
Abstract
Brain health is closely linked to bone homeostasis. Skeletal aging is characterized by inadequate bone formation and marrow adiposity, but whether the brain contributes to this imbalance remains unknown. This study shows that aged brain neurons, mainly those in the hippocampus and cerebral cortex, produce excess WD repeat and FYVE domain containing 1 (WDFY1) protein and transfer it to the bone via extracellular vesicles (EVs), leading to bone-fat imbalance and osteoporosis. Increasing brain Wdfy1 expression causes premature skeletal aging. Conversely, suppressing Wdfy1 in the whole brain, hippocampus or neurons, genetically deleting neuronal Wdfy1, and selectively inhibiting neuronal EV release all improve bone health. Mechanistically, WDFY1 binds to the retromer complex to promote the endosome-to-Golgi recycling of cathepsin D and peroxiredoxin 2, thus inhibiting osteogenesis and augmenting adipogenesis. This study identifies the role of aged brain neuronal EVs as an important messenger in triggering bone-fat imbalance by transferring WDFY1 to bone.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others

Data availability
This paper does not involve original code. There are restrictions to the availability of RVG-9R-siWdfy1 due to pending patent application. However, the data are available from the corresponding author upon reasonable request for the purpose of academic research and validation. Requestors may need to sign a confidentiality or material transfer agreement to prevent compromise of the pending patent.
Raw proteomic data eXchange Consortium (https://proteomecentral.proteomexchange.org) via the iProX partner repository51,52 with the dataset identifier PXD069570. All other data and materials supporting the results of this study are provided in the article, supplementary information file or source data or available from the corresponding authors upon reasonable request.
References
Mikail, N. et al. Imaging of the brain-heart axis: prognostic value in a European setting. Eur. Heart J. 45, 1613–1630 (2024).
Xia, W. et al. Damaged brain accelerates bone healing by releasing small extracellular vesicles that target osteoprogenitors. Nat. Commun. 12, 6043 (2021).
Zhang, X. et al. Brain control of humoral immune responses amenable to behavioural modulation. Nature 581, 204–208 (2020).
Ma, C. et al. Autonomic neural regulation in mediating the brain-bone axis: mechanisms and implications for regeneration under psychological stress. Qjm 117, 95–108 (2024).
Ducy, P. et al. Leptin inhibits bone formation through a hypothalamic relay: a central control of bone mass. Cell 100, 197–207 (2000).
Takeda, S. et al. Leptin regulates bone formation via the sympathetic nervous system. Cell 111, 305–317 (2002).
Otto, E. et al. Crosstalk of brain and bone-clinical observations and their molecular bases. Int. J. Mol. Sci. 21, 4946 (2020).
Zhang, F. & Zhang, W. Research progress in Alzheimer’s disease and bone-brain axis. Ageing Res. Rev. 98, 102341 (2024).
Xie, C., Wang, C. & Luo, H. Increased risk of osteoporosis in patients with cognitive impairment: a systematic review and meta-analysis. BMC Geriatr. 23, 797 (2023).
Ximerakis, M. et al. Single-cell transcriptomic profiling of the aging mouse brain. Nat. Neurosci. 22, 1696–1708 (2019).
Hu, S., Hu, Y. & Yan, W. Extracellular vesicle-mediated interorgan communication in metabolic diseases. Trends Endocrinol. Metab. 34, 571–582 (2023).
Bischoff, J. P., Schulz, A. & Morrison, H. The role of exosomes in intercellular and inter-organ communication of the peripheral nervous system. FEBS Lett. 596, 655–664 (2022).
Rao, S. et al. Extracellular vesicles from human urine-derived stem cells delay aging through the transfer of PLAU and TIMP1. Acta Pharm. Sin. B 14, 1166–1186 (2024).
Wang, Z. X. et al. Aged bone matrix-derived extracellular vesicles as a messenger for calcification paradox. Nat. Commun. 13, 1453 (2022).
Liu, X. et al. Brain-derived extracellular vesicles promote bone-fat imbalance in Alzheimer’s disease. Int. J. Biol. Sci. 19, 2409–2427 (2023).
Yu, B. & Wang, C. Y. Osteoporosis: the result of an ‘aged’ bone microenvironment. Trends Mol. Med. 22, 641–644 (2016).
Zhang, Y. et al. Neuronal induction of bone-Fat imbalance through osteocyte neuropeptide Y. Adv. Sci. 8, e2100808 (2021).
