
Abstract
How extracellular vesicles (EVs) may contribute to mechanisms of primary intracellular pathogenesis in Parkinson’s disease (PD) remains unknown. To critically advance our understanding of how EVs influence early-stage PD pathogenesis, we tested the hypothesis that rats acutely exposed to the PD neurotoxin rotenone would produce differential miRNAs in CSF/serum-derived EVs and that such modulation would be responsible for PD-relevant functional alterations in recipient neuronal cells. We discovered that acute rotenone treatment produced significant and specific serum miRNA alterations. Primary midbrain neurons treated with serum EVs from rotenone-exposed rats produced oxidative stress, mitochondrial toxicity, and cell loss in neuronal culture. These mechanisms were dependent on miR-30a-5p and miR-484. Thus, this study has elucidated that differential expression of miRNAs in circulating EVs from serum/CSF of rats is a potential early diagnostic marker for PD, and that the modulation of cellular functions and viability due to extracellular vesicles determines the pathological fate.

Similar content being viewed by others
Introduction
Parkinson’s disease (PD) is a debilitating neurodegenerative disorder that affects >6.1 million people worldwide, with incidence and prevalence rapidly rising in the past twenty years1. The hallmark motor features of PD include gait disturbances, tremors, bradykinesia, and muscle instability. The non-motor symptoms include sleep disorders, depression, hyposmia or constipation, may occur on an average a decade before the motor features, hence it is difficult to identify the disease at an early stage2. Hallmark PD pathology is the loss of dopaminergic neurons in the substantia nigra (SN) pars compacta region of the ventral midbrain, which in large part produces the motor phenotype3. At diagnosis, ~50–70% of dopaminergic neurons are lost in the nigra, and abnormal aggregation of α-synuclein is observed in the remaining neurons, which constitute the Lewy bodies4. The primary cellular mechanisms of PD have been extensively studied with much focus on neuroinflammation, mitochondrial dysfunction, oxidative stress, loss of proteostasis, and activation of apoptotic pathways, which ultimately lead to the death of the dopaminergic neurons5. The dysfunction of these cellular and molecular pathways is typically an outcome of interactions between ageing, genetic susceptibility, and environmental factors.
While intracellular mechanisms have been extensively studied, there are considerable gaps in our understanding of pathogenic spread across brain regions and systemically. Extracellular vesicles (EVs) are important for interneuronal communication and, importantly, can cross physiological barriers, including the blood–brain barrier (BBB) and the blood–CSF barrier (BCSFB), and recent reports have highlighted their role as potential circulating biomarkers for various pathologies, including cancer, heart diseases, diabetes, and neurodegenerative disorders, including PD6. EVs is a general term that refers to a wide group of “particles” that are released from cells, contained in lipid bilayers, and can be categorized into exosomes (30–150 nm), microvesicles (100–1000 nm), and apoptotic bodies (1000–5000 nm)7. All EVs do contain vital biological materials, like DNA, RNAs, like mRNAs, miRNAs, tRNAs and rRNAs, enzymes, lipids, etc.8,9. Exosomes, which is a class of EVs, are nano-sized vesicles, originating from the multivesicular bodies (MVBs) where ILVs (intra-luminal vesicles) are formed and recruit different cargo which are specifically packaged through the endolysosomal system10. The function of these vesicles is cell-cell communication, disposal of cellular waste, and, in case, of pathologies, transfer of pathogenic molecules11. The EV cargo varies according to different cell types, physiological conditions, and in response to pathological stimuli, which makes it an attractive target for novel prognosis/diagnosis, and drug carrier for other therapeutic possibilities. The levels of CSF-derived α-synuclein in plasma exosomes was found to be substantially higher in PD patients and this correlated with the severity of the disease12. Since in our current study we refer to a wide family of “particles”, and do not delve much into the subcellular origin of biogenesis of these vesicles, we will broadly refer to the term “extracellular vesicles” or EVs.
Small non-coding RNAs, specifically miRNAs, are stable in the body fluids and are important posttranscriptional regulators of mRNA for regulating the protein level during specific pathophysiological processes13. Several miRNAs are already reported to be differentially regulated in the plasma of PD patients as compared to healthy controls14. Several reports suggest that miRNAs are packaged inside the EVs and are internalized in specific cell types as important mediators of cellular outcomes. The circulating EV-miRNAs may also have a key role in the development and spread of PD pathogenesis. However, these pathways have not yet been well elucidated. The origin of EV-miRNA in the CSF remains unclear, with ongoing questions about whether they originate from neurons or other cell types in the brain, or if they are transferred from peripheral blood to the CSF crossing the blood–CSF barrier (BCSFB). Further, the role of these EV-miRNAs in determining the pathological fate by spreading the pathogenesis is not well understood. Circulating exosomal miRNAs in plasma as well as CSF have been identified in PD patients, which could be potentially used as biomarkers of disease progression15,16, however their release from specific cell types and abundance in the body fluids like serum and CSF needs to be further investigated.
The aim of this study was to address critical gaps in the existing literature by discovering whether rats acutely exposed to the Parkinson’s disease neurotoxin rotenone would exhibit distinct EV-miRNAs in CSF and serum, prior to downstream neuropathology. We hypothesized that the modulation of these EVs would play a role in functional changes observed in the recipient neuronal cells. While there are a number of models of PD, including genetic, neurotoxin/neurotoxicant, gene–environment interaction, and others, the rotenone model was chosen for environmental relevance and utility in modeling the earliest stages of the disease. Rotenone is a natural organic compound that is isolated from the roots of several plant species, and is widely used as a pesticide, an insecticide, and a piscicide and was identified as a potent mitochondrial complex I inhibitor of the electron transport chain17,18. Rotenone is selectively toxic to dopaminergic neurons at specific doses, both in vitro19 and in vivo18,20. A highly reproducible model of rotenone was also developed by daily intraperitoneal injections of rotenone to different age groups of male rats, which produced typical parkinsonism behavioral traits, including dopamine-dependent motor deficits and evidence of postural instability21. Moreover, equivalent loss of dopamine in the striatum was reported, along with the death of TH-positive neurons and formation of α-synuclein aggregates21. Finally, rotenone has been identified as a bona fide PD risk factor in humans22. The current study aimed at discovering the role of EV-associated miRNAs in PD pathogenesis through miRNA profiling of PD-rat models, functional assessments, and validation studies, ultimately exploring mechanisms of mitochondrial dysfunction and neuronal death in PD (Supplementary Fig. 1). Our findings demonstrated significant alterations in patterns of EV-miRNAs in the CSF and serum of rotenone-exposed rats. Further, incubation of the serum exosomes with primary neurons from rat embryos induced mitochondrial dysfunction and dopaminergic neuronal cell death. Our results have considerably advanced understanding of the primary role that EV-mediated intercellular communication has in PD pathogenesis.
Results
Rotenone toxicity in primary midbrain and cortical neuronal cultures alters exosome release
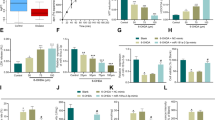
Extracellular vesicle release varies in different pathophysiological conditions, including different acute and chronic neuronal insults. However, this has not been systemically investigated. We first analyzed if rotenone toxicity in primary neuronal cultures from rat embryos influences the EV release. Primary midbrain and cortical cultures were first treated with different rotenone doses for 24 h (acute exposure) to determine the dose-dependent response and the lethal dose. Cell death assay by MTT indicated that significant cell death was observed at 25 nM (p = 0.0069, one-way ANOVA, n = 3) in primary midbrain cultures, whereas the primary cortical cultures required 60 nM (p ≤ 0.01, one-way ANOVA, n = 3)(Fig. 1(i)(a), (ii)(a)). The lethal dose was also confirmed by the DNA damage ELISA of different rotenone concentrations, which showed similar results (Fig. 1(i)(b), (ii)(b)).

(i) Primary midbrain neurons and (ii) primary cortical neurons were treated with different doses of rotenone as mentioned above for 24 h, and cell viability was assessed by a MTT cell viability assay at 570 nm and b cell death detection by ELISA at 405 nm. Asterisk (*), (**), and (***) indicates values statistically significant from control; p value <0.05, <0.01, and <0.001 (respectively), SEM of three independent experiments, data were analyzed using one-way ANOVA, followed by Dunnett’s multiple comparisons test.
Hence, for further experiments, we selected the dose of 25 nM rotenone for midbrain neurons and 60 nM for cortical neurons. Primary neurons were treated with rotenone, and the concentration of exosomes released was assessed by NTA. We observed that both primary midbrain cultures as well as primary cortical cultures treated with rotenone showed enhanced EV release and concentration increases (p ≤ 0.05, Student’s t-test, n = 3) (Fig. 2).

(i) Primary midbrain neuronal cultures and (ii) primary cortical neuronal cultures were treated with 25 and 60 nM of rotenone, respectively, for 24 h and the media was collected posttreatment and subsequently EVs were isolated. The concentration of the samples were determined using nanoparticle tracking analysis (NTA), which depicts the concentration of particles per ml of solution. Asterisk (*) indicates value statistically significant from control; p value <0.05, SEM of three independent experiments, data was analyzed using Student’s t-test. (iii) Percentage distribution of the population of extracellular vesicles (EVs) derived from (iii) primary midbrain cultures (untreated vs rotenone-treated) and (iv) primary cortical cultures (untreated vs rotenone-treated).
