
Abstract
Tumor-intrinsic adaptations with emerging resistant clones following T cell-targeted immunotherapies pose a major barrier to durable remissions in multiple myeloma. Through integrated genomic, transcriptomic and epigenomic interrogation of clonal plasma cells, we observed antigenic drift in 68.4% of relapsed cases following anti-GPRC5D T cell-engager (TCE) therapy (n = 21). These escape events were driven by three distinct mutational mechanisms involving (1) focal to large biallelic deletions at the GPRC5D gene locus; (2) monoallelic deletion coupled with GPRC5D single-nucleotide variants or insertions/deletions (indels) on the remaining allele; as well as (3) epigenetic GPRC5D promoter/enhancer silencing. Beyond biallelic deletions resulting in complete antigenic loss, we demonstrate that GPRC5D single-nucleotide variants and indels mutate anti-GPRC5D TCE-binding epitopes or more commonly affect G-protein-coupled receptor family conserved motifs critical for protein membrane trafficking resulting in endoplasmic reticulum GPRC5D trapping. Multiple subclones bearing distinct genomic alterations at GPRC5D locus co-emerged within individual cases, depicting their convergent evolutionary trajectories. Of note, anti-GPRC5D TCEs with varying epitope specificity, affinity and valency differentially targeted mutant subclones, underscoring their nonredundant functional roles in overcoming resistance.
Similar content being viewed by others


Main
The antitumoral efficacies of targeted immunotherapies, such as chimeric antigen receptor T cells (CAR T) and bispecific TCEs, hinge on the structural integrity and adequate expression of target antigens on tumor cell surface. In multiple myeloma (MM), B cell maturation antigen (BCMA) and G-protein-coupled receptor (GPCR) class C group 5 member D (GPRC5D) are among the key antigenic targets, with a wide array of anti-BCMA or -GPRC5D CAR T and TCE therapies being approved or undergoing clinical development1,2,3,4,5,6,7,8,9,10,11. Although these agents induce deep remissions in patients with relapsed refractory MM, resistance invariably develops in most patients1,2,3,4,5,6,7,8,9,10,11.
Immune-mediated selection of antigen escape clones is an inherent risk that limits the efficacy of single-epitope-targeting immunotherapies. Such antigenic escape mechanisms have been now well documented with anti-BCMA TCE12,13, and to a lesser extent post anti-BCMA CAR T5,14,15. With regard to GPRC5D, genomic analyses have been conducted on limited number cases to date, with biallelic deletions or epigenetic silencing documented post anti-GPRC5D CAR T (n = 8)16,17,18,19, as well as limited number of relapsed cases post anti-GPRC5D TCE demonstrating biallelic deletions (n = 2), single-nucleotide variants (SNVs)/indels with or without monoallelic deletions (n = 3) and epigenetic silencing (n = 2)13,20. While it is speculated that the emergence of these GPRC5D SNV/indel variants may represent immune-escape clones that precipitate MM resistance to anti-GPRC5D therapies, direct evidence supporting causality versus bystander mutation effect is lacking.
We here leveraged whole-genome sequencing (WGS) and single-cell RNA sequencing (scRNA-seq), in a large multi-institutional cohort of relapsing patients after anti-GPRC5D TCE, to investigate the genomic underpinnings of acquired resistance to talquetamab and linked the observed GPRC5D genomic alterations to outcomes through functional assays. In particular, we have identified a high frequency (68.4%) of GPRC5D mutational events at relapse-encompassing biallelic focal and large deletions, SNVs/indels and chromatin silencing at regulatory regions. Notably, GPRC5D SNVs/indels predominantly mapped to GPCR family conserved residues (regions required for the endoplasmic reticulum (ER)-to-cell membrane GPCR protein trafficking). We further demonstrate that the remaining mutations mapping outside these conserved GPCR residues led to functional epitope loss that differentially abrogated anti-GPRC5D TCE binding and cytotoxicity.
Results
Differential mutational patterns and convergent evolution of GPRC5D antigen escape clones post anti-GPRC5D TCE
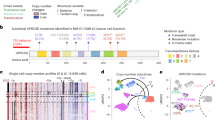
Patients with relapsed and refractory myeloma progressing on anti-GPRC5D TCE talquetamab (n = 21) were included in this analysis. A summary of the genomic analyses performed on each patient and their clinical characteristics are summarized in Fig. 1a,b and Supplementary Table 1. All patients were treated with talquetamab. All progression CD138+ samples were collected within 1–2 weeks of clinical confirmed disease progression by International Myeloma Working Group (IMWG) criteria21. Bulk WGS (100×) was performed on 20 patients (pre-therapy n = 8 and at relapse n = 20). For patients MM-03, MM-10 and MM-20, pre-therapy and relapse scATAC and/or scRNA-seq were performed in addition to WGS. Samples from patient MM-61 was subjected to targeted sequencing. In terms of clinical response, 14 of 21 cases had at least a very good partial response (VGPR) according to IMWG criteria21. Two patients (MM-65 and MM-67) had primary refractory disease (Fig. 1b). The median progression-free survival (PFS) for the overall group was 12.1 months (95% CI 8–18.1 months) (Fig. 1c).

a, Summary of patient cohort. Total 21 cases of anti-GPRC5D TCE relapse or patients with refractory MM were included. Sorted CD138+ plasma cells from bone-marrow aspirates and sorted CD3+ cells from peripheral blood samples were subjected to WGS, targeted sequencing, scATAC-seq and/ or scRNA-seq, as shown. Created in BioRender. Lee, H. (2026) https://BioRender.com/e28w8g. b, Swimmer plot showing PFS in months for 21 patients treated with anti-GPRC5D TCE talquetamab. Dotted vertical line indicates median PFS (12.1 months). Square label after each patient ID indicates types of samples collected for each patient. Circles indicate GPRC5D antigen escape status (red indicates yes, green indicates no). Numbers inside the circle refers to mechanisms of GPRC5D loss. Depth of response is annotated according to IMWG criteria. Case MM-65 and MM-67 had primary refractory MM. CR, complete response; LOH, loss of heterozygosity; PD, progressive disease; PR, partial response; sCR, stringent complete response; SD, stable disease; VGPR, very good partial response. c, Kaplan–Meier curve showing PFS in the overall cohort n = 21). Median PFS is reported. The gray shading area represents the 95% confidence interval (CI) around the survival probability estimate at each time point.
Genomic analyses revealed GPRC5D alterations including copy-number loss, structural variants, SNVs, indels and/or loss of chromatin accessibility at GPRC5D promoter/enhancer regions in 13 of 21 patients (61.9%). Excluding the two primary refractory cases, the GPRC5D mutational rate was 13 of 19 patients (68.4%). Among the patients with available baseline samples (n = 8), only one (MM-18) had pre-existing monoallelic loss of GPRC5D.
The three mutational patterns leading to GPRC5D antigen escape post-talquetamab included (1) clonal and subclonal GPRC5D SNVs/indels with or without copy-number changes in seven patients (MM-03, MM-31, MM-32, MM-61, MM-66, MM-68 and MM-69); (2) a focal to large biallelic deletion overlapping GPRC5D locus in five patients (MM-18, MM-19, MM-20, MM-54 and MM-62); and (3) epigenetic silencing coupled with GPRC5D frameshift deletion in one patient (MM-10). Monoallelic deletion of GPRC5D and copy-neutral loss of heterozygosity were each noted in patients MM-70 and MM-63, respectively. The remaining six patients (MM-64, MM-65, MM-67, MM-71, MM-72 and MM-73) showed no genomic events in GPRC5D by WGS.
Clonal and subclonal GPRC5D SNVs/indels
Among cases with SNVs/indels with or without monoallelic deletion involving GPRC5D, patient MM-03 previously received anti-BCMA TCE achieving complete response for 11 months13. CD138+ MM cells at progression harbored monoallelic deletion of TNFRSF17 coupled to a clonal point mutation in the extracellular domain of BCMA (p.Arg27Pro) abrogating anti-BCMA TCE cytotoxicity13. Subsequently, the patient was treated with talquetamab, daratumumab and pomalidomide (Talq-DP), and attained stringent complete response (sCR) lasting 13.8 months. scRNA-seq analysis of post-relapse CD138+ MM cells demonstrated retention of GPRC5D transcript (Fig. 2a). Pre-talquetamab WGS revealed a heterozygous germline single-nucleotide polymorphism (SNP) at GPRC5D Asp238 residue (c.714 G > A) (Fig. 1b, top left). At progression, two somatic GPRC5D SNVs were detected by WGS: a clonal missense mutation (p.Asp239Asn, c.715 C > T; cancer cell fractions (CCF) 1) in cis with the Asp238 SNP, and in trans GPRC5D nonsense mutation (p.Trp237Ter, c.710 C > T; CCF 0.82) affecting the second allele (Fig. 2b, bottom left). Therefore, as depicted in the phylogenetic tree in Fig. 2c (left), 82% of the cancer cells carried double-mutant GPRC5D alleles, whereas the residual 18% harbored p.Asp239Asn along with a wild-type GPRC5D allele. Flow cytometry showed reduced, but not absent, GPRC5D expression in the post-relapse CD138+ tumor cells (Fig. 2d,e). The GPRC5D mutants emerged from a common ancestral trunk clone harboring the clonal BCMA p.Arg27Pro mutant, selected post anti-BCMA TCE in the last line of therapy (Fig. 2b, right). This phylogenetic profile is consistent with the strong therapeutic bottleneck imposed by targeted immunotherapy (Fig. 2c, right panel and Extended Data Fig. 1a,b). Of note, a high APOBEC SBS COSMIC signature (SBS2 and 13) and mutational burden were observed in this sample at relapse as well as pre-therapy, as expected with the presence of MAF gene translocation (t(14;16)). (Extended Data Fig. 1c and Supplementary Fig. 1). In addition, the patient’s peripheral blood T cells at progression exhibited no evident functional T cell exhaustion as their peripheral blood mononuclear cells (PBMCs) effectively eradicated OPM2 MM cells in co-culture with talquetamab or elranatamab (comparable to co-culture with healthy donor PBMCs control) (Fig. 2f).

a, Uniform manifold approximation and projection (UMAP) of single-cell RNA transcripts (scRNA-seq) data from post-relapse CD138⁺ cells, showing TNFRSF17 (BCMA) and GPRC5D expressions. b, Integrative Genomics Viewer (IGV) snapshots of the GPRC5D and TNFRSF17 loci from pre-treatment and post-relapse CD138⁺ cells (WGS). c, A proposed phylogenetic tree depicting the evolutionary emergence of two GPRC5D mutants identified at progression (t2), with t0 representing pre-therapy and t1 representing an intermediate time point (left). Heterozygous GPRC5D p.D238 SNP is shown in green, in cis GPRC5D p.D239N mutation (purple) and in trans GPRC5D p.W237* mutation (yellow). Created in BioRender. Lee, H. (2026) https://BioRender.com/e28w8gr. Fish plot depicting clonal evolution across sequential lines of therapy, with each colored segment representing a genetically distinct subclone (right). Therapies administered at each time point are annotated along the timeline. d, Flow cytometry dot plots showing GPRC5D surface expression on CD138⁺ cells pre-treatment and at relapse as well as that of OPM2 MM cells. e, Histogram of flow cytometry results corresponding to d; numbers indicate median fluorescence intensity (MFI). f, Cytotoxicity assay of PBMCs at relapse, co-cultured with OPM2 MM cells at an effector:target (E:T) ratio of 10:1 for 24 h in the presence or absence of 10 nM talquetamab (talq) or elranatamab (elra). Viability of OPM2 MM cells was assessed by calcein AM⁺/propidium iodide⁻ staining. Healthy donor PBMCs were used as a positive control.