Men, Y. et al. Exosome reporter mice reveal the involvement of exosomes in mediating neuron to astroglia communication in the CNS. Nat. Commun. 10, 4136 (2019).
Krämer-Albers, E. M. Extracellular vesicles at CNS barriers: mode of action. Curr. Opin. Neurobiol. 75, 102569 (2022).
Ma, S. et al. Caloric restriction reprograms the single-cell transcriptional landscape of Rattus Norvegicus aging. Cell 180, 984–1001.e1022 (2020).
Lein, E. S. et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 445, 168–176 (2007).
Wang, X., Zhang, C., Szábo, G. & Sun, Q. Q. Distribution of CaMKIIα expression in the brain in vivo, studied by CaMKIIα-GFP mice. Brain Res. 1518, 9–25 (2013).
Ostrowski, M. et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 12, 19–30 (2010). sup pp 11-13.
Kumar, P. et al. Transvascular delivery of small interfering RNA to the central nervous system. Nature 448, 39–43 (2007).
Hu, Y. H. et al. WDFY1 mediates TLR3/4 signaling by recruiting TRIF. EMBO Rep. 16, 447–455 (2015).
Nandakumar, R. & Paludan, S. R. Catching the adaptor-WDFY1, a new player in the TLR-TRIF pathway. EMBO Rep. 16, 397–398 (2015).
Zhu, X. et al. The critical role of toll-like receptor 4 in bone remodeling of osteoporosis: from inflammation recognition to immunity. Front. Immunol. 15, 1333086 (2024).
Qi, C., Xiaofeng, X. & Xiaoguang, W. Effects of toll-like receptors 3 and 4 in the osteogenesis of stem cells. Stem Cells Int. 2014, 917168 (2014).
Seaman, M. N. The retromer complex - endosomal protein recycling and beyond. J. Cell Sci. 125, 4693–4702 (2012).
Cullen, P. J. & Steinberg, F. To degrade or not to degrade: mechanisms and significance of endocytic recycling. Nat. Rev. Mol. Cell Biol. 19, 679–696 (2018).
Xiong, L. et al. Retromer in osteoblasts interacts with protein phosphatase 1 regulator subunit 14C, terminates parathyroid hormone’s signaling, and promotes its catabolic response. EBioMedicine 9, 45–60 (2016).
Xia, W. F. et al. Vps35 loss promotes hyperresorptive osteoclastogenesis and osteoporosis via sustained RANKL signaling. J. Cell Biol. 200, 821–837 (2013).
Masson, O. et al. Cathepsin-D, a key protease in breast cancer, is up-regulated in obese mouse and human adipose tissue, and controls adipogenesis. PLoS One 6, e16452 (2011).
Kim, K. M. et al. Peroxiredoxin II negatively regulates BMP2-induced osteoblast differentiation and bone formation via PP2A Cα-mediated Smad1/5/9 dephosphorylation. Exp. Mol. Med. 51, 1–11 (2019).
Kim, M. H. et al. Peroxiredoxin 2 deficiency reduces white adipogenesis due to the excessive ROS generation. Cell Biol. Int. 44, 2086–2093 (2020).
Shi, H. & Chen, M. The brain-bone axis: unraveling the complex interplay between the central nervous system and skeletal metabolism. Eur. J. Med. Res. 29, 317 (2024).
Shi, T. et al. Osteocyte-derived sclerostin impairs cognitive function during ageing and Alzheimer’s disease progression. Nat. Metab. 6, 531–549 (2024).
Khrimian, L. et al. Gpr158 mediates osteocalcin’s regulation of cognition. J. Exp. Med. 214, 2859–2873 (2017).
Jiang, Y. L. et al. The protective effects of osteocyte-derived extracellular vesicles against Alzheimer’s disease diminished with aging. Adv. Sci. 9, e2105316 (2022).
Yang, Y., Hu, Y. H. & Liu, Y. Wdfy1 deficiency impairs Tlr3-mediated immune responses in vivo. Cell Mol. Immunol. 17, 1014–1016 (2020).
Yeo, I. J. et al. PRDX6 inhibits neurogenesis through downregulation of WDFY1-mediated TLR4 signal. Mol. Neurobiol. 56, 3132–3144 (2019).
Ouyang, L. et al. p39-associated Cdk5 activity regulates dendritic morphogenesis. Sci. Rep. 10, 18746 (2020).
Mei, Z. et al. Lowering hippocampal miR-29a expression slows cognitive decline and reduces beta-amyloid deposition in 5×FAD mice. Mol. Neurobiol. 61, 3343–3356 (2024).