EVs are known to be heterogeneous in nature, and different subpopulations are responsible for different biological functions. Therefore, we plotted the subpopulation of EVs based on the size difference into four kinds of groups: exomeres (~35 nm), Exo-S (60–80 nm), Exo-L (90–150 nm), large EVs (150–500 nm), and microvesicles (500–1500 nm). The exomeres, Exo-S and Exo-L particles are what can be categorized as exosomes, based on their size. An exomere population in the primary neuronal culture in untreated as well as in rotenone conditions was not detected. The major subpopulation in primary midbrain cultures was microvesicles, while large EVs were the dominant subpopulation in primary cortical cultures (Fig. 2). The populations Exo-L and large EVs was lower in rotenone conditions as compared to untreated in both of primary neuronal cultures; however, the microvesicle subpopulation increased (Fig. 2).
EV release is altered in acute rotenone-treated rat models
To advance our understanding of the role of signaling through extracellular vesicles in vivo, we tested whether acute rotenone toxicity induced release of EVs observed in vitro is recapitulated in vivo. For this, we generated an acute rotenone model of rats, treated with 3 mg/kg/day for 8 and 24 h. The body weights of the rats declined markedly or significantly after treatment with rotenone for 24 h (p ≤ 0.001, two-way ANOVA, n = 10), however no difference in body weight was observed after 8 h of rotenone treatment (Supplementary Fig. 2), consistent with prior reports21. Next, we quantified the EVs in the serum and cerebrospinal fluid of the rats treated with rotenone at both time points. Transmission electron microscopy (TEM) was performed on CSF- and serum-derived EVs, revealing the characteristic cup-like morphology of vesicles and confirming the purity of the EV samples (Supplementary Fig. 3). nanoparticle tracking analysis (NTA) further validated the size distribution of the EV population. For CSF-derived EVs, the mode size was ~183 nm, with an average size of 213 nm (Supplementary Fig. 4 (i), (iii)). Serum-derived EVs exhibited a mode size of 167 nm, with an average size of 295 nm (Supplementary Fig. 4 (ii), (iii)). Together, TEM and NTA provided robust characterization of the EVs, confirming their purity and size range, consistent with exosomal profiles. NTA analysis indicated an increase in the concentration of EVs obtained from CSF of rotenone-treated rats; however, it was not significant (Fig. 3(i)). The concentration of serum EVs from rotenone-treated rats was significantly higher as compared to that of untreated rats (p ≤ 0.05, Student’s t-test, n = 10) (Fig. 3(ii)).

Rotenone was administered at an acute concentration of 3 mg/kg to Sprague-Dawley male rats of 3 months of age for 8 and 24 h, and subsequently, EVs were isolated from the (i) CSF and (ii) serum of healthy controls as well as rotenone-treated rats post-euthanasia. The concentration of the samples were determined using nanoparticle tracking analysis (NTA), which depicts the concentration of particles per ml of solution. Asterisk (*) and (**) indicates values statistically significant from control; p value <0.05 and <0.01 (respectively), SEM of ten independent values, data were analyzed using Student’s t-test. (iii) Waffle chart depicting percentage distribution of the population of extracellular vesicles (EVs) derived from CSF and (iv) serum of acute-rotenone-exposed rats.
Further, the subpopulation analysis of the CSF EVs showed that the dominant subpopulation is large EVs. Exo-L subpopulation is higher in rotenone-treated rats; however, the concentration of microvesicles decreased in the rotenone-treated rats as compared to untreated (Fig. 3(iii)(b), (iv)(b)). Like CSF, large EVs is the dominant subpopulation in serum EVs as well. Surprisingly, all three subpopulations, exomeres, Exo-S, and Exo-L, increased in the serum of rotenone-treated rats as compared to untreated (Fig. 3(iv)). Large EVs are significantly lower in rotenone-treated rats as compared to untreated, while there is no difference in the microvesicle population.
Specific miRNA levels are altered in both primary cultures and EVs in PD stress conditions
We further analyzed specific EV-miRNA levels to acute rotenone treatment in primary cortical and primary midbrain cultures. The previous study from our group had identified specific miRNAs which are altered under PD stress conditions in neuronal cell lines through NGS23. Some of these specific miRNAs were validated in rotenone conditions in SK-N-SH and SH-SY5Y cells, which served as a rationale for the selection of the mentioned specific miRNAs for validation in the primary cultures and in vivo system in this study. We observed that primary neuronal cells treated with rotenone showed altered levels of miRNAs. The levels of miR-181c-5p (p = 0.016, one-way ANOVA, n = 3–4) and miR-484 (p ≤ 0.0001, one-way ANOVA, n = 3-4) were specifically significantly downregulated in the primary midbrain cultures upon rotenone treatment (Fig. 4(i)(a)). The regulation pattern in cortical cultures was broader, where several miRNAs are downregulated in the primary cortical cultures on rotenone exposure, including miR-30a-5p (p = 0.025, one-way ANOVA, n = 3–4), miR-181c-5p (p = 0.036, one-way ANOVA, n = 3–4), miR-484 (p = 0.03, one-way ANOVA, n = 3–4) and miR-93-5p (p = 0.03, one-way ANOVA, n = 3–4) (Fig. 4(ii)(a)). The levels of miR-30a-5p (p = 0.008, one-way ANOVA, n = 3–4), miR-320a-3p (p = 0.00077, one-way ANOVA, n = 3–4), miR-484 (p ≤ 0.0001, one-way ANOVA, n = 3–4), and miR-93-5p (p = 0.00145, one-way ANOVA, n = 3–4) decreased in EVs derived from primary midbrain cultures treated with rotenone (Fig. 4(i)(b)). However, the level of miR-484 increased in EVs from primary cortical cultures (p ≤ 0.0001, one-way ANOVA, n = 3–4). The level of miR-30a-5p (p = 0.00011, one-way ANOVA, n = 3–4) and miR-93-5p (p = 0.0014, one-way ANOVA, n = 3–4) decreased in EVs from cortical cultures due to rotenone treatment (Fig. 4(ii)(b)). Tables depicting the mean relative fold change in rotenone conditions have been shown along with the standard deviation and standard error of mean (Supplementary Table 1). The above results clearly indicated that specific miRNAs are significantly altered in EVs and cells in response to acute rotenone toxicity.

(i) Primary midbrain cultures and (ii) Primary cortical cultures were treated with 25 and 60 nM rotenone, respectively, for 24 h and subsequently EVs were isolated and differential expression of miRNAs were checked in a total cells and b EVs by RT-qPCR. Asterisk (*), (**), (***), and (****) indicates values statistically significant from control; p value <0.05, <0.01, <0.001, and <0.0001 (respectively), SEM of three or four independent experiments, data were analyzed using one-way ANOVA, followed by Dunnett’s multiple comparisons test.
EVs from serum and cerebrospinal fluid of acute rotenone-treated rats have differential expressions of miRNAs
Further, we analyzed the EV-miRNAs that were validated in the primary neuronal cultures, in the serum and CSF EVs of acute rotenone-treated rats by qRT-PCR. The level of miR-181c-5p (p = 0.047, one-way ANOVA, n = 10–11) and miR-30a-5p (p = 0.039, one-way ANOVA, n = 10–11) increased in EVs derived from CSF at 8 h rotenone treatment, which later decreased significantly after 24 h rotenone treatment (p ≤ 0.001, one-way ANOVA, n = 10–11). The levels of miR-93-5p are lower at both time points as compared to untreated rats (p = 0.000063, p = 0.0069, one-way ANOVA, n = 10–11) (Fig. 5(i)). Levels of both miR-30a-5p (p = 0.0297, p = 0.0062, one-way ANOVA, n = 10–11) and miR-484 (p = 0.007, p = 0.0004, one-way ANOVA, n = 10–11) were downregulated at both time points of rotenone in serum EVs as compared to the EVs from serum of untreated rats. However, the levels of miR-181c-5p (p = 0.0024, one-way ANOVA, n = 10–11) are low at 8 h rotenone treatment, which increased at 24 h rotenone treatment. However, the levels of miR-93-5p was high at 8 h (p = 0.028, one-way ANOVA, n = 10–11) and decreased at 24 h of rotenone treatment (p = 0.012, one-way ANOVA, n = 10–11) (Fig. 5(ii)). The above results indicated that the EV-miRNA levels are differentially sorted at early treatment time points of rotenone and is altered between 8 and 24 h time points of treatment. Specific miRNAs showed consistent changes; level of miR-93-5p decreased in the EVs from CSF and miR-30a-5p and miR-484 was decreased in EVs from serum of rotenone-treated rats.

Acute rotenone toxicity was administered to young adult male Sprague-Dawley rats of 3 months of age at a concentration of 3 mg/kg for 8 and 24 h. Biofluids were subsequently collected from the rats post-euthanasia and EVs were isolated from the biofluids. The differential expression of EV-miRNAs in the (i) CSF and (ii) serum were determined by RT-qPCR. Asterisk (*), (**), (***), and (****) indicates values statistically significant from control; p value <0.05, <0.01, <0.001, and <0.0001 (respectively), SEM of ten independent experiments, data were analyzed using one-way ANOVA, followed by Dunnett’s multiple comparisons test.
