific autism-like behavioral outcomes, highlighting the critical role of placental hormones in neurodevelopment.
Results
Placental ALLO reduction alters inflammatory pathways in the postnatal brain
We conducted RNA-sequencing (RNA-seq) analyses of the primary somatosensory cortex (S1), hippocampus (HP), and cerebellum (CB) at postnatal day 0, 7, 15, and 30 (P0, P7, P15, P30) on both male and female plKO and littermate controls (C) to characterize the regional, developmental and sex effects of placental ALLO reduction. Limma-Voom with duplicate correlation was used to account for multivariate data and align samples across regions from the same mouse8,26,27,28 (Supplementary Fig. 1a–c). After analyzing the significantly differentially expressed genes (DEGs) using Ingenuity Pathway Analysis (IPA, Qiagen), we identified common dysregulated canonical pathways across the brain: phagosome formation and S100 Family signaling. The CB showed the most significant expression changes of the regions examined, as indicated by its gene-set z-scores (Supplementary Fig. 1d), in parallel with the CB sex-specific myelination and behavioral alterations previously described in plKO mice5.
To define the molecular mechanisms underlying sex-specific cerebellar phenotypes, we analyzed the interaction between sex and genotype in CB differential gene expression across postnatal development. This analysis revealed a set of dysregulated pathways related to neuroinflammation (Fig. 1a). The specific gene lists were examined to identify the common target molecules within each pathway (Fig. 1b). For example, pathogen-induced cytokine storm pathway primarily contained genes in the interleukin, cytokine, and chemokine families, while the multiple sclerosis signaling pathway included inflammatory genes known to impact myelination and remyelination (Lif, Mbp, and Calpain gene family29,30) (Fig. 1b). Several toll-like receptor (Tlr) genes were also common to the identified pathways. TLRs are well-established contributors to anti-inflammatory signaling, including in ALLO’s anti-inflammatory activity11,12. The dynamic changes in the number of inflammation-related DEGs observed across postnatal development (Fig. 1c) suggest that decreased placental ALLO exposure changes the developmental neuroinflammatory profile in the developing CB.

a IPA analysis of differentially expressed genes (DEGs) comparing genotype and sex across postnatal development. The pathways were ranked by greatest –log(p-value). A right-tailed Fisher’s Exact Test was used with a cutoff of p < 0.05 and a 30% log2-fold change. Among the top 20 brain-related pathways identified, there was a prominent pattern of inflammation-related pathways. P0 (red), P7 (green), P15 (blue), and P30 (purple). b Diagram highlighting common genes in the canonical pathways identified by IPA. c Number of inflammation-associated DEGs (p-value < 0.05 and a 30% log2-fold change; (GO:0006954, “inflammatory response”; release 2022-07-01)) in the CB across postnatal development. Males in blue, females in red. Source data are provided as a Source Data file.
plKO cerebellum shows sexually divergent inflammatory profiles across the lifespan
The neuroinflammatory CB profiles in male and female mice were assayed, with a specific focus on the inflammatory molecules suggested by the expression changes in inflammation-associated genes in the postnatal plKO CB (Fig. 1). Key cytokines and chemokines from the RNA-seq (Fig. 1b, red; IL-11, LIF, and IL-12p70) as well as additional inflammatory molecules were measured by multiplex assay. Distinct inflammatory patterns were seen across CB development (Fig. 2a, b; Supplementary Fig. 2a–j). Pro- and anti-inflammatory indices were calculated using z-standardized concentrations of pro- (MCP-1, MIP1a, MIP3b, IP-10, and IL-12p70) and anti-inflammatory (6Ckine, IL-11, IL-9, m-CSF, and LIF) groups, respectively (Fig. 2a, b). At P30, plKO females showed a relative increase in anti-inflammatory cytokines compared to control females and plKO males (Fig. 2b). This finding demonstrates that, unlike males, females mount a more dynamic response to placental ALLO insufficiency, characterized by increased anti-inflammatory factors in the CB over a month after the initial insult.

a, b Cytokine and chemokine levels were measured by multiplex assay in whole cerebellar tissue across postnatal development. Data represent the means of z-normalized cytokine levels classified as pro-inflammatory (MCP-1, MIP1a, MIP3b, IP-10, and IL-12p70) or anti-inflammatory (6Ckine, IL-11, IL-9, m-CSF, and LIF). Data are presented as mean ± s.e.m. The number of independent biological replicates for each experiment is listed in the Supplementary Data File. Statistical analysis was performed using a three-way ANOVA comparing male and female cytokine values across genotype and age, followed by Tukey’s multiple comparison test. ∗p < 0.05, ∗∗p < 0.01 for genotype pairwise comparisons. For anti-inflammatory cytokines: P30 plKO Male vs. P30 plKO Female (p = 0.027); P30 C Female vs. P30 plKO Female (p = 0.008). Source data are provided as a Source Data file.
Placental inflammatory response to ALLO reduction differs by sex
Postnatal inflammatory phenotypes in the plKO CB could arise from prenatal or postnatal events, specifically from the placenta, which is the site of genetic manipulation in this model. To assess inflammatory cytokines at embryonic day (E) 15.5, coinciding with the most significant loss of Akr1c14 gene expression in the placenta5, paired embryonic brain and placenta tissues were collected. Protein level measurements of key inflammatory markers revealed a complex interplay of cytokine changes in both embryonic tissues. Placental ALLO insufficiency resulted in an increase in pro-inflammatory cytokines in both placenta and brain, with a larger increase in females overall (Fig. 3). IL-6 and IP-10 are unique as they show alterations across both sexes in both embryonic tissues (Fig. 3a, b). Other cytokines display responses that differ in either predominantly sex- or tissue-specific ways. Placental inflammatory responses are characterized by sex-divergent levels of TNFα and IL-5 production, but no similar divergence is seen in embryonic brain (Fig. 3c, d). Female embryonic plKO brain exhibits a larger response with increased levels of IL-17, IL-7, LIF, Eotaxin, and MCP-1 (Fig. 3e–i). Conversely, plKO males show a smaller response with dysregulated levels of IL-17, MCP-1, and M-CSF (Fig. 3e, i, j). To confirm that these sex-specific differences were due to ALLO loss and not ALLO-mediated changes in sex steroid hormone levels, we performed mass spectrometry, which revealed no significant alterations in estradiol or testosterone levels in plKO tissues despite the previously measured 50% decrease in ALLO5 (Supplementary Table 1). These findings demonstrate distinct inflammatory responses in both placenta and embryonic brain following placental ALLO insufficiency, with a particularly pronounced increase in pro-inflammatory cytokines in females.

a–j Inflammatory cytokine levels in paired placenta and embryonic brain at E15.5 were measured by multiplex assays. Box plots are presented with the median, interquartile range (25th–75th percentiles), and whiskers extending to the minimum and maximum values. Statistical analysis was performed using two-way ANOVAs comparing sex and genotype for each tissue, followed by Sidak multiple comparisons tests. C Male (n = 5), plKO Male (n = 7); C Female (n = 8), plKO Female (n = 5). # p < 0.05 for sex by genotype effect. ‡p < 0.05, ‡‡p < 0.01 for overall genotype effect. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 for genotype pairwise comparisons. Brain: IL-6: Genotype effect (p = 0.0050); C Female vs. plKO ( p= 0.0168). IP-10: Genotype effect (p = 0.0187). TNF-α: Sex and Genotype interaction effect ( p= 0.0218). IL-17: C Female vs. plKO (p = 0.0002). IL-7: C Female vs. plKO ( p= 0.0123). LIF: C Female vs. plKO (p = 0.0024). Eotaxin: C Female vs. plKO (P = 0.0092). MCP-1: Genotype effect (p = 0.0206). M-CSF: C Male vs. plKO (p = 0.0395). Placenta: IL-6: Sex vs. Genotype interaction (p = 0.0261); C Female vs. plKO (p = 0.0182). IL-5: Sex vs. Genotype interaction (p = 0.0185). Source data are provided as a Source Data file.
plKO mice show sex- and age-specific alterations in cerebellar white matter microglia
To begin to untangle the effects of this altered cytokine milieu in males and females, we examined the state and function of the local CB microglia. Microglia, the resident immune cells of the brain, play crucial roles in regulating inflammation, influencing brain sex differentiation, and orchestrating developmental pruning across multiple cell components, including myelin31. Microglial priming may also bridge the temporal gap between the initial embryonic insult and the postnatal plKO inflammatory signature32. Analysis of the microglial distribution throughout the different cerebellar layers revealed a significant effect of sex and genotype in the developing white matter (Fig. 4a, b). Specifically, the density of microglia in the cerebellar white matter was significantly decreased in female plKO at P7 and in male plKO mice at P30, compared to their littermate controls (Fig. 4c). Heatmaps of microglia-enriched genes across CB development showed striking differences by genotype and sex, beginning in the first postnatal week and persisting through adolescence (P30) in parallel to these changes in microglial density (Supplementary Fig. 3a–c). These data reveal a complex pattern of microglial alterations following decreased placental ALLO, with microglial reduction in the CB of plKO mice, with females showing an early decrease and males showing a later decrease.