Dutta, S. et al. Neuropilin-2 regulates endosome maturation and EGFR trafficking to support cancer cell pathobiology. Cancer Res. 76, 418–428 (2016).
Dutta, S. et al. NRP2 transcriptionally regulates its downstream effector WDFY1. Sci. Rep. 6, 23588 (2016).
Vagnozzi, A. N. & Praticò, D. Endosomal sorting and trafficking, the retromer complex and neurodegeneration. Mol. Psychiatry 24, 857–868 (2019).
Redlich, K. & Smolen, J. S. Inflammatory bone loss: pathogenesis and therapeutic intervention. Nat. Rev. Drug Discov. 11, 234–250 (2012).
Zhang, Y. Y. et al. Insights and implications of sexual dimorphism in osteoporosis. Bone Res. 12, 8 (2024).
Chen, C. Y. et al. Glucocorticoid-induced loss of beneficial gut bacterial extracellular vesicles is associated with the pathogenesis of osteonecrosis. Sci. Adv. 8, eabg8335 (2022).
Hou, J. et al. Aged bone marrow macrophages drive systemic aging and age-related dysfunction via extracellular vesicle-mediated induction of paracrine senescence. Nat. Aging 4, 1562–1581 (2024).
Ma, J. et al. iProX: an integrated proteome resource. Nucleic Acids Res. 47, D1211–D1217 (2019).
Chen, T. et al. iProX in 2021: connecting proteomics data sharing with big data. Nucleic Acids Res. 50, D1522–D1527 (2022).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (grants 82172501 to C.-Y.C., 82125023 to H.X., 82372479 to C.-Y.C., 82072504 to H.X., 81871822 to H.X., 82101463 to G.-W.H., and 82372471 to G.-W.H.), the China National Postdoctoral Program for Innovative Talents (grant BX2021383 to C.-Y.C.), the Natural Science Foundation of Hunan Province (grants 2023JJ10094 to C.-Y.C. and 2024JJ6668 to Z.W.), the Science and Technology Innovation Program of Hunan Province (grant 2022RC1211 to C.-Y.C.), the Central South University Innovation-Driven Research Programme (grant 2023CXQD001 to C.-Y.C.), the Jiangxi Province’s Science and Technology Agency Support Program (grant 20224ACB216007 to G.-W.H.), the China Postdoctoral Science Foundation (grants 2023T160739 to S.-S.R. and 2023M733956 to S.-S.R.), the Postdoctoral Fellowship Program of CPSF (grant No. GZB20230871 to S.-S.R.), the Youth Science Foundation of Xiangya Hospital (grants 2022Q07 to S.-S.R. and 2023Q15 to Z.W.) and the Postgraduate Innovative Project of Central South University (grants 2024ZZTS0925 to H.-J.Z. and 2025ZZTS0958 to Y.-X.D.). The schematic diagram in Fig. 8b was created using BioRender, with agreement number AC28V53YLH.
Author information
Authors and Affiliations
Contributions
H.X., C.-Y.C. and S.-S.R. designed the study. C.-Y.C., H.X., S.-S.R., Z.W., Y.L. and X.W. wrote the manuscript. C.-Y.C., S.-S.R., C.-G.H., Y.-J.T., Y.-X.D., Y.L., X.W., H.-J.Z., J.-Y.L., T.-F.W., H.Y., H.Z. and Z.-H.H. performed the experiments or/and analyzed the data. C.-Y.C., S.-S.R. and Z.W. prepared the figures. X.-X.L., Y.Z., Z.-G.W., X.-Y.H., G.-W.H., H.-L.L., Z.-X.W. and J.C. provided technical support.
Corresponding authors
Ethics declarations
Competing interests
H.X., C.-Y.C., S.-S.R., Y.L., Y.-X.D. and Z.W. are inventors of a submitted patent application related to this article. All other authors declare no competing interests.