As the population of miRNAs in CSF EVs is mainly influenced by the release of cellular miRNAs from different brain cell populations, we extensively analyzed miRNA expression at the cellular level. This was carried out in various rat brain regions subjected to acute rotenone treatment, using qRT-PCR. The miRNAs exhibited distinctive enrichment patterns across different brain regions and at various time points during treatment. Notably, the levels of miR-93-5p were significantly and exclusively elevated in the ventral midbrain region (a pivotal area of interest in PD due to its inclusion of the SNpc) (p ≤ 0.05, one-way ANOVA, n = 10–11), while comparatively lower levels were observed in other brain regions such as the cortex, dorsal midbrain, striatum, and hippocampus (Supplementary Fig. 5).
Conversely, miR-484, initially downregulated in the ventral midbrain at 8 h (p ≤ 0.05, one-way ANOVA, n = 10–11), exhibited an increase in expression at 24 h (p ≤ 0.05, one-way ANOVA, n = 10–11). This temporal shift in miR-484 levels aligns with its CSF EVs’ counterpart, where elevated levels were observed at 8 h followed by a significant decline at 24 h (Supplementary Fig. 5 and Fig. 5(i)). Brain regions like the cortex and hippocampus were designated as control regions, as they demonstrated no significant differences in miRNA levels under acute PD conditions, suggesting a relatively unaffected status in response to the treatment.
Serum EVs from acute rotenone-treated rats induce mitochondrial dysfunction and cell death in primary neuronal cultures from rat embryos
To study the effect of the EVs on primary cultures, serum EVs isolated from healthy rats (referred to as UN exo) and from rats treated with rotenone for 8 h (referred to as R8 exo) or 24 h (referred to as R24 exo) were incubated with primary midbrain and cortical cultures, and mitochondrial functions and cell viability were assessed. R8 exo significantly increased the total cellular ROS levels (p = 0.038, one-way ANOVA, n = 60) (Fig. 6(i)) as well as mitochondrial-specific ROS levels (p = 0.0275, one-way ANOVA, n = 60) (Fig. 6(ii)) in primary midbrain cultures as compared to the UN exo, while R24 exo only showed a stimulatory effect on mitochondrial ROS levels in these cultures. The total cellular ROS levels remained unaltered by the treatment of R8 and R24 exo to primary cortical cultures (Fig. 6(iii)). However, R8 exo significantly increased the mitochondrial ROS levels in primary cortical cultures (p = 0.0092, one-way ANOVA, n = 60) (Fig. 6(iv)).

Serum EVs derived from rotenone-treated rats were incubated with primary midbrain ((i) and (ii)) and cortical neurons ((iii) and (iv)), and total cellular ROS was determined by DCFDA, and mitochondrial ROS was assessed using MitoSox staining. UN exo: serum EVs derived from healthy rats, R8 exo: serum EVs derived from rats exposed to rotenone for 8 h, R24 exo: serum EVs derived from rats exposed to rotenone for 24 h. Each independent experiment consisted of a 96-well plate of primary culture, with each well treated with serum EVs from a different animal, resulting in an “n” of 12 per experiment. Five independent experiments were conducted, leading to a total n = 60. Asterisk (*) and (**) indicates values statistically significant from control; p value <0.05 and <0.01 (respectively), SEM of five independent experiments, data were analyzed using one-way ANOVA, followed by Dunnett’s multiple comparisons test.
Primary midbrain cultures showed decreased cell viability on incubation with both R8 exo (p = 0.0006, one-way ANOVA, n = 30–36) and R24 exo (p = 0.030, one-way ANOVA, n = 30–36) as measured using the MTT assay (Fig. 7(i)(a)). Similar results were obtained by the cell death detection ELISA, where increased enrichment of mono-and oligonucleosomes is observed in the cytoplasm, which is indicative of apoptotic cells, in the presence of R8 exo (p = 0.0145, one-way ANOVA, n = 30–36) and R24 exo (p = 0.0058, one-way ANOVA, n = 30–36) (Fig. 7(i)(b)). Primary cortical cultures incubated with R8 exo (but not R24 exo) showed cell death by MTT (p = 0.0266, one-way ANOVA, n = 30–36) (Fig. 7(ii)(a)), whereas analysis of these cultures by ELISA did not show any significant enrichment of mono-/oligonucleosomes on incubation with R8 or R24 serum EVs (Fig. 7(ii)(b)).

Serum EVs from rotenone-treated rat models of PD at both 8 and 24 h were incubated with primary neurons at a concentration of 50 μg/mL. Posttreatment, the cell viability was assessed in (i) primary midbrain neurons and (ii) primary cortical neurons by using a MTT cell viability assay and b cell death detection by DNA damage ELISA. (iii) a Serum EVs were treated to primary midbrain neurons as mentioned above, and the cells were fixed posttreatment, and immunocytochemistry was performed in the cells using TH (red) for specific dopaminergic neurons and MAP2 (green) as a general neuronal marker, and the cells were observed using confocal microscopy (merged image is displayed) and b % quantification of TH neurons over MAP2 was calculated by counting the cells using Nikon analysis software. The scale bar represents 100 µm. A minimum of 10 fields per well were considered, analyzing over 200 cells per field. UN exo: serum EVs derived from healthy rats, R8 exo: serum EVs derived from rats exposed to rotenone for 8 h, R24 exo: serum EVs derived from rats exposed to rotenone for 24 h. Each independent experiment consisted of a 48-well plate of primary culture, with each well treated with serum EVs from a different animal, resulting in an “n” of 6–12 per experiment. Three to five independent experiments were conducted, leading to a total n = 30–36. Asterisk (*), (**), and (***) indicates values statistically significant from control; p value <0.05, <0.01, and <0.001 (respectively), SEM of three and five independent experiments, data were analyzed using one-way ANOVA, followed by Dunnett’s multiple comparisons test.
Since significant loss in cell viability was observed in primary midbrain neuronal cultures, we further analyzed if serum EVs exhibited specific toxicity to the midbrain dopaminergic neurons. The ratio of TH neurons over MAP2 is significantly lower on incubation with R8 exo (p = 0.0065, one-way ANOVA, n = 48–60) and R24 exo (p = 0.0154, one-way ANOVA, n = 48–60) (Fig. 7(iii)) suggesting that the dopaminergic neuron population specifically is affected by the treatment of serum EVs.
Serum EVs elicit dopaminergic neuron cell death in primary rat midbrain cultures pretreated with rotenone
These data indicated that the serum EV cargo from rats treated with rotenone for 8 h (R8 exo) induced mitochondrial dysfunction and cell death in primary midbrain cultures and specifically the dopaminergic neurons. Therefore, we checked the effect of these EVs on primary midbrain cultures which were already pretreated with rotenone.
R8 exo was incubated with primary midbrain neuronal cultures that were pretreated with 25 nM of rotenone for 24 h and the cell death was assessed by MTT cell viability assay. A significant loss in cell viability was obtained in cultures treated with rotenone or R8 exo alone (p ≤ 0.05, two-way ANOVA, n = 48). The cell death was exacerbated when the R8 exo and rotenone is co-treated to the primary midbrain cultures (p ≤ 0.0001, two-way ANOVA, n = 48) (Fig. 8(i)). On performing the cell death detection by ELISA method, significant enrichment of mono-and oligonucleosomes was observed in the cells treated with both R8 exo and rotenone as compared to the individual treatments alone (p ≤ 0.0001, two-way ANOVA, n = 24–36) (Fig. 8(ii)).

Serum EVs from rotenone-treated rat models of PD at 8 h were incubated with primary neurons pretreated with 25 nM rotenone at a concentration of 50 µg/mL. Posttreatment, cell viability was assessed using (i) MTT cell viability assay and (ii) cell death detection by DNA damage ELISA. (iii) a Serum EVs were treated to primary midbrain neurons as mentioned above, and the cells were fixed posttreatment and immunocytochemistry was performed in the cells using TH (red) for specific dopaminergic neurons and HuC (green) as a general neuronal marker, and the cells were observed using confocal microscopy (merged image is displayed) and b % quantification of TH neurons over HuC was calculated by counting the cells using Nikon analysis software. The scale bar represents 100 µm. A minimum of 10 fields per well were considered, analyzing over 200 cells per field. UN exo: serum EVs derived from healthy rats, R8 exo: serum EVs derived from rats exposed to rotenone for 8 h. Each independent experiment consisted of a 96-well plate of primary culture, with each well treated with serum EVs from a different animal, resulting in an “n” of 6–8 per experiment. Three independent experiments were conducted, leading to a total n = 24–36. Asterisk (*), (***), and (****) indicates values statistically significant from control; p value <0.05, <0.001, and <0.0001 (respectively), SEM of three independent experiments, data were analyzed using two-way ANOVA, followed by Sidak’s and Tukey’s multiple comparisons test.
We also further checked whether the toxicity was specific to dopaminergic neurons of the midbrain. Dopaminergic neurons were tagged with TH (Tyrosine Hydroxylase), and a general neuronal marker, HuC, was used to tag all neurons, and the ratio of TH-positive neurons over HuC was calculated for the above-mentioned treatment conditions (Fig. 8(iii)(a)). There was a decrease in the ratio of TH neurons over HuC when cells were treated with rotenone or R8 exo alone (p ≤ 0.05, two-way ANOVA, n = 48), and the TH/HuC ratio was further reduced when these were treated together (p = 0.0201, two-way ANOVA, n = 48) (Fig. 8(iii)(b)).
