
Abstract
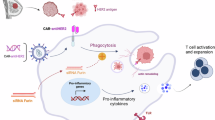
Chimeric antigen receptor (CAR) T cell therapies have successfully treated hematological malignancies. Macrophages have also gained attention as an immunotherapy owing to their immunomodulatory capacity and ability to infiltrate solid tumors and phagocytize tumor cells. The first-generation CD3ζ-based CAR-macrophages could phagocytose tumor cells in an antigen-dependent manner. Here we engineered induced pluripotent stem cell-derived macrophages (iMACs) with toll-like receptor 4 intracellular toll/IL-1R (TIR) domain-containing CARs resulting in a markedly enhanced antitumor effect over first-generation CAR-macrophages. Moreover, the design of a tandem CD3ζ-TIR dual signaling CAR endows iMACs with both target engulfment capacity and antigen-dependent M1 polarization and M2 resistance in a nuclear factor kappa B (NF-κB)-dependent manner, as well as the capacity to modulate the tumor microenvironment. We also outline a mechanism of tumor cell elimination by CAR-induced efferocytosis against tumor cell apoptotic bodies. Taken together, we provide a second-generation CAR-iMAC with an ability for orthogonal phagocytosis and polarization and superior antitumor functions in treating solid tumors relative to first-generation CAR-macrophages.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others
Data availability
scRNA-seq generated in this study has been deposited in the National Center for Biotechnology Information GEO database under the accession code GSE214140. Previously published single-cell and bulk RNA-Seq data that were re-analyzed here are available under accession codes E-MTAB-7407 and GSE55536. Source data are provided with this paper.
Code availability
The codes used for the analysis reported in this study are freely available at https://github.com/huayu1111/carmProj.
Change history
18 December 2023
A Correction to this paper has been published: https://doi.org/10.1038/s41590-023-01734-4
References
Gomes-Silva, D. et al. CD7-edited T cells expressing a CD7-specific CAR for the therapy of T-cell malignancies. Blood 130, 285–296 (2017).
Chuntova, P., Downey, K. M., Hegde, B., Almeida, N. D. & Okada, H. Genetically engineered T-cells for malignant glioma: overcoming the barriers to effective immunotherapy. Front. Immunol. 9, 3062 (2018).
Ma, L. et al. Enhanced CAR-T cell activity against solid tumors by vaccine boosting through the chimeric receptor. Science 365, 162–168 (2019).
Lynn, R. C. et al. c-Jun overexpression in CAR T cells induces exhaustion resistance. Nature 576, 293–300 (2019).
Adachi, K. et al. IL-7 and CCL19 expression in CAR-T cells improves immune cell infiltration and CAR-T cell survival in the tumor. Nat. Biotechnol. 36, 346–351 (2018).
Upadhyay, R. et al. A critical role for Fas-mediated off-target tumor killing in T-cell immunotherapy. Cancer Discov. 11, 599–613 (2021).
Hamieh, M. et al. CAR T cell trogocytosis and cooperative killing regulate tumour antigen escape. Nature 568, 112–116 (2019).
Martinez, M. & Moon, E. K. CAR T cells for solid tumors: new strategies for finding, infiltrating, and surviving in the tumor microenvironment. Front. Immunol. 10, 128 (2019).
Bonnardel, J. & Guilliams, M. Developmental control of macrophage function. Curr. Opin. Immunol. 50, 64–74 (2018).
Cassetta, L. & Pollard, J. W. Targeting macrophages: therapeutic approaches in cancer. Nat. Rev. Drug Discov. 17, 887–904 (2018).
Cortez-Retamozo, V. et al. Origins of tumor-associated macrophages and neutrophils. Proc. Natl Acad. Sci. USA 109, 2491–2496 (2012).
Qian, B. Z. & Pollard, J. W. Macrophage diversity enhances tumor progression and metastasis. Cell 141, 39–51 (2010).
Gambardella, V. et al. The role of tumor-associated macrophages in gastric cancer development and their potential as a therapeutic target. Cancer Treat. Rev. 86, 102015 (2020).
Chen, Y., Zhang, S., Wang, Q. & Zhang, X. Tumor-recruited M2 macrophages promote gastric and breast cancer metastasis via M2 macrophage-secreted CHI3L1 protein. J. Hematol. Oncol. 10, 36 (2017).
DeNardo, D. G. & Ruffell, B. Macrophages as regulators of tumour immunity and immunotherapy. Nat. Rev. Immunol. 19, 369–382 (2019).
