
Abstract
The gut-muscle axis represents a key regulatory pathway linking intestinal microbiota to muscle development and meat quality, yet its mechanistic basis in geese remains unclear. This study investigated how dietary perennial ryegrass modulates meat quality through microbiota reshaping and AMPK/Akt-mTOR signaling. Geese were fed a commercial diet (CD) or a ryegrass feeding (GD) for 30 days. GD markedly improved breast muscle morphology, increasing fiber density and reducing cross-sectional area, while enhancing meat color, texture, protein conformation, and reducing cooking loss and shear force. Amino acid and fatty acid enrichment, together with elevated antioxidant activity in serum, ileum and muscle, further supported quality improvement. 16S rRNA sequencing revealed increased Bacteroides, Akkermansia, Faecalibacterium, and Butyricicoccus abundances and higher short-chain fatty acid levels. Western blot and immunofluorescence confirmed AMPK activation and Akt-mTOR inhibition. These findings identify perennial ryegrass as a functional dietary strategy to enhance geese meat quality via gut-muscle metabolic crosstalk.
Similar content being viewed by others


Introduction
China is the world’s leading producer and consumer of geese, contributing approximately 5.5 million tons, representing about 76% of global consumption1. The species Anser cygnoides serves as an important livestock resource in the country, traditionally reared for its nutrient-dense meat. Goose meat is recognized for its low cholesterol content (52–76 mg/100 g), high protein and high polyunsaturated fatty acid (PUFA)2. Historically, geese have been raised under extensive systems using bulky forage-based diets; however, these traditional methods often compromise growth performance and production efficiency3. Moreover, geese raised without access to pasture tend to yield meat of lower nutritional quality compared to those supplemented with grass4. Therefore, integrating functional forages such as ryegrass into intensive production systems may offer a balanced solution for improving both growth efficiency and meat quality. Optimizing both growth performance and meat quality is an evolving priority in livestock production systems, driven by consumer demand for healthier, more sustainable protein sources5. Meat quality-related parameters, such as flavor, TPA profile, muscle fiber, color, oxidative stability, lipid metabolic profile, amino acids, protein secondary structure and fatty acid composition have recently attracted consumers’ attention6. Dietary strategies that enhance these traits, particularly through natural feed additives, are gaining traction. Perennial ryegrass (Lolium perenne), known for its digestibility and high nutritional value, also contains bioactive compounds such as polyphenols and flavonoids with potential systemic benefits7. Recent studies suggest that these benefits may be mediated through modulation of the gut microbiota, a critical regulator of host metabolism and immunity8.
The gut microbiota in the gastrointestinal tract (GIT) is crucial for nutrient metabolism, energy balance, and gut barrier integrity. Consumption of ryegrass has been shown to selectively enrich beneficial microbial taxa, such as Ruminococcus, Bacteroides, and Lactobacillus, which contribute to increased short-chain fatty acids (SCFAs) production and suppression of pathogenic bacteria8. These SCFAs serve not only as energy sources but also as signaling molecules that influence systemic metabolism, including the activation of adenosine monophosphate-activated protein kinase (AMPK)9.
The AMPK/protein kinase B-mammalian target of rapamycin (Akt-mTOR) signaling axis is crucial for regulating muscle protein synthesis, mitochondrial biogenesis, and fatty acid metabolism. Short chain fatty acids (SCFAs) can activate AMPK signaling in skeletal muscle and hepatic tissues, thereby enhancing oxidative metabolism, improving insulin sensitivity, and reducing lipogenesis10. Conversely, inhibition of the Akt-mTOR signaling by the AMPK pathway11 also downregulates excessive adipogenesis, promotes lean muscle development, and contributes to improved meat quality traits such as marbling, water-holding capacity (WHC), and tenderness12,13.
Despite increasing interest in functional forages and microbiota-targeted nutrition, the mechanistic link between ryegrass-induced microbiota shifts and host metabolic signaling particularly via the “AMPK/Akt-mTOR pathway” remains largely uncharacterized in geese. While prior research has highlighted the nutritional and microbial benefits of ryegrass, no study has yet elucidated how microbial metabolites interact with host signaling networks to influence meat quality outcomes in this species. This study aims to address this gap by evaluating the effects of perennial ryegrass supplementation on meat quality in geese, with a specific focus on gut microbiota composition, SCFAs production, and activation of the AMPK/Akt-mTOR signaling axis. Elucidating this gut-muscle communication may offer new avenues for sustainable meat production strategies that align animal performance with nutritional and health-driven consumer expectations.
Results
Growth performance
Goose body weight was affected by the addition of perennial ryegrass. Table 1 represented the growth performance and carcass traits. ADFI remained unaffected. Body weight (BW) was significantly lower (P < 0.05) in the ryegrass-fed group. Better feed conversion ratio (FCR) was observed in commercial feeding group (CD). There was significant reduction in abdominal fat as a result of ryegrass feeding. Overall growth performance data was presented in Table 1.
H&E analysis of breast muscle
Figure 1a represented breast muscle morphology. Surface area of breast muscle fiber in GD group was significantly lower (P < 0.01) as compared to commercial feeding group; however, muscle fiber density was significantly increased in GD group (Fig. 1b, c). Muscle fiber diameter and perimeter were significantly increased due to ryegrass feeding (Fig. 1d, e). PGC-1a, MyHC- I, MyHC-IIa, and MyHC-IIb play an important role in muscle fiber development. The mRNA expression level of PGC-1a, MyHC-I and MyHC-IIa was higher (P < 0.01) in the breast muscles of GD group, and MyHC-IIb showed lower expression as compared to CD group (Fig. 1f–i). Perennial ryegrass significantly lowered the mRNA expression levels of MSTN (negative regulator of muscle growth) and MURF-1 (muscle protein degrader) by increasing the mRNA expression levels of MyoG (key regulator of muscle cell differentiation and maturation), MyF4 (promotes the formation of muscle fibers) and MyBPH (protein involved in muscle contraction and structure maintaining muscle integrity) as shown in Fig. 1j–m.

a Representative H&E images of breast muscle and the genes regulating the muscle physiology between CD and GD group. Bars = 200 μm. b–e Muscle fiber cross sectional area of breast muscle, fiber density, fiber diameter and fiber perimeter, respectively. f–i PGC-1a (Peroxisome proliferator-activated receptor gamma coactivator 1-alpha), MyHC- I (myosin heavy chain type I), MyHC-II (myosin heavy chain type II) and MyHC-IIb (myosin heavy chain type IIb), respectively. j–m MSTN (myostatin), MyoG (myogenin), MyF4 (myogenic factor 4), and MyBPH (myosin binding protein H), respectively. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01.
