
Abstract
Premenopausal women preferentially store fat in subcutaneous depots, which provides protection against cardiometabolic disease. Their white adipose tissue also exhibits a more thermogenic, brown-like profile compared with that of men, yet the mechanisms underlying this sex-specific benefit remain unclear. Here, we show that Nuclear Receptor Coactivator 1 is highly expressed in human subcutaneous fat, with further induction during the conversion of white to beige adipocytes and in response to caloric restriction. Loss of Nuclear Receptor Coactivator 1 in subcutaneous adipocytes revealed a cell-autonomous role in promoting beiging by directly regulating the thermogenic factor Uncoupling Protein 1 and sex-dependent mitochondrial gene networks activated by cold and adrenergic stimulation. Acting together with GATA binding protein 3, it establishes elevated basal thermogenic tone in female subcutaneous fat. Female mice lacking this coregulator progressively developed obesity and glucose intolerance, and a male-like fat distribution, with increased visceral fat and reduced beige adipocytes. These findings identify Nuclear Receptor Coactivator 1 as a sex-specific determinant of adipose tissue remodelling and female metabolic resilience.
Similar content being viewed by others
Introduction
Several key aspects of energy homeostasis and metabolic regulation that contribute to susceptibility to the etiology of obesity-related cardiometabolic diseases, such as type 2 diabetes (T2D), are differentially regulated between women and men1,2,3,4. For example, clear sex differences have been reported in body composition, fat distribution, and utilization of carbohydrates and lipids as energy fuels5,6. Indeed, while women have a higher percentage of total fat mass than men, who have proportionally more muscle and lean mass, it has been reported that metabolically healthy premenopausal women tend to store fat in subcutaneous sites, particularly the gluteofemoral region, in contrast to the preferential fat deposition in visceral areas in men1,4,7,8. Importantly, such sex differences in metabolic traits translate into significant sex-specific health benefits, as young women are protected against the development of visceral obesity and associated metabolic complications, compared to men of the same age, whereas older postmenopausal women are at higher risk of developing visceral obesity and insulin resistance, leading to an increased incidence of T2D and cardiovascular disease5,9. However, the molecular networks underlying sexually dimorphic metabolic imprints and their health benefits remain largely elusive. Identifying the hubs responsible for the selective cardiometabolic protection of females could likely provide effective sex-based therapeutic interventions against metabolic disorders.
Adipose tissue plays a central role in the regulation of energy balance and metabolic homeostasis. White adipose tissue (WAT) is the primary site of triglyceride storage and release, and brown adipose tissue (BAT) contains specialized thermogenic cells that are essential for the regulation of body temperature and energy balance10,11. It is now well established that healthy adult humans possess active BAT, localized in small depots, at various anatomical sites (supraclavicular, cervical, paravertebral, suprarenal, pericoronary, and thoracic periaortic)12,13,14,15. BAT activity is inversely correlated with body mass index (BMI) and decreases with age, while whitening of brown adipocytes is promoted by a high-fat diet and ageing16,17,18. Subsequently, islands of brown-like adipocytes were identified within mammalian WAT depots, upon cold exposure or β3 adrenergic receptor stimulation. These adipocytes, termed brite (brown-in-white) or beige adipocytes, are also thermogenic19,20. The beiging process is associated with the induction of UCP1 (uncoupling protein 1), the key thermogenic player located in the inner mitochondrial membrane, as well as the presence of multilocular lipid droplets and a high density of mitochondria in adipocytes. Further studies have shown that human BAT is composed of both classic brown adipocytes and beige adipocytes, depending on the anatomical location21,22,23,24,25. From a clinical perspective, the robust capacity of beige adipocytes to burn not only lipids but also carbohydrates, as revealed by human positron emission tomography (PET) scans, could be considered as a promising therapeutic means to clear excess glucose and triglycerides from the blood of patients in pathological situations, such as T2D, obesity, and cardiovascular disease26,27.
Interestingly, rodent studies suggest that females have a greater ability to induce beiging in the WAT than males. Brown areas in the perigonadal fat pads were first described in female mice28 and a clear sex dimorphism in the activity of beige / brite islands was subsequently noted29,30,31. In humans, women exhibit a more brown-like adipose tissue profile within their WAT depots than men of the same age12,16,32,33. The aim of the present study was to identify molecular players involved in the sex-selective regulation of WAT remodeling, with a focus on the beiging process that occurs preferentially in subcutaneous fat of women and in female mouse models. Here, we show that the transcriptional coregulator Nuclear Receptor Coactivator 1 (Ncoa1) is a driver of this process, contributing to sex-specific adaptations in peripheral energy use and the fine-tuning of metabolic networks that protect females against age-related metabolic dysfunction.
Results
Human NCOA1 is highly expressed in female subcutaneous fat and increases with caloric restriction and beige adipocyte differentiation
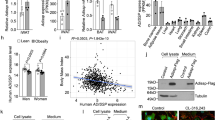
To identify the molecular players capable of mediating the beneficial metabolic effects that occur specifically in the subcutaneous white adipose tissue (SAT) of metabolically healthy women, we first interrogated data obtained from subcutaneous fat biopsies from non-diseased female donors of the Genotype-Tissue Expression (GTEx) project (n = 218; median age: 52.5 years)34, with a particular focus on transcriptional coregulators known to regulate large hierarchical sets of genes35,36,37. Interestingly, the coactivator NCOA1 showed the highest expression levels among the major coregulators with established key roles in WAT and/or BAT function, including PRDM16 (PR/SET domain 16)38,39, NRIP1 (nuclear receptor interacting protein 1)40, PPARGC1A (peroxisome proliferator activated receptor gamma coactivator 1 alpha)41, PPARGC1B (peroxisome proliferator activated receptor gamma coactivator 1 beta)42,43 and the other two members of the p160 coregulator family, NCOA2 and NCOA344,45,46 (Fig. 1a). Concurrently, a similar transcriptional profile with a predominant expression level of NCOA1 was also found following interrogation of a distinct human transcriptome carried out on an independent collection of SATs isolated from non-diabetic, non-obese premenopausal women (n = 12; median age: 43.8 + /− 1.9 years; median BMI: 26.0 + /− 1.5 kg/m2) (GSE11376447) (Fig. 1b). Interestingly, the same finding was not observed in human liver, another major metabolic tissue (Fig. S1a), thus suggesting the possibility of a selective effect of NCOA1 on SAT.

a Heat map of RNAseq analysis of subcutaneous adipose tissue (SAT) from female GTEx donors (n = 218; median age: 52.5 years). Data in TPM (Transcript Per Million). b Heat map of RNAseq analysis of SAT from non-diabetic, non-obese premenopausal women (n = 12; median age: 43.8 ±1.9 years; BMI 26.0 ±1.5 kg/m²). Dataset: GSE113764. Data in FPKM (Fragments Per Kilobase Million). c qRT-PCR of metabolic coregulators in adipocytes from gluteofemoral SAT of premenopausal (n = 7; median age: 31.4 ±1.6 years; median BMI: 26.0 ±1.2 kg/m²) and metabolically healthy postmenopausal women (n = 6; median age: 58.8 ±1.6 years; median BMI: 27.8 ±0.8 kg/m²) enrolled in ADIPENDO (Clinical trial: NCT02672826). Relative mRNA levels were normalized to ACTB and expressed as fold increase vs. ESR1. For premenopausal SAT, *** indicates p = 0.92e-6 for NCOA1 vs. NCOA2; p = 6.28e-6 for NCOA1 vs. NCOA3; p = 5.01e-6 for NCOA1 vs. PPARGC1A; p = 5.19e-6 for NCOA1 vs. PPARGC1B; and p = 4.08e-6 for NCOA1 vs. PRDM16. d - g RNAseq of SAT from female donors of the DiOGenes cohort (n = 93) at baseline, after an 8-week low-calorie diet (LCD; 800 kcal/day), and following a 6-month weight control diet (CD). Violin plots showing median and quartiles for NCOA1 (d), for NCOA2, NCOA3, and PPARGC1B (e), for PPARG (f), and for the beige adipocyte marker CPT1B (g). h Pearson’s correlation of NCOA1 and PPARG mRNA in SAT biopsies from all dietary groups of the DiOGenes cohort (n = 279). Two-tailed p values are reported. i Pearson’s correlation of NCOA1, NCOA2, NCOA3 with CPT1B in SAT biopsies from all dietary groups of DiOGenes (n = 279). Two-tailed p values are reported. j qRT-PCR of UCP1 and CPT1B in mature adipocytes from gluteofemoral SAT of premenopausal (n = 7) and postmenopausal (n = 6) women in ADIPENDO. k qRT-PCR of annotated genes during beige differentiation of hMADS cells over 14 days (n = 3 independent experiments, each performed in triplicates). l Gene expression of NCOA1, UCP1, NCOA2, NCOA3 in beige adipocytes derived from human SAT stromal vascular fraction from 3 female donors. Unpaired two-sided t-test was used for panels (c–g) and (j–l). Data are presented as mean ±SEM. Source data provided as a Source Data file.
As the SAT is a complex tissue entity comprising numerous distinct cell types11,48, we examined the mRNA levels of the same set of coregulators, in mature adipocytes isolated from the SAT of premenopausal (n = 7; median age: 31.4 + /− 1.6 years; median BMI: 26.0 + /− 1.2 kg/m2) and metabolically healthy postmenopausal women (n = 6; median age: 58.8 + /− 1.6 years; median BMI: 27.8 + /− 0.8 kg/m2) (Fig. S1b), enrolled in the ADIPENDO study (NCT0267282649). In all female subjects, regardless of their menopausal status, NCOA1 gene expression levels remained the highest, compared to all other key metabolic transcriptional coregulators tested (Fig. 1c). We also observed significantly higher expression of NCOA1 in metabolically healthy postmenopausal women compared with premenopausal women (Fig. 1c). Similarly, gene expression of NCOA2 and NCOA3, the other two members of the same family of transcriptional coactivators as NCOA1, was also found to be higher in metabolically healthy postmenopausal females than in premenopausal women (Fig. 1c).
To further evaluate the potential influence of NCOA1 on women’s metabolic health, its gene expression was analyzed in SAT biopsies from another cohort of obese, non-diabetic women (n = 93; median age: 42.0 + /− 0.62 years; median BMI: 34.5 + /− 0.5 kg/m2) from the Diet Obesity and Genes (DiOGenes) study, a pan-European, multicenter, randomized, controlled dietary intervention trial (NCT0039063750). Participants underwent an 8-week low calorie diet (LCD = 800 kcal/day), followed by a 6-month weight control diet (CD)50. Genome-wide transcriptomic analysis (RNAseq) was performed on SAT biopsies before (basal) and after the LCD diet, then after the follow-up diet (CD)51. As originally described, the LCD had a significant effect on BMI in obese women, with a clear beneficial metabolic adjustment on HOMA-IR and total cholesterol levels (Fig. S1c). Importantly, we found a significant increase in NCOA1 gene expression after LCD (Fig. 1d), with no difference for other coregulators such as NCOA2, NCOA3 and PPARGC1B (Fig. 1e). Concurrently, the gene expression of the master regulator of adipogenesis, PPARG (peroxisome proliferator activated receptor gamma) decreased after LCD (Fig. 1f), whereas the brown/beige adipocyte-associated gene, CPT1B (carnitine palmitoyltransferase 1B) was upregulated (Fig. 1g). We further observed a significant inverse correlation between NCOA1 and PPARG expression (Fig. 1h), independent of dietary interventions (Fig. S1d), while no clear correlation was found for NCOA2, NCOA3 or PPARGC1B (Fig. S1e). Conversely, a positive correlation was detected between NCOA1 and CPT1B expression (Fig. 1i), whereas NCOA2 and NCOA3 were inversely correlated with CPT1B (Fig. 1i) and PPARGC1B was not correlated with CPT1B (Fig. S1f). In support of these observations from the DiOGenes study, upregulation of UCP1 and CPT1B was also found in adipocytes from the SAT of metabolically healthy postmenopausal women from the ADIPENDO cohort compared to those from the SAT of premenopausal women (Fig. 1j). This finding is in line with that previously reported for NCOA1 (Fig. 1c), and confirms the close positive association between NCOA1 and key beiging players in distinct cohorts. Interestingly, in the non-diabetic obese men of the DiOGenes study (n = 46; median age: 43.3 ± 0.94 years; median BMI: 33.96 ± 0. 63 kg/m²), the SAT gene expression profile of NCOA1 (Fig. S1g), as well as the correlation patterns between NCOA1 and PPARG or CPT1B within the same fat depot (Fig. S1h), were remarkably similar to those observed in the SAT of non-diabetic women from the DiOGenes cohort. These findings provide compelling evidence supporting our previous observations regarding NCOA1 and the human SAT biology. Taken together, our results suggest a potentially important role for the coregulator NCOA1 in controlling the molecular adaptation of SAT, facilitating the transition from white adipocytes specialized for lipid storage to thermogenic adipocytes associated with enhanced beta-oxidation capacity.