Other patients with acquired GPRC5D SNVs included MM-61 with t(11;14) MM who achieved a 12 months complete response on talquetamab. At the time of relapse, targeted amplicon sequencing of the CD138+ tumor cells demonstrated monoallelic deletion of GPRC5D (Extended Data Fig. 2a) coupled with convergent evolution of nine distinct subclones each harboring genomic events on GPRC5D leading to mutations p.Val210AlafsTer51 (VAF 32.2%), p.Leu108PhefsTer39 (VAF 23.0%), p.Thr128del (VAF 21.2%), p.Leu174_Tyr176del (8.3%), p.Trp237Ter (VAF 7.7%), p.Tyr2ThrfsTer41 (VAF 7.2%), p.Leu108GlnfsTer34 (VAF 4.5%), p.Ile51AspfsTer96 (VAF 4.2%) and p.Arg233Ter (VAF 3.5%) (Extended Data Fig. 2b,c). In addition, the relapsed tumors harbored clonal TP53 mutation p.Pro72Arg (VAF 100%).
Convergent evolution of multiple GPRC5D subclones harboring SNVs/indels was also evident at talquetamab relapse in case MM-66. Post-relapse tumor cells harbored subclones with GPRC5D p.Lys189ProfsTer36 (CCF 0.37), p.Tyr176TrpfsTer45 (CCF 0.26), p.Ser105Leu (CCF 0.39) coupled with monoallelic deletion of chr12: 41,087-19,554,375 (CCF_CNV 0.53) (Extended Data Fig. 3a–c). Notably this post-relapse tumor also harbored monoallelic and biallelic TP53 (CCF_CNV 1) and CYLD deletions (CCF_CNV 1), respectively (Extended Data Fig. 3c).
MM-68, recipient of previous allogeneic stem cell transplantation, relapsed post-talquetamab; MM cells at progression carried a subclonal GPRC5D p.Trp57Ter (CCF 0.08) with subclonal copy-number loss at the GPRC5D locus (Extended Data Fig. 4a,b). Case MM-69 also demonstrated monoallelic deletion of GPRC5D (CCF_CNV 1) coupled with GPRC5D p.Leu255Ter (CCF 0.39) at the time of relapse post-talquetamab (Extended Data Fig. 4c,d).
We also previously reported on MM-31 with wild-type GPRC5D before talquetamab who relapsed with convergent evolution involving four subclones, all sharing a large deletion of 30.5 Mb in chromosome 12p coupled with one of a 356-kb focal deletion at GPRC5D locus (CCF_CNV 0.12), p.Arg233ter (CCF 0.45), p.Tyr257Ser (CCF 0.28) and rearrangement between chr2 and the GPRC5D locus on chr12 (CCF 0.15)13. In case MM-32, convergent evolution of GPRC5D mutant subclones emerged at relapse post-talquetamab, harboring biallelic deletion of chromosome 12p (CCF_CNV 0.07) and another subclone with monoallelic GPRC5D loss coupled with GPRC5D p.Glu146ter (CCF 0.35)13.
Focal to large biallelic deletion overlapping GPRC5D
A second pattern of GPRC5D antigenic escape resulting from biallelic deletion was observed in five cases. MM-20 received Talq-DP and achieved sCR for 42 months. Bulk WGS of post-relapse tumor demonstrated a chr12p monoallelic deletion with a focal 104-kb biallelic deletion (CCF_CNV 1) encompassing GPRC5D gene locus (chr12: 12,877,450–12,981,981), which were undetectable pre-therapy (Fig. 3a,b). scRNA-seq analysis confirmed loss of GPRC5D transcript in the post-relapse sample and chr12p copy-number loss (inferCNV) (Fig. 3c and Supplementary Fig. 2). Flow cytometry was also consistent with nearly absent membrane GPRC5D expression at relapse (Fig. 3d). In vitro cytotoxicity assay using patient PBMCs collected at progression showed efficient killing of OPM2 cells in the presence of 10 nM talquetamab (Fig. 3e). Of note, this patient’s peripheral blood T cell population was predominantly composed of CD8+ T cells (Supplementary Fig. 3a–c).

a, Circos plot of post-relapse tumor based on bulk WGS. The outer track runs clockwise from chromosome 1 to Y. Middle track indicates genes with clonal SNV/indel events (red lines and genes labeled inside the circle). Inner track represents copy-number alterations; gain (red) and loss (blue). Lines inside the circle represent translocations (black) with translocations involving chromosome 14 (IgH locus) marked in red. Large deletions greater than 1 Mb are indicated in dark red lines. b, Whole-genome copy-number analysis (CNVkit) showing focal biallelic deletion at the GPRC5D locus in post-relapse tumor. c, UMAP of scRNA-seq data showing TNFRSF17 (BCMA) and GPRC5D expression in pre-therapy and post-relapse tumor cells. Red shows CD138 negative, green shows CD138 positive. d, Dot plot of flow cytometry staining for CD138 and GPRC5D in post-relapse CD138⁺ sorted cells. OPM2 MM cell line was included as a positive control. e, Cytotoxicity assay of patient PBMCs at relapse, co-cultured with OPM2 cells at an effector:target ratio of 10:1 for 72 h in the presence or absence of 10 nM talquetamab. Viability of OPM2 cells was assessed by calcein AM⁺/propidium iodide⁻ staining. Healthy donor PBMCs were used as a positive control.
In cases MM-54 and MM-62, WGS on post-talquetamab relapse tumor cells also demonstrated a large monoallelic deletion overlapping the GPRC5D gene locus coupled with a focal clonal 93 kb (chr12: 12,928,898–13,022,283) (CCF_CNV 0.92) and focal 6 kb (chr12: 12,948,661–12,954,728) (CCF_CNV 0.96) resulting in GPRC5D biallelic deletions, respectively (Extended Data Figs. 5 and 6).
MM-18 demonstrated biallelic GPRC5D deletion (CCF_CNV 0.95) at progression along with a subclonal fraction (CCF 0.04) harboring frameshift deletion GPRC5D p.Leu174TrpfsTer180 (ref. 13). Similarly, MM-19 relapsed with a clonal GPRC5D biallelic deletion (CCF_CNV 1), as we previously reported13.
Epigenetic GPRC5D silencing
The last pattern of acquired resistance involved epigenetic silencing. Patient MM-10 previously received anti-BCMA TCE and progressed after 19 months with a clonal p.Pro34del BCMA mutation13. The patient then received Talq-DP and achieved sCR lasting 19 months. WGS of CD138+ cells at relapse on talquetamab (but not pre-therapy) identified a GPRC5D mutation p.Thr88LeufsTer21 (CCF 0.91) with no other CNVs or structural variants at this gene locus (Fig. 4a,b). By contrast, scRNA inferred CNV analysis (InferCNV)22 showed single-copy-number loss encompassing GPRC5D gene along with loss of GPRC5D transcript (Fig. 4c,d and Supplementary Fig. 4). Flow cytometry analysis of CD138+ cells collected on subsequent line of therapy post-talquetamab showed near absent GPRC5D expression (Supplementary Fig. 5). The absence of measurable GPRC5D protein expression, together with heterozygous copy-number loss (inferCNV) and discordant ploidy status at the GPRC5D locus between messenger RNA-inferred CNV and WGS, can only be explained by epigenetic silencing. Indeed, chromatin accessibility studies (scATAC-seq) on the pre- and post-samples revealed loss of chromatin accessibility in the gene body as well as the proximal and distal GPRC5D regulatory regions as shown in Fig. 4e and Supplementary Fig. 6. Therefore, GPRC5D antigen escape in this patient (MM-10) resulted from bimodal epigenetic silencing coupled with truncating frameshift mutation in GPRC5D.

a, Circos plot comparing pre-treatment and post-relapse tumor genomes based on bulk WGS. The outer track runs clockwise from chromosome 1 to Y. Middle track indicates genes with clonal SNV/indel events (red lines and genes labeled inside the circle). Inner track represents copy-number alterations; gain (red) and loss (blue). Lines inside the circle represent translocations (black) with translocations involving chromosome 14 (IgH locus) marked in red. b, IGV screenshot of the GPRC5D locus from WGS data, pre-therapy and post-relapse. c. UMAP of single-cell RNA-seq data showing TNFRSF17 (BCMA) and GPRC5D expression in pre-therapy and post-relapse tumor cells with retained TNFRSF17 mRNA expression but loss of GPRC5D. InferCNV Hidden Markov Model (HMM)-based CNV prediction analysis at the GPRC5D gene locus (GPRC5D_CNV) reveals one copy-number loss (inferred CNV state 2 indicates loss of one copy) of GPRC5D in post-relapse cells. d, InferCNV heatmap, red arrow indicating copy-number loss of chromosome 12p segment harboring GPRC5D in post-relapse tumor cells. e, Genome chromatin accessibility track (ATAC-seq) spanning the GPRC5D locus (chr12: 12,782,146–13,222,147) in pre versus post samples. Highlighted peaks in red indicate the loss chromatin accessibility at GPRC5D gene body and regulatory regions. Genes in this region are shown below the track, with red indicating transcription on the positive strand and blue on the negative strand.
Genomic alterations in each tumor sample, including purity, ploidy, variant allelic fractions (VAFs) and purity-corrected cancer cell fractions (CCFs) for SNVs/indels and CNVs, are provided in Supplementary Table 2. Figures summarizing the clonal and subclonal CCF distribution for each patient with antigen escape are included in Supplementary Fig. 7.
Antigen-independent acquired resistance
Two patients, MM-71 and MM-73, achieved sCR with durable remissions lasting 24.8 and 28.1 months, respectively (Fig. 1b). At progression, neither patient had evidence of GPRC5D mutations by WGS. While retained antigen expression could not be examined at progression due to lack of primary material, functional evaluation of peripheral blood T cell fitness revealed decreased CD4:CD8 ratio with reduced T cell-mediated cytotoxicity in both patients compared to healthy donors (Supplementary Fig. 3d,e and Supplementary Fig. 8).
Driver mutations and mutational signatures
Among eight individuals with paired pre-therapy and relapse samples, total mutational burden increased at relapse in five patients (MM-03, MM-18, MM-31, MM-63 and MM-65) (Supplementary Fig. 1). SNV and indel analysis of all relapse samples revealed predominantly missense C>T variants (Extended Data Fig. 7a,b), with TP53 and GPRC5D mutations observed in 45% and 40% of cases, respectively. Within relapsed samples, comparing patients with or without GPRC5D antigen escape, no clear enrichment pattern of co-occurring mutations was observed (Extended Data Fig. 7c). Notably, dNdScv analysis comparing pre- and post-relapse samples identified GPRC5D and TP53 as significantly mutated at relapse (q_global_cv < 1 × 10⁻⁵), but not in pre-therapy samples (Supplementary Tables 3 and 4).
Hyper APOBEC signature (SBS2 + SBS13 > 10%) was observed in four patients (19%): three of whom had GPRC5D antigen escape (MM-03 pre-therapy and relapse, MM-54 relapse and MM-66 relapse) and one without antigen loss (MM-67 relapse) (Supplementary Figs. 1 and 9). GISTIC23 analysis revealed enrichment in 1q21 gain, 1p32 loss, 12p13.1 loss (GPRC5D locus) and 17p13 loss (TP53 locus) in relapsed samples (Supplementary Fig. 10).
A summary of the frequency of GPRC5D events, together with sample-level genomic features, is provided in Supplementary Fig. 9.
Impaired cell membrane trafficking of GPRC5D mutants
We functionally investigated the impact of the observed GPRC5D SNVs/indels post-talquetamab relapse on antigen expression and anti-GPRC5D TCE-mediated cytotoxicity. Mutant GPRC5Ds including p.Arg233Ter (R233ter), p.Tyr257Ser (Y257S), p.Glu146ter (E146ter), p.Asp239Asn (D239N) and p.Leu174TrpfsTer180 (L174W), as well as wild-type (wt) GPRC5D and empty lentiviral vector were cloned and stably expressed in K562 cell line (Fig. 5a–c). GPRC5D constructs were N-terminally tagged with a human influenza hemagglutinin (HA) tag to allow the detection of GPRC5D variants with both anti-HA and anti-GPRC5D antibodies. Wild-type GPRC5D and GPRC5D missense variants (p.Tyr257Ser and p.Asp239Asn) were detectable by western blotting at their expected molecular mass (34 kDa), whereas the three GPRC5D truncating or frameshift mutants (p.Arg233ter, p.Glu146ter or p.Leu174TrpfsTer180) resulted in smaller-sized bands (Fig. 5d). Confocal microscopy of fixed and permeabilized wild-type and mutant GPRC5D-expressing K562 cells demonstrated that, except for wild-type and the p.Asp239Asn mutant, all other GPRC5D mutants colocalized with intracellular calnexin staining, indicating their entrapment in the ER (Fig. 5e and Supplementary Fig. 11). Loss of cell surface expression of the GPRC5D mutants, with the exception of p.Asp239Asn, was also confirmed by flow cytometry (Fig. 5f). The expression patterns of these mutant GPRC5D clones were also validated in the GPRC5D-low-expressing human MM cell lines JJN3 (The Human Protein Atlas, https://www.proteinatlas.org/; Supplementary Fig. 12a–c). Of note, we confirmed that the N-terminal HA tagging of the GPRC5D transgenes did not interfere with membrane GPRC5D protein expression (Supplementary Fig. 13a,b).