Meat quality assessment
Figure 2a showed the breast meat quality indicators. Perennial ryegrass group showed low L* value (P < 0.05), while “a” and “b” values showed no significant difference (Fig. 2b). GD group exhibited a significant decline in cooking loss as compared to commercial feeding group (Fig. 2c). There was no significant difference in drip loss (Fig. 2d). However, there was a significant decrease (P < 0.05) in shear force as result of ryegrass feeding in the GD group (Fig. 2e). The pH values of the breast muscle after 45 min and 24 h of slaughter showed significant difference between the two feeding groups (Fig. 2f). Water holding capacity (WHC) showed significant difference in both groups (Fig. 2g). Regarding water distribution and mobility, the immobile water (T21) was significantly higher in GD group, while bound water (T2b) remained unaffected (Fig. 2h). In addition, free water (T22) content was higher in CD group (Fig. 2h). The TPA analysis exhibited a decreasing trend in terms of chewiness, hardness, cohesiveness, adhesiveness, gumminess and springiness in the GD group (Fig. 2i). Gumminess, cohesiveness and free water were positively correlated with TPA profile as shown by the pearson correlation heatmap (Fig. 2j).

a Breast meat quality indicators in geese. b Color values “L”, “a” and “b”. c Cooking loss. d Drip loss. e Shear force. f pH value at 45 min and pH value after 24 h. g Water holding capacity (WHC). (h) Bound water, immobile water and free water, respectively. TPA profile i hardness, cohesiveness, adhesiveness, gumminess, springiness, and chewiness respectively. j Correlation between WHC profile and meat quality indicators. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01.
Serum biochemistry profile
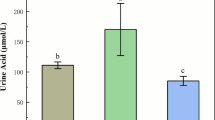
Table 2 showed the biochemical indicators. Perennial ryegrass (GD) significantly altered serum biochemical indicators. The triglycerides (TG), blood urea nitrogen (BUN), glucose (GLU), total cholesterol (TC) and low-density lipoprotein (LDL) were significantly lower (P < 0.01; P < 0.05) in GD group. However, high-density lipoprotein (HDL) showed an increasing trend as a result of perennial ryegrass feeding.
Effect of perennial ryegrass on antioxidant activity
The concentrations of antioxidant enzymes (MDA, TBARS, SOD, CAT, T-AOC and GSH-PX) and the genes responsible for antioxidant activity (Nrf2, HO-1, NQO-1, GCLC, GCLM, and Keap-I) were quantified through ELISA and RT-qPCR, respectively. Perennial ryegrass (GD) enhanced (P < 0.01) the activity of SOD, GSH-PX, CAT, and T-AOC in ileum and breast meat by decreasing the levels of MDA and TBARS as compared to commercial feeding group (CD), as shown in Table 2. Genes responsible for antioxidant activities (Nrf2, HO-1, NQO-1, GCLC, and GCLM) showed significantly higher mRNA expression levels in GD group by lowering Keap-I levels as compared to CD group (Table 2).
Proximate analysis of breast muscle
Crude protein (CP), crude fiber (CF), ash, and dry matter (DM) are crucial meat quality attributes. Breast meat had lower CF and high CP content in the GD group, while showing no difference in terms of ash and DM between the two groups (Table 3).
Fatty acid profile of breast muscle
Perennial ryegrass (GD) had lower saturated fatty acids (SFAs) e.g., C17:0, C20:0, C8:0, and C18:0 content in breast muscle. C14:1 and C16:1 were higher as a result of ryegrass feeding. Perennial ryegrass significantly improved (P < 0.01) the C18:2n6t content of the goose breast muscle (Table 3). Regarding the overall fatty acid percentage, SFAs and n-6/n-3 ratio were significantly lower in the GD group. Perennial ryegrass significantly increased (P < 0.01) monounsaturated fatty acids (MUFA) of the breast muscle of geese (Table 3). Enhanced β-oxidation contributes to improved meat leanness and nutritional quality in poultry and livestock. GD upregulated the mRNA expressions of SIRT1, PPARα, FISL, and CPT-1 (Fig. 3a). Modulating lipogenic gene expression through diet can thus be an effective strategy to enhance meat quality in poultry and livestock. Perennial ryegrass significantly increased the mRNA expression of FADS1, while downregulating the mRNA expression of SREBP-1, FABP, PPAR-γ, and FASN (Fig. 3b).

a SIRT-1 (Sirtuin 1), PPAR-a (peroxisome proliferator-activated receptor alpha), PGC-1a (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), FISL (fatty acid transport protein 1), and CPT-1 (carnitine palmitoyltransferase 1). b SREBP-1 (sterol regulatory element-binding protein 1), FABP (fatty acid-binding protein), PPAR-γ (peroxisome proliferator-activated receptor gamma), FASN (fatty acid synthase), and FADS-1 (fatty acid desaturase 1). c SLC7A9 (solute carrier family 7 member 9), SLC6A19 (solute carrier family 6 member 19), SLC7A1 (solute carrier family 7 member 1), SLC7A5 (solute carrier family 7 member 5), and SLC43A2 (Solute Carrier Family 43 Member 2). d β-sheets, α-helix, β-turn and random coil, respectively. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01.
Amino acid profile of breast muscle
Perennial ryegrass feeding (GD) improved the amino acids namely aspartic acid, glutamate, serine, arginine, glycine, tyrosine, proline, alanine, valine, methionine, cysteine, isoleucine, phenylalanine, histidine, lysine and tyrosine of goose breast muscle (Table 3). Amino acid transporters (SLC7A9, SLC6A19, SLC7A1, SLC7A5, and SLC43A2) expression showed an increasing trend in the GD group as compared to CD group (Fig. 3c).
Analysis of the protein secondary structure of myofibrillar in breast muscle
Figure 3d represented the protein secondary structure analysis. Raman spectroscopy results showed that decreased β-sheet ratio, while α-helix and β-turn ratio were increased as a result of perennial ryegrass (GD) strategy supplementation. However, random coil ratio remained unaffected between the two groups.
Pearson correlation
Figure 4a, b showed the ring heatmap correlation between two groups with edges representing correlation among breast meat quality indicators and fatty acid profile. The b* value was positively correlated with CAT (r = −0.414; P < 0.05) and bound water (r = 0.621; P < 0.01). Chewiness and drip loss showed negative correlation (r = −0.418; P < 0.05). There was a positive correlation between cooking loss, gumminess and cohesiveness (r = 0.627; P < 0.05, r = 0.519; P < 0.05, respectively), while a negative correlation was observed with immobile water and T-AOC (r = −0.674, P = 0.031 −0.450, and r = −0.450; P = 0.02). The bound water was positively associated with the MUFA, antioxidant enzymes and protein secondary structure (r = 0.512; P = 0.011, r = 0.312; P = 0.041, r = 0.499; P = 0.015), while showing a negative correlation with free and immobile water of breast muscle. These results collectively demonstrated that perennial ryegrass might improve by meat quality by improving these meat quality parameters.