To further substantiate the positive association between NCOA1 abundance and the beiging program, we next used human multipotent adipose-derived stem cells (hMADS), derived from the umbilical fat pad of a 31-month-old healthy female donor52 and induced their differentiation into beige adipocytes, as indicated by UCP1 gene upregulation (Fig. S1i). During the activation of the beige adipogenic program, a potent rise in NCOA1 gene expression was observed over time (Fig. 1k), mimicking the profile of the incremental transcriptional escalation of key coregulators of adipogenesis such as PPARGC1A, PPARGC1B and PRDM16 (Fig. 1k), whose absolute levels of gene expression, however, remained much lower than those of NCOA1 (Fig. 1k).
Finally, human adipose tissue stromal vascular fraction (SVF) cells, obtained from abdominal subcutaneous adipose tissue of three non-obese female donors, were differentiated into white or beige adipocytes53,54. The induction of NCOA1 mRNA levels was higher upon differentiation into beige adipocytes than into white adipocytes (Fig. 1l). This occurred concomitantly with upregulation of the key thermogenic gene UCP1 (Fig. 1l). No significant change was observed for NCOA2 and NCOA3 expression (Fig. 1l). Taken together, our observations in human biological samples suggest that NCOA1 has great potential to play a role in human white adipocytes, with a plausible involvement in the beiging process.
Ncoa1 activity exerts cell-autonomous control over energy dissipation during beiging, primarily by regulating the key thermogenic marker Ucp1 in a sex-selective manner
To further investigate the nature of the putative influence of Ncoa1 on the white-to-brown adipocyte conversion, we used SVF cells of SAT isolated from 16 to 20-week-old female Ncoa1 knockout (Ncoa1 KO) mice or their sex-matched wild-type (WT) littermates. Cells were treated with appropriate white or beige differentiation cocktails and then exposed to the beta-adrenergic agonist isoproterenol to fully activate the beiging transcriptional program. White adipocyte differentiation of SVF from Ncoa1 KO mice proceeded normally compared with WT, as evidenced by the transcriptional response of genes encoding the isoform 2 of peroxisome proliferative-activated receptor gamma (Pparg2) and the lipid droplet-associated protein perilipin 1 (Plin1) in Ncoa1 KO adipocytes, compared with WT cells (Fig. S2a). However, the expression of several key genes involved in beiging, including carnitine palmitoyltransferase 1b (Cpt1b), iodothyronine deiodinase 2 (Dio2), phosphoenolpyruvate kinase (Pepck), pyruvate dehydrogenase kinase isozyme 4 (Pdk4), peroxisome proliferative activated receptor alpha (Ppara), and pparg coactivator 1 beta (Ppargc1b), was significantly reduced after beige adipocyte differentiation of the SVF from Ncoa1 KO mice compared with WT controls, notably under conditions not supplemented with isoproterenol (Fig. 2a and S2b, c). An even greater drop in mRNA levels, compared with those of WT cells, was observed for the key thermogenic player, Ucp1, in the absence of Ncoa1, particularly during beiging (Fig. 2b). The transcriptional decline of Ucp1 expression in beige adipocytes lacking Ncoa1, was fully confirmed at the protein level, by Western blot analysis (Fig. 2c). When beige differentiation was performed over time using isoproterenol, Ucp1 protein levels were found to be strongly diminished in the absence of Ncoa1 (Fig. 2d). Assessment of global oxygen consumption in the same cellular model of primary mouse adipocytes (Fig. 2e–h), revealed a clear reduction in uncoupled energy dissipation in Ncoa1 KO beige adipocytes, compared with differentiated beige WT cells (Fig. 2g), confirming the apparent defect in Ucp1 activity, in the absence of Ncoa1. Since both the transcriptional regulation and the activity of Ucp1 are known to be controlled by free fatty acids, which are primarily released by lipolysis during beiging, the observed reduction in Ucp1 activity could possibly have resulted from a defect in lipolysis in Ncoa1 KO adipocytes. However, analysis of glycerol levels, in the presence or absence of isoproterenol, showed no difference between WT and Ncoa1 KO white or beige adipocytes (Fig. S2d). Concurrently, using Seahorse-based bioenergetics analysis, we found that the full respiratory activity of Ncoa1 KO beige adipocytes was significantly reduced, compared with that of WT cells (Fig. 2h). We therefore examined the transcriptional profiles of key genes involved in mitochondrial activity. Structural mitochondrial genes, such as Timm23 and Tomm20, had similar expression levels in Ncoa1-null adipocytes, compared with WT cells (Fig. 2i). Consistently, the mitochondrial mass, assessed by mitochondrial DNA analysis, was similar in Ncoa1 KO and WT beige adipocytes (Fig. S2e). This suggests that the absence of Ncoa1 had no effect on global mitochondrial biogenesis. In contrast, mRNA levels of genes involved in the mitochondrial respiratory chain complexes, such as cytochrome c oxidase subunit 5B (Cox5b), cytochrome c oxidase subunit 8B (Cox8b) and cytochrome C (Cycs) and genes involved in the TCA cycle such as citrate synthase (Cs), were significantly lower in Ncoa1-deficient adipocytes, compared with those of WT cells, specifically during the beiging adaptation (Fig. 2i). At the protein level, a significant reduction of all the five respiratory chain complexes was observed during beiging of Ncoa1 KO adipocytes (Fig. 2j, k), highlighting the key cell-autonomous transcriptional action of Ncoa1 in controlling mitochondrial function and respiratory chain complex activity. We next decided to test whether the same cell-autonomous effects of Ncoa1 depletion could be observed in male subcutaneous adipocytes. Following adipocyte differentiation of SVF cells isolated from the SAT of 16- to 20-week-old male Ncoa1 KO mice or their sex-matched WT littermates, we found that Ncoa1 deletion had no significant effect on Ucp1 gene expression or protein activity after beige conversion (Figs. S2f–h). Similarly, the expression of other key thermogenic genes, including Cpt1b, Pepck, and Plin5, remained unchanged (Fig. S2i).

a qRT-PCR analysis of annotated genes in white or beige adipocytes differentiated from stromal vascular fraction (SVF) cells of subcutaneous adipose tissue (SAT) of 16–20-week-old female Ncoa1 KO and WT mice, under basal conditions (Ctrl) or after 1 μM isoproterenol (Iso, 2 h). n = 3 biological replicates per condition were used. b qRT-PCR analysis of Ucp1 in white or beige adipocytes from SAT SVF of Ncoa1 KO and WT mice, with or without isoproterenol. n = 3 biological replicates per condition were used. c Western blot analysis of Ucp1 protein levels in white or beige adipocytes from SAT SVF of Ncoa1 KO and WT mice, in Ctrl or Iso conditions. Two biological replicates were loaded for each condition on the same blot. Plin1 was used as a loading control. This was repeated n = 3 independent times with similar results. d Western blot monitoring Ucp1 protein accumulation kinetics after Iso treatment in beige adipocytes from Ncoa1 KO and WT mice. Amidoblack staining served as a loading control. This was repeated n = 3 independent times with similar results. e Representative Seahorse experiments assessing oxygen consumption rate (OCR) in white or beige adipocytes from SAT SVF of WT and Ncoa1 KO mice. Cells were sequentially exposed to Iso, oligomycin (Oligo; ATP synthase inhibitor), FCCP (uncoupler), and rotenone (Rot) + antimycin A (Ant; electron transport inhibitors). n = 10 – 11 biological replicates per condition. This was repeated twice independently with similar results. f–h Quantification of basal, uncoupled, and maximal respiration from Seahorse data in (e). i qRT-PCR of genes involved in mitochondrial function in white or beige adipocytes from SAT SVF of 12-week-old female Ncoa1 KO and WT mice, in Ctrl and Iso conditions. n = 3 biological replicates per condition were used. j Western blot analysis of proteins from the five mitochondrial complexes in beige adipocytes from Ncoa1 KO and WT mice, with or without Iso. Amidoblack served as the loading control. Two biological replicates were loaded for each condition on the same blot. This was repeated 2 independent times with similar results. k Quantification of Western blot in (j) using Image Lab 5.0. Data represent mean ±SEM. Unpaired two-sided t-test was applied for panels a, b, e–i, and k. Source data provided as a Source Data file.
To further substantiate the sex-specific, cell-autonomous role of Ncoa1 in regulating thermogenic gene expression in differentiated SVF-derived adipocytes, we systematically profiled a panel of canonical thermogenic genes (Ucp1, Cidea, Cpt1b, Pepck, Dio2, Ppara, Ppargc1a, and Ppargc1b) across both sexes and genotypes. To more effectively induce thermogenic differentiation, we applied a higher concentration of isoproterenol (10 µM; Iso), simulating the robust thermogenic activation observed in vivo. Under these conditions, Ucp1 expression was consistently and significantly reduced in Ncoa1 KO female adipocytes across all stimulatory settings (white + Iso, beige + vehicle (control), and beige + Iso) relative to WT female cells (Fig. S2j). In contrast, Ncoa1 KO male adipocytes exhibited no reduction in Ucp1 expression compared to WT males (Fig. S2j). Similarly, several other thermogenic genes (Cidea, Cpt1b, Pepck, Dio2, Ppara, and Ppargc1b) were significantly downregulated in Ncoa1 KO female adipocytes, particularly under beige + Iso conditions (Fig. S2j). By comparison, Ncoa1 KO male adipocytes showed either no significant changes or increased expression of select thermogenic genes, including Ucp1 (white + Iso) and Cidea (Beige ± Iso) (Fig. S2j), suggesting possible compensatory or competitive regulatory mechanisms.
Collectively, these findings provide strong additional support for a sex-dependent, cell-autonomous role of Ncoa1 in orchestrating the thermogenic transcriptional program in subcutaneous beige adipocytes.
Ncoa1 activity controls the transcriptional program of WAT beiging, by regulating the expression levels of key mitochondrial genes, in a sex-selective manner
To consolidate our cellular findings, we decided to pharmacologically induce WAT beiging in vivo, using systemic administration of the β3 agonist CL316,243 in 16- to 20-week-old Ncoa1 KO and WT mice of both sexes. Unbiased RNAseq analysis of the SAT from treated (CL316,243) or untreated (NaCl) animals was further used to obtain a clear picture of the role of the coregulator Ncoa1 in the transcriptional gene orchestration of the beiging program in both sexes. Importantly, our transcriptomic data strongly suggested that Ncoa1 influences the beiging program in both males and females, but with an apparent sex bias and a greater effect in females (Fig. 3a–f). Indeed, the total number of differentially expressed genes (DEGs) in response to CL316,243 in Ncoa1 KO versus WT SAT, particularly the downregulated ones that would likely reflect a loss of function in beiging, was significantly greater in females than in males (570 vs 420, respectively) (Fig. 3a, d). Volcano plots of DEGs and principal component analysis (PCA) confirmed the differences between the sexes (Fig. 3a, d, S3a). In females, KEGG pathway enrichment analysis of downregulated SAT genes in response to CL316,243 revealed a robust effect of Ncoa1 on thermogenesis (q value = 1.07e-32; rich factor = 0.22) and, more selectively, on oxidative phosphorylation (q value = 5.22e-32; rich factor = 0.29), the two pathways being known to play crucial roles in beiging in vivo (Fig. 3b). Gene set enrichment analysis (GSEA) strongly confirmed our KEGG findings (Fig. 3c). In contrast, the top affected KEGG pathways in Ncoa1 KO males, following β3 agonist administration, differed from those in females of the same genotype, and included fatty acid degradation (q value = 5.95e-6; rich factor = 0.19), carbon metabolism (q value = 6.59e-6; rich factor = 0.10) and fatty acid metabolism (q value = 8.27e-6; rich factor = 0.16) (Fig. 3d–f). In contrast to females, oxidative phosphorylation was not significantly impaired in male Ncoa1 KO mice (q value = 0.074; rich factor = 0.05), and thermogenesis was only moderately affected (q value = 0.022; rich factor = 0.05) (Fig. 3e, f).

a Volcano plot and total number of differentially expressed genes (DEGs) in SAT of female Ncoa1 KO vs. WT mice treated with CL316,243 (CL), analyzed by DESeq2 (log2FC ≥ 0.25; q ≤ 0.05). n = 4 animals per group, except for females WT NaCl, Ncoa1 KO NaCl, and Ncoa1 KO CL: n = 3 per group. b KEGG pathway enrichment bubble plot of downregulated genes in female Ncoa1 KO vs. WT mice after CL treatment. Bubble size indicates number of genes, color indicates Q-value, and rich factor represents DEG proportion per pathway. c Gene set enrichment analysis (GSEA) plots showing thermogenesis (KEGG:04714; top) and oxidative phosphorylation (KEGG:00190; bottom) in SAT of female Ncoa1 KO vs. WT mice treated with CL. d Volcano plot of DEGs in SAT of male Ncoa1 KO vs. WT mice after CL treatment (DESeq2, log2FC ≥ 0.25; q ≤ 0.05). n = 4 males per group. e KEGG pathway enrichment bubble plot of downregulated SAT genes in male Ncoa1 KO vs. WT mice after CL treatment. f GSEA plots of thermogenesis and oxidative phosphorylation in SAT of male Ncoa1 KO vs. WT mice after CL. g, h Expression of genes encoding proteins of the respiratory chain complexes (g) and mitochondrial proteins (h) in SAT of male or female Ncoa1 KO and WT mice treated with CL or vehicle (NaCl). n = 4 animals per group, except for females WT NaCl, Ncoa1 KO NaCl, and Ncoa1 KO CL: n = 3 per group. i Representative H&E staining of SAT from WT and Ncoa1 KO mice treated with CL or NaCl. Scale bar: 100 μm. j Heatmap of thermogenic gene expression in SAT from male and female WT and Ncoa1 KO mice treated with CL or NaCl. n = 4 animals per group, except for females WT NaCl, Ncoa1 KO NaCl, and Ncoa1 KO CL: n = 3 per group. k Ucp1 mRNA levels in SAT from female and male WT and Ncoa1 KO mice after 7 days of CL or NaCl. n = 4 animals per group, except for females WT NaCl, Ncoa1 KO NaCl, and Ncoa1 KO CL: n = 3 per group. l Representative Ucp1 immunohistochemistry in SAT from male and female WT and Ncoa1 KO mice after CL. Scale bar: 200 μm. m RNAseq analysis of beiging genes (Cidea, Cpt1b) in SAT from male and female WT and Ncoa1 KO mice treated with CL or NaCl. n = 4 animals per group, except for females WT NaCl, Ncoa1 KO NaCl, and Ncoa1 KO CL: n = 3 per group. Values represent mean ±SEM. Data analyzed by two-way ANOVA with Fisher’s LSD test in panels g, h, k, and m. Source data provided as a Source Data file.