a, b Representative parasagittal sections of control and plKO P30 male CB immunostained for IBA1 imaged by confocal microscopy. Scale bar: 200 μm. a’, b’ Zoomed images of cerebellar white matter microglia at P30 immunostained for IBA1 imaged by confocal microscopy. Scale bar: 40 μm. c Quantification of IBA1+ cell densities in the cerebellar white matter (WM). Data are presented as mean ± s.e.m. Statistical analysis was performed using a two-way ANOVA for sex and genotype effects, followed by Sidak multiple comparisons tests. Male: C P7 (n = 4), P15 (n = 3), P30 (n = 4); plKO: P7 (n = 4), P15 (n = 3), P30 (n = 4). Female: C P7 (n = 4), P15 (n = 3), P30 (n = 5); plKO: P7 (n = 3), P15 (n = 4), P30 (n = 4). ∗p < 0.05 for genotype pairwise comparison. P30: Sex vs. Genotype Interaction (p = 0.0247); C Female vs. plKO Female (p= 0.0414); C Male vs. plKO Male ( p= 0.0442). d–g Representative cleared whole-brain light-sheet microscopy images. Section planes are indicated by dash-dot lines and labeled i (coronal section through SSC and CC), i’ (coronal section through CB), and i” (sagittal section including SSC, CC, HP, and CB). YoPro nuclear stain is yellow, and IBA1+ microglia are blue. Scale bars: d–f (1.7 mm), g (3 mm). h, i”, Heatmaps showing the average IBA1+ cell density differences between plKO and C brains in P30 males on coronal sections i and i’ and sagittal sections i”. j Quantification of IBA1+ cell densities in selected brain regions. Data are presented as mean ± s.e.m. Statistical comparisons were performed using multiple two-sided unpaired t-tests with Welch correction (n = 5 C and n = 6 plKO brains). ∗∗∗FDR(qvalue) <0.005 (q = 0.003196). The experiment was repeated twice with independent samples. CB cerebellum, CC corpus callosum, HP hippocampus, SSC somatosensory cortex. Source data are provided as a Source Data file.
To determine whether changes in microglial density were specific to the CB, whole-brain IBA1 immunofluorescence analysis using light-sheet microscopy on cleared P30 male brains was used. This confirmed the significant reduction of IBA1+ cell density in the CB of plKO male mice (Fig. 4d–j). In contrast, other brain regions, such as the somatosensory cortex (SSC), hippocampus (HP), and corpus callosum (CC), did not show significant differences in IBA+ cell density, indicating that placental ALLO insufficiency results in region-specific effects.
plKO mice show sex and genotype-associated differences in microglial phagocytosis machinery
Microglia developmentally regulate myelination through phagocytosis of oligodendrocytes and myelin33. Given the changes in CB myelination that occur in postnatal plKO mice, we assessed the phagocytic capacity of cerebellar microglia in C and plKO mice ex vivo. CD11b+ fluorescence-activated cell sorting (FACS) was used to isolate microglia from the cerebella, and phagocytosis was measured using pH-sensitive pHrodo-conjugated, inactivated, unopsonized E. coli. Increased pHrodo fluorescence (pHrodoHigh) indicates phagocytosis with lysosomal acidification. At P15, both male and female plKO mice displayed a transient genotype-based increase in pHrodoHigh microglia (Fig. 5a, b; Supplementary Fig. 4a–d), suggesting increased, but not necessarily functional or appropriate, phagocytosis.

a Representative gating for FACS of CD11b and pHrodo fluorescence intensity. Thresholds used to categorize High versus Low pHrodo cells are shown. b Percent of pHrodoHigh CD11b+ cells in males and females across development. Data are presented as mean ± s.e.m. Statistical analysis was performed using two-way ANOVAs for sex and genotype, followed by Sidak multiple comparison tests. Male: Control P7 (n = 7), P15 (n = 8), P30 (n = 6); plKO P7 (n = 8), P15 (n = 8), P30 (n = 7). Female: Control P7 (n = 7), P15 (n = 7), P30 (n = 9); plKO P7 (n = 8), P15 (n = 7), P30 (n = 8). ‡‡p < 0.01 for the overall genotype effect (P = 0.0048). c Heatmap representing log2-fold expression changes in a curated list of phagocytosis-related genes determined by high-throughput RT-qPCR on sorted microglia. Statistical comparisons were performed using two-sided, Welch’s multiple t-tests between genotypes at each age. Results are presented as log2-fold change (Log2FC). The number of independent biological replicates for each experiment is listed in the Supplementary Data File. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001. d RT-qPCR for Sirpa expression in sorted microglia. Data are presented as mean ± s.e.m. Statistical analysis was performed using two-way ANOVAs for sex and genotype, followed by Sidak multiple comparison tests. Male: Control P7 (n = 7), P15 (n = 8), P30 (n = 6); plKO P7 (n = 8), P15 (n = 8), P30 (n = 7). Female: C P7 (n = 7), P15 (n = 7), P30 (n = 9); plKO P7 (n = 8), P15 (n = 7), P30 (n = 8). ∗∗p < 0.01 for the pairwise comparison (p = 0.0092). Source data are provided as a Source Data file.
To characterize the molecular mechanisms underlying altered phagocytosis suggested by the shift in pHrodoHigh microglia, gene expression analysis on sorted CD11b+ microglia was done, focusing on genes regulating phagocytic targeting, engulfment, and lysosome acidification. Consistent with the FACS data, phagocytosis-associated gene expression was most significantly altered at P15, particularly in males. While many genes involved in phagosome acidification, such as Lamp1, Ctsd, and Cd68, exhibited sustained expression elevation, many of the genes critical to microglial phagocytosis, such as Axl, Mertk, Tyrobp, and Lgals1/3, showed a decrease in expression by P15 in male plKO (Fig. 5c).
We further analyzed the expression of Sirpa, a key gene that inhibits microglial phagocytosis by preventing necessary cytoskeletal rearrangement in CD11b+ cells34,35. At P15, a significant upregulation of Sirpa expression was observed in male plKO microglia compared to female (Fig. 5d). Microglial SIRPα binds to CD47 on phagocytic targets and inhibits all other “eat me” signals necessary for phagocytosis34, suggesting impairment of the phagocytic machinery in plKO microglia predominantly in males despite the overall increase in pHrodoHigh microglia.
Male plKO microglia engulf fewer myelin components
Based on these changes in phagocytic machinery, we hypothesized that the phagocytosis of myelin by plKO microglia would be altered in a sex-divergent manner, contributing to the excess of plKO cerebellar myelin in males. Given that myelin proteins, such as myelin basic protein (MBP), are translated locally36, we reasoned that microglia actively engulfing myelin would contain detectable levels of oligodendrocyte-produced Mbp mRNA. In male plKO microglia, we observed a significant decrease in Mbp transcript detectable in isolated microglia at P15 and P30 (Fig. 6a), congruent with the period in which phagocytosis genes were most dysregulated (Fig. 5c). In contrast, female plKO microglia showed a trend towards an increase in Mbp mRNA within sorted microglia at P7 (p = 0.0671). When combining all sorted microglial samples, independent of age, linear regression analysis demonstrated a significant negative correlation between Sirpa gene expression and Mbp mRNA content in microglia (Fig. 6b). This is consistent with our hypothesis that SIRPα expression plays a role in myelin uptake, with higher expression of Sirpa associated with less engulfed myelin (Fig. 6c).