Serum-derived miR-484 and miR-30a-5p alleviate rotenone-induced neurotoxicity in SH-SY5Y neuronal cells
The findings indicate that serum EVs, when exposed to rotenone-treated primary midbrain cultures, intensified cell death. To investigate the role of miR-484 and miR-30a-5p—found to be downregulated in serum EVs—we transfected SH-SY5Y cells with synthetic miRNA mimics. We aimed to elucidate whether restoring these miRNAs can modulate or mitigate rotenone-induced neurotoxicity, potentially uncovering a protective role in dopaminergic neuron survival. To assess the role of miR-484 and miR-30a-5p in mitigating rotenone toxicity, a dose-dependent study determined 50 nM rotenone (p < 0.0001, one-way ANOVA, n = 12) as sufficient to induce significant cell death in SH-SY5Y cells (Supplementary Fig. 6). Cells transfected with miR-484 or miR-30a-5p, alongside control mimics (CM), were treated with 50 nM rotenone. Both miR-484 and miR-30a-5p effectively alleviated rotenone-induced cellular (p < 0.0001, two-way ANOVA, n = 16) (Fig. 9a) and mitochondrial ROS (p < 0.0001, two-way ANOVA, n = 16) (Fig. 9b) in SH-SY5Y cells. We further assessed ROS levels when both miR-484 and miR-30a-5p were co-transfected. While each miRNA individually reduced ROS levels, their combined expression yielded a similar reduction without any additional enhancement in mitigating ROS levels (p = 0.0015, p = 0.0035, two-way ANOVA, n = 16) (Supplementary Fig. 7).

SH-SY5Y cells were transfected with control mimic (CM) and miR-30a-5p or miR-484 mimics and were subsequently treated with 50 nM rotenone. a Total cellular ROS levels were quantified using DCFDA staining, and b mitochondrial ROS levels were assessed by MitoSox staining. The cellular viability was assessed using c MTT assay and d DNA damage ELISA. Asterisk (*), (**), and (****) indicates values statistically significant from control; p value <0.05, <0.01, and <0.0001 (respectively), SEM of eight independent experiments, data were analyzed using two-way ANOVA, followed by Tukey’s multiple comparisons test.
To further assess the protective effects of miR-484 and miR-30a-5p against rotenone-induced toxicity, cell viability was measured. MTT assay demonstrated that cells transfected with the CM showed a significant reduction in viability upon rotenone exposure (p < 0.0001, two-way ANOVA, n = 16). However, miR-484 and miR-30a-5p mimic transfection alleviated cell death (p < 0.0001, two-way ANOVA, n = 16) (Fig. 9c). These findings were corroborated using the cell death detection ELISA, which revealed reduced enrichment of mono- and oligonucleosomes in the cytoplasm of miR-484 (p = 0.0491, two-way ANOVA, n = 16) and miR-30a-5p (p = 0.0067, two-way ANOVA, n = 16) transfected cells compared to CM-transfected rotenone-treated cells, further confirming their protective role (Fig. 9d). A putative bioinformatic analysis of the targets of miR-30a-5p and miR-484 was performed using Starbase, focusing on genes related to cell death and mitochondrial functions. The target genes were visualized using Cytoscape software (Supplementary Fig. 8). The analysis revealed that both miRNAs shared common targets, including BNIP3L and HNF1A, as well as unique targets involved in apoptotic processes and mitochondrial functions (Supplementary Fig. 8). Additionally, all combined targets of the two miRNAs were analyzed using DAVID, which indicated that the most significantly affected biological processes were regulation of cell communication and apoptosis regulation (Supplementary Fig. 9a). The molecular functions of these targets were primarily related to protein binding (Supplementary Fig. 9b), and their cellular localization was predominantly cytosolic, though they were also distributed across various neuronal substructures, including the synapse and axon (Supplementary Fig. 9c).
Discussion
The mechanisms driving PD pathogenesis, including its spread from dopaminergic neurons to other brain regions, remain incompletely understood. Specifically, while extracellular vesicles (EVs) are implicated in various conditions, their involvement in PD pathogenesis and systemic spread has not been systematically explored. Moreover, early diagnostic biomarkers for PD are lacking. Our studies identified new roles of EVs as mediators of intercellular communication in early PD stages. Addressing this gap, we identified critical alterations in EV-miRNAs in an acute systemic PD model, suggesting their role in driving pathogenic spread across brain regions and systemically, advancing understanding of PD progression.
Previously, reports indicated that both genetic or sporadic PD cases exhibit abnormalities in mitochondrial function and pathways24,25. It is well known that PD pathology extends beyond the dopaminergic neurons of SN, and EVs may also contribute to this spread of pathogenesis. However, it is not known if mitochondrial dysfunction in dopaminergic neurons and neurons of other brain regions leads to alterations in vesicular release and changes in EV-miRNA signatures. We utilized rotenone as a model to induce mitochondrial dysfunction, a cardinal feature of PD, and investigated EV release, monitored EV levels in serum and CSF, and explored the neurotoxic potential of released EVs. Previous reports suggest that PD stress (rotenone) can enhance the exosome release in neuroblastoma as well as glial cell lines26. We examined the EV concentration that is released from primary midbrain and cortical cultures, since the midbrain region is particularly susceptible in PD vs cortical neurons, on exposure to acute rotenone toxicity. We found EV concentration to be increased due to rotenone treatment of primary neurons which is concordant with the previous results26. To understand the complexity of the heterogenous nature of these nanovesicles, various studies have attempted to isolate different subpopulations of these vesicles with different biodistributions and functions27,28. One group identified exosome subpopulations based on size by using asymmetric flow field-flow fractionation (AF4) technology29. Through NTA analysis, we identified and categorized the extracellular vesicles (as mentioned in the Results section). While the large EVs and microvesicles comprised most of the population of the vesicles, surprisingly, we did not detect “exomere” population in the primary neurons, which are reported to be enriched in metabolic enzymes, and are involved in specific pathways, like glycolysis and mTOR signaling29. Interestingly, the total concentration of EVs microvesicle population increased significantly from primary neurons. A recent study demonstrated that the microvesicle population of EVs transfers active mitochondria from brain endothelial cells to recipient cells, elevating the ATP levels, indicating increased cell survival30. Our study has demonstrated changes in the mitochondrial functions in the treatment of EVs, which supports the above report hinting at various cellular functions that might be affected in PD conditions.
PD detection currently relies on clinical symptoms, which appear only in later stages. Early detection is crucial, as therapeutic interventions are most effective at this stage. However, no validated diagnostic biomarker exists. Non-invasive or minimally invasive biofluids (e.g., blood, saliva, urine, and CSF) are ideal for identifying clinically relevant biomarkers. The use of CSF was considered ideal for the detection of an experimental biomarker, because of its proximity to the central nervous system (CNS)31. However, α-synuclein levels in blood or CSF can lack specificity32, hence there is a need to develop alternative biomarkers for the detection of PD. The circulating miRNAs in the body fluids then were beginning to be explored as promising biomarkers for PD33,34. We examined the EV-miRNAs in primary neurons on treatment with rotenone. A previous study from our lab conducted a comprehensive analysis of exosomal miRNA alterations under PD stress conditions in SH-SY5Y neuroblastoma cells. From this, we identified and validated a subset of miRNAs significantly altered during PD stress. Notably, only a few of these miRNAs showed relevance in modulating mitochondrial functions, as supported by prior studies and bioinformatics predictions. Bioinformatics analysis revealed that these miRNAs target distinct or overlapping mitochondrial pathways, corroborated by other research. This focused refinement rationalized the use of miRNAs in our current study, emphasizing miRNAs not only altered in PD cellular models but also implicated in mitochondrial regulation. Due to this reason, we chose miRNAs identified in the previous study for validation in the rotenone model of PD in this study. We found particularly two miRNAs–hsa-miR-30a-5p and hsa-miR-181c-5p that were significantly downregulated in both primary midbrain and cortical neurons, which recapitulates the results that we previously obtained in the in vitro model of PD in SH-SY5Y neuroblastoma cells23.
Subsequently, we examined the circulating EV-miRNAs in the biofluids of the acute rotenone-rat models of PD to validate our previous findings. To address volume constraints associated with our biofluids (CSF and serum), we selected EV isolation protocols based on Thermo Fisher exosome isolation kits. According to the MISEV guidelines, “Extracellular particles (EPs)” is the umbrella term for multimolecular assemblies in the nanometer to micron size range, outside of the cell, including both EVs and non‐vesicular extracellular particles (NVEPs)7. While “extracellular vesicles” (EVs) specifically refers to membrane-enclosed structures, “extracellular particles” (EPs) is a broader term encompassing both vesicular and non-vesicular components. Although a minor fraction of non-vesicular material may be present in the isolate, the totality of our NTA data, plus TEM analysis confirmed the presence of membrane-bound vesicles, indicating that the majority of the sample consists of EVs. This supports the use of the term “EVs”, suggesting non-vesicular contributions were likely negligible, though it is impossible to fully rule out. To strengthen our characterization of the EVs, we conducted TEM on biofluid-derived EVs to assess their structural integrity, purity, and defined size range. The cup-like bilayer morphology observed through TEM corroborated the EV characterization, while the NTA profiles confirmed that their size range aligns with the typical dimensions expected for EVs. NTA analysis of CSF EVs of rats showed no significant changes, however serum EVs were significantly elevated in rotenone-treated rats. Unlike primary neurons, the CSF and serum EV population showed the presence of exomeres. Exo-S represent canonical exosomes, and Exo-L, on the other hand, are non-canonical exosomes/sEVs of different subcellular origin; both the types of EVs contain miRNAs and other small RNA populations29.