To further define the sex-specific, cell-autonomous role of Ncoa1 in regulating the beige adipocyte transcriptional program, we investigated mitochondrial gene responses to β3-adrenergic stimulation both in vivo and in vitro. In subcutaneous fat from Ncoa1 KO females treated with CL316,243, we observed a broad and significant downregulation of nuclear-encoded genes across all five oxidative phosphorylation complexes: Complex I (Ndufa3, Ndufa7, Ndufb7, Ndufb9), Complex II (Coq8a, Coq9, Sdhb), Complex III (Uqcr10, Uqcr11), Complex IV (Cox6a1, Cox8b), and Complex V (Atp5d, Atp5e, Atp5o), relative to WT females (Fig. 3g). This repression extended to key mitochondrial regulators including Phb1, Phb2, Bola3, and Polrmt (Fig. 3h), as well as genes encoding mitochondrial ribosomal subunits (e.g., Mrpl34, Mrpl42, Mrpl10, Mrpl16, Mrpl38, Mrpl58 and Mrps21, Mrps7, Mrps18b) (Fig. 3h, S3b-c) and other key genes involved in mitochondrial protein translation and maintennance of cristae junctions (e.g. TUFm and Chchd10 respectively) (Fig. S3f). Notably, these CL316,243-dependent transcriptional responses were more robust in WT female SAT than in WT male SAT, and this sex difference was nearly abolished in the absence of Ncoa1, implicating Ncoa1 as a key mediator of female-specific mitochondrial gene activation in subcutaneous fat. To demonstrate that these effects are cell-autonomous, we profiled a subset of these sexually dimorphic genes in SVF-derived adipocytes differentiated under white and beige conditions. The sex-biased expression observed in vivo was faithfully recapitulated in vitro, with higher expression in WT female beige adipocytes compared to WT male beige fat cells (Fig. S3d). Consistent with in vivo findings, Ncoa1 deletion selectively impaired the induction of these genes in female beige adipocytes, but not in males, further supporting a sex-specific, cell-autonomous role for Ncoa1 in coordinating mitochondrial gene programs during beige adipocyte differentiation.
At the histological level, hematoxylin and eosin staining of SAT depots from CL316,243-treated WT animals revealed large areas of brown-like remodeling with small multilocular adipocytes, whereas female and male Ncoa1 KO mice showed obvious defects in adipose tissue browning (Fig. 3i). The lack of WAT remodeling in response to CL316,243 in Ncoa1-deficient conditions, was fully confirmed at the molecular level, as the β3 agonist-dependent expression of Ucp1 was drastically impaired in Ncoa1 KO mice of both sexes, at the mRNA and protein levels (Fig. 3j–l and Fig. S3e). Importantly, a key observation was made for Ucp1 and two other major beiging genes, Cidea and Cpt1b, which showed distinct sex-specific profiles of their basal mRNA levels as well as different amplitudes of transcriptional responses to CL316,243. Indeed, WT females exhibited higher mRNA levels of Ucp1, Cidea and Cpt1b genes under basal conditions (i.e., NaCl), compared with those seen in WT males (Fig. 3k, m), and subsequently showed a lower amplitude of transcriptional response to CL316,243 than WT males. The absence of Ncoa1 in females, under basal conditions, resulted in a drastic decrease in the basal mRNA levels of Ucp1, Cidea, and Cpt1b, compared with those of WT female littermates (Fig. 3k, m), highlighting the critical role of Ncoa1 in controlling the basal levels of key endogenous genes of beiging in females. In contrast, the latter observation could not be made in males under basal conditions, as Ucp1, Cidea and Cpt1b mRNA levels in Ncoa1 KO males were all nearly undetectable, as in WT male mice (Fig. 3k, m and Fig. S3e). In males, the effect of Ncoa1 on the mRNA levels of these three beiging genes could only be measured under conditions of β3 agonist administration, where Ncoa1 depletion resulted in a marked reduction in the transcriptional response to CL316,243, compared with that seen in treated WT male littermates (Fig. 3k, m).
To further support our observations, cold room challenges were carried out to induce thermogenesis in vivo in a more physiological manner than β3-agonist administration (CL316,243). Thus, we were able to reproduce all our previous results, again with a strong impact of Ncoa1 depletion on the SAT thermogenesis genetic program, in a sex selective manner (Fig. S3g–j). In contrast, the expression of genes involved in BAT thermogenesis was only moderately reduced in Ncoa1 KO mice compared to WT mice, in both sexes (Fig. S3i). Interestingly, however, Fgf21 gene expression in BAT showed greater induction in female Ncoa1 KO mice than in female WT mice, with no significant difference observed between male Ncoa1 KO and WT mice (Fig. S3i). This result is consistent with the fact that Fgf21 is a mitokine known for its role in fine-tuning inter-organ metabolic communication, which may be activated in mouse models of mitochondrial dysfunction to induce energy expenditure and promote beiging compensation55.
Ncoa1 maintains elevated basal expression levels of key players of beiging in female subcutaneous WAT
Since our main objective was to unravel the molecular nodes responsible for the female-selective protection in metabolism and energy dissipation, we aimed to dissect the mechanisms underlying the marked sex-specific influence of Ncoa1 on the basal transcriptional activity of key beiging genes in SAT depots (Fig. 3k, m and Fig. S3g). We therefore searched our transcriptomic data for differences in gene signatures in SAT between female and male WT mice, under basal conditions (i.e., NaCl administration). Interestingly, under these conditions, without CL316,243 stimulation, PCA analysis confirmed a clear sexual dimorphism (Fig. 4a). The number of differentially expressed genes in the SAT of WT animals was also different, with 1590 genes being more highly expressed in females than in males, whereas 1230 genes showed a higher expression in males than in females (Fig. S4a). KEGG enrichment analysis revealed that, among the genes with higher expression in WT females, the fatty acid elongation program was the most significantly enriched (q value = 6.1e-4; rich factor = 0.325). In addition, the oxidative phosphorylation (q value = 0.0078; rich factor = 0.16) and thermogenesis (q value = 0.05; rich factor = 0.125) gene programs were found to be significantly enriched in WT females compared to WT males (Fig. 4b). In contrast, genes with higher expression in WT males included sets of genes with common cellular functions, such as ribosome biogenesis or cell cycle action (Fig. 4c). In the case of Ncoa1 KO animals, differential analysis of SAT transcriptomic data between females and males under basal conditions (NaCl) revealed a lower total number of DEGs than that observed when comparing dimorphic genes in WT mice (Fig. S4a, b). Furthermore, in Ncoa1 KO females exposed to control conditions (NaCl), the top KEGG pathways did not belong to any of the metabolic networks that were strongly affected in WT females, namely oxidative phosphorylation (q value = 0.99; rich factor = 0.013), thermogenesis (q value = 0.99; rich ratio = 0.008) or the fatty acid elongation program (q value = 0.90; rich ratio = 0.075) (Fig. 4b and S4c), confirming the high specificity of the transcriptional action of Ncoa1 in maintaining the constitutive expression of these three metabolic pathways in females.

a PCA of differentially expressed genes (DEGs) in SAT of WT females (n = 3) vs. WT males (n = 4) under basal (NaCl) conditions. b, c KEGG pathway enrichment bubble plots of DEGs in SAT of WT mice under basal conditions, comparing females (n = 3) vs. males (n = 4). Genes upregulated in WT females (b) and WT males (c). d RNAseq expression analysis of Elovl family genes in SAT of Ncoa1 KO and WT mice (n = 4 animals per group, except for females WT NaCl, Ncoa1 KO NaCl, and Ncoa1 KO CL: n = 3 per group). e Luciferase assays monitoring the transcriptional activity of the Ucp1 promoter (− 3750 / + 490 bp) in HeLa cells co-transfected with Ncoa1 and the indicated coregulators. Data are mean of 3 independent experiments, each performed in triplicates. f RNAseq expression analysis of Gata family genes in SAT of Ncoa1 KO and WT mice (same n as in (d)). g, h Luciferase assays monitoring the effect of Gata3 and Ncoa1 on Ucp1 promoter activity. Data are mean of 3 independent experiments, each performed in triplicates. i ChIP assay showing Ncoa1 and Gata3 binding to the proximal Ucp1 promoter in SAT of WT or Ncoa1 KO females and WT males. Data are mean of 3 independent experiments performed in 2 to 4 replicates. j GATA3 expression (RNAseq) in SAT and VAT from GTEx donors: women (SAT: n = 218; VAT: n = 170) and men (SAT: n = 445; VAT: n = 371). Data are in TPM (Transcript Per Million). k qRT-PCR analysis of Gata3 expression in SAT and BAT of female and male WT and Ncoa1 KO mice (n = 4 per group). l ATAC-seq at the Gata3 locus in both sexes (n = 2 WT mice per sex). Left: genome browser tracks; Right: average peak quantification. m–r Energy expenditure measured by indirect calorimetry during day (white) and night (gray) in WT and Ncoa1 KO mice: females (WT: n = 4; KO: n = 6) (m–o), and males (WT: n = 5; KO: n = 6) (p–r). s Locomotor activity (total 24 h distance) in 12-month-old KO vs. WT mice: females (KO: n = 7; WT: n = 5) and males (KO: n = 6; WT: n = 4). t Body weight of WT (n = 5 mice per sex group) and Ncoa1 KO mice (females n = 7 and males n = 5). Values are mean ±SEM. Two-way ANOVA with Fisher’s LSD test was applied (panels d, f); unpaired two-sided t-test used for panels e, and g–t. Source data provided as a Source Data file.
To further evaluate the sex-specific control of Ncoa1 on the constitutive transcriptional program of beiging within the SAT depot, we examined the expression levels of genes of the top enriched KEGG pathway showing sexual dimorphism in WT animals, namely the fatty acid elongase family (Elovl1-7) (Fig. 4d). As expected, four of the seven members of the family, i.e., Elovl 3, 5, 6 and 7, showed significantly higher expression in WT females, under basal conditions (NaCl), compared with age-matched WT males, with Elovl3 and Elovl6 displaying the most striking sex differences (Fig. 4d). However, the dimorphic pattern of the constitutive expression was completely lost in Ncoa1 KO females, as Ncoa1 deletion in females drastically reduced the basal mRNA levels of Elovl3 and Elovl6, compared to WT females, but had no effect on males (Fig. 4d). The amplitude of the CL316,243-dependent transcriptional response of Elovl6 and Elovl3 was substantially higher in WT males, compared with age-matched WT females, due to their low basal mRNA levels (Fig. 4d).
To capture the precise molecular sequences orchestrated by Ncoa1 in controlling the constitutive expression of key genes of beiging in the female SAT depot, we focused our analysis on the major marker of beiging, Ucp1. We carried out transient transfections of the well-characterized minimal promoter (-3750/ + 490 bp) of the Ucp1 gene into cells. A strong induction of the transcriptional activity of the Ucp1 reporter gene occurred following co-transfection with Ncoa1, in the absence of any ligand or exogenous inducer (Fig. 4e). This transcriptional response was significantly greater than that obtained with Ppargc1a (alias Pgc1α), a well-established coactivator of Ucp1 (Fig. 4e). Furthermore, by transfecting different DNA sequences of the Ucp1 promoter, we were able to localize the Ncoa1 responsive sequence in the very proximal part of the Ucp1 promoter (-502 / + 490 bp), strongly supporting an effect of Ncoa1 on the basal activity of the Ucp1 gene (Fig. S4d).