a Normalized RT-qPCR data of Mbp mRNA (to Actb, beta actin; Fth1, ferritin heavy chain 1) found within sorted microglia. Data are presented as mean fold change ± s.e.m. Statistical analysis was performed using two-way ANOVAs for sex and genotype, followed by Sidak multiple comparison tests. Male: C P7 (n = 6), P15 (n = 8), P30 (n = 4); plKO P7 (n = 7), P15 (n = 6), P30 (n = 6). Female: C P7 (n = 7), P15 (n = 7), P30 (n = 7); plKO P7 (n = 8), P15 (n = 5), P30 (n = 8). ∗p < 0.05 (P15 C Male vs. plKO Male (p= 0.0371); P30 C Male vs. plKO Male (p= 0.0293)). b Two-sided Pearson R correlation analysis between Sirpa mRNA levels and corresponding Mbp mRNA transcripts in microglia from all studied samples (all sexes, genotypes, and ages included). n = 76 samples. c Schematic of SIRPα function in binding CD47 and overriding phagocytic signals. SIRPα on the surface of microglia is proposed to bind to CD47 on myelin and inhibit the necessary cytoskeletal rearrangement that promotes phagocytosis34. Created in BioRender. https://BioRender.com/c91i906. Adapted from “CD47- Targeting in Cancer Immunotherapy.” Retrieved from https://app.biorender.com/biorender-templates. d Representative images of cells from the imaging flow cytometer. Cells were imaged at 60× magnification. e Quantification of the flow cytometry imaging in P15 males for internalized MBP within CD11b+ microglia (n = 4 C, n = 6 plKO independent mice). Data are presented as mean % cells ± s.e.m. Statistical analysis was performed using a two-sided Mann–Whitney test. f Quantification of the co-localization of MBP with the lysosomal marker, LAMP1 (lysosomal-associated membrane protein 1). Data are presented as mean % cells ± s.e.m. (n = 4 C, n = 6 plKO independent mice). g Quantification of the flow cytometry imaging in P7 females for internalized MBP stain within CD11b+ microglia. Data are presented as mean % cells ± s.e.m. C (n = 5) and plKO (n = 4) were compared using a two-sided Mann–Whitney test. ∗p < 0.05 (p = 0.0333). h Percent pHrodoHigh CD11b+ microglia following 7-day treatment with RRx-001 (10 mg/kg/day in 5% DMSO and sesame oil; P7-P14; s.c.) compared to plKO administered vehicle. Data are presented as mean ± s.e.m. One-way ANOVA for treatment with Tukey’s multiple comparison tests. Male: C (+ Veh) (n = 11), plKO (+ Veh) (n = 12), plKO + RRx-001 (n = 8). Female: C (+ Veh) (n = 11), plKO (+ Veh) (n = 10), plKO + RRx-001 (n = 7). ∗∗∗p < 0.001. Male plKO (+ Veh) vs. plKO + RRx-001 (p = 0.0002); Female plKO (+veh) vs. plKO + RRx-001 (p = 0.0002). i Mbp mRNA within sorted CD11b+ microglia following 7-day treatment with RRx-001 compared to vehicle-administered plKO mice. Data are presented as mean ± s.e.m. Statistical analysis was performed using a two-way ANOVA, followed by Sidak multiple comparison tests. Male: plKO (+ Veh) (n = 6), plKO + RRx-001 (n = 6). Female: plKO (+ Veh) (n = 4), plKO + RRx-001 (n = 7). ∗p < 0.05. Female plKO (+ Veh) vs. plKO + RRx-001 (p = 0.0368). j CD47 concentration (pg/mg of total protein) in purified cerebellar myelin measured by ELISA. Data are presented as mean levels ± s.e.m. Statistical analysis was performed using a two-way ANOVA for genotype and sex, followed by Sidak multiple comparison tests. Male: C (n = 6), plKO (n = 6). Female: C (n = 6), plKO (n = 3). ∗∗p < 0.01. plKO Male vs. plKO Female (p = 0.0097). Source data are provided as a Source Data file.
To validate the decreased myelin internalization by male plKO microglia, we conducted imaging flow cytometry on male plKO and C microglia at P15 (Fig. 6d, Supplementary Fig. 4e–i), the age when male plKO Mbp mRNA internalization was most altered. MBP staining within CD11b+ cells showed a strong trend towards less internalized myelin in plKO males at P15 compared to littermate controls (p = 0.0571) (Fig. 6e). Co-localization of LAMP1 and MBP revealed that myelin fragments within microglia were primarily in lysosomes, confirming microglial engulfment of myelin for subsequent degradation (Fig. 6f). Since female plKO showed changes in internalized Mbp mRNA at P7, we repeated this experiment with P7 females, which confirmed a significant increase in internalized MBP protein inside CD11b+ microglia compared to control females (Fig. 6g). There was minimal co-localization with LAMP1 at this age, suggesting that the observed phagocytic activity is not strongly associated with lysosomal processing of MBP at this time. This pattern mimics our Mbp mRNA data that showed decreased Mbp transcripts in male plKO microglia at P15 and a trend towards increased Mbp mRNA in female plKO microglia at P7, reinforcing the link between Mbp mRNA levels and myelin internalization. These sex differences in myelin component uptake may underlie the previously reported sex-divergent myelination changes in this model5.
To assess the inhibitory role of the SIRPα-CD47 axis on myelin phagocytosis, we treated plKO mice with the small molecule inhibitor, RRx-001, previously reported to improve macrophage phagocytosis by reducing SIRPα and CD47 expression37. Replication of our pHrodo flow cytometry experiment showed that RRx-001 treatment significantly elevated levels of phagocytic microglia (Fig. 6h), suggesting that inhibiting the SIRPα-CD47 axis enhances overall phagocytic activity. However, reassessment of Mbp mRNA levels within sorted microglia following RRx-001 treatment revealed an increase in engulfed Mbp mRNA in females only (Fig. 6i). The increased general phagocytic activity in male plKO microglia without a corresponding increase in Mbp mRNA suggests that the reduced myelin uptake in males is not solely due to increased microglial SIRPα expression.
The failure of RRX-001 treatment to increase the uptake of myelin in plKO males despite increased phagocytosis raises the possibility of a myelin-specific defect that renders it less susceptible to engulfment. The SIRPα “don’t eat me” signal on microglia relies on the expression of its ligand, CD47, on myelin to inhibit phagocytosis34. To determine whether CD47 levels were altered in cerebellar myelin, we performed a CD47 ELISA on purified myelin. This revealed a significant increase of CD47 in plKO male myelin compared to plKO female myelin (Fig. 6j). This demonstrates that while all plKO mice show a genotype-based increase in microglial phagocytosis, the male-specific upregulation of the SIRPa-CD47 axis may contribute to the disruption of the necessary crosstalk for myelin targeting and engulfment. Overall, combining this myelin phagocytosis data with the pHrodo analysis suggests that the type of material being targeted for phagocytosis may be different between the sexes at different timepoints, resulting in sex-dependent differences in brain architecture.
Cerebellar PGE2-EP4 receptor modulation influences sex-associated inflammation, myelination, and ASD-like behaviors in plKO mice
The sex differences in the inflammatory milieu and microglial functioning of cerebellar plKO could be regulated by common upstream mechanisms. To explore potential mediators of these divergent developmental trajectories, we returned to our RNA-seq IPA analysis to investigate upstream regulators and identified Ptger4, Cx3cl1, and Trem2, genes with well-recognized roles in inflammation, particularly concerning microglial pathology38,39,40. The most consistent upstream regulator candidate was the prostaglandin E2 (PGE2) receptor EP4 (Ptger4). Prostaglandins are critical in microglial sexual differentiation, particularly in the CB, where PGE2 promotes estradiol synthesis40,41. The EP4 receptor, highly expressed in microglia, reduces inflammatory stimuli and promotes microglial phagocytosis42. Comparing female plKO CB to male plKO CB transcriptomes, IPA projections indicated a dynamic sex-specific pattern of Ptger4 pathway activation: higher in males at P0 and P30 (positive z-scores) and higher in females at P7 and P15 (negative z-scores) (Fig. 7a). Direct measurement of cerebellar PGE2 levels via ELISA revealed reduced levels at P15 in male plKOs and increased levels in females (Fig. 7b). Embryonic (E15.5) plKO brain showed an increase in PGE2 in both sexes with no accompanying changes in the placenta (Supplementary Fig. 5a, b). This suggests the divergence in PGE2 signaling begins postnatally along with cerebellar sex-linked differentiation.

a Z-score plot for gene families regulated by Ptger4 at P0, P7, P15, and P30 comparing plKO Male (positive z-score) to plKO Female (negative z-score). b PGE2 levels in the cerebellum at P7, P15 and P30 measured by ELISA. Data are presented as mean concentration ± s.e.m. Statistical analysis was performed using two-way ANOVAs for sex and genotype, followed by Sidak multiple comparison tests. Male: CP7 (n = 7), P15 (n = 8), P30 (n = 8); plKO P7 (n = 8), P15 (n = 8), P30 (n = 6). Female: C P7 (n = 9), P15 (n = 8), P30 (n = 7); plKO P7 (n = 7), P15 (n = 7), P30 (n = 9). #p < 0.05 for the sex by genotype effect (p = 0.0173). c Schematic representation of L-902,688 treatment on cytokine expression at P30 following 6-day injection series of PGE2 agonist, L-902,688 (1 mg/kg/day; P7–P12; in saline; s.c.). Created in BioRender. https://BioRender.com/uto032r. d–h L-902,688 treatment on cytokine expression at P30 following 6-day injection series of PGE2 agonist assessed by multiplex assay. Data are presented as mean ± s.e.m. The number of independent biological replicates for each experiment is listed in the Supplementary Data File. Statistical analysis was performed using two-way ANOVAs comparing sex and genotype, followed by Sidak multiple comparisons tests. ‡p < 0.05, genotype effect; *p < 0.05, **p < 0.01, pairwise comparison. IFN-β1: Male plKO + Veh vs. plKO + L-902,688 (p = 0.0023); Female plKO + Veh vs. plKO + L-902,688 (p = 0.0248). IFNγ: Genotype effect (p = 0.0253). IL-1α: Genotype effect (p = 0.0438). IL-10: Genotype effect (p = 0.0150). M-CSF: Male plKO + Veh vs. plKO + L-902,688 (p = 0.0041); Female plKO + Veh vs. plKO + L-902,688 (p= 0.0033). Source data are provided as a Source Data file.