Validation of EV-miRNAs revealed differential regulation at different time points of rotenone treatment. We observed a high variability in the levels of the miRNAs at different time points. This suggests that miRNA changes under different acute conditions of rotenone is highly dynamic, which is in consonance with earlier studies35,36. Exosomal miRNAs are dynamically regulated in response to acute exercise versus sustained exercise35, as well as in response to different antitumor therapies (chemotherapy versus surgery) for lung cancer36. Previously, brain-specific miR-93-5p has been reported to increase in small EVs of serum in patients with ischemic stroke37. In our study, miR-93-5p was downregulated in EVs derived from both primary midbrain and cortical neurons, and further was even consistently decreased in CSF-derived EVs, making miR-93-5p a potentially significant biomarker for PD. Additionally, a report has shown that exosomal miRNA profiles in serum and cerebrospinal fluid found that miR-93-3p, which is derived from the same precursor as miR-93-5p, was significantly downregulated in the CSF of patients with neurosyphilis38. A study profiling blood miRNAs expression in sporadic Creutzfeldt–Jakob disease (sCJD) identified miR-93-5p as differentially expressed compared to healthy controls. Interestingly, miR-93-5p was downregulated in sCJD patients but upregulated in Alzheimer’s disease (AD) patients39. This contrasting expression pattern also suggested that miR-93-5p could serve as a biomarker to differentiate between sCJD and AD39. Moreover, steady low levels of miR-30a-5p and miR-484 in the serum of rotenone-treated rats could serve as potential biomarkers for an early stage of PD. While there aren’t reports of alterations of miR-484 in EVs, the miRNA is known to be upregulated in the cortex and hippocampus regions of AD mouse models and plasma of AD patients40. Hence, exosomal miR-484 shows promise as a biomarker for various neurodegenerative diseases, and the downregulation of miR-484 in serum EVs corroborates with our hypothesis. Our previous report demonstrated the critical role of miR-30a-5p downregulation in modulating in vitro PD conditions23, and the findings from serum EVs in this study align with those results. Our current report, along with various previous studies, has now proven that modulation of miRNAs in EVs reflects disease-specific changes and regulates key cellular processes, making them valuable biomarkers for early diagnosis and monitoring of diseases like PD. Interestingly, CSF EVs encompass the population of brain-derived exosomes41, which is why we extended our analysis to include the levels of miRNAs in the brain tissues as well (the primary source of brain-derived exosomes). Examining miRNA levels and their modulation in the brain becomes imperative due to the potential insights it offers into the pathogenesis and its implications in functions. Interestingly, we observed significantly low levels of miR-93-5p in CSF. Notably, the ventral midbrain region stands out with a remarkably enriched level of miR-93-5p, a finding that may bear significance in the context of pathogenesis. Further exploration of the functions of this particular miRNA could provide valuable insights into its potential role in the underlying mechanisms of the observed pathogenesis.
The CSF is separated from the blood by the presence of a selective physiological barrier, the blood-BCSFB, which modulates small molecule exchange between the two biofluids. Interestingly, exosome transport through the physiological barrier, the blood–brain barrier (BBB) has been well-studied hence exosomal cargo from brain can be transferred to sites outside the brain giving rise to extra neuronal phenotype in PD42,43. A study reported the presence of astrocyte-specific exosome protein cargo in the blood of rats, demonstrating the bidirectional movement of exosomes across BBB44. The miRNA changes we observed in the rotenone-rat model indicated that miR-181c-5p is significantly high in CSF at 8 h of rotenone treatment, but its levels in serum are low. Contrarily, at 24 h of rotenone treatment, the miR-181c-5p levels drop significantly in the CSF and are elevated in the serum, which could be indicative of the bidirectional pathway of the exosomal cargo across the BCSFB.
Emerging reports suggest that exosomal miRNAs are vital in mediating mitochondrial functions and affect the mitochondrial activities in the cell, which is a distinct hallmark of PD45. Both R8 exo and R24 exo induced increased cellular ROS as well as mitochondrial-specific ROS in primary midbrain cultures. A similar study has shown that serum exosomes from PD mouse models contain elevated miR-137 levels which affects the oxidative stress in neurons46. Two independent studies have shown that microglial exosomes induced by LPS and α-synuclein, respectively, induce neuronal apoptosis leading to neurodegeneration47,48. However, the effect of rotenone-induced serum exosomes had not been explored for their functional capabilities. On observing significant loss in cell viability due to the treatment of serum exosomes, we confirmed that the cell death observed in the primary midbrain neuronal cultures is specific to the dopaminergic neuron population.
The impact of exosomes on neurons already sensitized to rotenone remains unclear. Since R8 exosomal cargo induced mitochondrial alterations and cell death, subsequent experiments focused on treating rotenone-exposed primary midbrain neurons with R8 exosomes. DNA damage ELISA and cell viability tests showed exacerbated cell death with combined treatments compared to individual ones. Additionally, specific dopaminergic neurons were significantly reduced in the combined treatment, aligning with PD’s hallmark dopaminergic neuron loss. These findings suggest that R8 exosome cargo may accelerate dopaminergic degeneration under rotenone-induced stress.
To investigate whether the observed neuronal cell death and changes were driven by serum-derived EV-miRNAs, we focused on miR-30a-5p and miR-484, which were consistently downregulated in serum EVs. The synthesized mimics of these miRNAs were transfected in SH-SY5Y neuroblastoma cells to assess their ability to counteract the observed effects. Strikingly, the transfection of these miRNAs alleviated rotenone-induced ROS production and mitigated cell death. miR-484 has been previously reported to play a role in neurodegeneration because it is known to regulate mitochondrial fission by targeting Fis1, which is crucial for maintaining mitochondrial function and apoptosis49. Moreover, it has also been reported to be involved in the regulation of ER stress, oxidative stress, and inflammation50.
However, the combined transfection of both miRNAs 30a-5p and 484 did not provide an additive rescue effect, suggesting that both miRNAs may target a shared pathway involved in mitochondrial function rather than distinct pathways. On performing putative bioinformatic analysis, we identified numerous mRNA targets of these miRNAs, suggesting their involvement in mitochondrial processes and apoptotic cell death. The major pathways modulated by the targets included cell-cell communication, regulation of apoptosis, stress response, and cell death. Notably, the primary localization of these targets was identified in axons, synapses, and the dendritic tree.
Importantly, while the synthetic miRNA mimics demonstrated protective effects in this controlled setting, the physiological outcome could differ. In a natural context, the regulation of cellular functions involves the intricate interplay of numerous miRNAs, each fine-tuned to maintain homeostasis. PD-related toxicity, such as that induced by rotenone, likely disrupts the balance of multiple miRNAs simultaneously, contributing to mitochondrial dysfunction and subsequent neurodegeneration.
While we attempted various methods to characterize the EVs isolated from their respective sources, including TEM imaging to confirm membrane-bound structures and NTA analysis to classify them by size, a difficult-to-address limitation of this study is that due to volumetric constraints, the isolation techniques used resulted in a mixture of EVs and non-vesicular components. TEM data provided strong evidence of membrane-bound vesicles, supporting our use of the term “EVs” throughout this manuscript, and suggested that the presence of non-vesicular particles was negligible. However, the inclusion of some non-vesicular components in our isolates remains unavoidable. Future studies should further classify these extracellular particles based on size, cellular origin, or mode of biogenesis, as these characteristics have significant implications for understanding their functional roles. The size difference reflects distinct formation mechanisms, offering insights into cellular processes and potential functions51. Different-sized EVs often carry unique molecular cargoes, which can indicate their cellular origin and influence their biological effects52,53. Incorporating such advanced characterization techniques enhances the ability to distinguish vesicular from non-vesicular components, ensuring more precise classification and functional interpretation of these extracellular particles.
While our study successfully identifies the functional outcomes of serum exosomes on primary midbrain neurons, we have identified important additional gaps to be addressed in the near future. Current limitations include a lack of understanding of a specific miRNA or pathway responsible for these effects, though our studies have identified potential targets. The observed outcome is a culmination of multiple miRNAs undergoing alterations in PD. It becomes imperative to explore combinatorial strategies for a comprehensive understanding of pathogenesis. Furthermore, our findings highlight the dynamic nature of miRNA changes. Therefore, it is crucial to assess specific miRNA levels in both acute and chronic conditions, spanning various age groups. This approach is essential for developing a robust potential biomarker for PD. Moreover, in future studies, time courses will be essential to track exosomal changes with pathological stage and the emergence of functional deficits, where multiple interception pathways and time points can also be tested.