a Heatmap correlation of TPA profile, antioxidant status, meat color protein secondary structure and water distribution between CD and GD group. b Heatmap correlation of fatty acid profile between CD and GD groups.The different rectangles were colored based on the Pearson correlation coefficients between meat quality assessment indicators. The intensity of color represents the degree of correlation, blue represents positive correlation, red represents negative correlation. The edges indicated the correlation between breast muscle ratios and the meat quality assessment index, blue for the positive correlation and red for the negative correlation.
Diversity and composition of gut microbiota in geese
As shown in Fig. 5a, the α-diversity indices (Shannon, Simpson, Chao1, and Ace) and observed species of intestinal flora showed no significant difference between the two groups (P > 0.05). The coverage of the samples exceeded 99.99%, which could be used for subsequent analyses. The β-diversity of the bacterial communities was analyzed by principal component analysis (PCA) and non-metric multidimensional scaling (NMD) to compare the similarity in microbial composition between the two groups (Fig. 5b). The PCA plot revealed a clear separation in the communities between the CD and GD groups, suggesting that GD intervention altered the gut microbial profile of meat geese. Beta diversity exhibited a significant difference (P < 0.05) between the CD and GD groups. PCA and NMDS analysis were presented in Fig. 5b.

a α-diversity. b β-diversity analysis. c Relative abundance of microbiota at phylum level. d Relative abundance at genus level. e Differential bacteria at phylum level. f Differential bacteria at genus level. g Metabolic pathways associated with gut microbiota. h Cladogram. i Fisher’s exact test bar plot (Genus). j Pearson correlation between gut microbiota and meat fatty acids. k Pearson correlation between gut microbiota and meat quality indicators. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01.
The relative abundance of gut microbiota was further analyzed at the phylum and genus levels (Fig. 5c, d). Firmicutes and Bacteroidetes were the dominant phyla in both groups, accounting for approximately 90% of the total abundance (Fig. 5e). The abundance of Verrucomicrobiota was affected by perennial ryegrass (GD group). The GD group has higher abundance of Verrucomicrobiota. At the genus level, GD exhibited a significant modulation in terms of microbial community (Fig. 5e). Figure 5f showed significant differences in gut microbial composition between two groups. Notably, Bacteriodales, norank clostridia_UCG-014, Prevotellaceae_Ga6A1 and Faecalibacterium, a are significantly more abundant in the GD group, as indicated by Fig. 5i. These patterns suggested that the gut microbiota differs markedly between the two groups, possibly reflecting underlying differences in health status, diet, or other environmental factors associated with CD and GD.
We sorted 6 major bacteria at the genus level related to meat quality. Faecalbacterium, Akkermansia, Blautia, Allistipes, Prevotellaceae_Ga6A1 and Butyricoccus showed high relative abundance in GD group. In contrast, Ruminococcus_torques_group and were more abundant in the CD group (Fig. 6f). The LEfSe cladogram displayed the taxa with significant differential abundance between the CD and GD groups. In this analysis, members of the phylum Fusobacteriota (blue) were significantly enriched in the GD group, suggesting their higher relative abundance in this cohort. Conversely, several taxa within the Firmicutes phylum (red) were more abundant in the CD group, including Clostridia_UCG-014, Ruminococcaceae, and Lachnospiraceae lineages, emphasizing clusters that contribute to the compositional differences between the two groups (Fig. 5h). The GD group exhibited higher relative abundances in metabolic pathways related to carbohydrate and amino acid metabolism (glycolysis/gluconeogenesis, carbon metabolism, biosynthesis of amino acids), suggesting enhanced microbial metabolic activity. In contrast, the CD group showed lower functional potential across most pathways, indicating reduced microbial functional diversity or activity. These findings implied distinct metabolic capacities of the gut microbiota between the two groups (Fig. 5g).

a Overview of SCFA profile in ileum. b Comparison of individual SCFAs content. c Correlation analysis between SCFAs and gut microbiota. d Relative expression levels of GPR43/41/109. The asterisks symbol indicates significant differences *P < 0.05, **P < 0.01.
Correlation between gut microbiota and meat quality indicators
Pearson correlation analysis between gut microbiota, fatty acids, and meat quality indicators revealed a significant relationship with breast muscle. Akkermansia, Allistipes, Blautia, Prevotellaceae_Ga6A1, and Butyricoccus were negatively correlated with C8:0, C16:0, C18:,0 and C20:0 and positively correlated with C14:1, C16:1, C17:1, C18:2n6, and C18:2n6t (Fig. 5j). Faecalbacterium was negatively correlated with meat color b* (Fig. 5k). Akkermansia was negatively correlated with TPA indicators (hardness, adhesiveness, springiness, cohesiveness, Gumminess, and chewiness) free water, and positively correlated with bound and immobile water. Alistipes showed the similar trend as Akkermensia. Blautia was positively correlated with bound and immobile water, while showing a negative correlation with free water (Fig. 5k).
Short chain fatty acids (SCFAs) profile
SCFAs improve meat quality by enhancing flavor, tenderness, and nutritional content. The contents of propionic, butyric, isobutyric, isovaleric, and caproic acids and total SCFAs were markedly elevated by ryegrass feeding (P < 0.001) as shown in Fig. 6a, b. As shown in Fig. 6c, mantel correlation analysis revealed that acetic and butyric acids were positively correlated with Butyricoccus (P < 0.05). Propionic and butyric acids were positively correlated with Prevotella and Akkermansia (P < 0.05). Propionic acid was positively correlated with Faecalibacterium (P < 0.05). Additionally, GPR43 and GPR109a expression levels in the GD group were significantly higher than those in the CD group (Fig. 6d).
AMPK/Akt-mTOR signaling mechanism and meat quality
The AMPK and Akt-mTOR signaling pathways play crucial but opposing roles in regulating cellular energy balance, muscle growth, and meat quality. AMPK is activated under conditions of energy stress, promoting fat oxidation and inhibiting anabolic processes like muscle protein synthesis to conserve energy13. Perennial ryegrass increased the mRNA expression of AMPK and its regulating gene (FOXO3) in breast muscles by decreasing expression level of Akt/mTOR and their regulating genes namely P70S6 and 4EBP1 (Fig. 7a–f). Western blot further proved this signaling mechanism as shown in Fig. 7g–k.

a–f AMPK (AMP-activated protein kinase), FOXO3 (forkhead box O3), Akt (protein kinase B), mTOR (mammalian target of rapamycin), P70S6K (ribosomal protein S6 kinase), and 4EBP,1 respectively. g–k Western blot of Akt, mTOR, AMPK, and FOXO3, respectively. l Immunofluorescence (IF) images of AMPK, Akt, and mTOR. Rabbit anti-AMPK (1:500/1:200) showing nuclear translocation of AMPK. Blue: nucleus (DAPI); Red: AMPK staining; Merge; combination of blue and red indicating nuclear translocation of AMPK, scale bar = 200 μm. Immunofluorescence (IF) analysis using Rabbit Anti-Akt (1:500/1:200) showing nuclear translocation of Akt in the breast muscle tissue of geese. Blue: nucleus (DAPI); Red: Akt staining; Merge; combination of blue and red indicating nuclear translocation of Akt, scale bar = 200 μm. Immunofluorescence (IF) analysis using Rabbit Anti-mTOR (1:500/1:200) showing nuclear translocation of mTOR in the breast muscle tissue of geese. Blue: nucleus (DAPI); Red: Akt staining; Merge; combination of blue and red indicating nuclear translocation of mTOR, scale bar = 200 μm. m Correlation between key signaling genes and meat quality indicators.