a, Lollipop plot depicting the four exons of GPRC5D (noncoding exon 1 and coding exons 2–4) and the genomic coordinates of identified SNVs or indels. b, Topological domains of GPRC5D protein based on UniProt51 (Q9NZD1, GPC5D_HUMAN1); mutation sites identified in this study which were cloned and functionally validated are marked with red arrows. c, Cell membrane topology of GPRC5D illustrated using Protter52; SNV/indels cloned and functionally validated in this study are highlighted in red. d, Western blot analysis of GPRC5D expression in K562 cells transduced with wild-type or mutant constructs, and in empty vector control cells (K562_EV). Shown is the representative blot among three independent western blots. e, Confocal microscopy of K562 cells stained with anti-HA (GPRC5D) and anti-calnexin (ER marker) antibodies. Cells were fixed and permeabilized. TL, transmitted light. Image gain was adjusted to reflect maximal expression. Shown are the representative images from two independent experiments. f, Flow cytometry histograms of live transduced K562 cells stained with anti-HA or anti-GPRC5D antibodies. MFI is indicated. g, Schematic illustration of talquetamab (monovalent GPRC5D-binding TCE) and forimtamig (bivalent GPRC5D-binding TCE). h,i, Binding assays for talquetamab (h) and forimtamig (i). Transduced K562 cells were incubated with or without 100 nM of talquetamab or forimtamig, followed by staining with fluorophore-conjugated secondary antibodies specific for the Fc isotype of each TCE (anti-IgG4-FITC for talquetamab; anti-IgG1-PE for forimtamig). Flow cytometry was used to assess binding; MFI values are shown.
Rescue of intracellularly entrapped GPCRs has been reported by exposing the cells to lower temperatures or using various chemical or pharmacological chaperones24,25,26. We therefore investigated whether the mis-localized mutant GPRC5D proteins in the transduced K562 cells can be enforced to traffic to the cell membrane. Culturing the cells at 27 °C for 24 h or treatment with 1% dimethyl sulfoxide (DMSO), 4% glycerol or 10 μM all-trans retinoic acid (ATRA) for 24 or 48 h did not lead to detectable changes in membrane GPRC5D expression (Supplementary Fig. 14). Prolonged overnight staining with anti-GPRC5D antibodies also failed to demonstrate detectable surface expression (Supplementary Fig. 15a,b). Last, in the cycloheximide chase assay, we did not observe differences in protein stability among the various GPRC5D mutants (Supplementary Fig. 16).
Clustering of GPRC5D mutations within conserved GPCR family membrane-trafficking motifs
The GPRC5D SNV sites detected at relapse post-talquetamab, including Glu146, Leu174, Asp239 and Tyr257 are conserved residues across members (GPRC5A, GPRC5B, GPRC5C and GPRC5D) of the GPCR RAIG1 family (Extended Data Fig. 8). These conserved residues contain canonical motifs involved in ER-to-membrane trafficking25. In particular, GPRC5D p.Try257Ser and p.Leu174TrpfsTer180 alter the known tyrosine- and dileucine-based GPCR trafficking motifs, respectively24,27,28. Furthermore, the membrane-proximal C terminus of GPCRs is essential for their export from the ER to the cell surface28. Consistent with this, our in vitro observations showed that the identified nonsense and frameshift mutations (p.Arg233Ter, p.Glu146Ter, p.Leu174TrpfsTer180 and p.Trp237Ter), which introduce premature termination codons and truncate this critical C-terminal signaling domain, result in loss of antigen cell surface expression (Fig. 5e and Supplementary Fig. 11).
Differential sensitivity profiles of GPRC5D mutants to anti-GPRC5D TCEs
We next examined the binding and cytolytic activities of two anti-GPRC5D TCEs, talquetamab29 (monovalent anti-GPRC5D binding) and forimtamig30 (bivalent anti-GPRC5D binding) (Fig. 5g) against GPRC5D mutants. We first confirmed that the HA tagging for wild-type GPRC5D did not impede the binding or the cytolytic activity of anti-GPRC5D TCEs (Supplementary Fig. 13c,d). Treatment of p.Asp239Asn GPRC5D expressing K562 cells with 100 nM of talquetamab or forimtamig demonstrated efficient cell membrane binding of both TCEs (Fig. 5h, i). By contrast, neither talquetamab nor forimtamig binding (relative to anti-IgG control) was observed in K562 cells expressing p.Arg233Ter, p.Glu146Ter, or p.Leu174TrpfsTer180 GPRC5D mutants, consistent with their lack of measurable GPRC5D surface expression. One exception is the p.Tyr257Ser GPRC5D mutant where minimal binding of forimtamig (relative to anti-IgG control) was detectable despite its lack of flow cytometry measurable GPRC5D surface expression (Fig. 5i).
GPRC5D p.Asp239Asn mutant emerged at progression after a durable 13.8 months sCR in patient MM-03 who was treated with biweekly talquetamab (0.8 mg kg−1 body weight). The reported clinically achievable serum concentrations of talquetamab ranges from 1,000–2,700 ng ml−1 (6.9–18.6 nM)31. Therefore we evaluated in a dose–response assay the binding of anti-GPRC5D TCE to p.Asp239Asn mutant at concentrations ranging from 0.01–100 nM. Despite comparable surface expression of p.Asp239Asn to the wild-type GPRC5D (Fig. 6a), talquetamab failed to bind to the mutant p.Asp239Asn at concentrations lower than 68 nM, while displaying low level binding at higher concentrations (≥68 nM). At equimolar concentrations, binding of the bivalent anti-GPRC5D forimtamig to p.Asp239Asn was unperturbed (Fig. 6b). Consistent with their respective binding profiles, in a cytolytic assay against p.Asp239Asn, talquetamab log10-transformed half-maximum inhibitory concentration (IC50)) increased nearly 275-fold (relative to wild-type GPRC5D), whereas the log10[IC50] of forimtamig was relatively unchanged (Fig. 6c).

a, Flow cytometry histograms of K562 cells expressing wild-type GPRC5D (K562_wtGPRC5D) or K562_p.Asp239Asn, stained with anti-HA or anti-GPRC5D antibodies. b, Dose–response binding assay for anti-GPRC5D TCEs. K562_wtGPRC5D and K562_p.Asp239Asn cells were incubated with a range of talquetamab or forimtamig concentrations as indicated, followed by staining with a common fluorophore-conjugated anti-Fc (APC) secondary antibody to enable direct comparison of binding. Flow cytometry was used to assess binding; MFI values are shown. c, Dose–response cytotoxicity assay. K562_wtGPRC5D and K562_p.Asp239Asn cells were co-cultured with healthy donor PBMCs at an effector:target ratio of 10:1 for 48 h in the presence or absence of anti-GPRC5D TCEs. Target cell viability is shown on the y axis; log10[TCE (nM)] concentrations are on the x axis. Data are presented as mean ± s.d. from triplicate biologically independent experiments (n = 3 per cell line). P values represented in the figure correspond specifically to the comparison of dose–response curves between K562_p.Asp239Asn and K562_wtGPRC5D cells treated with talquetamab. Comparisons were performed using ordinary two-way analysis of variance (ANOVA) with factors for TCE concentration and cell line, followed by Tukey’s multiple comparison test with a single pooled variance to adjust for multiple comparisons using Prism. P values are two-sided and multiplicity adjusted (Supplementary Table 6 provides full results). NS, non-significant. d, Comparative dose–response cytotoxicity assay assessing the cytolytic activity of talquetamab and forimtamig across all transduced K562 cell lines. Co-cultures were performed at an effector:target ratio of 5:1 for 48 h. Data are presented as mean ± s.d. Raw data are from triplicate biologically independent experiments (n = for each cell line) and P values are available in Supplementary Table 7. e, Structural modeling of the p.Asp239Asn variant. Cryo-EM structure of wild-type GPRC5D (yellow) in complex with talquetamab (blue) (PDB: 9IMA), showing Asp239 forming an ion pair with Arg102 of talquetamab (top). AlphaFold2 Multimer model predicts that the p.Asp239Asn mutation disrupts this interaction (bottom). CDR H3, complementarity-determining region 3 of the heavy chain; ECL, extracellular loop; TM, transmembrane domain. f, Structural modeling of the p.Tyr257Ser variant. Tyr257 in wild-type GPRC5D forms a stabilizing hydrogen-bond network with Gln63 and His110 within the transmembrane core (top). AlphaFold2 Multimer model suggests this network is disrupted in the p.Tyr257Ser variant, potentially compromising receptor integrity (bottom).
We also evaluated the cytolytic activity of the talquetamab and forimtamig against the GPRC5D mutants (p.Arg233Ter, p.Tyr257Ser, p.Glu146Ter and p.Leu174TrpfsTer180) lacking measurable surface GPRC5D expression. The truncating mutants, p.Arg233Ter and p.Glu146Ter, as well as the frameshift mutant leading to early stop codon p.Leu174TrpfsTer180, were completely resistant to both anti-GPRC5D TCEs (Fig. 6d). Of interest, the p.Tyr257Ser mutant displayed minimal sensitivity to high-dose talquetamab (log10[IC50] 1.92, ~20,000-fold increase relative to wild-type GPRC5D), while retaining a partial sensitivity to forimtamig (log10[IC50] −0.16, ~66-fold increase relative to wild-type GPRC5D) (Fig. 6d). This minimal to partial sensitivity to a high dose of anti-GPRC5D TCEs is consistent with very low surface expression of p.Tyr257Ser, well below the limit of detection thresholds for flow cytometry and standard confocal microscopy. Therefore, we next used an optimized confocal imaging staining protocol (no permeabilization step) that permits the detection of low-density membrane proteins. Under these conditions, cell surface expression of p.Tyr257Ser GPRC5D mutant was detected (Extended Data Fig. 9).
GPRC5D-low-expressing escape clones retain sensitivity to a higher dose intensity of anti-GPRC5D TCE
Although p.Tyr257Ser GPRC5D K562 cells displayed undetectable surface GPRC5D by flow cytometry, they remained sensitive to forimtamig-mediated cytolysis as well as to high doses of talquetamab. This, together with detectable p.Tyr257Ser GPRC5D cell surface expression by enhanced confocal microscopy imaging, supported that this mutant GPRC5D protein is expressed at low levels sufficient for TCE engagement, despite being below the detection threshold of standard flow-based assays. To confirm that the cell killing resulted from an on-target effect, we compared the cytotoxic response of the p.Tyr257Ser GPRC5D mutant cells to that of GPRC5D-null K562 cells (K562_empty vector) and to MM cell line JJN3, which have undetectable GPRC5D levels by flow cytometry assays (Supplementary Fig. 17a). Despite the lack of flow cytometry measurable GPRC5D expression or TCE binding (Supplementary Fig. 17b,c), the cytotoxicity assays demonstrated that JJN3 exhibited partial sensitivity to anti-GPRC5D TCE-mediated killing comparable to that observed with p.Tyr257Ser GPRC5D mutant cells. By contrast, K562_empty vector cells remained viable at all tested TCE concentrations (0.01–20 nM) ruling out an off-target cytotoxicity effect (Supplementary Fig. 17d).