Vitale, I., Manic, G., Coussens, L. M., Kroemer, G. & Galluzzi, L. Macrophages and metabolism in the tumor microenvironment. Cell Metab. 30, 36–50 (2019).
Song, M. et al. Tumor derived UBR5 promotes ovarian cancer growth and metastasis through inducing immunosuppressive macrophages. Nat. Commun. 11, 6298 (2020).
Cioni, B. et al. Androgen receptor signalling in macrophages promotes TREM-1-mediated prostate cancer cell line migration and invasion. Nat. Commun. 11, 4498 (2020).
Huang, R. et al. CCL5 derived from tumor-associated macrophages promotes prostate cancer stem cells and metastasis via activating β-catenin/STAT3 signaling. Cell Death Dis. 11, 234 (2020).
Salvagno, C. et al. Therapeutic targeting of macrophages enhances chemotherapy efficacy by unleashing type I interferon response. Nat. Cell Biol. 21, 511–521 (2019).
Wiehagen, K. R. et al. Combination of CD40 agonism and CSF-1R blockade reconditions tumor-associated macrophages and drives potent antitumor immunity. Cancer Immunol. Res. 5, 1109–1121 (2017).
Kang, M. et al. Nanocomplex-mediated in vivo programming to chimeric antigen receptor-M1 macrophages for cancer therapy. Adv. Mater. 33, e2103258 (2021).
Ries, C. H. et al. Targeting tumor-associated macrophages with anti-CSF-1R antibody reveals a strategy for cancer therapy. Cancer Cell 25, 846–859 (2014).
Akkari, L. et al. Dynamic changes in glioma macrophage populations after radiotherapy reveal CSF-1R inhibition as a strategy to overcome resistance. Sci.Transl. Med. 12, eaaw7843 (2020).
Zhang, L. et al. Pluripotent stem cell-derived CAR-macrophage cells with antigen-dependent anti-cancer cell functions. J. Hematol. Oncol. 13, 153 (2020).
Klichinsky, M. et al. Human chimeric antigen receptor macrophages for cancer immunotherapy. Nat. Biotechnol. 38, 947–953 (2020).
Morrissey, M. A. et al. Chimeric antigen receptors that trigger phagocytosis. eLife 7, e36688 (2018).
Takata, K. et al. Induced-pluripotent-stem-cell-derived primitive macrophages provide a platform for modeling tissue-resident macrophage differentiation and function. Immunity 47, 183–198 (2017).
Zhang, H. & Reilly, M. P. Human induced pluripotent stem cell-derived macrophages for unraveling human macrophage biology. Arter. Thromb. Vasc. Biol. 37, 2000–2006 (2017).
Lachmann, N. et al. Large-scale hematopoietic differentiation of human induced pluripotent stem cells provides granulocytes or macrophages for cell replacement therapies. Stem Cell Rep. 4, 282–296 (2015).
Kawai, T. & Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 34, 637–650 (2011).
Lancaster, G. I. et al. Evidence that TLR4 is not a receptor for saturated fatty acids but mediates lipid-induced inflammation by reprogramming macrophage metabolism. Cell Metab. 27, 1096–1110 (2018).
Fan, W. et al. FoxO1 regulates Tlr4 inflammatory pathway signalling in macrophages. EMBO J. 29, 4223–4236 (2010).
Fitzgerald, K. A. & Kagan, J. C. Toll-like receptors and the control of immunity. Cell 180, 1044–1066 (2020).
Kawai, T. & Akira, S. The role of pattern-recognition receptors in innate immunity: update on toll-like receptors. Nat. Immunol. 11, 373–384 (2010).
Kondo, T., Kawai, T. & Akira, S. Dissecting negative regulation of toll-like receptor signaling. Trends Immunol. 33, 449–458 (2012).
Schappe, M. S. et al. Chanzyme TRPM7 mediates the Ca2+ influx essential for lipopolysaccharide-induced toll-like receptor 4 endocytosis and macrophage activation. Immunity 48, 59–74 (2018).
Kagan, J. C. et al. TRAM couples endocytosis of toll-like receptor 4 to the induction of interferon-β. Nat. Immunol. 9, 361–368 (2008).
Yamamoto, M. et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 301, 640–643 (2003).