Since no significant sexual dimorphism in Ncoa1 gene expression was observed in human SAT (Fig. S4e) and mouse SAT (Fig. S4f), we searched for potential sex-biased candidates that could act in concert with Ncoa1 to specifically control basal expression of the Ucp1 gene in female subcutaneous adipocytes. Knowing that Ncoa1 preferentially interacts with nuclear receptors to control selective target genes45, we first examined the gene expression profiles of the entire family of nuclear receptors, including well-characterized sex steroid transcription factors such as estrogen receptor alpha (encoded by Esr1) or β (encoded by Esr2) and androgen receptor (encoded by Ar). We observed no expression of Esr2 in the SAT depot, no sexual dimorphism for Ar and only a modest twofold difference between the sexes for Esr1 (Fig. S4f). None of these factors were regulated by Ncoa1 or induced by CL316,243 (Fig. S4f). No major differences were observed for any member of the nuclear receptor family known to be important for SAT functions (Fig. S4f–g), except for Ppara, which showed moderately higher levels of gene expression in females compared to males (Fig. S4f). Extending our search to other transcription factors, we finally found an interesting sex-biased candidate, Gata3, whose basal gene expression in the SAT was higher in WT female mice than in WT male mice (Fig. 4f), and whose action in beiging has been reported in vivo56. To further characterize the molecular mechanism underlying the actions of Ncoa1 and Gata3 on basal thermogenesis gene activity, we first showed that Gata3 was able to induce the proximal promoter of Ucp1 (Fig. 4g), by acting synergistically with the coactivator Ncoa1 to functionally drive the basal transcription of Ucp1 gene (Fig. 4h). Furthermore, we found that in fully differentiated adipocytes, Ncoa1-dependent induction of Ucp1 gene expression was significantly reduced by selective knockdown of Gata3 gene expression (Fig. S4h). Co-immunoprecipitation assays demonstrated the physical presence of Gata3 in Ncoa1 protein complexes (Fig. S4i). Finally, chromatin immunoprecipitation (ChIP) experiments performed on SAT deposits from WT mice confirmed the co-occupancy of Ncoa1 and Gata3 on the proximal promoter of the endogenous Ucp1 gene specifically in females and not in males (Fig. 4i) and successfully demonstrated the in vivo physical colocalization of Ncoa1 with Gata3 on this promoter (Fig. S4j). Interestingly, in the SAT of female Ncoa1 KO mice, Gata3 binding was drastically reduced but not abolished (Fig. 4i), confirming the dynamic role of these two transcriptional actors in their mutual in vivo binding to the proximal Ucp1 promoter. Concurrent analysis of the human GTEx data revealed higher expression of the human GATA3 gene in the SAT of women compared to men (Fig. 4j), with no significant difference in the visceral WAT (VAT) (Fig. 4j). The gene expression profiles of human GATA3 detected in mature adipocytes isolated from the SAT of premenopausal and menopausal women from the ADIPENDO study (NCT0267282649), were similar to those observed for human NCOA1, with very similar mRNA levels (Fig. S4k). In female WT mice, Gata3 gene expression was markedly lower in the BAT than in the SAT, with near-zero expression in the BAT of male WT mice (Fig. 4k). We then decided to check whether the chromatin organization of the endogenous Gata3 promoter region in SAT could explain the marked dimorphic expression of this transcription factor. In particular, using ATAC-seq profiling of SAT from WT mice, we found that the proximal region of Gata3 promoter displayed a more accessible chromatin conformation in females than in males (Fig. 4l), providing a molecular explanation for Gata3 marked dimorphic expression in the subcutaneous fat depot. While the overall distribution of open chromatin regions across the genome appears to be very similar in SAT from female and male WT mice (Fig. S4l), the total number of peaks was twice as high in SAT from females as in SAT from males (Fig. S4l), demonstrating a significant sex difference in the overall accessibility of chromatin in the SAT.
Finally, to confirm the physiological relevance of our observations, the effect of Ncoa1 deletion on global energy expenditure was assessed in vivo using indirect calorimetry in 16- to 20-week-old male and female mice. Interestingly, the absence of Ncoa1 resulted in a modest but significant decrease in energy expenditure in females compared to their WT female littermates (Fig. 4m–o), but no noticeable effect was observed in males (Fig. 4p–r), except for a slight, non-significant increase at night (Fig. 4p). Locomotor activity and body weight in 16- to 20-week-old Ncoa1 KO animals remained unchanged in both sexes (Fig. 4s, t).
Female Ncoa1 KO mice are overweight and exhibit a male-like profile of WAT, with a fat distribution in visceral depots and a marked decrease in beiging
To investigate whether sex-specific effects of Ncoa1 might be associated with variations in more global metabolic traits such as body weight and/or fat mass, we first monitored the body weight of female and male Ncoa1 KO and WT mice over a 12-month period. The mice were fed regular chow diet ad libitum and were housed at 22-24 °C, to maintain steady activation of the beiging program in WAT depots. At 12 months of age, female Ncoa1 KO mice, but not male mice, exhibited a net increase in their body weight, compared with age-matched WT female littermates (Fig. 5a, b). Next, we analyzed the whole body composition of 12-month-old female Ncoa1 KO and WT mice of both sexes, using quantitative magnetic resonance imaging (MRI). A marked increase in total fat mass was observed in female Ncoa1 KO mice compared to age-matched WT females, with no change in male mice between genotypes (Fig. 5c). In contrast, lean mass was unchanged in both sexes and genotypes (Fig. 5c). Advanced micro-computed tomography (micro-CT) imaging revealed that the increase in fat mass in female Ncoa1 KO mice was essentially due to a marked accumulation of fat in the VAT (Fig. 5d, f), suggesting a potential role for the coactivator Ncoa1 in fat distribution in vivo. As expected from our MRI data, no differences were observed in male mice (Fig. 5g, h). Histological analysis of VAT from Ncoa1 KO females did not reveal any significant change in adipocyte size (Fig. S5a–c), whereas the expression of key adipogenic genes such as peroxysome proliferator receptor gamma (Pparg), fatty acid binding protein 4 (Fabp4) and glucose transporter 4 (Glut4) was upregulated, as assessed by qRT-PCR (Fig. S5d), suggesting that the increase in total fat volume was mainly due to adipocyte hyperplasia. In contrast, in the SAT of 12-month-old Ncoa1 KO females, adipocyte size was significantly increased (Fig. S5e–g), but not accompanied by any change in the expression levels of key adipogenic markers (Fig. S5h), highlighting the occurrence of adipocyte hypertrophy in the SAT of aged female mice lacking Ncoa1. Notably, in both SAT and VAT, the expression of key pro-inflammatory cytokines, such as tumor necrosis factor alpha (Tnf) and interleukin-1 beta (Il1ß) was increased in 12-month-old Ncoa1 KO females, compared with WT littermates (Fig. S5d–h).

a Body weight gain of 12-month-old female (n = 8 / genotype) and male (n = 12 / genotype) Ncoa1 KO mice compared with sex-matched WT littermates. b Body weight of 12-month-old female (n = 8) and male (n = 12) WT and Ncoa1 KO mice (left) and representative image of 12-month-old female WT and KO mice (right). c MRI quantification of fat and lean mass in 12-month-old female (WT: n = 4; Ncoa1 KO: n = 4) and male mice (WT: n = 4; Ncoa1 KO: n = 5). d X-ray CT images of fat mass (orange) in 12-month-old female WT (n = 6) and Ncoa1 KO (n = 7) mice. Sagittal 3D reconstructions at L2/L4 and L5/S1 and longitudinal images from L1 to S4 are shown. e, f X-ray CT quantification of SAT and VAT in 12-month-old female WT (n = 6) and Ncoa1 KO (n = 7) mice. g, h X-ray CT quantification of SAT and VAT in 12-month-old male WT (n = 4) and Ncoa1 KO (n = 4) mice. i Hematoxylin–eosin staining of SAT from 12-month-old female WT (n = 6) and Ncoa1 KO (n = 7) mice. Arrows indicate multilocular adipocytes. Scale bar: 50 μm. j qRT-PCR analysis of Ucp1 (females: WT n = 4, Ncoa1 KO n = 6; males: WT n = 3, Ncoa1 KO n = 4) and Cidea (females: WT n = 4, Ncoa1 KO n = 6; males: WT n = 6, Ncoa1 KO n = 4) expression in SAT from 12-month-old female and male WT and Ncoa1 KO mice. k qRT-PCR analysis of Ucp1 (females: WT n = 5, Ncoa1 KO n = 6) and Cidea (females: WT n = 5, Ncoa1 KO n = 7) expression in VAT from 12-month-old female mice. l qRT-PCR in differentiated adipocytes derived from SAT SVF of 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). Values are mean ±SEM. One-way ANOVA with Tukey’s post hoc test was applied for panels a - b; unpaired two-sided t-test for panels c, e–h, and j–l. Source data provided as a Source Data file.
Hematoxylin and eosin tissue staining of the SAT of 12-month-old female Ncoa1 KO mice revealed the absence of a beige profile, whereas islets of multilocular cells were clearly visible in the SAT of WT females of the same age (Fig. 5i). As seen in younger animals (Fig. 3j–l), a significant decrease in Ucp1 and Cidea mRNA levels was observed in the SAT of 12-month-old female Ncoa1 KO mice, compared with age-matched WT females (Fig. 5j), whereas males of the same age showed no change. A significant decrease in Ucp1 and Cidea gene expression was also observed in the visceral perigonadal WAT depot of aged female Ncoa1 KO mice (Fig. 5k). To exclude any potential influence of other tissue compartments under conditions associated with ageing, SVF cells from SAT depots of 12-month-old Ncoa1 KO females and their sex-matched WT littermates were differentiated with the appropriate white and beige differentiation cocktails, as previously done in younger mice (Fig. 2a–k). A clear effect of Ncoa1 on Ucp1 and Cpt1b gene expression (Fig. 5l) was again observed, with no effect on Plin1 gene expression (Fig. S5i), clearly demonstrating a cell-autonomous effect of Ncoa1 on the beiging program.
Overweight female Ncoa1 KO mice develop glucose intolerance and liver steatosis with age
To fully explore the mechanisms contributing to the progressive weight gain observed in female Ncoa1 KO mice, food intake, energy expenditure, and locomotor activity were monitored in 12-month-old female Ncoa1 KO and WT mice using metabolic cages over three consecutive days. No significant difference in food intake was observed between the two genotypes (Fig. 6a–c), although a slight reduction in energy expenditure was detected in 12-month-old Ncoa1 KO females compared to their sex-matched WT counterparts (Fig. 6d). Additionally, we found a significant reduction in locomotor activity in female Ncoa1 KO mice compared to sex-matched WT mice (Fig. 6e, f and S6a, b). Histological examination of BAT from 12-month-old female Ncoa1 KO mice showed no visible fat accumulation (Fig. 6g). Expression profiles of key players involved in BAT biology, such as Ppargc1a and Prdm16, were further assessed by qRT-PCR and no obvious differences were observed, compared with those of WT females (Fig. 6h). We found, however, a significant transcriptional upregulation of Tfam, Pparg and Plin1 and, even more surprisingly, an induction of Ucp1, and Fgf21 gene expression in the BAT of 12-month-old Ncoa1-deprived female mice (Fig. 6h). Such transcriptional responses from genes involved in energy dissipation in the BAT were most likely indicative of compensatory mechanisms to limit weight gain and/or maintain body temperature, since BAT activity was not found to be intrinsically altered in Ncoa1 KO females (Fig. 6i, j and S6c). Indeed, the body temperature of Ncoa1-deficient female mice was similar to that of sex-matched WT littermates in several physiological situations, including overnight fasting (16 h), and when subjected to a cold room challenge (Fig. 6i, j).

a, b Food intake measured over 3 days (white) and 3 nights (gray) in 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). c Food intake measured across 7 consecutive days in 12-month-old female WT and Ncoa1 KO mice (n = 4 per group). d Energy expenditure recorded over 24 h in 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). e, f Locomotor activity (average total distance over 24 h) in 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). White/black bars represent day/night. g Representative H&E staining of BAT from 12-month-old female WT and Ncoa1 KO mice. Scale bar: 50 μm. h qRT-PCR analysis of metabolic genes in BAT from 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). i Rectal temperature after 16 h fasting in 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). j Cold chamber test in 12-month-old female WT (n = 5) and Ncoa1 KO (n = 7) mice for 3 h. k Glucose tolerance test (GTT) showing impaired glucose tolerance in 12-month-old female Ncoa1 KO mice vs. WT (n = 6 per group). l GTT in 12-month-old male WT and Ncoa1 KO mice showing no difference (n = 6 per group). m–o Plasma glucose in (m) (WT n = 5, Ncoa1 KO n = 6), insulin in (n) (n = 6 per group), and cholesterol in (o) (n = 6 per group) in 12-month-old female WT and Ncoa1 KO mice. p Representative H&E and Oil Red O staining of liver from 12-month-old female WT and Ncoa1 KO mice, showing steatosis in Ncoa1 KO mice (n = 6 per group). Scale bar: 50 μm. q Liver triglyceride quantification in 12-month-old female WT (n = 6) and Ncoa1 KO (n = 7) mice. r qRT-PCR of lipid and cholesterol metabolism genes in liver from 12-month-old female WT and Ncoa1 KO mice (n = 6 per group). s Western blot of Akt and p-Akt (Ser473) in liver of 12-month-old female WT and Ncoa1 KO mice, with or without insulin injection (1.0 U/kg, 10 min). Two independent biological replicates were loaded for each condition on the same blot. Hsp90 served as a loading control. Data are mean ±SEM. Unpaired two-sided t-test used in panels a–c, e–f, h–o, and q, r. ns: not significant. Source data provided as a Source Data file.