To determine if manipulating the PGE2 receptor could modify the sex-specific inflammatory profiles and their associated phenotypes in plKO mice, we investigated cytokine levels following treatment with the selective EP4 agonist, L-902,688. Mice were treated from P7 to P14, a previously identified sensitive period for sex-specific responses to inflammation in the CB43,44. Following L-902,688 treatment, P30 plKO mice displayed genotype-specific cytokine alterations, characterized by a significant increase in pro-inflammatory IFNγ and IL-1α and a decrease in anti-inflammatory IFNβ1, IL-10, and M-CSF, relative to vehicle-injected controls (Fig. 7c–h).
Next, we studied the potential of L-902,688 treatment to mitigate the plKO male-specific hypermyelination and ASD-like behaviors. Cerebellar MBP contents determined by ELISA were significantly decreased following L-902,688 treatment in P30 plKO males and females (Fig. 8a). Evaluating the effect of L-902,688 treatment on the male plKO autism-like behaviors5, revealed complete normalization of autism composite scores at P30 (Fig. 8b) which include sociability deficits and spontaneous stereotypical behaviors (e.g., gnawing and digging) (Fig. 8d). Female plKO, with previously reported low levels of CB MBP and no accompanying ASD-like behaviors, were unaffected by pharmacological increase in EP4 signaling (Fig. 8c, e). In summary, increased plKO myelination in male CB and ASD-like behaviors were associated with decreased PGE2 levels, while plKO females with decreased myelination exhibited increased levels of PGE2. By increasing EP4 signaling via L-902,688 treatment, MBP levels were reduced independent of sex, and male ASD-like behaviors were normalized. These results support the role of EP4 signaling in the sex divergent myelination anomalies in plKO mice, suggest that excess myelination, as measured by MBP levels, may impair specific behaviors, and that both EP4/PGE2 and MBP levels may be biomarkers for abnormal neurodevelopment.

a Cerebellar MBP concentration at P30 following L-902,688 treatment (1 mg/kg/day; P7–P12; in saline; s.c.) measured by ELISA. Data are presented as mean concentration ± s.e.m. Statistical analysis was performed using a two-way ANOVA comparing sex and genotype, followed by Sidak multiple comparison tests. Male: plKO + Veh (n = 9), plKO + L-902,688 (n = 10). Female: plKO + Veh (n = 9), plKO + L-902,688 (n = 5). ‡p < 0.05 for the genotype effect (p = 0.0397). ASD composite severity score in males (b) and females (c) at P30 following a 6-day injection series of PGE2 agonist, L-902,688. Data are presented as mean ± s.e.m. One-way ANOVA with Tukey’s multiple comparisons test. Male: C = 20, plKO = 41, plKO+ L-902,688 = 7, Female:C = 15, plKO = 11, plKO+ L-902,688= 6. *p < 0.05, ****p < 0.0001 (plKO vs plKO+ L-902,688 p = 0.023). z-standardized single significant behavior readouts that are integrated into the autism composite score in males (d) and females (e) at P30 following a 6-day injection series of L-902,688. Data are presented as mean ± s.e.m. Statistical analysis was performed using a one-way ANOVA with Tukey’s multiple comparisons test. Male: C (n = 20), plKO (n = 41), plKO + L-902,688 (n = 7). Female: C (n = 15), plKO (n = 11), plKO + L-902,688 (n = 6). Sociability: C vs. plKO (p < 0.0001), plKO vs. plKO+L-902,688 (p = 0.0153). Gnawing: C vs. plKO (p < 0.0001), plKO vs. plKO+ L-902,688 (p = 0.0153). Digging: C vs. plKO (p < 0.0001), plKO vs. plKO+ L-902,688 (p = 0.0153). Independent mice per group. In the “plKO (+Veh)” group, mixed populations from non-injected and saline-injected animals were pooled as they showed no significant behavioral differences. C and plKO group scores were published in Vacher et al.5. Source data are provided as a Source Data file.
Discussion
The sexually divergent neuroanatomical and behavioral phenotypes seen in offspring following insufficient placental ALLO exposure, particularly in the CB, are striking. In the experiments described here, we have identified changes in neuroinflammatory microglial cells and brain cytokine levels that are key contributors to these sex-dependent alterations.
The placenta plays a critical role in regulating the intrauterine immunological state during pregnancy. Placental trophoblasts secrete cytokines and chemokines that maintain a balance between the relative immunosuppression needed for semi-allogeneic fetal development and an active immune response against potential intrauterine infections45,46,47. Maternal infection, placental inflammation, and elevated levels of pro-inflammatory cytokines are associated with neonatal brain damage and neurodevelopmental disorders3. Pregnancy pathologies like preeclampsia and preterm delivery are also associated with alterations in placental immune status48,49,50,51. High serum levels of IL-6 increase the risk of ASD in children, as well as in rodent models of maternal immune activation52,53,54. Our study suggests that normal placental ALLO production is important in maintaining this immunological balance and may help to prevent prenatal inflammation-associated neurodevelopmental disorders, especially for male offspring. Our study provides strong evidence that non-infection-related inflammation can be generated, both in the placenta and the brain, by specific placental hormone insufficiency. These placental changes can then impact the inflammatory milieu of the developing brain through altered microglia function, leading to long-term behavioral alteration. Further investigation is needed to address whether the acute inflammatory response originates primarily in the placenta following a decrease in local ALLO exposure with subsequent transmission of inflammatory signals to the developing brain, or if both the placenta and CB respond directly to the loss of placental ALLO.
Prior studies have shown that under normal conditions, female rodents and humans have less inflammatory signaling in the developing brain, while inflammation in the male brain increases during the process of brain masculinization55. We hypothesized that decreased placental ALLO exposure would further increase these sex-specific developmental differences in the CB. Indeed, insufficient placental ALLO resulted in increased anti-inflammatory cytokines in postnatal females, while male plKO cytokine/chemokine levels showed no such response. Many studies have suggested that exposure to placental inflammation brings male brains closer to a threshold of vulnerability that can be easily crossed if inflammation occurs during a sensitive developmental period, translating into a higher risk of neurodevelopmental disorders56,57. Our findings demonstrate that altered placental endocrine support can trigger distinct and opposing inflammatory responses in the developing male and female CB. This work points to a previously unrecognized mechanism of placental endocrine influence on the sexually dimorphic bias in neurodevelopmental disorders.
Microglia not only mediate and regulate neuroinflammation but also play a key role in shaping the sex differences in the brain31. Clusters of microglia are found in the developing white matter, where they secrete factors regulating OL lineage cell survival, maturation, and apoptosis39,58,59,60. They also regulate myelin production by phagocytosing excess OLs and improperly formed myelin membranes. Our flow cytometry results indicate elevated phagocytic activity in the microglia of plKO CB, accompanied by a sex difference with reduced internalization of myelin components in male plKOs and an increase in female plKO microglia. However, our experiments did not address whether degenerative demyelination contributes to the sex differences. The differential response to the SIRPα-CD47 inhibitor, RRx-001, further supports a significant microglial sex difference. RRx-001 treatment increased microglial phagocytosis in both sexes, but only the females exhibited an associated increase in Mbp mRNA engulfment. SIRPα signaling relies on the expression of CD47 on microglial phagocytic targets, in this case on myelin, and the failure of RRx-001 to restore male myelin phagocytosis suggests male plKO myelin defects. The increased CD47 expression measured on purified myelin from male plKO mice is consistent with this interpretation. Both male and female plKO mice respond to the altered postnatal inflammatory landscape by increasing microglial phagocytic activity; however, upregulated SIRPα-CD47 signaling in male plKO prevents effective targeting of myelin by microglia. This divergent signaling in plKO microglia is likely to contribute to excess CB myelination in males and less in females in this model5.
Our study further identified prostaglandins, specifically the PGE2-EP4 signaling pathway, as mediators of sex-specific inflammatory responses and altered behavioral outcomes following placental ALLO loss. The EP4 receptor is highly expressed in microglia and typically restricts their response to inflammatory stimuli61. The rise in cerebellar PGE2 in plKO females precedes the increase in anti-inflammatory cytokine profile, consistent with previous reports of cerebellar PGE2 signaling contributing to downstream changes in cytokine secretion and in sexually dimorphic behavior41. PGE2 promotes estradiol synthesis in the CB, particularly during the first two postnatal weeks of development when the myelination deficits develop in our mice44,62. Here, we show that activating EP4 signaling in male plKO mice during this developmental window prevents the onset of ASD-like behaviors. Moreover, EP4 agonist treatment altered cytokine responses and reduced overall MBP levels in the CB. This implicates impaired EP4 prostaglandin signaling in the expression of male-specific neurobehavioral alterations and suggests a new pathway that could be therapeutically targeted to reduce ASD-like outcomes. Ultimately, our findings underscore the need for further investigation into mechanisms underlying sex differences in neurodevelopmental disorders to better tailor therapeutic interventions following pregnancy complications.