In conclusion, this study showed the enhanced extracellular vesicle release by rotenone-induced mitochondrial dysfunction from both primary and midbrain neurons as well as acute rotenone model of rat in vivo. Further, specific miRNAs (like miR-93-5p) were consistently downregulated in CSF, and the levels of miR-30a-5p and miR-484 were differentially present in the EVs from primary and rat model. These findings provide insight into the exploitation of EV-miRNAs in biofluids as potential diagnostic/prognostic biomarkers for early detection of PD. Further, serum EVs, when treated to primary midbrain neurons, elicited mitochondrial defects in terms of ROS generation, leading to neuronal cell death. This cell death was also evident (and in some cases more pronounced) when the recipient cells were pretreated with rotenone along with receiving EVs from the serum. In conclusion, the EV cargo changes the physiological status of the recipient cell that they are uptaken in, thereby altering the pathological outcome in PD.
Methods
Acute neurotoxic PD-rat model
Sprague-Dawley rats were bred in-house for the study. Male rats aged 3 months (young adults) were used for the study. All the rats were housed under a standard 12 h light cycle, with free access to water, and fed ad libitum. All the methods used in the study were approved by the Purdue Animal Care and Use Committee (PACUC). Rats were divided into three following groups: treated with vehicle (V) (n = 11), treated with rotenone for 8 h (R8) (n = 10), and treated with rotenone for 24 h (R24) (n = 11). The rotenone dose (3 mg/kg) and acute time points were chosen to capture preclinical changes prior to cell loss and behavioral phenotypes. Repeated rotenone dosing produces a well-defined end-stage phenotype (weeks of exposure, where timeline is age dependent). Here, we focused on evaluation at acute time points where known mitochondrial dysfunction, mitophagy alterations, and protein aggregations occur in the nigrostriatal dopamine system and other brain regions affected in early-stage PD, each of which are known primary pathogenic pathways in PD antecedent to DA cell death. Importantly, at these time points, these preclinical alterations occur in the absence of late-stage pathology21,54.
The rotenone solution was prepared fresh in 98% Miglyol 812N and 2% DMSO (dimethylsulfoxide), which on vortexing, produces a stable emulsion of DMSO containing Miglyol 812N and rotenone. The rotenone was administered to the rats intraperitoneally and sacrificed 8 or 24 h after injections. Rats were anaesthetized by isoflurane gas, after which CSF collection was performed from the cisterna magna by direct puncture as described in the protocol55. After CSF collection, blood from rats was collected using cardiac puncture. Post-euthanasia, the brain was rapidly removed, and brain regions, like dorsal and ventral striatum, dorsal and ventral midbrain, cerebral cortex, and hippocampus were dissected on ice, flash frozen, and then stored at −80 °C for processing of miRNA analysis.
Isolation of primary neurons and treatment of rotenone
Female pregnant Sprague-Dawley rats were euthanized using methods approved by the Purdue Animal Care and Use Committee, and embryonic E17 brains were dissected56,57,58. The cortex and mesencephalic region, which contains the substantia nigra and ventral tegmental area, were isolated stereoscopically. The isolated tissue was treated with papain (20 units/mL in 1X HBSS), and a dissociated cell suspension was generated by vigorous pipetting. The cell suspension was plated on poly-D-Lysine-coated plates in media containing Neurobasal, 2% B-27 supplement, 5% fetal bovine serum (FBS), Glutamax, and 1% (w/v) Pen-Strep. The plating media is replaced after 24 h by the complete Neurobasal media supplemented with B-27 and Glutamax but lacking FBS. The neurons were maintained in this media for at least 6 days (DIV-6) before the commencement of treatments.
Isolation of EVs from CSF and serum
A starting volume of 10 µl CSF was used for EV extraction specifically for NTA analysis. Furthermore, 100 µl CSF was used for isolating EVs for EV-miRNA analysis. CSF was centrifuged at 2000×g for 30 min at 4 °C to remove cells and debris, and supernatant was collected and centrifuged for an additional 30 min at 10,000×g at 4 °C. Clarified CSF was transferred to a new tube, and an equal volume of the Total Exosome Isolation reagent (from other body fluids) (Thermo Fisher Scientific, Invitrogen, USA; Cat No: 4484453) was added and mixed thoroughly and was incubated at 4 °C for 1 h. After incubation, the samples were centrifuged at 10,000×g for 1 h at 4 °C, and the exosome pellet was collected and processed further accordingly.
A starting volume of 25 and 500 µl serum was used for isolating EVs for NTA and EV-miRNA analysis, respectively. The serum was centrifuged at 2000×g for 30 min at 4 °C to remove cellular debris. Total Exosome Isolation reagent (from serum) (Thermo Fisher Scientific, Invitrogen, USA; Cat No: 4478360) was added in 5:1 ratio volume and incubated at 4 °C for 30 min. After incubation, the samples were centrifuged at 10,000×g for 30 min at 4 °C to obtain the pellet.
Isolation of EVs from primary neurons
Primary cortical and midbrain neurons were seeded at a density of 6 × 105 and 8 × 105 cells per well in six-well plates, respectively (containing 2 ml of media per well) and were treated with rotenone in neurobasal media (lacking FBS). About 2 ml of media was collected 24 h posttreatment to isolate exosomes. The media was centrifuged at a low speed of 2000×g for 30 min to remove the cell debris and larger particles. The resultant supernatant was collected and total exosome isolation (from cell culture media) (Thermo Fisher Scientific, Invitrogen, USA; Cat No: 4478359) was added in 2:1 ratio volume. The mixture was incubated overnight at 4 °C. Following incubation, the EV pellet is obtained by centrifuging the mixture at 10,000×g for 1 h at 4 °C. The EV pellet was either used for further downstream analysis and/or was kept at −80 °C for long-term storage.
Nanoparticle tracking analysis (NTA)
Primary cortical and midbrain neurons were seeded at a density of 3 × 105 and 4 × 105 cells/well in 12 well plates, respectively, and treated with rotenone. The media was collected, and EVs were isolated by the method described above. The resultant pellet was resuspended and dissolved in 1 ml of 1X PBS (phosphate-buffered saline) solution and was analyzed by the NanoSight LM10 (Malvern Panalytical, UK), which shows both the particle size as well as concentration.
The EVs from the biofluids were isolated according to the method described above and were resuspended in 1 ml of 1X PBS solution and were analyzed by NTA. The resultant data were plotted as a concentration of particles per ml of the solution resuspended. The subpopulation data were obtained by manually calculating the concentrations of populations from each size range obtained in the experiment summary data file from the NanoSight NTA 3.2 analysis tool and plotted as the relative percentage of each subpopulation.
Transmission electron microscopy of EVs from CSF and serum
EVs from CSF and serum were isolated using the procedure mentioned above. The resultant EV pellet was resuspended and washed in 2% paraformaldehyde (PFA) in PBS and again filtered with a 0.2-micron filter. MISEV guidelines were followed to ensure the protocol was adapted to EVs7. The protocol for TEM was followed from previous reports with slight modifications59,60. Briefly, 10 µL of EV suspension was deposited onto glow-discharged formvar-carbon-coated EM grids and allowed to adsorb for 10 min. The grids were rinsed five times with 100 µL of PBS to remove unbound material, followed by transfer to a 1% glutaraldehyde solution for 5 min to fix the EVs. The grids were then washed with distilled water six to eight times to ensure thorough removal of glutaraldehyde. For contrasting and embedding, the grids were transferred to a 50 µL solution of uranyl oxalate for 5 min and subsequently placed in a mixture of 4% uranyl acetate and 2% methylcellulose (100 µL/900 µL, respectively) for final contrast and embedding60. This procedure preserved the bilayer morphology of the EVs. Finally, the excess stain was gently blotted off, and the grids were air-dried before imaging with a TEM at 80–120 kV to confirm the characteristic size and morphology of the EVs.
RNA isolation and quantitation
Total RNA was isolated from the primary neurons, brain tissue, and as well as EV samples using RNAzol reagent (Molecular Research Center, Inc., USA). The resultant RNA was subjected to Poly-A tailing (for small RNA) using E. coli Poly-A Polymerase (New England Biolabs, UK) at 37 °C for 30 min. cDNA was synthesized using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA), and using the oligo dT primers. Primers for miRNAs were designed according to previous reports (Table 1)61. Real-time PCR of the samples was performed using the THUNDERBIRDTM SYBR qPCR mix (Toyobo, Japan).
To ensure reliable normalization of exosomal miRNA expression, we selected an appropriate endogenous control based on previous literature. Reports indicate that small nucleolar RNAs are present in EVs, albeit in low quantities, confirming their existence62,63. Moreover, many studies64,65, including those investigating biomarkers in CSF from AD and PD patients15, have used small nuclear RNAs as endogenous controls for exosomal miRNA analysis in patient biofluids. To select an appropriate endogenous control for our study, we referred to two reports that analyzed small RNAs in rat models of endometrial cancer and retinopathy66,67. Based on these findings and the established presence of small nuclear RNAs in EVs, we initially considered 4.5S RNA(H) A, snoRNA, U87, and RNU6B (U6 snRNA) as potential candidates. However, 4.5S RNA(H) A and snoRNA were undetectable in our EV fractions, and RNU6B exhibited variability across samples (data not included in the manuscript). Given its consistent expression and stability, U87 was selected as the most suitable endogenous control for our study. The levels of miRNAs were determined by the 2−ΔΔCT method. The PCR reaction conditions were as follows: 95 °C for 2 min followed by 50 cycles of 95 °C for 5 s and 60 °C for 1 min. The melt curves were also acquired.