Finally, to determine whether the weight gain observed in 12-month-old female Ncoa1 KO mice affected glucose homeostasis in vivo, we performed glucose tolerance tests. A clear difference in glucose utilization was observed in female Ncoa1 KO mice, compared with sex-matched WT littermates (Fig. 6k). In males, no difference was observed between Ncoa1 KO and WT animals (Fig. 6l). Under fasting conditions, insulin and glucose levels were clearly elevated in the blood of female Ncoa1 KO mice, compared with those of WT littermates, suggesting an apparent glucose intolerance (Fig. 6m, n). In addition, serum total and HDL cholesterol levels were increased in female Ncoa1 KO mice compared with WT controls (Fig. 6o). Furthermore, steatosis was detected in the livers of 12-month-old female Ncoa1 KO mice, as evidenced by hematoxylin-eosin, Oil Red O staining, along with elevated intrahepatic triglycerides levels (Fig. 6p, q). This was accompanied by a marked transcriptional upregulation of lipogenic genes, including Srebp-1c, Fasn, and Pparg (Fig. 6r). Interestingly, the expression of key fatty acid oxidation genes, such as Cpt1a, Acadm, Acadl, and Ppara, was also increased in the livers of 12-month-old female Ncoa1 KO mice compared to WT (Fig. S6d). Additionally, hepatic insulin resistance was indicated by a significant reduction in the phosphorylation status of Akt (Ser473) protein in the liver following insulin injection (Fig. 6s) and under random fed conditions (Fig. S6e).
Discussion
The role of sex is an important issue in the incidence and evolution of type 2 diabetes, obesity, and metabolic syndrome1,57,58. However, despite increased attention in recent years, many clinical trials continue to neglect sex-specific considerations, and preclinical research relies heavily on male animals and cells, obscuring important sex differences that could guide clinical trials59. As women appear to be protected against metabolic syndrome and related diseases, at least before menopause5, we speculated that a close examination of biological events occurring in females would likely unveil major circuits involved in providing these protective effects. The present study uncovered a pivotal sex-specific action exerted by the transcriptional coregulator Ncoa1 on the control of energy utilization, with implications for mammalian physiology and human health. Genetic deletion of Ncoa1 had a profound impact on energy homeostasis in females, with only a modest effect in males. In contrast to age-matched WT females, female Ncoa1 KO mice gradually developed obesity, hepatic steatosis, and glucose intolerance, while males grew normally. As a plausible molecular explanation for the dimorphic metabolic traits of our Ncoa1 KO mice, we deciphered a cell-autonomous role of Ncoa1 in WAT remodeling and energy use, primarily through sex-selective transcriptional regulation of the key thermogenic player Ucp1 and other important metabolic genes, including Cidea and Cpt1b. We have shown that, in the female SAT, Ncoa1 sustains a heightened constitutive transcriptional activity of the key genes involved in the beiging program, whereas in males, Ncoa1 is exclusively involved in the acute beta-adrenergic-dependent transcriptional induction of genes. Thus, our findings provide a major clarification to explain the sex-linked mechanisms of beige adipocyte emergence in WAT. Although reported several years earlier in mice28, the sex dimorphism of beiging was first demonstrated at the molecular level by the report of an in vivo activity of a Ucp1 reporter transgene occurring exclusively in female SAT, after cold exposure29. More recently, browning activity in mice was found to be higher in female peri-ovarian WAT than in male epididymal WAT31. In humans, UCP1 gene expression has also been shown to be sex-dependent in SAT, with higher UCP1 expression in women than in men12,32,33. At first, the sex disparity in UCP1 expression levels could be viewed as an evolutionary protective advantage for maintaining body temperature, as non-obese premenopausal women have, on average, a higher surface-area-to-volume ratio than men, requiring more effective thermogenic capacity to maintain body temperature within physiological range. However, the contribution of beige adipose tissue to systemic thermoregulation remains controversial, especially in humans23,60,61. Recently, it has been hypothesized that WAT beiging may primarily mobilize fatty acids to provide energy to other organs, in response to negative energy balance62. For example, during exercise or muscular activity, WAT beiging is induced to fuel skeletal muscle with lipids, allowing for increased metabolic capacity63,64. In this regard, it is well established that females rely more heavily on fat oxidation during exercise than males65. The preferential use of fat during exercise in females occurs in parallel with an increase in mitochondrial volume and intrinsic fatty acid oxidation in skeletal muscle fibers65. It should be added that female rodents are more active than males66. Thus, it can be hypothesized that the more pronounced beiging profile of WAT in females may also be due to more efficient lipid mobilization to fuel skeletal muscle, potentially compensating for the deficit in total skeletal muscle mass found in women compared to men, and/or the greater physical activity found in female compared to male mice.
The coactivator NCOA1 (also known as SRC-1 or RIP160) was one of the first transcriptional coactivators identified to interact with sex steroid receptors to enhance their transactivation in a ligand-dependent manner67. As reported in the GTEx data, human NCOA1 is highly expressed in reproductive organs, particularly in women (ovary: TPM = 45.52; endocervix: TPM = 41.9; uterus: TPM = 39.13; Fallopian tube: TPM = 38.25), in some regions of the brain (cerebellar hemisphere: TPM = 42.07; cerebellum: TPM = 38.55) and in the SAT (TPM = 30.56), but is expressed at much lower levels in the renal cortex (TPM = 10.10) and pancreas (TPM = 8.09). In mice, a similar tissue expression profile was observed across several tested tissues (Fig. S6f). Ncoa1 is also an important modulator of the activity of Ppargc1a (alias Pgc1α), a key coregulator involved in thermogenesis, working in tandem to fine-tune target gene transcriptional activity and modulate the epigenomic chromatin landscape, including promoters and enhancers68. The metabolic role of Ncoa1 was first highlighted by the characterization of a novel Pparg modulator, FMOC-L-Leu69. When administered in vivo, FMOC-L-Leu improves insulin sensitivity in diet-induced and db/db glucose-intolerant mice, to a similar extent as the thiazolidinediones. Further support for the role of Ncoa1 in metabolic control came from the characterization of Ncoa1-deficient animals, as male Ncoa1 KO mice are prone to obesity on a high-calorie diet, with residual impairment of BAT activity despite increased caloric intake compared to WT males. Altogether, Ncoa1 possesses multiple intrinsic molecular properties that allow it to act as a pleiotropic player capable of finely orchestrating sex-specific differences in energy balance. Indeed, we have provided evidence that depletion of Ncoa1 strongly affects sex-dependent biological characteristics of the WAT. Firstly, a marked decrease in the beige adipocyte program was observed in the subcutaneous fat of female Ncoa1 KO mice, compared to age-matched WT female littermates, knowing that WAT beiging is much more active in females than in males, both in rodents and humans30,32. Secondly, female Ncoa1 KO mice showed an overall fat distribution in favor of visceral depots, which is a typical feature of males70. Third, while genes encoding key proinflammatory mediators, such as tumor necrosis factor alpha (Tnf) and interleukin 1 beta (Il1b), were not expressed in the SAT of young female mice, in line with the known protection provided by the SAT against inflammatory insults, particularly in females71,72, these proinflammatory markers were, detected in the SAT of aged Ncoa1 KO females, which thus resembled male profiles. Taken together, our observations lend support to the pivotal role played by Ncoa1 as a selective gatekeeper of the most important sex-dependent metabolic features of WAT depots.
As Ncoa1 has no apparent intrinsic dimorphic properties, we searched for potential molecular partners capable of mediating its sex-dependent effect on the SAT beiging transcriptional program. An unbiased RNAseq analysis performed in female and male SAT depots revealed the transcription factor Gata3 as a potential candidate. Indeed, with a pronounced ‘female-biased’ dimorphic gene expression profile in the subcutaneous WAT depot (Fig. 4f), Gata3 can synergistically transactivate Ucp1 promoter activity with Ncoa1, in a cellular context (Fig. 4g, h) and selectively bind the endogenous Ucp1 proximal promoter in female mouse adipocytes (Fig. 4i). Known as a master regulator of immune cell differentiation, particularly Th2 cell commitment73, Gata3 has previously been described as a regulator of white adipogenesis74. More recently, Gata3 has been implicated in the activation of the thermogenic program in vivo through a functional interaction with Ppargc1a (alias Pgc1α), both acting directly on the proximal Ucp1 promoter56. Interestingly, Gata3 gene expression, protein levels and nuclear localization are strongly stimulated by 17β estradiol (E2) through the nuclear receptor ER alpha in mature adipocytes75. Thus, with a well-characterized pivotal role in key biological functions such as immune function and adipocyte differentiation, but also being directly stimulated by female sex steroid hormones in adipocytes, Gata3 is a very attractive candidate for fine-tuning sex-dependent hierarchical genetic programs, such as the beiging adaptation of the SAT.
While investigating the physiology of the BAT, the main tissue involved in thermoregulation in rodents, we unexpectedly observed a marked upregulation in female Ncoa1 KO mice of key genes involved in the adaptive thermogenic program, namely Tfam, Pparγ, and Ucp1. This finding most likely indicates the existence of compensatory mechanisms aimed at restoring “normal” energy homeostasis throughout the body of female Ncoa1 KO mice. Previous independent studies have clearly shown that defects in BAT activity are able to induce compensatory mechanisms that result in the induction of beiging adaptation in subcutaneous WAT76. Consistently, we found significantly increased levels of Fgf21 gene expression in the BAT of female Ncoa1 KO mice, a factor known to be released in high levels by the BAT to promote thermogenesis and metabolite oxidation77. In addition, Fgf21 has been shown to be the major endogenous peptide that causes the paradoxical resistance to obesity in Ucp1 KO mice78. Interestingly, Fgf21 gene expression was found to be dimorphic, since in the BAT of mice fed an obesogenic diet, the Fgf21 gene was induced more in males than in females79. Taken together, these observations confirm the key role of Ncoa1 in energy homeostasis in female mice, but also highlight the existence of additional underappreciated sex-specific mechanisms underlying energy balance adaptation.
Our study also revealed that upon β3 agonist induction, the coregulator Ncoa1 acts as an essential transcriptional coordinator of large hierarchical sets of genes involved in mitochondrial functions. Indeed, our unbiased RNAseq screen has identified that Ncoa1 orchestrates the selective β3 induction of (i) genes encoding multiple subunits of all the respiratory chain complexes; (ii) genes involved in the formation and the stability of the respiratory chain complexes, such as Bola3, Phb1 and Phb2; (iii) genes involved in cristae formation and stabilization, such as Chchd10 (Fig. S3d); (iv) central genes directly acting on the mitochondrial transcription and translation, such as Polrmt and Tufm (Fig. S3e); and finally (v) several genes encoding mitoribosomes. Notably, a functional link between NCOA1 and mitochondrial transcription in humans has already been suggested, after transcriptional profiling of lymphocytes from large donor cohorts identified NCOA1 as the top gene with the strongest association with human mitochondrial content, a surrogate measure of mitochondrial integrity (p value = 7.4e-8; rank # 1)80. It is noteworthy that the pleiotropic transcriptional role of Ncoa1 is similar to the molecular mode of action of Ppargc1α, the master coregulator involved in mitochondrial biogenesis41,81. Coordination by key transcriptional coregulators such as Ppargc1α and Ncoa1 following β3 adrenergic stimulation is likely required to fine-tune the broad transcriptional induction of hundreds of nuclear genes responsible for the global metabolic interconversion between white and beige adipocytes. The pronounced effect of Ncoa1 on mitochondrial gene expression levels showed a clear sexual dimorphism and was mainly observed in the SAT of female mice, with only a modest effect in males. Furthermore, we found that the transcription of almost all Ncoa1 target genes was more robustly induced by the β3 agonist in WT female mice than in WT males. This is consistent with previous studies reporting that women and female mice have increased expression of genes involved in mitochondrial function in WAT30,33 and that the sex-dependent mitochondrial gene signature in females appears to be specific to adipose tissue33. In addition, the metabolic rate per kilogram of SAT was shown to be significantly higher in women than in men, a difference partly explained by an increase in the expression levels of the beige adipocyte marker UCP1 in WAT in females compared with males33. It should be noted that this difference is independent of sex steroids and fat distribution33. Although adipose tissue is clearly not the major determinant of energy expenditure compared to skeletal muscle, brain, and heart, the limited impact of sex differences on the contribution of fat mass to basal or induced metabolic rate may be significant in slowly progressive clinical conditions, such as type 2 diabetes or cardiovascular disease. As it has become undeniable that mitochondrial dysfunction contributes to a wide range of human disorders, including metabolic diseases, delineating the underlying molecular mechanisms of sex dimorphic regulation of mitochondrial genes, to define where sex makes a difference, should offer the possibility of identifying protective agents that could help fight metabolic diseases.
In our current effort to investigate the functional role of the transcriptional coregulator Ncoa1 during WAT beiging in vivo, we used a mouse model with global genetic disruption of Ncoa1. The cell-autonomous action of Ncoa1 that we report in isolated mature subcutaneous white adipocytes unequivocally highlights its pivotal role in the sexually dimorphic control of adipose beiging. However, we emphasize that these findings do not exclude potential contributions from other surrounding tissues to the progressive development of the metabolic phenotype observed in aged female Ncoa1-depleted mice. It is important to note that there is currently no transgenic mouse strategy to ablate a gene specifically in subcutaneous WAT, as all existing Cre recombinase mouse strains (namely Adiponectin-Cre and Ucp1-Cre mice) disrupt target genes in both WAT and BAT. Furthermore, obesity and insulin resistance are complex pathophysiological conditions involving multiple interrelated organs and cannot be fully recapitulated by deleting a single gene in a single tissue. Additionally, we acknowledge that we have not investigated the upstream signal(s) that may modulate the sexually dimorphic effects of Ncoa1 during beiging adaptation. Thyroid hormones, natural circulating steroid hormones (such as estradiol and glucorticoids), and other factors have been implicated in the control of adipose tissue formation and could potentially contribute to the effects of Ncoa1 observed in this study. The role of intra-adipogenic steroids in this process also cannot be ruled out. While defining the exact hormones and/or metabolites involved in the specific action of Ncoa1 is beyond the scope of this study, it certainly warrants further investigation in the future. Finally, we have provided strong evidence supporting a key role for Gata3, in concert with Ncoa1, in the selective modulation of basal thermogenic tone in female mice. However, we recognize that further work is needed to functionally validate these findings in humans.