In conclusion, our study provides compelling evidence that placental ALLO insufficiency induces sex-specific alterations in microglial function, leading to divergent cerebellar myelination patterns in males and females (Fig. 9). We have demonstrated that this placental endocrine dysfunction alters neuroinflammatory pathways and microglial phagocytic activity in a sex-dependent manner, with significant implications for neurodevelopmental outcomes. Together, these results establish a critical link between placental hormone production, microglial function, and sex-biased neurodevelopmental trajectories, advancing our understanding of how prenatal factors may contribute to the sex bias observed in conditions such as autism spectrum disorder. This work highlights the importance of considering sex as a biological variable in both basic neurodevelopmental research and clinical approaches to neurodevelopmental disorders. Future studies are needed to explore how exogenous inflammatory challenges interact with ALLO insufficiency, potentially identifying novel therapeutic targets for preventing or mitigating neurodevelopmental impairments in high-risk pregnancies.

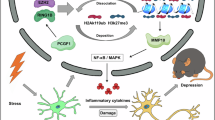
Embryonically, placental ALLO is reduced in our plKO mouse model, resulting in unique inflammatory signatures in the male and female placenta and brain. By postnatal day 7, the cerebellar phenotype of plKO males and females begins to diverge, with females exhibiting increased engulfment of myelin components. At P15, plKO microglial phagocytosis is increased in both sexes. Despite this increase, myelin phagocytosis by microglia is impaired in males via disrupted SIRPα-CD47 signaling. In parallel, PGE2 signaling begins to diverge at P15 with associated cytokine alterations. By P30, plKO males exhibit a pro-inflammatory profile, hypermyelination, and ASD-like behavior, while females show an anti-inflammatory profile, reduced myelination, and lack similar behavioral alterations. Created in BioRender. https://BioRender.com/5gs7mkd.
Methods
Materials
Mice. All the procedures on experimental animals were performed in accordance with the protocol approved by the Institutional Animal Care and Use Committee at Columbia University Medical Center (protocol nos. AC-AABE 6553 (PI: Penn) and AC-AABF 5550 (PI: Yang)). Mice were housed in a climate-controlled facility on a 12-h light/dark cycle and had ad libitum access to food and water. The Akr1c14-floxed mouse was constructed with inGenious Targeting Laboratories as previously described5. The Cyp19-Cre mouse line(#5912 line), expressing Cre exclusively in trophoblast cells, was obtained from Dr. Gustavo Leone, Ohio State University. Akr1c14fl/fl/Cre+ dams were bred with homozygous Akr1c14fl/fl/Cre- males, yielding mixed litters of plKO mice and littermate controls. Model validation was previously described5.
Drug injections
To pharmacologically test the contribution of PGE2 EP4 on plKO sex-specific behavioral phenotype, the PGE2-EP4 agonist, L-902,688 (Cayman Chemicals: Cat #10007712), was ordered in a special formulation of neat solid. It was then reconstituted in 0.9% Sodium Chloride. Pups were injected subcutaneously with either 1 mg/kg of drug or saline vehicle, once daily from P7 to P12. To pharmacologically test SIRPα-CD47 signaling, RRx-001 (MedChemExpress: Cat #925206-65-1) was suspended in 5% DMSO and sesame oil. Mice were injected once a day subcutaneously at 10 mg/kg from P7-P14.
Methods
RNA-sequencing
From each mouse, the S1, the HP, and CB were collected and stored in −80 °C for RNA extraction. RNA-sequencing was conducted by New York Genome Center using Illumina TruSeq Stranded mRNA Library prep (Illumina, catalog number 20020595) for mRNA library preparation in accordance with manufacturer recommendations, and using IDT for Illumina TruSeq DNA UD Indices (Illumina, catalog number 20022370) for adapters. A total of 500 ng of RNA underwent purification and mRNA fragmentation. The purified mRNA underwent both first and second-strand cDNA synthesis. Subsequently, the cDNA was adenylated, ligated to Illumina sequencing adapters, and subjected to PCR amplification (8 cycles). Quantification of the cDNA libraries was performed using the Fragment Analyzer 5300 kit FA-NGS-HS (Agilent, catalog number DNF-474-1000) and Spectramax M2 kit Picogreen (Life Technologies, catalog number P7589). The libraries were sequenced on an Illumina NovaSeq sequencer, employing 2 × 100 bp cycles. rRNA abundance was measured by mapping with Bowtie2, and quality control was performed using Picard (v1.83) and RSeQC (v2.6.1). The reads were aligned with STAR (version 2.5.2a), and genes annotated in Gencode vM16 and quantified with featureCounts (v1.4.3-p1). For the analysis, Quantro was used to test for global differences in distributions, and smooth quantile (qsmooth) normalization was used63,64. Differential expression was assessed using Limma-Voom with duplicate correlation26,27,28. Duplicate correlation was used to adjust for the brain region from the same mouse27. The transformed counts were fitted to a linear model, and Empirical Bayes statistics were used to generate DEG lists with log fold change and p-values. DEG lists from each group comparison were then entered into IPA (Qiagen) for pathway analysis. Cutoffs for DEGs were set to Log2-Fold Change of ±20% (downregulated −0.514537 and upregulated 0.378512) and a p-value of <0.05. The sample size was n = 4 for all groups, with the following exceptions: n = 5 for the plKO Female P7 S1 and HP, as well as for the C Female P15 CB groups. For the plKO Female P15 S1 and all plKO Male P7 groups, the n = 3. Additionally, the hippocampus was not collected at the P0 time point.
RT-PCR/ biomark experiments
For whole CB experiments, tissues were homogenized in TRIzol Reagent (Thermo Fisher Scientific) and total RNA was extracted with the RNeasy Mini Kit (Qiagen, no. 74104) according to manufacturer instructions. To extract RNA from sorted microglia cells were sorted into TRIzol Reagent and mixed with 400 µm of phenol:chloroform, pH 4.5 (Invitrogen, no. AM9722) for phase-separation. After a 15-min centrifugation at 20,000 × g, the top aqueous phase was transferred and mixed with 450 µm of isopropanol and 2 µl of GlycoBlue (Thermo Fisher no. AM9515) for coprecipitation. After a 10-min incubation, samples are centrifuged at 12,000 × g for 1 h. RNA was washed twice with 75% ethanol, and the pellet was resuspended in 15 µm of water. For all RT-PCR experiments, 1 µg of RNA for whole CB or 300 µg microglia samples was used to make cDNA with the iScript cDNA Synthesis Kit (Bio-Rad, no. 1708891). In whole CB samples, RT-PCR experiment was performed on cDNA samples using PowerUp SYBR Green Master Mix (Thermo Fisher Scientific, no. 1725271) with specific primers at 100 nM using the CFX96 Touch Real-Time PCR Detection System (Bio-Rad). For sorted microglia, high-throughput RT-PCR was performed using the Biomark™ HD System (Standard Biotools), which supports the 96.96 Dynamic Array™ IFC. The microglial cDNA was pre-amplified using the Fluidigm Preamp Master Mix (Fluidigm PN 100-5580 or PN 100-5581) according to manufacturer instructions (Quick reference 100-5875). All samples were diluted 5-fold. Samples and primers were loaded according to the protocol described in Gene Expression with the 96.96 IFC Using Delta Gene Assays Quick Reference (100-9792) using SsoFast EvaGreen Supermix with low ROX (Bio-Rad, 172-5211) and 20X DNA Binding Dye (Fluidigm, 100-7609) for samples and 2X Assay Loading Reagent (Fluidigm100-7611) for primers. All primer pairs were designed and validated in-house for efficiency and specificity. The cDNA-generated signals for target genes were internally corrected with phosphoglycerate kinase 1 (Pgk1) in mouse postnatal brains, tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta (Ywhaz) in mouse placentas, and Ferritin Heavy Chain 1 (Fth1) and beta actin (Actb) for sorted microglia. The regulation fold changes were determined with the 2−∆∆Cq method65. Primer pair sequences have been summarized in the Supplementary Information.
Histology
Animals were anesthetized using isoflurane (Isothesia, Henry Schein Animal Health) and transcardially perfused with 1× PBS followed by 4% PFA. Brains were post-fixed for 24 h in 4% PFA and cryoprotected in 20% sucrose in PBS. Serial 40-μm-thick sagittal cerebellar sections were collected using a freezing microtome. Immunohistochemistry was performed as previously described5. Briefly, sections were incubated with goat anti-IBA1 (Novus Biologicals 1:500) overnight at 4 °C, containing 0.3% Triton and 10% normal donkey serum. Sections were then incubated with secondary antibody, donkey anti-goat Alexa-647 (Invitrogen, no. A-21447,1:500) together with DAPI (Invitrogen, no. D1306, 1:1000) for 2 h at room temperature. Floating sections were mounted and cover-slipped using ProLong Gold Antifade Mountant (Thermo Fisher Scientific, P36930) before imaging.