Immunofluorescence (IF) exploration of AMPK and Akt-mTOR localization in breast muscle tissue
Immunofluorescence analysis was performed to evaluate nuclear translocation and further confirm the modulation of AMPK and Akt-mTOR signaling in the leg muscle tissue. Confocal microscopy revealed stronger nuclear localization of AMPK in the GD group compared to the CD group, while Akt-mTOR showed higher nuclear expression in the CD group (Fig. 7l). These results suggested that perennial ryegrass intake may enhance meat quality by modulating the AMPK/Akt-mTOR signaling balance.
Correlation between meat quality and signaling genes
A correlation heatmap was established explaining the relationship between the key signaling pathway (APMK/Akt-mTOR) and meat quality indices. Akt-mTOR and Keap-I signaling were negatively correlated with WD (bound and immobile water), protein secondary structure (α-helix and β-turn), antioxidant enzymes activity (SOD, GSH-PX, CAT, and T-AOC), and MUFA, and positively correlated with TPA profile. AMPK and Nrf2 were positively correlated with WD (water distribution), α-helix, β-turn, random coil, antioxidant activity, and MUFA, while showing a negative correlation with TPA profile, MDA, TBAR, SFA, and n-6/n-3 (Fig. 7m).
Discussion
As consumer preference for healthier and more flavorful meat continues to grow, breast meat has gained prominence due to its nutrient content e.g., high protein and low fat14. In this study, dietary supplementation with perennial ryegrass was shown to significantly improve several key meat quality parameters in geese, including myofibrillar protein secondary structure, water distribution, textural profile, and antioxidant capacity. These enhancements collectively contributed to superior meat quality in the ryegrass-fed group. Furthermore, this study provides compelling evidence that dietary perennial ryegrass supplementation modulates gut microbiota composition and regulates host muscle metabolism via the AMPK/Akt-mTOR signaling pathway, thereby promoting a systems-level improvement in meat quality. The findings provide a holistic view of how functional forage-based diets can shape animal physiology and product quality through the gut-muscle axis, offering novel insights into sustainable production practices15,16.
Although geese fed the ryegrass-based diet (GD group) had slightly lower body weight compared to those on the commercial diet (CD group), they exhibited significantly lower abdominal fat deposition. This shift in body composition is desirable from a consumer health perspective and aligns with growing market preferences for leaner meat17. The observed trade-off suggests that dietary ryegrass may redirect metabolic energy from adipogenesis toward lean tissue development18. This hypothesis is supported by histological evidence showing increased muscle fiber density and smaller fiber cross-sectional area in the GD group, indicative of a shift toward oxidative muscle fibers19. At the molecular level, upregulation of myogenic regulatory factors such as MyoG, Myf4, and MyBPH, along with the downregulation of inhibitory genes like MSTN and MURF-1, further supports the role of ryegrass and its polyphenolic constituents in promoting muscle differentiation and growth20.
Furthermore, the expression patterns of myosin isoforms, namely heavy chain (MyHC), characterized by increased MyHC-I and MyHC-IIa and reduced MyHC-IIb expression, suggest a fiber-type shift toward oxidative metabolism in the GD group19,21. This transition is widely associated with improved meat texture profile, improved stability of meat color, and increased WHC, traits that are directly linked to consumer preference and product quality22. In line with this, the lightness (L*) value of breast meat was significantly higher in ryegrass-fed geese, indicating greater visual appeal and freshness, which are essential attributes influencing consumer acceptance23. These outcomes reinforce the role of perennial ryegrass in modulating muscle fiber composition and promoting superior meat quality through enhanced oxidative fiber development and improved visual characteristics. Nutritional composition analysis further highlighted the benefits of ryegrass supplementation. The GD group showed significantly higher CP content and lower intramuscular fat (IMF), yielding a leaner, protein-rich product with enhanced nutritional value24. Additionally, the increased concentrations of both essential and non-essential amino acids suggest an upregulation of protein synthesis and amino acid in muscle tissue. Consistently, the upregulation of amino acid transporter genes such as SLC7A1 and SLC6A19 in the GD group points to enhanced nutrient absorption and utilization capacity. These molecular adaptations are in line with improved amino acid availability in muscle, which not only supports growth but also contributes to meat flavor and quality25.
At the structural protein level, Raman spectroscopy revealed a shift in secondary structure of myofibrillar proteins, with higher β-turn and lower β-sheet content in the GD group. This structural profile is associated with increased protein flexibility and improved meat tenderness and juiciness26. The observed decreases in hardness and chewiness in the GD group are consistent with prior studies, supporting the notion that the modification enhances product tenderness and palatability27. Enhanced water retention capacity, demonstrated by LF-NMR analysis, provided further evidence of improved meat quality. The GD group exhibited a higher proportion of immobilized water and reduced cooking loss, reflecting superior water-binding capacity, likely due to increased oxidative fiber content and improved protein-water interactions.
Moreover, shear force values, an objective indicator of tenderness, were significantly lower in the GD group, reinforcing the conclusion that ryegrass supplementation contributes to improved textural quality28. Interestingly, despite the reduction in IMF, meat from the GD group still exhibited superior tenderness. This suggests that structural features such as muscle fiber composition and myofibrillar protein organization may exert a more dominant influence on tenderness than IMF in this context29. Overall, ryegrass-fed geese produced meat with improved nutritional, structural, and sensory attributes, supporting the utility of functional forage-based feeding strategies in enhancing meat quality. Serum biochemical analysis revealed that geese in the ryegrass-fed group (GD) exhibited significantly lower levels of TC, TG, and LDL indicating systemic improvements in lipid metabolism24,28. These physiological shifts were supported by hepatic gene expression patterns, which showed downregulation of key lipogenic genes (SREBP-1, FASN, PPARγ) and upregulation of genes promoting fatty acid oxidation (CPT1, PGC-1α, SIRT1)9. Together, these findings suggest that perennial ryegrass supplementation reprograms hepatic lipid metabolism toward a more catabolic and energy-efficient state, which supports lean tissue development and reduces fat deposition.