Structural modeling of GPRC5D SNVs and talquetamab
The structural interaction of membrane bound GPRC5D homodimer with the anti-GPRC5D Fab fragment of talquetamab has been characterized previously32, wherein Asp239 of GPRC5D was defined to be a critical residue recognized by talquetamab. Amino acid Arg102 within the complementarity-determining region 3 (CDR3) of the heavy chain of talquetamab protrudes into the shallow pocket in the extracellular domain of GPRC5D and forms an ion pair with Asp239 of GPRC5D32. GPRC5D mutation p.Asp239Asn emerged in patient MM-03 at progression on talquetamab as described earlier. Indeed, in silico modeling revealed that p.Asp239Asn missense GPRC5D led to the abrogation of the ion pair between Arg102 of talquetamab and GPRC5D Asn239, destabilizing the interaction between these two molecules (Fig. 6e). In the case of the p.Tyr257Ser GPRC5D missense mutation (identified in patient MM-32), Tyr257 directly interacts with Gln63 and His110 of the GPRC5D molecules. Tyr257 side-chain is tightly packed in the hydrophobic core of GPRC5D by directly interacting with Gln63 and His110. Mutation of this GPRC5D residue to serine (p.Tyr257Ser) is predicted to destabilize the molecule and affect its membrane localization (Fig. 6f).
GPRC5D epigenetic silencing in t(11;14) myeloma
We observed reduced chromatin accessibility at the GPRC5D promoter and enhancer in patient MM-10 with t(11;14) myeloma. Epigenetic silencing of GPRC5D was recently reported in patients with MM progressing after anti-GPRC5D TCE and CAR T16,20. While t(11;14) MM is recognized to be associated with reduced expression of GPRC5D at the transcript and protein levels, the biological basis for this association is unknown30,33,34,35. Given the known plasma-cell-restricted expression pattern of GPRC5D35,36 and the B cell-like transcriptional and epigenetic program of t(11;14)37,38,39, we re-analyzed our previously published cohort with single-cell chromatin accessibility (scATAC-seq) coupled with single-cell transcriptome analysis (scRNA-seq) from CD138+ t(11;14) and non-t(11;14) MM39. Pre-treatment bone-marrow CD138+ tumor samples were used to compare GPRC5D gene expression and chromatin accessibility between t(11;14) and non-t(11;14) tumor cells. t(11;14) tumors (PM01, 05, 07, 09, 10, 11) demonstrated lower GPRC5D transcript levels. Notably, scATAC-seq revealed reduced chromatin accessibility in t(11;14) samples at the GPRC5D gene body and promoter compared to non-t(11;14) samples (Extended Data Fig. 10a,b). Peak-to-gene (Peak2Gene40) analysis also identified key enhancer regions 5′ and 3′ to GPRC5D gene noting reduced chromatin accessibility in these regulatory regions only in t(11;14) tumor samples (Extended Data Fig. 10a and Supplementary Table 9). Furthermore, high-throughput chromosome conformation capture (Hi-C) analysis in the GM12878 lymphoblastoid cell line41 revealed a topologically associating domain encompassing the GPRC5D-HEBP1 locus, with enriched intra-domain interactions consistent with coordinated regulatory activity. Notably, off-diagonal contact enrichment revealed long-range chromatin looping between EMP1 and GPRC5D, supporting distal enhancer-promoter communication consistent with our Peak2Gene analysis of scATAC-seq data (blue highlighted peaks in Extended Data Fig. 10A and Supplementary Fig. 18).
Discussion
Antigen escape through GPRC5D loss represents a major hurdle to the sustained efficacy of GPRC5D-targeted immunotherapies in MM. Biallelic deletions of GPRC5D is emerging as a mechanism by which MM cells evade TCE and CAR T cell therapies13,19,20. In this study, we demonstrate that antigen loss is not limited to structural variant with copy-number alterations, but rather, a broader spectrum of genetic alterations including SNVs, small indels within GPRC5D, as well as epigenetic silencing mechanisms. Our findings highlight a more complex and heterogeneous landscape of immune evasion than previously appreciated. We have identified frequent GPRC5D mutational events resulting in functional antigenic loss in nearly two-thirds of anti-GPRC5D TCE-treated patients. These events consisted of three main patterns: (1) focal (as small as 6 kb) to large biallelic deletion overlapping GPRC5D; (2) nonsense and missense SNVs and indels involving GPRC5D gene with or without hemizygous copy-number losses; and (3) epigenetic silencing of GPRC5D promoter and enhancer regions.
While the loss of TCE activity resulting from target gene biallelic deletion is evident, the impact of SNVs and indels on TCE efficacy is unclear. SNVs and indels were the most common events involving the GPRC5D gene (9 of 21), followed by structural variants and copy-number alterations. These missense and nonsense SNVs/indels involved extracellular, transmembrane and intracellular domains of GPRC5D. Of note, all events within the cytoplasmic domain consisted of nonsense or frameshift mutations leading to protein truncation. SNVs involving transmembrane domain predominantly clustered within GPCR family conserved motifs involved in ER to membrane trafficking leading to GPRC5D ER trapping. Mutations involving the extracellular domain grouped into deletions, nonsense, frameshift mutations leading to protein truncation or missense mutations resulting in functional epitope loss (Extended Data Fig. 8b).
Notably GPRC5D protein expression patterns by flow cytometry or standard confocal imaging did not always correlate with clinical TCE resistance nor with their in vitro cytotoxic activity. Therefore caution is warranted when adopting these approaches as clinical screening platforms for patient selection and response prediction to anti-GPRC5D therapies. Of note, ER trapping is well established as the most common mechanisms of GPCR protein inactivation28,42. Indeed in our series, some GPRC5D mutants, in particular those involving ER-to-membrane-trafficking motifs (p.Arg233Ter, p.Glu146Ter and p.Leu174WfsTer180) led to GPRC5D ER trapping with complete loss of surface expression and ensuing loss of monovalent and bivalent anti-GPRC5D TCE cytotoxicity. In contrast, other missense mutations such as p.Tyr257Ser retained sensitivity to bivalent anti-GRPC5D TCE (and high-dose monovalent TCE) despite undetectable surface GPRC5D expression by flow cytometry and standard confocal microscopy. This discrepancy between the lack of detectable p.Tyr257Ser GPRC5D on the cell surface and the observed sensitivity of p.Tyr257Ser-expressing K562 cells to forimtamig suggests that such mutations may partially impair, rather than completely block, GPRC5D trafficking to the cell surface. This is consistent with the detectable GPRC5D expression by our enhanced confocal imaging. An alternative explanation is that p.Tyr257Ser GPRC5D mutant undergoes dynamic or transient surface presentation, which is difficult to capture using static detection methods such as flow cytometry, yet sufficient to enable T cell-mediated cytotoxicity by high avidity anti-GPRC5D TCE. Similar to our observation, prior studies have demonstrated that tumor cells with very low levels of target antigen (<100 molecules per cell), below the detection threshold of flow cytometry or immunohistochemistry, may remain sensitive to CAR T cell-based therapies43.
We also observed GPRC5D SNVs with fully retained surface expression but loss of clinical response and in vitro resistance to talquetamab. This is the case for the missense mutation p.Asp239Asn which disrupts a key GPRC5D epitope required for talquetamab binding as confirmed by both in vitro and in silico analyses. Notably, cell lines expressing the GPRC5D p.Asp239Asn variant exhibited a flow cytometry immunolabelled anti-GPRC5D fluorescence signal indistinguishable from wild-type GPRC5D. These findings underscore the necessity of incorporating genomic sequencing-based strategies into clinical workflows, rather than solely relying on antibody-based assays, for the detection of specific epitope changes.
The GPRC5D mutations p.Tyr257Ser and p.Asp239Asn, which reduce surface expression and alters the functional epitope binding respectively, confer dose-dependent resistance to anti-GPRC5D TCEs. These mutants require higher TCE concentrations for effective therapeutic targeting compared to wild-type GPRC5D. Therefore, while balancing on-target off-tumor toxicities, it is critical to avoid sub-therapeutic dosing or premature dose de-escalation of anti-GPRC5D TCE, as such adjustments may undermine treatment efficacy in patients harboring these mutations. Alongside maintaining adequate TCE concentrations, other strategies include the development of multivalent and biparatopic anti-GPRC5D TCEs. These designs, which target unique, nonoverlapping epitopes on GPRC5D, may provide a means to bypass the effects of these mutants, via their multivalency (distinct epitope targets) and/or increased avidity toward GPRC5D. Notably, the cytotoxic activity of forimtamig, anti-GPRC5D TCE with two asymmetrically arranged GPRC5D-binding domains with affinity dissociation constant (Kd) of 11 nM and avidity Kd of 0.2 nM (ref. 30), was unaffected by the p.Asp239Asn mutation, and less impacted by the p.Tyr257ser mutation, while the cytolytic activity of talquetamab (single GPRC5D binding domain, affinity Kd of 4.21 nM (ref. 44) was impaired by both mutants. Alternatively, dual antigen targeting approaches with trispecific antibodies45 or CAR T cells targeting BCMA and GPRC5D46, may eliminate these emerging single antigen mutant clones.
The post-talquetamab relapse in MM-10 with t(11;14) MM, along with previous studies16,20, implicate epigenetic silencing as a mechanism of acquired resistance to GPRC5D-targeted immunotherapy. Chromatin accessibility studies demonstrate that t(11;14) MM, characterized by a transcriptional and epigenetic B cell-like landscape37,39, exhibits reduced GPRC5D chromatin accessibility and expression at baseline. Furthermore, the restricted expression of GPRC5D to plasma cells supports a dynamic epigenetic regulation of GPRC5D expression along the B cell to plasma-cell developmental trajectory. These findings raise several critical questions: does subtype-specific epigenetic repression of GPRC5D as in t(11;14) MM contribute to primary or acquired resistance? In the context of relapse, does GPRC5D silencing reflect a broader epigenetic reprogramming event, wherein MM cells reprogram to a more B cell-like or less plasma-cell-like state? Clinically available data to date have not investigated the outcomes for the t(11;14) MM subgroup with anti-GPRC5D targeting therapies. Future clinical studies will need to further investigate whether MM subtype-specific epigenetic repression of GPRC5D contributes to resistance. Dissecting whether resistance is driven by lineage plasticity versus clonal selection has important implications for patient stratification, the reversibility of antigen loss and the future development of combination strategies aimed at restoring target expression by agents such as chromatin modifiers16.
Antigenic loss was not observed in 6 of 19 patients who progressed after initial response to anti-GPRC5D TCE. Of interest, two of these patients had durable remissions lasting more than 2 years with no detectable mutant GPRC5D clones at progression. Emerging data suggest that continuous TCE therapy may lead to loss of TCE antitumor efficacy47,48. While the current analysis strongly supports antigenic loss as the main driver of resistance to TCEs, this does not preclude the possible co-occurrence of reduced T cell fitness with prolonged TCE therapy.
The genomic drivers of multiclonal convergent evolution under selective immune pressure remain to be elucidated. Of note, several GPRC5D mutations emerging on anti-GPRC5D TCE therapy have also been reported in germline as well as somatic non-MM tumor samples49,50. It is therefore plausible that these GPRC5D mutations arise stochastically and endow the mutant clones with a survival advantage only under sustained anti-GPRC5D immune selective pressures. Such paradigm of immune evasion warrants investigating whether finite TCE therapy exposure can limit stochastic clone selection reducing the risk of antigenic drift and resistance.
Methods
Patient CD138+ MM sample collection
This study was approved by the Conjoint Health Research Ethics Board at the University of Calgary (ethics ID HREBA.CC-21-0248), the Health Research Ethics Board at the University Hospital of Würzburg (Würzburg EK 8/21), Heidelberg University (S-096/2017), Kapodistrian University of Athens (116/08-02-2018) and University of Toulouse (DC-2012-1654), and are consistent with the Declaration of Helsinki. Bone marrow and peripheral blood samples were obtained after written informed consent. All samples with adequate cell counts or somatic DNA material from patients relapsing after anti-GPRC5D TCE (talquetamab) therapy at participating institutions in this collaborative study were included in this analysis. Therefore, no formal sample size calculation was required before the study. Patients treated with anti-GPRC5D TCE (talquetamab) (n = 21) with study protocol 64407564MMY1002/ClinicalTrials.gov identifier NCT04108195 (n = 13), study protocol TALMMY1001-PT3/ClinicalTrials.gov identifier NCT04634552 (n = 3) and nonclinical trial access (n = 5) were included in the study (Supplementary Table 1).