Fluorescence imaging and cell counting
Cerebellar sagittal sections were imaged using a Nikon Ti Eclipse inverted microscope with an A1 scanning confocal unit under a ×20 objective. Z-stack images were acquired with a step size of 2.0μm and viewed using NIH ImageJ 1.53c (http://imagej.nih.gov/ij). The different cerebellar layers were delineated manually using the freehand selection tool in ImageJ and measured for area. Cropped sections of IBA1-stained CB were uploaded to Imaris 9.90 (Oxford Instruments) and processed through the machine learning-based spot creation wizard workflow. The algorithm for spot detection was trained with a nucleus size of 8 μm. Using an autogenerated grid overlaid on the image, microglial nuclei were identified within seven randomly selected grid boxes, and any errors were corrected. The trained algorithm was subsequently applied to images.
Whole-brain imaging analysis
Thirty-day-old C and plKO mice underwent transcardial perfusion using ice-cold 1X PBS containing 10 U/mL heparin until clear perfusate was observed, followed by ice-cold 4% PFA. Following brain extraction, tissues were post-fixed in 4% PFA for 24 h before transfer to PBS containing 0.02% sodium azide. LifeCanvas Technologies (Cambridge, MA) performed whole-brain clearing and imaging as a contracted service. Using previously established protocols (PMID: 39080289), whole brains underwent SHIELD treatment for protein preservation, followed by clearing and IBA1 immunolabeling using SmartBatch+ technology. Brain imaging was conducted using a LifeCanvas SmartSPIM Light Sheet Microscope, with subsequent cell quantification and heatmap generation performed via SmartAnalytics following Allen Brain Atlas registration.
Protein extraction and cytokine assay
Proteins were extracted from the whole CB homogenized in 150 µm of extraction buffer containing: 20 mM Tris HCl (pH 7.5), 0.5% Tween 20, 150 mM NaCl, with 1 tablet cOmplete, Mini, EDTA-free protease inhibitor dissolved per 10 ml of buffer solution (Roche 11836170001). For protein extraction of L-902,688 or Veh treated samples, and embryonic tissue (placenta and brain), tissues were homogenized in 250 μl of radioimmunoprecipitation assay lysis buffer consisting of (in mM) 50 Tris-HCl, pH 7.4, 150 NaCl, 2 EDTA, 50 NaF, 1 Na3VO4, 1% Triton X-100, 0.1% SDS, 0.5% Na-deoxycholate and a Protease/Phosphatase Inhibitor Cocktail (Santa Cruz Biotechnology). Homogenate was then centrifuged at 10,000 × g for 10 min at 4 °C, and the supernatant was frozen at −80 °C. Protein concentrations were assessed using the Pierce BCA total protein assay and normalized for concentration. For the cytokine multiplex assay, samples were shipped to Eve Technologies (Calgary, Alberta, Canada) for the Mouse Cytokine/Chemokine 44-Plex Discovery Assay® Array (MD44). Cytokine levels were assessed by Luminex 200 instruments running on Bio-Plex Manager™ software (with compatible Bio-Rad calibration kits).
ELISA assays
All ELISAs were completed according to the manufacturer’s instructions. PGE2 was assessed using a competitive enzyme immunoassay ELISA kit (Mybiosource, MBS732453). Briefly, the assay sample and buffer were incubated together with PGE2-HRP conjugate in an anti-PGE2 pre-coated plate for one hour at 37 °C. Wells were then washed five times with wash buffer and incubated with HRP enzyme substrate for 30 min at 37 °C. To assess the concentration of CD47/IAP, a double-antibody sandwich ELISA was used (Mybiosource, MBS2097217). Similarly, samples were added to a pre-coated microtiter plate containing an HRP-conjugated antibody specific to IAP and incubated at 37 °C for one hour, followed by a one-hour incubation with detection reagent. After five washes with the wash buffer, the TMB substrate solution was added. To determine the MBP concentration, a sandwich ELISA was used (Mybiosource, MBS2501572). Samples were added to the anti-MBP pre-coated plate, followed by an MBP-biotinylated detection antibody and Avidin HRP conjugate. For all ELISA kits, the enzyme-substrate reaction was stopped using the stop solution, and color intensity was spectrophotometrically measured at 450 nm.
Mass spectrometry
Mass spectrometry was conducted by Columbia’s Irving Institute for Clinical and Translational Reseach Biomarkers Core Laboratory (BCL). Estradiol, and testosterone were measured in mouse brain and placental tissues by high-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS) on a platform comprising of Waters Xevo TQS triple quadrupole mass spectrometer integrated with a Waters Acquity UHPLC (Waters, Milford, MA). Tissue samples were homogenized in LCMS grade water using the Precyllis lysing kits with a bead homogenizer (Benchmark, NH). The steroids were extracted from the tissue homogenates spiked with deuterated internal standards by liquid-liquid extraction using a mixture of hexane/dichloromethane (3:2). Chromatographic separation was performed on a Waters C18 BEH column (2.1 × 100 mm, 1.7 µm) maintained at 50 °C, employing gradient elution using water and acetonitrile with 0.1% formic acid as mobile phases. Positive ESI-MS/MS under MRM mode was performed using the following transitions: estradiol 506.2 > 171.1; testosterone 289.2 > 109.1. The steroid concentration was measured by comparing integrated peak areas against known amounts of dexamethasone using Targetlynx V4.2. The lower limit of quantitation of the assay was 2.5 pg/ml. The mean intra-and inter-assay imprecision was 1.01%.
Flow cytometry
Cell suspension of microglia for flow cytometry was performed according to an adapted protocol from Bohlen et al.66. Briefly, mice were perfused with ice-cold PBS, and CB were extracted and placed into 1 mL of sample buffer containing 1× PBS and DNAse for homogenization. Samples were centrifuged at 9300 × g for 1 min and pellets suspended in 0.9 mL MACS buffer (Miltenyi Biotec MACS® BSA Stock Solution, 1X PBS, Fetal Bovine Serum, 0,5 M EDTA, 1 M HEPES) with 100 µm of myelin removal beads (Miltenyi Biotec, 130-096-433). Samples were incubated for 15 min with gentle flicking every 5 min to mix the beads and diluted with 1 mL of MACS buffer and centrifuged at 9300 × g for 1 min. Samples were then resuspended in MACS buffer and applied to LS columns (Miltenyi Biotec, 130-042-401) on the QuadroMACS™ Separator. Columns were washed to elute any remaining cells. Flow-through was collected, centrifuged, and resuspended in PBS pHrodo™ Red (Invitrogen P36600), CD11b Monoclonal Antibody (M1/70), FITC (11-0112-82), for 30 min. Lastly, samples were spun down, and pellets were resuspended in flow buffer containing DAPI (Invitrogen, D1306) and Vybrant™ DyeCycle™ Ruby Stain (Invitrogen, V10309). Microglial sorting was done on the Sony MA900 Multi-application Cell Sorter (Sony Biotechnology). Microglial population statistics were obtained on the NovoCyte Quanteon (Agilent).
Image stream
Cells were suspended as previously described above. Cells were first stained with CD11b Monoclonal Antibody (M1/70), FITC (11-0112-82) for 30 min followed by 20 min of fixation in 4% paraformaldehyde solution. Paraformaldehyde was decanted, and cells were resuspended in permeabilization buffer with 0.3% Triton X-100 with 0.5% BSA 10 min. Cells were then stained with intracellular stain, including PE-Dazzle 594-LAMP1 (Biolegend, 121624, 1:100), Alexa Fluor 647-MBP (Biolegend, 850910, 1:200), and DAPI for an additional 30 min. Cells were resuspended in 35 µm of 1X PBS for acquisition on the ImageStreamX Mk II imaging cytometer using Channel 1 (Brightfield for Camera 1), Channel 2 (FITC, CD11b), Channel 4 (PE-Dazzle 594-LAMP1), Channel 6 (Side Scatter), Channel 7 (DAPI), Channel 9 (Brightfield for Camera 2), and Channel 11 (Alexa Fluor 647-MBP). The laser powers were set up, measuring the Raw Max Pixel feature for each active channel, ensuring no saturation. Images were acquired at 60x magnification and recorded on a plot of Area against Aspect Ratio for brightfield (Channel 1) to exclude Speed Beads from the file. Compensation was performed using the compensation wizard in INSPIRE, and Anti-Rat Ig, κ/Negative Control (BSA) Compensation Plus (7.5 µm) Particles Set (BD Bioscience Cat# 560499) for the antibody conjugates, and a control was prepared using cells for DAPI. Amnis IDEAS software was used to analyze data, which was analyzed first by selecting focused cells using the Gradient RMS feature of Channel 1(Camera 1 brightfield). Then, single cells were selected in a plot of Area against Aspect Ratio of Channel 1. DAPI+ and CD11b+ objects (microglia) were selected using the Intensity feature of each channel. To assess the presence of myelin inside microglia, the IDEAS Internalization Wizard was used.
Myelin isolation
Myelin was isolated from the plKO CB of P15 mice according to the previously described protocol67. Briefly, CB was Dounce homogenized in a solution containing 10 mM HEPES, 5 mM EDTA, 0.32 M sucrose, and protease inhibitor. The homogenate was then separated using a sucrose gradient, first layered on top of 0.85 M sucrose/HEPES-EDTA buffer and centrifuged at 75,000 × g for 30 min. The myelin layer was collected and then centrifuged at 75,000 × g for 15 min in water. Ice-cold water was then used to further purify the pellet by hypo-osmotic shock at 12,000 × g for 10 min. This was repeated twice, and the pellet was resuspended in HEPES-EDTA buffer and stored at −80 °C.