Peer review
Peer review information
Nature Aging thanks Xiaochun Bai, Hirotaka Iijima and Mone Zaidi for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Detrimental effects of AB-EVs on bone phenotypes.
a, Ex vivo fluorescent imaging of tissues from DiR-labeled YB-EVs- or AB-EVs-treated mice for 3 h by intracerebroventricular (I.C.V.) or intravenous (I.V.) injection. Scale bar: 6 mm. b–d, Femoral μCT images (b), bone microstructural parameters (c), and ultimate load (d) in vehicle-, YB-EVs-, or AB-EVs-treated 4-month-old young female mice. Scale bar: 1 mm. n = 8 (YB-EVs or AB-EVs) or 9 (vehicle) per group. e, f, qRT-PCR for Runx2 (e) and Pparγ (f), and ELISA for serum OCN (e) in 4-month-old young male mice receiving different treatments. qRT-PCR: n = 8 (YB-EVs) or 9 (vehicle or AB-EVs) per group; ELISA: n = 10 per group. g–i, Tartrate-resistant acid phosphatase (TRAP) staining images (g), osteoclast (OC) numbers (h), and ELISA for serum C-terminal telopeptides of type I collagen (CTX-I; i) in 4-month-old young male mice with different treatments. Scale bar: 50 μm. n = 10 per group. j–l, qRT-PCR for Runx2 (j) and Pparγ (k), ELISA for serum OCN (j) and CTX-I (l), and TRAP+ osteoclast numbers (l) in 15-month-old aged male mice with different treatments. n = 9 (qRT-PCR) or 10 (ELISA and TRAP) per group. m, n, Representative images (m) and quantification for the mean intensity (n) of the Dil (red)-labeled YB-EVs and AB-EVs in BMSCs. Scale bar: 50 μm. n = 5 per group. Data are mean ± SD. Statistics: one-way ANOVA with Bonferroni post hoc test (c–f, h, i, and n); unpaired, two-tailed Student’s t-test (j–l).
Extended Data Fig. 2 Plasma EVs derived from aged people inhibit osteogenesis and promote adipogenesis of BMSCs.
a, b, ARS and ORO staining images (a) and quantification of ARS+ and ORO+ areas (b) in BMSCs treated with vehicle or plasma EVs from aged or young people (A-Pla-EVs or Y-Pla-EVs) under osteogenic or adipogenic induction. Scale bar: 200 μm (ARS) or 100 μm (ORO). n = 3 per group. Data are mean ± SD. Statistics: one-way ANOVA with Bonferroni post hoc test (b).
Extended Data Fig. 3 Effects of different doses of WDFY1 on osteoclast formation.
a, b, TRAP staining images of RAW264.7 cells receiving different treatments under osteoclastic induction (a) and quantification of osteoclast numbers per well in a 48-well plate (b). Scale bar: 50 μm. n = 3 per group. Data are mean ± SD. Statistics: one-way ANOVA with Bonferroni post hoc test (b).
Extended Data Fig. 4 Effects of brain Wdfy1 overexpression or downregulation on osteogenesis, adipogenesis, and inflammation.
a–d, qRT-PCR for Runx2 (a) and Pparγ (b) expression in femur tissues, and ELISA for serum OCN (c) and pro-inflammatory factors including interleukin-1α (IL-1α), IL-1β, IL-6, and tumor necrosis factor α (TNF-α) (d) in 4-month-old young female mice treated with AAV2-Con or AAV2-Wdfy1 by intracerebroventricular injection. n = 14 (AAV2-Con) or 10 (AAV2-Wdfy1) per group. e–h, qRT-PCR for Runx2 (e) and Pparγ (f) expression in femur tissues, and ELISA for serum OCN (g) and pro-inflammatory factors (h) in 15-month-old male mice treated with AAV2-shCon or AAV2-shWdfy1. n = 6 per group. Data are mean ± SD. Statistics: unpaired, two-tailed Student’s t-test.
Extended Data Fig. 5 Hippocampal Wdfy1 overexpression or downregulation alters bone phenotypes and the bone metabolism-regulatory role of Ser-EVs.