Additionally, the fatty acid composition of breast muscle was notably altered as a result of perennial ryegrass feeding (GD group), showing increased MUFAs and decreased SFAs and favorable n-6/n-3 ratio14,25. These shifts are nutritionally significant, as MUFAs are associated with cardiovascular health benefits and improved sensory quality of meat30. The upregulation of lipid metabolism-related genes such as LPL, FADS1, and FADS2 further supports enhanced fatty acid processing and deposition of healthier fat types in muscle tissue31. These findings highlight the novel potential of perennial ryegrass feeding as a natural dietary strategy to enhance meat quality by modulating fatty acid profiles and promoting the deposition of health-beneficial fats in muscle tissue.
The observed improvements in meat texture and composition were accompanied by enhanced antioxidant capacity. The GD group exhibited higher levels of serum and muscle antioxidant enzymes along with upregulated expression of Nrf2 and its downstream targets (HO-1, NQO1)24. Concurrently, lipid peroxidation markers such as malondialdehyde (MDA) and TBARS were significantly reduced32. These findings indicate reduced oxidative stress and improved oxidative stability of the meat, likely contributing to better shelf-life, color stability, and overall meat quality33. The antioxidant effects likely stem from the fiber content and bioactive polyphenolic compounds in ryegrass and their interaction with gut microbiota and its metabolites.
The 16S rRNA sequencing further elucidated the impact of ryegrass on gut microbial composition. The GD group showed a high abundance of beneficial genera such as Akkermansia, Ruminococcus, Faecalibacterium, and Lactobacillus, well-established producers of SCFAs. These microbes are associated with enhanced gut barrier function, metabolic efficiency, and nutrient absorption34. The altered β-diversity and predicted metagenomic profiles indicated not only a shift in microbial composition but also influence host metabolism and muscle development35.
Among these taxa, Bacteroides exhibited particularly strong associations with meat quality traits. Its negative correlation with TPA parameters namely gumminess, cohesiveness, and chewiness, suggests a potential role in modulating muscle structure and postmortem proteolysis, possibly through microbial-mediated metabolic or inflammatory pathways36. Additionally, Bacteroides showed inverse correlation with SFA content and a positive association with MUFA levels, consistent with its known role in lipid metabolism regulation37. Its positive correlations with protein secondary structure elements (α-helix and β-turn) and antioxidant enzyme markers (T-AOC, SOD, GSH-PX) further indicate its involvement in maintaining redox homeostasis and structural protein integrity, ultimately enhancing meat quality.
Faecalibacterium, a prominent butyrate-producing bacterium, showed a negative correlation with meat color parameter b (yellowness) and a positive correlation with protein secondary structures. These associations are consistent with its known anti-inflammatory and metabolic functions, which may enhance protein stability and reduce lipid oxidation in muscle tissues38. Similarly, Akkermansia, known for reinforcing gut barrier integrity and promoting metabolic health, was negatively correlated with cooking loss, free water content, and hardness. These associations point to its potential role in enhancing WHC and tenderness, key determinants of juiciness and palatability. Its inverse relationship with the n-6/n-3 ratio further underscores its anti-inflammatory potential and contribution to meat with improved nutritional and sensory properties39,40. Alistipes also demonstrated favorable correlations, being negatively associated with SFA and the n-6/n-3 ratio and positively correlated with MUFA, reflecting its lipid-lowering and anti-inflammatory roles41. Collectively, these findings underscore the pivotal role of gut microbiota in shaping meat quality through mechanisms involving lipid metabolism, antioxidant defense, and muscle structural remodeling. Targeted modulation of the gut microbiota via dietary strategies such as perennial ryegrass supplementation holds significant promise for enhancing both the nutritional value and sensory appeal of meat in sustainable livestock production systems.
The AMPK and Akt-mTOR signaling pathways are pivotal regulators of cellular energy homeostasis and exert opposing roles in skeletal muscle metabolism, ultimately influencing meat quality traits. AMPK serves as a cellular energy sensor activated under conditions of metabolic stress or low ATP availability, initiating catabolic processes such as fatty acid oxidation while suppressing anabolic pathways like protein synthesis42. In contrast, the Akt-mTOR axis is predominantly activated in response to sufficient nutrient and insulin availability, ultimately promoting protein synthesis and muscle hypertrophy through downstream effectors like p70S6 kinase (P70S6K) and eukaryotic translation initiation factor 4E-binding protein 1 (4EBP1)43,44. The reciprocal regulation of these pathways is essential for balancing muscle development and lipid metabolism10,11. In poultry, heightened AMPK activity has been linked to reduced intramuscular fat and improved lipid profiles, while modulation of the Akt-mTOR pathway is critical for optimal muscle growth and texture45.
Our findings demonstrate that perennial ryegrass supplementation promotes AMPK activation and concurrently suppresses Akt-mTOR signaling in geese breast muscle. This molecular shift toward catabolic dominance likely underpins the observed improvements in muscle composition, reduced fat deposition, and enhanced meat quality traits (Fig. 8). The coordinated regulation of these pathways suggests that dietary ryegrass supports a metabolic environment conducive to leaner, more nutritionally valuable meat. To our knowledge, this is the first study to elucidate a mechanistic link between dietary perennial ryegrass supplementation and improved meat quality in geese through gut microbiota modulation and activation of the AMPK/Akt-mTOR signaling axis. These findings provide novel insights into how fiber-rich forages can serve not only as nutrient sources but also as functional modulators of host metabolism via the gut-muscle axis. This research opens new avenues for developing sustainable, health-oriented feeding strategies in poultry production.

Potential mechanism of perennial ryegrass improving meat quality in meat geese via gut microbiota-mediated SCFAs/AMPK/Akt-mTOR signaling pathway.
Material and methods
Animals and experimental design
A total of 300 Wanpu geese, aged 60 days, were procured from a commercial farm and randomly allocated into two experimental groups. The control group was designated as the in-house commercial feeding group (CD), while the treatment group was designated as the pasture grazing group (GD). Each group comprised 150 geese, subdivided into six replicates of 25 birds each. Beginning at day 60, geese in the CD group were fed a standard finisher diet (as detailed in Table 4). The GD group was subjected to a pasture-based feeding regime, grazing on perennial ryegrass from 6:00 am to 6:00 p.m., and supplemented with a commercial feed once daily at 7:00 p.m. Perennial ryegrass nutritional composition was provided in Supplementary Table 1. The trial spanned a 30-day period. Experimental procedures for animal handling were conducted in accordance with the guidelines approved by the Animal Care Committee of Henan Agricultural University, Zhengzhou, China (Approval No. HNND2023072608). An overview of the experimental design was shown in Fig. 9.

An overview of experimental design from geese feeding, sampling to index determination.
Sample collections and preparations
At 90 d, selected geese were anesthetized with sodium pentobarbital (30 mg/kg BW, intravenous) prior to euthanasia and slaughtered in accordance with halal (Muslim) procedures. To calculate the FCR, average daily feed intake (ADFI) and average daily gain (ADG) were recorded. Pasture intake was estimated using a simplified method as described46. Tissues were rinsed with PBS (phosphate-buffered saline) and stored in the liquid nitrogen at −80 °C for subsequent analyses.