Primary neuron treatment with EVs
The serum-derived EVs were isolated using the method described above, and the protein was quantified using BCA (Bicinchoninic) assay. About 50 μg/mL of EVs were incubated with primary cortical and midbrain neurons at a density of 75,000 and 100,000 cells/per well in 48-well plates. EVs were first washed with sterile phosphate-buffered saline (PBS) and filtered through a 0.2-µm membrane filter to ensure sterility. Subsequently, the primary neurons were incubated with the EVs for a period of 24 h. Viability was assessed by multiple methods indicated below.
MTT assay
Primary cortical and midbrain neurons were plated at the density of 75,000 and 100,000 cells/per well in 48-well plates. After treatment with either rotenone or serum-derived EVs for 24 h, an MTT assay was performed as described previously68. Briefly, the cells were first incubated with MTT (0.1 mg/ml) at 37 °C for 1 h. The visible violet crystals were solubilized using DMSO, and absorbance at 570 nm was recorded using a microplate reader. The results are displayed as the relative cell viability percentage.
Cell death detection by ELISA
The cell death detection by ELISA was performed using a kit-based assay (Roche, Germany). Briefly, primary cortical and midbrain neurons were plated in 48-well plates at a density of 75,000 and 100,000 cells/per well and treated with the mentioned treatments for 24 h. Cells were harvested posttreatment and lysed using the kit’s lysis buffer for 30 min and centrifuged at 8000×g for 10 min to settle down the cell debris. The supernatant containing cytoplasmic proteins was incubated on streptavidin-coated 96-well plates, along with biotin-tagged anti-histone and anti-DNA-POD antibodies. After 2 h of incubation, the mixture was aspirated from the plates, plates were washed thrice, and substrate solution from the kit was added, for which a blue-purple-colored solution was developed. The reaction was stopped using a stop buffer, and absorbance was measured at 405 and 490 nm. The enrichment factor was calculated using the formula: mU sample/mU control.
Immunocytochemistry for midbrain dopaminergic neurons
The specific dopaminergic neurotoxicity in primary midbrain neurons was assessed by immunocytochemistry. Primary midbrain neurons were plated at a density of 2 × 105 cells/per well in a 24-well plate and were given the mentioned treatments for 24 h. After treatment, the cells were fixed using 4% PFA (paraformaldehyde) for 30 min at 4 °C. After fixing, the cells were rinsed with PBS and incubated with the primary antibodies: Tyrosine hydroxylase (TH) (AB9983) (Millipore, Sigma, USA), which was specific to dopaminergic neurons and MAP2 (AB5622) (Millipore, Sigma, USA), a general neuronal marker at 4 °C overnight. The primary antibody was prepared in 1X PBS containing 10% NDS and 0.3% Triton-X 100. After overnight incubation with primary antibody, the cells were washed with PBS buffer, thrice for 5 min. Following, the cells were incubated with the respective fluorescent-tagged secondary antibody for 2 h at room temperature and were imaged by confocal microscope (Nikon) using a 20X objective.
Transfection of synthetic miRNA mimics in in vitro SH-SY5Y cells and treatment of rotenone
SH-SY5Y cells were initially treated with varying concentrations of rotenone to determine the optimal dose, which was found to be 50 nM. Based on this, 50 nM rotenone was used for all subsequent treatments. Pre-made miRNA mimics from Thermo Fisher were used for the experiments: hsa-miR-30a-5p assay ID: MC11062; has-miR-484 assay ID: MC10379; Cat No: 4464066; mirVana™ miRNA Mimic, Negative Control #1; Cat No: 4464058). Mimics were transfected into the cells using Lipofectamine RNAiMax (Invitrogen, USA) (Cat No: 13778150), with the transfection allowed to proceed overnight. The following day, the media was changed, and the cells were treated with rotenone for 24 h. Mitochondrial assays and cell death assays were then performed as previously described.
Bioinformatic analysis
MiRNA-mRNA target interactions were analyzed using the ENCORI/starBase database (http://starbase.sysu.edu.cn/)69. To determine the functional pathways associated with the selected miRNAs, all miRNAs within each category were considered, combining three target prediction tools and ClipSeq data with low stringency, applying a corrected p value threshold of <0.05. Gene Ontology (GO) terms and pathways related to these miRNA targets were retrieved70. The GO terms were categorized by selecting the top ten terms with the highest gene involvement from the mRNA targets. The selection was further refined by a significance threshold, specifically choosing terms with a p value of <0.05. Cytoscape software was utilized to visualize the cellular targets of the two selected miRNAs.
Statistical analysis and figure creation
Data were shown as mean ± SEM for the number of times the experiment was repeated. Comparisons for columns (two groups) were performed using student's t-test for repeated measurements to determine the levels of significance for each group. Comparisons between groups was done by either one-way or two-way ANOVA, using Dunnett’s (comparison back to control), Tukey’s (comparison of all groups), or Sidak’s (where only select comparisons were scientifically warranted) multiple comparisons test. The experiments were performed a minimum of three times independently and p < 0.05 was considered statistically significant. GraphPad Prism (version 9) software was used to perform all the statistical analysis and create graphs. The graphical abstract and Supplementary Fig. 1 is original artwork created using Biorender. This software has both freely available and purchasable versions. Here, we utilized the premium version, which includes publishing and licensing rights.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Bloem, B. R., Okun, M. S. & Klein, C. Parkinson’s disease. Lancet 397, 2284–2303 (2021).
Armstrong, M. J. & Okun, M. S. Diagnosis and treatment of Parkinson disease: a review. JAMA 323, 548 (2020).
Oertel, W. & Schulz, J. B. Current and experimental treatments of Parkinson disease: a guide for neuroscientists. J. Neurochemistry 139, 325–337 (2016).
Gibb, W. R. & Lees, A. J. The relevance of the Lewy body to the pathogenesis of idiopathic Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 51, 745–752 (1988).
Jamebozorgi, K. et al. Cellular and molecular aspects of Parkinson treatment: future therapeutic perspectives. Mol. Neurobiol. 56, 4799–4811 (2019).
Properzi, F., Logozzi, M. & Fais, S. Exosomes: the future of biomarkers in medicine. Biomark. Med. 7, 769–778 (2013).
Welsh, J. A. et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J. Extracell. vesicles 13, e12404 (2024).
Harding, C. V., Heuser, J. E. & Stahl, P. D. Exosomes: looking back three decades and into the future. J. Cell Biol. 200, 367 (2013).
Li, J., Lu, S., Chen, F. & Zhu, H. Unveiling the hidden role of extracellular vesicles in brain metastases: a comprehensive review. Front. Immunol. 15, 1388574 (2024).
Hessvik, N. P. & Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 75, 193–208 (2018).
De Toro, J., Herschlik, L., Waldner, C. & Mongini, C. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front. Immunol. 6, 203 (2015).
Shi, M. et al. Plasma exosomal α-synuclein is likely CNS-derived and increased in Parkinson’s disease. Acta Neuropathol. 128, 639–650 (2014).
Majidinia, M. et al. The roles of non-coding RNAs in Parkinson’s disease. Mol. Biol. Rep. 43, 1193–1204 (2016).
Quinlan, S., Kenny, A., Medina, M., Engel, T. & Jimenez-Mateos, E. M. MicroRNAs in neurodegenerative diseases. Int. Rev. Cell Mol. Biol. 334, 309–343 (2017).
Gui, Y., Liu, H., Zhang, L., Lv, W. & Hu, X. Altered microRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 6, 37043–37053 (2015).
Yao, Y., Qu, M., Li, G., Zhang, F. & Rui, H. Circulating exosomal miRNAs as diagnostic biomarkers in Parkinson’s disease. Eur. Rev. Med. Pharm. Sci. 22, 5278–5283 (2018).
Ravanel, P., Tissut, M. & Douce, R. Effects of rotenoids on isolated plant mitochondria. Plant Physiol. 75, 414–420 (1984).
Betarbet, R. et al. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 3, 1301–1306 (2000).
Huang, J. et al. Involvement of glyceraldehyde-3-phosphate dehydrogenase in rotenone-induced cell apoptosis: relevance to protein misfolding and aggregation. Brain Res. 1279, 1–8 (2009).
Xiong, N. et al. Stereotaxical infusion of rotenone: a reliable rodent model for Parkinson’s disease. PLoS ONE 4, e7878 (2009).
Cannon, J. R. et al. A highly reproducible rotenone model of Parkinson’s disease. Neurobiol. Dis. 34, 279–290 (2009).
Tanner, C. M. et al. Rotenone, paraquat, and Parkinson’s disease. Environ. Health Perspect. 119, 866–872 (2011).
Currim, F. et al. Neuronal exosomal miRNAs modulate mitochondrial functions and cell death in bystander neuronal cells under Parkinson’s disease stress conditions. Neurotoxicology 101, 102–116 (2024).
Ryan, B. J., Hoek, S., Fon, E. A. & Wade-Martins, R. Mitochondrial dysfunction and mitophagy in Parkinson’s: from familial to sporadic disease. Trends Biochem. Sci. 40, 200–210 (2015).
Bogaerts, V., Theuns, J. & Van Broeckhoven, C. Genetic findings in Parkinson’s disease and translation into treatment: a leading role for mitochondria? Genes Brain Behav. 7, 129–151 (2008).