Wermuth, P. J. & Jimenez, S. A. Gadolinium compounds signaling through TLR4 and TLR7 in normal human macrophages: establishment of a proinflammatory phenotype and implications for the pathogenesis of nephrogenic systemic fibrosis. J. Immunol. 189, 318–327 (2012).
Scharping, N. E. et al. Mitochondrial stress induced by continuous stimulation under hypoxia rapidly drives T cell exhaustion. Nat. Immunol. 22, 205–215 (2021).
Park, J. E. et al. Hypoxia-induced tumor exosomes promote M2-like macrophage polarization of infiltrating myeloid cells and microRNA-mediated metabolic shift. Oncogene 38, 5158–5173 (2019).
Liu, Y. N. et al. Hypoxia induces mitochondrial defect that promotes T cell exhaustion in tumor microenvironment through MYC-regulated pathways. Front. Immunol. 11, 1906 (2020).
Xu, J. et al. Hypoxic glioma-derived exosomes promote M2-like macrophage polarization by enhancing autophagy induction. Cell Death Dis. 12, 373 (2021).
Moynagh, P. N. TLR signalling and activation of IRFs: revisiting old friends from the NF-κB pathway. Trends Immunol. 26, 469–476 (2005).
Zhang, H. et al. Functional analysis and transcriptomic profiling of iPSC-derived macrophages and their application in modeling Mendelian disease. Circ. Res. 117, 17–28 (2015).
Mehrotra, P. & Ravichandran, K. S. Drugging the efferocytosis process: concepts and opportunities. Nat. Rev. Drug Discov. 21, 601–620 (2022).
Franklin, R. A. et al. The cellular and molecular origin of tumor-associated macrophages. Science 344, 921–925 (2014).
Chen, Y. et al. CAR-macrophage: a new immunotherapy candidate against solid tumors. Biomed. Pharmacother. 139, 111605 (2021).
Sadelain, M., Brentjens, R. & Rivière, I. The promise and potential pitfalls of chimeric antigen receptors. Curr. Opin. Immunol. 21, 215–223 (2009).
Huang, X. et al. Fast, long-term, super-resolution imaging with Hessian structured illumination microscopy. Nat. Biotechnol. 36, 451–459 (2018).
Zhao, W. et al. Sparse deconvolution improves the resolution of live-cell super-resolution fluorescence microscopy. Nat. Biotechnol. 40, 606–617 (2022).
Douvaras, P. et al. Directed differentiation of human pluripotent stem cells to microglia. Stem Cell Rep. 8, 1516–1524 (2017).
Popescu, D. M. et al. Decoding human fetal liver haematopoiesis. Nature 574, 365–371 (2019).
Liu, B. et al. An entropy-based metric for assessing the purity of single cell populations. Nat. Commun. 11, 3155 (2020).
Acknowledgements
We acknowledge Dan Kaufman (UCSD), Huang Zhu (UCSD) and Jin Zhang lab members for discussion of the manuscript. We also thank Chong Liu (Zhejiang University) for providing the U87MG cell line and valuable suggestions and Xuanming Yang (Shanghai Jiao Tong University) for providing the reagents such as the JurkatTCRαβ-KO cell line. This work was funded by the National Natural Science Foundation of China (82373238, 31871453 and 91857116 to J.Z.), the National Key Research and Development Program of China (2018YFA0107103 and 2018YFC1005002 to J.Z.), Zhejiang Innovation Team (grant 2019R01004 to J.Z.), the Zhejiang Natural Science Foundation (LR19C120001 to J.Z.), the Key Research and Development Program of Zhejiang Province (2023C03036) and research fund from CellOrigin and Qilu Pharmaceutical, the Zhejiang Natural Science Foundation of China (LQ22H160001 to A.L.) and the China Postdoctoral Science Foundation (2019M662035 to A.L.). We thank Guangzhou CSR Biotech for live-cell imaging by using their commercial super-resolution microscope (HIS-SIM), data acquisition, SR image reconstruction, analysis and discussion.
Author information
Authors and Affiliations
Contributions
J.Z. and A.L. designed the study. A.L. S.L., H.L., X.D., T.T., X.W., H.Z., S.S., M.Z., L.T., D. X., W.Z., S.Z., Y.C., W.X., L.Z., Y.Z. and J.Z. performed the experiments. H.Y. contributed to the NGS data analysis. W.J. contributed to the culture of U87MG cells. G.C., F.C. and Z.G. provided important discussions. J.Z. and A.L. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
J.Z. is a scientific cofounder of CellOrigin. The other authors declare no competing interests.