CD138+ MM cells were isolated as follows. Mononuclear cell fraction from peripheral blood or bone-marrow aspirates were collected by Ficol gradient centrifugation (30 min at 400g with slow acceleration and deceleration). Cells were incubated with CD138+ magnetic beads (Miltenyi Biotec, 130-051-301) for 15 min on ice, washed with MACS buffer and separated using MS magnetic columns (Miltenyi Biotec, 130-042-201) according to the manufacturer’s instructions. Isolated CD138+ cells were cryopreserved in freezing buffer (90% FBS with 10% DMSO) and stored at −80 °C. Details regarding reagents and antibodies used in this study are listed in Supplementary Table 10.
Whole-genome sequencing and analysis
WGS was performed as previously reported13. Libraries were prepared using the Truseq DNA Nano Library Preparation kit (Illumina, 20015965). Then, 100 ng of DNA were sheared using a Covaris LE220 sonicator (adaptive focused acoustics), and processed with bead-based size selection, end-repaired, adenylated, ligated to Illumina sequencing adaptors and amplified. Final libraries were quantified using the Qubit Fluorometer (Life Technologies) or Spectramax M2 (Molecular Devices) and Fragment Analyzer (Advanced Analytical) or Agilent 2100 BioAnalyzer. Libraries were sequenced on Illumina Novaseq6000 sequencer or NovaseqX using 2 × 150-bp cycles. The New York Genome Center (NYGC) pipeline was used for post-sequencing analysis and somatic variant calls. SNVs were called integrating Mutect2 (v.4.2.5.0), Strelka2 (v.2.9.3), Lancet (v.1.0.7); and indels using Mutect2 (v.4.2.5.0), Manta (v.1.4.0), Strelka2 (v.2.9.3), Lancet (v.1.0.7), SvABA (v.1.2.0); SV using Manta (v.1.4.0), SvABA (v.1.2.0) and Lumpy (v.0.2.1). Complex SV were defined according to our previously published criteria53,54. CNV analysis was conducted using BICSEQ2 (ref. 55) (v.0.2.6) and CNVkit56 (v.0.9.12) (https://github.com/etal/cnvkit).
The reported CCFs in Supplementary Table 2 were calculated using ASCAT57 (v.3.2.0) (https://github.com/VanLoo-lab/ascat) and Palimpsest58 (https://github.com/FunGeST/Palimpsest). Mutational SBS COSMIC signatures analyses were performed using SigProfilerAssignmentR (v.0.2.3) (https://github.com/AlexandrovLab/SigProfilerAssignment) and mmsig59 (https://github.com/evenrus/mmsig). Mutational burden was analyzed using mutationalpatterns60 (v.3.18.0) (https://github.com/UMCUGenetics/MutationalPatterns).
dNdScv R package (v.0.1.0) (https://github.com/im3sanger/dndscv)61 was used to estimate gene-level dN:dS ratios and identify genes under positive selection in the pre-therapy and post-relapse sample cohort.
FACETS62 (v.0.6.2) (https://github.com/mskcc/facets) was used to estimate CCFs for copy-number alterations.
Genomic variant (SNV) summary plots and oncoplots were generated with R packages maftools63 (https://github.com/PoisonAlien/maftools) and ComplexHeatmap64 (https://github.com/jokergoo/ComplexHeatmap).
GISTIC2 (ref. 23) (v.2.0.23) (https://github.com/broadinstitute/gistic2) was used to identify significantly recurrent copy-number alterations across the study cohort subgroups.
Further details of the NYGC somatic variant analysis can be found at https://www.nygenome.org/bioinformatics/wp-content/uploads/2019/06/SomaticPipeline_v6.0_Human_WGS.pdf.
For patient samples collected at the University Hospital Center Würzburg, WGS libraries and analysis were conducted as previously reported13. Bone-marrow-sorted plasma cells from patient MM-61 from Heidelberg University were subjected to targeted sequencing consisted of commonly mutated MM genes, including GPRC5D and TNFRSF17.
Matched normal was available for all except two patients (MM-62 and MM-63) as summarized in Supplementary Table 1. For these samples (MM-62 and MM-63), genomic DNA from the contemporary normal NA12878 (from the Coriell Cell Repository GM12878 lymphoblastoid cell line, part of the 1000 Genomes Project) was used as germline control. NA12878, MM-62 and MM-63 were sequenced on the same Illumina platform and aligned to hg38. Germline WGS (NA12878) was processed with the same pipeline as the somatic DNA from MM-62 and MM-63 and used to (1) build a CNVkit WGS reference for copy-number normalization, and (2) create a limited Mutect2 panel of normals to filter recurrent technical artifacts. Somatic variant calls were further filtered using gnomAD (hg38) to mitigate germline contamination due to the mismatched normal.
scRNA-seq and analysis
Unbiased scRNA-seq of bone-marrow-sorted CD138+ MM cells were conducted as previously published13. In brief, primary CD138+ sorted MM cells were processed using 10x Genomics reagent kits (CG00052 v.2 Chemistry), and partitioned into nanoliter-scale gel bead-in-emulsions (GEMs) via 10x GemCode Technology. The GEMs contained primers with an Illumina R1 sequence, 16-bp 10x barcode, 10-bp unique molecular identifier (UMI) and poly(dT) sequence, which were used to generate barcoded full-length complementary DNA from polyadenylated mRNA. Silane magnetic beads removed excess reagents, followed by PCR amplification of the cDNA. Enzymatic fragmentation and size selection were performed to optimize cDNA amplicon size. The R1 primer was added during GEM incubation, whereas P5, P7, i7 and R2 primers were added during library construction via end repair, A-tailing, adaptor ligation and PCR.
The KAPA Library Quantification qPCR kit (Roche) was used on a Bio-Rad qPCR instrument to quantify libraries. A pooled library with equimolar amounts of each sample was subjected to cluster formation and sequencing on an Illumina NextSeq 500 using a high-output v.2.5 150-cycle RNA-seq kit following standard Illumina protocols. After sequencing, bcl data were converted to fastq files using the Illumina BCL2FASTQ utility. Genomic reads were processed with Cell Ranger (v.7.1.0), aligning to the GRCh38 reference genome.
For gene expression analysis, filtered HDF5 matrices from the Cell Ranger (https://www.10xgenomics.com/support/software/cell-ranger/latest/release-notes) pipeline were imported into Seurat (v.5.3.0) for normalization, scaling, integration, clustering, dimensionality reduction and differential expression analysis. Cells with <1,000 UMIs, >25,000 UMIs or >25% mitochondrial reads were excluded. Cell annotation was conducted Azimuth. We used the InferCNV (v.1.20.0) package (https://github.com/broadinstitute/infercnv) with default parameters to derive virtual copy-number profiles and identify tumor subclones.
scATAC-seq and analysis
Frozen cells were thawed at 37 °C, resuspended in RPMI 1640 medium (Gibco) and washed twice by centrifugation at 400g for 5 min. The desired number of nuclei (2,000–8,000) was targeted and processed according to the Nuclei Isolation for Single Cell ATAC sequencing (CG000169, 10x Genomics). Nuclei isolation was performed as indicated in the nuclei isolation protocol for Single Cell ATAC Sequencing (10x Genomics). Based on the starting number of cells and desired final nuclei concentration, primary MM cells were lysed and resuspended in an appropriate volume of chilled Diluted Nuclei Buffer. The resulting nuclei were immediately used to generate scATAC-seq libraries using the Chromium Single Cell ATAC reagent kit (CG000168, 10x Genomics) according to the manufacturer’s protocol. Quality control and quantification of libraries were performed using the KAPA Library Quantification qPCR kit (Roche) on a Bio-Rad qPCR instrument. Final libraries were then sequenced on the Illumina NextSeq 1000 sequencer using a high-output v.2.5 150-cycle sequencing kit per standard Illumina protocols. After sequencing, the bcl data were converted to fastq data files using the Illumina BCL2FASTQ utility. Samples were processed with CellRanger-ATAC (v.2.1.0) and downstream analyses were performed as indicated below.
scATAC-seq analysis was performed as previously reported39 using ArchR (v.1.0.3)40. The plotBrowserTrack function from ArchR was used to plot the GPRC5D trackplot. The plot region window was adjusted by modifying the number of base pairs in the downstream (270,000 bp) and upstream (170,000 bp) of the GPRC5D transcription start site to allow the visualization of all gene regions.
Peak-to-gene linkage
Peak-to-gene analysis was performed on MM samples from our previously published data39. The data are available via the Gene Expression Omnibus (GEO) public repository GSE199373, consisting of GSE199268 for scATAC-seq and GSE199359 for scRNA-seq. After data processing and scRNA-seq integration with ArchR40, cells from the post-venetoclax treatment samples were removed from the ArchR project using the subsetArchRProject function and saved in a new ArchR project. A total of 15 samples (15 patients), 46,237 cells, with a median transcription start site of 11.997 and frags of 13,448 were obtained. The plotBrowserTrack function from ArchR was used to plot the GPRC5D trackplot using the scRNA-seq integrated data (useMatrix = GeneIntegrationMatrix) and default parameters. The plot region window was adjusted by modifying the number of base pairs in the downstream (270,000 bp) and upstream (170,000 bp) of the GPRC5D transcription start site to allow the visualization of all gene regions and identified peak-to-gene associations. Only peaks associated with GPRC5D were plotted on the trackplot.
Cell lines and cloning
GPRC5D expressing K562 cell lines were generated using the following protocol. Human GPRC5D cDNA sequence was taken from the NCBI Consensus Coding Sequence (CCDS) database (CCDS ID:CCDS8658.1)65. Custom DNA oligonucleotide and primers were ordered from Integrated DNA Technologies (IDT). GPRC5D insert sequences were designed with and without human influenza hemagglutinin (HA) sequence (amino acids YPYDVPDYA) at the N terminus.
The insert sequences from IDT were cloned into a pLX307 lentiviral plasmid backbone (Addgene, 117734) under an EF-1α promoter. The cloned wild-type and mutant GPRC5D sequences are indicated in Supplementary Table 11. Restriction enzymes Nhe1 and EcoRV were used for insert DNA and vector plasmid digestion, followed by overnight ligation using T4 DNA ligase. NEB stable competent Escherichia coli cells (NEB, C3040H) were used for transformation per manufacturer protocol. Then, 100 ng of plasmid DNA was added to the cell mixture, incubated on ice for 30 min, placed in water bath at 42 °C for 30 s, then placed back on ice for 5 min. Then, 950 μl of NEB 10-β/Stable Outgrowth Medium was added to the mixture and incubated at 30 °C for 1 h. Cells were then spread onto an ampicillin selection plate and incubated overnight at 37 °C. Bacterial colonies selected from the ampicillin agar plate were processed with a QIAprep Spin Miniprep kit (QIAGEN, 27104) as per the manufacturer’s instructions. Extracted DNA was validated by Sanger sequencing (Molecular Services Core, Univ. Calgary). Transfer plasmid was generated from positive clones using an EndoFree Plasmid Maxi kit (QIAGEN, 12362).
Lentiviral packaging of the transfer plasmid, psPAX2 packaging plasmid, (Addgene, 12260) and pMD2.G envelope plasmid (Addgene, 12259) was performed in HEK 293T cells using calcium phosphate transfection (Thermo Fisher, K278001). HEK 293T cells were cultured in DMEM with 10% FBS and 1% penicillin–streptomycin. Cells were plated 1 day before transfection and transfected the next day using a standard calcium phosphate method with 2× HEPES-buffered saline. The cell medium was removed after 18 h followed by incubation in fresh medium. Cells were collected 72 h after transfection, washed with PBS and pelleted at 400g. Supernatant was filtered using 0.45-μm filter, followed by ultracentrifugation at 90,000g. The lentiviral pellet was resuspended in PBS and stored in −80 °C.