Analysis of breast muscle meat color
The instrumental color attributes of the meat samples were quantitatively analyzed using the CIE Lab color space, encompassing parameters for lightness (L), red-green component (a*), yellow-blue component (b*), and chroma (c*). Color measurements were performed at the geometric center of each sample using an SR-68 spectrocolorimeter (Shenzhen Threenh Technology Co., Ltd, Guangdong, China) under controlled conditions at 15 ± 2 °C. The pH of the breast meat samples was determined using a digital pH meter (model Testo 206, Tetu Instruments, Shanghai, China). Proximate analysis (DM, ash, crude fat, and CP) of breast meat was determined following the methods outlined in AOAC (1990) standards: 934.01 for ash, 990.03 for DM, and 920.39 for CP47. Shear force, an indicator of tenderness, was measured using a C-LM4 digital muscle tenderness meter (Tenovo, Beijing, China) at 200 mm/min crosshead speed. Drip loss was measured as: drip loss (%) = [(initial wt − final wt)/initial wt] × 100.
Breast meat texture profile analysis
Texture analysis was performed using a Rapid TA+ texture analyzer (Supertech Instruments Ltd., Beijing, China) fitted with a spherical probe to achieve 75% sample compression. The test parameters were set to a compression speed of 1 mm/s, a trigger force of 5 N, and a probe contact duration of 2 s.
Raman spectroscopy analysis of protein structure
Protein secondary structure of the breast meat was analyzed using Raman spectroscopy. Samples were subjected to confocal Raman micro-spectrometry using 532.8 nm laser as the excitation source. Calibration was conducted using silicon wafers (Si), with the 520.7 cm-1 peak as the reference. From 400 to 1800 cm−1, spectra were picked with a resolution of 1 cm−1, an integration time of 30 s (3 times), and maximum laser intensity (50 mW). A total of 12 random spectra were acquired per sample. Normalization of spectra was performed as previously described48. The protein secondary structure (α-helix, β-sheet, β-turn, and random coils) was measured49.
Histological examination with H&E staining
Breast meat tissues were fixed in 10% PBS, paraffin-embedded, and sectioning was performed for H&E staining. Using an optical microscope, images of the muscle cross-sections were taken. For each section, at least five non-overlapping images were captured, and muscle fiber cross-sectional area, denoted by CSA was measured (except for the fibers with blurred outlines that could not be recognized by the software). The average CSA across the five images was calculated. Fiber density (fibers/mm²) was computed using the formula: Fiber density (fibers/mm²) = Area of the field /No. of fibers in field.
LF-NMR transverse relaxation for water distribution
Breast meat samples were prepared by removing surface fat and connective tissue, then cutting them into uniform pieces approximately 1 cm3 in size. The transverse relaxation time (T₂) of water in the samples was measured using a low-field nuclear magnetic resonance (LF-NMR) analyzer operating at 20 MHz. The measurements were performed using the Carr–Purcell–Meiboom–Gill pulse sequence, with the probe temperature maintained at 32 ± 1 °C. The key acquisition parameters were set as follows: repetition time (TR) of 2000 ms, echo time (TE) of 200 µs, and 8000 echoes, with 4–8 scans performed per sample to enhance signal accuracy. Each sample was placed in a 10 mm NMR tube and inserted into the instrument for measurement. The obtained T₂ relaxation data were processed using inverse Laplace transformation to generate T₂ relaxation distributions. MultiExp Inv Analysis program version 4.08 was used to process data from the LF-NMR analysis (Niumag Co., Ltd, Shanghai, China).
Determination of amino acids
Amino acid profile of the goose breast meat was determined according to the method used by Okruszek et al.50.
Lipid profile
TC, TG, HDL, LDL, GLU, and BUN were measured using commercial ELISA kits (Crescent Diagnostic Lab) according to the manufacturer’s instructions. Measurements were performed using a spectrophotometric method.
Fatty acid analysis
The fatty acid composition of breast meat was analyzed by first extracting total lipids from freeze-dried samples using the chloroform-methanol (1:1, v/v) method. Fatty acid methyl esters (FAMEs) were subsequently synthesized using methanolic KOH, in accordance with procedures51. Separation and quantification of FAMEs were conducted using an Agilent 7890B gas chromatograph (GC) equipped with a flame ionization detector (FID) (Agilent Technologies Inc., Santa Clara, CA, USA), following the analytical method52. Fatty acids were identified by comparing their retention times with those of certified reference standards (Sigma-Aldrich, St. Louis, MO, USA) and expressed as percentages of the total identified fatty acids.
Determination of antioxidant enzymes
Ileum and breast muscle samples (n = 5 per group) were used to evaluate the activity of key antioxidant enzymes. Superoxide dismutase (SOD), glutathione peroxidase (GSH-PX), catalase (CAT), total antioxidant capacity (T-AOC), malondialdehyde (MDA), and thiobarbituric acid reactive substances (TBARS) were measured using ELISA kits according to the company guidelines.
Quantification of short chain fatty acids (SCFAs)
Approximately 0.5 g of ileal content was homogenized with 2 mL of ultrapure water and allowed to settle for 30 min. The mixture was then centrifuged at 3000 × g for 15 min. One milliliter of the resulting supernatant was combined with 0.2 mL of ice-cold 25% (w/v) metaphosphoric acid and incubated at 4 °C for 30 min to precipitate proteins and stabilize the SCFAs. The samples were subsequently centrifuged at 11,000 × g for 10 min. The SCFA including acetate, propionate and butyrate were quantified using gas chromatography (GC) with a Varian CP-3800 system (Varian Inc., USA)53.
Real-time quantitative PCR
Total RNA was isolated from pulverized liver and leg tissues using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA), in accordance with the manufacturer's protocol. Complementary DNA (cDNA) was synthesized via reverse transcription, followed by quantitative real-time PCR (qRT-PCR) conducted on an ABI 7900HT Fast Real-Time PCR System (Applied Biosystems, CA, USA). Gene-specific primers were designed using Primer3 software (refer to Supplementary Table 2). Relative gene expression levels were calculated using the comparative Ct method, normalized to the expression of housekeeping genes GAPDH54.
Western blotting
Tissues from the breast muscles of experimental groups were homogenized in RIPA lysis buffer (#CW2333S, CWBIO, Jiangsu, China). The homogenates were then centrifuged at 12,000 × g for 5 min to extract total protein. Protein concentrations were determined using a BCA protein assay kit, following the manufacturer’s instructions. Equal amounts of protein (40 μg per sample) were separated on 4–20% (w/v) SDS-PAGE gels and transferred onto PVDF membranes. The membranes were incubated overnight at 4 °C with primary antibodies targeting Akt, mTOR, AMPK, FOXO3 and GAPDH. After washing with TBST for 30 min, membranes were incubated with the appropriate secondary antibodies for 2 h at room temperature. Following another 30-min TBST wash, protein bands were visualized using an AI600 imaging system (CE, USA). All blots originated from the same experiment and were processed under identical conditions (Source Data). Band intensities were quantified using ImageJ software (v1.8.0, NIH).