By revealing Ncoa1-dependent effects on energy balance and health, our study reaffirms the idea that targeting transcriptional coregulators and/or their selective transcription factors in metabolic tissues of interest represents an attractive therapeutic avenue to combat metabolic dysfunction82. In this regard, an independent gene expression profiling study in large mammals demonstrated that Ncoa1 gene expression is significantly downregulated during the first phase of postnatal beige-to-white adipose tissue transformation in sheep perineal fat83, a complex tissue remodeling similar to the transition occurring in human infants11,83. Furthermore, recent exome sequencing identified 15 rare human variants of NCOA1 in severe childhood-onset obesity84. An additional confirmatory study expanded the number of NCOA1 genetic variants in human obesity85. Clearly, these observations, combined with our own results, highlight a unique role for the transcriptional coregulator Ncoa1 in energy homeostasis to fine-tune energy fluxes in vivo. Finally, the identification of underappreciated sex-specific differences in metabolic function and disease incidence provides additional arguments to pave the way for sex-based personalized medicine.
Methods
Human subjects and adipose tissue samples
Subcutaneous gluteofemoral adipose tissue was obtained from premenopausal (n = 8; median age: 31.4 + /- 1.6 years; median BMI: 26.0 + /- 1.2 kg/m2) and metabolically healthy postmenopausal women (n = 6; median age: 58.8 + /- 1.6 years; median BMI: 27.8 + /- 0.8 kg/m2), programmed for esthetic dermolipectomy at the University Hospital of Toulouse and enrolled in the interventional cohort study ADIPENDO, registered under the unique identifier: NCT02672826 on ClinicalTrials.gov (https://www.clinicaltrials.gov). This study was approved by the Institutional Review Boards. All subjects gave written informed consent. The protocol used to isolate mature adipocytes from subcutaneous fat was previously described in detail in ref. 49. Additional human RNA seq data were obtained from: (1) the Genotype-Tissue Expression (GTEx) project (https://www.gtexportal.org/home/;34); (2) the transcriptomic dataset GSE11376447, which specifically includes the analysis of total RNA from SAT isolated from the supraclavicular region of non-diabetic, non-obese and premenopausal women (n = 12; median age: 43.8 + /- 1.9 years; median BMI: 26.0 + /- 1.5 kg/m2); and (3) the DiOGenes study50, a pan-European, randomized, and controlled trial investigating the longitudinal effects of dietary intervention on weight gain, metabolic and cardiovascular risk factors in obese and overweight non-diabetic families in eight European centers. Participants underwent an 8-week low-calorie diet (LCD = 800 kcal/day), followed by a 6-month weight control diet (CD). DiOGenes was registered in ClinicalTrials.gov (NCT00390637). We closely analyzed RNA sequencing data from obese, non-diabetic women (n = 93; median age: 42.0 + /- 0.62 years; median BMI: 34.5 kg/m2 +/- 0.5) and obese non-diabetic men (n = 46; median age: 43.3 ±0.94 years; median BMI: 33.96 kg/m² ± 0. 63) from the DiOGenes study. Total RNA was prepared from frozen abdominal subcutaneous adipose tissue needle biopsies. RNA extraction and data processing are described in detail in ref. 51. Expression data are available in the Gene Expression Omnibus under accession GSE95640.
Animal care
Ncoa1 KO mice and WT mice were generated from heterozygous breeders and maintained as littermates on a pure C57BL/6JRj background. Genotyping was performed as previously described86. Mice were group-housed by sex under controlled conditions (22–24 °C; 12-hour light/dark cycle) with ad libitum access to water and standard rodent chow (SAFE® D04, UAR, Villemoisson-sur-Orge, France). All experimental procedures complied with European guidelines for the care and use of laboratory animals and were approved by the French Ministry of Agriculture’s National Committees (protocol agreement no. 02470.02; animal facility agreement no. B06-088-20).
Mouse and human adipose tissue SVF preparation and differentiation
Mouse subcutaneous WAT depots were harvested, washed in PBS, and minced. Adipose tissue samples were digested for 45 min at 37 °C in DMEM containing 2 mg/ml collagenase A (Roche Diagnostics, Meylan, France) and 20 mg/ml bovine serum albumin (Sigma-Aldrich Chimie). Cell lysates were successively filtered through 250, 100, and 27 µm nylon sheets, and centrifuged at 500 g for 5 min at room temperature. The pellet containing stromal vascular fraction (SVF) cells was subjected to a red blood cell lysis procedure. The SVF cells were plated and maintained in DMEM containing 10 % FBS until confluence. Differentiation was induced in the same medium supplemented with 1 µM dexamethasone, 0.5 mM isobutylmethylxanthine, and 170 nM insulin and 2 nM triiodothyronine for 2 days. The cells were then maintained for 10 days in the presence of 10 nM insulin for white adipogenesis or 10 nM insulin, 1 µM rosiglitazone, and 2 nM triiodothyronine for beige adipogenesis. Cells were exposed to 1 µM isoproterenol for 2 hours (Iso) or without isoproterenol (Ctrl). Human adipose tissue SVF cells from abdominal subcutaneous fat were collected, isolated according to the procedure described for mice (see above) and differentiated in serum-free medium as previously described53,54.
Human multipotent adipose-derived stem cell (hMADS) culture
The establishment and characterization of hMADS cells have been well described previously52. Briefly, cells were seeded at a density of 5000 cells/cm2 in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS, 15 mM Hepes, and 2.5 ng/ml hFGF2. hFGF2 was removed when cells reached confluence. Cells were induced to differentiate at day 2 post confluence (referred to as day 0) in DMEM/Ham’s F12 (1:1) media supplemented with 10 μg/ml transferrin, 10 nM insulin, 0.2 nM triiodothyronine, 1 μM dexamethasone, and 500 μM isobutyl-methylxanthine. Two days later, the medium was refreshed, but dexamethasone and isobutyl-methylxanthine were removed, and 100 nM rosiglitazone was added for twelve days.
Cell culture and transient transfections
Hela cells (RRID:CVCL_0030) were obtained from the American Type Culture Collection (ATCC). Transient transfections were performed using Transfectin reagent (Bio-Rad) diluted in OptiMEM medium (Thermo Fisher Scientific, MA, USA) with the indicated expression vectors and reporter plasmids. Reporter gene luciferase levels were determined 48 hours after transfection, according to the manufacturer’s instructions (Promega). Results were normalized to total proteins using the BCA protein assay kit (Pierce, Rockford, IL, USA). 3T3-L1CARdelta1 cells were obtained by direct request87. Cells were plated and grown to confluence in DMEM containing 10 % fetal bovine serum. Differentiation was induced for 2 days in the same medium supplemented with 1 µM dexamethasone, 0.5 mM isobutylmethylxanthine, and 860 nM insulin, and then maintained for 7 days in the presence of 860 nM insulin and 1 µM rosiglitazone. Gata3 knock-down was realized using Lipofectanine RNAiMax reagent (Thermo Fisher Scientific, MA, USA) diluted in OptiMEM medium (Thermo Fisher Scientific, MA, USA) with ON-TARGETplus mouse Gata3 SMARTPool siRNA (Horizon Discovery) and ON-TARGETplus non-targeting control siRNAs (Scramble). For over-expression, Ncoa1 adenovirus (Applied Biological Materials, Canada) was used at 109vp/ml for 48 h.
Metabolic phenotyping
Systemic administration of the ß3-receptor agonist consisted of a daily intraperitoneal injection of CL316,243 (0.5 mg/kg body weight, in saline solution (NaCl)) for 7 consecutive days. For cold experiments, mice were placed in a 6 °C cold chamber (A-Box 80, Noirot, Suresnes, France) for 4 h or 7 days or kept at room temperature (24 °C), and their rectal temperature was assessed. Indirect calorimetry was used to determine energy expenditure, food intake and locomotor activity. The mice were individually housed in calorimetric tidal cages (OxyletPro Physiocage, Panlab-Bioseb, Vitrolles, France) and acclimatized for 24 h before the experimental measurements. They were then monitored for 24 h or 48 h with free access to food and water. For the cold tolerance test, mice were exposed to 4 °C for 3 h, and their rectal temperatures were monitored every 1 h with a digital thermometer. For glucose tolerance tests (GTT), mice were fasted for 6 h and tail blood glucose concentrations were measured using a commercially available OneTouch Ultra blood glucose meter and test strips (LifeScan). Mice received an i.p. injection of glucose solution (Sigma-Aldrich) at a dose of 1.5 g glucose/kg body weight. Tail blood glucose was measured at 0, 15, 30, 60, and 120 min post injection. Serum total cholesterol, LDL-cholesterol and HDL-cholesterol levels were measured using Biosentec kits and a Mindray analyzer (BS-240Pro). Plasma glucose and insulin were measured as described in ref. 88. For lipolysis measurements, differentiated adipocytes were insulin deprived overnight. Fresh medium was added, and the cells were immediately exposed to 1 µM isoproterenol for 2 hours. Sampled medium was used to measure glycerol release using free glycerol reagent (Sigma-Aldrich) according to the manufacturer’s instructions. The results were normalized to the amount of protein using the BCA protein assay kit (Pierce, Rockford, IL).
MRI imaging and micro-computed tomography
Body weight was monitored, and body composition was analyzed using a Lunar PIXImus densitometer (Lunar Corp., Madison, WI) equipped for dual energy X-ray absorptiometry (DEXA). Anatomical MRI experiments were performed using a Bruker Pharmascan 7.0-T spectrometer, 16 cm bore, horizontal imaging system (Bruker Biospin, Billerica, MA) with a 38 mm volume resonator. All imaging sessions were breath controlled. During imaging, animal body temperature was maintained at 37.0 °C using an animal heating system (SA Instruments, Stony Brook, NY). For each mouse, images were acquired for 20 axial slices and 20 coronal slices at a thickness of 1.0 mm. To obtain the best contrast for adipose tissue, a T1-weighted spin-echo sequence was used with the following parameters: repetition time = 500 ms; echo time = 10.7 ms; field of view = 50.0 mm × 45.0 mm (coronal slices) and 45.0 × 45.0 mm (axial slices); matrix size of 256 × 256; two signal averages were acquired.
Adipose tissue quantification of anesthetized mice was performed using a SkyScan-1178 X-ray micro-computed tomography (micro-CT) analysis. Briefly, mice were scanned using the following parameters: 104 μm voxel size, 49 kV, 0.5 mm thick aluminum filter, and a rotation step of 0.9°. Total adipose tissue volume was measured between lumbar vertebra 1 (L1) and caudal vertebra 4 (C4), while intra-abdominal and subcutaneous adipose tissue areas were measured on one slice at lumbar 5 (L5) level. Adipose tissue areas and volumes were analyzed using NRecon and CTAn software and 3D visualizations using CTVox software (Bruker microCT, Kontich).
Bulk RNA sequencing
Frozen mouse subcutaneous adipose tissue was homogenized in RLT buffer (Qiagen), and RNA extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s instructions (Qiagen). Total RNA were submitted to BGI genomics (Hong Kong, China; https://www.bgi.com/global) for library preparation and sequencing on a BGISEQ-500 platform (Paired-end mode, 30 M reads per sample). Briefly, oligo(dT)-tagged magnetic beads were used to purify mRNA. Purified mRNA was fragmented into small pieces using the fragment buffer at the appropriate temperature. First-strand cDNA was generated in the first-strand reaction system by random hexamer-primed reverse transcription, and second-strand cDNA was generated in the strand synthesis reaction system (using dUTP instead of dTTP). The reaction product was purified by magnetic beads, and then A-Tailing mix and RNA index adapters were added by incubation to complete the repair. The cDNA fragments with adapters obtained from the previous step were amplified by multiple rounds of PCR, to digest the U-labeled second-strand template with UDG enzyme. The PCR products were purified with XP Beads and dissolved in EB solution. The library was quality assessed to ensure the high quality of the sequencing data and the fragment size distribution was checked using the Agilent 2100 Bioanalyzer. The double-stranded PCR products were heat-denatured and circularized using the splint oligo sequence. The single-stranded circular DNA (ssCir DNA) was formatted as the final library. The library was amplified with phi29 to produce DNA nanoballs (DNBs) with more than 300 copies of a molecule. The DNBs were loaded onto the patterned nanoarray, and pair-end 150-base reads were sequenced by combinatorial probe-anchor synthesis (cPAS) on DNBseq. Raw reads were quality controlled, and the raw data were filtered using the filtering software SOAPnuke (v1.5.2). The filtered clean reads were aligned to the reference genome sequence (mouse genome GRCm38 (mm10)) using HISAT2 software (v2.0.4). Bowtie2 software was used to align the clean reads to the reference gene sequences, and RSEM were used to calculate the gene expression levels in each sample. Fragments per kilobase of exon model per million mapped fragments (FPKM) and TPM values were used to estimate gene expression levels, and the genes in each group with a mean FPKM value greater than 0.5 were considered to be expressed in the group and incorporated into downstream statistical analysis. Differential gene expression was analyzed with the DESeq2 algorithm, which applies the Benjamini-Hochberg technique for multiple testing correction to compute adjusted p-values89 and differentially expressed genes (DEG) were identified with the following parameters: (1) |log2FC | ≥ 0,25 and (2) q-value ≤ 0.05). KEGG pathway enrichment and gene set enrichment analysis (GSEA) were further performed on BGI’s Dr.Tom system (http://biosys.bgi.com) with the software package provided by the Broad Institute, based on the MSigDB (Molecular Signatures Database). Normalized enrichment score (NES) and false discovery rate (FDR) were used to quantify the enrichment magnitude and statistical significance, respectively ( | NES| is greater than 1.0, nominal p-value below 5%, and the false detection rate (FDR) below 25%).