K562 and JJN3 cells were resuspended in RPMI medium supplemented with 10% FBS, 1% penicillin–streptomycin, 0.2% normocin and lentivirus suspension was added for transduction. Transduction efficiency was confirmed by checking GPRC5D expression by anti-HA or anti-GPRC5D flow cytometry. Transduced cell lines were cultured in RPMI medium at 37 °C/5% CO2 with puromycin selection (1 μg ml−1). The newly generated GPRC5D expressing K562 and JJN3 cell lines and associated plasmids are available upon reasonable request. Requests can be directed to the corresponding author. CLASTR short tandem repeat profiling66 results for the authentication of MM cell lines used in this study are provided in Supplementary Tables 12 (OPM2) and 13 (JJN3).
Flow cytometry
CD138⁺ patient-derived cells obtained from the tissue bank were thawed and washed twice with RPMI medium. Cells were then resuspended in cell-staining buffer (CSB) and incubated with either anti-HA or anti-GPRC5D antibodies at 4 °C for 30 min. Antibodies were used at a 1:50 dilution (2 μl antibody per 100 μl staining volume) unless otherwise specified. A complete list of antibodies used for flow cytometry is provided in Supplementary Table 10. After staining, cells were washed and resuspended in CSB for analysis. For cell line experiments, 10,000 live events were recorded based on forward and side-scatter gating. In the case of primary CD138⁺ tumor samples, the maximum number of viable events was collected, depending on sample viability and yield. Gating strategies are summarized in Supplementary Fig. 19. Flow cytometry was performed using a Beckman CytoFLEX instrument, and all data analysis and figure generation were carried out using Kaluza Analysis Software v.2.1 (Beckman Coulter).
For low-temperature and pharmacological chaperone treatments, cells were either incubated at 27 °C for 24 h or treated at 37 °C with one of the following agents: 1% DMSO, 4% glycerol or 10 μM ATRA for 24 or 48 h. After treatment, cells were collected and assessed for GPRC5D surface expression by flow cytometry.
For the overnight cell-staining protocol, cells were seeded in 96-well plates (200 μl per well) in RPMI medium supplemented with 10% FBS and incubated with 4 μl per well of anti-GPRC5D-PE antibody at 37 °C for 18 h. The following morning, cells were washed with PBS and resuspended in CSB for flow cytometry analysis.
Western blot
A total of 5 million cells were collected, centrifuged, washed with PBS and incubated in RIPA buffer supplemented with protease and phosphatase inhibitors for 30 min on ice. Cell lysates were centrifuged at 14g for 10 min at 4 °C. Collected lysates were stored at −80 °C until use for western blot. Protein quantification was performed using DC Protein Assay kit (Bio-Rad, 5000111). Equal amounts of protein were mixed with 4× Laemmli sample buffer, boiled at 95 °C for 5 min and separated on NuPAGE 4–12% gel, and transferred to nitrocellulose membrane using wet transfer at 100 V for 85 min at 4 °C. Membranes were blocked for 1 h at room temperature in 5% nonfat milk in TBS-T (Tris-buffered saline with 0.1% Tween-20), then incubated overnight at 4 °C with primary antibodies diluted in blocking buffer. Primary antibodies used in this manuscript were diluted at 1:1,000. Supplementary Table 10 provides a list of antibodies. After washing three times with TBS-T, membranes were incubated with HRP-conjugated secondary antibodies for 1 h at room temperature. Secondary antibodies used for western blotting were diluted at 1:3,000. Protein bands were visualized using Clarity Western ECL Substrate (Bio-Rad, 1705061) and imaged using a chemiluminescence detection system (Bio-Rad ChemiDoc Imaging System).
Immunofluorescence staining
For standard immunofluorescence staining, cells were formaldehyde-fixed by applying 4% (v/v) PFA in PBS at room temperature for 20 min. The formaldehyde was then quenched with 50 mM NH4Cl for 10 min at room temperature. Cells were then washed three times with PBS and then permeabilized with 0.4% (v/v) Triton X-100 in PBS for 15 min, followed by three washes with PBS. Primary antibodies (anti-HA and anti-calnexin) were added (1:500 dilution) to the cells for 1 h in PBS + 2% FBS (v/v) at room temperature. Cells were then washed three times with PBS + 2% FBS, secondary antibodies were added for 1 h in PBS + 2% FBS at room temperature, followed by three washes with PBS. Cells were left in PBS, plated onto coverslipped-bottom eight-well dishes (Ibidi, 80826) and confocal images were acquired to visualize the localization of immunolabelled HA and calnexin. Images were acquired on a Leica SP8 confocal microscope using a HC APO ×63/1.40 oil CS2 objective.
The immunofluorescence staining protocol was optimized for membrane GPRC5D expression detection by eliminating the permeabilization step in p.Tyr257Ser-expressing K562 cell line. Live cells were spun onto coverslip using cytospin at 26g for 3 min, followed by fixation as described above. CytoLiner dye (Biotium cat. no. 30133A) was used to label the plasma membrane per the manufacturer’s protocol (incubation for 10 min with 1 ml dye in 500 ml buffer). Cells were washed with PBS 3 times and then incubated with the anti-HA antibody (1:500 dilution) overnight at 4 °C. The next day cells were washed with PBS 3 times and secondary antibody (rabbit IgG-AF648) was added for 1 h in PBS + 2% FBS at room temperature, followed by washing three times with PBS. The coverslip was then mounted onto glass slides with PBS. Confocal image stacks were acquired at 0.2 µm intervals with a Nikon A1R confocal system (acquisition software NIS elements software v.5.42.04) using a Nikon Plan Apo l ×60/1.40 NA objective. The immunolabelled HA fluorescence and CytoLiner were imaged sequentially with 561 and 638 nm excitation, with the corresponding emission detection at 583/46 nm and >640 nm, single-labeled controls were used to check for cross-talk and bleed-through. Images shown in were visualized using identical brightness and contrast.
TCE-binding assay
Talquetamab and forimtamig used for the TCE-binding assays were obtained from Janssen Pharmaceuticals and Roche, respectively. K562 cells were resuspended in 100 μl of CSB with 0.01–100 nM of TCE for 15 min at room temperature. The cells were washed with PBS, resuspended in CSB and incubated with anti-IgG4 Fc or anti-IgG1 Fc flow antibodies (1:50 dilution) for 15 min at 4 °C. Cells were washed with CSB before flow cytometry analysis. Then, 10,000 live single events (based on forward- and side-scatter plots) were recorded and anti-IgG antibody expression levels were determined based on MFI on single parameter histograms. Gating strategies are summarized in Supplementary Fig. 10. All flow experiments in this manuscript were performed using the Beckman CytoFLEX flow cytometer. Figures for flow cytometry results were generated using Kaluza Analysis Software v.2.1 (Beckman Coulter).
TCE cytotoxicity assay
Healthy donor or patient PBMCs were collected from Ficoll gradient centrifugation and washed with PBS for their use as effector cells in co-culture assays. Target K562 or MM cells were stained with CellTrace Violet (CTV) for 20 min at 37 °C protected from light followed by washing with PBS. PBMCs were co-cultured with CTV stained K562 cells in RPMI medium (containing 10% FBS, 1% penicillin–streptomycin and 0.2% normocin) with or without TCE at varying concentrations. Co-culture experiments were incubated for 24 or 72 h at 37 °C/5% CO2.
Calcein AM and propidium iodide (PI) were used to stain the cells before flow cytometry as per manufacturer protocols. Then, 10,000 events of CTV-positive cells were collected per treatment condition. After gating on CTV-positive cells (BV421 channel), the cells were displayed on two parameter dot plot (mCherry for PI and FITC for calcein AM). Cells staining strongly positive for calcein AM were considered viable and their gated percentage was used to compare target cell viabilities between treatment conditions. Gating strategies are summarized in Supplementary Fig. 19.
In silico structural modeling
We generated models for two GPRC5D mutants that associated with GPRC5D antigen escape using AlphaFold2 Multimer67. Specifically, we utilized the AlphaFold2_multimer_v3 version via ColabFold (v.1.5.5)68 with the number of relaxations set to 1 to perform Amber (Assisted Model Building with Energy Refinement) energy minimization. To facilitate prediction, the wild-type GPRC5D complex (PDB 9IMA) was used as a template. The number of recycles was set to 3, and the predicted template modeling (PTM) score, which evaluates the quality of the multimer model, was calculated. Both the PTM score and pLDDT values were considered to rank the five predictions, and the rank 1 model was selected as the final model for analyses. The models were checked and drawn using COOT (v.0.9.8.92)69 and UCSF Chimera X (v.1.8)70, respectively.
Statistical analysis
Due to restricted availability of primary CD138+-sorted bone-marrow cells and PBMCs from patients, experiments involving these samples were only conducted once. Experiments involving cell lines were conducted in independent triplicate experiments.
Cytotoxicity data and dose–response curves were analyzed using GraphPad Prism v.9. TCE concentrations were log-transformed and nonlinear regression was applied to generate log(inhibitor) versus response curves. For IC50 determination, response values were normalized to percent viability relative to PBMC control (set as 100%) and maximal inhibition with TCE (set as 0%), and curves were fitted using a four-parameter logistic model with the top and bottom constrained to 100 and 0, respectively, and the Hill slope fitted freely. Absolute IC50 values and 95% CIs were derived from the best-fit model using the profile-likelihood method.
For comparative analysis of K562 target cell viability in TCE and CAR T cytotoxicity assays, an ordinary two-way ANOVA was performed with factors for treatment condition and target cell construct. Tukey’s multiple comparison test was applied to assess all pairwise differences, using a single pooled variance and multiplicity-adjusted two-sided P values. Reported values in Supplementary Tables 5–8 include exact P values (up to 15 decimal places), alongside mean differences and 95% CIs. A family-wise significance threshold of α = 0.05 was applied.
Kaplan–Meier survival curves were generated using the R packages Survival (v.3.8-3) and Survminer (v.0.5.0).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
WGS data have been deposited in the European Genome–phenome Archive (EGA) under accession number EGAD50000001643. In accordance with EGA policies and the European Union General Data Protection Regulation, access to the dataset is controlled. Data can be obtained upon request through the designated Data Access Committee (EGAC00001002170). Requests are reviewed promptly, and once complete and compliant documentation is provided, access is granted within 10 business days.
For the GPRC5D chromatin accessibility studies for patients with t(11;14) MM, scATAC-seq and scRNA-seq data were deposited to the NCBI’s GEO repository under GEO SuperSeries accession code GSE199373, accession numbers GSE199268 for scATAC-seq and GSE199359 for scRNA-seq. Additional scATAC-seq and scRNA-seq data are available under accession no. GSE301287 and GSE301286, respectively. Source data are provided with this paper.
Code availability
All bioinformatic tools used in this study are indicated in the relevant sections of the Methods, with versions and modified parameters.
References
Moreau, P. et al. Teclistamab in relapsed or refractory multiple myeloma. New Engl. J. Med. 387, 495–505 (2022).
Lesokhin, A. M. et al. Elranatamab in relapsed or refractory multiple myeloma: phase 2 MagnetisMM-3 trial results. Nat. Med. 29, 2259–2267 (2023).
Bahlis, N. J. et al. Elranatamab in relapsed or refractory multiple myeloma: the MagnetisMM-1 phase 1 trial. Nat. Med. 29, 2570–2576 (2023).
Chari, A. et al. Talquetamab, a T-cell-redirecting gprc5d bispecific antibody for multiple myeloma. New Engl. J. Med. 387, 2232–2244 (2022).
Munshi, N. C. et al. Idecabtagene vicleucel in relapsed and refractory multiple myeloma. New Engl. J. Med. 384, 705–716 (2021).
Berdeja, J. G. et al. Ciltacabtagene autoleucel, a B-cell maturation antigen-directed chimeric antigen receptor T-cell therapy in patients with relapsed or refractory multiple myeloma (CARTITUDE-1): a phase 1b/2 open-label study. Lancet 398, 314–324 (2021).
D’Souza, A. et al. A phase I first-in-human study of ABBV-383, a B-cell maturation antigen × CD3 bispecific T-cell redirecting antibody, in patients with relapsed/refractory multiple myeloma. J. Clin. Oncol. 40, 3576–3586 (2022).
Cooper, D. et al. Safety and preliminary clinical activity of REGN5458, an anti-BCMA x anti-CD3 bispecific antibody, in patients with relapsed/refractory multiple myeloma. Blood 134, 3176 (2019).