a, b, Femoral μCT images (a) and bone microstructural parameters (b) in AAV2-Con- or AAV2-Wdfy1-treated 4-month-old young female mice, and from AAV2-shCon- or AAV2-shWdfy1-treated 15-month-old male mice. H.I.P: hippocampal injection. Scale bar: 1 mm. n = 10 (young) or 7 (aged) per group. c, Three-point bending test of femoral ultimate load. n = 8 (young) or 7 (aged) per group. d, qRT-PCR for femoral Runx2 expression and ELISA for serum OCN. qRT-PCR: n = 9 (young) or 7 (aged) per group. ELISA: n = 9 (young: AAV2-Con-H.I.P), 8 (young: AAV2-Wdfy1-H.I.P), or 7 (aged: both groups). e, ORO staining images and quantification of ORO+ areas in distal femurs. Scale bar: 50 μm. n = 10 (young) or 7 (aged) per group. f, qRT-PCR for femoral Pparγ expression. n = 9 (young: AAV2-Con-H.I.P), 10 (young: AAV2-Wdfy1-H.I.P), or 7 (aged: both groups). g–i, qRT-PCR for femoral Runx2 (g) and Pparγ (h), and ELISA for serum OCN (g) and CTX-I (i) in vehicle- or WDFY1-treated 4-month-old young female mice by intramedullary injection. n = 9 (qRT-PCR for Runx2 in WDFY1 group) or 10 (others) per group. j–l, Femoral μCT images (j), bone microstructural parameters (k), and ultimate load (l) in 4-month-old young female mice treated with Ser-EVs from different groups. Scale bar: 1 mm. n = 10 per group. Data are mean ± SD. Statistics: unpaired, two-tailed Student’s t-test (b–i, k, and l).
Extended Data Fig. 6 Neuronal Wdfy1 deletion increases osteogenesis, decreases adipogenesis, suppresses inflammation, and downregulates AB-EVs’ function in aged mice.
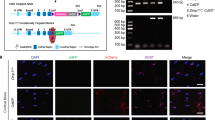
a, Co-staining for GFAP/WDFY1, IBA1/WDFY1, and MBP/WDFY1 in the hippocampus and cerebral cortex from 18-month-old aged mice. Scale bar: 50 μm. b, Schematic diagram of the targeting strategy for generating mice bearing the conditional Wdfy1 allele, with exon 2 flanked by loxP sites. c, Representative PCR gels showing genotyping for the loxP site (left), Camk2a-CreERT2 (middle), and Map2-CreERT2 (right) alleles across the different mouse lines. d–g, qRT-PCR for femoral Runx2 (d) and Pparγ (e), and ELISA for serum OCN (d), CTX-I (f), and pro-inflammatory factors (IL-1α, IL-1β, IL-6, and TNF-α; g) in mice of different genotypes receiving tamoxifen at 15-month-old and left for 2 months. n = 6 (Wdfy1fl/fl and Camk2a-CreERT2; Wdfy1fl/fl) or 8 (Map2-CreERT2; Wdfy1fl/fl) per group. h–k, Levels of hematological and organ function parameters including hemoglobin and red blood cells (h), neutrophil and lymphocyte percentages (i), and indicators of liver (j) or kidney (k) function. ALT: alanine transaminase; AST: aspartate aminotransferase; CREA: creatinine; BUN: blood urea nitrogen. n = 6 (Wdfy1fl/fl and Camk2a-CreERT2; Wdfy1fl/fl) or 8 (Map2-CreERT2; Wdfy1fl/fl) per group. l, m, ARS and ORO staining images (l) and quantification of ARS+ and ORO+ areas (m) in BMSCs with different treatments. Scale bar: 200 μm (ARS) or 100 μm (ORO). n = 3 per group. Data are mean ± SD. Statistics: one-way ANOVA with Bonferroni post hoc test (c–k and m).
Extended Data Fig. 7 Inhibition of neuronal EV release in aged mice increases osteogenesis, decreases adipogenesis, and impairs AB-EVs’ function.
a, Schematic diagram of the targeting strategy for generating mice bearing the floxed Rab27b allele, with exon 4 flanked by loxP sites. b, Representative PCR gels showing genotyping for the loxP site (up) and Map2-CreERT2 (bottom) alleles across the different mouse lines. c–e, qRT-PCR for femoral Runx2 (c) and Pparγ (d), and ELISA for serum OCN (c) and CTX-I (e) in Rab27bfl/fl and Map2-CreERT2; Rab27fl/fl mice receiving tamoxifen at 15-month-old and left for 2 months. n = 8 (all parameters in Rab27bfl/fl mice), 9 (Pparγ in Map2-CreERT2; Rab27fl/fl mice), or 10 (other parameters in Map2-CreERT2; Rab27fl/fl mice) per group. f, Protein contents of AB-EVs normalized to brain weight in the indicated mouse lines. n = 5 per group. g, Western blotting for WDFY1 levels in AB-EVs from mice of the indicated genotypes. n = 3 per group. h, i, ARS and ORO staining images (h) and quantification of ARS+ and ORO+ areas (i) in BMSCs receiving different treatments. Scale bar: 200 μm (ARS) or 100 μm (ORO). n = 3 per group. Data are mean ± SD. Statistics: unpaired, two-tailed Student’s t-test (c–f, and i).