Animal behavior
Stereotyped behavior
To test one aspect of ASD-like behavior, an assessment of spontaneous repetitive behavior was done. After a 30-min habituation period, mice were placed in a new, empty cage with a lateral-view video camera. 15 min of spontaneous behavior was recorded and manually scored for time spent, digging, gnawing, and grooming.
Sociability
The three-chamber test (Crawley’s paradigm68) was used to assess social behavior as previously described69. A clear, Plexiglas box with three chambers was used with small openings to allow for free movement. The lateral chambers were each equipped with an identical wire cup. Following 10 min of habituation to the testing arena (session 1), an unfamiliar adult mouse of similar weight, age, sex, and coloration was placed within the wire cup in one of the side chambers, and an ‘object’ was placed within the wire cup in the opposite chamber for a 10-min test session (session 2). Social preference was then analyzed using the social preference index (SPI). SPI was calculated as follows: if time spent with the mouse (social) is S and the object (non-social) is NS, then SPI = S/(S + NS).
Statistics
All data analyses were conducted blinded to group allocation. Mouse assignment to groups was not randomized as knowledge of genotype (plKO versus C) is required. Mice from a minimum 2 litters were used per group. All sample sizes are presented in legends. Interquartile range (IQR = Q3 – Q1) was used to assess each group for outliers in qPCR, flow cytometry, multiplex cytokine assays, and microscope images. ROUT outlier analysis was completed for ELISAs. All analyses were evaluated for normality, and if not normal, non-parametric tests were used. Comparison between two groups (C and plKO) in Biomark high-throughput RT-qPCR was analyzed using an unpaired Welch’s t-test. Two-way ANOVA was used to compare means across two or more independent variables (i.e., sex and genotype) with Sidak multiple comparisons testing. For comparisons with three factors (i.e., age, sex, and genotype), a three-way ANOVA with Tukey’s multiple comparisons testing was used. For comparison of means with one independent variable, such as treatment with RRx-001, one-way ANOVA or non-parametric Kruskal–Wallis with Dunn’s Multiple Comparisons was used. Summary data are presented in the text as mean ± s.e.m. from n animals. Differences were considered significant at p < 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The RNA-sequencing data discussed in this publication have been deposited in NCBI’s Gene Expression Omnibus and are accessible through the GEO Series accession number GSE306583. All data necessary for the conclusions described in this study are included in the source data provided with this paper. Source data are provided with this paper.
References
May, T., Adesina, I., McGillivray, J. & Rinehart, N. J. Sex differences in neurodevelopmental disorders. Curr. Opin. Neurol. 32, 622–626 (2019).
Wardinger, J. E. & Ambati, S. Placental insufficiency. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, (2020).
Liu, D., Liu, D., Wang, Q. & Xiong, T. Placental dysfunction: the core mechanism for poor neurodevelopmental outcomes in the offspring of preeclampsia pregnancies. Placenta 126, 224–232 (2022).
Kratimenos, P. & Penn, A. A. Placental programming of neuropsychiatric disease. Pediatr. Res. 86, 157–164 (2019).
Vacher, C. M. et al. Placental endocrine function shapes cerebellar development and social behavior. Nat. Neurosci. 24, 1392–1401 (2021).
Brunton, P. J., Russell, J. A. & Hirst, J. J. Allopregnanolone in the brain: protecting pregnancy and birth outcomes. Prog. Neurobiol. 113, 106–136 (2014).
Hosie, A. M., Wilkins, M. E., da Silva, H. M. & Smart, T. G. Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites. Nature 444, 486–489 (2006).
Schumacher, M. et al. Revisiting the roles of progesterone and allopregnanolone in the nervous system: resurgence of the progesterone receptors. Prog. Neurobiol. 113, 6–39 (2014).
Mellon, S. H. Neurosteroid regulation of central nervous system development. Pharmacol. Ther. 116, 107–124 (2007).
Rupprecht, R., Berning, B., Hauser, C. A., Holsboer, F. & Reul, J. M. Steroid receptor-mediated effects of neuroactive steroids: characterization of structure-activity relationship. Eur. J. Pharmacol. 303, 227–234 (1996).
Balan, I. et al. Neurosteroid allopregnanolone (3α, 5α-THP) inhibits inflammatory signals induced by activated MyD88-dependent toll-like receptors. Transl. Psychiatry 11, 145 (2021).
Balan, I., Beattie, M. C., O’Buckley, T. K., Aurelian, L. & Morrow, A. L. Endogenous neurosteroid (3α, 5α) 3-hydroxypregnan-20-one inhibits toll-like-4 receptor activation and pro-inflammatory signaling in macrophages and brain. Sci. Rep. 9, 1220 (2019).
Fitzgerald, K. A. et al. Mal (MyD88-adapter-like) is required for Toll-like receptor-4 signal transduction. Nature 413, 78–83 (2001).
Gay, N. J., Symmons, M. F., Gangloff, M. & Bryant, C. E. Assembly and localization of Toll-like receptor signalling complexes. Nat. Rev. Immunol. 14, 546–558 (2014).
He, J., Evans, C. O., Hoffman, S. W., Oyesiku, N. M. & Stein, D. G. Progesterone and allopregnanolone reduce inflammatory cytokines after traumatic brain injury. Exp. Neurol. 189, 404–412 (2004).
Jolivel, V. et al. Microglial cell morphology and phagocytic activity are critically regulated by the neurosteroid allopregnanolone: a possible role in neuroprotection. Cells 10, 698 (2021).
Liao, G. et al. Allopregnanolone treatment delays cholesterol accumulation and reduces autophagic/lysosomal dysfunction and inflammation in Npc1-/- mouse brain. Brain Res. 1270, 140–151 (2009).
Yilmaz, C. et al. Neurosteroids as regulators of neuroinflammation. Front. Neuroendocrinol. 55, 100788 (2019).
Hirst, J. J., Yawno, T., Nguyen, P. & Walker, D. W. Stress in pregnancy activates neurosteroid production in the fetal brain. Neuroendocrinology 84, 264–274 (2006).
Westcott, K. T., Hirst, J. J., Ciurej, I., Walker, D. W. & Wlodek, M. E. Brain allopregnanolone in the fetal and postnatal rat in response to uteroplacental insufficiency. Neuroendocrinology 88, 287–292 (2008).
Wenzel, P. L. & Leone, G. Expression of Cre recombinase in early diploid trophoblast cells of the mouse placenta. Genesis 45, 129–134 (2007).
Hertig, A. et al. Steroid profiling in preeclamptic women: evidence for aromatase deficiency. Am. J. Obstet. Gynecol. 203, 477. e471–477.e479 (2010).
Pecks, U. et al. Maternal serum lipid, estradiol, and progesterone levels in pregnancy, and the impact of placental and hepatic pathologies. Geburtshilfe Frauenheilkd. 76, 799–808 (2016).
Yawno, T., Yan, E., Walker, D. W. & Hirst, J. J. Inhibition of neurosteroid synthesis increases asphyxia-induced brain injury in the late gestation fetal sheep. Neuroscience 146, 1726–1733 (2007).
Bakalar, D. et al. Lack of placental neurosteroid alters cortical development and female somatosensory function. Front. Endocrinol. 13, 972033 (2022).
Ritchie, M. E. et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Aacids Res. 43, e47–e47 (2015).
Smyth, G. K., Michaud, J. & Scott, H. S. Use of within-array replicate spots for assessing differential expression in microarray experiments. Bioinformatics 21, 2067–2075 (2005).
Law, C. W., Chen, Y., Shi, W. & Smyth, G. K. voom: precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 15, 1–17 (2014).
Marriott, M. P. et al. Leukemia inhibitory factor signaling modulates both central nervous system demyelination and myelin repair. Glia 56, 686–698 (2008).
Schaecher, K., Shields, D. & Banik, N. Mechanism of myelin breakdown in experimental demyelination: a putative role for calpain. Neurochem. Res. 26, 731–737 (2001).
Lenz, K. M., Nugent, B. M., Haliyur, R. & McCarthy, M. M. Microglia are essential to masculinization of brain and behavior. J. Neurosci. 33, 2761–2772 (2013).
Han, V. X. et al. Maternal acute and chronic inflammation in pregnancy is associated with common neurodevelopmental disorders: a systematic review. Transl. Psychiatry 11, 71 (2021).
Santos, E. N. & Fields, R. D. Regulation of myelination by microglia. Sci. Adv. 7, eabk1131 (2021).
Gitik, M., Liraz-Zaltsman, S., Oldenborg, P. A., Reichert, F. & Rotshenker, S. Myelin down-regulates myelin phagocytosis by microglia and macrophages through interactions between CD47 on myelin and SIRPalpha (signal regulatory protein-alpha) on phagocytes. J. Neuroinflammation 8, 24 (2011).