ATAC-seq
Frozen mouse SATs were submitted to Active Motif for the ATAC-seq assay. They were initially ground to a fine powder using liquid nitrogen and resuspended in an appropriate lysis buffer (Active Motif). Isolated nuclei were quantified using the Countess II FL automated cell counter (Thermo Fisher) and 100,000 nuclei were labeled as previously described90, with some modifications based on91, using the enzyme and buffer provided in the ATAC-Seq kit (Active Motif). The labeled DNA was then purified using the MinElute PCR purification kit (Qiagen), amplified with 10 cycles of PCR, and purified using Agencourt AMPure SPRI beads (Beckman Colter). The resulting material was quantified using Qubit (Invitrogen) and TapeStation (Agilent) and sequenced using PE42 sequencing on the NovaSeq 6000 (Illumina). Reads were aligned using the BWA algorithm (mem mode; default settings). Duplicate reads were removed, and only reads corresponding to matched pairs and unique reads (mapping quality ≥ 1) were used for further analysis. Alignments were extended in silico at their 3’ ends to a length of 200 bp and assigned to 32 nt bins along the genome. The resulting histograms (genomic “signal maps”) were stored in bigWig files. Peaks were identified using the MACS 2.1.0 algorithm with a p-value cut-off of 1e-7, without a control file and with the -nomodel option. Peaks on the ENCODE blacklist of known false ChIP-Seq peaks were removed. Signal maps and peak locations were used as input data to the Active Motif proprietary analysis program, which generates Excel tables with detailed information on sample comparison, peak metrics, peak locations, and gene annotations.
Histological and immunostaining analysis
Pieces of mouse white, brown adipose, and skeletal muscle tissues were fixed in 10 % formalin solution at room temperature, dehydrated, and then embedded in paraffin. Tissue blocks were then sectioned at 5 µm thickness. Sections were deparaffinized, rehydrated, and stained with hematoxylin and eosin. For immunostaining, sections were permeabilized with 0.1% Triton X-100 for 10 min, and then incubated with primary antibody (anti-UCP1, Abcam, ab10983, RRID:AB_2241462, dilution 1:100) overnight at 4 °C and with the secondary antibody coupled to HRP (Invitrogen, dilution 1:500) for 1 hour at RT. The staining was visualized with DAB substrate. Finally, the sections were mounted and visualized with a microscope. Oil red O staining was performed on frozen sections as previously described44.
Chromatin immunoprecipitation (ChIP) and re-chIP
Subcutaneous WAT depots were harvested, washed in PBS, and minced into small pieces. Tissue fixation, homogenization, and chromatin shearing were performed as previously described92. All other chromatin immunoprecipitation steps were performed using the EZ ChIP kit (Millipore) according to the manufacturer’s protocol. The Ncoa1 antibody was from Cell Signaling Technology (#2191; RRID:AB_2196189; dilution 1:1000) and the Gata3 antibody from Abcam; (#ab199428; clone EPR16651; RRID:AB_2819013; 5 μg). qPCR for ChIP was performed using SYBR Green technology (Applied Biosystems) with sequence-specific primers. Results were normalized to the input. Primer sequences are available on request.
Western blot
Proteins were extracted using RIPA lysis buffer with Complete protease inhibitor cocktail (Roche Inc.). Protein concentration was determined using a BCA protein assay kit (Pierce, Rockford, IL). Twenty micrograms of protein from each biological sample was separated by SDS-PAGE and electrotransferred to a nitrocellulose membrane for immunoblot analysis. The following antibodies were used against: Ucp1 (Abcam, ab10983; RRID:AB_2241462; dilution 1:1000); Ncoa1 (Cell Signaling, #2191; RRID:AB_2196189; dilution 1:1000); Akt (Cell Signaling Technology, #9272; RRID:AB_329827; dilution 1:1000) and Phospho-Akt 473Ser (Cell Signaling Technology, #9271; RRID:AB_329825; dilution 1:1000); Hsp90 (Santa Cruz Biotechnology, SC-13119; RRID:AB_675659; dilution 1:500); β-tubulin (Sigma #T5201; RRID:AB_609915; dilution 1:2000); HRP-conjugated anti-mouse (GE Healthcare UK Limited, 1:10,000); anti-rabbit (GE Healthcare UK Limited, 1:10,000). The SuperSignal West Pico Chemiluminescence Kit (Pierce) was used as substrate. For mitochondrial complex quantification, equal amounts of cell proteins were separated by SDS-PAGE gradient (10–20%) and blotted onto nitrocellulose membranes. Saturated membranes were incubated overnight with a 1:1000 dilution of a total OXPHOS human western blot antibody cocktail (Abcam, ab110413; RRID:AB_2629281) followed by incubation with HRP-conjugated anti-mouse immunoglobulins for 60 min. The chemiluminescence obtained after the addition of Clarity Western ECL substrate (Bio-Rad, Marnes-la-Coquette, France) was detected using a ChemiDoc MP imaging system (Bio-Rad) and quantified using Image Lab 5.0 software (Bio-Rad).
Quantification of mitochondrial DNA
Genomic DNA was extracted using a DNA extraction kit (Macherey-Nagel EURL, France). 2 ng of total DNA was used for qPCR analysis, and mitochondrial DNA content was calculated from the ratio of mitochondrial NADH dehydrogenase subunit 1 gene DNA and nuclear lipoprotein lipase nuclear gene DNA. Oligonucleotide sequences are available on request.
Analysis of mitochondrial respiration
SVF cells isolated from mouse SAT were seeded in a 24-well plate and differentiated, as described herein. The oxygen consumption rate (OCR) of differentiated cells was determined using the Seahorse XF24 Extracellular Flux Analyzer (Agilent, France). Isoproterenol (100 nM) was used to characterize β adrenergic inducible respiration. Uncoupled and maximal OCR were determined with oligomycin (1.2 µM) and FCCP (1 µM). Rotenone (1 µM) and antimycin A (1 µM) were used to inhibit complex I- and III-dependent respiration, respectively.
RNA analysis and real-time PCR
Standard RNA extraction procedures (Qiagen RNeasy Mini Kit) were used. Reverse transcription was performed using a high-capacity cDNA reverse transcription kit (Applied Biosystems). For gene expression analysis, qRT-PCR was performed using sequence-specific primers and probes from Roche (Universal Probe Library). The gene RPLP0 (ribosomal protein lateral stalk subunit P0; alias 36B4) was used as an internal control for all gene expression assays. All qPCR primer sequences are available on request.
Co-immunoprecipitation assays
For co-immunoprecipitation, 293 T cells were transiently transfected with NCOA1 and GATA3 plasmid vectors using Transfectin reagent (Bio-Rad) diluted in OptiMEM medium (Thermo Fisher Scientific). Cell lysates were then incubated with anti-GATA3 (Abcam; #ab199428; clone EPR16651; RRID:AB_2819013; 5μg). The immune complexes were eluted and subjected to SDS-PAGE with anti- Ncoa1 (Cell Signaling, #2191; RRID:AB_2196189; dilution 1:1000).
Statistical analysis
All biological experiments were performed in triplicates and repeated at least three times independently with reproducible results. Statistical analyses were performed with GraphPad Prism v.9.5.0. All data are presented as mean ±SEM. Unpaired Student’s t-test was used for pairwise comparisons. Two-way ANOVA was used for comparisons between more than two groups, followed by Fisher’s LSD test or Bonferroni’s post-hoc test. The statistical parameters and sample size used for each experiment are indicated in the corresponding figure legends. Statistical significance was determined when values were equal to or less than 0.05.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
RNA-seq and ATAC-seq data are available in NCBI’s Gene Expression Omnibus under accession numbers GSE308798 and GSE309045, respectively. All data supporting the findings described in this manuscript are available in the article and in the Supplementary Information and from the corresponding authors upon request. Source data are provided with this paper.
References
Clegg, D. J. & Mauvais-Jarvis, F. An integrated view of sex differences in metabolic physiology and disease. Mol. Metab. 15, 1–2 (2018).
Kautzky-Willer, A., Leutner, M. & Harreiter, J. Sex differences in type 2 diabetes. Diabetologia 66, 986–1002 (2023).
Mauvais-Jarvis, F. Gender differences in glucose homeostasis and diabetes. Physiol. Behav. 187, 20–23 (2018).
Mauvais-Jarvis, F. et al. Sex and gender: modifiers of health, disease, and medicine. Lancet 396, 565–582 (2020).
Tramunt, B. et al. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia 63, 453–461 (2020).
Vague, J. Sexual differentiation; Factor determining forms of obesity]. Presse Med (1893) 55, 339 (1947).
Goossens, G. H., Jocken, J. W. E. & Blaak, E. E. Sexual dimorphism in cardiometabolic health: the role of adipose tissue, muscle and liver. Nat. Rev. Endocrinol. 17, 47–66 (2021).
Gourdy, P. et al. Estrogen receptor subcellular localization and cardiometabolism. Mol. Metab. 15, 56–69 (2018).
Louet, J. F., LeMay, C. & Mauvais-Jarvis, F. Antidiabetic actions of estrogen: insight from human and genetic mouse models. Curr. Atheroscler. Rep. 6, 180–185 (2004).
Cypess, A. M. Reassessing Human Adipose Tissue. N. Engl. J. Med 386, 768–779 (2022).
Sakers, A., De Siqueira, M. K., Seale, P. & Villanueva, C. J. Adipose-tissue plasticity in health and disease. Cell 185, 419–446 (2022).
Cypess, A. M. et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med 360, 1509–1517 (2009).
Saito, M. et al. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes 58, 1526-1531, db09-0530 [pii] (2009).
van Marken Lichtenbelt, W. D. et al. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med 360, 1500–1508 (2009).
Virtanen, K. A. et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med 360, 1518–1525 (2009). 360/15/1518 [pii].
Becher, T. et al. Brown adipose tissue is associated with cardiometabolic health. Nat. Med 27, 58–65 (2021).
Shimizu, I. et al. Vascular rarefaction mediates whitening of brown fat in obesity. J. Clin. Invest 124, 2099–2112 (2014).
Shimizu, I. & Walsh, K. The whitening of brown fat and its implications for weight management in obesity. Curr. Obes. Rep. 4, 224–229 (2015).
Petrovic, N. et al. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem. 285, 7153–7164 (2010).
Wu, J. et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 150, 366–376 (2012).
Cypess, A. M. et al. Anatomical localization, gene expression profiling and functional characterization of adult human neck brown fat. Nat. Med 19, 635–639 (2013).
Jespersen, N. Z. et al. A classical brown adipose tissue mRNA signature partly overlaps with brite in the supraclavicular region of adult humans. Cell Metab. 17, 798–805 (2013).
Kiefer, F. W. The significance of beige and brown fat in humans. Endocr. Connect 6, R70–R79 (2017).
Lidell, M. E. et al. Evidence for two types of brown adipose tissue in humans. Nat. Med 19, 631–634 (2013).
Nedergaard, J. & Cannon, B. How brown is brown fat? It depends where you look. Nat. Med 19, 540–541 (2013).
Chen, K. Y. et al. Opportunities and challenges in the therapeutic activation of human energy expenditure and thermogenesis to manage obesity. J. Biol. Chem. 295, 1926–1942 (2020).
Giordano, A., Frontini, A. & Cinti, S. Convertible visceral fat as a therapeutic target to curb obesity. Nat. Rev. Drug Discov. 15, 405–424 (2016).
Young, P., Arch, J. R. & Ashwell, M. Brown adipose tissue in the parametrial fat pad of the mouse. FEBS Lett. 167, 10–14 (1984).
Cassard-Doulcier, A. M. et al. Tissue-specific and beta-adrenergic regulation of the mitochondrial uncoupling protein gene: control by cis-acting elements in the 5’-flanking region. Mol. Endocrinol. 7, 497–506 (1993).
Norheim, F. et al. Gene-by-Sex Interactions in Mitochondrial Functions and Cardio-Metabolic Traits. Cell Metab. 29, 932–949.e934 (2019).
Wang, H. et al. A dual Ucp1 reporter mouse model for imaging and quantitation of brown and brite fat recruitment. Mol. Metab. 20, 14–27 (2019).