Peer review
Peer review information
Nature Immunology thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: N. Bernard, in collaboration with the Nature Immunology team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Examining phagocytosis of CAR-iMACs against the antigen-expressing cancer cells.
a, Diagrammatic drawing for an EGFRvIII-expressing lentivirus vector. b, Western blotting showing the expression of EGFRvIII in wild-type U87MG (WT-U87MG) cells and lentivirus-infected U87MG cells (U87MGEGFRvIII). c, Western blotting showing the expression of the total EGFR in wild-type U87MG cells and lentivirus-infected U87MG cells. d, Immunofluorescence showing expression and localization of EGFRvIII in U87MGEGFRvIII cells. EGFRvIII exhibited prominently high expression and evident localization on the membrane of cells (indicated by the white arrows) of U87MGEGFRvIII. e, Co-incubating experiment showing that EGFP-marked EGFRvIII-targeting CAR-iMACs (truncated, CD3ζ- and TIR-CAR-iMACs) adhered to the tdTomato-expressed U87MGEGFRvIII cells more closely than EGFP-marked WT-iMACs after co-culturing for 4 hours. f, Both CD3ζ-CAR-iMACs and TIR-CAR-iMACs showed enhanced cancer cell phagocytosis activity against U87MGEGFRvIII cells compared to truncated CAR-iMACs and WT-iMACs after co-culturing for 12 hours.
Extended Data Fig. 2 CD3ζ-TIR-CAR-iMACs exhibited enhanced and persistent antitumor activity.
a, An overview of the antitumor assay of EGFRvIII-targeting CAR-iMACs in vivo. Four hours after injection of FFluc+U87MGEGFRvIII cells into NOD-SCID mice, EGFRvIII-targeting truncated CAR-iMACs, CD3ζ-CAR-iMACs, TIR-CAR-iMACs, and CD3ζ-TIR-CAR-iMACs without IFN-γ/LPS-polarization pretreatment were injected into the tumor-bearing mice. Two hours after the CAR-iMACs injection, animal imagings were conducted to detect the signal of BLI from tumor cells. On the following day 1, day 3, and day 7, bioluminescent imaging was performed at the same time during the days. b, Visual presentation of the tumor signal from the tumor-bearing mice after treatment by the four types of CAR-iMACs. c, Statistics analysis of tumor BLI signal from b (n = 5 mice from truncated CAR-iMAC group, n = 5 mice from CD3ζ-CAR-iMAC group, n = 10 mice from TIR-CAR-iMAC group, n = 10 mice from CD3ζ-TIR-CAR-iMAC group). Significance was calculated with a two-way ANOVA analysis and are presented as mean±s.e.m. p-value: ns, not significant, *P < 0.05, **P < 0.01. d, The Kaplan-Meier survival curve of CAR-iMACs treated mice over 29 days from b. Significance was calculated with two-tailed log-rank Mantel-Cox test (CD3ζ-CAR-iMAC group versus truncated CAR-iMAC group, P = 0.0999. TIR-CAR-iMAC group versus truncated CAR-iMAC group, P = 0.0003. CD3ζ-TIR-CAR-iMAC group versus truncated CAR-iMAC group, P = 0.0003) e, FCM analysis of CD80-positive population and CD163-positive population of EGFRvIII-targeting CD3ζ-CAR-iMACs, TIR-CAR-iMACs, and CD3ζ-TIR-CAR-iMACs isolated from the mice treated in the same way as in b for 48 hours. The FCM experiments were carried out in triplicates and the results were processed using FlowJo. f, Histograph analysis of e (n = 3 biologically independent samples per group). The significance was calculated with a two-way ANOVA analysis and are presented as mean±s.e.m. p-value: ns, not significant, *P < 0.05, **P < 0.01. All the statistical data above was shown by Graphpad Prism 8.2.1.