AMPK and Akt-mTOR immunofluorescence (IF) imaging
For immunofluorescence analysis, tissue sections were fixed in 4% paraformaldehyde for 10 min. Paraffin-embedded samples were dewaxed in xylene for 15 min, followed by sequential rehydration through a graded ethanol series and rinsing with distilled water. Antigen retrieval was performed by boiling the rehydrated leg muscle sections in citrate-EDTA buffer for 2 min. Non-specific binding was blocked by incubating the sections in PBS supplemented with 10% fetal bovine serum (FBS) and 0.3% Triton X-100 for 30 min at room temperature. Primary antibodies were applied and incubated overnight at 4 °C to detect target proteins. Following washing steps, the sections were treated with fluorophore-conjugated secondary antibodies, and fluorescence images were acquired using a fluorescence microscope.
16S rRNA gene sequencing
DNA was extracted from ileal content utilizing the PSP Spin Stool DNA plus Kit. Before amplifying the 16sRNA gene, DNA quality and fragment size were evaluated, specifically targeting the V3-V4 hypervariable regions with the help of primer pair F515/R806 in a polymerase chain reaction (PCR). Amplicon sequencing was done using Illumina PE250 platform (BGI, Shenzhen, China). Raw sequence data underwent quality control procedures, including trimming and filtering using the DADA2 pipeline in R (version 4.3.1), enabling the identification of ASVs. Paired-end reads were merged, and chimeric sequences were filtered out using the DADA2 pipeline with either default or optimized parameter settings. Using a SILVA reference database, ASV Taxonomic classification was carried out (version 138.1). ASVs with relative abundances below 0.01% were removed using the Microbiota Process package (version 3.17) in R to minimize noise from low-abundance taxa. Alpha diversity metrics, such ACE, Chao, and Shannon indexes were calculated with Mothur (version 1.30.1). For the analysis of Beta diversity, PCA was performed in R using the Vegan package (version 2.5-3) based on Bray-Curtis dissimilarity. Genus-level taxonomic distributions were visualized via a bar plot. Fisher’s exact test bar plot analysis was performed to identify significantly different taxa using a linear discriminant analysis (LDA), showing a cladogram.
Statistical analysis
All data were analyzed through one-way analysis of variance (ANOVA) using IBM SPSS Statistics for Windows (version 20.0). Data were presented as mean ± standard deviation (SD). A statistical power exceeding 0.80 was maintained, assuming a minimum effect size of 1.0. The Shapiro–Wilk test and Levene’s test were employed to assess the assumptions of normality and homogeneity of variance, respectively. Unpaired T-test was employed determining differences among groups, based on the following statistical model:
Pearson correlation was used to identify the relationship between parameters. Differences were identified at P < 0.05 and P < 0.01, respectively.
Data availability
All data supporting the findings of this study are available within the manuscript and its Supplementary Information files.
References
Dumlu, B. The global goose meat production quantity forecast for the 2023–2027 years. Selcuk J. Agr. Food Sci. 38, 326–341 (2024).
Cui, L. L. et al. Analysis of meat flavor compounds in pedigree and two-strain Yangzhou geese. Poult. Sci. 94, 2266–2271 (2015).
Song, Y. et al. Effects of forage feeding versus grain feeding on the growth performance and meat quality of Yangzhou geese. Br. Poult. Sci. 58, 397–401 (2017).
Liu, H. W. & Zhou, D. W. Influence of pasture intake on meat quality, lipid oxidation, and fatty acid composition of geese. J. Anim. Sci. 91, 764–771 (2013).
Thornton, P. K. Livestock production: recent trends, future prospects. Philos. Trans. R. Soc. B 365, 2853–2867 (2010).
Dalle Zotte, A. & Szendrő, Z. The role of rabbit meat as functional food. Meat Sci. 88, 319–331 (2011).
Kagan, I. A. Soluble phenolic compounds of perennial ryegrass (Lolium perenne L.): potential effects on animal performance, and challenges in determining profiles and concentrations. Anim. Feed Sci. Technol. 277, 114960 (2021).
Ali, Q. et al. Pasture intake protects against commercial diet-induced lipopolysaccharide production facilitated by gut microbiota through activating intestinal alkaline phosphatase enzyme in meat geese. Front. Immunol. 13, 1041070 (2022).
Den Besten, G. et al. Short-chain fatty acids protect against high-fat diet–induced obesity via a PPARγ-dependent switch from lipogenesis to fat oxidation. Diabetes 64, 2398–2408 (2015).
He, J. et al. Short-chain fatty acids and their association with signalling pathways in inflammation, glucose and lipid metabolism. Int. J. Mol. Sci. 21, 6356 (2020).
Sartori, R., Romanello, V. & Sandri, M. Mechanisms of muscle atrophy and hypertrophy: implications in health and disease. Nat. Commun. 12, 330 (2021).
Underwood, K. R. et al. AMP-activated protein kinase is negatively associated with intramuscular fat content in longissimus dorsi muscle of beef cattle. Meat Sci. 79, 394–402 (2008).
Thomson, D. M. The role of AMPK in the regulation of skeletal muscle size, hypertrophy, and regeneration. Int. J. Mol. Sci. 19, 3125 (2018).
Pillai, K., Sambandam, R., Nagarajan, M. & Harishchandra, R. Nutritional interventions to improve breast meat yield in broilers - review. Int. J. Livest. Res. 9, 49–61 (2019).
Kang, L. et al. Effects of feeding patterns on production performance, lipo-nutritional quality and gut microbiota of Sunit sheep. Meat Sci. 218, 109642 (2024).
Liu, B. et al. Gut microbiota modulate rabbit meat quality in response to dietary fiber. Front. Nutr. 9, 849429 (2022).
Van Elswyk, M. E. & McNeill, S. H. Impact of grass/forage feeding versus grain finishing on beef nutrients and sensory quality: the U.S. experience. Meat Sci. 96, 535–540 (2014).
Islam, A., Civitarese, A. E., Hesslink, R. L. & Gallaher, D. D. Viscous dietary fiber reduces adiposity and plasma leptin and increases muscle expression of fat oxidation genes in rats. Obesity 20, 349–355 (2012).
Gagaoua, M., Monteils, V., Couvreur, S. & Picard, B. Identification of biomarkers associated with the rearing practices, carcass characteristics, and beef quality: an integrative approach. J. Agric. Food Chem. 65, 8264–8278 (2017).
Nagai, A. et al. Quercetin induces a slow myofiber phenotype in engineered human skeletal muscle tissues. FASEB J. 38, e70009 (2024).