Gitik, M., Kleinhaus, R., Hadas, S., Reichert, F. & Rotshenker, S. Phagocytic receptors activate and immune inhibitory receptor SIRPα inhibits phagocytosis through paxillin and cofilin. Front. Cell. Neurosci. 8, 104 (2014).
Müller, C., Bauer, N. M., Schäfer, I. & White, R. Making myelin basic protein -from mRNA transport to localized translation. Front. Cell. Neurosci. 7, 169 (2013).
Cabrales, P. RRx-001 acts as a dual small molecule checkpoint inhibitor by downregulating CD47 on cancer cells and SIRP-α on monocytes/macrophages. Transl. Oncol. 12, 626–632 (2019).
Poliani, P. L. et al. TREM2 sustains microglial expansion during aging and response to demyelination. J. Clin. Invest. 125, 2161–2170 (2015).
Nemes-Baran, A. D., White, D. R. & DeSilva, T. M. Fractalkine-dependent microglial pruning of viable oligodendrocyte progenitor cells regulates myelination. Cell Rep. 32, 108047 (2020).
Perez-Pouchoulen, M., Yu, S. J., Roby, C. R., Bonsavage, N. & McCarthy, M. M. Regulatory control of microglial phagocytosis by estradiol and prostaglandin E2 in the developing rat cerebellum. Cerebellum 18, 882–895 (2019).
Dean, S. L. et al. Prostaglandin E2 stimulates estradiol synthesis in the cerebellum postnatally with associated effects on Purkinje neuron dendritic arbor and electrophysiological properties. Endocrinology 153, 5415–5427 (2012).
Woodling, N. S. et al. Suppression of Alzheimer-associated inflammation by microglial prostaglandin-E2 EP4 receptor signaling. J. Neurosci. 34, 5882–5894 (2014).
Hoffman, J. F., Wright, C. L. & McCarthy, M. M. A critical period in Purkinje cell development is mediated by local estradiol synthesis, disrupted by inflammation, and has enduring consequences only for males. J. Neurosci. 36, 10039–10049 (2016).
VanRyzin, J. W., Pickett, L. A. & McCarthy, M. M. Microglia: driving critical periods and sexual differentiation of the brain. Dev. Neurobiol. 78, 580–592 (2018).
Vacher, C. M., Bonnin, A., Mir, I. N. & Penn, A. A. Editorial: Advances and perspectives in neuroplacentology. Front. Endocrinol. 14, 1206072 (2023).
Mor, G., Cardenas, I., Abrahams, V. & Guller, S. Inflammation and pregnancy: the role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 1221, 80–87 (2011).
Hussain, T. et al. Understanding the immune system in fetal protection and maternal infections during pregnancy. J. Immunol. Res. 2022, 7567708 (2022).
Afkham, A. et al. Toll-like receptors signaling network in pre-eclampsia: an updated review. J. Cell Physiol. 234, 2229–2240 (2019).
Aldo, P. B. et al. A novel three-dimensional in vitro system to study trophoblast-endothelium cell interactions. Am. J. Reprod. Immunol. 58, 98–110 (2007).
Goldenberg, R. L., Hauth, J. C. & Andrews, W. W. Intrauterine infection and preterm delivery. N. Engl. J. Med. 342, 1500–1507 (2000).
Rodrigues-Duarte, L., Pandya, Y., Neres, R. & Penha-Goncalves, C. Fetal and maternal innate immunity receptors have opposing effects on the severity of experimental malaria in pregnancy: beneficial roles for fetus-derived toll-like receptor 4 and type i interferon receptor 1. Infect Immun. https://doi.org/10.1128/IAI.00708-17 (2018).
Goines, P. E. et al. Increased midgestational IFN-gamma, IL-4 and IL-5 in women bearing a child with autism: a case-control study. Mol. Autism 2, 13 (2011).
Gumusoglu, S. B. et al. Chronic maternal interleukin-17 and autism-related cortical gene expression, neurobiology, and behavior. Neuropsychopharmacology 45, 1008–1017 (2020).
Smith, S. E., Li, J., Garbett, K., Mirnics, K. & Patterson, P. H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 27, 10695–10702 (2007).
Werling, D. M. The role of sex-differential biology in risk for autism spectrum disorder. Biol. Sex. Differ. 7, 58 (2016).
Bronson, S. L. & Bale, T. L. Prenatal stress-induced increases in placental inflammation and offspring hyperactivity are male-specific and ameliorated by maternal antiinflammatory treatment. Endocrinology 155, 2635–2646 (2014).
Osman, H. C. et al. Impact of maternal immune activation and sex on placental and fetal brain cytokine and gene expression profiles in a preclinical model of neurodevelopmental disorders. J. Neuroinflammation 21, 118 (2024).
Hughes, A. N. & Appel, B. Microglia phagocytose myelin sheaths to modify developmental myelination. Nat. Neurosci. 23, 1055–1066 (2020).
Wlodarczyk, A. et al. A novel microglial subset plays a key role in myelinogenesis in developing brain. EMBO J. 36, 3292–3308 (2017).
Hagemeyer, N. et al. Microglia contribute to normal myelinogenesis and to oligodendrocyte progenitor maintenance during adulthood. Acta Neuropathol. 134, 441–458 (2017).
Shi, J. et al. The prostaglandin E2 E-prostanoid 4 receptor exerts anti-inflammatory effects in brain innate immunity. J. Immunol. 184, 7207–7218 (2010).
McCarthy, M. M. How to study the origins of sex differences in brain and behavior. Neuronline. https://neuronline.sfn.org/-/media/Project/Neuronline/PDFs/2019/How-to-Study-the-Origins-of-Sex-Differences-in-Brain-and-Behavior.pdf (2019).
Hicks, S. C. & Irizarry, R. A. Quantro: a data-driven approach to guide the choice of an appropriate normalization method. Genome Biol. 16, 1–8 (2015).
Hicks, S. C. et al. Smooth quantile normalization. Biostatistics 19, 185–198 (2018).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods 25, 402–408 (2001).
Bohlen, C. J., Bennett, F. C. & Bennett, M. L. Isolation and culture of microglia. Curr. Protoc. Immunol. 125, e70 (2019).
Larocca, J. N. & Norton, W. T. Isolation of myelin. Curr. Protoc. cell Biol. 33, 3.25. 21–23.25. 19 (2006).
Nadler, J. J. et al. Automated apparatus for quantitation of social approach behaviors in mice. Genes Brain Behav. 3, 303–314 (2004).
Silverman, J. L., Yang, M., Lord, C. & Crawley, J. N. Behavioural phenotyping assays for mouse models of autism. Nat. Rev. Neurosci. 11, 490–502 (2010).
Acknowledgements
We thank Prof. Margaret M. McCarthy (U. Maryland, Baltimore) for her insightful recommendations on our manuscript. We thank Dr. G. Leone for the use of the Cyp19a-Cre mouse line. Additionally, we extend our appreciation to Dr. M. Yang (Neurobehavioral Core at Columbia University Medical Center) and to Columbia’s Institute for Comparative Medicine. Special thanks are due to Dr. Richard Friedman from the Herbert Irving Comprehensive Cancer Center Biomedical Informatics Shared Resource for his invaluable guidance in the analysis of our time-course RNA-sequencing experiments. We are also grateful to Elliot Brandwein for coding support and to Parssa Akbari for technical assistance. All confocal microscopic analyses were conducted at the Herbert Irving Comprehensive Cancer Center (HICCC) Confocal and Specialized Microscopy Core. Research reported in this publication using the ImageStreamX MK II imaging cytometer was performed in the Columbia University Stem Cell Initiative (CSCI) Flow Cytometry core facility at Columbia University Irving Medical Center and was supported by the Office of the Director, National Institutes of Health under Award Number S10OD026845. We acknowledge the University Paris Paris-Saclay (France) for supporting Dr. Vacher’s research leave. This work was funded by National Institutes of Health grants R01HD092593 (A.A.P.), T32NS064928 (J.S.), R21HD109623 (A.A.P.), and the Simons Foundation (SFARI Pilot Award no. 572832; A.A.P.).
Author information
Authors and Affiliations
Contributions
J.S., C.-M.V. and A.A.P. conceived the project and designed the experiments. J.S. and J.O. performed the RNA-sequencing analysis. H.L. contributed to the coding. J.S. and H.L. performed RT-qPCRs. J.S. performed histological experiments and cell counting. J.S. and M.K. performed the flow cytometry experiments. J.G. and C.-M.V. conducted the behavioral experiments and analyses. J.G. performed the genotyping. J.S. and H.L. completed the drug injections and subsequent tissue collection. J.S., C.-M.V. and A.A.P. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors no competing interests.
Peer review
Peer review information
Nature Communications thanks James Cuffe and the other anonymous reviewer(s) for their contribution to the peer review of this work. [A peer review file is available.]
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Salzbank, J., Lacaille, H., Gaby, J. et al. Microglia alter sex-specific cerebellar myelination following placental hormone loss. Nat Commun 16, 9846 (2025). https://doi.org/10.1038/s41467-025-64814-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-64814-z