Extended Data Fig. 3 The CD3ζ-TIR-CAR-iMACs showed potent antitumor activity against intracranial GBM.
a, An overview of the anti-GBM assay of EGFRvIII-targeting CAR-iMACs in vivo. 1 × 104 FFluc+U87MGEGFRvIII cells were injected into the intracranial corpus striatum of NOD-SCID mice six days before immune cell therapy. On day -1, live animal imaging was conducted to monitor the formation of GBM. On day 0, EGFRvIII-targeting truncated CAR-iMACs and CD3ζ-TIR-CAR-iMACs with no IFN-γ/LPS-pretreatment were injected into the GBM lesion. On the following day 1, day 2, day 3, day 5, and day 10, the bioluminescent imaging was performed at the same time point to record the change of signal from the GBM. b, Visual presentation of the tumor signal from the intracranial GBM after treatment by the truncated CAR-iMACs and CD3ζ-TIR-CAR-iMACs. The PBS treatment group was designed as a negative control. c, Statistics analysis of tumor BLI signal from b (n = 5 mice from PBS group, n = 5 from truncated CAR-iMAC group, n = 5 mice from CD3ζ-TIR-CAR-iMAC group). The significance was calculated with the two-way ANOVA analysis and are presented as mean±s.e.m. p-value: ns, not significant, ***P < 0.001. d, The Kaplan-Meier survival curve of CAR-iMACs treated mice over 17 days from b. Significance was calculated with two-tailed log-rank Mantel-Cox test (CD3ζ-TIR-CAR-iMAC group versus PBS group, P = 0.0003. CD3ζ-TIR-CAR-iMAC group versus truncated CAR-iMAC group, P = 0.0003) All the statistic data above was shown by Graphpad Prism 8.2.1. e, The IHC staining showing the EGFRvIII protein expression of FFluc+U87MGEGFRvIII-derived GBM on day 8 and day 16 after the tumor cell inoculation, indicating the persistent expression of EGFRvIII antigen.
Extended Data Fig. 4 GPC3-targeting CAR-iMACs showed enhanced antitumor capacity against HCC.
a, The flow cytometry analysis was performed to determine the expression of GPC3-CARs on truncated CAR-iMACs, TIR-CAR-iMAC, and CD3ζ-TIR-CAR-iMACs by an APC-conjugated rabbit antibody specific to F(ab’)2 of human IgG. Shown is a representative result from three biological replicates. The results were processed using FlowJo. The numbers are mean±s.e.m. b, Real-time imaging by an Incucyte2022 A Live-Cell Analysis Instrument showed the signals of the tdTomato+ HepG2 cells (white arrow) during a time course of 138 hours after incubating GPC3-targeting truncated CAR-iMACs, TIR-CAR-iMAC, and CD3ζ-TIR-CAR-iMACs (bright field) with the tumor cells at the E/T ratio of 10/1. c, The TIR-based CAR-iMACs cooperating with CD47-antibody cured HepG2-derived hepatocellular neoplasm in tumor-bearing mice. 5 × 105 FFluc+HepG2 cells were intraperitoneally injected to B-NDG mouse. 4 days after that, GPC3-targeting truncated CAR-iMACs, TIR-CAR-iMAC, and CD3ζ-TIR-CAR-iMACs mixed with CD47-antibody were injected intraperitoneally into the tumor-bearing mice. 2 hours after the treatment, live animal imaging was performed to monitor the growth of HepG2 cells in vivo. The treatment group with only CD47-antibody was designed as a control. d, The statistics of change of tumor burden from the treated mice from c (n = 5 mice from CD47 antibody group, n = 5 mice from truncated CAR-iMAC + CD47 antibody group, n = 5 mice from TIR-CAR-iMAC + CD47 antibody group, n = 10 mice from CD3ζ-TIR-CAR-iMAC + CD47 antibody group). e, The statistics of change of weight of the mice from c. All the above data was shown by GraphPad Prism 8.2.1. The significance was calculated with a two-way ANOVA analysis with multiple comparisons and are presented as mean±s.e.m. p-value: ns, not significant, *P < 0.05,***P < 0.001,****P < 0.0001.
Extended Data Fig. 5 The second-generation CAR improved the TME.