Barquissau, V. et al. Caloric restriction and diet-induced weight loss do not induce browning of human subcutaneous white adipose tissue in women and men with obesity. Cell Rep. 22, 1079–1089 (2018).
Nookaew, I. et al. Adipose tissue resting energy expenditure and expression of genes involved in mitochondrial function are higher in women than in men. J. Clin. Endocrinol. Metab. 98, E370–E378 (2013).
Consortium, G. T. The genotype-tissue expression (GTEx) project. Nat. Genet 45, 580–585 (2013).
O’Malley, B. W. Origins of the field of molecular endocrinology: a personal perspective. Mol. Endocrinol. 30, 1015–1018 (2016).
Spiegelman, B. M. & Heinrich, R. Biological control through regulated transcriptional coactivators. Cell 119, 157–167 (2004).
Stallcup, M. R. & Poulard, C. Gene-specific actions of transcriptional coregulators facilitate physiological plasticity: evidence for a physiological coregulator code. Trends Biochem Sci. 45, 497–510 (2020).
Ohno, H., Shinoda, K., Spiegelman, B. M. & Kajimura, S. PPARgamma agonists induce a white-to-brown fat conversion through stabilization of PRDM16 protein. Cell Metab. 15, 395–404 (2012).
Seale, P. et al. Transcriptional control of brown fat determination by PRDM16. Cell Metab. 6, 38–54 (2007).
Leonardsson, G. et al. Nuclear receptor corepressor RIP140 regulates fat accumulation. Proc. Natl Acad. Sci. USA 101, 8437–8442 (2004).
Puigserver, P. et al. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 92, 829–839 (1998).
Enguix, N. et al. Mice lacking PGC-1beta in adipose tissues reveal a dissociation between mitochondrial dysfunction and insulin resistance. Mol. Metab. 2, 215–226 (2013).
Kressler, D., Schreiber, S. N., Knutti, D. & Kralli, A. The PGC-1-related protein PERC is a selective coactivator of estrogen receptor alpha. J. Biol. Chem. 277, 13918–13925 (2002).
Louet, J. F. et al. Oncogenic steroid receptor coactivator-3 is a key regulator of the white adipogenic program. Proc. Natl Acad. Sci. USA 103, 17868–17873 (2006).
Louet, J. F. & O’Malley, B. W. Coregulators in adipogenesis: what could we learn from the SRC (p160) coactivator family?. Cell Cycle 6, 2448–2452 (2007).
Picard, F. et al. SRC-1 and TIF2 control energy balance between white and brown adipose tissues. Cell 111, 931–941 (2002).
Din, Mu. et al. Postprandial oxidative metabolism of human brown fat indicates thermogenesis. Cell Metab. 28, 207–216.e203 (2018).
Massier, L. et al. An integrated single cell and spatial transcriptomic map of human white adipose tissue. Nat. Commun. 14, 1438 (2023).
Aupetit, A. et al. Endothelial DLL4 is an adipose depot-specific fasting sensor regulating fatty acid fluxes. Arterioscler Thromb. Vasc. Biol. 43, 684–696 (2023).
Larsen, T. M. et al. Diets with high or low protein content and glycemic index for weight-loss maintenance. N. Engl. J. Med 363, 2102–2113 (2010).
Armenise, C. et al. Transcriptome profiling from adipose tissue during a low-calorie diet reveals predictors of weight and glycemic outcomes in obese, nondiabetic subjects. Am. J. Clin. Nutr. 106, 736–746 (2017).
Rodriguez, A. M. et al. Transplantation of a multipotent cell population from human adipose tissue induces dystrophin expression in the immunocompetent mdx mouse. J. Exp. Med 201, 1397–1405 (2005).
Giroud, M. et al. miR-125b affects mitochondrial biogenesis and impairs brite adipocyte formation and function. Mol. Metab. 5, 615–625 (2016).
Zaragosi, L. E. et al. Activin a plays a critical role in proliferation and differentiation of human adipose progenitors. Diabetes 59, 2513–2521 (2010).
Geng, L., Lam, K. S. L. & Xu, A. The therapeutic potential of FGF21 in metabolic diseases: from bench to clinic. Nat. Rev. Endocrinol. 16, 654–667 (2020).
Son, M. J. et al. GATA3 induces the upregulation of UCP-1 by directly binding to PGC-1alpha during adipose tissue browning. Metabolism 109, 154280 (2020).
Hevener, A. L., Clegg, D. J. & Mauvais-Jarvis, F. Impaired estrogen receptor action in the pathogenesis of the metabolic syndrome. Mol. Cell Endocrinol. 418, 306–321 (2015).
Mauvais-Jarvis, F. Elucidating sex and gender differences in diabetes: a necessary step toward personalized medicine. J. Diabetes Complications 29, 162–163 (2015).
Clayton, J. A. & Collins, F. S. Policy: NIH to balance sex in cell and animal studies. Nature 509, 282–283 (2014).
Kozak, L. P. Brown fat and the myth of diet-induced thermogenesis. Cell Metab. 11, 263–267 (2010).
Shabalina, I. G. et al. UCP1 in brite/beige adipose tissue mitochondria is functionally thermogenic. Cell Rep. 5, 1196–1203 (2013).
Jastroch, M. & Seebacher, F. Importance of adipocyte browning in the evolution of endothermy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375, 20190134 (2020).
Lehnig, A. C. & Stanford, K. I. Exercise-induced adaptations to white and brown adipose tissue. J Exp Biol 221, https://doi.org/10.1242/jeb.161570 (2018).
Stanford, K. I., Middelbeek, R. J. & Goodyear, L. J. Exercise Effects on White Adipose Tissue: Beiging and Metabolic Adaptations. Diabetes 64, 2361–2368 (2015).
Montero, D., Madsen, K., Meinild-Lundby, A. K., Edin, F. & Lundby, C. Sexual dimorphism of substrate utilization: Differences in skeletal muscle mitochondrial volume density and function. Exp. Physiol. 103, 851–859 (2018).
Lightfoot, J. T. Sex hormones’ regulation of rodent physical activity: a review. Int J. Biol. Sci. 4, 126–132 (2008).
Onate, S. A., Tsai, S. Y., Tsai, M. J. & O’Malley, B. W. Sequence and characterization of a coactivator for the steroid hormone receptor superfamily. Science 270, 1354–1357 (1995).
Puigserver, P. et al. Activation of PPARgamma coactivator-1 through transcription factor docking. Science 286, 1368–1371 (1999).
Rocchi, S. et al. A unique PPARgamma ligand with potent insulin-sensitizing yet weak adipogenic activity. Mol. Cell 8, 737–747 (2001).
Jeffery, E. et al. The adipose tissue microenvironment regulates depot-specific adipogenesis in obesity. Cell Metab. 24, 142–150 (2016).
Bloor, I. D. & Symonds, M. E. Sexual dimorphism in white and brown adipose tissue with obesity and inflammation. Horm. Behav. 66, 95–103 (2014).
Lee, M. J. & Fried, S. K. Sex-dependent depot differences in adipose tissue development and function; role of sex steroids. J. Obes. Metab. Syndr. 26, 172–180 (2017).
Trujillo-Ochoa, J. L., Kazemian, M. & Afzali, B. The role of transcription factors in shaping regulatory T cell identity. Nat. Rev. Immunol. 23, 842–856 (2023).
Tong, Q. et al. Function of GATA transcription factors in preadipocyte-adipocyte transition. Science 290, 134–138 (2000).
Ahluwalia, A., Hoa, N., Ge, L., Blumberg, B. & Levin, E. R. Mechanisms by Which Membrane and Nuclear ER Alpha Inhibit Adipogenesis in Cells Isolated From Female Mice. Endocrinology 161, https://doi.org/10.1210/endocr/bqaa175 (2020).
Schulz, T. J. et al. Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 495, 379–383 (2013).
Keipert, S. et al. Genetic disruption of uncoupling protein 1 in mice renders brown adipose tissue a significant source of FGF21 secretion. Mol. Metab. 4, 537–542 (2015).
Keipert, S. et al. Endogenous FGF21-signaling controls paradoxical obesity resistance of UCP1-deficient mice. Nat. Commun. 11, 624 (2020).
Chukijrungroat, N., Khamphaya, T., Weerachayaphorn, J., Songserm, T. & Saengsirisuwan, V. Hepatic FGF21 mediates sex differences in high-fat high-fructose diet-induced fatty liver. Am. J. Physiol. Endocrinol. Metab. 313, E203–E212 (2017).
Curran, J. E. et al. Genetic determinants of mitochondrial content. Hum. Mol. Genet 16, 1504–1514 (2007).
Wu, Z. et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 98, 115–124 (1999).
Hinney, A., Korner, A. & Fischer-Posovszky, P. The promise of new anti-obesity therapies arising from knowledge of genetic obesity traits. Nat. Rev. Endocrinol. 18, 623–637 (2022).
Basse, A. L. et al. Global gene expression profiling of brown to white adipose tissue transformation in sheep reveals novel transcriptional components linked to adipose remodeling. BMC Genomics 16, 215 (2015).
Yang, Y. et al. Steroid receptor coactivator-1 modulates the function of Pomc neurons and energy homeostasis. Nat. Commun. 10, 1718 (2019).
Cacciottolo, T. M. et al. Obesity due to steroid receptor coactivator-1 deficiency is associated with endocrine and metabolic abnormalities. J. Clin. Endocrinol. Metab. 107, e2532–e2544 (2022).
Xu, J. et al. Partial hormone resistance in mice with disruption of the steroid receptor coactivator-1 (SRC-1) gene. Science 279, 1922–1925 (1998).
Ross, S. A., Song, X., Burney, M. W., Kasai, Y. & Orlicky, D. J. Efficient adenovirus transduction of 3T3-L1 adipocytes stably expressing coxsackie-adenovirus receptor. Biochem Biophys. Res Commun. 302, 354–358 (2003).
Louet, J. F. et al. The coactivator SRC-1 is an essential coordinator of hepatic glucose production. Cell Metab. 12, 606–618 (2010).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Buenrostro, J. D., Giresi, P. G., Zaba, L. C., Chang, H. Y. & Greenleaf, W. J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 10, 1213–1218 (2013).
Corces, M. R. et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nat. Methods 14, 959–962 (2017).
Castellano-Castillo, D. et al. Chromatin immunoprecipitation improvements for the processing of small frozen pieces of adipose tissue. PLoS One 13, e0192314 (2018).
Kong, X. et al. IRF4 is a key thermogenic transcriptional partner of PGC-1alpha. Cell 158, 69–83 (2014).
Acknowledgements
The authors thank Drs. Jianming Xu and Bert W. O’Malley for providing the Ncoa1 KO mouse strains, as well as the animal facilities of Baylor College of Medicine and Inserm U1065 for housing the mice used in this study. The mouse Ucp1 promoter constructs were kindly provided by Dr Evan Rosen93 and the Ppargc1a and Ppargc1b constructs were generously shared by Dr Johan Auwerx. The authors also extend their gratitude to Pr. Grolleau-Raoux and the Department of Plastic Surgery (Hôpital Universitaire Toulouse Rangueil, Toulouse, France) for collecting subcutaneous gluteofemoral adipose tissue from women participating in the ADIPENDO interventional cohort study. The isolation of mature human adipocytes was conducted under the PHRC interregional 2012 ADIPENDO N° 1206708, CHU de Toulouse (Toulouse, France). This work was carried out as part of the Adipo-Cible Research Study Group, supported by the French government through the France 2030 Investment Plan managed by the National Research Agency (ANR), as part of the Initiative of Excellence of Université Côte d’Azur under reference number ANR-15-IDEX-01. The study was supported by the Institut National de la Santé et de la Recherche Médicale (Inserm) and Côte d’Azur University; by grants from the Société Francophone du Diabète (SFD) to J-F.L.; by the French National Research Agency (ANR) through the investments for the future Labex SIGNALIFE (ANR-11-LABX-0028-01) to J-F.T.; J-F.L. and J-F.T. are researchers at the Centre National de la Recherche Scientifique (CNRS). M.T-L. is an Inserm researcher and was funded by the Inserm, CNRS, Institut National du Cancer (PLBIO INCa 2018-1-PLBIO-05) and the Association de Recherche contre le Cancer (ARC PJA 20171206570).
Author information
Authors and Affiliations
Contributions
Conceptualization: J.-F.L. and M.T.L.; Methodology: J.-F.L., M.T.L., D.P., E.M., A.B., D.L., N.V., P.G., E-Z.A.; Experimentation: J.-F.L., M.T.L., D.P., H.B., A.Z.; M.B., N.L., B.C., E-Z.A., N.G., B.C.; Data analysis: J.-F.L., M.T.L., D.P., E.M.; J.-F. T., M.C., P.G., E-Z.A.; Writing original draft: J.-F.L. and M.T.L.; Writing review & editing: J.-F.L., M.T.L., D.P., E.M., N.V.; J.-F.T., M.C., P.G., E-Z.A.; D.L. Funding acquisition: J.-F. L., M.T.L., J-F.T.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Tannour-Louet, M., Pisani, D.F., Bouguerra, H. et al. NCOA1 is a gatekeeper of the sexually dimorphic thermogenic activity of white adipose tissue. Nat Commun 17, 717 (2026). https://doi.org/10.1038/s41467-025-65229-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-65229-6