Carlo-Stella, C. et al. RG6234, a GPRC5DxCD3 T-cell engaging bispecific antibody, is highly active in patients (pts) with relapsed/refractory multiple myeloma (RRMM): updated intravenous (iv) and first subcutaneous (sc) results from a phase I dose-escalation study. Blood 140, 397–399 (2022).
Harrison, S. J. et al. Efficacy of forimtamig, a GPRC5D×CD3 bispecific antibody, in patients with relapsed/refractory multiple myeloma: analysis of patient- and disease-related factors associated with responses. Clin. Lymphoma Myeloma Leuk. 23 (S3–S4), Abstract OA-05 (2023).
Schinke, C. D. et al. Pivotal phase 2 MonumenTAL-1 results of talquetamab (tal), a GPRC5DxCD3 bispecific antibody (BsAb), for relapsed/refractory multiple myeloma (RRMM). J. Clin. Oncol. 41, 8036 (2023).
Truger, M. S. et al. Single- and double-hit events in genes encoding immune targets before and after T cell-engaging antibody therapy in MM. Blood Adv. 5, 3794–3798 (2021).
Lee, H. et al. Mechanisms of antigen escape from BCMA- or GPRC5D-targeted immunotherapies in multiple myeloma. Nat. Med. 29, 2295–2306 (2023).
Da Vià, M. C. et al. Homozygous BCMA gene deletion in response to anti-BCMA CAR T cells in a patient with multiple myeloma. Nat. Med. 27, 616–619 (2021).
Samur, M. K. et al. Biallelic loss of BCMA as a resistance mechanism to CAR T cell therapy in a patient with multiple myeloma. Nat. Commun. 12, 868 (2021).
Ma, S. et al. Genetic and epigenetic mechanisms of GPRC5D loss after anti-GPRC5D CAR T-cell therapy in multiple myeloma. Blood 146, 178–190 (2025).
Mailankody, S. et al. GPRC5D-targeted CAR T cells for myeloma. New Engl. J. Med. 387, 1196–1206 (2022).
Jurgens, E. M. et al. Phase I trial of MCARH109, a G protein-coupled receptor class C group 5 member D (GPRC5D)-targeted chimeric antigen receptor T-cell therapy for multiple myeloma: an updated analysis. J. Clin. Oncol. 43, 498–504 (2025).
Mi, X., Penson, A., Abdel-Wahab, O. & Mailankody, S. Genetic basis of relapse after GPRC5D-targeted CAR T cells. New Engl. J. Med. 389, 1435–1437 (2023).
Derrien, J. et al. Acquired resistance to a GPRC5D-directed T-cell engager in multiple myeloma is mediated by genetic or epigenetic target inactivation. Nat. Cancer 4, 1536–1543 (2023).
Kumar, S. et al. International Myeloma Working Group consensus criteria for response and minimal residual disease assessment in multiple myeloma. Lancet. Oncol. 17, e328–e346 (2016).
Patel, A. P. et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 344, 1396–1401 (2014).
Mermel, C. H. et al. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol. 12, R41 (2011).
Dong, C., Filipeanu, C. M., Duvernay, M. T. & Wu, G. Regulation of G protein-coupled receptor export trafficking. Biochim. Biophys. Acta 1768, 853–870 (2007).
Inoue, S., Nambu, T. & Shimomura, T. The RAIG family member, GPRC5D, is associated with hard-keratinized structures. J. Invest. Dermatol. 122, 565–573 (2004).
Tao, Y. X. & Conn, P. M. Chaperoning G protein-coupled receptors: from cell biology to therapeutics. Endocr. Rev. 35, 602–647 (2014).
Tan, C. M., Brady, A. E., Nickols, H. H., Wang, Q. & Limbird, L. E. Membrane trafficking of G protein-coupled receptors. Annu. Rev. Pharmacol. Toxicol. 44, 559–609 (2004).
Stoy, H. & Gurevich, V. V. How genetic errors in GPCRs affect their function: possible therapeutic strategies. Genes Dis. 2, 108–132 (2015).
Pillarisetti, K. et al. A T-cell-redirecting bispecific G-protein-coupled receptor class 5 member D x CD3 antibody to treat multiple myeloma. Blood 135, 1232–1243 (2020).
Eckmann, J. et al. Forimtamig, a novel GPRC5D-targeting T-cell bispecific antibody with a 2+1 format, for the treatment of multiple myeloma. Blood 145, 202–219 (2025).
Center for Drug Evaluation and Research. NDA/BLA Multi-disciplinary Review and Evaluation {BLA 761342}, application number 761342Orig1s000 (2022).
Jeong, J., Park, J., Young Mo, G., Shin, J. & Cho, Y. Structural basis for the recognition of GPRC5D by talquetamab, a bispecific antibody for multiple myeloma. J. Mol. Biol. 436, 168748 (2024).
Tomita, U. et al. A novel T cell-redirecting anti-GPRC5D×CD3 bispecific antibody with potent antitumor activity in multiple myeloma preclinical models. Sci. Rep. 14, 5135 (2024).
Verkleij, C. P. M. et al. Preclinical activity and determinants of response of the GPRC5D×CD3 bispecific antibody talquetamab in multiple myeloma. Blood Adv. 5, 2196–2215 (2021).
Smith, E. L. et al. GPRC5D is a target for the immunotherapy of multiple myeloma with rationally designed CAR T cells. Sci. Transl. Med. 11, eaau7746 (2019).
Jelinek, T. et al. Depletion of mature B cells and of normal plasma cells (PC) behind the higher incidence of infections after anti-BCMA VS anti-GPRC5D bispecific antibodies (bsAb) in relapsed refractory multiple myeloma (RRMM). Blood 144, 1934 (2024).
Gupta, V. A. et al. Venetoclax sensitivity in multiple myeloma is associated with B-cell gene expression. Blood 137, 3604–3615 (2021).
Kitadate, A. et al. Multiple myeloma with t(11;14)-associated immature phenotype has lower CD38 expression and higher BCL2 dependence. Cancer Sci. 112, 3645–3654 (2021).
Leblay, N. et al. Integrated epigenetic and transcriptional single-cell analysis of t(11;14) multiple myeloma and its BCL2 dependency. Blood 143, 42–56 (2024).
Granja, J. M. et al. ArchR is a scalable software package for integrative single-cell chromatin accessibility analysis. Nat. Genet. 53, 403–411 (2021).
Rao, S. S. et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 159, 1665–1680 (2014).
Tao, Y. X. Inactivating mutations of G protein-coupled receptors and diseases: structure-function insights and therapeutic implications. Pharmacol. Ther. 111, 949–973 (2006).
Nerreter, T. et al. Super-resolution microscopy reveals ultra-low CD19 expression on myeloma cells that triggers elimination by CD19 CAR-T. Nat. Commun. 10, 3137 (2019).
Li, F. et al. Abstract 1875: A novel T-cell-engaging therapeutical antibody against GPRC5D&BCMA for multiple myeloma treatment. Cancer Res. 83, 1875–1875 (2023).
Donk, N. W. C. J. V. D. et al. First-in-human study of JNJ-79635322 (JNJ-5322), a novel, next-generation trispecific antibody (TsAb), in patients (pts) with relapsed/refractory multiple myeloma (RRMM): initial phase 1 results. J. Clin. Oncol. 43, 7505–7505 (2025).
Fernández de Larrea, C. et al. Defining an optimal dual-targeted CAR T-cell therapy approach simultaneously targeting BCMA and GPRC5D to prevent BCMA escape-driven relapse in multiple myeloma. Blood Cancer Discov. 1, 146–154 (2020).
Duffy, A. M. et al. Early CD4+ T-cell proliferative burst and chronic T-cell engagement impact myeloma outcomes following T-cell engager therapy. J. Clin. Invest. https://doi.org/10.1172/jci192927 (2025).
Cortes-Selva, D. et al. Correlation of immune fitness with response to teclistamab in relapsed/refractory multiple myeloma in the MajesTEC-1 study. Blood 144, 615–628 (2024).
Tate, J. G. et al. COSMIC: the catalogue of somatic mutations in cancer. Nucleic Acids Res. 47, D941–D947 (2019).
Sudlow, C. et al. UK biobank: an open access resource for identifying the causes of a wide range of complex diseases of middle and old age. PLoS Med. 12, e1001779 (2015).
Consortium, T. U. UniProt: the Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 51, D523–D531 (2022).
Omasits, U., Ahrens, C. H., Müller, S. & Wollscheid, B. Protter: interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 30, 884–886 (2013).
Rustad, E. H. et al. Revealing the impact of structural variants in multiple myeloma. Blood Cancer Discov. 1, 258–273 (2020).
Maclachlan, K. H. et al. Copy number signatures predict chromothripsis and clinical outcomes in newly diagnosed multiple myeloma. Nat. Commun. 12, 5172 (2021).
Xi, R., Lee, S., Xia, Y., Kim, T. M. & Park, P. J. Copy number analysis of whole-genome data using BIC-seq2 and its application to detection of cancer susceptibility variants. Nucleic Acids Res. 44, 6274–6286 (2016).
Talevich, E., Shain, A. H., Botton, T. & Bastian, B. C. CNVkit: genome-wide copy number detection and visualization from targeted DNA sequencing. PLoS Comput. Biol. 12, e1004873 (2016).
Raine, K. M. et al. ascatNgs: identifying somatically acquired copy-number alterations from whole-genome sequencing data. Curr. Protoc. Bioinform. 56, 15.19.11–15.19.17 (2016).
Shinde, J. et al. Palimpsest: an R package for studying mutational and structural variant signatures along clonal evolution in cancer. Bioinformatics 34, 3380–3381 (2018).
Rustad, E. H. et al. mmsig: a fitting approach to accurately identify somatic mutational signatures in hematological malignancies. Commun. Biol. 4, 424 (2021).
Blokzijl, F., Janssen, R., van Boxtel, R. & Cuppen, E. MutationalPatterns: comprehensive genome-wide analysis of mutational processes. Genome Med. 10, 33 (2018).
Martincorena, I. et al. Universal patterns of selection in cancer and somatic tissues. Cell 171, 1029–1041.e1021 (2017).
Shen, R. & Seshan, V. E. FACETS: allele-specific copy number and clonal heterogeneity analysis tool for high-throughput DNA sequencing. Nucleic Acids Res. 44, e131 (2016).
Mayakonda, A., Lin, D. C., Assenov, Y., Plass, C. & Koeffler, H. P. Maftools: efficient and comprehensive analysis of somatic variants in cancer. Genome Res. 28, 1747–1756 (2018).
Gu, Z., Eils, R. & Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 32, 2847–2849 (2016).
Pruitt, K. D. et al. The Consensus Coding Sequence (CCDS) project: identifying a common protein-coding gene set for the human and mouse genomes. Genome Res. 19, 1316–1323 (2009).
Robin, T., Capes-Davis, A. & Bairoch, A. CLASTR: the Cellosaurus STR similarity search tool - a precious help for cell line authentication. Int. J. Cancer 146, 1299–1306 (2020).
Evans, R. et al. Protein complex prediction with AlphaFold-Multimer. Preprint at bioRxiv https://doi.org/10.1101/2021.10.04.463034 (2022).
Mirdita, M. et al. ColabFold: making protein folding accessible to all. Nat. Methods 19, 679–682 (2022).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D 60, 2126–2132 (2004).
Pettersen, E. F. et al. UCSF Chimera–a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Acknowledgements
The authors are grateful to the patients who enrolled in the study. H.L. was supported by grant support from the Multiple Myeloma Research Foundation, Leukemia Lymphoma Society Canada and International Myeloma Society (IMS). N.J.B. was supported by the Paula and Rodger Riney Foundation, IMS, IMS Translational Research Award, Blood Cancer United (formerly, The Leukemia Lymphoma Society), The Leukemia Lymphoma Society Canada and the Terry Fox Foundation Marathon of Hope Cancer Center Networks. M.S.R. was funded by IMS Translational Research Award. L.R. was funded by the Paula and Rodger Riney Foundation and the Bundesministerium für Bildung und Forschung. L.H.B. was funded by the Paula and Rodger Riney Foundation and Blood Cancer United (formerly, The Leukemia Lymphoma Society). P.N. was funded by the Paula and Rodger Riney Foundation. N.W. was funded by the Advanced Medical Scientist Program of Heidelberg University, Faculty of Medicine. H.E. was supported by the German Research Foundation, CRC/TRR 338, Z02. K.M.K. was funded by the German Research Foundation and Stifterverband. F.M. was supported by a Memorial Sloan Kettering Cancer Center National Cancer Institute Core Grant (P30 CA 008748). C.R. was supported by a fellowship (project number ZZ-40) of the Interdisciplinary Center for Clinical Research. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript. We thank the Snyder Institute for support of the Live Cell Imaging Laboratory, University of Calgary (RRID:SCR_024748).