a, The schematic diagram of constructs of GPC3-targeting CARs specific to mouse cells. Compared to the human-specific second-generation CAR, we replaced the intracellular signal transduction domain with mouse-specific TIR and CD3ζ domains in tandem, and then named the new CAR G2-CAR. The truncated CAR (G1-CAR) without an intracellular signal domain was designed as a negative control. b, The imaging by fluorescence microscope showed the successful transduction of G2-CAR into mouse BMDMs. c and d, The flow cytometry analysis was performed to detect the expression level of the EGFP protein (c) and the expression level of the CAR (d). The results were from one experiment and were processed using FlowJo e, Confocal imaging showed the phagocytosis of EGFP-labeled CAR-BMDMs against tdTomato+ Hepa1-6 cells expressing human GPC3 (Hepa1-6GPC3) in vitro. The experiment was performed for three times. f, The statistic of CAR-BMDMs that have phagocytized the tumor cells from e (n = 11 BMDM samples per group from three independent experiments). g, The solid tumors were harvested nine days after subcutaneous injection of Hepa1-6GPC3 cells in C57BL/6 mice and six days after intratumoral CAR-BMDM treatment. The PBS treatment group was designed as a negative control. h and i, The change of tumor size (h) and body weight (i) were recorded from day 3 (CAR-BMDM treatment) to day 9 (n = 6 samples from PBS group, n = 6 samples from G1-CAR-BMDM group, n = 6 samples from G2-CAR-BMDM group). The data was shown by GraphPad Prism 8.2.1. The significance was calculated with a two-way ANOVA analysis with multiple comparisons and are presented as mean±s.e.m. p-value: **P < 0.01. j–n, The population of CD86-positive macrophages (j), CD206-positive macrophages (k), DC cells (l), NK cells (m), and CD69-positive T cells (n) were detected respectively by flow cytometry analysis from tumor samples in g (n = 6 independent samples per group). The data was shown by GraphPad Prism 8.2.1. The significance was calculated with a two-tailed/non-paired t-test and are presented as mean±s.e.m. p-value: ns, not significant, *P < 0.05, **P < 0.01.
Extended Data Fig. 6 IF assays showed subcellular localization of NF-κB/P65 in CAR-iMACs.
a, NF-κB/P65 exhibited nuclear localization in response to LPS stimulation to EGFRvIII-targeting CAR-iMACs (indicated by the white arrow). The red circle indicating monocyte-like iMACs showed high expression of NF-κB/P65 in the absence of LPS stimulation. b, 4 hours of incubation of U87MGEGFRvIII cells with four types of CAR-iMACs without IFN-γ/LPS pretreatment showed limited distribution of NF-κB/P65 in the nuclei of CAR-iMACs (indicated by the white arrow). The experiment was performed three times.
Extended Data Fig. 7 scRNA-seq analysis revealed the features of sub-populations of CAR-iMACs.
a, A tSNE plot of 10× Genomic single-cell RNA-seq data showing six clusters from the EGFRvIII-targeting truncated CAR-iMACs group after being incubated with U87MGEGFRvIII cells for 24 hours in vitro. b, Cell type annotations of the different sub-populations or clusters from a obtained by an entropy-based computational approach (Liu et al., 2020). c, A tSNE plot of the six main clusters derived from the TIR-CAR-iMACs group after being incubated with U87MGEGFRvIII cells for 24 hours in vitro. d, Cell type annotation of different clusters from c using an entropy-based computational approach. e, A tSNE plot of six main clusters in the CD3ζ-TIR-CAR-iMACs group after being incubated with U87MGEGFRvIII cells for 24 hours in vitro. f, Cell type annotation of different clusters from e. All the scales in b, d, and f were designed to indicate the percentage of a specific cell type.
Extended Data Fig. 8 scRNA-seq analysis revealed similar expression profiles between TIR-based CAR-iMACs and M1-like macrophages.
a, The heatmap comparing the gene expression profile of a set of M1 and M2-associated genes for EGFRvIII-targeting truncated CAR-iMACs, TIR-CAR-iMACs, and CD3ζ-TIR-CAR-iMACs to that of stimulated M1-like, M2-like and unstimulated iMACs. b, Hierarchical cluster tree showing the relationship of the truncated CAR-iMACs, TIR-CAR-iMACs, and CD3ζ-TIR-CAR-iMACs with stimulated M1-like, M2-like, and unstimulated iMACs. c, The heatmap comparing the gene expression profile of a set of M1 and M2-associated genes in CD80+ and nonCD80 cells in truncated CAR-iMACs, TIR-CAR-iMACs, and CD3ζ-TIR-CAR-iMACs to that of stimulated M1-like, M2-like and unstimulated iMACs. d, Hierarchical cluster tree showing the relationship of CD80+ and nonCD80 cells in truncated CAR-iMACs, TIR-CAR-iMACs, and CD3ζ-TIR-CAR-iMACs with stimulated M1-like, M2-like, and unstimulated iMACs. The data of IPSDM MAC, IPSDM M1, and IPSDM M2 were obtained from the GEO database (GSE55536).
Extended Data Fig. 9 A schematic illustration of the second-generation CAR.