Yu, M. et al. Different dietary starch sources alter the carcass traits, meat quality, and the profile of muscle amino acid and fatty acid in finishing pigs. J. Anim. Sci. Biotechnol. 11, 78 (2020).
Weng, K. et al. Fiber characteristics and meat quality of different muscular tissues from slow- and fast-growing broilers. Poult. Sci. 101, 101537 (2022).
Yüksel, S. et al. Effects of different finishing systems on carcass traits, fatty acid composition, and beef quality characteristics of young Eastern Anatolian Red bulls. Trop. Anim. Health Prod. 44, 1521–1528 (2012).
Ali, Q. et al. Integrated transcriptome and metabolome profiles reveal the regulatory molecular mechanisms of meat quality in pasture-fed goose. Food Biosci. 61, 104846 (2024).
Guo, B. et al. Research note: effect of diet with different proportions of ryegrass on breast meat quality of broiler geese. Poult. Sci. 99, 2500–2507 (2020).
Fowler, S. M., Schmidt, H., Van De Ven, R. & Hopkins, D. L. Preliminary investigation of the use of Raman spectroscopy to predict meat and eating quality traits of beef loins. Meat Sci. 138, 53–58 (2018).
Fruet, A. P. B. et al. Effects of feeding legume-grass pasture and different concentrate levels on fatty acid profile, volatile compounds, and off-flavor of the M. longissimus thoracis. Meat Sci. 140, 112–118 (2018).
Zhang, Y. et al. Comparison of growth performance, meat quality, and blood biochemical indexes of Yangzhou goose under different feeding patterns. Poult. Sci. 103, 103349 (2024).
Huo, W. et al. Effect of muscle fiber characteristics on meat quality in fast- and slow-growing ducks. Poult. Sci. 100, 101264 (2021).
Valentini, J. et al. Chemical composition, lipid peroxidation, and fatty acid profile in meat of broilers fed with glycerol monolaurate additive. Food Chem. 330, 127187 (2020).
Jiao, A. et al. Infusion of short chain fatty acids in the ileum improves the carcass traits, meat quality and lipid metabolism of growing pigs. Anim. Nutr. 7, 94–100 (2021).
Funaro, A. et al. Comparison of meat quality characteristics and oxidative stability between conventional and free-range chickens. Poult. Sci. 93, 1511–1522 (2014).
Insani, E. M. et al. Oxidative stability and its relationship with natural antioxidants during refrigerated retail display of beef produced in Argentina. Meat Sci. 79, 444–452 (2008).
Zulfiqar, Z. et al. Zinc glycine supplementation improves bone quality in meat geese by modulating gut microbiota, SCFAs, and gut barrier function through Wnt10b/NF-κB axis. Poult. Sci. 104, 104925 (2025).
Tremaroli, V. & Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 489, 242–249 (2012).
Bischof, G. et al. Metabolic, proteomic and microbial changes postmortem and during beef aging. Crit. Rev. Food Sci. Nutr. 64, 1076–1109 (2024).
Brown, E. M., Clardy, J. & Xavier, R. J. Gut microbiome lipid metabolism and its impact on host physiology. Cell Host Microbe 31, 173–186 (2023).
Balasubramanian, B. & Liu, W.-C. Editorial: gut microbiota: allied with livestock nutrition, health, and welfare. Front. Vet. Sci. 11, 1413671 (2024).
Simopoulos, A. P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 233, 674–688 (2008).
Zhao, Y. et al. Akkermansia muciniphila: a promising probiotic against inflammation and metabolic disorders. Virulence 15, 2375555 (2024).
Parker, B. J., Wearsch, P. A., Veloo, A. C. M. & Rodriguez-Palacios, A. The genus Alistipes: gut bacteria with emerging implications to inflammation, cancer, and mental health. Front. Immunol. 11, 906 (2020).
Hardie, D. G., Ross, F. A. & Hawley, S. A. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 13, 251–262 (2012).
Ogasawara, R. et al. The role of mTOR signalling in the regulation of skeletal muscle mass in a rodent model of resistance exercise. Sci. Rep. 6, 31142 (2016).
Saxton, R. A. & Sabatini, D. M. mTOR signaling in growth, metabolism, and disease. Cell 169, 361–371 (2017).
Du, M., Wang, B., Fu, X., Yang, Q. & Zhu, M.-J. Fetal programming in meat production. Meat Sci. 109, 40–47 (2015).
Cartoni Mancinelli, A. et al. Rearing Romagnola geese in vineyard: pasture and antioxidant intake, performance, carcass and meat quality. Ital. J. Anim. Sci. 18, 372–380 (2019).
A.O.A.C. Official Methods of Analysis 15th edn (Association of Official Analytical Chemist, Washington DC, 1990).
Zhao, G. et al. The effect of shower time, electrolyte treatment, and electrical stimulation on meat quality of cattle longissimus thoracis muscle in cold weather. Meat Sci. 184, 108664 (2022).
Susi, H. & Byler, D. M. Fourier deconvolution of the amide I Raman band of proteins as related to conformation. Appl. Spectrosc. 42, 819–826 (1988).
Okruszek, A., Wołoszyn, J., Haraf, G., Orkusz, A. & Wereńska, M. Chemical composition and amino acid profiles of goose muscles from native Polish breeds. Poult. Sci. 92, 1127–1133 (2013).
Yu, M. et al. Use of Hermetia illucens larvae as a dietary protein source: effects on growth performance, carcass traits, and meat quality in finishing pigs. Meat Sci. 158, 107837 (2019).
Li, F. et al. Effects of dietary n-6:n-3 PUFA ratio on fatty acid composition, free amino acid profile and gene expression of transporters in finishing pigs. Br. J. Nutr. 113, 739–748 (2015).
Qin, S. et al. Dietary administration of resistant starch improved caecal barrier function by enhancing intestinal morphology and modulating microbiota composition in meat duck. Br. J. Nutr. 123, 172–181 (2020).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
Acknowledgements
This study was financially supported by the China Agriculture Research System of MOF and MARA (No. CARS-34) and the Science and Technology Innovation Leading Talent in Central Plains (No. 244200510010).
Author information
Authors and Affiliations
Contributions
Zeshan Zulfiqar performed the experiment and wrote the original draft. Xiaoyan Zhu, Zhichang Wang, and Hao Sun did formal analysis. Boshuai Liu supervised the experiment. Yinghua Shi and Yalei Cui conceptualized and structured the manuscript. Muhammad Arslan Asif and Han Yao supported in lab analysis. Shucheng Huang and Khawla Alharbi revised the manuscript. All the authors have read and approved the final version of the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zulfiqar, Z., Asif, M.A., Alharbi, K. et al. Dietary perennial ryegrass improves meat quality: insights into the gut-muscle axis in geese. npj Sci Food 9, 258 (2025). https://doi.org/10.1038/s41538-025-00642-1
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41538-025-00642-1