Extended Data Fig. 8 Selectively suppressing neuronal Wdfy1 expression enhances osteogenesis and reduces adipogenesis.
a–c, qRT-PCR for Runx2 (a) and Pparγ (b) expression in femur tissues and ELISA for serum OCN (a) and CTX-I (c) in 16-month-old aged male mice treated with RV-MAT-9R-siCon, RV-MAT-9R-siWdfy1, RVG-9R-siCon, or RVG-9R-siWdfy1. n = 7 (RVG-9R-siWdfy1), 8 (RV-MAT-9R-siWdfy1), or 9 (RV-MAT-9R-siCon and RVG-9R-siCon groups). Data are mean ± SD. Statistics: two-way ANOVA with Bonferroni post hoc test.
Extended Data Fig. 9 WDFY1 exerts harmful effects on bone independent of TLR3/4 signaling.
a, b, Femoral μCT images (a) and quantification of Tb. BV/TV, Tb. N, Tb. Th, and Ct. Th (b) in 12-month-old Tlr3−/− male mice treated with AAV2-Con or AAV2-Wdfy1. Scale bar: 1 mm. n = 9 (AAV2-Con) or 10 (AAV2-Wdfy1) per group. c, d, Femoral μCT images (c) and quantification of Tb. BV/TV, Tb. N, Tb. Th, and Ct. Th (d) in 12-month-old Tlr4−/− male mice treated with AAV2-Con or AAV2-Wdfy1. Scale bar: 1 mm. n = 6 (AAV2-Con) or 7 (AAV2-Wdfy1) per group. Data are mean ± SD. Statistics: unpaired, two-tailed Student’s t-test (b and d).
Extended Data Fig. 10 WDFY1 binds to the retromer VPS proteins independent of the FYVE domain.
a, Direct binding between WDFY-Flag and four VPS proteins (VPS29-His, VPS26A-His, VPS26B-His, and VPS35-His) determined by anti-Flag-pull-down assay. b, Western blot analysis of Flag and WDFY1 expression in CHO cells transfected with the plasmids carrying full-length Wdfy1, one or several WD domains-deleted (ΔWD) mutant Wdfy1, or ΔFYVE mutant Wdfy1. c, Co-immunoprecipitation of full-length WDFY1-Flag or ΔFYVE mutant WDFY1-Flag with four VPS proteins after transfection of plasmids into CHO cells.
Supplementary information
Supplementary Information (download PDF )
Supplementary Tables 3–7.
Supplementary Table 1 (download XLSX )
Differentially expressed proteins between AB-EVs and YB-EVs.
Supplementary Table 2 (download XLSX )
Proteins differentially pulled down by WDFY1-Flag compared to controls.
Supplementary Data (download XLSX )
Source Data for Supplementary Table 7: Clinical information for human donors.
Source data
Source Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Fig. 1 (download PDF )
Unprocessed images and western blots.
Source Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Fig. 2 (download PDF )
Unprocessed images and western blots.
Source Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Fig. 3 (download PDF )
Unprocessed images and western blots.
Source Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Fig. 4 (download PDF )
Unprocessed images.
Source Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Fig. 5 (download PDF )
Unprocessed images.
Source Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Fig. 6 (download PDF )
Unprocessed images and western blots.
Source Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Fig. 7 (download PDF )
Unprocessed images and western blots.
Source Data Fig. 8 (download PDF )
Unprocessed western blots.
Source Data Extended Data Fig. 1 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 1 (download PDF )
Unprocessed images.
Source Data Extended Data Fig. 2 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 2 (download PDF )
Unprocessed images.
Source Data Extended Data Fig. 3 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 3 (download PDF )
Unprocessed images.
Source Data Extended Data Fig. 4 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 5 (download PDF )
Unprocessed images.
Source Data Extended Data Fig. 6 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 6 (download PDF )
Unprocessed images and gels.
Source Data Extended Data Fig. 7 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 7 (download PDF )
Unprocessed images, gels, and western blots.
Source Data Extended Data Fig. 8 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 9 (download XLSX )
Statistical source data.
Source Data Extended Data Fig. 9 (download PDF )
Unprocessed images.
Source Data Extended Data Fig. 10 (download PDF )
Unprocessed western blots.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chen, CY., Wang, Z., Hong, CG. et al. Brain neuron-derived WDFY1 induces bone loss. Nat Aging 6, 329–348 (2026). https://doi.org/10.1038/s43587-025-01032-8
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s43587-025-01032-8