All the CARs comprise an extracellular scFv domain recognizing a specific epitope, a transmembrane (TM) domain and a hinge region from CD8α. The truncated CAR was designed as the negative control without the intracellular signaling domain. The CD3ζ-CAR containing a single intracellular CD3ζ signaling domain was defined as the first-generation CAR corresponding to the first-generation T-CAR. The TIR-CAR contains an intracellular TIR signal transduction domain. The CD3ζ-TIR-CAR containing CD3ζ and TIR in the intracellular domain was defined as the second-generation CAR and possesses higher proinflammatory activity than the first-generation CAR upon binding to a specific antigen.
Supplementary information
Supplementary Video 1 (download MP4 )
A real-time video reveals the events of apoptosis and efferocytosis of HepG2 cells induced by CD3ζ-TIR-CAR-iMACs. The highlighted white arrows and numbers indicate the following events: the 1 arrow indicates the anchoring event of a GPC3-targeting CD3ζ-TIR-CAR-iMAC cell to a HepG2 cell; the 2 arrow points to the aggregation of tumor cell apoptotic bodies, which are gradually engulfed by the adjacent CD3ζ-TIR-CAR-iMAC cell; the 3 arrow indicates the apoptosis process of a living HepG2 cell induced by the CD3ζ-TIR-CAR-iMAC cell, and the apoptotic bodies are gradually taken up by the CAR-iMAC cell. The imaging was performed every 2 min.
Supplementary Video 2 (download MP4 )
A real-time video shows the apoptosis and efferocytosis of U87MGEGFRvIII cells mediated by CD3ζ-TIR-CAR-iMACs. The highlighted white arrows and numbers indicate the following events: the 1 arrow indicates the anchoring event of a tdTomato-labeled U87MGEGFRvIII cell to an adjacent EGFP-marked EGFRvIII-targeting CD3ζ-TIR-CAR-iMAC cell; the 1 and 2 arrows point to the apoptosis event of the tumor cell indicated by the formed granules, and the tumor cell is eventually eliminated; the 3 arrow and the white box show the debris from tdTomato-labeled U87MGEGFRvIII cell taken up by the CD3ζ-TIR-CAR-iMAC cells; the 4 arrow indicates that the apoptotic bodies of tumor cells are eventually eradicated after in contact with a CAR-iMAC cell; the 5 arrow indicates that a WT iMAC without the CAR and EGFP expression moves around without being anchored. The imaging was performed every 2 min.
Supplementary Data (download XLSX )
Sequence of oligonucleotides.
Source data
Source Data Fig. 1 (download XLSX )
Primary pictures and gating strategy of FCM.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download XLSX )
Statistical source data and gating strategy of FCM.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download XLSX )
Statistical source data and primary pictures of FCM.
Source Data Fig. 7 (download XLSX )
Statistical source data, gating strategy of FCM and primary pictures of FCM.
Source Data Extended Data Fig. 1 (download PDF )
Unprocessed western blots of Extended Data Fig.1b,c.
Source Data Extended Data Fig. 1 (download XLSX )
Primary pictures of Extended Data Fig. 1d,f.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data and gating strategy of FCM.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data and gating strategy of FCM.
Source Data Extended Data Fig. 4 (download XLSX )
Statistical source data, gating strategy of FCM and primary pictures.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data, gating strategy of FCM and primary pictures.
Source Data Extended Data Fig. 6 (download XLSX )
Primary pictures.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lei, A., Yu, H., Lu, S. et al. A second-generation M1-polarized CAR macrophage with antitumor efficacy. Nat Immunol 25, 102–116 (2024). https://doi.org/10.1038/s41590-023-01687-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41590-023-01687-8
This article is cited by
-
CAR-engineered cell therapies: current understandings and future perspectives
Molecular Biomedicine (2026)
-
Viscoelastic hydrogel primed CAR-macrophage for pulmonary fibrosis treatment
Nature Communications (2026)
-
Optimization of THP-1-CAR monocytes utilizing CD32a signaling phagocytosis for antigen-specific T cell activation
Scientific Reports (2026)
-
tRNA m1A modification orchestrates STING translation in macrophages to enhance antitumor immunity and CAR-macrophage immunotherapy
Cellular & Molecular Immunology (2026)
-
Fasting-mimicking diet induces IFNβ secretion in tumor-associated macrophages via NRF1-mediated ubiquitin-dependent proteolysis of Trex1
British Journal of Cancer (2026)