Currim, F. et al. Exosome release is modulated by the mitochondrial-lysosomal crosstalk in Parkinson’s disease stress conditions. Mol. Neurobiol. 58, 1819–1833 (2021).
Willms, E. et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci. Rep. 6, 22519 (2016).
Colombo, M. et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J. Cell Sci. https://doi.org/10.1242/jcs.128868 (2013).
Zhang, H. et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 20, 332–343 (2018).
D’Souza, A. et al. Microvesicles transfer mitochondria and increase mitochondrial function in brain endothelial cells. J. Control. Release 338, 505–526 (2021).
Parnetti, L. et al. CSF and blood biomarkers for Parkinson’s disease. Lancet Neurol. 18, 573–586 (2019).
Goldman, J. G. et al. Cerebrospinal fluid, plasma, and saliva in the BioFIND study: relationships among biomarkers and Parkinson’s disease features. Mov. Disord. 33, 282–288 (2018).
Ding, H. et al. Identification of a panel of five serum miRNAs as a biomarker for Parkinson’s disease. Parkinsonism Relat. Disord. 22, 68–73 (2016).
Cardo, L. F. et al. Profile of microRNAs in the plasma of Parkinson’s disease patients and healthy controls. J. Neurol. 260, 1420–1422 (2013).
Li, Y. et al. Dynamic regulation of circulating microRNAs during acute exercise and long-term exercise training in basketball athletes. Front. Physiol. 9, 282 (2018).
Ponomaryova, A. A. et al. Dynamic changes in circulating miRNA levels in response to antitumor therapy of lung cancer. Exp. Lung Res. 42, 95–102 (2016).
Zhou, X. et al. Acute cerebral ischemia increases a set of brain-specific miRNAs in serum small extracellular vesicles. Front. Mol. Neurosci. 15, 874903 (2022).
Chen, H. et al. Exosomal microRNA profiles from serum and cerebrospinal fluid in neurosyphilis. Sex. Transm. Infect. 95, 246–250 (2019).
Norsworthy, P. J. et al. A blood miRNA signature associates with sporadic Creutzfeldt-Jakob disease diagnosis. Nat. Commun. 11, 3960 (2020).
Wingo, T. S. et al. Brain microRNAs associated with late-life depressive symptoms are also associated with cognitive trajectory and dementia. NPJ Genom. Med. 5, 6 (2020).
Patz, S. et al. More than cell dust: microparticles isolated from cerebrospinal fluid of brain injured patients are messengers carrying mRNAs, miRNAs, and proteins. J. Neurotrauma 30, 1232–1242 (2013).
Alvarez-Erviti, L. et al. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 29, 341–345 (2011).
Abdelsalam, M., Ahmed, M., Osaid, Z., Hamoudi, R. & Harati, R. Insights into exosome transport through the blood–brain barrier and the potential therapeutical applications in brain diseases. Pharmaceuticals 16, 571 (2023).
Gómez-Molina, C. et al. Small extracellular vesicles in rat serum contain astrocyte-derived protein biomarkers of repetitive stress. Int. J. Neuropsychopharmacol. 22, 232–246 (2019).
Wang, Y., Jin, P., Liu, J. & Xie, X. Exosomal microRNA-122 mediates obesity-related cardiomyopathy through suppressing mitochondrial ADP-ribosylation factor-like 2. Clin. Sci. 133, 1871–1881 (2019).
Jiang, Y. et al. Serum secreted miR-137-containing exosomes affects oxidative stress of neurons by regulating OXR1 in Parkinson’s disease. Brain Res. 1722, 146331 (2019).
Chen, X. et al. A20 protects neuronal apoptosis stimulated by lipopolysaccharide-induced microglial exosomes. Neurosci. Lett. 712, 134480 (2019).
Chang, C. et al. Exosomes of BV-2 cells induced by alpha-synuclein: important mediator of neurodegeneration in PD. Neurosci. Lett. 548, 190–195 (2013).
Wang, K. et al. miR-484 regulates mitochondrial network through targeting Fis1. Nat. Commun. 3, 781 (2012).
Jia, Y., Liu, J., Wang, G. & Song, Z. miR-484: a potential biomarker in health and disease. Front. Oncol. 12, 830420 (2022).
van de Wakker, S. I. et al. Size matters: functional differences of small extracellular vesicle subpopulations in cardiac repair responses. J. Extracell. Vesicles 13, 12396 (2024).
Collino, F. et al. Exosome and microvesicle-enriched fractions isolated from mesenchymal stem cells by gradient separation showed different molecular signatures and functions on renal tubular epithelial cells. Stem Cell Rev. Rep. 13, 226–243 (2017).
Kumar, M. A. et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct. Target. Ther. 9, 27 (2024).
Wise, J. P. Jr., Price, C. G., Amaro, J. A. & Cannon, J. R. Autophagy disruptions associated with altered optineurin expression in extranigral regions in a rotenone model of Parkinson’s disease. Front. Neurosci. 12, 289 (2018).
Li, Y., Zhang, B., Liu, X.-W., Liu, M. & Huang, S.-M. An applicable method of drawing cerebrospinal fluid in rats. J. Chem. Neuroanat. 74, 18–20 (2016).
Volpicelli-Daley, L. A., Luk, K. C. & Lee, V. M. Addition of exogenous α-synuclein preformed fibrils to primary neuronal cultures to seed recruitment of endogenous α-synuclein to Lewy body and Lewy neurite–like aggregates. Nat. Protoc. 9, 2135–2146 (2014).
de Rus Jacquet, A. et al. Neuroprotective mechanisms of red clover and soy isoflavones in Parkinson’s disease models. Food Funct. 12, 11987–12007 (2021).
Landeck, N. et al. Two C-terminal sequence variations determine differential neurotoxicity between human and mouse α-synuclein. Mol. Neurodegeneration 15, 1–23 (2020).
VainIo, S (ed.) Cell-Secreted Vesicles: Methods and Protocols Vol. 2668 (Springer, 2023).
Théry, C., Amigorena, S., Raposo, G. & Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 30 (2006).
Ingrid, B. Cirera, S. & Peter K. Busk. Specific and sensitive quantitative RT-PCR of miRNAs with DNA primers. BMC biotechnology 1, 1-11 (2011).
Huang, X. et al. Characterization of human plasma-derived exosomal RNAs by deep sequencing. BMC Genomics 14, 1–14 (2013).
Zhao, F. et al. Characterization of serum small extracellular vesicles and their small RNA contents across humans, rats, and mice. Sci. Rep. 10, 4197 (2020).
Aushev, V. N. et al. Comparisons of microRNA patterns in plasma before and after tumor removal reveal new biomarkers of lung squamous cell carcinoma. PLoS ONE 8, e78649 (2013).
Munch, E. M. et al. Transcriptome profiling of microRNA by Next-Gen deep sequencing reveals known and novel miRNA species in the lipid fraction of human breast milk. PLoS ONE 8, e50564 (2013).
Jurcevic, S., Olsson, B. & Klinga-Levan, K. Validation of suitable endogenous control genes for quantitative PCR analysis of microRNA gene expression in a rat model of endometrial cancer. Cancer Cell Int. 13, 1–6 (2013).
Tea, M., Michael, M. Z., Brereton, H. M. & Williams, K. A. Stability of small non-coding RNA reference gene expression in the rat retina during exposure to cyclic hyperoxia. Mol. Vis. 19, 501 (2013).
Shukla, S. et al. hsa-miR-320a mediated exosome release under PD stress conditions rescue mitochondrial ROS and cell death in the recipient neuronal and glial cells. Int. J. Biochem. Cell Biol. 162, 106439 (2023).
Yang, J.-H. et al. starBase: a database for exploring microRNA–mRNA interaction maps from argonaute CLIP-seq and degradome-seq data. Nucleic Acids Res. 39, D202–D209 (2011).
Dennis, G. et al. DAVID: database for annotation, visualization, and integrated discovery. Genome Biol. 4, 1–11 (2003).
Acknowledgements
Authors acknowledge financial support by the National Institutes of Health to JRC (R01ES035019 and R01ES025750) and J.H.S. (R01ES033173). The authors also acknowledge the SERB-OVDF (Overseas Visiting Doctoral Fellowship) for the manpower funding to work at Purdue University, West Lafayette, Indiana, USA, and acknowledge the Branfman Foundation. This work is a part of the Ph.D. thesis of Fatema Currim. The graphical abstract is created with https://BioRender.com. (Created in BioRender. Currim, F. (2025) https://BioRender.com/u25k757).
Author information
Authors and Affiliations
Contributions
F.C.: Investigation, validation, formal data analysis, writing—original draft, methodology, and resources. J.B.-L., T.S., M.C., and S.S.: Formal data analysis, methodology, and resources. J.H.S., J.-C.R., and W.Q.: Methodology and resources. P.B.: Formal data analysis. R.S.: Conceptualization, visualization, writing—original draft, critical revision of the article, and supervision. J.H.S.: Conceptualization, methodology, resources, writing—original draft, visualization, supervision, and funding acquisition. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Currim, F., Brown-Leung, J., Syeda, T. et al. Rotenone induced acute miRNA alterations in extracellular vesicles produce mitochondrial dysfunction and cell death. npj Parkinsons Dis. 11, 59 (2025). https://doi.org/10.1038/s41531-025-00917-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41531-025-00917-0