Author information
Authors and Affiliations
Contributions
H.L. conceptualized the study, designed and performed all in vitro experiments, analyzed and interpreted data, generated the figures and co-wrote the paper. S.A. performed single-cell experiments. G.G., D.G., J.C. and P.C. performed confocal imaging, cell-staining protocol optimization, generated figures and aided in the design and interpretation of optimized immunofluorescence confocal imaging. N.L. performed analysis of scATAC and scRNA-seq data. E.B. performed primary tumor sorting. S.B., M.P., E.H., D.J. and F.M. analyzed data. M.T., C.R., H.E., M.K., S.E., J.H.F., M.R., L.R., J.C., H.A., K.T., E.K. and P.N. provided patient samples and analyzed data. N.W. and L.H.B. contributed to data interpretation and study design. J.J. and Y.C. performed in silico analyses and generated the figures. J.E., J.S., D.V. and S.S. provided patient samples, study reagents and analyzed data. N.J.B conceptualized, designed and supervised the study, analyzed and interpreted scRNA and WGS data, generated figures and co-wrote the paper.
Corresponding author
Ethics declarations
Competing interests
N.J.B. has received research funding from Pfizer, and received speaker’s bureau honoraria from Amgen, BMS, Sanofi, Pfizer and Janssen and is a consultant/advisory board member for BMS, Janssen and Pfizer. P.N. received speaker’s bureau honoraria from BMS, Janssen and Sanofi and is a consultant/advisory board member for BMS and Janssen. D.V. and S.S. are employees of Johnson & Johnson and may hold stock, stock options or equity in Johnson & Johnson. All other authors declare no competing interests.
Peer review
Peer review information
Nature Medicine thanks Bruno Paiva, Marta Chesi and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Primary Handling Editor: Ulrike Harjes, in collaboration with the Nature Medicine team. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Case MM-03 undergoes stepwise antigen escape evolution through sequential BCMA- and GPRC5D-directed TCE therapies.
A. Stepwise clonal evolution of the dual antigen escape clone. Created in BioRender. Lee, H. (2026) https://BioRender.com/e28w8gr. B. Kernel density plot depicting the clonal distribution of the mutated gene variants with VAF depicting variant allelic frequency. X axis represent VAF and Y axis represents the density of various clusters. Shown in red dotted lines are VAFs for the TNFRSF17 and the two GPRC5D mutants. C. Cosmic mutational signature SBS 1, 2, 5, 13 for patient MM-03 relapse sample (left panel), showing enrichment of APOBEC signature (SBS2 + SBS13) and high mutational load (middle and right panels).
Extended Data Fig. 2 Post-relapse targeted sequencing in case MM-61.
A. Targeted amplicon sequencing with CNVkit analysis revealing single-copy loss of GPRC5D (coordinates aligned to hg19). Shown in the table are the targeted probes mapping to GPRC5D gene locus with corresponding genomic coordinates, log2 fold change, and depth of sequencing. B. Fish plot illustrating clonal evolution with the trunk clonal t(11;14) translocation, clonal TP53 mutation (p.Pro72Arg), and nine branching clones carrying various GPRC5D mutations. C. IGV screenshots showing distinct GPRC5D mutant subclones detected at relapse.
Extended Data Fig. 3 Post-relapse CD138 + MM WGS data for case MM-66.
A. Circos plot of post-relapse genome based on bulk whole-genome sequencing (WGS). The outer track runs clockwise from chromosome 1 to Y. Middle track indicates genes with clonal SNV/indel events (red lines and genes labeled inside the circle). Inner track represents copy number alterations — gain (red), loss (blue), neutral (gray). Lines inside the circle represent translocations (black) with translocations involving chromosome 14 (IgH locus) marked in red. Large deletions greater than 1 Mb are in dark red. B. IGV screenshot displaying three GPRC5D mutant clones with variant allele frequencies (VAFs). C. Copy number loss at GPRC5D, TP53, and CYLD loci detected by CNVkit at relapse post-talquetamab.
Extended Data Fig. 4 WGS data for cases MM-68 and MM-69.
A. CNVkit analysis of MM-68 post-relapse tumor genome (WGS) depicting copy number loss at GPRC5D gene locus. B. Lollipop plot (Maftool) of MM-68 post-relapse tumor representing a truncating GPRC5D p.Trp57Ter mutation. 7tm_3 indicates Pfam database annotation of 7-transmembrane domain family #3 (rhodopsin-like GPCR family). C. MM-69 post-relapse tumor clonal (CCF_CNV 1) monoallelic copy number loss at GPRC5D locus by CNVkit WGS analysis. D. MM-69 IGV screenshot demonstrating GPRC5D frameshift deletion p.Leu255Ter with CCF 0.39.
Extended Data Fig. 5 Post-relapse CD138 + MM whole-genome sequencing (WGS) data for case MM-54.
A. IGV screenshot with biallelic deletion (93 kb) spanning chr12:12,928,898–13,022,283 (GRCh38), encompassing the GPRC5D locus. B. Circos plot of post-relapse tumor based on bulk whole-genome sequencing (WGS). The outer track runs clockwise from chromosome 1 to Y. Middle track represents copy number alterations—gain (red) and loss (blue). Genes are marked in the inner most track. The plot depicts biallelic deletion of the GPRC5D locus, biallelic deletion of TNFRSF17 (chr16:11,924,238-12,021,653), as well as CDKN2C biallelic deletion on chr1p. Also shown are on monoallelic loss of TP53 on chr17p and BCL2 gain on chr18q. C. CNV plots generated by CNVkit at the GPRC5D locus (yellow) on chr12p. The top panel depicts monoallelic deletion spanning chr12p, while the bottom panel depicts focal 93 Kb deletion at GPRC5D gene locus (chr12: 12,928,898–13,022,283).
Extended Data Fig. 6 6. Post-relapse CD138 + MM whole-genome sequencing (WGS) data for case MM-62.
A. IGV screenshot showing focal deletion (6 Kb) spanning chr12:12,948,661-12,954,728 (GRCh38), encompassing GPRC5D locus. B. CNVkit copy number plot of post-relapse WGS data, showing a broad monoallelic deletion on chr12p that includes GPRC5D (top panel), along with a focal biallelic deletion restricted to the GPRC5D locus (bottom panel).
Extended Data Fig. 7 Summary of mutational landscape detected by WGS in post-talquetamab relapse CD138+ tumor samples (n = 20).
A. Variant classification summary across tumor samples (n = 20 independent biological samples). Bar graphs show the number of variants, with missense mutations, SNP (single-nucleotide polymorphisms), and C > T conversions being the most frequent. Insertions (INS), deletions (DEL), and di- or tri-nucleotide polymorphisms (DNP, TNP) are also indicated. B. Mutation substitution spectrum and transition (Ti) versus transversion (Tv) analysis across 20 independent biological samples. Boxplots indicate the interquartile range (boxes; 25th-75th percentile), median (horizontal line), and whiskers extending to 1.5x the interquartile range. No technical replicates were included. C. Oncoplot summarizing the mutation landscape of top mutated genes in all relapse samples (top panel, n = 20) compared to cases with GPRC5D antigen escape mutations (bottom panel, n = 12). TP53 is among the most frequently mutated genes across both groups. Case MM-61 with targeted sequencing only (instead of WGS) was excluded from this analysis.
Extended Data Fig. 8 Sequence alignment and annotation of GPRC5D mutations.
A.Alignment of GPCR family proteins (GPRC5A-D). GPRC5D mutations reported in this manuscript (red). GPRC5D transmembrane domains highlighted in blue; talquetamab binding residues reported by Jeong et al.32 indicated with green dots. Bar heights indicate residue conservation. Figure generated using SnapGene software (www.snapgene.com) sequence alignment. B. Full-length GPRC5D annotated with talquetamab binding sites and mutations. Amino acid residues of GPRC5D are shown (Figure generated using Protter52). Green highlights indicate reported talquetamab binding domains (Jeong et al.32). Red highlights mark all SNV/ indel mutation sites identified in this study. *D239 represents talquetamab binding site that is also mutated. n indicates number of patients harboring same mutations.
Extended Data Fig. 9 Membrane localization of wild-type and p.Tyr257Ser GPRC5D in K562 cells.
Figures illustrate representative confocal images of the fluorescence emission of A. wild-type GPRC5D and B. p.Tyr257Ser GPRC5D expressing K562 cells labeled with a membrane dye (CytoLiner) as well as indirect immunofluorescence of the HA tag (GPRC5D). Cyan represents the Cytoliner fluorescence emission and magenta represents the corresponding immunofluorescence emission from the indirect antibody labeling of HA. In each panel, cyan is the leftmost image, magenta is the middle, and the rightmost shows a composite image with both channels overlaid. The side view (a cut through the z-stack) is also shown for each channel. The fluorescence emission overlaps for both the membrane and HA, supporting membrane localization of the fusion protein. Scale bar, 5 µm. The figure is representative of 2 independent experiments.
Extended Data Fig. 10 Reduced chromatin accessibility and transcriptional silencing of GPRC5D in t(11;14) MM.
A. Genome accessibility track of the GPRC5D locus (chr12: 12,782,146–13,222,147) visualized with peak-to-gene links (Peak2Gene Links) identified using ArchR40. Arc color denotes correlation strength between peak accessibility and GPRC5D expression; darker arcs indicate stronger correlations. Single-cell GPRC5D mRNA gene expression across patient samples is shown to the right of the trackplot. t(11;14) MM samples are highlighted with blue boxes. The lower portion of the trackplot shows annotated genes within the region, with red labels for genes on the positive strand (TSS left) and blue for genes on the negative strand (TSS right). Genomic coordinates and associated false discovery rates (FDR) for peak-to-gene correlations are provided in Supplementary Table 9. Highlighted in blue are peaks differentially accessible between t(11;14) and non-t(11;14) cases denoting reduced chromatin accessibility at GPRC5D gene body and other regulatory regions. B. Comparison of GPRC5D transcript expression in t(11;14) versus non-t(11;14) multiple myeloma patient samples. Box plots display the median (horizontal line), interquartile range (boxes; 25th-75th percentile), and whiskers extending to 1.5 times the IQR. Individual dots represent values outside this range. Analysis was performed comparing the ArchR GPRC5D mRNA expression using a two-sided Wilcoxon rank-sum test on samples from patients with t(11;14) (n = 6) and non-t(11;14) (n = 9). Shown is the Benjamini-Hochberg adjusted P value for this comparison (FDR = 1.04219e-153).
Supplementary information
Supplementary Information (download PDF )
Supplementary Figs. 1–19, figure legends and table legends.
Supplementary Tables (download XLSX )
Excel file including 13 Supplementary Tables.
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig. 12 (top left).
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig. 12 (top right).
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig. 12 (bottom left).
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig16 (top left).
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig. 16 (top middle).
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig. 16 (top right).
Supplementary Data (download JPG )
Unprocessed western blot for Supplementary Fig. 16 (bottom).
Source data
Source Data Fig. 1 (download JPG )
Unprocessed western blot for Fig. 5d (top).
Source Data Fig. 2 (download JPG )
Unprocessed western blot for Fig. 5d (bottom).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lee, H., Ahn, S., Gonzales, G.A. et al. Multimodal antigenic escape to GPRC5D-targeted T cell engagers in multiple myeloma. Nat Med 32, 964–977 (2026). https://doi.org/10.1038/s41591-025-04175-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41591-025-04175-8

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}